Submitted:
11 June 2025
Posted:
13 June 2025
You are already at the latest version
Abstract
Plant-derived vesicle-like nanoparticles (PDVLNs) are bioactive nanovesicles secreted by plant cells, emerging as a novel therapeutic tool for tissue repair and regeneration due to their low immunogenicity, intrinsic bioactivity, and potential as drug delivery carriers. This review examines PDVLNs’ biogenesis mechanisms, isolation techniques, and compositional diversity, emphasizing their roles in promoting essential regenerative processes—cell proliferation, differentiation, migration, immune modulation, and angiogenesis. We explore their therapeutic applications across multiple tissue types, including skin, bone, neural, liver, gastrointestinal, cardiovascular, and dental tissues, using both natural and engineered PDVLNs in various disease models. Compared to mammalian exosomes, PDVLNs offer advantages such as reduced immune rejection and ethical concerns, enhancing their sustainability and appeal for regenerative medicine. However, challenges in clinical translation, including scalability, standardization, and safety, remain. This paper consolidates current knowledge on PDVLNs, highlighting their versatility and providing insights into engineering strategies to optimize efficacy, ultimately outlining future research directions to advance their clinical potential.
Keywords:
plant-derived vesicle-like nanoparticles
; tissue repair
; regenerative medicine
; nanotherapeutics
; drug delivery
-
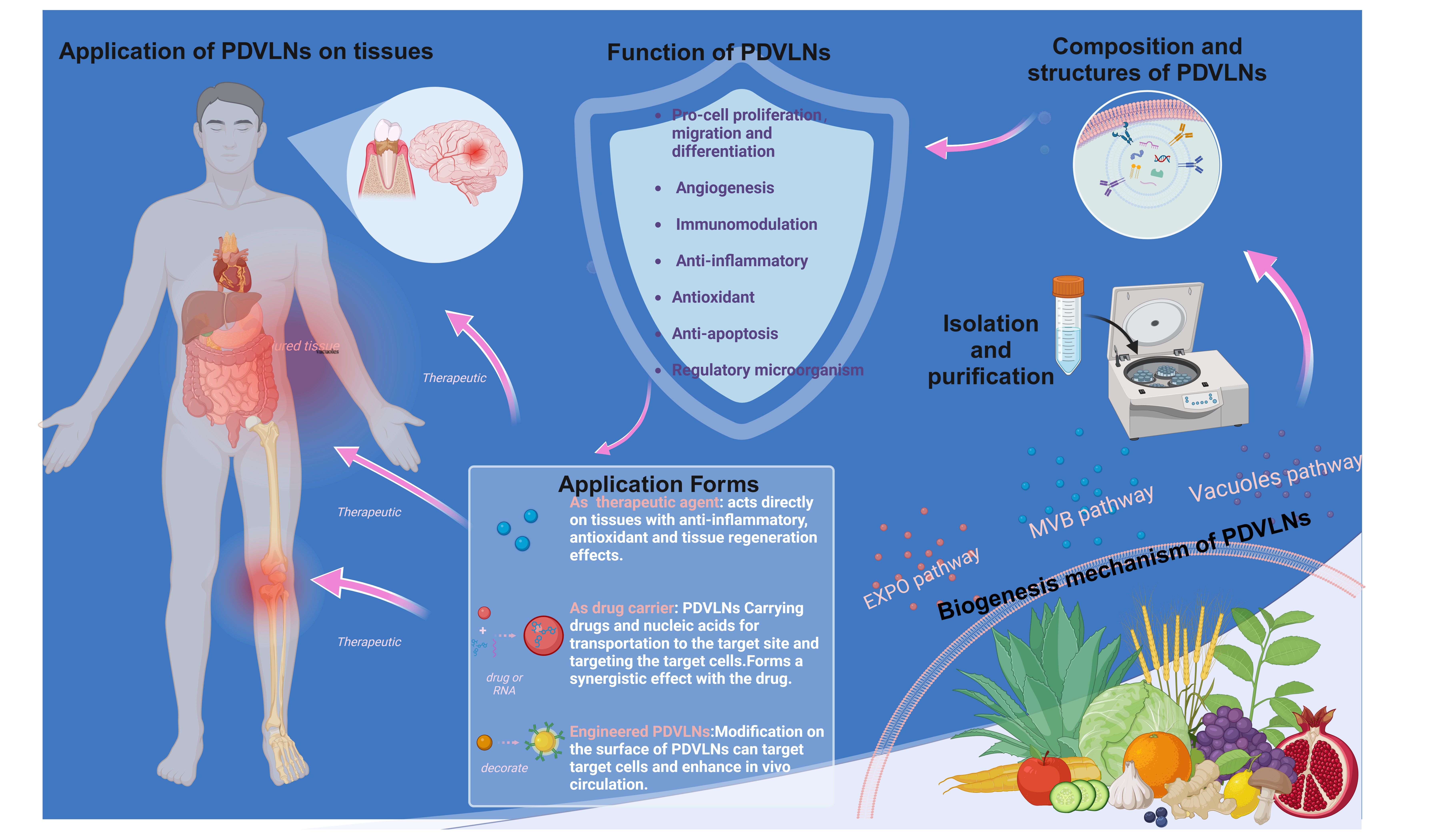
TOC (created with biorender.com)
 Plant vesicle-like nanoparticles (PDVLNs) may become a new avenue for the treatment of tissue injury, promoting tissue repair and regeneration through their intrinsic bioactivity or as drug delivery carriers. In addition, PDVLNs can be engineered and modified to achieve better results.
Plant vesicle-like nanoparticles (PDVLNs) may become a new avenue for the treatment of tissue injury, promoting tissue repair and regeneration through their intrinsic bioactivity or as drug delivery carriers. In addition, PDVLNs can be engineered and modified to achieve better results.
1. Introduction
Human tissue is the basic structural unit of the body. Its complex structure and function are coordinated to maintain normal physiological function, and different types of tissues work in concert to ensure effective organ and system functioning [1] However, external trauma, disease, or congenital defects can cause tissue damage or defects in the human body, affecting the health and function of the whole organism [2]. Effective treatment is essential for restoring the physiological function of tissues and organs [3]. Traditional treatments, including surgical repair, transplantation, and physical therapy, face limitations such as donor scarcity, immune rejection, and surgical complications [3,4]. Stem cell therapy promotes regeneration and repair of damaged tissues by implanting pluripotent stem cells or inducing cell differentiation [5,6]. Nevertheless, unregulated proliferation and aberrant differentiation of stem cells could result in unpredictable therapeutic outcomes or tumor formation [7]. To reduce risk, cell-free therapies have become alternative therapeutic strategies [8], which utilize cell-secreted biomolecules to promote tissue repair and regeneration without directly using the cells themselves [9,10]. Extensive research conducted on mammalian-originated exosomes in the context of tissue injury repair highlights their pivotal role in cell-free therapeutic strategies. However, potential risks are associated with exosomes of mammalian origin, such as immune reactions and disease transmission [11].
Plant-derived vesicle-like nanoparticles (PDVLNs) are produced by plant cells through specific biological pathways and are often involved in plant cell–environment interactions, such as plant growth, development, and fighting pathogenic microorganisms [12]. PDVLNs harbor a plethora of biologically active molecules alongside plant-specific growth factors and hormones. These constituents confer diverse functionalities to PDVLNs, including antioxidant, immunomodulatory, and cellular proliferative properties, enabling their participation in and regulation of a variety of physiological processes [13]. Compared with mammalian exosomes, PDVLNs have a lower risk of immunogenicity and immune rejection [14]. Furthermore, PDVLNs circumvent ethical concerns associated with animal experimentation or embryo donation, rendering them more sustainable and socially acceptable. As their use in regenerative medicine evolves, numerous studies have shown their significant potential in areas such as tissue engineering, drug delivery, and disease therapeutics [15,16,17].
This review consolidates PDVLNs’ mechanisms, applications, and engineering potential, critically evaluating their advantages over existing regenerative agents and outlining a roadmap for overcoming current limitations.
2. Overview of PDVLNs
2.1. Biogenesis Mechanism of PDVLNs
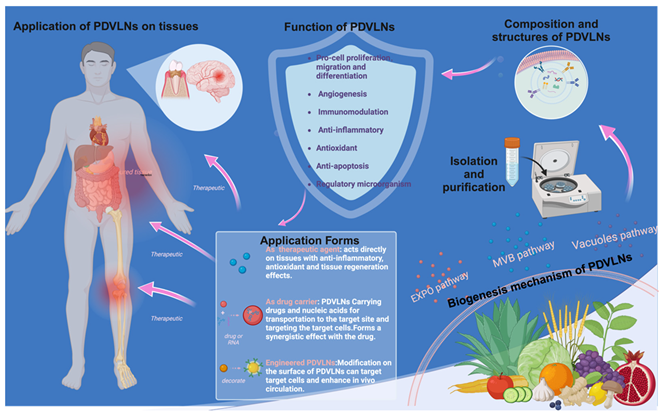
PDVLNs were initially identified in carrot cells through electron microscopy (EM) during the 1960s [18]. Despite numerous successful studies on the isolation and characterization of PDVLNs, their biogenesis remains poorly understood [19]. Three potential biogenesis pathways include the multivesicular body (MVB), EXPO, and vesicular pathways [16]. The three biogenesis pathways are shown in Figure 1.
MVBs are late nuclear endosomes that arise from the formation of trans-Golgi networks (TGNs) or early endosomes [20]. These networks or early endosomes further facilitate the maturation of MVBs, leading to the formation of characteristic intraluminal vesicles (ILVs) within them, which are enriched with biologically active molecules such as RNA, lipids, and proteins [16]. The subsequent fusion of MVBs with the plasma membrane (PM) leads to exocytosis of ILVs into the extracellular space, eliminating intracellular waste products and enabling efficient delivery of bioactive molecules [21]. The MVB pathway, as a core mechanism, exhibits inherent diversity under a wide range of conditions [22]. Initially, during plant–microbe symbiosis, the fusion of MVBs with host-derived plexiform membranes (PAMs) is facilitated [23]. PAM in continuity with the PM releases ILVs into the clumped peripheral region between plants and fungi, potentially modulating disease resistance signaling and the inhibition of pathogenic microorganisms [24], as evidenced in Arabidopsis infected with turnip mosaic virus and in radish mosaic virus infection [25,26]. Second, MVBs mediate antifungal defense responses by fusing with membrane structures at the site of fungal infection [27]. In barley powdery mildew fungal infestation, MVBs may be released both by fusion with the PM and by secretion to specific sites of infection to release exosomes. Ultimately, MVBs may release PDVLNs extracellularly through a mechanism involving direct fusion with the PM [28,29,30]. The regulatory mechanisms of the MVB pathway under various environmental conditions equip plants with crucial molecular strategies to cope with external environmental challenges and combat pathogenic microorganisms, culminating in the formation and release of PDVLNs [29].
Recent advancements have elucidated the structural and functional properties of vesicles, showing that they contain ILVs and that specific MVBs can deliver ILVs to vesicles through a fusion mechanism with the vesicles [31]. Ultimately, upon the fusion of vesicles with the cytoplasmic membrane, the enclosed ILVs are extruded from the cell and enriched with sRNAs and proteins implicated in defense responses [32]. Under conditions of bacterial infection, MVBs are translocated to the central vesicle and degraded via endocytosed vesicles [33]. Concurrently, parietal cells utilize their membrane architecture to merge with the cytoplasmic membrane, enabling the secretion of vesicles into the extracellular milieu [26]. For example, the structure of MVBs with ILVs was observed in the epidermal cells of grapefruit in their PM and in the cytoplasm surrounding the central vesicle [34]. This is an important defense mechanism for plants against pathogen invasion and a pathway for the formation of released PDVLNs.
In Arabidopsis thaliana and tobacco cells, EXPO, an exocyst-positive organelle, aids in cytokinesis by facilitating the transfer of cytoplasmic contents to the cell wall, particularly during the fusion of PDVLNs with the cytoplasmic membrane [35]. The shape of EXPO is significantly different from MVB [36], with high-pressure frozen samples shown to have a spherical double-membrane structure [37]. The Exo70E2 protein in Arabidopsis plays a central role in EXPO formation [35]. This extracapsular protein is uniquely localized on discrete punctate structural domains of the cytoplasmic membrane and the double membrane architecture of EXPO, which is distinct from the inner membrane markers of the Golgi apparatus, TGN, and MVB [24]. During PDVLN release, EXPO fuses with the PM, liberating its inner single-membrane vesicle into the cell wall. Markers on both the inner surface of the vesicle and in its peripheral region indicate eventual rupture and release of Exo70E2 into the cell wall [26,37]. EXPO is not restricted to a specific cell type and is observed in diverse plant structures, and it represents a distinct form of secretion exclusive to plants [38]. Although the biological function of EXPO-mediated secretion of PDVLNs requires further exploration, this finding provides a completely new perspective for understanding the secretory pathway in plant cells.
