Submitted:
27 June 2025
Posted:
30 June 2025
You are already at the latest version
Abstract
The sustainable management of farmland soils is fundamental to addressing the intertwined challenges of food security, environmental degradation, and climate change. This review synthesizes current knowledge on key management practices—crop rotation, no-tillage agriculture, organic amendments (specifically farmyard manure), and soil microbiome regulation—and their synergistic effects on soil health and crop productivity. Crop rotation disrupts pest and disease cycles, enhances nutrient cycling, and stabilizes yields. No-tillage improves soil physical properties, promotes carbon sequestration, and supports more diverse and resilient microbial communities. Organic amendments enrich soil organic matter, stimulate microbial-mediated nutrient cycling, and improve soil fertility over the long term. Targeted management of soil microbiomes further boosts plant stress resistance, nutrient acquisition, and disease suppression, offering powerful avenues for ecosystem resilience. Critically, the integration of these practices amplifies their individual benefits. Systems that combine rotation with no-tillage, or organic amendments with conservation practices, demonstrate superior performance in enhancing soil structure, nutrient dynamics, biological diversity, and carbon storage. Precision agriculture technologies and microbiome-based interventions are poised to refine these integrated systems further, enabling site-specific optimization. Despite technical and operational challenges—such as early-stage yield variability and management complexities—synergistic soil health management offers a clear pathway toward regenerative, climate-resilient agriculture. Future research must focus on understanding microbial functional dynamics, advancing real-time soil health monitoring, and developing holistic, scalable strategies that align productivity goals with ecological stewardship. An integrated, ecosystem-based approach to farmland management is essential to achieve sustainable agricultural development and global carbon neutrality targets.
Keywords:
crop rotation
; no-tillage agriculture
; organic amendments
; soil microbiome
; sustainable farmland management
; carbon sequestration
1. Introduction
Global agriculture is facing unprecedented challenges in sustaining productivity while preserving soil and environmental health. Over the past few decades, intensive agricultural practices have led to widespread soil degradation, nutrient depletion, and escalating pressures from pests and diseases. A combination of factors—including population growth, climate change, and unsustainable land management—has exacerbated the vulnerability of agroecosystems worldwide. According to the United Nations Food and Agriculture Organization (FAO), an estimated 33% of global soils are moderately to highly degraded due to erosion, loss of organic carbon, salinization, acidification, compaction, and contamination. Soil degradation not only threatens food security but also undermines ecosystem services critical for human well-being, such as carbon sequestration, water filtration, and biodiversity conservation [1]. Nutrient depletion is a particularly pressing issue. Intensive cropping systems continuously remove large amounts of nitrogen (N), phosphorus (P), and potassium (K) from the soil without adequate replenishment, leading to declining soil fertility over time[2]. Moreover, soil organic carbon (SOC) stocks have been significantly reduced in many agricultural regions, impairing soil structure, water retention capacity, and microbial activity[3]. Simultaneously, pest and disease pressures have intensified under monoculture regimes, causing frequent crop losses and increasing reliance on chemical pesticides [4]. This situation demands transformative approaches to agricultural management that can reconcile productivity goals with environmental stewardship.
Traditional monoculture and heavy tillage practices have played a central role in accelerating soil degradation processes. Monoculture—the continuous cultivation of a single crop species on the same land—disrupts nutrient cycling, homogenizes microbial communities, and creates ecological niches favorable to specialized pests and pathogens [5]. Long-term monocropping systems, such as continuous maize or soybean, have been linked to increased incidences of soil-borne diseases like Fusarium wilt and root rots, leading to yield declines and higher input costs [6]. Heavy tillage, involving repeated plowing and soil inversion, was historically adopted to control weeds, incorporate residues, and prepare seedbeds. However, it has profound negative impacts on soil health. Intensive tillage disrupts soil structure, increases erosion rates, accelerates organic matter mineralization, and reduces microbial habitat stability [7]. Research shows that tillage-induced soil degradation results in lower aggregate stability, diminished water infiltration, and increased vulnerability to drought and flood stresses [8]. Furthermore, the breakdown of soil aggregates releases previously stabilized organic carbon into the atmosphere as carbon dioxide (CO₂), contributing to climate change [9].
The cumulative consequences of monoculture and heavy tillage systems include nutrient imbalances, declining biological diversity, reduced resilience to environmental shocks, and diminishing economic returns over time. Hence, there is an urgent need to transition toward diversified and conservation-oriented agricultural systems that enhance soil health and ecosystem functioning. A suite of sustainable land management practices has emerged as promising solutions to the challenges outlined above. Among these, four practices are particularly noteworthy: crop rotation, no-tillage (NT), organic amendments, and the strategic management of soil microbiomes.
Crop rotation, the sequential planting of different crop species on the same land, is a time-honored practice known to enhance nutrient use efficiency, break pest and disease cycles, and improve soil structure [10]. Rotations that include legumes can biologically fix atmospheric nitrogen, reducing the need for synthetic fertilizers [11]. Diversified rotations have also been shown to increase SOC, enhance microbial diversity, and stabilize yields across variable climatic conditions [12,13]. No-tillage agriculture minimizes soil disturbance by planting crops directly into residues from previous crops without plowing. By preserving soil structure and maintaining continuous residue cover, no-tillage practices reduce erosion, promote carbon sequestration, and improve water infiltration[14]. However, no-tillage systems often require complementary strategies, such as cover cropping and integrated weed management, to address potential challenges like weed proliferation and initial yield lags [15,16]. Organic amendments, such as animal manure, compost, and crop residues, are critical for replenishing soil organic matter and providing essential nutrients. Organic inputs not only improve physical soil properties (aggregation, porosity, water holding capacity) but also stimulate microbial activity and functional diversity [17]. Integrated nutrient management—combining organic and inorganic inputs—has been shown to optimize crop yields while maintaining long-term soil fertility[18]. Soil microbiomes, the complex communities of bacteria, fungi, archaea, and protozoa inhabiting soils, are increasingly recognized as fundamental drivers of soil health and plant productivity. Beneficial microbes facilitate nutrient cycling, suppress pathogens, enhance stress tolerance, and contribute to ecosystem stability [19,20]. Emerging approaches, such as microbial inoculation, microbiome engineering, and precision microbiome management, offer exciting opportunities to further optimize soil biological functioning[21,22].
Collectively, these four strategies represent complementary pillars of sustainable agriculture. Their integration can produce synergistic effects greater than the sum of their individual contributions, offering pathways to resilient, productive, and climate-smart agroecosystems.
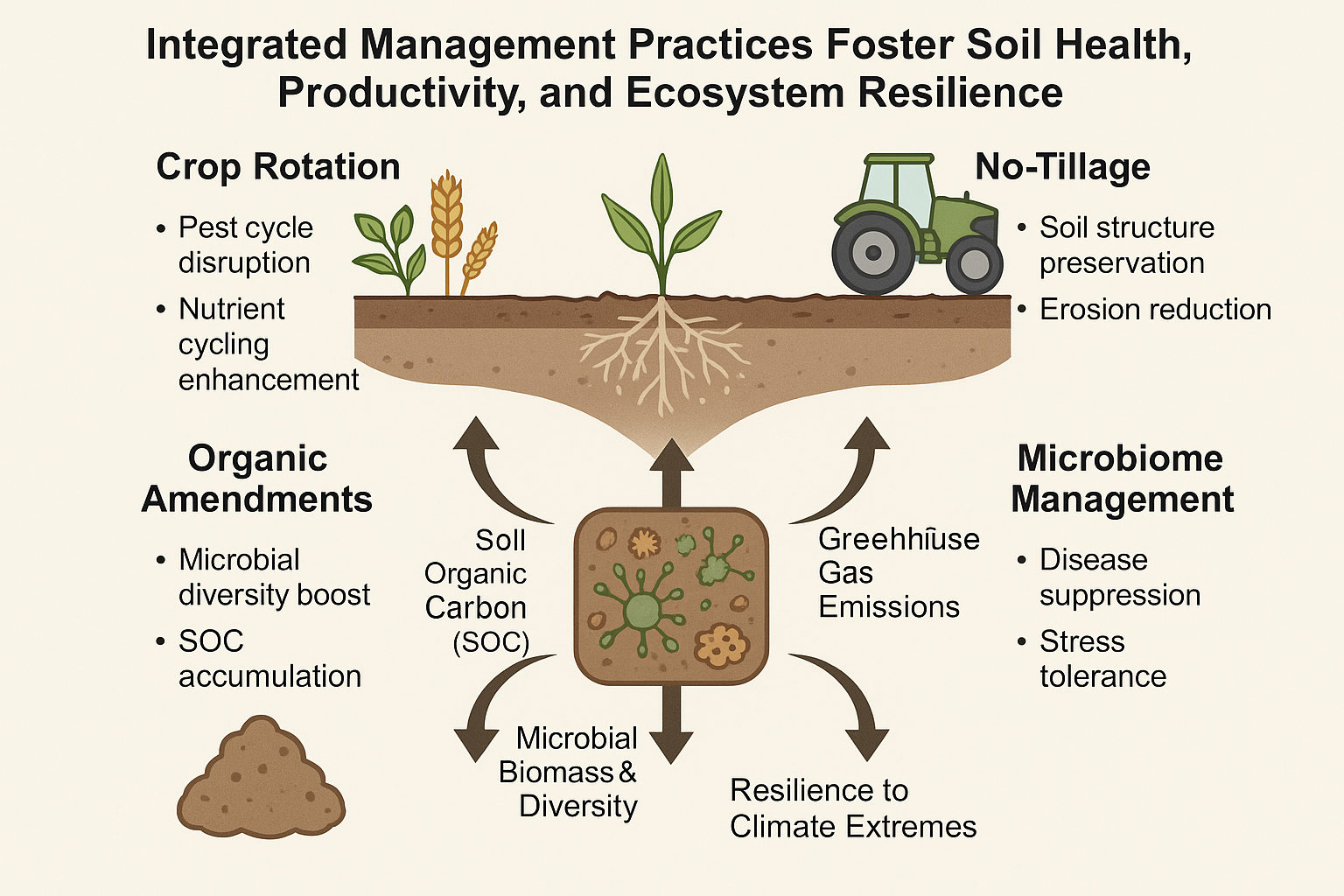
Despite the growing body of research on sustainable farming practices, most studies examine individual strategies (e.g., rotation alone, or no-till alone) in isolation. Few comprehensive syntheses exist that integrate crop rotation, no-tillage, organic amendments, and soil microbiome management into a unified framework for sustainable farmland management. To summarize, sustainable agricultural practices such as crop rotation, no-tillage, organic amendments, and microbiome management contribute synergistically to soil health and ecosystem resilience (Figure 1). This conceptual framework provides the foundation for the present study’s investigation into Soil-Health-Based Sustainable Farming Strategies.
The primary objectives of this review are therefore fourfold:
- To synthesize recent (2001–2025) advances in understanding how crop rotation, no-tillage, organic amendments, and microbiome management individually and synergistically contribute to soil health and agricultural sustainability.
- To critically evaluate the ecological mechanisms underpinning these practices, including nutrient cycling, soil structure formation, pest and disease suppression, and microbial dynamics.
- To identify key challenges, knowledge gaps, and trade-offs associated with implementing integrated soil management systems under diverse agroecological conditions.
- To propose future research directions and practical strategies for optimizing farmland management in the context of climate change and resource constraints.
By comprehensively linking agronomic, soil ecological, and microbiological perspectives, this review aims to provide a holistic foundation for advancing sustainable agriculture. It highlights that truly sustainable farmland management must move beyond single-practice solutions toward integrated, systems-level approaches that restore and harness the natural functioning of soils.
Ultimately, achieving agricultural sustainability is not only a technical endeavor but a necessity for ensuring global food security, preserving environmental integrity, and meeting international sustainability goals such as the United Nations Sustainable Development Goals (SDGs) [23].
2. Review Methodology
To ensure a comprehensive and unbiased synthesis of current knowledge, we adopted a systematic review framework grounded in the PRISMA-ScR (Preferred Reporting Items for Systematic Reviews and Meta-Analyses for Scoping Reviews) guidelines. Our review focused on literature published between January 2001 and June 2025, supplemented by the selective inclusion of influential earlier studies where necessary for conceptual or methodological context. We searched across multiple scholarly databases, including Web of Science, Scopus, and Google Scholar, using combinations of the terms “Crop rotation” and “No-tillage agriculture”. These keywords were chosen to capture a broad and representative range of research related to sustainable agricultural practices and their impacts on soil, crops, and the environment.
Our inclusion criteria emphasized peer-reviewed journal articles and high-quality conference proceedings that reported empirical findings, methodologies, or reviews focusing on the effects of crop rotation and no-tillage systems. Eligible studies covered themes such as soil health, carbon sequestration, biodiversity, crop productivity, and system resilience under different environmental or management conditions. Reference lists of selected articles were also screened (snowball sampling) to identify additional relevant publications. This rigorous search yielded an initial pool of approximately 1757 studies, from which over 120 met the established criteria and were retained for detailed evaluation.
The selected literature spans a wide array of geographical contexts, climatic zones, and cropping systems, ranging from temperate cereal-based systems to tropical legume-rotation practices. We systematically extracted information related to the experimental or observational design, the nature and duration of the rotation or tillage treatments, measured outcomes such as soil organic carbon, microbial biomass, nutrient cycling, crop yield, and methodological approaches including field trials, meta-analyses, and remote sensing assessments. Additionally, we noted any studies that linked agronomic outcomes to broader policy frameworks, such as climate-smart agriculture or carbon neutrality goals.
In synthesizing the findings, we organized the literature according to several core dimensions: types of agronomic practices (rotation schemes, tillage intensity), biophysical responses (e.g., soil structure, erosion control, GHG emissions), productivity trade-offs, and environmental sustainability indicators. Where applicable, we identified areas of consensus—such as the generally positive impacts of diversified rotations on soil aggregation—and flagged ongoing debates, including the performance variability of no-tillage under different soil moisture regimes or pest pressures. This allowed us to draw attention to both robust patterns and knowledge gaps across temporal and spatial scales.
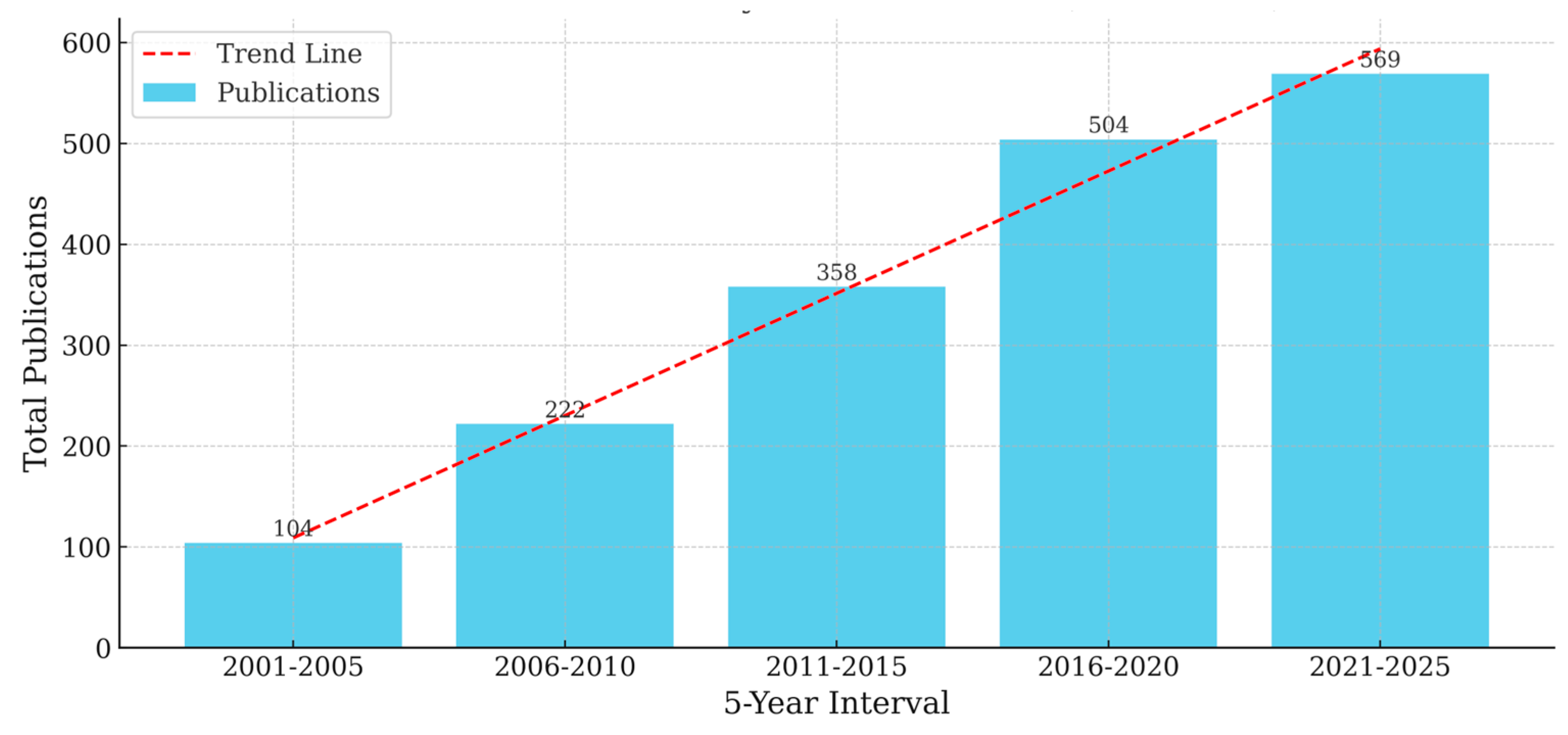
Temporal distribution of publication counts in five-year intervals (2001–2025), showing an increasing research trend with a fitted linear regression (Figure 2).