PDVLN biogenesis has far-reaching implications for multifaceted scientific research. This study contributes to a deeper understanding of molecular interactions and regulatory networks within plant cells, uncovering the functions and roles of PDVLNs in a variety of biological processes. Moreover, this investigation provides new insights for the artificial preparation of PDVLNs by leveraging natural generation mechanisms and utilizing biological systems to drive their production. Upon successful synthesis, further analysis of their structure and properties can be conducted to facilitate clinical translation. Through an in-depth comparison with natural PDVLNs, the differences between artificially prepared PDVLNs and natural PDVLNs can be identified. This study is anticipated to provide pivotal technology for large-scale production and enable extensive applications of PDVLNs in various domains, including regenerative medicine.
2.2. Composition of PDVLNs
PDVLNs comprise lipids, nucleic acids, proteins, and other constituents [39]. Given their complexity and heterogeneity, there is no specialized database for PDVLN components [40]. This heterogeneity is reflected in two main aspects: (1) PDVLNs from different sources have different compositional components. (2) Vesicle-like nanoparticles (VLNs) from distinct parts of the same plant exhibit varying compositions. The biological functions of PDVLNs vary depending on preparation and isolation methods, highlighting the importance of investigating their composition.
2.2.1. Lipids
Lipids play a pivotal role in the membrane structure of PDVLNs, influencing their formation, release, biological functionality, and cellular internalization [41]. Secretion of PDVLNs was reduced in Arabidopsis leaf cells lacking glycosylinositol phosphate ceramide (GIPC). In contrast, stimulation of Arabidopsis leaves with exogenous GIPC significantly increased PDVLN formation and release [42]. PDVLN lipids are mainly categorized into glycerolipids and phospholipids, which have significantly less cholesterol than animal cells do [43]. In grape-derived VLNs (GDVLNs), up to 98% of the lipids are phospholipids, with typical plant lipids, such as galactolipids, accounting for the remaining 2%, suggesting precise lipid sorting during Gp-DVLN biogenesis [44,45]. This precise lipid sorting was reflected in nanovesicles from different sources, such as the varying contents of phosphatidic acid (PA) and monogalactosylglycerol in VLNs of ginger or grape origin [46,47].
Notably, lipids are also active compounds in PDVLNs. In ginger-derived VLNs (Gg-DVLNs), denaturing proteins, removing RNA, or extracting lipids and reassembling them into liposomes inhibit the activity of NOD-like receptor protein 3 (NLRP3) inflammatory vesicles [48]. PAs in Gg-DVLNs, which make up 50% of phospholipids, interact with mammalian target of rapamycin (mTOR), an unconventional serine/threonine kinase pivotal in orchestrating the regulation of cell growth and proliferation [49]. Lipids also induce specific cellular responses in recipient cells, such as lipids within ginger-derived nanoparticles that upregulate Foxa2 expression in intestinal epithelial cells, independent of RNA or protein influence [50]. Despite the anti-inflammatory and pro-cell proliferation functions of lipids, our understanding of them is limited; therefore, a more comprehensive lipidomic analysis of different PDVLNs is needed.
2.2.2. Carbohydrates
PDVLNs are rich in protein, ranging from cellular communication to the delivery of bioactive substances, and their presence adds a new dimension to cell biology [51,52]. In conventional protein secretion, proteins originate from the endoplasmic reticulum, traverse the Golgi apparatus and TGN, and are ultimately secreted extracellularly. In unconventional protein secretion (UPS), however, proteins do not need to pass through these organelles, a pathway by which the UPS is formed in PDVLNs [53]. PDVLNs are also rich in membrane proteins that dominate and regulate communication between cells [54]. Moreover, the proteins on these vesicles contain specific markers, such as the PEN1 protein and TET8-GFP of PDVLNs in transgenic Arabidopsis, that reflect the origin and localization of the vesicles and the secretion mechanism [27]. Additional investigations are warranted to determine whether PEN1 or TET8 can function as universal protein biomarkers across diverse PDVLNs [55].
In addition to proteins that regulate cellular activity and function, the proteome of PDVLNs also includes enzymes related to carbohydrate and lipid metabolism and lattice protein chains and ATPases [56]. The presence of these proteins further corroborates the multifunctionality of PDVLNs [57]. More importantly, specific proteins in PDVLNs, such as water channel proteins, provide better stability to vesicles, enabling PDVLNs to be used as carriers for drug delivery [58]. Therefore, an in-depth understanding of the protein composition of PDVLNs reveals their biological importance and may also pave the way for their application in drug transportation in regeneration.
2.2.3. Nucleic Acids
PDVLNs loaded with nucleic acids also show specificity, with constituent RNAs, especially miRNAs, of particular interest. PDVLNs exhibit a selective packaging mechanism for RNAs, with specific miRNAs and siRNAs preferentially loaded into PDVLNs. This is evident from the distinct RNA profiles observed between plant tissues and PDVLNs [59]. For example, the types and amounts of miRNAs in strawberry- and ginger-derived PDVLNs differ significantly from those present in their parental tissues [60,61]. The RNA composition of various plants may vary depending on the species, as demonstrated by the total RNA content extracted from fruit tissues or fruit-derived PDVLNs, such as grapefruit and grape, being significantly lower than that extracted from vegetable tissues or vegetable-derived PDVLNs, such as carrot and ginger [30]. This selective RNA packaging reveals the diverse biological activities of PDVLNs and the importance of RNA delivery in intercellular communication.
miRNAs perform a diverse array of regulatory functions within cells. They can bind to target mRNAs, thereby degrading them or inhibiting their translation [62]. PDVLNs provide a platform for carrying and delivering these functional miRNAs. For instance, Ginseng-derived VLNs (Gs-DVLNs) delivered 20 miRNAs to bone marrow mesenchymal stem cells (BMSCs), modulating genes linked to neural differentiation, maturation, and function. Both the GO and KEGG analyses underscore the significant potential of miRNAs within Gs-DVLNs to regulate neurally relevant processes [63]. Moreover, honey-derived VLNs efficiently suppressed NLRP3 inflammatory vesicle activity through MiR-4057 [64]. Compared with animal miRNAs, plant miRNAs undergo natural 20-O-methylations, increasing their stability and protection against degradation and uridylation in various environments. These findings highlight the potential of plant miRNAs as drug carriers for therapeutic applications [65].
PDVLNs can serve as vehicles for RNA delivery, transferring RNA to mammalian cells to modulate gene expression, thus enabling plant–animal cellular communication. The RNA composition of PDVLNs varies among different plant species, necessitating further research to understand these differences and leverage them for the advancement of tissue repair and regeneration applications.
2.3. Preparation and Isolation Methods for PDVLNs
The process of preparing PDVLNs begins with the extraction of sap from the plant material. The appropriate extraction method is chosen on the basis of the plant species. For juice-rich fruit plants such as grapes and apples, the tissue lysis method is used, which releases abundant juice by breaking the plant tissue cells and provides ideal raw materials for the subsequent preparation of PDVLNs. In contrast, for rhizomes such as ginseng, because of their denser tissues, the tissue osmosis method is used, in which the solvent gradually penetrates the interior of the plant tissues through osmosis to efficiently extract the desired sap. These two methods ensure efficient and accurate extraction of the raw materials required for the preparation of PDVLNs according to plant characteristics, laying a solid foundation for subsequent steps. In addition to the direct isolation of PDVLNs from fresh plants, boiled herbal tonics have been shown to be effective sources. Recent studies have demonstrated that herbal broths contain abundant lipids and substantial quantities of sRNA [66]. An in-depth analysis of the composition of the broth revealed that vesicular nanoparticles loaded with sRNAs exhibited antifibrotic and anti-inflammatory potential. These findings suggest that herbal tonics hold great promise for obtaining and isolating PDVLNs [67]. The main methods for the preparation and isolation of PDVLNs are shown in Figure 2.
Segregation involves the separation of PDVLN and non-PDVLN components and the separation of distinct types of PDVLNs [68]. Separation methods are fundamental to the study of nanoparticles and have an important impact on subsequent extensive preparation and characterization work. Various separation techniques may result in discrepancies in the size, shape, and surface characteristics of nanoparticles, consequently influencing their biological activity and applicability [69]. Consequently, exploring isolation techniques for PDVLNs is crucial for their potential applications in tissue repair and regeneration. The main methods for isolating PDVLNs and their respective advantages and limitations are summarized in Table 1.

Table 1.
Isolation methods for the PDVLN.
| Separation method |
Principle | Advantages | Drawbacks | Make superior | Reference |
|---|---|---|---|---|---|
| Ultracentrifugation | Separation of particles with large differences in size and density | Simple and economical | Formation of proteins, exosome aggregates; expensive equipment and expertise required | Combining density gradient centrifugation to obtain purer nanoparticles | [70,71,72,73,74,75] |
| Dense differential velocity gradient centrifugation | Centrifugal settling or sedimentation equilibrium in an inert gradient medium results in the formation of different separation zones. | Improved separation purity | Time-consuming, requires expensive equipment and specialized knowledge | Combine with differential ultracentrifugation | [69,73,74,75,76] |
| Ultrafiltration | Separation of large and small molecules by pressure membranes | Does not affect exosome activity, economical | Filter membranes prone to clogging, affecting recycling rates | Ultrafiltration pre-concentration of samples combined with size exclusion chromatography | [77,78,79,80,81,82] |
| Size exclusion chromatography (SEC) |
Based on particle size differences | High purity, high yield | Inability to selectively extract specific size exosomes | Combine with ultrafiltration pre-concentration | [83,84,85] |
| Asymmetric flow field flow separation | Field effects cause particles to move at different speeds in different channels | Wide range of separated nanoparticles | Inability to distinguish between differently shaped aggregates | Immunoaffinity capture after preliminary separation using asymmetric flow field flow separation method | [84,85,86] |
| Immunoaffinity capture method | Utilizes high affinity antigen-antibody binding | Highly selective | Requires specific antibodies or magnetic beads, may affect biological function | Combine with asymmetric flow field flow separation | [55,87,88,89,90,91,92] |
| PEG-based precipitation | Utilizes the intrinsic negative charge property on the surface of PDVLNs | Simultaneously removing impurities such as nucleic acids and proteins | Non-specificity, not suitable suitable for proteins that are sensitive to the presence of PEG | Combine with electrophoresis and dialysis | [93] |
In the preparation of PDVLNs, in addition to the selection of the separation method, the preparation conditions, including temperature, pH, centrifugation speed, and solution concentration, are equally crucial and significantly influence the characteristics and yield of PDVLNs [94]. Suresh et al. reported that PEG precipitation at pH values of 4 and 5 significantly increased the yield of PDVLNs [95]. The colloidal stability of VLNs of ginseng origin can be significantly enhanced by optimizing the preparation conditions and using a combinatorial approach when extracting the vesicles to obtain PDVLNs with the desired properties [96]. Therefore, the isolation method should be selected, and the preparation conditions should be optimized according to the plant materials and target properties to obtain high-quality PDVLNs samples. With the continuous development of nanotechnology, the purity and quality of nanoparticles are increasing. However, current isolation methods are unable to balance recovery and specificity. Therefore, new and efficient separation methods must be explored to achieve breakthroughs in the application of PDVLNs.
2.4. Characterization Methods for PDVLNs
The physical properties of PDVLNs are specific in two ways: 1. PDVLNs from different parts of the same plant may contain various particle subtypes with varying potentials. 2. Artificial factors, such as preparation and isolation methods and characterization instruments, may also lead to differences in the characterization of PDVLNs.
2.4.1. Particle Size Distribution of PDVLNs
Commonly employed techniques for assessing the particle size distribution of PDVLNs include optical microscopy (OM), dynamic light scattering (DLS), nanoparticle tracking analysis (NTA), and laser diffractometry (LD) [97]. Multiparametric surface labeling analysis with fluorescent dyes and antibodies facilitates the quantification of specific PDVLN subtypes. Moreover, both the NanoFCM Nanoanalyzer and Particle Metrix ZetaView Fluorescent Nanoparticle Tracking Analyzer were optimized for PDVLN labeling [98].
2.4.2. Morphology of PDVLNs
EM, a high-resolution imaging technique, is often used to explore the morphological features of PDVLNs. However, different imaging techniques can affect the morphology of PDVLNs; therefore, careful consideration is required when selecting an imaging method. Scanning electron microscopy (SEM) and transmission electron microscopy (TEM), two of the most prevalent EM techniques, require thorough sample fixation, dehydration, and staining, which may induce dehydration and deformation of vesicles, resulting in a cup-shaped morphology [99]. In low-temperature EM, PDVLNs typically exhibit a spherical morphology, suggesting that the cup-shaped structures observed via other EM techniques may result from dehydration. Additionally, EM enables precise characterization of PDVLN morphology and size and provides crucial insights into the mechanisms of their biogenesis. For example, Cai et al. observed the fusion of MVBs with the PM and the release of EVs via EM, supporting the presence of extracellular membrane structures in the plasma ectodomain [27].
2.4.3. Potentiation of PDVLNs
DLS and NTA provide information on the surface charge state of PDVLNs [100]. The zeta potential is a metric for evaluating the stability of nanoparticle colloids. Typically, nanoparticles with zeta potential values between -30 mV and +30 mV exhibit enhanced stability [101]. PDVLNs exhibit diverse ranges of ζ-potentials, indicating their ability to exist independently without aggregation. Furthermore, the surface charge of PDVLNs varies, ranging from nearly neutral to approximately -50 mV, potentially attributable to variations in the components present in PDVLNs sourced from different plants.