Through this structured and transparent approach, our review captures the evolution and diversity of research on crop rotation and no-tillage systems over the past two decades. It also reflects the growing relevance of these practices in global discussions around sustainable land use, food security, and climate mitigation. Figure 2 illustrates the upward trajectory of scholarly publications in this domain from 2001 to 2025, signaling increased academic engagement driven by advances in agroecological research and policy interest in regenerative farming strategies.
3. Ecological Benefits and Mechanisms of Crop Rotation
Crop rotation is a foundational practice that enhances nutrient cycling, yield stability, and resilience to climate stress. Diversifying crops in sequence improves the availability and balance of nutrients in soil (Figure 3). For instance, long-term field studies across North America show that including different crop functional types (e.g. legumes or forages) in rotation can significantly boost grain yields while reducing reliance on synthetic fertilizers[24,25]. Rotational diversity disrupts pest and disease life cycles, lowering pathogen carryover and weed pressure. A recent study in the U.S. Corn Belt found that even a simple two-year corn–soybean rotation increases corn and soybean yields by ~1.0 and 0.2 t/ha, respectively, compared to continuous monoculture [26]. Notably, the yield benefits of rotation become most pronounced under adverse weather conditions like drought. This suggests rotation is an important strategy for climate change adaptation, buffering crops against climate extremes[26]. In addition, introducing legumes in rotations enhances biological nitrogen fixation, reducing fertilizer needs, while deep-rooted crops mobilize phosphorus and potassium from deep soil layers. Crop rotation also fosters a more diverse soil food web and disease-suppressive microbiome [27]. For example, rotating peanuts with non-host crops enriched beneficial rhizosphere microbes that suppressed peanut root rot, whereas continuous monocropping led to depletion of those protective microbes[27]. In summary, crop rotations act as ecological regulators—improving nutrient cycling, breaking pest and disease cycles, and increasing yield stability — all of which contribute to enhanced soil health and farm productivity[25].
3.1. Effects on Nutrient Cycling
Crop rotation profoundly influences soil nutrient dynamics, enhancing both nutrient availability and overall soil fertility. Rotational diversity affects the acquisition, retention, and cycling of key macronutrients—particularly nitrogen (N), phosphorus (P), and potassium (K)—by modifying plant-soil-microbe interactions and optimizing resource use efficiency. To enhance our understanding of the role of crop rotation in soil health, recent studies have provided robust quantitative evidence. For instance, a recent meta-analysis demonstrated that diversified crop rotations significantly improved soil physical health indicators, reduced bulk density, and enhanced aggregate stability, thereby optimizing water infiltration and retention capacity[28]. In the North China Plain, a six-year field trial found that incorporating legumes and economic crops in rotation increased grain-equivalent yield by up to 38% while reducing nitrogen fertilizer usage and nitrous oxide emissions by 39%, highlighting a win–win scenario for productivity and environmental sustainability [12]. Similarly, multi-year experiments on the Tibetan Plateau showed that rotating barley with wheat or rapeseed increased barley yields by 17% and 12%, respectively, and improved soil quality indices (including soil organic matter and microbial biomass nitrogen) by 11–21% [29]. These findings further support the concept that crop rotation enhances yield stability and productivity by improving soil structure, biodiversity, and nutrient cycling. Notably, the positive effects of rotation on soil microbial communities have also been confirmed: rotations significantly increased the abundance of beneficial functional microbes such as nitrogen fixers while suppressing the accumulation of certain pathogens [27]. These results reinforce previous statements in the article about how crop rotation breaks pest cycles and promotes nutrient cycling, providing expanded support across diverse agroecological contexts.
Biological Nitrogen Fixation is one of the most significant contributions of crop rotation systems that include legumes. Leguminous crops such as soybean, chickpea, and clover harbor symbiotic rhizobia bacteria within root nodules, enabling the fixation of atmospheric nitrogen into plant-available forms. Recent meta-analyses have demonstrated that legume-inclusive rotations can reduce the requirement for synthetic nitrogen fertilizers by 30–60% without sacrificing yields, thereby offering both economic and environmental benefits [30]. For instance, a six-year rotation study in the North China Plain showed that peanut–wheat–maize rotations could reduce synthetic N inputs by 41% while maintaining comparable yields to conventional monocultures [31]. Moreover, biological nitrogen fixed during legume phases not only benefits the legumes themselves but also enhances soil residual N for subsequent cereal crops, a phenomenon known as the “N credit effect” [32].
Phosphorus Mobilization is another critical benefit of diversified rotations. Different crop species exude distinct profiles of organic acids, enzymes, and other exudates that mobilize sparingly soluble P from soil minerals. Cover crops such as lupins, for example, release citric acid that solubilizes phosphate, improving P availability for subsequent crops[33]. Furthermore, root turnover and microbial decomposition of diverse crop residues facilitate phosphorus recycling in the soil-plant system, leading to more sustainable P dynamics over time.
Potassium Redistribution and Uptake is also influenced by crop rotation. Deep-rooted crops like oilseed radish and sunflowers can scavenge potassium from subsoil layers inaccessible to shallow-rooted species, redistributing nutrients vertically within the soil profile [34,35]. Rotations that include such species help mitigate nutrient stratification and enhance long-term potassium use efficiency. Nutrient Balance Restoration is perhaps one of the most holistic benefits of rotation. In monoculture systems, certain nutrients become severely depleted while others accumulate excessively. Crop rotation balances nutrient extraction patterns by alternating between high-demand and low-demand crops, maintaining soil nutrient homeostasis [36,37]. For example, alternating between cereals and legumes moderates nitrogen dynamics, while including root or tuber crops can diversify potassium and micronutrient demand patterns.
In summary, crop rotations act as ecological nutrient management strategies, enhancing natural nutrient acquisition processes, reducing dependence on chemical fertilizers, and contributing to the long-term sustainability of agricultural soils.
3.2. Mechanisms of Pest and Disease Reduction
One of the most profound ecological benefits of crop rotation lies in its ability to suppress pests, diseases, and weeds through both direct and indirect mechanisms. Unlike monocultures, where pathogens and herbivorous pests find continuous hosts season after season, diversified rotations disrupt their life cycles, diminish inoculum sources, and enhance soil and plant health, thus reducing the reliance on chemical controls.
3.2.1. Host Disruption and Inoculum Dilution
Continuous cropping of the same species provides a stable, predictable environment for specialized pests and pathogens. Crop rotation disrupts this cycle by introducing non-host species that hinder pest survival, reproduction, and spread. For example, the corn rootworm, one of the most damaging maize pests in North America, is effectively managed by rotating maize with soybean, thereby starving rootworm larvae of suitable roots[38]. Studies have shown that even a single year of soybean in rotation can reduce rootworm larval densities by over 90% compared to continuous maize systems [39].
Similarly, soil-borne diseases such as Fusarium wilt, Verticillium wilt, and Rhizoctonia damping-off are greatly reduced under diverse rotations. Pathogen populations decline in the absence of their preferred hosts, lowering soil inoculum levels over time. A six-year study in rice–wheat systems demonstrated that rotating rice with leguminous crops significantly reduced the incidence of Fusarium and Pythium-induced root rot in wheat[40].
3.2.2. Induction of Disease-Suppressive Soils
Beyond mere host absence, rotations actively foster beneficial soil microbial communities that can antagonize pathogens—a phenomenon known as disease-suppressive soils. Diverse rotations promote microbial diversity, functional redundancy, and the establishment of antagonistic bacteria and fungi such as Pseudomonas, Bacillus, and Trichoderma species[41]. These beneficial microbes outcompete or inhibit pathogens through mechanisms like antibiotic production, resource competition, parasitism, and induced systemic resistance (ISR) in plants [42].
Recent metagenomic studies revealed that rotated soils harbor a higher abundance of genes related to antifungal compounds (e.g., phenazines, DAPG) compared to monocultures, indicating a microbiome-mediated defense enhancement[43]. Furthermore, disease-suppressive capacity was found to increase with the complexity and temporal heterogeneity of crop rotations, suggesting that longer and more diversified rotations build stronger biotic resistance.
3.2.3. Weed Pressure Management
Crop rotation also indirectly suppresses weed communities by altering disturbance regimes, resource availability, and competitive hierarchies. Continuous monocultures create selection pressure favoring a narrow group of weed species adapted to specific cropping calendars and practices. In contrast, diversified rotations introduce variability in planting dates, canopy structures, and soil cover, thereby preventing any single weed species from dominating [15,44].
For instance, rotating cereals with broadleaf crops disrupts the life cycle of grass weeds like Avena fatua (wild oat), while including cover crops can smother winter annuals such as Stellaria media (chickweed) [45]. Moreover, diversified rotations reduce the selective pressure that drives herbicide resistance evolution, an increasingly urgent concern in intensive agricultural systems.
3.2.4. Enhancement of Plant Vigor and Stress Tolerance
Healthier soils resulting from crop rotation practices often translate into more vigorous plant growth, which itself can confer partial resistance to pests and diseases. Plants growing in nutrient-rich, biologically active soils are better able to withstand infections or herbivory. Additionally, rotation-induced changes in soil microbiomes can prime plant immune responses, enhancing resilience against a broad range of biotic stresses[46].
A recent experiment demonstrated that maize following soybean had significantly higher expression of defense-related genes compared to continuous maize, suggesting microbiome-mediated induced resistance triggered by the previous legume phase [47].
3.2.5. Limitations and Adaptations of Pests
It must be noted that some pests, like the “rotation-resistant” variant of the western corn rootworm, have evolved to overcome traditional rotational strategies by adapting their egg-laying behavior[48]. Therefore, relying solely on simple two-crop rotations may not be sufficient over the long term. Integrating rotations with additional strategies such as cover cropping, diverse species mixes, and occasional disruptive tillage can help counteract adaptive pest responses[49].
3.3. Impacts on Soil Structure and Microbial Communities
Crop rotation exerts profound and multifaceted effects on soil physical structure and microbial community composition, both of which are critical determinants of soil health, nutrient cycling, and long-term agricultural sustainability. By varying root architectures, residue inputs, and rhizosphere interactions over time, diversified rotations help maintain or enhance the biological and physical integrity of soils.
3.3.1. Improvements in Soil Aggregation and Physical Properties
Soil structure, particularly the formation and stability of aggregates, is crucial for water retention, gas exchange, and resistance to erosion. Root systems of different crops contribute distinctively to soil aggregation: fibrous-rooted species like cereals produce fine root networks that entangle soil particles, while tap-rooted species such as legumes and oilseeds penetrate deeper soil layers, promoting vertical microporosity[50].
Multiple studies have demonstrated that diversified rotations significantly improve aggregate stability compared to continuous monocultures. For example, a 12-year field experiment showed that rotations involving maize, soybean, and wheat increased water-stable macroaggregates by 25–40% relative to continuous maize, primarily due to increased root biomass and diversified residue chemistry[51]. Enhanced aggregation not only improves soil porosity but also protects soil organic matter (SOM) by physically occluding it within stable aggregates, thereby promoting long-term carbon sequestration.
Management practices—particularly the incorporation of grain legumes and the selection of tillage systems—modulated the effects of crop diversification on soil physical health, with notable variation across different subgroups. Specifically, the inclusion of grain legumes, relative to cereal-only rotations, led to a significant reduction in soil bulk density (−1.7%; 95% CI: −3.1% to −0.2%), a marked increase in aggregate stability (20.8%; 95% CI: 6.3% to 35.3%), and improvements in both soil porosity (4.1%; 95% CI: 1.7% to 6.5%) and saturated hydraulic conductivity (119.5%; 95% CI: 52.1% to 186.9%). However, no statistically significant enhancement was observed in infiltration rates (120.0%; 95% CI: −7.3% to 247.2%)[28].
Additionally, rotations improve soil bulk density and infiltration rates. Systems incorporating diverse crops were shown to reduce surface compaction and enhance rainfall infiltration capacity, reducing runoff and erosion risks under both temperate and tropical conditions.
3.3.2. Enhancements in Soil Organic Matter Dynamics
Crop rotation is a key lever for building and maintaining soil organic matter stocks. Different crops contribute variable quantities and qualities of organic residues—ranging from C:N ratios to lignin contents—that influence decomposition rates and SOM stabilization [52]. Including high-biomass-producing or deep-rooting species in rotations increases organic matter inputs into both surface and subsoil layers.
Long-term trials have indicated that crop rotations can increase soil organic carbon (SOC) levels by 10–20% compared to monoculture systems over a decade[53]. Furthermore, diverse rotations lead to more gradual and continuous residue input patterns, promoting a more resilient and active soil food web. This continuous carbon flow is vital for supporting microbial communities and sustaining nutrient cycling.
3.3.3. Impacts on Soil Microbial Diversity and Functionality
The soil microbiome responds sensitively to crop rotational diversity. Each plant species shapes its rhizosphere microbiome through unique patterns of root exudates, secondary metabolites, and residue chemistry[54]. Therefore, changing crops seasonally introduces temporal heterogeneity into the soil environment, fostering higher microbial diversity and preventing dominance by pathogenic or opportunistic organisms. Recent high-throughput sequencing studies revealed that crop rotations consistently increase microbial alpha diversity (species richness within a sample) and beta diversity (differences between communities) relative to continuous monocultures[55]. Moreover, rotations favor beneficial microbial groups, such as nitrogen-fixing bacteria (Bradyrhizobium, Azospirillum), phosphate-solubilizing bacteria (Pseudomonas spp.), and mycorrhizal fungi (Glomeromycota), which enhance nutrient availability and plant growth[56].
Functional analyses suggest that crop rotations promote microbial genes associated with carbon degradation, nitrogen fixation, and disease suppression[57,58]. Notably, metagenomic comparisons between continuous wheat and wheat–legume rotations demonstrated higher abundances of genes involved in denitrification, cellulose degradation, and biocontrol traits in the rotated systems[59,60].
3.3.4. Disease Suppressiveness and Soil Resilience
Higher microbial diversity associated with crop rotations contributes to the natural suppressiveness of soils against diseases and pests. Complex microbial networks provide functional redundancy, ensuring that critical ecosystem services such as pathogen antagonism, nutrient cycling, and organic matter turnover are maintained under stress conditions [61].
Experimental evidence indicates that soils under diversified rotations are more resilient to drought, salinity, and pathogen invasion compared to those under monoculture [62]. For instance, rotating soybean with maize or wheat reduced the relative abundance of soil-borne Fusarium spp. and increased populations of Bacillus and Streptomyces, genera known for producing antifungal compounds[63,64].
3.3.5. Integration with Soil Health Indicators
Due to their broad effects on physical and biological properties, crop rotations are increasingly used as benchmarks for soil health assessment frameworks. Indicators such as microbial biomass carbon, soil respiration rates, extracellular enzyme activities (e.g., β-glucosidase, urease), and aggregate stability often score higher under diversified rotations compared to monocultures [65].
Thus, integrating crop rotation practices is not merely a yield-enhancement strategy but a foundational pillar for regenerating soil health, enhancing ecosystem multifunctionality, and building resilience against future environmental stresses.
3.4. Case Studies of Typical Rotation Systems
To translate the ecological theories of crop rotation into practice, it is essential to examine real-world examples across major cropping systems. Case studies of widely implemented rotation sequences—such as soybean–corn and rice–wheat systems—offer valuable insights into the agronomic, ecological, and economic outcomes associated with rotational diversity.
3.4.1. Soybean–Corn Rotation Systems
The soybean–corn rotation is one of the most common cropping systems in North America, covering millions of hectares across the Midwest and beyond. This simple two-crop rotation has demonstrated substantial benefits over continuous monoculture of either crop.
Nutrient dynamics are significantly improved through the inclusion of soybean, a leguminous crop capable of biological nitrogen fixation. Studies show that the nitrogen contribution from soybean residues can partially satisfy the nitrogen requirements of subsequent corn crops, reducing fertilizer needs by 30–50%[32]. Moreover, this rotation improves soil organic carbon accumulation, as alternating C3 (soybean) and C4 (corn) plant residues contribute different chemical compositions to the soil carbon pool [66].
Pest and disease suppression is another major advantage. Rotating corn with soybean effectively disrupts the lifecycle of the corn rootworm, reducing larval populations and associated root damage[38]. However, the emergence of rotation-resistant rootworm populations in some regions has prompted more complex rotations or additional integrated pest management strategies[49].
Yield stability and resilience are also enhanced in the soybean–corn system. Research indicates that this rotation improves corn yields by 5–15% compared to continuous corn, largely due to reduced disease pressure and better soil health parameters[67]. Moreover, rotated systems exhibit greater resilience under drought conditions because of improved soil water-holding capacities associated with diversified root systems and organic matter inputs.
3.4.2. Rice–Wheat Rotation Systems
In South and East Asia, the rice–wheat rotation dominates major agricultural landscapes, particularly in the Indo-Gangetic Plains. Despite its importance for regional food security, the intensive nature of this system has raised concerns about sustainability.
Soil health challenges such as declining organic matter, micronutrient deficiencies (e.g., zinc, iron), and soil compaction are prevalent in continuous rice–wheat systems [68]. However, strategic diversification within the rice–wheat framework—such as integrating legumes (e.g., mungbean, chickpea) or oilseeds (e.g., mustard) into the rotation—can restore soil fertility and structure[69].
Weed dynamics are a critical issue, particularly the proliferation of Phalaris minor (little seed canary grass) in wheat fields. Diversifying rotations or introducing short-duration legumes between rice and wheat has been shown to disrupt Phalaris seedbanks and reduce herbicide resistance evolution [70].
Carbon sequestration and greenhouse gas mitigation are emerging focuses. Traditional flooded rice paddies contribute significantly to methane emissions. Introducing aerobic crops into the system (e.g., upland rice varieties, legumes) or adopting conservation agriculture practices within rice–wheat rotations can significantly lower greenhouse gas footprints while maintaining productivity[71].
Yield advantages have been recorded where rice–wheat rotations are diversified or adjusted for local climatic conditions. Rotations incorporating legumes improved wheat yields by 10–20% compared to rice–wheat monocultures, attributed to better nitrogen supply, soil health, and reduced pest pressures[72].
3.4.3. Other Notable Rotational Systems
Other well-studied systems also illustrate the power of rotation:
- Barley–canola rotations in Australia have been associated with improved soil nitrogen dynamics and reduced blackleg disease in canola[73].
- Sugar beet–wheat–maize rotations in Europe enhance soil microbial activity and prevent disease buildup in sugar beet [74].
- Cotton–wheat–mungbean rotations in South Asia have improved water productivity, nitrogen use efficiency, and smallholder profitability[75].
Each of these systems highlights how region-specific adaptations of rotation principles can generate profound agronomic and ecological benefits.
3.4.4. Lessons Learned and Future Directions
While simple two-crop rotations like soybean–corn or rice–wheat have clear advantages over monoculture, growing evidence suggests that increasing rotation complexity—adding a third or fourth species, integrating cover crops, or alternating functional crop groups—delivers even greater benefits. Longer and more diversified rotations tend to support higher soil microbial diversity, better nutrient cycling, and greater resilience to climatic extremes [76]. Precision agriculture technologies, such as remote sensing and soil health diagnostics, now allow for dynamic, site-specific rotation planning that maximizes benefits while accounting for spatial heterogeneity in soil and climatic conditions.
Future research and policy efforts should therefore encourage not only the practice of rotation itself but also its intensification and smart optimization for sustainability and resilience.
4. The Central Role of Soil Microbiomes in Farmland Management
No-tillage (NT) agriculture provides significant benefits for soil structure, carbon sequestration, and erosion control. Eliminating plowing allows soils to develop stable aggregates and protective surface cover. A global meta-analysis of 89 studies confirmed that no-till consistently increases the formation of larger soil aggregates and raises soil organic carbon within those aggregates, relative to conventional tillage [77]. Pore continuity and mean aggregate size under no-till were found to improve by ~20%, indicating better soil structure and permeability. These physical changes reduce soil compaction and runoff, thereby decreasing erosion. Importantly, no-till farming can also contribute to carbon sequestration in soil, though results vary with context. A recent meta-analysis showed that no-till alone yielded modest SOC gains (+4–5%), but when combined with residue retention (e.g. leaving crop straw on fields), SOC increases were greater. In China, integrating no-till with full straw return led to measurable SOC increases over time [78]. No-till’s climate benefits extend beyond carbon – it also significantly cuts fuel use and can lower soil nitrous oxide emissions by ~12–14% through improved soil structure and reduced microbial denitrification hotspots. Over the long term, NT farming supports a richer soil biota (including earthworms and mycorrhizae) by minimizing soil disturbance. This biological boost further reinforces aggregation and nutrient cycling. Studies have observed that combining no-till with organic amendments amplifies these advantages: for example, adding compost in a no-till system increased aggregate stability and accumulation of microbial residues (glomalin), leading to more resilient soil structure [79]. While no-till may sometimes initially reduce yields or require adaptations in weed management, its overall benefits for soil physical health and carbon retention are well-documented. When implemented with complementary practices (residue management, cover crops), no-tillage is a cornerstone of sustainable soil management that builds soil organic matter and mitigates climate change.
No-tillage agriculture enhances soil structure, stabilizes aggregates, and promotes microbial diversity through residue retention and minimal soil disturbance, thereby supporting carbon sequestration and reducing erosion. In contrast, conventional tillage disrupts aggregates, increases leaching and erosion, and elevates fuel costs, leading to greater soil degradation. These contrasting mechanisms highlight the central role of tillage practices in shaping soil microbiome function and sustainable farmland management (Figure 4).
4.1. Microbial Contributions to Nutrient Transformations (N, P, K)
Soil microbiomes serve as critical regulators of nutrient transformations that underpin plant productivity. Microbial-driven cycling of nitrogen (N), phosphorus (P), and potassium (K) sustains soil fertility, reduces reliance on synthetic fertilizers, and enhances agroecosystem resilience.
4.1.1. Nitrogen Transformations
Biological nitrogen fixation (BNF) remains the most direct microbial contribution to farmland nitrogen pools. Symbiotic rhizobia in leguminous crops fix atmospheric N₂ into ammonium (NH₄⁺), providing a sustainable nitrogen source [30]. Free-living diazotrophs such as Azospirillum, Klebsiella, and Paenibacillus further supplement soil nitrogen, particularly under low-fertility conditions[80]. Beyond fixation, nitrifying bacteria (e.g., Nitrosomonas spp.) and archaea (e.g., Nitrosopumilus spp.) oxidize NH₄⁺ into nitrate (NO₃⁻), a form readily absorbed by plants but prone to leaching losses [81]. Denitrifiers, in turn, reduce NO₃⁻ to gaseous N₂O and N₂, regulating nitrogen retention versus loss pathways.
Recent metagenomic studies reveal that crop rotation intensifies the abundance of nifH genes associated with nitrogen fixation while tempering nitrifier populations, suggesting that management practices modulate nitrogen-cycling communities [82].
4.1.2. Phosphorus Solubilization
Microbial mediation of phosphorus availability centers on two processes: solubilization of inorganic phosphates and mineralization of organic P. Phosphate-solubilizing bacteria (PSB), such as Pseudomonas fluorescens and Bacillus megaterium, excrete organic acids (e.g., gluconic acid, citric acid) that release P from insoluble mineral complexes[83]. Simultaneously, phosphatase-producing microbes degrade organic P compounds into bioavailable forms.
Field experiments demonstrate that inoculation with PSB can enhance soil-available P by 20–50%, depending on soil texture and parent material [84]. Moreover, microbial community diversity strongly correlates with P turnover rates, emphasizing the need to preserve belowground biodiversity.
4.1.3. Potassium Mobilization
While traditionally viewed as geochemically driven, potassium (K) cycling also involves biological components. Potassium-solubilizing microorganisms (KSMs), such as Bacillus mucilaginosus and Frateuria aurantia, release chelating substances and organic acids that liberate K from feldspar, mica, and illite minerals [85]. Recent findings suggest that KSM activity can augment potassium availability by 15–30% in depleted soils [86].
Furthermore, endophytic bacteria associated with deep-rooted crops have been implicated in facilitating subsoil K uptake, linking root architecture with microbial K mobilization[87].
In sum, soil microbiomes act as dynamic engineers of nutrient cycling, coordinating transformations across N, P, and K pools. Harnessing this functional potential through microbiome management is vital for the sustainable intensification of agriculture.
4.2. Relationships between Microbiomes and Crop Stress Resistance
The soil microbiome acts as a pivotal mediator of plant resilience to abiotic and biotic stresses. Through intricate interactions in the rhizosphere, beneficial microbes enhance plant tolerance to drought, salinity, pathogens, and nutrient deficiencies, thereby safeguarding yields under increasingly variable environmental conditions.
4.2.1. Microbial Enhancement of Abiotic Stress Tolerance
Certain microbial taxa, particularly plant growth-promoting rhizobacteria (PGPRs) such as Azospirillum, Bacillus, and Pseudomonas spp., induce systemic tolerance mechanisms in crops. These microbes modulate phytohormone pathways—such as upregulating abscisic acid (ABA) during drought or enhancing ethylene signaling under salinity stress—thereby priming plant physiological defenses[88].
Inoculation experiments demonstrate that Bacillus subtilis strains can increase wheat biomass under water-limited conditions by enhancing root hydraulic conductivity and antioxidant enzyme activity[89]. Similarly, Pseudomonas putida promotes osmotic balance and maintains ion homeostasis in saline soils by stimulating proline accumulation in host plants[90].
4.2.2. Microbiome-Mediated Disease Suppression
The microbial community structure profoundly influences plant susceptibility to soilborne pathogens. Beneficial microbes compete with, inhibit, or induce plant resistance against invaders such as Fusarium spp., Rhizoctonia spp., and nematodes [41]. For instance, communities rich in Streptomyces and Trichoderma species correlate with natural suppression of root rot and damping-off diseases [91].
Moreover, diversified soil microbiomes enhance “community resilience,” ensuring that even after pathogen outbreaks, the microbial assemblage can rapidly recover and re-establish disease suppression functions [42].
4.2.3. Holobiont Perspective
Emerging frameworks view the plant and its associated microbiome as a single ecological unit—or “holobiont”—with co-evolved strategies for stress resistance [92]. Recognizing crops as holobionts shifts management goals toward fostering beneficial plant-microbe partnerships rather than solely breeding stress-resistant genotypes.
Ultimately, soil microbiomes represent a living reservoir of resilience traits, and manipulating these communities offers a powerful complement to conventional plant breeding and agrochemical approaches.
4.3. Regulation of Soil Microbial Communities by Agricultural Practices
Agricultural management practices exert a decisive influence on the structure, function, and resilience of soil microbial communities. Through alterations in disturbance regimes, resource availability, and habitat conditions, farming decisions shape the trajectory of microbiome assembly and its ecological roles.
4.3.1. Tillage and Soil Microbiomes
Conventional tillage disrupts soil physical structure, fragmenting microbial habitats and reducing community stability. Comparative studies have shown that no-tillage systems consistently support higher microbial biomass, greater fungal-to-bacterial ratios, and enhanced functional diversity relative to tilled soils[93]. Reduced mechanical disturbance allows the establishment of spatially structured biofilms and promotes the accumulation of beneficial symbionts such as mycorrhizal fungi [94].
However, the response of microbial communities to tillage is context-dependent, varying with soil type, climate, and crop rotation schemes. Transitional periods following a switch to conservation tillage may initially decrease microbial diversity due to abrupt habitat shifts[95].
4.3.2. Organic Amendments and Fertilization
Organic amendments—such as manure, compost, and green manures—enrich microbial activity by providing complex carbon substrates and micronutrients. Application of organic matter not only boosts microbial abundance but also enhances the prevalence of taxa associated with nutrient cycling and disease suppression[96]. For instance, amendment with compost increased the abundance of phosphate-solubilizing Pseudomonas spp. and nitrogen-fixing Rhizobium spp. in maize fields[97].
Conversely, long-term exclusive reliance on synthetic fertilizers, particularly high doses of nitrogen, tends to reduce microbial richness and select for copiotrophic, fast-growing taxa, potentially destabilizing nutrient cycling processes[98].
4.3.3. Crop Rotation and Diversification
Rotation practices, especially those involving functional diversity (e.g., cereals-legumes-oilseeds), create temporal heterogeneity in rhizodeposition patterns, fostering dynamic and diverse microbiomes[99]. Studies indicate that complex rotations (three or more crops) enhance soil microbial evenness and promote keystone taxa critical for nitrogen cycling and disease resistance [100].
In contrast, continuous monoculture often leads to microbiome homogenization and the buildup of pathogenic organisms, increasing vulnerability to disease outbreaks and yield instability.
4.3.4. Pesticide and Herbicide Applications
Chemical inputs, while critical for pest management, have unintended consequences for microbial communities. Some broad-spectrum herbicides, such as glyphosate, can inhibit beneficial soil bacteria and alter community composition, although effects vary based on dosage and soil properties[101]. Integrated pest management strategies that minimize chemical inputs while maintaining microbial habitat integrity are thus pivotal for sustainable farmland management.
4.3.5. Towards Microbiome-Conscious Farming
Emerging “microbiome engineering” frameworks advocate deliberate agricultural interventions—such as targeted inoculations, selective fertilization, and minimal disturbance—to steer microbiomes toward desired functional outcomes [102]. Recognizing soil microbiomes as manageable assets rather than passive components opens new frontiers for resilient, low-input agriculture.
No-tillage (NT) agriculture provides significant benefits for soil structure and carbon retention. Recent frontier studies further emphasize the central role of soil microbiomes in enhancing the resilience of farming systems. For example, researchers propose that manipulating plant root exudates and microbial communities could facilitate the breeding of "climate-smart" crops [103]. This approach optimizes plant–microbiome interactions to improve nutrient use efficiency and stress tolerance without increasing external inputs, while also accelerating soil carbon sequestration[103]. Additionally, machine learning analyses reveal that nutrient source management (organic vs. synthetic fertilizers) exerts a more profound effect on soil microbial community structure and interaction networks than tillage or cover crops [104]. The application of organic amendments (e.g., farmyard manure) fosters fungal-dominated microbial networks in the 0–30 cm soil layer, enhancing community stability. In contrast, deep plowing disrupts microbial interactions in the 0–20 cm layer, while the effect of cover crops is relatively minor [104]. These findings align with earlier statements about how reduced soil disturbance and organic inputs support microbial richness and emphasize the importance of organic fertilization from a microbiome perspective. Furthermore, long-term field trials demonstrate that intensified crop rotation and organic fertilization increase the abundance of nitrogen fixation-related genes and suppress nitrifier populations, leading to more efficient nitrogen cycling [105].
4.4. Application Potentials of Functional Microbial Groups (PGPRs, Biocontrol Agents)
Harnessing functional microbial groups represents a frontier strategy for sustainable agriculture. Plant Growth-Promoting Rhizobacteria (PGPRs) and biocontrol agents offer eco-friendly alternatives to synthetic fertilizers and pesticides, enhancing crop performance while mitigating environmental impacts.
Plant Growth-Promoting Rhizobacteria (PGPRs)
PGPRs such as Azospirillum, Bacillus, Pseudomonas, and Rhizobium species stimulate plant growth through multiple pathways: nitrogen fixation, phosphate solubilization, phytohormone modulation, and siderophore production[106]. Meta-analyses reveal that PGPR inoculation can increase crop yields by an average of 20–30%, with greater effects under abiotic stress conditions[107]. Recent formulations combining multiple PGPR strains exhibit synergistic effects, promoting not only growth but also enhancing systemic resistance against pathogens and environmental stresses[108].
Biocontrol Agents Against Pests and Diseases
Biocontrol agents—such as Trichoderma, Bacillus subtilis, and Streptomyces spp.—suppress soilborne pathogens through antibiosis, mycoparasitism, competition, and induction of plant immune responses[108]. Field applications of Trichoderma harzianum have demonstrated consistent reductions in Fusarium wilt incidence across diverse cropping systems[109].
Furthermore, next-generation biocontrol strategies increasingly exploit microbiome engineering approaches, introducing beneficial consortia rather than single strains to stabilize pathogen-suppressive functions[110].
Challenges and Prospects
Despite promising outcomes, variability in field efficacy remains a major hurdle, driven by soil heterogeneity, native microbiome competition, and environmental factors[111]. Precision agriculture technologies—such as microbiome diagnostics and targeted delivery systems—hold potential to tailor microbial applications to site-specific conditions. Ultimately, integrating functional microbial groups into holistic soil health frameworks offers a path toward resilient, productive, and environmentally sound farmland systems.