3. Mechanisms and Applications of PDVLNs in Tissue Repair and Regeneration
Tissue repair and regeneration are complex biological processes involving multiple cellular and molecular mechanisms. Plant-derived vesicle-like nanoparticles (PDVLNs) exhibit a range of biological functions—such as promoting cell proliferation, migration, differentiation, angiogenesis, immunomodulation, antioxidation, antiapoptosis, and microbiota regulation—that underpin their therapeutic potential across various disease models [102,103]. These functions facilitate intercellular communication, modulate recipient cell behavior, and create a microenvironment conducive to repair and regeneration [104,105]. Below, we systematically explore how these mechanisms translate into therapeutic applications, organized by specific tissue types and associated diseases.
3.1. Skin: Wound Healing and Facial Rejuvenation
Skin injuries, whether acute (e.g., trauma, burns) or chronic (e.g., diabetic ulcers), benefit from PDVLNs’ ability to regulate inflammation, promote cellular activity, and enhance vascularization. These nanoparticles promote cell proliferation, migration, and differentiation, key processes in skin r epair [106]. For instance, aloe saponin-derived VLNs (AS-DVLNs) stimulate human dermal fibroblast (HDF) proliferation, while ginseng-derived VLNs (Gs-DVLNs) enhance proliferation of human keratin-forming (HaCaT) cells and human umbilical vein endothelial cells (HUVECs) [107,108]. Wheat-derived VLNs simultaneously promote proliferation of HDFs, HUVECs, and HaCaT cells, underscoring their biocompatibility and potential natural targeting mechanisms [109]. Cell migration, another critical step, is enhanced by Aloe vera peel-derived VLNs (Avp-DVLNs), which accelerate HaCaT and HDF movement to promote wound closure, and by Gs-DVLNs, which support HaCaT and HUVEC migration [110,111].
Therapeutically, these functions translate into accelerated wound healing. In acute wounds, wheat- and ginseng-derived VLNs regulate HaCaT and HUVEC proliferation via ERK and Akt/mTOR pathways, attenuate inflammation, and promote angiogenesis in HUVECs, enhancing local microcirculation and nutrient delivery for re-epithelialization [108,109]. In chronic wounds, excessive matrix metalloproteinase (MMP) activity and reactive oxygen species (ROS) impede healing [112,113]. Grapefruit-derived VLNs (Gf-DVLNs) reduce oxidative stress in HaCaT cells, upregulate collagen type I and fibronectin expression, and promote proliferation and migration, expediting repair [113]. Avp-DVLNs activate the Nrf2 pathway, bolstering antioxidant defenses and enhancing migratory capacity, further supporting wound closure (Figure 3 and Figure 4) [114].
Panel I: ADVLNs doubled collagen protein levels, reduced integrin/adhesion molecule expression threef-old (A), decreased metalloproteinase expression (B), and doubled collagen chain expression (C).
Panel II: CDVLNs enhanced skin permeation, varying by formulation (A), and doubled penetration depth of lipophilic substitutes vs. buffer (B).
Panel III: ADVLNs exerted anti-inflammatory effects, downregulating NF-κB pathway genes (A, B) and IL-1 family factors/receptors (C), with KEGG analysis showing IL-1β/NF-κB pathway changes (D).
Panel IV: WDVLNs boosted proliferation and migration in HDFs, HUVECs, and HaCaT cells (A-C).
Beyond wound healing, PDVLNs address skin aging, characterized by chronic inflammation and extracellular matrix (ECM) degradation. Apple-derived VLNs (ADVLNs) downregulate the NF-κB proinflammatory pathway by inhibiting TLR4, reducing inflammation and increasing collagen synthesis (e.g., COL3A1, COL1A2) while suppressing MMPs (e.g., MMP1, MMP8), slowing ECM breakdown [115]. Wheat-derived VLNs boost type I collagen mRNA levels in fibroblasts, enhance proliferation and migration of endothelial, epithelial, and dermal cells, and suppress apoptosis, offering antiaging effects (see Figure 4) [108]. Unlike traditional antiaging products, PDVLNs’ natural composition minimizes allergic risks, and their small size enhances skin penetration, though further animal and clinical studies are needed.
3.2. Bone: Osteogenesis and Osteoporosis Treatment
Bone regeneration relies on cellular differentiation and proliferation, processes effectively promoted by plant-derived vesicle-like nanoparticles (PDVLNs). Yam-derived VLNs (YDVLNs) upregulate osteogenic markers such as osteopontin (OPN), alkaline phosphatase (ALP), and collagen type I (COL I) in ovariectomy-induced osteoporotic mice, facilitating bone regeneration [116]. Apple-derived VLNs (ADVLNs) enhance osteogenic gene and protein expression in MC3T3-E1 osteoblasts, supporting bone formation [117]. Tomato-derived VLNs increase chondrogenic markers and extracellular matrix (ECM) proteins in human adipose-derived mesenchymal stem cells (MSCs), outperforming lemon-derived VLNs and suggesting potential for cartilage repair in osteoarthritis [46].
Panel I: YDVLNs upregulate OPN, ALP, and COL I in osteoporotic mice, enhancing bone density (A-C).
Panel II: ADVLNs increase osteogenic gene expression in MC3T3-E1 cells (A), with elevated protein levels (B).
Panel III: Tomato-derived VLNs boost chondrogenic markers in MSCs, supporting cartilage repair (A-B).
Therapeutically, these mechanisms translate into treatments for bone-related disorders. YDVLNs’ induction of osteogenic differentiation supports their use in osteoporosis, restoring bone density in preclinical models (see Figure 5) [116]. ADVLNs’ effects on osteoblasts highlight their potential in fracture repair, while tomato-derived VLNs’ chondrogenic promotion offers a novel approach to osteoarthritis, addressing a condition with limited regenerative therapies. Additionally, Pueraria lobata-derived VLNs rebalance intestinal microbiota by metabolizing trimethylamine-N-oxide (TMAO), enhancing osteoblast differentiation and mineralization to alleviate osteoporosis, demonstrating a microbiota-mediated repair mechanism [118].
3.3. Nervous System: Neural Differentiation and Neuroprotection
Neural repair requires differentiation and protection against oxidative stress, both facilitated by PDVLNs [119]. Ginseng-derived VLNs (Gs-DVLNs) are internalized by bone marrow mesenchymal stem cells (BMSCs) in a time-dependent manner, delivering miRNAs enriched in neural-related signaling pathway genes [120]. This induces neural differentiation, evidenced by electrophysiological properties and neuron-like synaptic extensions, potentially via PI3K and transcriptional pathways, offering a novel strategy for stem cell-directed neural repair.
In neurodegenerative contexts, PDVLNs’ antioxidant and anti-inflammatory properties are critical. Blueberry-derived VLNs (Bb-DVLNs) inhibit ROS production and apoptosis in rotenone-treated HepG2 cells, a model relevant to Parkinson’s disease, where oxidative stress drives dopaminergic neuron loss [121]. Carrot-derived VLNs, rich in antioxidants, show promise for treating Parkinson’s and myocardial infarction by mitigating oxidative damage [122]. Gs-DVLNs’ miRNA delivery further supports their potential in neural regeneration, though specific uptake mechanisms by neural cells remain unclear [120].
3.4. Liver: Anti-Inflammatory and Antioxidant Effects
Liver repair benefits from PDVLNs’ immunomodulatory, anti-inflammatory, and antioxidant functions. Shiitake mushroom- and honey-derived VLNs inhibit NLRP3 inflammasome activation, reducing inflammation and promoting liver tissue repair [64,123]. Ginger-derived VLNs (Gg-DVLNs) similarly suppress NLRP3 activity, laying a foundation for treating Alzheimer’s and type 2 diabetes, conditions linked to liver dysfunction. Antioxidant effects are evident in Bb-DVLNs and Gg-DVLNs, which regulate Nrf2 distribution to mitigate hepatocyte apoptosis [124].
Therapeutically, these properties address liver injury. Lemon-derived VLNs enhance Lactobacillus rhamnosus GG (LGG) bile tolerance, upregulating LGG to support liver repair, demonstrating microbiota regulation’s role in regeneration [125]. Gg-DVLNs’ anti-inflammatory and antioxidant effects further position them as candidates for liver disease management, though optimal dosing requires further study.
3.5. Gastrointestinal Tract: Inflammation Control and Mucosal Repair
In the gastrointestinal (GI) tract, PDVLNs regulate inflammation and microbiota to promote repair. Ginger-derived VLNs (Gg-DVLNs) suppress NLRP3 inflammasome activity, reducing inflammation linked to inflammatory bowel disease (IBD) [126]. Blueberry-derived VLNs (Bb-DVLNs) modulate NF-κB and TLR4 expression in Caco-1 cells, downregulating IL-8β and IL-5 to mitigate intestinal inflammation [127]. Gg-DVLNs also increase probiotic Lactobacillus rhamnosus GG (LGG13A) populations, elevating IL-22 via the aryl hydrocarbon receptor (AHR) pathway to enhance antimicrobial immunity and mucosal repair at barrier surfaces [46].
Therapeutically, these mechanisms support GI repair (see Figure 6). Gg-DVLNs’ anti-inflammatory and microbiota-modulating effects offer potential for IBD treatment, while Bb-DVLNs’ regulatory actions suggest broader applications in intestinal inflammation. By addressing dysbiosis-related damage, PDVLNs provide a promising approach to mucosal regeneration, though further studies on dosing and specificity are needed.
3.6. Cardiovascular System: Antioxidant and Angiogenic Support
Cardiovascular repair hinges on angiogenesis and oxidative stress mitigation, both enhanced by PDVLNs. AS-DVLNs promote angiogenesis in HUVECs, supporting nutrient delivery in skin wound healing, a mechanism applicable to ischemic tissues [107,128]. Carrot-derived VLNs’ antioxidant capacity counters myocardial hypoxia/reoxygenation-induced oxidative stress, protecting against cardiac hypertrophy and heart failure [122]. CDVLNs inhibit Nrf2 reduction in cardiomyocytes, further safeguarding against oxidative damage.
Therapeutically, these effects position PDVLNs as candidates for cardiovascular diseases. Carrot-derived VLNs’ high antioxidant yield suggests potential in myocardial infarction treatment, while their angiogenic properties could enhance revascularization in ischemic conditions, warranting further preclinical validation.
3.7. Dental Tissue: Periodontal Repair
PDVLNs contribute to periodontal repair through anti-inflammatory and microbiota-regulating functions, addressing damage in oral tissues. Ginger-derived VLNs (Gg-DVLNs) specifically target Porphyromonas gingivalis, suppressing virulence factor expression, which reduces inflammation and bone loss in periodontal tissues. Their immunomodulatory effects, such as decreasing proinflammatory cytokines (e.g., IL-6, IL-7) and increasing anti-inflammatory IL-10, further support tissue repair [129,130]. Additionally, Gg-DVLNs enhance probiotic populations like LGG13A, promoting a balanced oral microbiota conducive to regeneration [131].
Gg-DVLNs inhibit Porphyromonas gingivalis virulence, reducing inflammation and bone loss, while enhancing IL-10 and LGG13A levels to support periodontal tissue repair.
Therapeutically, Gg-DVLNs show promise in treating periodontal disease by reducing inflammation and supporting alveolar bone preservation (see Figure 7). Their ability to mitigate oxidative stress and regulate microbial dysbiosis positions them as a novel alternative to conventional therapies, though clinical validation is pending.
Table 2.
Role of PDVLNs in promoting tissue repair and regeneration in different tissue and disease models.
Table 2.
Role of PDVLNs in promoting tissue repair and regeneration in different tissue and disease models.
| Tissue | Source | Method of administration | In vivo or in vitro models | Potential mechanism | Key findings | Main ingredients | Reference |
|---|---|---|---|---|---|---|---|
| Skin | Aloe-saponaria | co-incubation | In vitro: HDFs, HUVECs | Proliferation and migration of HDFs, Angiogenesis of HUVECs, Angiogenesis of HUVECs, Antioxidant | Promotes chronic wounds healing | unknown | [132] |
| Ginseng | co-incubation | In vitro: HaCaT, HUVECs, BJ | Enhanced cell migration and angiogenesis, Increased secretion of extracellular matrix proteins, Regulation of proliferation and inflammatory response via ERK and Akt/mTOR pathways | Promotes wound healing | ginsenoside | [109] | |
| Wheat | co-incubation | In vitro: HDFs, HUVECs, HaCaT | Enhancement of wound healing-related gene expression; activation of fibroblast function, coordination of vascularization process | Promotes wound healing | unknown | [108] | |
| BFR | co-incubation | In vitro: HDFs | Down-regulation of pro-inflammatory cytokines; Promotion of HDF proliferation and migration | Promotes wound healing | unknown | [133] | |
| Aloe vera Peels | co-incubation | In vitro: HaCaT, HDF | Decreased levels of ROS within HaCaT cells, Increased migration capacity of cells, Increased expression of mRNA for Nrf2, HO-1, CAT, and SOD genes | Promotes wound healing | unknown | [114] | |
| Grapefruit | co-incubation | In vitro: HaCaT, HUVEC | Reduction of ROS levels in HaCaT cells, Enhancement of proliferation- and migration-related gene expression in HaCaT, Enhancement of HUVEC cell tube formation | Promotes chronic wound healing | unknown | [134] | |
| Apple | co-incubation | In vitro: HDF | Inhibition of TLR4 activity, Down-regulation of NF-κB pro-inflammatory pathway, Enhancement of collagen synthesis, Inhibition of metalloproteinase production | Anti-aging of the skin | unknown | [115] | |
| Liver | Shiitake mushroom | none | In vivo: mouse model of acute liver injury (GaiN, LPS induced) | Inhibition of NLRP3 inflammatory vesicle activation, Decrease in IL-6 activity | Combating FHF; preventing GaIn/LPS-induced acute liver injury | unknown | [135] |
| Ginger | oral | In vivo: mouse model of alcoholic liver disease | Activation of Nrf2, Triggering of hepatic detoxification/antioxidant gene expression, Inhibition of ROS production | Protective effect against alcoholic liver injury in mice | shogaol | [136] | |
| Garlic | Oral or intravenous administration | In vivo: mice with acute liver injury | Inhibition of NLRP3 inflammatory vesicle activation pathway, Reduction in cysteinyl asparagin-1 autocleavage, inhibition of cytokine release and pyroptosis cell death in primary macrophages | Attenuating inflammation in chemically induced acute liver injury | DLPC | [123] | |
| Honey | none | In vivo: mice with acute liver injury | Inhibition of NLRP3 inflammatory vesicle activation, Reduction of IL-1β, IL-3, IL-6 and TNFα levels | Amelioration of inflammation and liver injury in acute liver injury | MiR-4057 | [65] | |
| Blueberry | intragastric administration | In vitro: HepG2 In vivo: male C57BL/6 mice (6-8 weeks) |
Acceleration of Nrf2 translocation, Decrease in AST and ALT levels, Improvement of insulin resistance, Inhibition of FAS and ACC1 expression | Treatment of NAFLD | unknown | [121] | |
| Lemon | co-incubation | In vitro: LGG | Inhibition of Msp1 and Msp3 production by RNase P-mediated degradation of specific tRNAs, Improvement of LGG tolerance to bile, Increase in LGG percentage | Regulates LGG to promote liver tissue repair | unknown | [125] | |
| Bone | Plum | co-incubation | In vitro: MC3T3-E1 cells, mouse bone marrow primary osteoblasts | Modulation of BMP-2/MAPK/Smad-1 pathway | Promotes osteoblast activation, Reduces osteoclast differentiation | unknown | [137] |
| Ginseng | co-incubation | In vitro: bone marrow-derived osteoclasts | Inhibits RANKL-induced signaling pathways, Regulates osteoclast maturation genes, Inhibits osteoclast differentiation | Anti-osteoporosis | ginsenoside | [138] | |
| Yam | oral | In vivo: mouse model of osteoporosis | Activates BMP-2/p-p38-dependent Runx2 pathway, Increases bone differentiation markers. | Promotes bone regeneration, Promotes osteoblast differentiation and mineralization | unknown | [116] | |
| Apple | none | In vitro: MC3T3-E1 | Regulates BMP3/Smad3 pathway, Activates ERK and JNK-related signaling, Promotes osteoblast growth and differentiation. | Anti-osteoporosis | unknown | [117] | |
| Pueraria lobata | co-incubation | In vitro: hBMSC In vivo: Ovariectomy (OVX)-induced osteoporosis in rats |
Promotes hBMSC differentiation and mineralization, Degradation of TMAO promotes autophagy. | Promotes bone formation and reduces bone resorption. | unknown | [118] | |
| Colon | Grapefruit | oral | In vivo: DSS-induced colitis model in mouse | Up-regulation of HO-1 expression, Inhibition of IL-1β and TNF-α production in intestinal macrophages | Anti-inflammatory, immune-modulatory, maintenance of intestinal macrophage homeostasis | unknown | [139] |
| Ginger | oral | In vivo: colitis model in mouse | Increased survival and proliferation of IECs, Decreased pro-inflammatory cytokines, Increased anti-inflammatory cytokines | Prevents and protects against IBD | 6-gingerol | [49] | |
| Ginger | oral | In vitro: Lactobacillus rhamnosus (LGG) In vivo: mouse model |
Activation of the AHR pathway, Induction of IL-22 expression | LGG-mediated inhibition of colitis in mice | unknown | [140] | |
| Mulberry bark | oral | In vitro: MC38 and human Caco2 colon cancer cell lines In vivo: mouse colitis model |
Promoting HSPA8-mediated activation of the AhR signaling pathway and anti-inflammatory via AhR-COPS8 | Prevention of colitis in mice | HSPA8 | [141] | |
| Grape | oral | In vivo: rat | Modulating stem cell microenvironment, Promotes proliferation of Lgr5+ intestinal stem cells |
Prevention of colitis in rats | unknown | [142] | |
| Grape | oral | In vivo: DSS-induced mouse colitis model | Induction of intestinal stem cell growth factor gene expression through the Wnt/β-catenin signaling pathway, Promotes proliferation of Lgr5+ intestinal stem cells | Promotes self-renewal of intestinal epithelium and accelerates the recovery of intestinal structure | unknown | [143] | |
| Tea leaves | oral | In vivo: mouse colitis model | Reduces ROS production, Inhibits pro-inflammatory cytokines, and Increases macrophage secretion of anti-inflammatory cytokines | Prevention of colitis-related colon cancer, antioxidant | galactose (CH2O6) | [144] | |
| Broccoli | oral | In vivo: mouse colitis model | Inhibition of pro-inflammatory cytokines and activation of AMPK are involved in inhibiting DC pro-inflammatory factor activation | Preventing DC activation and inducing tolerance to DC | unknown | [145] | |
| Turmeric | oral | In vitro: primitive 264.7 murine macrophages, NCM 460 and HT-29 colonic epithelial cells In vivo: ICR mice |
Inhibits pro-inflammatory cytokine expression, promotes antioxidant gene levels, regulates gut microbiota composition and abundance, and remodels the immune microenvironment | Anti-inflammatory, antioxidant, relieves symptoms of intestinal inflammation | unknown | [146] | |
| Carrot | oral | In vitro: intestinal macrophages and stem cells. In vivo: B6.Cg-Tg(BAT-lacZ)3Picc/J mice |
Promotion of Nrf2 nuclear translocation, induction of anti-inflammatory cytokines, antioxidant and activation of Wnt signaling gene expression | Anti-inflammatory, antioxidant, maintains intestinal homeostasis | unknown | [31] | |
| Pomegranate | co-incubation | In vitro: THP-1, Caco-2 cells | Improved Caco-2 cell survival at 5 ug/ml | Anti-inflammatory, antioxidant, promotes healing of intestinal epithelium | unknown | [147] | |
| Nervous system | Garlic | oral | In vivo: mouse high-fat diet model | Inhibition of cGAS/STING/IDO1/AHR inflammatory signaling cascade response | Alleviating brain inflammation and obesity in mice | PA(36:4) | [148] |
| Momordica charantia | none | In vivo: MCAO model in male SD rats | Up-regulation of AKT/GSK3β signaling pathway, inhibition of MMP-9, attenuation of BBB injury, and inhibition of neuronal apoptosis | Improving neurologic dysfunction in cerebral ischemia-reperfusion injury | Possibly related to miR5266 | [149] | |
| Cardiovascular system | M charantia | intraperitoneal injection | In vitro: H9C2 cardiomyocytes In vivo: thoracic-irradiated 5-6 week old BALB/c nude mice |
Promotes cell proliferation, inhibits apoptosis, attenuates DNA damage, scavenges mitochondrial ROS, and regulates p-AKT/AKT and p-ERK/ERK ratios | Reduces myocardial damage and fibrosis and is potentially protective against radiation-induced heart disease | unknown | [150] |
| Citrus limon (L.) Osbeck | sublingual administration | In vivo: healthy participants | Effect on waist circumference and LDL-C | Cardiometabolic risk factors for management of normolipidemic participants | unknown | [151] | |
| Carrot | co-incubation | In vitro: H2C5 cardiomyoblasts and SH-SY2Y cells | Inhibits decreased expression of antioxidant molecules, protects cells from oxidative stress, and inhibits ROS production and ROS-induced apoptosis | Novel drug candidates for treatment of myocardial infarction and Parkinson’s disease | unknown | [122] | |
| Oral cavity | Ginger | oral | In vitro: Porphyromonas gingivalis surface In vivo: mouse model of chronic periodontitis |
Acts with heme-binding protein 35 (HBP35) to inhibit the pathogenicity of Porphyromonas gingivalis, reduce the expression of anti-inflammatory factors in periodontal tissues, and reduce alveolar bone loss | Prevention/treatment of chronic periodontitis | PA(36:2) | [129] |
4. Therapeutic Advantages and Comparisons of PDVLNs in Regenerative Medicine
Plant-derived vesicle-like nanoparticles (PDVLNs) offer a compelling alternative to conventional regenerative therapies due to their unique biological and practical advantages [152]. Unlike stem cell therapies, which carry risks of tumorigenesis and immune rejection, PDVLNs provide a cell-free approach with low immunogenicity and minimal ethical concerns, stemming from their plant origin [153]. Their sustainability—bypassing animal-derived sources or embryo use—further enhances their appeal over mammalian exosomes, which, while effective in tissue repair , pose risks of disease transmission and immune activation [154]. For example, ginger-derived VLNs (Gg-DVLNs) suppress NLRP3 inflammasome activity without eliciting adverse responses , a feat less consistently achieved with mammalian counterparts [155].
Compared to synthetic nanoparticles, PDVLNs possess intrinsic bioactivity from their diverse cargos—lipids, proteins, and miRNAs—eliminating the need for extensive synthetic loading. Plant-derived compounds like quercetin or 6-gingerol exhibit pharmacological effects (e.g., antioxidant, neurogenic [156,157]) but suffer from poor solubility and bioavailability. PDVLNs overcome these barriers by encapsulating such bioactives within vesicular structures, as seen with apple-derived VLNs (ADVLNs) enhancing collagen synthesis in skin aging models [115]. Table 2 illustrates PDVLNs’ multifaceted roles across tissues—promoting angiogenesis in skin wounds, osteogenesis in bone defects, and neuroprotection in neural models—highlighting their superiority over single-target agents in addressing complex diseases.
This combinatorial efficacy, coupled with biocompatibility and scalability, positions PDVLNs as a promising platform in regenerative medicine. However, their full potential hinges on overcoming limitations in standardization and mechanistic understanding, as explored in subsequent sections.
5. Engineering PDVLNs for Enhanced Tissue Repair and Regeneration
To maximize PDVLNs’ regenerative potential, engineering strategies can enhance their stability, specificity, and delivery efficiency, building on their natural advantages (Section 4). Surface modification, inspired by mammalian exosome engineering, can improve targeting precision. [158] For instance, conjugating PDVLNs with antibodies against periodontal pathogens like Porphyromonas gingivalis [Figure 7] could amplify Gg-DVLNs’ efficacy in dental repair (Table 2) [129,130,131]. Lipid reconstitution—reassembling PDVLNs with synthetic lipids—enhances membrane stability and uptake, as demonstrated by ginger-derived liposomes retaining anti-inflammatory activity [49].
Cargo enrichment offers another avenue, enabling PDVLNs to deliver tailored therapeutics. Loading with miRNAs (e.g., miR-162a for osteogenesis [159] and miRNAs for intestinal tract [160]) or small molecules (e.g., quercetin [161]) could bolster effects like Gs-DVLNs’ neural differentiation in BMSCs [63]. Controlled release systems, such as encapsulation in hydrogels [162], could sustain delivery in chronic conditions like diabetic ulcers or osteoporosis, extending PDVLNs’ impact beyond natural kinetics. Bioinspired synthesis—mimicking biogenesis pathways like MVB fusion (Section 2.1)—promises scalable production, with reconstituted grape-derived VLNs maintaining bioactivity [45].
These approaches, while transformative, must preserve PDVLNs’ low immunogenicity and biocompatibility. Advances in nanotechnology and bioengineering, paired with deeper insights into uptake mechanisms, will be pivotal to translating engineered PDVLNs into clinical tools, addressing limitations outlined in the next section.
6. Challenges and Future Directions in PDVLN-Based Regenerative Medicine
Despite PDVLNs’ therapeutic promise (Section 3, Section 4 and Section 5), significant challenges impede their clinical translation. Biogenesis mechanisms—e.g., MVB fusion, EXPO secretion (Section 2.1)—remain incompletely understood, obscuring cargo sorting and artificial synthesis potential. Uptake by mammalian cells, whether receptor-mediated or passive, is equally unclear, complicating biodistribution predictions. Standardization is a critical hurdle; variability in composition across plant sources (Section 2.2) and isolation methods (Section 2.3) undermines reproducibility, with techniques like ultracentrifugation struggling to balance yield and purity (Table 1). Preservation methods also lag, risking stability under storage or physiological conditions.
Therapeutically, PDVLNs lack the adaptability of stem cells, limiting responsiveness to dynamic pathologies. Preclinical efficacy (Table 2) in models like wound healing and neurodegeneration awaits clinical validation, with safety, dosing, and regulatory pathways undefined. Addressing these requires integrated solutions. Multi-omics (e.g., lipidomics, proteomics) and advanced imaging (e.g., cryo-EM [Section 2.4]) could decode biogenesis and uptake, guiding engineering (Section 5). Hybrid isolation protocols (e.g., ultrafiltration plus SEC) and stabilization techniques (e.g., lyophilization) may standardize production, while engineered PDVLNs—targeted or cargo-enriched—could overcome therapeutic limitations.