5. Mechanisms of Organic Amendments (Farmyard Manure) Effects
Applying organic amendments (such as farmyard manure, compost, or crop residues) is a proven strategy to replenish soil organic matter and improve a range of soil properties. Organic inputs directly add carbon and nutrients to soil, stimulating soil biological activity and aggregation. Global syntheses indicate that long-term organic amendment can increase topsoil organic carbon by ~24% on average. This contributes not only to soil fertility and crop yields, but also to carbon sequestration as a climate mitigation measure. In fact, rebuilding depleted soil carbon stocks through practices like manure application is estimated to account for roughly 60% of the agriculture sector’s potential in natural climate solutions [112]. Beyond carbon, organic amendments greatly benefit the soil microbiome and nutrient cycling. A 2023 meta-analysis found that adding organic amendments consistently increased soil bacterial diversity (richness and Shannon index) while also enhancing soil enzyme activities for nitrogen and phosphorus cycling [113]. By providing fresh substrates, amendments shift microbial communities toward copiotrophic (fast-growing) organisms that mineralize nutrients, thus improving nutrient availability to plants. These inputs also supply organic acids and humic compounds that can improve soil structure. Research shows that regular manure or compost applications lead to a higher proportion of water-stable macroaggregates in soil. In one long-term study, manure-amended plots had 2–3 times more stable macroaggregates than plots receiving only mineral fertilizer. Such improvements in aggregation are often mediated by increased fungal hyphae and microbial polysaccharides. For example, experiments in Central Europe demonstrated that compost and digestate applications raised aggregate stability, accompanied by greater abundance of arbuscular mycorrhizal fungi and their glomalin production[79]. The net effect of organic amendments is a positive feedback loop: higher soil organic matter supports a thriving microbiome, which in turn builds soil structure and nutrient reserves. Over time, soils under organic amendment management exhibit improved porosity, water retention, and buffering capacity against acidification. Crop productivity benefits as well – a recent analysis noted an average 27% increase in yields with organic amendments compared to mineral fertilizers alone, owing to improvements in soil health[114]. In summary, organic amendments are essential for restoring degraded soils: they increase soil organic carbon, nurture beneficial microbial communities, enhance soil structure, and thus drive the long-term productivity and sustainability of agroecosystems.
Organic amendments such as farmyard manure enhance soil health through the addition of carbon and nutrients, stimulation of microbial activity, and promotion of water-stable macroaggregates. These processes improve soil structure (up to 24% increase in topsoil stability), enhance microbial-mediated nutrient cycling, and ultimately boost crop productivity by approximately 27%. Fungal hyphae and microbial polysaccharides play a key role in the stabilization of soil aggregates, supporting long-term soil fertility and ecosystem resilience (Figure 5).
5.1. Improvements in Soil Physical and Chemical Properties
Farmyard manure (FYM) fundamentally enhances soil health by improving physical structure and chemical fertility. Organic inputs increase soil organic matter (SOM), promoting aggregation and reducing bulk density, which in turn enhances porosity and water retention[115]. Long-term application of FYM has been shown to significantly raise cation exchange capacity (CEC), pH buffering, and the retention of macro- and micronutrients[116]. Moreover, manure decomposes slowly, acting as a sustained-release nutrient source, buffering crops against nutrient fluctuations often observed under synthetic fertilizer regimes.
Figure 6.
Ten-year effects of contrasting organic fertilization strategies on crop yield and yield stability in Eastern Austria[117]. Copyright 2025 Elsevier.
Figure 6.
Ten-year effects of contrasting organic fertilization strategies on crop yield and yield stability in Eastern Austria[117]. Copyright 2025 Elsevier.
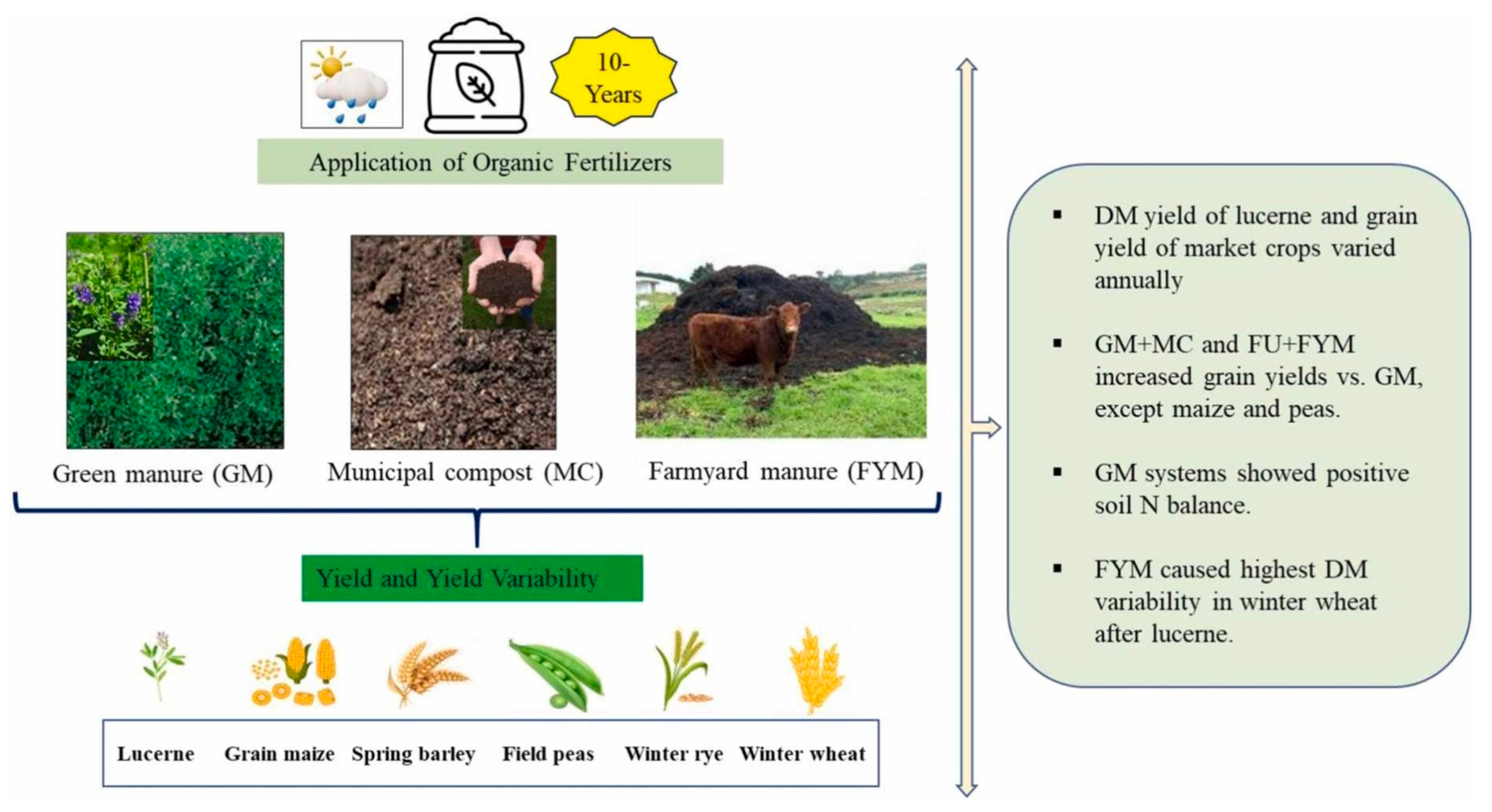
Notably, FYM contributes complex organic compounds—humic and fulvic acids—that enhance soil mineral stability and influence metal chelation processes [118]. These physicochemical improvements underpin better root growth environments and long-term soil productivity. Long-term organic fertilization systems impact crop productivity and sustainability (Figure 6) [117].
5.2. Enhancement of Microbial Activity and Diversity
Organic amendments serve as rich carbon and energy sources, stimulating microbial growth and functional diversity. Application of FYM consistently increases microbial biomass carbon, enzymatic activities (e.g., dehydrogenase, urease), and respiration rates across soil types[119]. Decadal datasets show farmyard manure doubled the share of water-stable macro-aggregates relative to mineral N [120]. High-throughput sequencing reveals that FYM-treated soils harbor more diverse bacterial and fungal communities, including beneficial groups such as nitrogen-fixers (Bradyrhizobium spp.) and phosphate-solubilizers (Pseudomonas spp.)[121]. FYM addition also promotes keystone taxa associated with carbon sequestration and disease suppression, enhancing ecosystem multifunctionality.
Such microbial shifts improve nutrient cycling efficiency, pathogen suppression, and the resilience of the soil microbiome under environmental stresses.
5.3. Synergistic Effects Between Organic Amendments and Chemical Fertilizers
Combining FYM with chemical fertilizers exploits the strengths of both inputs: the immediate nutrient supply from synthetic sources and the soil conditioning benefits of organic matter. Meta-analyses show that integrated nutrient management (INM) increases crop yields by 10–20% and improves nutrient use efficiency compared to chemical fertilization alone[122]. FYM buffers the acidifying effects of nitrogen fertilizers, stabilizes microbial communities under fertilization stress, and mitigates nitrate leaching and ammonia volatilization [123]. Moreover, synchronized nutrient release from FYM and mineral fertilizers ensures steady crop nutrition throughout critical growth stages. Thus, judicious integration of organic and inorganic sources optimizes soil health and sustains agricultural productivity.
5.4. Case Studies from Long-Term Field Trials (e.g., Chinese Mollisol Regions)
Long-term experiments in the Chinese Mollisol region, a key agricultural belt, provide compelling evidence for the benefits of FYM. The Gongzhuling and Hailun stations have monitored treatments for over 30 years, revealing that FYM application significantly enhanced SOC stocks (by 25–35%), microbial biomass, and aggregate stability compared to solely fertilized soils[124].
Furthermore, FYM improved nitrogen mineralization rates, reducing dependency on external nitrogen inputs while maintaining or increasing crop yields[125].
These findings underscore that sustained FYM application is crucial for restoring degraded soils, mitigating greenhouse gas emissions, and ensuring resilient agroecosystems under intensive management regimes.
6. No-Tillage Agriculture and Soil Health
6.1. Improvements in Soil Physical Properties
No-tillage (NT) systems fundamentally alter the soil’s physical environment by minimizing mechanical disturbance. Unlike conventional tillage, which disrupts soil aggregates and promotes erosion, NT maintains a stable surface structure, enhancing aggregate stability, porosity, and infiltration[8].
Long-term NT systems reduce surface crusting, improve water retention, and lower bulk density, promoting deeper root growth and drought resilience [126].
Meta-analyses indicate that NT can increase infiltration rates by 30–50% compared to plowed soils, particularly in dryland systems where water conservation is critical[127]. However, compaction may occur in certain soil types, necessitating periodic subsoiling or rotational practices to alleviate mechanical impedance.
6.2. Contributions to Soil Organic Matter Accumulation and Carbon Sequestration
No-tillage practices promote the buildup of soil organic matter (SOM) by reducing organic carbon mineralization and protecting residues on the soil surface. Surface residues shield soil from erosive forces, while undisturbed microbial habitats foster stable carbon pools[128].
Recent syntheses suggest that NT increases topsoil organic carbon (SOC) by 10–20% within two decades, particularly when combined with residue retention and diversified rotations [129]. Moreover, NT enhances the formation of particulate organic matter (POM) and mineral-associated organic matter (MAOM), two key fractions critical for long-term carbon stabilization[130]. Thus, NT agriculture plays a pivotal role in mitigating climate change through enhanced carbon sequestration, although benefits vary with climate, soil texture, and management intensity.
6.3. Impacts on Pest, Disease, and Weed Dynamics
Pest Insects
Residue retention under NT provides overwintering habitats for certain insect pests, including Diabrotica virgifera (corn rootworm) and Helicoverpa zea (bollworm). However, diversified rotations and biological control enhancement can offset these risks, emphasizing the need for integrated pest management (IPM) strategies.
Soil-borne Diseases
Undisturbed residues may harbor pathogens like Fusarium, Rhizoctonia, and Pythium species, potentially elevating soil-borne disease risks[131]. Yet, improved soil biological activity and competition from saprotrophic microbes under NT often suppress pathogen establishment over time[132].
Weed Community Shifts
Reduced soil disturbance shifts weed communities toward perennial and rhizomatous species, such as Convolvulus arvensis (bindweed) and Cirsium arvense (Canada thistle) [133]. Herbicide reliance may initially increase, but integrating cover crops and diversified rotations can mitigate weed pressure and herbicide resistance evolution.
Integrated Pest and Weed Management Strategies
Successful NT systems incorporate biological controls, rotation planning, and mechanical interventions (e.g., roller crimping cover crops) to manage pests and weeds sustainably[134]. Transitioning to NT thus demands a holistic ecological approach rather than simple input substitution.
6.4. No-Tillage and Soil Microbial Diversity and Community Shifts
No-tillage profoundly reshapes soil microbial communities by preserving habitat continuity and enhancing carbon substrates near the soil surface. Studies consistently report higher microbial biomass, greater fungal dominance, and increased microbial functional diversity under NT compared to conventional tillage [135].
Fungal-to-bacterial ratios, an indicator of soil food web complexity, tend to rise under NT, promoting efficient nutrient cycling and aggregate formation [136]. Furthermore, NT favors keystone taxa associated with disease suppression, nitrogen fixation, and carbon stabilization [137]. Recent metagenomic analyses show that NT enriches genes involved in nutrient transformation, antibiotic production, and stress resilience[138]. These microbial shifts not only enhance soil fertility but also build system-wide resilience against biotic and abiotic stressors.
This section discusses the effects of NT on soil structure and organic carbon. A recent global meta-analysis quantified NT’s impact on yield and carbon stocks, finding that NT slightly reduced upland crop yields (~5%) or maintained yield levels while significantly improving soil organic carbon content [139]. This study also noted that NT’s impact on yield varied by region and management intensity: yield benefits were pronounced in arid regions due to improved moisture retention, while no significant yield differences were observed under high-input systems. NT increased topsoil carbon by ~6%, with greater benefits under straw retention[139]. Furthermore, NT adoption has expanded to over 205 million hectares globally, representing 14.7% of all croplands—a 92.9% increase over the past decade. Adoption rates exceed 70% in South America and Oceania [140]. Finally, recent studies have evaluated the interaction between NT and fertilization practices. Under low nitrogen input systems, combining NT with diverse cropping significantly enhances yields and nutrient use efficiency. In contrast, marginal benefits are reduced under high-input regimes[141].
7. Synergistic Effects Among Management Practices
7.1. Crop Rotation + No-Tillage: Coupled Improvement of Soil Health
Integrating crop rotation with no-tillage (NT) amplifies the benefits of each practice, creating compounded improvements in soil structure, organic matter retention, and biological activity.
Rotation diversifies root exudates and residue inputs, enriching microbial diversity and functional redundancy, while NT preserves physical soil integrity by minimizing disturbance[142]. Long-term experiments reveal that the combination of NT and diversified rotations increases soil organic carbon (SOC) stocks by 20–40% more than either practice alone[143]. Additionally, this integration enhances soil aggregation, reduces bulk density, and boosts water infiltration, leading to improved drought resilience and nutrient cycling efficiency [144]. Thus, coupling crop rotation with NT represents a foundational strategy for regenerative soil management.
7.2. Organic Amendments + No-Tillage: Promotion of Microbial Carbon Cycling
The co-application of organic amendments, particularly farmyard manure (FYM), with NT systems synergistically accelerates microbial-mediated carbon cycling.
FYM supplies complex carbon substrates, while NT maintains continuous microbial habitats, enhancing the establishment of functionally diverse decomposer communities[145]. Metagenomic analyses demonstrate that NT combined with organic inputs enriches genes involved in carbon degradation (e.g., cellulase, ligninase) and stabilizes soil aggregates via extracellular polymeric substances (EPS) production [146].
This synergy fosters the buildup of particulate organic matter (POM) and mineral-associated organic matter (MAOM), enhancing both short-term nutrient supply and long-term carbon sequestration[147]. Moreover, the combination reduces greenhouse gas emissions compared to conventional tillage with synthetic fertilizers, positioning it as a key strategy for climate-smart agriculture.
7.3. Integrated Systems for Enhanced Crop Productivity and Ecosystem Services
Truly sustainable farmland management demands integrated systems that combine crop rotation, NT, organic amendments, and microbiome stewardship. Multifactorial trials demonstrate that such integrated approaches outperform single-practice systems, achieving higher yields (+15–30%), greater soil biodiversity, enhanced nutrient cycling, and improved resilience to climatic stresses [76]. Beyond productivity, integrated systems deliver vital ecosystem services: carbon sequestration, water regulation, pest and disease suppression, and biodiversity conservation[148]. Precision agriculture technologies—such as soil sensors, microbiome diagnostics, and dynamic rotation modeling—further enable site-specific tailoring of integrated practices, maximizing synergistic benefits.