Looking forward, PDVLNs’ multitarget capabilities position them for combination therapies against complex diseases (e.g., IBD, osteoporosis). Interdisciplinary collaboration—spanning bioengineering, nanotechnology, and clinical research—will be essential to realize this vision, establishing PDVLNs as a cornerstone of regenerative medicine.
Author Contributions
QW designed the content and wrote the first draft; QW, ZH, JG and ZT produced the figures; YH, MH and HL produced the table; and QW, MW, WC and YM revised the manuscript. BJ was responsible for supervision of the entire project as the corresponding author.
Funding
This work was supported by Horizontal topics of the Dental Hospital of Southern Medical University (NYKQ-JG(HX)-202405, NYKQ-JG(HX)-202402, NYKQ-JG(HX)-202401, 2023ZDZX2009, PY2022015).
Ethics Approval
Not applicable.
Consent to Participate
Not applicable.
Consent for Publication
Not applicable.
Data Availability Statement
Not applicable.
Code Availability
Not applicable.
Conflicts of Interest
References
- Mostafa, H. K. K. Different Cells of the Human Body: Categories and Morphological Characters. J. Microsc. Ultrastruct. 2022, 10(2), 40. [Google Scholar] [CrossRef] [PubMed]
- Şeker, Ş.; Elçin, A. E.; Elçin, Y. M. Advances in Regenerative Medicine and Biomaterials. In Gene, Drug, and Tissue Engineering; Pereira, G. C., Ed.; Methods in Molecular Biology; Springer US: New York, NY, 2023; Vol. 2575, pp. 127–152. [Google Scholar] [CrossRef]
- Farley, A.; McLafferty, E.; Hendry, C. Cells, Tissues, Organs and Systems. Nurs. Stand. R. Coll. Nurs. G. B. 1987 2012, 26(52), 40–45. [Google Scholar] [CrossRef]
- Matai, I.; Kaur, G.; Seyedsalehi, A.; McClinton, A.; Laurencin, C. T. Progress in 3D Bioprinting Technology for Tissue/Organ Regenerative Engineering. Biomaterials 2020, 226, 119536. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ma, X.-J.-N.; Fei, Y.-Y.; Han, H.-T.; Xu, J.; Cheng, L.; Li, X. Stem Cell Therapy in Liver Regeneration: Focus on Mesenchymal Stem Cells and Induced Pluripotent Stem Cells. Pharmacol. Ther. 2022, 232, 108004. [Google Scholar] [CrossRef]
- Zhang, D.; Wei, G.; Li, P.; Zhou, X.; Zhang, Y. Urine-Derived Stem Cells: A Novel and Versatile Progenitor Source for Cell-Based Therapy and Regenerative Medicine. Genes Dis. 2014, 1(1), 8–17. [Google Scholar] [CrossRef]
- Tumorigenicity as a clinical hurdle for pluripotent stem cell therapies | Nature Medicine. Available online: https://www.nature.com/articles/nm.3267 (accessed on 17 March 2025).
- L. , P. K.; Kandoi, S.; Misra, R.; S., V.; K., R.; Verma, R. S. The Mesenchymal Stem Cell Secretome: A New Paradigm towards Cell-Free Therapeutic Mode in Regenerative Medicine. Cytokine Growth Factor Rev. 2019, 46, 1–9. [Google Scholar] [CrossRef]
- Welsh, J. A.; Goberdhan, D. C. I.; O’Driscoll, L.; Buzas, E. I.; Blenkiron, C.; Bussolati, B.; Cai, H.; Di Vizio, D.; Driedonks, T. A. P.; Erdbrügger, U.; Falcon-Perez, J. M.; Fu, Q.-L.; Hill, A. F.; Lenassi, M.; Lim, S. K.; Mahoney, M. G.; Mohanty, S.; Möller, A.; Nieuwland, R.; Ochiya, T.; Sahoo, S.; Torrecilhas, A. C.; Zheng, L.; Zijlstra, A.; Abuelreich, S.; Bagabas, R.; Bergese, P.; Bridges, E. M.; Brucale, M.; Burger, D.; Carney, R. P.; Cocucci, E.; Crescitelli, R.; Hanser, E.; Harris, A. L.; Haughey, N. J.; Hendrix, A.; Ivanov, A. R.; Jovanovic-Talisman, T.; Kruh-Garcia, N. A.; Ku’ulei-Lyn Faustino, V.; Kyburz, D.; Lässer, C.; Lennon, K. M.; Lötvall, J.; Maddox, A. L.; Martens-Uzunova, E. S.; Mizenko, R. R.; Newman, L. A.; Ridolfi, A.; Rohde, E.; Rojalin, T.; Rowland, A.; Saftics, A.; Sandau, U. S.; Saugstad, J. A.; Shekari, F.; Swift, S.; Ter-Ovanesyan, D.; Tosar, J. P.; Useckaite, Z.; Valle, F.; Varga, Z.; van der Pol, E.; van Herwijnen, M. J. C.; Wauben, M. H. M.; Wehman, A. M.; Williams, S.; Zendrini, A.; Zimmerman, A. J.; MISEV Consortium; Théry, C.; Witwer, K. W. Minimal Information for Studies of Extracellular Vesicles (MISEV2023): From Basic to Advanced Approaches. J. Extracell. Vesicles 2024, 13(2), e12404. [Google Scholar] [CrossRef]
- Nam, G.-H.; Choi, Y.; Kim, G. B.; Kim, S.; Kim, S. A.; Kim, I.-S. Emerging Prospects of Exosomes for Cancer Treatment: From Conventional Therapy to Immunotherapy. Adv. Mater. Deerfield Beach Fla 2020, 32(51), e2002440. [Google Scholar] [CrossRef]
- Vizoso, F. J.; Eiro, N.; Cid, S.; Schneider, J.; Perez-Fernandez, R. Mesenchymal Stem Cell Secretome: Toward Cell-Free Therapeutic Strategies in Regenerative Medicine. Int. J. Mol. Sci. 2017, 18(9), 1852. [Google Scholar] [CrossRef]
- Roomi, S.; Masi, A.; Conselvan, G. B.; Trevisan, S.; Quaggiotti, S.; Pivato, M.; Arrigoni, G.; Yasmin, T.; Carletti, P. Protein Profiling of Arabidopsis Roots Treated With Humic Substances: Insights Into the Metabolic and Interactome Networks. Front. Plant Sci. 2018, 9, 1812. [Google Scholar] [CrossRef]
- Qiao, S. A.; Roth, R. Messenger and Message: Uncovering the Roles, Rhythm and Regulation of Extracellular Vesicles in Plant Biotic Interactions. Curr. Opin. Plant Biol. 2025, 83, 102672. [Google Scholar] [CrossRef] [PubMed]
- Al-Masawa, M. E.; Alshawsh, M. A.; Ng, C. Y.; Ng, A. M. H.; Foo, J. B.; Vijakumaran, U.; Subramaniam, R.; Ghani, N. A. A.; Witwer, K. W.; Law, J. X. Efficacy and Safety of Small Extracellular Vesicle Interventions in Wound Healing and Skin Regeneration: A Systematic Review and Meta-Analysis of Animal Studies. Theranostics 2022, 12(15), 6455–6508. [Google Scholar] [CrossRef] [PubMed]
- Cong, M.; Tan, S.; Li, S.; Gao, L.; Huang, L.; Zhang, H.-G.; Qiao, H. Technology Insight: Plant-Derived Vesicles-How Far from the Clinical Biotherapeutics and Therapeutic Drug Carriers? Adv. Drug Deliv. Rev. 2022, 182, 114108. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Liu, K. Plant-Derived Extracellular Vesicles as Oral Drug Delivery Carriers. J. Control. Release Off. J. Control. Release Soc. 2022, 350, 389–400. [Google Scholar] [CrossRef]
- Samadi, P.; Sheykhhasan, M.; Mokhtari, K.; Yang, P.; Maghool, F.; Kalhor, N. Chapter 13 - Extracellular Vesicles: Unlocking Therapeutic Potential in Regenerative Medicine. In Extracellular Vesicles for Therapeutic and Diagnostic Applications; Anand, K., Vadivalagan, C., Gangadaran, P., Muthu, S., Peacock, B., Eds.; Nanotechnology in Biomedicine; Elsevier, 2025; pp 397–435. [CrossRef]
- Halperin, W.; Jensen, W. A. Ultrastructural Changes during Growth and Embryogenesis in Carrot Cell Cultures. J. Ultrastruct. Res. 1967, 18(3), 428–443. [Google Scholar] [CrossRef]
- Kalra, H.; Drummen, G. P. C.; Mathivanan, S. Focus on Extracellular Vesicles: Introducing the Next Small Big Thing. Int. J. Mol. Sci. 2016, 17(2), 170. [Google Scholar] [CrossRef]
- Scheuring, D.; Viotti, C.; Krüger, F.; Künzl, F.; Sturm, S.; Bubeck, J.; Hillmer, S.; Frigerio, L.; Robinson, D. G.; Pimpl, P.; Schumacher, K. Multivesicular Bodies Mature from the Trans-Golgi Network/Early Endosome in Arabidopsis. Plant Cell 2011, 23(9), 3463–3481. [Google Scholar] [CrossRef]
- Ung, T. H.; Madsen, H. J.; Hellwinkel, J. E.; Lencioni, A. M.; Graner, M. W. Exosome Proteomics Reveals Transcriptional Regulator Proteins with Potential to Mediate Downstream Pathways. Cancer Sci. 2014, 105(11), 1384–1392. [Google Scholar] [CrossRef]
- Cui, Y.; Zhao, Q.; Hu, S.; Jiang, L. Vacuole Biogenesis in Plants: How Many Vacuoles, How Many Models? Trends Plant Sci. 2020, 25(6), 538–548. [Google Scholar] [CrossRef]
- Ho, P. T. B.; Clark, I. M.; Le, L. T. T. MicroRNA-Based Diagnosis and Therapy. Int. J. Mol. Sci. 2022, 23(13), 7167. [Google Scholar] [CrossRef]
- Poulsen, C. P.; Dilokpimol, A.; Mouille, G.; Burow, M.; Geshi, N. Arabinogalactan Glycosyltransferases Target to a Unique Subcellular Compartment That May Function in Unconventional Secretion in Plants. Traffic Cph. Den. 2014, 15(11), 1219–1234. [Google Scholar] [CrossRef] [PubMed]
- Gayral, M.; Arias Gaguancela, O.; Vasquez, E.; Herath, V.; Flores, F. J.; Dickman, M. B.; Verchot, J. Multiple ER-to-Nucleus Stress Signaling Pathways Are Activated during Plantago Asiatica Mosaic Virus and Turnip Mosaic Virus Infection in Arabidopsis Thaliana. Plant J. Cell Mol. Biol. 2020, 103(3), 1233–1245. [Google Scholar] [CrossRef] [PubMed]
- Movahed, N.; Cabanillas, D. G.; Wan, J.; Vali, H.; Laliberté, J.-F.; Zheng, H. Turnip Mosaic Virus Components Are Released into the Extracellular Space by Vesicles in Infected Leaves. Plant Physiol. 2019, 180(3), 1375–1388. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Qiao, L.; Wang, M.; He, B.; Lin, F.-M.; Palmquist, J.; Huang, S.-D.; Jin, H. Plants Send Small RNAs in Extracellular Vesicles to Fungal Pathogen to Silence Virulence Genes. Science 2018, 360(6393), 1126–1129. [Google Scholar] [CrossRef]
- Wang, F.; Shang, Y.; Fan, B.; Yu, J.-Q.; Chen, Z. Arabidopsis LIP5, a Positive Regulator of Multivesicular Body Biogenesis, Is a Critical Target of Pathogen-Responsive MAPK Cascade in Plant Basal Defense. PLoS Pathog. 2014, 10(7), e1004243. [Google Scholar] [CrossRef]
- Cui, Y.; Shen, J.; Gao, C.; Zhuang, X.; Wang, J.; Jiang, L. Biogenesis of Plant Prevacuolar Multivesicular Bodies. Mol. Plant 2016, 9(6), 774–786. [Google Scholar] [CrossRef]
- Mu, J.; Zhuang, X.; Wang, Q.; Jiang, H.; Deng, Z.-B.; Wang, B.; Zhang, L.; Kakar, S.; Jun, Y.; Miller, D.; Zhang, H.-G. Interspecies Communication between Plant and Mouse Gut Host Cells through Edible Plant Derived Exosome-like Nanoparticles. Mol. Nutr. Food Res. 2014, 58(7), 1561–1573. [Google Scholar] [CrossRef]