The future of sustainable agriculture lies in designing and managing holistic, diversified systems that restore and leverage the natural functionalities of soils and agroecosystems.
To strengthen this section’s relevance, we suggest referencing comprehensive new datasets and field experiments. A global assessment of cover crop strategies showed that while short-term cover cropping improves yield and carbon sequestration, it may increase GHG emissions temporarily. In contrast, implementing cover crops for ≥5 years in combination with NT and diversified rotations enhances multiple ecosystem services simultaneously [149]. The optimal protocol—mixing leguminous and non-leguminous cover species, crimping them 25 days before planting, and retaining surface mulch—yielded 1.25% higher multifunctionality compared to conventional systems. This integrated strategy could add 98 million tons of food production, sequester 2.17 billion tons of CO₂, and reduce 2.41 billion tons of soil erosion annually by 2100 [149]. In low-fertility soils, combining deep-rooted crops, rotations, and organic inputs substantially improved soil resilience and crop adaptation [150]. Additionally, long-term trials show that the performance of integrated practices depends on nutrient input levels—under low-input conditions, they act complementarily to boost yields, whereas under high inputs, their role shifts toward maintaining ecosystem stability (Figure 7) [141].
8. Challenges and Future Perspectives
8.1. Bottlenecks in Current Applications
Despite notable successes, significant challenges hinder the widespread implementation of integrated farmland management systems. One major bottleneck is early-stage yield variability, especially during the transition from conventional to conservation-oriented practices such as no-tillage and diversified rotations [151]. Adoption is further constrained by technical complexities, including the need for specialized machinery, precise residue management, and adaptive pest control strategies [152].
Additionally, knowledge gaps among farmers regarding microbial ecology and soil health often delay the uptake of biological-based management approaches. Closing these gaps requires targeted education, extension services, and supportive policy frameworks.
8.2. Trends in Precision Management and Smart Agriculture
Advances in precision agriculture are poised to transform farmland management by enabling real-time monitoring and targeted interventions.
Sensor-based microbial monitoring technologies, including soil DNA metabarcoding and volatile organic compound (VOC) profiling, allow dynamic assessment of soil health and pathogen pressures [153]. Coupled with remote sensing, machine learning, and Internet of Things (IoT) networks, these innovations facilitate site-specific management, optimizing fertilizer application, microbial inoculation, and rotation planning to match field heterogeneity[154]. Smart agriculture thus holds the promise of operationalizing complex ecological principles at scale, enhancing both productivity and environmental outcomes.
8.3. Role in Farmland Carbon Neutrality and Carbon Credit Mechanisms
Farmland systems adopting conservation tillage, organic amendments, and microbial optimization play a critical role in achieving carbon neutrality goals. By enhancing soil organic carbon sequestration, reducing greenhouse gas emissions, and improving nutrient use efficiency, these systems contribute substantially to climate mitigation strategies[155].
Emerging carbon credit markets now recognize soil carbon as a tradable asset, offering financial incentives to farmers who implement verified practices [3]. However, robust, standardized measurement protocols are essential to ensure credibility, permanence, and equitable participation in such schemes.
8.4. Future Research Directions
Key future research avenues include targeted regulation of functional microbial groups, such as designing consortia tailored to specific soil-climate-crop systems[156].
Synthetic microbial communities (SynComs) offer promising platforms to fine-tune nutrient cycling, disease suppression, and stress resilience in a predictable manner. Moreover, longitudinal field studies integrating multi-omics (metagenomics, metabolomics, transcriptomics) with agronomic data are urgently needed to unravel causal mechanisms and optimize practice design[157].
Building resilient, multifunctional soils through ecological engineering of microbiomes represents a frontier for sustainable agriculture, demanding interdisciplinary collaboration across microbiology, agronomy, ecology, and data science.
9. Conclusions
Soil health is the foundation of sustainable agricultural production, and the practices discussed—crop rotation, organic amendments, no-tillage agriculture, and soil microbiome management—each independently contribute critical benefits.
Crop rotation diversifies biological interactions, disrupts pest and pathogen cycles, and improves nutrient cycling efficiency. Organic amendments enrich soil organic matter, stimulate microbial activity, and enhance long-term fertility. No-tillage preserves soil structure, reduces erosion, and promotes carbon sequestration. Microbiome-targeted strategies amplify nutrient acquisition, stress resilience, and disease suppression.
While each practice offers measurable advantages when implemented alone, their synergistic integration—particularly through combined systems such as crop rotation with no-tillage or organic amendments with conservation practices—magnifies ecological functions and agronomic outcomes. Integrated management approaches stabilize yields, enhance soil biodiversity, build resilience against climatic stresses, and contribute to global climate mitigation through increased soil carbon stocks.
Moving forward, integrated soil health management must become the core paradigm for agricultural development. Through the coupling of ecological principles with precision technologies, farmers can optimize productivity while preserving the vital soil ecosystem services that underpin food security and environmental sustainability.
Integrated, synergistic management is not merely an option but a critical pathway toward regenerative and climate-resilient farming systems for the 21st century and beyond.
Author Contributions
Author Contributions: Conceptualization, X.L. and S.Y.; Methodology, X.L.; Software, X.L.; Validation, X.L., S.Y., and Y.J.; Formal Analysis, X.L.; Investigation, X.L.; Resources, X.L.; Data Curation, X.L.; Writing—Original Draft Preparation, X.L.; Writing—Review and Editing, X.L., S.Y., D.G., Y.J., and Y.W.; Visualization, X.L.; Supervision, S.Y.; Project Administration, S.Y.; Funding Acquisition, Y.J. All authors have read and agreed to the published version of the manuscript.
Funding
This article was funded by the Heilongjiang Provincial Natural Science Foundation of China (LH2023C066).
Data Availability Statement
Data are available through request to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Lal, R. Soil degradation as a reason for inadequate human nutrition. Food Security 2009, 1, 45-57. [CrossRef]
- Amundson, R.; Berhe, A.A.; Hopmans, J.W.; Olson, C.; Sztein, A.E.; Sparks, D.L. Soil and human security in the 21st century. Science 2015, 348, 1261071. [CrossRef]
- Paustian, K.; Lehmann, J.; Ogle, S.; Reay, D.; Robertson, G.P.; Smith, P. Climate-smart soils. Nature 2016, 532, 49-57.
- Scheierling, S.; Treguer, D.O.; Booker, J.F. Water Productivity in Agriculture: Looking for Water in the Agricultural Productivity and Efficiency Literature. Water Economics and Policy 2016, 02, 1650007, doi:10.1142/S2382624X16500077. [CrossRef]
- Nguyen, T.T.; Do, T.T.; Harper, R.; Pham, T.T.; Linh, T.V.K.; Le, T.S.; Thanh, L.B.; Giap, N.X. Soil Health Impacts of Rubber Farming: The Implication of Conversion of Degraded Natural Forests into Monoculture Plantations. Agriculture 2020, 10, 357.
- Liu, Z.; Liu, J.; Yu, Z.; Yao, Q.; Li, Y.; Liang, A.; Zhang, W.; Mi, G.; Jin, J.; Liu, X.; et al. Long-term continuous cropping of soybean is comparable to crop rotation in mediating microbial abundance, diversity and community composition. Soil and Tillage Research 2020, 197, 104503, https://doi.org/10.1016/j.still.2019.104503. [CrossRef]
- Hobbs, P.R.; Sayre, K.; Gupta, R. The role of conservation agriculture in sustainable agriculture. Philosophical Transactions of the Royal Society B: Biological Sciences 2008, 363, 543-555.
- Blanco-Canqui, H.; Ruis, S.J. No-tillage and soil physical environment. Geoderma 2018, 326, 164-200. [CrossRef]
- Oldfield, E.E.; Bradford, M.A.; Wood, S.A. Global meta-analysis of the relationship between soil organic matter and crop yields. Soil 2019, 5, 15-32. [CrossRef]
- Smith, M.E.; Bommarco, R.; Vico, G. Rethinking Crop Rotational Benefits Under Climate Change: Beyond the Growing Season. Glob Chang Biol 2025, 31, e70012, doi:10.1111/gcb.70012. [CrossRef]
- Srivastava, R.K.; Yetgin, A.; Srivastava, S. The role of legume roots in carbon sequestration, soil health enhancement, and salinity mitigation under climate change: A comprehensive review. Soil and Tillage Research 2025, 253, 106656, https://doi.org/10.1016/j.still.2025.106656. [CrossRef]
- Yang, X.; Xiong, J.; Du, T.; Ju, X.; Gan, Y.; Li, S.; Xia, L.; Shen, Y.; Pacenka, S.; Steenhuis, T.S.; et al. Diversifying crop rotation increases food production, reduces net greenhouse gas emissions and improves soil health. Nature Communications 2024, 15, 198, doi:10.1038/s41467-023-44464-9. [CrossRef]
- Sun, Y.; Yang, X.; Elsgaard, L.; Du, T.; Siddique, K.H.M.; Kang, S.; Butterbach-Bahl, K. Diversified crop rotations improve soil microbial communities and functions in a six-year field experiment. Journal of Environmental Management 2024, 370, 122604, https://doi.org/10.1016/j.jenvman.2024.122604. [CrossRef]
- Derpsch, R.; Friedrich, T.; Kassam, A.; Li, H. Current status of adoption of no-till farming in the world and some of its main benefits. International journal of agricultural and biological engineering 2010, 3, 1-25.
- Oys, E.; Krupek, F.S.; Proctor, C.; Koehler-Cole, K.; Basche, A. Exploring how multi-year cover crop use alters above and belowground weed communities in limited tillage corn–soybean systems. Frontiers in Agronomy 2025, Volume 7 - 2025. [CrossRef]
- Dhakal, M.; Rui, Y.; Benson, A.R.; Hinson, P.O.; Delate, K.; Afshar, R.K.; Luck, B.; Smith, A. Cover crop management strategies affect weeds and profitability of organic no-till soybean. Renewable Agriculture and Food Systems 2024, 39, e3, doi:10.1017/S1742170523000522. [CrossRef]
- Wu, B.; Zhang, M.; Zhai, Z.; Dai, H.; Yang, M.; Zhang, Y.; Liang, T. Soil organic carbon, carbon fractions, and microbial community under various organic amendments. Scientific Reports 2024, 14, 25431, doi:10.1038/s41598-024-75771-w. [CrossRef]
- Soman, C.; Li, D.; Wander, M.M.; Kent, A.D. Long-term fertilizer and crop-rotation treatments differentially affect soil bacterial community structure. Plant and Soil 2017, 413, 145-159. [CrossRef]
- Pandey, K.; Saharan, B.S. Soil microbiomes: a promising strategy for boosting crop yield and advancing sustainable agriculture. Discover Agriculture 2025, 3, 54, doi:10.1007/s44279-025-00208-5. [CrossRef]
- Wang, X.; Chi, Y.; Song, S. Important soil microbiota's effects on plants and soils: a comprehensive 30-year systematic literature review. Front Microbiol 2024, 15, 1347745, doi:10.3389/fmicb.2024.1347745. [CrossRef]
- Papin, M.; Philippot, L.; Breuil, M.C.; Bru, D.; Dreux-Zigha, A.; Mounier, A.; Le Roux, X.; Rouard, N.; Spor, A. Survival of a microbial inoculant in soil after recurrent inoculations. Scientific Reports 2024, 14, 4177, doi:10.1038/s41598-024-54069-x. [CrossRef]
- Ugwu, C.; Eze, V.; Ogenyi, F. Microbiome Engineering for Sustainable Agriculture. 2024, 3, 96-100.
- Denning, G. Sustainable intensification of agriculture: the foundation for universal food security. npj Sustainable Agriculture 2025, 3, 7, doi:10.1038/s44264-025-00047-3. [CrossRef]
- Bowles, T.M.; Mooshammer, M.; Socolar, Y.; Calderón, F.; Cavigelli, M.A.; Culman, S.W.; Deen, W.; Drury, C.F.; Garcia y Garcia, A.; Gaudin, A.C.M.; et al. Long-Term Evidence Shows that Crop-Rotation Diversification Increases Agricultural Resilience to Adverse Growing Conditions in North America. One Earth 2020, 2, 284-293, https://doi.org/10.1016/j.oneear.2020.02.007. [CrossRef]
- Tamburini, G.; Bommarco, R.; Wanger, T.C.; Kremen, C.; van der Heijden, M.G.A.; Liebman, M.; Hallin, S. Agricultural diversification promotes multiple ecosystem services without compromising yield. Science Advances 2020, 6, eaba1715, doi:doi:10.1126/sciadv.aba1715. [CrossRef]
- Zhou, J.; Zhu, P.; Kluger, D.M.; Lobell, D.B.; Jin, Z. Changes in the Yield Effect of the Preceding Crop in the US Corn Belt Under a Warming Climate. Global Change Biology 2024, 30, e17556, https://doi.org/10.1111/gcb.17556. [CrossRef]
- Zhou, Y.; Yang, Z.; Liu, J.; Li, X.; Wang, X.; Dai, C.; Zhang, T.; Carrión, V.J.; Wei, Z.; Cao, F.; et al. Crop rotation and native microbiome inoculation restore soil capacity to suppress a root disease. Nature Communications 2023, 14, 8126, doi:10.1038/s41467-023-43926-4. [CrossRef]
- Iheshiulo, E.M.-A.; Larney, F.J.; Hernandez-Ramirez, G.; St. Luce, M.; Liu, K.; Chau, H.W. Do diversified crop rotations influence soil physical health? A meta-analysis. Soil and Tillage Research 2023, 233, 105781, https://doi.org/10.1016/j.still.2023.105781. [CrossRef]
- Wu, H.; Liu, E.; Jin, T.; Liu, B.; Gopalakrishnan, S.; Zhou, J.; Shao, G.; Mei, X.; Delaplace, P.; De Clerck, C. Crop rotation increases Tibetan barley yield and soil quality on the Tibetan Plateau. Nature Food 2025, 6, 151-160, doi:10.1038/s43016-024-01094-8. [CrossRef]
- Peoples, M.B.; Herridge, D.F.; Ladha, J.K. Biological nitrogen fixation: an efficient source of nitrogen for sustainable agricultural production? In Proceedings of the Management of Biological Nitrogen Fixation for the Development of More Productive and Sustainable Agricultural Systems: Extended versions of papers presented at the Symposium on Biological Nitrogen Fixation for Sustainable Agriculture at the 15th Congress of Soil Science, Acapulco, Mexico, 1994, 1995; pp. 3-28.
- Yang, L.; Wang, L.; Chu, J.; Zhao, H.; Jie, Z.; Zang, H.; Yang, Y.; Zeng, Z. Improving soil quality and wheat yield through diversified crop rotations in the North China Plain. Soil and Tillage Research 2024, 244, 106231, doi:10.1016/j.still.2024.106231. [CrossRef]
- Drinkwater, L.E.; Snapp, S. Nutrients in agroecosystems: rethinking the management paradigm. Advances in agronomy 2007, 92, 163-186.
- Shen, J.; Yuan, L.; Zhang, J.; Li, H.; Bai, Z.; Chen, X.; Zhang, W.; Zhang, F. Phosphorus dynamics: from soil to plant. Plant physiology 2011, 156, 997-1005. [CrossRef]
- Vanderhasselt, A.; Steinwidder, L.; D’Hose, T.; Cornelis, W. Opening up the subsoil: Subsoiling and bio-subsoilers to remediate subsoil compaction in three fodder crop rotations on a sandy loam soil. Soil and Tillage Research 2024, 237, 105956, https://doi.org/10.1016/j.still.2023.105956. [CrossRef]
- Akchaya, K.; Parasuraman, P.; Pandian, K.; Vijayakumar, S.; Thirukumaran, K.; Mustaffa, M.R.A.F.; Rajpoot, S.K.; Choudhary, A.K. Boosting resource use efficiency, soil fertility, food security, ecosystem services, and climate resilience with legume intercropping: a review. Frontiers in Sustainable Food Systems 2025, Volume 9 - 2025, doi:10.3389/fsufs.2025.1527256. [CrossRef]
- Zhao, J.; Chen, J.; Beillouin, D.; Lambers, H.; Yang, Y.; Smith, P.; Zeng, Z.; Olesen, J.E.; Zang, H. Global systematic review with meta-analysis reveals yield advantage of legume-based rotations and its drivers. Nature Communications 2022, 13, 4926, doi:10.1038/s41467-022-32464-0. [CrossRef]
- Zou, Y.; Liu, Z.; Chen, Y.; Wang, Y.; Feng, S. Crop Rotation and Diversification in China: Enhancing Sustainable Agriculture and Resilience. Agriculture 2024, 14, 1465. [CrossRef]
- Gray, M.E.; Sappington, T.W.; Miller, N.J.; Moeser, J.; Bohn, M.O. Adaptation and invasiveness of western corn rootworm: intensifying research on a worsening pest. Annual review of entomology 2009, 54, 303-321. [CrossRef]
- Onstad, D.; Spencer, J.; Guse, C.; Levine, E.; Isard, S. Modeling evolution of behavioral resistance by an insect to crop rotation. Entomologia experimentalis et Applicata 2001, 100, 195-201. [CrossRef]
- S.F, H.; Ahmed, H.; Gossen, B.; Kutcher, H.; Brandt, S.; Strelkov, S.; Chang, K.-F.; Turnbull, G.D. Effect of Crop Rotation on the Soil Pathogen Population Dynamics and Canola Seedling Establishment. Plant Pathology Journal 2009, 8, doi:10.3923/ppj.2009.106.112. [CrossRef]
- Mendes, R.; Kruijt, M.; De Bruijn, I.; Dekkers, E.; Van Der Voort, M.; Schneider, J.H.; Piceno, Y.M.; DeSantis, T.Z.; Andersen, G.L.; Bakker, P.A. Deciphering the rhizosphere microbiome for disease-suppressive bacteria. Science 2011, 332, 1097-1100. [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.; Bakker, P.A. The rhizosphere microbiome and plant health. Trends in plant science 2012, 17, 478-486. [CrossRef]
- Carrión, V.J.; Perez-Jaramillo, J.; Cordovez, V.; Tracanna, V.; De Hollander, M.; Ruiz-Buck, D.; Mendes, L.W.; van Ijcken, W.F.; Gomez-Exposito, R.; Elsayed, S.S. Pathogen-induced activation of disease-suppressive functions in the endophytic root microbiome. Science 2019, 366, 606-612. [CrossRef]
- Weisberger, D.; Nichols, V.; Liebman, M. Does diversifying crop rotations suppress weeds? A meta-analysis. PLOS ONE 2019, 14, e0219847, doi:10.1371/journal.pone.0219847. [CrossRef]
- Owen, M.D.; Zelaya, I.A. Herbicide-resistant crops and weed resistance to herbicides. Pest Management Science: formerly Pesticide Science 2005, 61, 301-311.
- Du, Y.; Han, X.; Tsuda, K. Microbiome-mediated plant disease resistance: recent advances and future directions. Journal of General Plant Pathology 2025, 91, 1-17, doi:10.1007/s10327-024-01204-1. [CrossRef]
- Xun, W.; Liu, Y.; Ma, A.; Yan, H.; Miao, Y.; Shao, J.; Zhang, N.; Xu, Z.; Shen, Q.; Zhang, R. Dissection of rhizosphere microbiome and exploiting strategies for sustainable agriculture. New Phytologist 2024, 242, 2401-2410, https://doi.org/10.1111/nph.19697. [CrossRef]
- Sappington, T.W.; Spencer, J.L. Movement Ecology of Adult Western Corn Rootworm: Implications for Management. Insects 2023, 14, 922. [CrossRef]
- Meinke, L.J.; Reinders, J.D.; Dang, T.B.; Krumm, J.T.; Pilcher, C.D.; Carroll, M.W.; Head, G.P. Resistance management and integrated pest management insights from deployment of a Cry3Bb1+ Gpp34Ab1/Tpp35Ab1 pyramid in a resistant western corn rootworm landscape. PLOS ONE 2024, 19, e0299483, doi:10.1371/journal.pone.0299483. [CrossRef]
- Blanco-Canqui, H.; Ruis, S. Cover crop impacts on soil physical properties: A review. Soil Science Society of America Journal 2020, 84, doi:10.1002/saj2.20129. [CrossRef]
- Guo, X.; Wang, H.; Yu, Q.; Ahmad, N.; Li, J.; Wang, R.; Wang, X. Subsoiling and plowing rotation increase soil C and N storage and crop yield on a semiarid Loess Plateau. Soil and Tillage Research 2022, 221, 105413, https://doi.org/10.1016/j.still.2022.105413. [CrossRef]
- Cassman, K.G.; Dobermann, A.; Walters, D.T. Agroecosystems, nitrogen-use efficiency, and nitrogen management. AMBIO: A Journal of the Human Environment 2002, 31, 132-140.
- Govaerts, B.; Sayre, K.D.; Deckers, J. Stable high yields with zero tillage and permanent bed planting? Field crops research 2005, 94, 33-42.
- Chaparro, J.M.; Sheflin, A.M.; Manter, D.K.; Vivanco, J.M. Manipulating the soil microbiome to increase soil health and plant fertility. Biology and Fertility of Soils 2012, 48, 489-499. [CrossRef]
- Chaparro, J.M.; Badri, D.V.; Vivanco, J.M. Rhizosphere microbiome assemblage is affected by plant development. The ISME journal 2014, 8, 790-803. [CrossRef]
- Hanif, M.S.; Tayyab, M.; Baillo, E.H.; Islam, M.M.; Islam, W.; Li, X. Plant microbiome technology for sustainable agriculture. Frontiers in Microbiology 2024, Volume 15 - 2024, doi:10.3389/fmicb.2024.1500260. [CrossRef]
- Jansson, J.K.; Hofmockel, K.S. Soil microbiomes and climate change. Nature Reviews Microbiology 2020, 18, 35-46, doi:10.1038/s41579-019-0265-7. [CrossRef]
- Kaur, R.; Gupta, S.; Tripathi, V.; Bharadwaj, A. Unravelling the secrets of soil microbiome and climate change for sustainable agroecosystems. Folia Microbiologica 2025, 70, 19-40, doi:10.1007/s12223-024-01194-9. [CrossRef]
- Liu, Y.; Wen, M.; Hu, R.; Zhao, F.; Wang, J. Regulation of wheat yield by soil multifunctionality and metagenomic-based microbial degradation potentials under crop rotations. J Environ Manage 2024, 370, 122897, doi:10.1016/j.jenvman.2024.122897. [CrossRef]
- Maul, J.E.; Cavigelli, M.A.; Vinyard, B.; Buyer, J.S. Cropping system history and crop rotation phase drive the abundance of soil denitrification genes nirK, nirS and nosZ in conventional and organic grain agroecosystems. Agriculture, Ecosystems & Environment 2019, 273, 95-106, https://doi.org/10.1016/j.agee.2018.11.022. [CrossRef]
- Wagg, C.; Bender, S.F.; Widmer, F.; Van Der Heijden, M.G. Soil biodiversity and soil community composition determine ecosystem multifunctionality. Proceedings of the National Academy of Sciences 2014, 111, 5266-5270. [CrossRef]
- Bach, E.M.; Baer, S.G.; Meyer, C.K.; Six, J. Soil texture affects soil microbial and structural recovery during grassland restoration. Soil biology and biochemistry 2010, 42, 2182-2191. [CrossRef]
- Chang, X.; Yan, L.; Naeem, M.; Khaskheli, M.I.; Zhang, H.; Gong, G.; Zhang, M.; Song, C.; Yang, W.; Liu, T.; et al. Maize/Soybean Relay Strip Intercropping Reduces the Occurrence of Fusarium Root Rot and Changes the Diversity of the Pathogenic Fusarium Species. Pathogens 2020, 9, doi:10.3390/pathogens9030211. [CrossRef]
- Chang, X.; Wei, D.; Zeng, Y.; Zhao, X.; Hu, Y.; Wu, X.; Song, C.; Gong, G.; Chen, H.; Yang, C.; et al. Maize-soybean relay strip intercropping reshapes the rhizosphere bacterial community and recruits beneficial bacteria to suppress Fusarium root rot of soybean. Frontiers in Microbiology 2022, Volume 13 - 2022, doi:10.3389/fmicb.2022.1009689. [CrossRef]
- Meng, X.; Wang, B.; Zhang, X.; Liu, C.; Ji, J.; Hao, X.; Yang, B.; Wang, W.; Xu, D.; Zhang, S.; et al. Long-Term Crop Rotation Revealed the Relationship Between Soil Organic Carbon Physical Fraction and Bacterial Community at Aggregate Scales. Microorganisms 2025, 13, doi:10.3390/microorganisms13030496. [CrossRef]
- Stewart, C.E.; Plante, A.F.; Paustian, K.; Conant, R.T.; Six, J. Soil carbon saturation: linking concept and measurable carbon pools. Soil Science Society of America Journal 2008, 72, 379-392. [CrossRef]
- Seifert, C.A.; Roberts, M.J.; Lobell, D.B. Continuous Corn and Soybean Yield Penalties across Hundreds of Thousands of Fields. Agronomy Journal 2017, 109, 541-548, https://doi.org/10.2134/agronj2016.03.0134. [CrossRef]
- Ladha, J.K.; Pathak, H.; Krupnik, T.J.; Six, J.; van Kessel, C. Efficiency of fertilizer nitrogen in cereal production: retrospects and prospects. Advances in agronomy 2005, 87, 85-156.
- Zhang, L.; Yuan, J.; Zhang, M.; Zhang, Y.; Wang, L.; Li, J. Long term effects of crop rotation and fertilization on crop yield stability in southeast China. Scientific Reports 2022, 12, 14234, doi:10.1038/s41598-022-17675-1. [CrossRef]
- Chauhan, B.S.; Mahajan, G.; Sardana, V.; Timsina, J.; Jat, M.L. Productivity and sustainability of the rice–wheat cropping system in the Indo-Gangetic Plains of the Indian subcontinent: problems, opportunities, and strategies. Advances in agronomy 2012, 117, 315-369.
- Kandpal, A.; T M, K.; Pal, S. Does conservation agriculture promote Sustainable intensification in the rice–wheat System of the Indo-Gangetic plains in India? Empirical evidences from on-farm studies. Current Science 2023, 124, 1188-1193, doi:10.18520/cs/v124/i10/1188-1193. [CrossRef]
- Zhao, J.; Chen, J.; Beillouin, D.; Lambers, H.; Yang, Y.; Smith, P.; Zeng, Z.; Olesen, J.E.; Zang, H. Global systematic review with meta-analysis reveals yield advantage of legume-based rotations and its drivers. Nat Commun 2022, 13, 4926, doi:10.1038/s41467-022-32464-0. [CrossRef]
- O'Donovan, J.T.; Grant, C.A.; Blackshaw, R.E.; Harker, K.N.; Johnson, E.N.; Gan, Y.; Lafond, G.P.; May, W.E.; Turkington, T.K.; Lupwayi, N.Z.; et al. Rotational Effects of Legumes and Non-Legumes on Hybrid Canola and Malting Barley. Agronomy Journal 2014, 106, 1921-1932, https://doi.org/10.2134/agronj14.0236. [CrossRef]
- Van der Werf, H.M.; Petit, J. Evaluation of the environmental impact of agriculture at the farm level: a comparison and analysis of 12 indicator-based methods. Agriculture, Ecosystems & Environment 2002, 93, 131-145.
- Liang, J.; He, Z.; Shi, W. Cotton/mung bean intercropping improves crop productivity, water use efficiency, nitrogen uptake, and economic benefits in the arid area of Northwest China. Agricultural Water Management 2020, 240, 106277, doi:10.1016/j.agwat.2020.106277. [CrossRef]
- Davis, A.S.; Hill, J.D.; Chase, C.A.; Johanns, A.M.; Liebman, M. Increasing cropping system diversity balances productivity, profitability and environmental health. 2012. [CrossRef]
- Liu, X.; Wu, X.; Liang, G.; Zheng, F.; Zhang, M.; Li, S. A global meta-analysis of the impacts of no-tillage on soil aggregation and aggregate-associated organic carbon. Land Degradation & Development 2021, 32, 5292-5305, doi:10.1002/ldr.4109. [CrossRef]
- Meng, X.; Meng, F.; Chen, P.; Hou, D.; Zheng, E.; Xu, T. A meta-analysis of conservation tillage management effects on soil organic carbon sequestration and soil greenhouse gas flux. Science of The Total Environment 2024, 954, 176315, https://doi.org/10.1016/j.scitotenv.2024.176315. [CrossRef]
- Řezáčová, V.; Czakó, A.; Stehlík, M.; Mayerová, M.; Šimon, T.; Smatanová, M.; Madaras, M. Organic fertilization improves soil aggregation through increases in abundance of eubacteria and products of arbuscular mycorrhizal fungi. Scientific Reports 2021, 11, 12548, doi:10.1038/s41598-021-91653-x. [CrossRef]
- Hafeez, F.Y.; Yasmin, S.; Ariani, D.; Renseigné, N.; Zafar, Y.; Malik, K.A. Plant growth-promoting bacteria as biofertilizer. Agronomy for sustainable development 2006, 26, 143-150. [CrossRef]
- Prosser, J.I.; Nicol, G.W. Archaeal and bacterial ammonia-oxidisers in soil: the quest for niche specialisation and differentiation. Trends in microbiology 2012, 20, 523-531. [CrossRef]
- Jin, X.; Yang, X.; Peng, S.; Ma, E.; Zhang, H.; Lin, X.; Wang, Y.; Li, J. Cropping rotation improved the bacterial diversity and N-cycling genes in tobacco fields through a 19-year long-term experiment. Applied Soil Ecology 2024, 193, 105165, https://doi.org/10.1016/j.apsoil.2023.105165. [CrossRef]
- de Werra, P.; Péchy-Tarr, M.; Keel, C.; Maurhofer, M. Role of gluconic acid production in the regulation of biocontrol traits of Pseudomonas fluorescens CHA0. Appl Environ Microbiol 2009, 75, 4162-4174, doi:10.1128/aem.00295-09. [CrossRef]
- Sharma, S.B.; Sayyed, R.Z.; Trivedi, M.H.; Gobi, T.A. Phosphate solubilizing microbes: sustainable approach for managing phosphorus deficiency in agricultural soils. SpringerPlus 2013, 2, 1-14.
- Parmar, P.; Sindhu, S. Potassium solubilization by rhizosphere bacteria: influence of nutritional and environmental conditions. J Microbiol Res 2013, 3, 25-31.
- Meena, V.S.; Maurya, B.R.; Verma, J.P.; Meena, R.S. Potassium solubilizing microorganisms for sustainable agriculture; Springer: 2016; Volume 331.
- Cipriano, M.A.P.; Freitas-Iório, R.d.P.; Dimitrov, M.R.; de Andrade, S.A.L.; Kuramae, E.E.; Silveira, A.P.D.d. Plant-Growth Endophytic Bacteria Improve Nutrient Use Efficiency and Modulate Foliar N-Metabolites in Sugarcane Seedling. Microorganisms 2021, 9, 479.
- Misra, S.; Chauhan, P.S. ACC deaminase-producing rhizosphere competent Bacillus spp. mitigate salt stress and promote Zea mays growth by modulating ethylene metabolism. 3 Biotech 2020, 10, 119, doi:10.1007/s13205-020-2104-y. [CrossRef]
- Vurukonda, S.S.K.P.; Vardharajula, S.; Shrivastava, M.; SkZ, A. Enhancement of drought stress tolerance in crops by plant growth promoting rhizobacteria. Microbiological research 2016, 184, 13-24. [CrossRef]
- Alhoqail, W.A. ACC-Deaminase producing Pseudomonas putida RT12 inoculation: A promising strategy for improving Brassica juncea tolerance to salinity stress. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 2024, 52, 13550, doi:10.15835/nbha52113550. [CrossRef]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—opportunistic, avirulent plant symbionts. Nature reviews microbiology 2004, 2, 43-56.