- Maricchiolo, E.; Panfili, E.; Pompa, A.; De Marchis, F.; Bellucci, M.; Pallotta, M. T. Unconventional Pathways of Protein Secretion: Mammals vs. Plants. Front. Cell Dev. Biol. 2022, 10, 895853. [Google Scholar] [CrossRef]
- Bruns, C.; McCaffery, J. M.; Curwin, A. J.; Duran, J. M.; Malhotra, V. Biogenesis of a Novel Compartment for Autophagosome-Mediated Unconventional Protein Secretion. J. Cell Biol. 2011, 195(6), 979–992. [Google Scholar] [CrossRef]
- Gu, Y.; Innes, R. W. The KEEP ON GOING Protein of Arabidopsis Regulates Intracellular Protein Trafficking and Is Degraded during Fungal Infection. Plant Cell 2012, 24(11), 4717–4730. [Google Scholar] [CrossRef]
- Ito, Y.; Taniguchi, K.; Kuranaga, Y.; Eid, N.; Inomata, Y.; Lee, S.-W.; Uchiyama, K. Uptake of MicroRNAs from Exosome-Like Nanovesicles of Edible Plant Juice by Rat Enterocytes. Int. J. Mol. Sci. 2021, 22(7), 3749. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ding, Y.; Wang, J.; Hillmer, S.; Miao, Y.; Lo, S. W.; Wang, X.; Robinson, D. G.; Jiang, L. EXPO, an Exocyst-Positive Organelle Distinct from Multivesicular Endosomes and Autophagosomes, Mediates Cytosol to Cell Wall Exocytosis in Arabidopsis and Tobacco Cells. Plant Cell 2010, 22(12), 4009–4030. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D. G.; Ding, Y.; Jiang, L. Unconventional Protein Secretion in Plants: A Critical Assessment. Protoplasma 2016, 253(1), 31–43. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Bermúdez, P.; Blesa, J.; Soriano, J. M.; Marcilla, A. Extracellular Vesicles in Food: Experimental Evidence of Their Secretion in Grape Fruits. Eur. J. Pharm. Sci. Off. J. Eur. Fed. Pharm. Sci. 2017, 98, 40–50. [Google Scholar] [CrossRef]
- Cai, Q.; He, B.; Wang, S.; Fletcher, S.; Niu, D.; Mitter, N.; Birch, P. R. J.; Jin, H. Message in a Bubble: Shuttling Small RNAs and Proteins Between Cells and Interacting Organisms Using Extracellular Vesicles. Annu. Rev. Plant Biol. 2021, 72, 497–524. [Google Scholar] [CrossRef]
- Jeppesen, D. K.; Fenix, A. M.; Franklin, J. L.; Higginbotham, J. N.; Zhang, Q.; Zimmerman, L. J.; Liebler, D. C.; Ping, J.; Liu, Q.; Evans, R.; Fissell, W. H.; Patton, J. G.; Rome, L. H.; Burnette, D. T.; Coffey, R. J. Reassessment of Exosome Composition. Cell 2019, 177(2), 428–445.e18. [Google Scholar] [CrossRef]
- Alfieri, M.; Leone, A.; Ambrosone, A. Plant-Derived Nano and Microvesicles for Human Health and Therapeutic Potential in Nanomedicine. Pharmaceutics 2021, 13(4), 498. [Google Scholar] [CrossRef]
- Zhou, S.; Cao, Y.; Shan, F.; Huang, P.; Yang, Y.; Liu, S. Analyses of Chemical Components and Their Functions in Single Species Plant-Derived Exosome like Vesicle. TrAC Trends Anal. Chem. 2023, 167, 117274. [Google Scholar] [CrossRef]
- Liu, N.-J.; Wang, N.; Bao, J.-J.; Zhu, H.-X.; Wang, L.-J.; Chen, X.-Y. Lipidomic Analysis Reveals the Importance of GIPCs in Arabidopsis Leaf Extracellular Vesicles. Mol. Plant 2020, 13(10), 1523–1532. [Google Scholar] [CrossRef]
- Yang, C.; Zhang, M.; Merlin, D. Advances in Plant-Derived Edible Nanoparticle-Based Lipid Nano-Drug Delivery Systems as Therapeutic Nanomedicines. J. Mater. Chem. B 2018, 6(9), 1312–1321. [Google Scholar] [CrossRef]
- Laulagnier, K.; Motta, C.; Hamdi, S.; Roy, S.; Fauvelle, F.; Pageaux, J.-F.; Kobayashi, T.; Salles, J.-P.; Perret, B.; Bonnerot, C.; Record, M. Mast Cell- and Dendritic Cell-Derived Exosomes Display a Specific Lipid Composition and an Unusual Membrane Organization. Biochem. J. 2004, 380 Pt 1, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Record, M. Exosome-like Nanoparticles from Food: Protective Nanoshuttles for Bioactive Cargo. Mol. Ther. J. Am. Soc. Gene Ther. 2013, 21(7), 1294–1296. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Viennois, E.; Prasad, M.; Zhang, Y.; Wang, L.; Zhang, Z.; Han, M. K.; Xiao, B.; Xu, C.; Srinivasan, S.; Merlin, D. Edible Ginger-Derived Nanoparticles: A Novel Therapeutic Approach for the Prevention and Treatment of Inflammatory Bowel Disease and Colitis-Associated Cancer. Biomaterials 2016, 101, 321–340. [Google Scholar] [CrossRef] [PubMed]
- Szatanek, R.; Baj-Krzyworzeka, M.; Zimoch, J.; Lekka, M.; Siedlar, M.; Baran, J. The Methods of Choice for Extracellular Vesicles (EVs) Characterization. Int. J. Mol. Sci. 2017, 18(6), 1153. [Google Scholar] [CrossRef]
- Chen, X.; Zhou, Y.; Yu, J. Exosome-like Nanoparticles from Ginger Rhizomes Inhibited NLRP3 Inflammasome Activation. Mol. Pharm. 2019, 16(6), 2690–2699. [Google Scholar] [CrossRef]
- Goul, C.; Peruzzo, R.; Zoncu, R. The Molecular Basis of Nutrient Sensing and Signalling by mTORC1 in Metabolism Regulation and Disease. Nat. Rev. Mol. Cell Biol. 2023, 24(12), 857–875. [Google Scholar] [CrossRef]
- Kumar, A.; Sundaram, K.; Teng, Y.; Mu, J.; Sriwastva, M. K.; Zhang, L.; Hood, J. L.; Yan, J.; Zhang, X.; Park, J. W.; Merchant, M. L.; Zhang, H.-G. Ginger Nanoparticles Mediated Induction of Foxa2 Prevents High-Fat Diet-Induced Insulin Resistance. Theranostics 2022, 12(3), 1388–1403. [Google Scholar] [CrossRef]
- Xu, F.; Mu, J.; Teng, Y.; Zhang, X.; Sundaram, K.; Sriwastva, M. K.; Kumar, A.; Lei, C.; Zhang, L.; Liu, Q. M.; Yan, J.; McClain, C. J.; Merchant, M. L.; Zhang, H.-G. Restoring Oat Nanoparticles Mediated Brain Memory Function of Mice Fed Alcohol by Sorting Inflammatory Dectin-1 Complex Into Microglial Exosomes. Small Weinh. Bergstr. Ger. 2022, 18(6), e2105385. [Google Scholar] [CrossRef]
- Hu, X.-L.; Lu, H.; Hassan, M. M.; Zhang, J.; Yuan, G.; Abraham, P. E.; Shrestha, H. K.; Villalobos Solis, M. I.; Chen, J.-G.; Tschaplinski, T. J.; Doktycz, M. J.; Tuskan, G. A.; Cheng, Z.-M. M.; Yang, X. Advances and Perspectives in Discovery and Functional Analysis of Small Secreted Proteins in Plants. Hortic. Res. 2021, 8(1), 130. [Google Scholar] [CrossRef]
- Wang, X.; Chung, K. P.; Lin, W.; Jiang, L. Protein Secretion in Plants: Conventional and Unconventional Pathways and New Techniques. J. Exp. Bot. 2017, 69(1), 21–37. [Google Scholar] [CrossRef]
- Tang, Y.-T.; Huang, Y.-Y.; Zheng, L.; Qin, S.-H.; Xu, X.-P.; An, T.-X.; Xu, Y.; Wu, Y.-S.; Hu, X.-M.; Ping, B.-H.; Wang, Q. Comparison of Isolation Methods of Exosomes and Exosomal RNA from Cell Culture Medium and Serum. Int. J. Mol. Med. 2017, 40(3), 834–844. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wang, S.; Cai, Q.; Jin, H. Effective Methods for Isolation and Purification of Extracellular Vesicles from Plants. J. Integr. Plant Biol. 2021, 63(12), 2020–2030. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Lin, J.; Ye, J.; Wang, R.; Yang, C.; Gong, J.; Liu, Y.; Deng, C.; Liu, P.; Chen, C.; Cheng, Y.; Deng, X.; Zeng, Y. A Comprehensive Proteomic Analysis of Elaioplasts from Citrus Fruits Reveals Insights into Elaioplast Biogenesis and Function. Hortic. Res. 2018, 5, 6. [Google Scholar] [CrossRef] [PubMed]
- Kim, S. Q.; Kim, K.-H. Emergence of Edible Plant-Derived Nanovesicles as Functional Food Components and Nanocarriers for Therapeutics Delivery: Potentials in Human Health and Disease. Cells 2022, 11(14), 2232. [Google Scholar] [CrossRef]
- Ryu, H.; Fuwad, A.; Yoon, S.; Jang, H.; Lee, J. C.; Kim, S. M.; Jeon, T.-J. Biomimetic Membranes with Transmembrane Proteins: State-of-the-Art in Transmembrane Protein Applications. Int. J. Mol. Sci. 2019, 20(6), 1437. [Google Scholar] [CrossRef]
- Baldrich, P.; Rutter, B. D.; Karimi, H. Z.; Podicheti, R.; Meyers, B. C.; Innes, R. W. Plant Extracellular Vesicles Contain Diverse Small RNA Species and Are Enriched in 10- to 17-Nucleotide “Tiny” RNAs. Plant Cell 2019, 31(2), 315–324. [Google Scholar] [CrossRef]
- Perut, F.; Roncuzzi, L.; Avnet, S.; Massa, A.; Zini, N.; Sabbadini, S.; Giampieri, F.; Mezzetti, B.; Baldini, N. Strawberry-Derived Exosome-Like Nanoparticles Prevent Oxidative Stress in Human Mesenchymal Stromal Cells. Biomolecules 2021, 11(1), 87. [Google Scholar] [CrossRef]
- Yin, L.; Yan, L.; Yu, Q.; Wang, J.; Liu, C.; Wang, L.; Zheng, L. Characterization of the MicroRNA Profile of Ginger Exosome-like Nanoparticles and Their Anti-Inflammatory Effects in Intestinal Caco-2 Cells. J. Agric. Food Chem. 2022, 70(15), 4725–4734. [Google Scholar] [CrossRef]
- Wu, S.; Yang, J.; Xu, H.; Wang, X.; Zhang, R.; Lu, W.; Yang, J.; Li, X.; Chen, S.; Zou, Y.; Nan, A. Circular RNA circGLIS3 Promotes Bladder Cancer Proliferation via the miR-1273f/SKP1/Cyclin D1 Axis. Cell Biol. Toxicol. 2022, 38(1), 129–146. [Google Scholar] [CrossRef]
- Veerapandian, M.; Ramasundaram, S.; Jerome, P.; Chellasamy, G.; Govindaraju, S.; Yun, K.; Oh, T. H. Drug Delivery Application of Functional Nanomaterials Synthesized Using Natural Sources. J. Funct. Biomater. 2023, 14(8), 426. [Google Scholar] [CrossRef]
- Chen, X.; Liu, B.; Li, X.; An, T. T.; Zhou, Y.; Li, G.; Wu-Smart, J.; Alvarez, S.; Naldrett, M. J.; Eudy, J.; Kubik, G.; Wilson, R. A.; Kachman, S. D.; Cui, J.; Yu, J. Identification of Anti-Inflammatory Vesicle-like Nanoparticles in Honey. J. Extracell. Vesicles 2021, 10(4), e12069. [Google Scholar] [CrossRef] [PubMed]
- Saiyed, A. N.; Vasavada, A. R.; Johar, S. R. K. Recent Trends in miRNA Therapeutics and the Application of Plant miRNA for Prevention and Treatment of Human Diseases. Future J. Pharm. Sci. 2022, 8(1), 24. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Liang, Z.; Xu, J.; Zhao, Y.; Li, X.; Zhang, Y.; Zhao, D.; Chen, R.; Liu, Y.; Joshi, T.; Chang, J.; Wang, Z.; Zhang, Y.; Zhu, J.; Liu, Q.; Xu, D.; Jiang, C. Plant-Derived Phosphocholine Facilitates Cellular Uptake of Anti-Pulmonary Fibrotic HJT-sRNA-M7. Sci. China Life Sci. 2019, 62(3), 309–320. [Google Scholar] [CrossRef]