- Vandenkoornhuyse, P.; Quaiser, A.; Duhamel, M.; Le Van, A.; Dufresne, A. The importance of the microbiome of the plant holobiont. New Phytologist 2015, 206, 1196-1206. [CrossRef]
- van Capelle, C.; Schrader, S.; Brunotte, J. Tillage-induced changes in the functional diversity of soil biota–A review with a focus on German data. European Journal of Soil Biology 2012, 50, 165-181. [CrossRef]
- Schmidt, J.E.; Bowles, T.M.; Gaudin, A.C. Using ancient traits to convert soil health into crop yield: impact of selection on maize root and rhizosphere function. Frontiers in Plant Science 2016, 7, 373. [CrossRef]
- Guan, Y.; Xu, B.; Zhang, X.; Yang, W. Tillage Practices and Residue Management Manipulate Soil Bacterial and Fungal Communities and Networks in Maize Agroecosystems. Microorganisms 2022, 10, doi:10.3390/microorganisms10051056. [CrossRef]
- Ouyang, Y.; Reeve, J.R.; Norton, J.M. The quality of organic amendments affects soil microbiome and nitrogen-cycling bacteria in an organic farming system. Frontiers in Soil Science 2022, Volume 2 - 2022, doi:10.3389/fsoil.2022.869136. [CrossRef]
- Govaerts, B.; Sayre, K.D.; Deckers, J. A minimum data set for soil quality assessment of wheat and maize cropping in the highlands of Mexico. Soil and tillage research 2006, 87, 163-174. [CrossRef]
- Zhou, J.; Jiang, X.; Wei, D.; Zhao, B.; Ma, M.; Chen, S.; Cao, F.; Shen, D.; Guan, D.; Li, J. Consistent effects of nitrogen fertilization on soil bacterial communities in black soils for two crop seasons in China. Scientific Reports 2017, 7, 3267, doi:10.1038/s41598-017-03539-6. [CrossRef]
- Benitez, M.-S.; Ewing, P.M.; Osborne, S.L.; Lehman, R.M. Rhizosphere microbial communities explain positive effects of diverse crop rotations on maize and soybean performance. Soil Biology and Biochemistry 2021, 159, 108309, https://doi.org/10.1016/j.soilbio.2021.108309. [CrossRef]
- Peralta, A.L.; Sun, Y.; McDaniel, M.D.; Lennon, J.T. Crop rotational diversity increases disease suppressive capacity of soil microbiomes. Ecosphere 2018, 9, e02235. [CrossRef]
- Newman, M.M.; Hoilett, N.; Lorenz, N.; Dick, R.P.; Liles, M.R.; Ramsier, C.; Kloepper, J.W. Glyphosate effects on soil rhizosphere-associated bacterial communities. Science of the Total Environment 2016, 543, 155-160. [CrossRef]
- Fitzpatrick, C.R.; Salas-González, I.; Conway, J.M.; Finkel, O.M.; Gilbert, S.; Russ, D.; Teixeira, P.J.P.L.; Dangl, J.L. The plant microbiome: from ecology to reductionism and beyond. Annual review of microbiology 2020, 74, 81-100. [CrossRef]
- Ge, A.H.; Wang, E. Exploring the plant microbiome: A pathway to climate-smart crops. Cell 2025, 188, 1469-1485, doi:10.1016/j.cell.2025.01.035. [CrossRef]
- Mo, Y.; Bier, R.; Li, X.; Daniels, M.; Smith, A.; Yu, L.; Kan, J. Agricultural practices influence soil microbiome assembly and interactions at different depths identified by machine learning. Communications Biology 2024, 7, 1349, doi:10.1038/s42003-024-07059-8. [CrossRef]
- Liu, C.; Wang, J.; Wang, Y.; Li, L.; Feng, Z.; Xian, Y.; Jiang, Y.; Yu, J.; Tong, T.; Li, X.; et al. Crop rotation and fertilization shape the microbiomes of maize rhizosphere soil with distinct mechanisms. Plant and Soil 2025, 507, 89-108, doi:10.1007/s11104-024-06847-9. [CrossRef]
- Timofeeva, A.M.; Galyamova, M.R.; Sedykh, S.E. Plant Growth-Promoting Soil Bacteria: Nitrogen Fixation, Phosphate Solubilization, Siderophore Production, and Other Biological Activities. Plants 2023, 12, 4074. [CrossRef]
- Li, J.; Wang, J.; Liu, H.; Macdonald, C.A.; Singh, B.K. Application of microbial inoculants significantly enhances crop productivity: A meta-analysis of studies from 2010 to 2020. Journal of Sustainable Agriculture and Environment 2022, 1, 216-225, https://doi.org/10.1002/sae2.12028. [CrossRef]
- Saleem, M.; Nawaz, F.; Hussain, M.B.; Ikram, R.M. Comparative Effects of Individual and Consortia Plant Growth Promoting Bacteria on Physiological and Enzymatic Mechanisms to Confer Drought Tolerance in Maize (Zea mays L.). Journal of Soil Science and Plant Nutrition 2021, 21, 3461-3476, doi:10.1007/s42729-021-00620-y. [CrossRef]
- Haque, Z.; Nawaz, S.; Haidar, L.; Ansari, M.S.A. Development of novel Trichoderma bioformulations against Fusarium wilt of chickpea. Scientific Reports 2025, 15, 9564, doi:10.1038/s41598-025-86984-y. [CrossRef]
- Minchev, Z.; Kostenko, O.; Soler, R.; Pozo, M.J. Microbial Consortia for Effective Biocontrol of Root and Foliar Diseases in Tomato. Frontiers in Plant Science 2021, Volume 12 - 2021, doi:10.3389/fpls.2021.756368. [CrossRef]
- Haskett, T.L.; Tkacz, A.; Poole, P.S. Engineering rhizobacteria for sustainable agriculture. Isme j 2021, 15, 949-964, doi:10.1038/s41396-020-00835-4. [CrossRef]
- Bossio, D.A.; Cook-Patton, S.C.; Ellis, P.W.; Fargione, J.; Sanderman, J.; Smith, P.; Wood, S.; Zomer, R.J.; von Unger, M.; Emmer, I.M.; et al. The role of soil carbon in natural climate solutions. Nature Sustainability 2020, 3, 391-398, doi:10.1038/s41893-020-0491-z. [CrossRef]
- Cui, J.; Yang, B.; Zhang, M.; Song, D.; Xu, X.; Ai, C.; Liang, G.; Zhou, W. Investigating the effects of organic amendments on soil microbial composition and its linkage to soil organic carbon: A global meta-analysis. Science of The Total Environment 2023, 894, 164899, https://doi.org/10.1016/j.scitotenv.2023.164899. [CrossRef]
- Luo, G.; Li, L.; Friman, V.-P.; Guo, J.; Guo, S.; Shen, Q.; Ling, N. Organic amendments increase crop yields by improving microbe-mediated soil functioning of agroecosystems: A meta-analysis. Soil Biology and Biochemistry 2018, 124, doi:10.1016/j.soilbio.2018.06.002. [CrossRef]
- Whalen, J.K.; Chang, C.; Clayton, G.W.; Carefoot, J.P. Cattle manure amendments can increase the pH of acid soils. Soil Science Society of America Journal 2000, 64, 962-966. [CrossRef]
- Dhaliwal, S.S.; Sharma, V.; Shukla, A.K.; Verma, V.; Kaur, M.; Singh, P.; Gaber, A.; Hossain, A. Effect of addition of organic manures on basmati yield, nutrient content and soil fertility status in north-western India. Heliyon 2023, 9, e14514, doi:10.1016/j.heliyon.2023.e14514. [CrossRef]
- Salehi, A.; Surböck, A.; Gollner, G.; Eitzinger, J.; Friedel, J.K.; Wohlmuth, M.-L.; Freyer, B. The effects of organic fertilization systems on yield and yield variability of lucerne and market crops: Insights from a 10-year field experiment in Austria. European Journal of Agronomy 2025, 169, 127662, https://doi.org/10.1016/j.eja.2025.127662. [CrossRef]
- Canellas, L.P.; Olivares, F.L. Physiological responses to humic substances as plant growth promoter. Chemical and Biological Technologies in Agriculture 2014, 1, 1-11. [CrossRef]
- Heidari, G.; Mohammadi, K.; Sohrabi, Y. Responses of Soil Microbial Biomass and Enzyme Activities to Tillage and Fertilization Systems in Soybean (Glycine max L.) Production. Front Plant Sci 2016, 7, 1730, doi:10.3389/fpls.2016.01730. [CrossRef]
- Das, S.; Liptzin, D.; Maharjan, B. Long-term manure application improves soil health and stabilizes carbon in continuous maize production system. Geoderma 2023, 430, 116338, https://doi.org/10.1016/j.geoderma.2023.116338. [CrossRef]
- Bolo, P.; Mucheru-Muna, M.W.; Mwirichia, R.K.; Kinyua, M.; Ayaga, G.; Kihara, J. Influence of Farmyard Manure Application on Potential Zinc Solubilizing Microbial Species Abundance in a Ferralsol of Western Kenya. Agriculture 2023, 13, 2217. [CrossRef]
- Jin, Y.; Zhang, N.; Chen, Y.; Wang, Q.; Qin, Z.; Sun, Z.; Zhang, S. Quantitative Evaluation of the Crop Yield, Soil-Available Phosphorus, and Total Phosphorus Leaching Caused by Phosphorus Fertilization: A Meta-Analysis. Agronomy 2023, 13, 2436. [CrossRef]
- Guo, J.H.; Liu, X.J.; Zhang, Y.; Shen, J.; Han, W.; Zhang, W.; Christie, P.; Goulding, K.; Vitousek, P.; Zhang, F. Significant acidification in major Chinese croplands. science 2010, 327, 1008-1010. [CrossRef]
- Yang, F.; Tian, J.; Fang, H.; Gao, Y.; Xu, M.; Lou, Y.; Zhou, B.; Kuzyakov, Y. Functional Soil Organic Matter Fractions, Microbial Community, and Enzyme Activities in a Mollisol Under 35 Years Manure and Mineral Fertilization. Journal of Soil Science and Plant Nutrition 2019, 19, 430-439, doi:10.1007/s42729-019-00047-6. [CrossRef]
- Maitlo, A.A.; Zhang, S.; Ahmed, W.; Jangid, K.; Ali, S.; Yang, H.; Bhatti, S.M.; Duan, Y.; Xu, M. Potential Nitrogen Mineralization and Its Availability in Response to Long-Term Fertilization in a Chinese Fluvo-Aquic Soil. Agronomy 2022, 12, 1260. [CrossRef]
- Strudley, M.W.; Green, T.R.; Ascough II, J.C. Tillage effects on soil hydraulic properties in space and time: State of the science. Soil and Tillage Research 2008, 99, 4-48.
- Liao, K.; Feng, J.; Lai, X.; Zhu, Q. Effects of environmental factors on the influence of tillage conversion on saturated soil hydraulic conductivity obtained with different methodologies: a global meta-analysis. SOIL 2022, 8, 309-317, doi:10.5194/soil-8-309-2022. [CrossRef]
- West, T.O.; Post, W.M. Soil organic carbon sequestration rates by tillage and crop rotation: a global data analysis. Soil Science Society of America Journal 2002, 66, 1930-1946.
- Alvarez, R.; Steinbach, H.S. A review of the effects of tillage systems on some soil physical properties, water content, nitrate availability and crops yield in the Argentine Pampas. Soil and tillage research 2009, 104, 1-15. [CrossRef]
- Prairie, A.M.; King, A.E.; Cotrufo, M.F. Restoring particulate and mineral-associated organic carbon through regenerative agriculture. Proc Natl Acad Sci U S A 2023, 120, e2217481120, doi:10.1073/pnas.2217481120. [CrossRef]
- Yin, C.; McLaughlin, K.; Paulitz, T.C.; Kroese, D.R.; Hagerty, C.H. Population Dynamics of Wheat Root Pathogens Under Different Tillage Systems in Northeast Oregon. Plant Dis 2020, 104, 2649-2657, doi:10.1094/pdis-03-19-0621-re. [CrossRef]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The rhizosphere microbiome: significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS microbiology reviews 2013, 37, 634-663. [CrossRef]
- Caamal-Maldonado, J.A.; Jiménez-Osornio, J.J.; Torres-Barragán, A.; Anaya, A.L. The use of allelopathic legume cover and mulch species for weed control in cropping systems. Agronomy journal 2001, 93, 27-36. [CrossRef]
- Wallace, J.M.; Williams, A.; Liebert, J.A.; Ackroyd, V.J.; Vann, R.A.; Curran, W.S.; Keene, C.L.; VanGessel, M.J.; Ryan, M.R.; Mirsky, S.B. Cover crop-based, organic rotational no-till corn and soybean production systems in the mid-Atlantic United States. Agriculture 2017, 7, 34. [CrossRef]
- Helgason, B.; Walley, F.; Germida, J. No-till soil management increases microbial biomass and alters community profiles in soil aggregates. Applied Soil Ecology 2010, 46, 390-397. [CrossRef]
- Spedding, T.; Hamel, C.; Mehuys, G.; Madramootoo, C. Soil microbial dynamics in maize-growing soil under different tillage and residue management systems. Soil Biology and Biochemistry 2004, 36, 499-512. [CrossRef]
- Costa, O.Y.; Raaijmakers, J.M.; Kuramae, E.E. Microbial extracellular polymeric substances: ecological function and impact on soil aggregation. Frontiers in microbiology 2018, 9, 1636. [CrossRef]
- Chukwuneme, C.F.; Ayangbenro, A.S.; Babalola, O.O. Metagenomic Analyses of Plant Growth-Promoting and Carbon-Cycling Genes in Maize Rhizosphere Soils with Distinct Land-Use and Management Histories. Genes 2021, 12, 1431. [CrossRef]
- Cui, Y.; Zhang, W.; Zhang, Y.; Liu, X.; Zhang, Y.; Zheng, X.; Luo, J.; Zou, J. Effects of no-till on upland crop yield and soil organic carbon: a global meta-analysis. Plant and Soil 2024, 499, 363-377, doi:10.1007/s11104-022-05854-y. [CrossRef]
- Kassam, A.; Friedrich, T.; Derpsch, R. Successful Experiences and Lessons from Conservation Agriculture Worldwide. Agronomy 2022, 12, 769. [CrossRef]
- MacLaren, C.; Mead, A.; van Balen, D.; Claessens, L.; Etana, A.; de Haan, J.; Haagsma, W.; Jäck, O.; Keller, T.; Labuschagne, J.; et al. Long-term evidence for ecological intensification as a pathway to sustainable agriculture. Nature Sustainability 2022, 5, 770-779, doi:10.1038/s41893-022-00911-x. [CrossRef]
- Blanco-Canqui, H.; Shaver, T.M.; Lindquist, J.L.; Shapiro, C.A.; Elmore, R.W.; Francis, C.A.; Hergert, G.W. Cover crops and ecosystem services: Insights from studies in temperate soils. Agronomy journal 2015, 107, 2449-2474. [CrossRef]
- Maiga, A.; Alhameid, A.; Singh, S.; Polat, A.; Singh, J.; Kumar, S.; Osborne, S. Responses of soil organic carbon, aggregate stability, carbon and nitrogen fractions to 15 and 24 years of no-till diversified crop rotations. Soil Research 2019, 57, 149-157, https://doi.org/10.1071/SR18068. [CrossRef]
- Franzluebbers, A. Soil organic matter stratification ratio as an indicator of soil quality. Soil and Tillage Research 2002, 66, 95-106. [CrossRef]
- Srour, A.Y.; Ammar, H.A.; Subedi, A.; Pimentel, M.; Cook, R.L.; Bond, J.; Fakhoury, A.M. Microbial Communities Associated With Long-Term Tillage and Fertility Treatments in a Corn-Soybean Cropping System. Front Microbiol 2020, 11, 1363, doi:10.3389/fmicb.2020.01363. [CrossRef]
- Liang, C.; Schimel, J.P.; Jastrow, J.D. The importance of anabolism in microbial control over soil carbon storage. Nature microbiology 2017, 2, 1-6. [CrossRef]
- Lehmann, J.; Kleber, M. The contentious nature of soil organic matter. Nature 2015, 528, 60-68. [CrossRef]
- Tamburini, G.; Bommarco, R.; Wanger, T.C.; Kremen, C.; van der Heijden, M.G.A.; Liebman, M.; Hallin, S. Agricultural diversification promotes multiple ecosystem services without compromising yield. Sci Adv 2020, 6, doi:10.1126/sciadv.aba1715. [CrossRef]
- Qiu, T.; Shi, Y.; Peñuelas, J.; Liu, J.; Cui, Q.; Sardans, J.; Zhou, F.; Xia, L.; Yan, W.; Zhao, S.; et al. Optimizing cover crop practices as a sustainable solution for global agroecosystem services. Nature Communications 2024, 15, 10617, doi:10.1038/s41467-024-54536-z. [CrossRef]
- Zhang, W.-P.; Surigaoge, S.; Yang, H.; Yu, R.-P.; Wu, J.-P.; Xing, Y.; Chen, Y.; Li, L. Diversified cropping systems with complementary root growth strategies improve crop adaptation to and remediation of hostile soils. Plant and Soil 2024, 502, 7-30, doi:10.1007/s11104-023-06464-y. [CrossRef]
- Pittelkow, C.M.; Liang, X.; Linquist, B.A.; Van Groenigen, K.J.; Lee, J.; Lundy, M.E.; Van Gestel, N.; Six, J.; Venterea, R.T.; Van Kessel, C. Productivity limits and potentials of the principles of conservation agriculture. Nature 2015, 517, 365-368. [CrossRef]
- Knowler, D.; Bradshaw, B. Farmers’ adoption of conservation agriculture: A review and synthesis of recent research. Food policy 2007, 32, 25-48. [CrossRef]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proceedings of the National Academy of Sciences 2006, 103, 626-631. [CrossRef]
- Lawes, R.; Mata, G.; Richetti, J.; Fletcher, A.; Herrmann, C. Using remote sensing, process-based crop models, and machine learning to evaluate crop rotations across 20 million hectares in Western Australia. Agronomy for Sustainable Development 2022, 42, 120, doi:10.1007/s13593-022-00851-y. [CrossRef]
- Lal, R. Soil carbon sequestration impacts on global climate change and food security. science 2004, 304, 1623-1627. [CrossRef]
- Toju, H.; Peay, K.G.; Yamamichi, M.; Narisawa, K.; Hiruma, K.; Naito, K.; Fukuda, S.; Ushio, M.; Nakaoka, S.; Onoda, Y. Core microbiomes for sustainable agroecosystems. Nature plants 2018, 4, 247-257. [CrossRef]
- Tariq, A.; Guo, S.; Farhat, F.; Shen, X. Engineering Synthetic Microbial Communities: Diversity and Applications in Soil for Plant Resilience. Agronomy 2025, 15, 513. [CrossRef]
Figure 1.