- Li, X.; Liang, Z.; Du, J.; Wang, Z.; Mei, S.; Li, Z.; Zhao, Y.; Zhao, D.; Ma, Y.; Ye, J.; Xu, J.; Zhao, Y.; Chang, J.; Qin, Y.; Yu, L.; Wang, C.; Jiang, C. Herbal Decoctosome Is a Novel Form of Medicine. Sci. China Life Sci. 2019, 62(3), 333–348. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K. W.; Aikawa, E.; Alcaraz, M. J.; Anderson, J. D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G. K.; Ayre, D. C.; Bach, J.-M.; Bachurski, D.; Baharvand, H.; Balaj, L.; Baldacchino, S.; Bauer, N. N.; Baxter, A. A.; Bebawy, M.; Beckham, C.; Bedina Zavec, A.; Benmoussa, A.; Berardi, A. C.; Bergese, P.; Bielska, E.; Blenkiron, C.; Bobis-Wozowicz, S.; Boilard, E.; Boireau, W.; Bongiovanni, A.; Borràs, F. E.; Bosch, S.; Boulanger, C. M.; Breakefield, X.; Breglio, A. M.; Brennan, M. Á.; Brigstock, D. R.; Brisson, A.; Broekman, M. L.; Bromberg, J. F.; Bryl-Górecka, P.; Buch, S.; Buck, A. H.; Burger, D.; Busatto, S.; Buschmann, D.; Bussolati, B.; Buzás, E. I.; Byrd, J. B.; Camussi, G.; Carter, D. R.; Caruso, S.; Chamley, L. W.; Chang, Y.-T.; Chen, C.; Chen, S.; Cheng, L.; Chin, A. R.; Clayton, A.; Clerici, S. P.; Cocks, A.; Cocucci, E.; Coffey, R. J.; Cordeiro-da-Silva, A.; Couch, Y.; Coumans, F. A.; Coyle, B.; Crescitelli, R.; Criado, M. F.; D’Souza-Schorey, C.; Das, S.; Datta Chaudhuri, A.; de Candia, P.; De Santana Junior, E. F.; De Wever, O.; del Portillo, H. A.; Demaret, T.; Deville, S.; Devitt, A.; Dhondt, B.; Di Vizio, D.; Dieterich, L. C.; Dolo, V.; Dominguez Rubio, A. P.; Dominici, M.; Dourado, M. R.; Driedonks, T. A.; Duarte, F. V.; Duncan, H. M.; Eichenberger, R. M.; Ekström, K.; EL Andaloussi, S.; Elie-Caille, C.; Erdbrügger, U.; Falcón-Pérez, J. M.; Fatima, F.; Fish, J. E.; Flores-Bellver, M.; Försönits, A.; Frelet-Barrand, A.; Fricke, F.; Fuhrmann, G.; Gabrielsson, S.; Gámez-Valero, A.; Gardiner, C.; Gärtner, K.; Gaudin, R.; Gho, Y. S.; Giebel, B.; Gilbert, C.; Gimona, M.; Giusti, I.; Goberdhan, D. C.; Görgens, A.; Gorski, S. M.; Greening, D. W.; Gross, J. C.; Gualerzi, A.; Gupta, G. N.; Gustafson, D.; Handberg, A.; Haraszti, R. A.; Harrison, P.; Hegyesi, H.; Hendrix, A.; Hill, A. F.; Hochberg, F. H.; Hoffmann, K. F.; Holder, B.; Holthofer, H.; Hosseinkhani, B.; Hu, G.; Huang, Y.; Huber, V.; Hunt, S.; Ibrahim, A. G.-E.; Ikezu, T.; Inal, J. M.; Isin, M.; Ivanova, A.; Jackson, H. K.; Jacobsen, S.; Jay, S. M.; Jayachandran, M.; Jenster, G.; Jiang, L.; Johnson, S. M.; Jones, J. C.; Jong, A.; Jovanovic-Talisman, T.; Jung, S.; Kalluri, R.; Kano, S.; Kaur, S.; Kawamura, Y.; Keller, E. T.; Khamari, D.; Khomyakova, E.; Khvorova, A.; Kierulf, P.; Kim, K. P.; Kislinger, T.; Klingeborn, M.; Klinke II, D. J.; Kornek, M.; Kosanović, M. M.; Kovács, Á. F.; Krämer-Albers, E.-M.; Krasemann, S.; Krause, M.; Kurochkin, I. V.; Kusuma, G. D.; Kuypers, S.; Laitinen, S.; Langevin, S. M.; Languino, L. R.; Lannigan, J.; Lässer, C.; Laurent, L. C.; Lavieu, G.; Lázaro-Ibáñez, E.; Le Lay, S.; Lee, M.-S.; Lee, Y. X. F.; Lemos, D. S.; Lenassi, M.; Leszczynska, A.; Li, I. T.; Liao, K.; Libregts, S. F.; Ligeti, E.; Lim, R.; Lim, S. K.; Linē, A.; Linnemannstöns, K.; Llorente, A.; Lombard, C. A.; Lorenowicz, M. J.; Lörincz, Á. M.; Lötvall, J.; Lovett, J.; Lowry, M. C.; Loyer, X.; Lu, Q.; Lukomska, B.; Lunavat, T. R.; Maas, S. L.; Malhi, H.; Marcilla, A.; Mariani, J.; Mariscal, J.; Martens-Uzunova, E. S.; Martin-Jaular, L.; Martinez, M. C.; Martins, V. R.; Mathieu, M.; Mathivanan, S.; Maugeri, M.; McGinnis, L. K.; McVey, M. J.; Meckes Jr, D. G.; Meehan, K. L.; Mertens, I.; Minciacchi, V. R.; Möller, A.; Møller Jørgensen, M.; Morales-Kastresana, A.; Morhayim, J.; Mullier, F.; Muraca, M.; Musante, L.; Mussack, V.; Muth, D. C.; Myburgh, K. H.; Najrana, T.; Nawaz, M.; Nazarenko, I.; Nejsum, P.; Neri, C.; Neri, T.; Nieuwland, R.; Nimrichter, L.; Nolan, J. P.; Nolte-’t Hoen, E. N.; Noren Hooten, N.; O’Driscoll, L.; O’Grady, T.; O’Loghlen, A.; Ochiya, T.; Olivier, M.; Ortiz, A.; Ortiz, L. A.; Osteikoetxea, X.; Østergaard, O.; Ostrowski, M.; Park, J.; Pegtel, D. M.; Peinado, H.; Perut, F.; Pfaffl, M. W.; Phinney, D. G.; Pieters, B. C.; Pink, R. C.; Pisetsky, D. S.; Pogge von Strandmann, E.; Polakovicova, I.; Poon, I. K.; Powell, B. H.; Prada, I.; Pulliam, L.; Quesenberry, P.; Radeghieri, A.; Raffai, R. L.; Raimondo, S.; Rak, J.; Ramirez, M. I.; Raposo, G.; Rayyan, M. S.; Regev-Rudzki, N.; Ricklefs, F. L.; Robbins, P. D.; Roberts, D. D.; Rodrigues, S. C.; Rohde, E.; Rome, S.; Rouschop, K. M.; Rughetti, A.; Russell, A. E.; Saá, P.; Sahoo, S.; Salas-Huenuleo, E.; Sánchez, C.; Saugstad, J. A.; Saul, M. J.; Schiffelers, R. M.; Schneider, R.; Schøyen, T. H.; Scott, A.; Shahaj, E.; Sharma, S.; Shatnyeva, O.; Shekari, F.; Shelke, G. V.; Shetty, A. K.; Shiba, K.; Siljander, P. R.-M.; Silva, A. M.; Skowronek, A.; Snyder II, O. L.; Soares, R. P.; Sódar, B. W.; Soekmadji, C.; Sotillo, J.; Stahl, P. D.; Stoorvogel, W.; Stott, S. L.; Strasser, E. F.; Swift, S.; Tahara, H.; Tewari, M.; Timms, K.; Tiwari, S.; Tixeira, R.; Tkach, M.; Toh, W. S.; Tomasini, R.; Torrecilhas, A. C.; Tosar, J. P.; Toxavidis, V.; Urbanelli, L.; Vader, P.; van Balkom, B. W.; van der Grein, S. G.; Van Deun, J.; van Herwijnen, M. J.; Van Keuren-Jensen, K.; van Niel, G.; van Royen, M. E.; van Wijnen, A. J.; Vasconcelos, M. H.; Vechetti Jr, I. J.; Veit, T. D.; Vella, L. J.; Velot, É.; Verweij, F. J.; Vestad, B.; Viñas, J. L.; Visnovitz, T.; Vukman, K. V.; Wahlgren, J.; Watson, D. C.; Wauben, M. H.; Weaver, A.; Webber, J. P.; Weber, V.; Wehman, A. M.; Weiss, D. J.; Welsh, J. A.; Wendt, S.; Wheelock, A. M.; Wiener, Z.; Witte, L.; Wolfram, J.; Xagorari, A.; Xander, P.; Xu, J.; Yan, X.; Yáñez-Mó, M.; Yin, H.; Yuana, Y.; Zappulli, V.; Zarubova, J.; Žėkas, V.; Zhang, J.; Zhao, Z.; Zheng, L.; Zheutlin, A. R.; Zickler, A. M.; Zimmermann, P.; Zivkovic, A. M.; Zocco, D.; Zuba-Surma, E. K. Minimal Information for Studies of Extracellular Vesicles 2018 (MISEV2018): A Position Statement of the International Society for Extracellular Vesicles and Update of the MISEV2014 Guidelines. J. Extracell. Vesicles 2018, 7(1), 1535750. [Google Scholar] [CrossRef]
- Jia, Y.; Yu, L.; Ma, T.; Xu, W.; Qian, H.; Sun, Y.; Shi, H. Small Extracellular Vesicles Isolation and Separation: Current Techniques, Pending Questions and Clinical Applications. Theranostics 2022, 12(15), 6548–6575. [Google Scholar] [CrossRef]
- Bobrie, A.; Colombo, M.; Krumeich, S.; Raposo, G.; Théry, C. Diverse Subpopulations of Vesicles Secreted by Different Intracellular Mechanisms Are Present in Exosome Preparations Obtained by Differential Ultracentrifugation. J. Extracell. Vesicles 2012, 1. [Google Scholar] [CrossRef]
- Mathivanan, S.; Fahner, C. J.; Reid, G. E.; Simpson, R. J. ExoCarta 2012: Database of Exosomal Proteins, RNA and Lipids. Nucleic Acids Res. 2012, 40(Database issue), D1241–1244. [Google Scholar] [CrossRef]
- Ali, N. B.; Abdull Razis, A. F.; Ooi, D. J.; Chan, K. W.; Ismail, N.; Foo, J. B. Theragnostic Applications of Mammal and Plant-Derived Extracellular Vesicles: Latest Findings, Current Technologies, and Prospects. Mol. Basel Switz. 2022, 27(12), 3941. [Google Scholar] [CrossRef]
- Zou, Z.; Li, H.; Xu, G.; Hu, Y.; Zhang, W.; Tian, K. Current Knowledge and Future Perspectives of Exosomes as Nanocarriers in Diagnosis and Treatment of Diseases. Int. J. Nanomedicine 2023, 18, 4751–4778. [Google Scholar] [CrossRef]
- Greening, D. W.; Xu, R.; Ji, H.; Tauro, B. J.; Simpson, R. J. A Protocol for Exosome Isolation and Characterization: Evaluation of Ultracentrifugation, Density-Gradient Separation, and Immunoaffinity Capture Methods. Methods Mol. Biol. Clifton NJ 2015, 1295, 179–209. [Google Scholar] [CrossRef]
- Pertoft, H. Fractionation of Cells and Subcellular Particles with Percoll. J. Biochem. Biophys. Methods 2000, 44(1–2), 1–30. [Google Scholar] [CrossRef] [PubMed]
- Brakke, M. K. Density Gradient Centrifugation: A New Separation Technique1. ACS Publications. [CrossRef]
- Buschmann, D.; Mussack, V.; Byrd, J. B. Separation, Characterization, and Standardization of Extracellular Vesicles for Drug Delivery Applications. Adv. Drug Deliv. Rev. 2021, 174, 348–368. [Google Scholar] [CrossRef]
- Lobb, R. J.; Becker, M.; Wen, S. W.; Wong, C. S. F.; Wiegmans, A. P.; Leimgruber, A.; Möller, A. Optimized Exosome Isolation Protocol for Cell Culture Supernatant and Human Plasma. J. Extracell. Vesicles 2015, 4, 27031. [Google Scholar] [CrossRef]
- Xu, D.; Di, K.; Fan, B.; Wu, J.; Gu, X.; Sun, Y.; Khan, A.; Li, P.; Li, Z. MicroRNAs in Extracellular Vesicles: Sorting Mechanisms, Diagnostic Value, Isolation, and Detection Technology. Front. Bioeng. Biotechnol. 2022, 10, 948959. [Google Scholar] [CrossRef]
- Alzhrani, G. N.; Alanazi, S. T.; Alsharif, S. Y.; Albalawi, A. M.; Alsharif, A. A.; Abdel-Maksoud, M. S.; Elsherbiny, N. Exosomes: Isolation, Characterization, and Biomedical Applications. Cell Biol. Int. 2021, 45(9), 1807–1831. [Google Scholar] [CrossRef]
- Contreras-Naranjo, J. C.; Wu, H.-J.; Ugaz, V. M. Microfluidics for Exosome Isolation and Analysis: Enabling Liquid Biopsy for Personalized Medicine. Lab. Chip 2017, 17(21), 3558–3577. [Google Scholar] [CrossRef]
- Zhong, Y.; Wang, X.; Zhao, X.; Shen, J.; Wu, X.; Gao, P.; Yang, P.; Chen, J.; An, W. Multifunctional Milk-Derived Small Extracellular Vesicles and Their Biomedical Applications. Pharmaceutics 2023, 15(5), 1418. [Google Scholar] [CrossRef]
- Garzo, E.; Sánchez-López, C. M.; Fereres, A.; Soler, C.; Marcilla, A.; Pérez-Bermúdez, P. Isolation of Extracellular Vesicles from Phloem Sap by Size Exclusion Chromatography. Curr. Protoc. 2023, 3(10), e903. [Google Scholar] [CrossRef]
- Monguió-Tortajada, M.; Gálvez-Montón, C.; Bayes-Genis, A.; Roura, S.; Borràs, F. E. Extracellular Vesicle Isolation Methods: Rising Impact of Size-Exclusion Chromatography. Cell. Mol. Life Sci. CMLS 2019, 76(12), 2369–2382. [Google Scholar] [CrossRef]
- Martínez-Greene, J. A.; Hernández-Ortega, K.; Quiroz-Baez, R.; Resendis-Antonio, O.; Pichardo-Casas, I.; Sinclair, D. A.; Budnik, B.; Hidalgo-Miranda, A.; Uribe-Querol, E.; Ramos-Godínez, M. D. P.; Martínez-Martínez, E. Quantitative Proteomic Analysis of Extracellular Vesicle Subgroups Isolated by an Optimized Method Combining Polymer-Based Precipitation and Size Exclusion Chromatography. J. Extracell. Vesicles 2021, 10(6), e12087. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Lyden, D. Asymmetric-Flow Field-Flow Fractionation Technology for Exomere and Small Extracellular Vesicle Separation and Characterization. Nat. Protoc. 2019, 14(4), 1027–1053. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Cai, Q.; Qiao, L.; Huang, C.-Y.; Wang, S.; Miao, W.; Ha, T.; Wang, Y.; Jin, H. RNA-Binding Proteins Contribute to Small RNA Loading in Plant Extracellular Vesicles. Nat. Plants 2021, 7(3), 342–352. [Google Scholar] [CrossRef]
- Liangsupree, T.; Multia, E.; Riekkola, M.-L. Modern Isolation and Separation Techniques for Extracellular Vesicles. J. Chromatogr. A 2021, 1636, 461773. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; He, B.; Jin, H. Isolation of Extracellular Vesicles from Arabidopsis. Curr. Protoc. 2022, 2(1), e352. [Google Scholar] [CrossRef]
- Ackermann, B. L. Understanding the Role of Immunoaffinity-Based Mass Spectrometry Methods for Clinical Applications. Clin. Chem. 2012, 58(12), 1620–1622. [Google Scholar] [CrossRef]
- Mondal, S. K.; Whiteside, T. L. Immunoaffinity-Based Isolation of Melanoma Cell-Derived and T Cell-Derived Exosomes from Plasma of Melanoma Patients. Methods Mol. Biol. Clifton NJ 2021, 2265, 305–321. [Google Scholar] [CrossRef]
- Liu, W.-Z.; Ma, Z.-J.; Kang, X.-W. Current Status and Outlook of Advances in Exosome Isolation. Anal. Bioanal. Chem. 2022, 414(24), 7123–7141. [Google Scholar] [CrossRef]
- He, B.; Cai, Q.; Qiao, L.; Huang, C.-Y.; Wang, S.; Miao, W.; Ha, T.; Wang, Y.; Jin, H. RNA-Binding Proteins Contribute to Small RNA Loading in Plant Extracellular Vesicles. Nat. Plants 2021, 7(3), 342–352. [Google Scholar] [CrossRef]
- Staubach, S.; Bauer, F. N.; Tertel, T.; Börger, V.; Stambouli, O.; Salzig, D.; Giebel, B. Scaled Preparation of Extracellular Vesicles from Conditioned Media. Adv. Drug Deliv. Rev. 2021, 177, 113940. [Google Scholar] [CrossRef]
- Suresh, A. P.; Kalarikkal, S. P.; Pullareddy, B.; Sundaram, G. M. Low pH-Based Method to Increase the Yield of Plant-Derived Nanoparticles from Fresh Ginger Rhizomes. ACS Omega 2021, 6(27), 17635–17641. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.; Jeong, H.; Jang, E.; Kim, E.; Yoon, Y.; Jang, S.; Jeong, H.-S.; Jang, G. Isolation of High-Purity and High-Stability Exosomes from Ginseng. Front. Plant Sci. 2022, 13, 1064412. [Google Scholar] [CrossRef] [PubMed]
- Filipe, V.; Hawe, A.; Jiskoot, W. Critical Evaluation of Nanoparticle Tracking Analysis (NTA) by NanoSight for the Measurement of Nanoparticles and Protein Aggregates. Pharm. Res. 2010, 27(5), 796–810. [Google Scholar] [CrossRef]
- Fortunato, D.; Mladenović, D.; Criscuoli, M.; Loria, F.; Veiman, K.-L.; Zocco, D.; Koort, K.; Zarovni, N. Opportunities and Pitfalls of Fluorescent Labeling Methodologies for Extracellular Vesicle Profiling on High-Resolution Single-Particle Platforms. Int. J. Mol. Sci. 2021, 22(19), 10510. [Google Scholar] [CrossRef]
- Sharma, S.; Gillespie, B. M.; Palanisamy, V.; Gimzewski, J. K. Quantitative Nanostructural and Single-Molecule Force Spectroscopy Biomolecular Analysis of Human-Saliva-Derived Exosomes. Langmuir ACS J. Surf. Colloids 2011, 27(23), 14394–14400. [Google Scholar] [CrossRef]
- Koliqi, R.; Breznica, P.; Daka, A.; Koshi, B. Application of Design of Expert Software for Evaluating the Influence of Formulation Variables on the Encapsulation Efficacy, Drug Content and Particle Size of PEO-PPO-PEO/Poly (DL-Lactide-Co-Caprolactone) Nanoparticles as Carriers for SN-38. Med. Pharm. Rep. 2021, 94(4), 483–497. [Google Scholar] [CrossRef]
- Bommakanti, V.; Banerjee, M.; Shah, D.; Manisha, K.; Sri, K.; Banerjee, S. An Overview of Synthesis, Characterization, Applications and Associated Adverse Effects of Bioactive Nanoparticles. Environ. Res. 2022, 214 Pt 2, 113919. [Google Scholar] [CrossRef]
- Nagelkerke, A.; Ojansivu, M.; van der Koog, L.; Whittaker, T. E.; Cunnane, E. M.; Silva, A. M.; Dekker, N.; Stevens, M. M. Extracellular Vesicles for Tissue Repair and Regeneration: Evidence, Challenges and Opportunities. Adv. Drug Deliv. Rev. 2021, 175, 113775. [Google Scholar] [CrossRef]
- Chen, F.-M.; Wu, L.-A.; Zhang, M.; Zhang, R.; Sun, H.-H. Homing of Endogenous Stem/Progenitor Cells for in Situ Tissue Regeneration: Promises, Strategies, and Translational Perspectives. Biomaterials 2011, 32(12), 3189–3209. [Google Scholar] [CrossRef]
- Jahani, M.; Rezazadeh, D.; Mohammadi, P.; Abdolmaleki, A.; Norooznezhad, A.; Mansouri, K. Regenerative Medicine and Angiogenesis; Challenges and Opportunities. Adv. Pharm. Bull. 2020, 10(4), 490–501. [Google Scholar] [CrossRef]
- Ding, J.-Y.; Chen, M.-J.; Wu, L.-F.; Shu, G.-F.; Fang, S.-J.; Li, Z.-Y.; Chu, X.-R.; Li, X.-K.; Wang, Z.-G.; Ji, J.-S. Mesenchymal Stem Cell-Derived Extracellular Vesicles in Skin Wound Healing: Roles, Opportunities and Challenges. Mil. Med. Res. 2023, 10(1), 36. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Liang, J.; Liu, H.; Zhang, K.; Li, J. Nanomaterials for Angiogenesis in Skin Tissue Engineering. Tissue Eng. Part B Rev. 2020, 26(3), 203–216. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Tao, R.; Chen, L.; Xiong, Y.; Xue, H.; Hu, L.; Yan, C.; Xie, X.; Lin, Z.; Panayi, A. C.; Mi, B.; Liu, G. Exosomes Derived from Pioglitazone-Pretreated MSCs Accelerate Diabetic Wound Healing through Enhancing Angiogenesis. J. Nanobiotechnology 2021, 19(1), 150. [Google Scholar] [CrossRef]
- Şahin, F.; Koçak, P.; Güneş, M. Y.; Özkan, İ.; Yıldırım, E.; Kala, E. Y. In Vitro Wound Healing Activity of Wheat-Derived Nanovesicles. Appl. Biochem. Biotechnol. 2019, 188(2), 381–394. [Google Scholar] [CrossRef]
- Yang, S.; Lu, S.; Ren, L.; Bian, S.; Zhao, D.; Liu, M.; Wang, J. Ginseng-Derived Nanoparticles Induce Skin Cell Proliferation and Promote Wound Healing. J. Ginseng Res. 2023, 47(1), 133–143. [Google Scholar] [CrossRef]
- Hu, Y.; Tao, R.; Chen, L.; Xiong, Y.; Xue, H.; Hu, L.; Yan, C.; Xie, X.; Lin, Z.; Panayi, A. C.; Mi, B.; Liu, G. Exosomes Derived from Pioglitazone-Pretreated MSCs Accelerate Diabetic Wound Healing through Enhancing Angiogenesis. J. Nanobiotechnology 2021, 19(1), 150. [Google Scholar] [CrossRef]
- Yang, S.; Lu, S.; Ren, L.; Bian, S.; Zhao, D.; Liu, M.; Wang, J. Ginseng-Derived Nanoparticles Induce Skin Cell Proliferation and Promote Wound Healing. J. Ginseng Res. 2023, 47(1), 133–143. [Google Scholar] [CrossRef]
- Slavkovsky, R.; Kohlerova, R.; Tkacova, V.; Jiroutova, A.; Tahmazoglu, B.; Velebny, V.; Rezačová, M.; Sobotka, L.; Kanta, J. Zucker Diabetic Fatty Rat: A New Model of Impaired Cutaneous Wound Repair with Type II Diabetes Mellitus and Obesity. Wound Repair Regen. Off. Publ. Wound Heal. Soc. Eur. Tissue Repair Soc. 2011, 19(4), 515–525. [Google Scholar] [CrossRef]
- Lobmann, R.; Ambrosch, A.; Schultz, G.; Waldmann, K.; Schiweck, S.; Lehnert, H. Expression of Matrix-Metalloproteinases and Their Inhibitors in the Wounds of Diabetic and Non-Diabetic Patients. Diabetologia 2002, 45(7), 1011–1016. [Google Scholar] [CrossRef]
- Kim, M. K.; Choi, Y. C.; Cho, S. H.; Choi, J. S.; Cho, Y. W. The Antioxidant Effect of Small Extracellular Vesicles Derived from Aloe Vera Peels for Wound Healing. Tissue Eng. Regen. Med. 2021, 18(4), 561–571. [Google Scholar] [CrossRef]
- Trentini, M.; Zanolla, I.; Zanotti, F.; Tiengo, E.; Licastro, D.; Dal Monego, S.; Lovatti, L.; Zavan, B. Apple Derived Exosomes Improve Collagen Type I Production and Decrease MMPs during Aging of the Skin through Downregulation of the NF-κB Pathway as Mode of Action. Cells 2022, 11(24), 3950. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.-H.; Park, Y.-S.; Kim, H.-S.; Kim, D.; Lee, S.-H.; Lee, C.-H.; Lee, S.-H.; Kim, J.-E.; Lee, S.; Kim, H. M.; Kim, H.-W.; Kim, J.; Seo, W.; Kwon, H.-J.; Song, B.-J.; Kim, D.-K.; Baek, M.-C.; Cho, Y.-E. Yam-Derived Exosome-like Nanovesicles Stimulate Osteoblast Formation and Prevent Osteoporosis in Mice. J. Controlled Release 2023, 355, 184–198. [Google Scholar] [CrossRef] [PubMed]
- Sim, Y.; Seo, H.-J.; Kim, D.-H.; Lee, S.-H.; Kwon, J.; Kwun, I.-S.; Jung, C.; Kim, J.-I.; Lim, J.-H.; Kim, D.-K.; Baek, M.-C.; Cho, Y.-E. The Effect of Apple-Derived Nanovesicles on the Osteoblastogenesis of Osteoblastic MC3T3-E1 Cells. J. Med. Food 2023, 26(1), 49–58. [Google Scholar] [CrossRef] [PubMed]
- Zhan, W.; Deng, M.; Huang, X.; Xie, D.; Gao, X.; Chen, J.; Shi, Z.; Lu, J.; Lin, H.; Li, P. Pueraria Lobata-Derived Exosome-like Nanovesicles Alleviate Osteoporosis by Enhacning Autophagy. J. Control. Release Off. J. Control. Release Soc. 2023, 364, 644–653. [Google Scholar] [CrossRef]
- Yu, Y.; Xu, Z.; Xu, L.; Lu, D.; Tang, Y.; Mai, H. Plant Extracellular Vesicles as Emerging Neuroprotective Agents for