Overview of Soil-Health-Based Sustainable Farming Strategies.
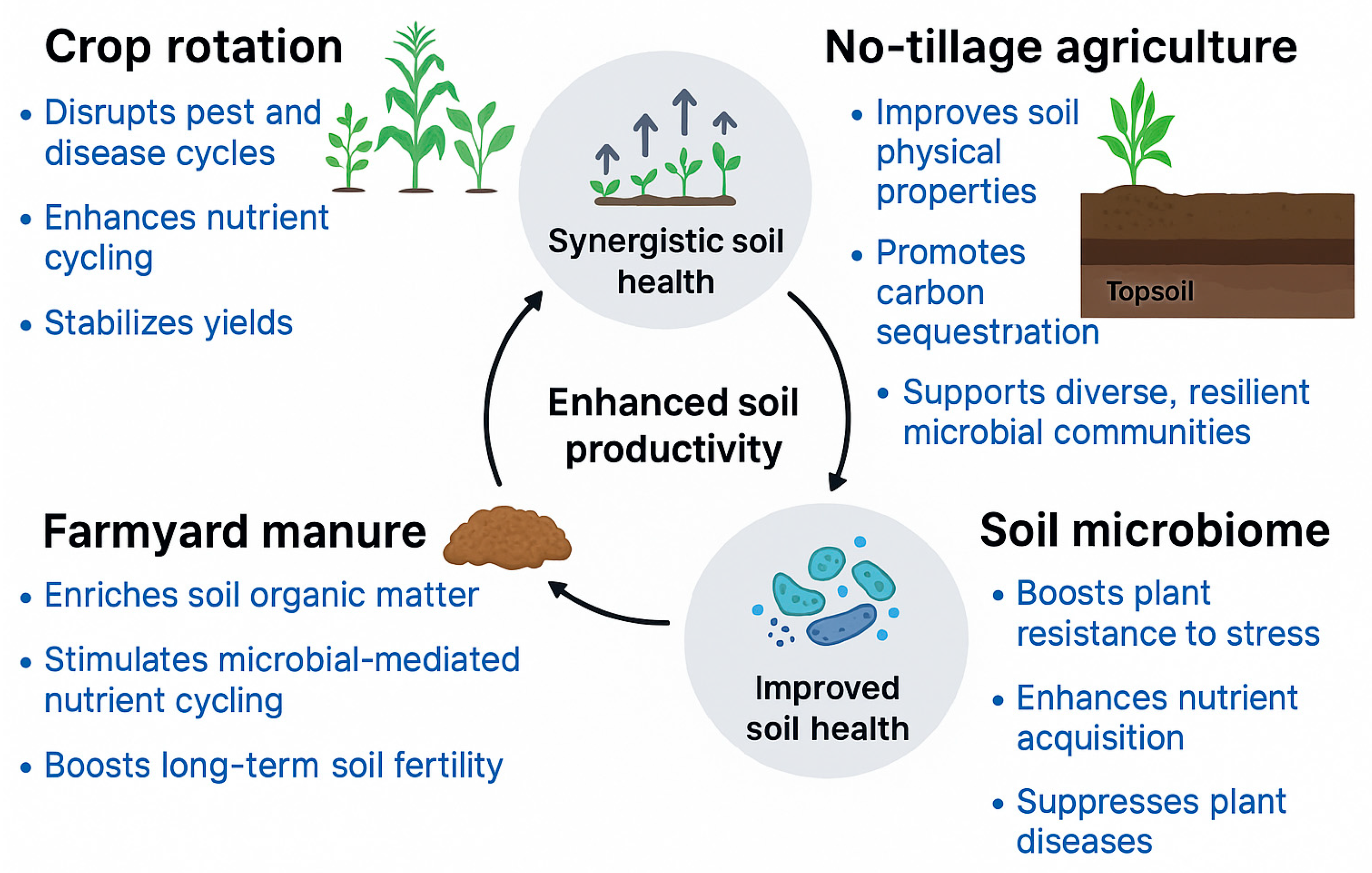
Figure 3.
Ecological Roles of Crop Rotation in Sustainable Agriculture.
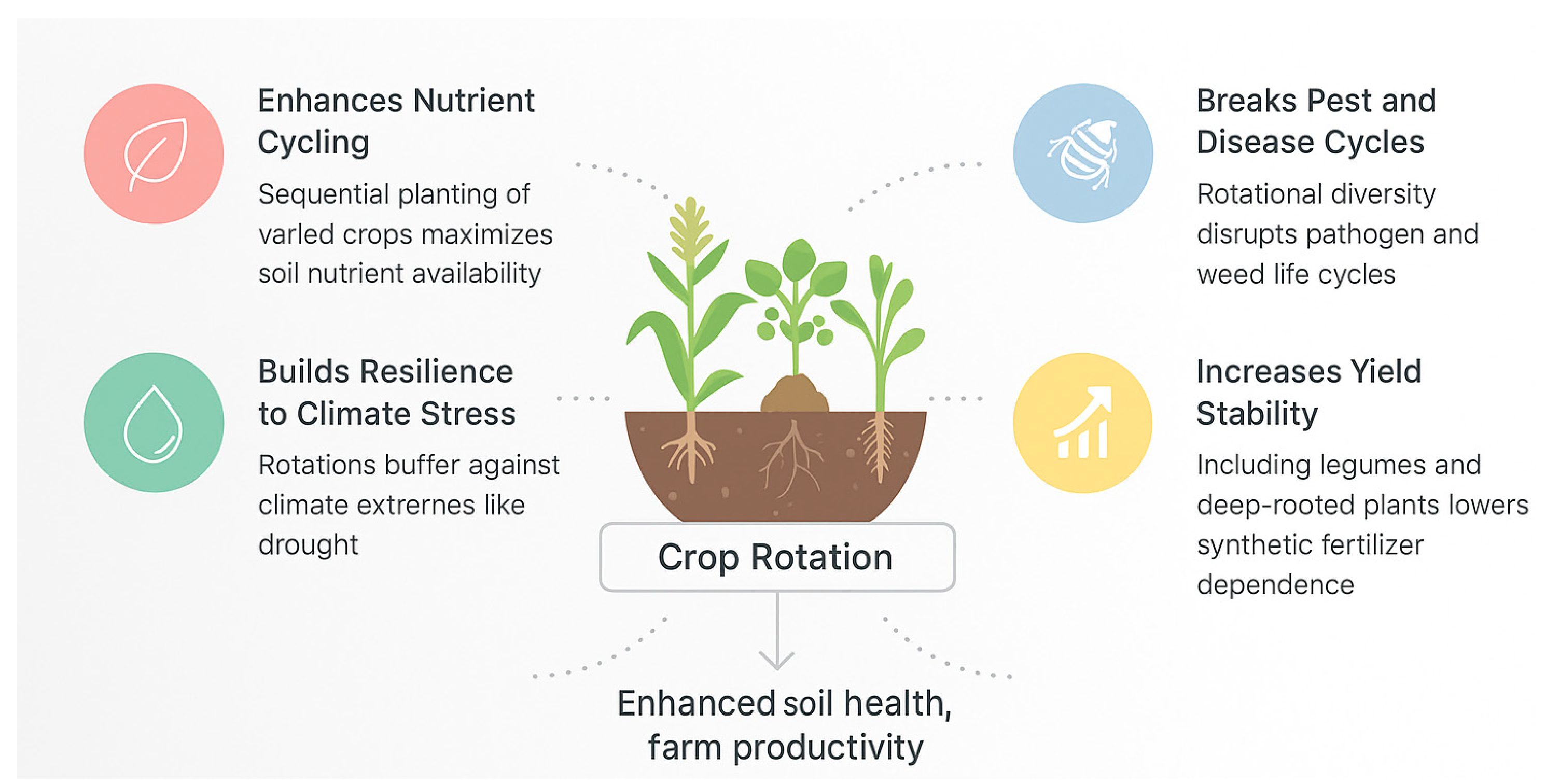
Figure 4.
Effects of Tillage Practices on Soil Aggregates and Ecosystem Functions.
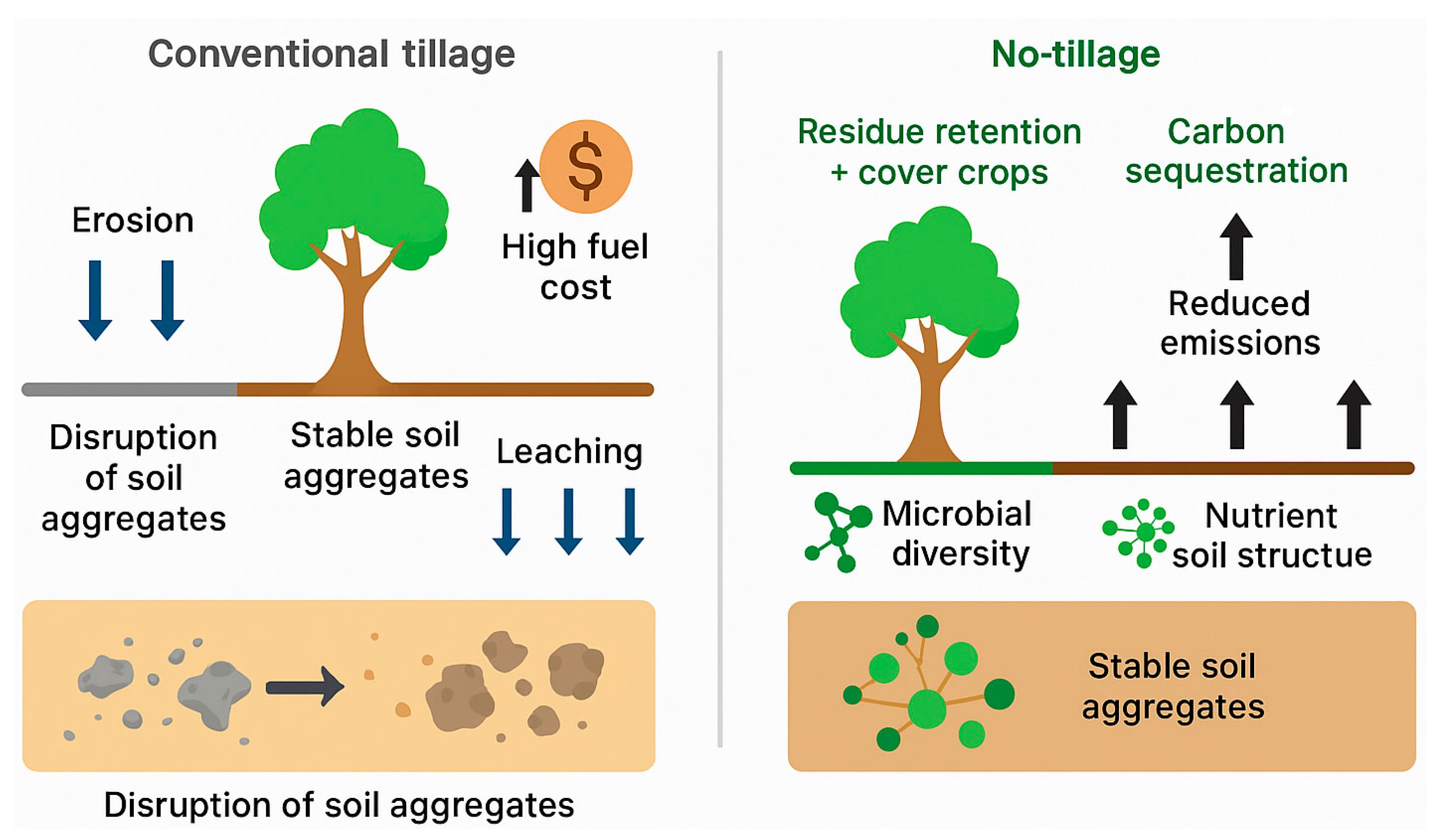
Figure 5.
Mechanisms by which farmyard manure improves soil structure, nutrient cycling, and crop productivity.
Figure 5.
Mechanisms by which farmyard manure improves soil structure, nutrient cycling, and crop productivity.
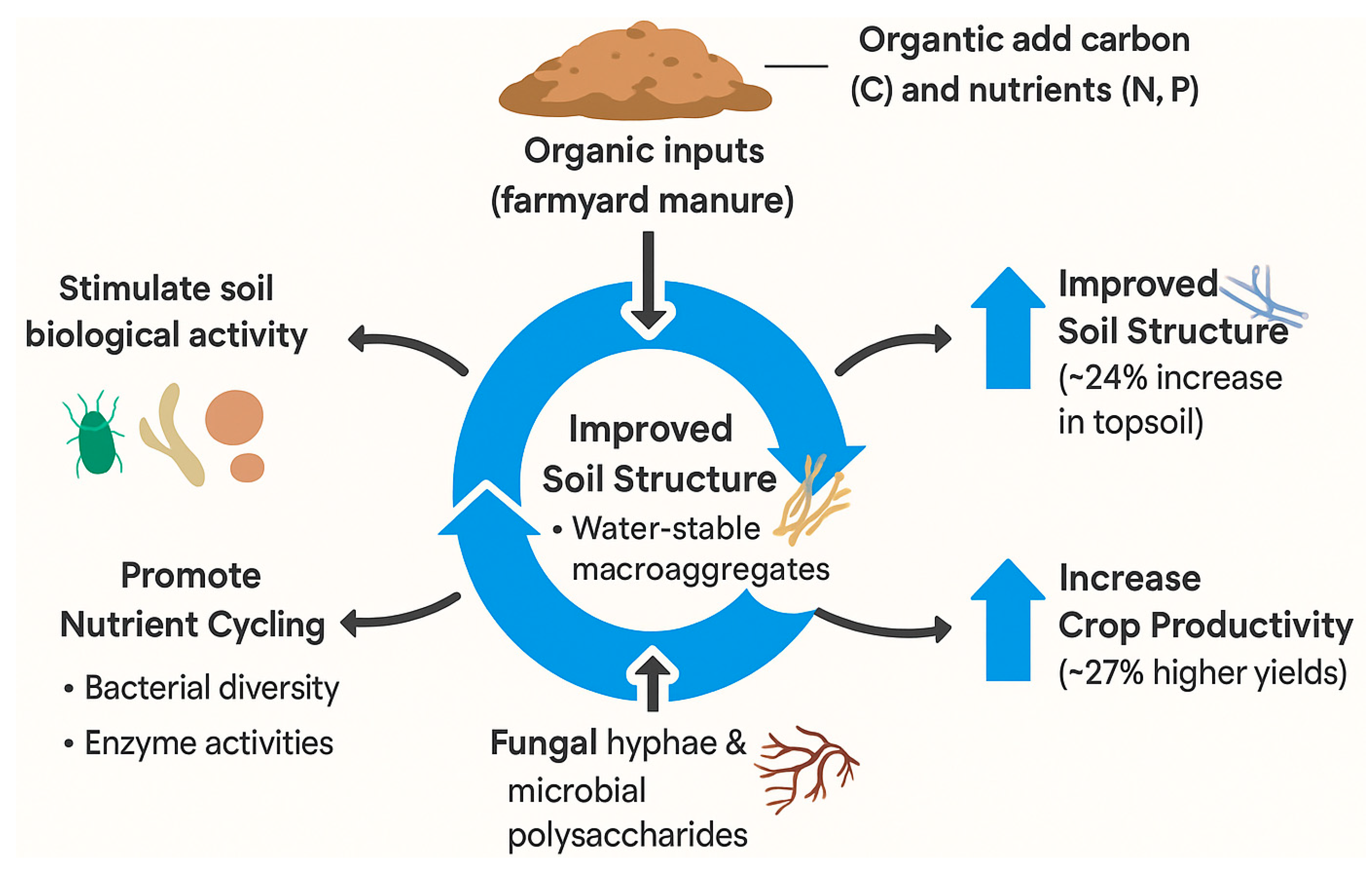
Figure 7.
Ecosystem Benefits of Integrated Soil Management Practices.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



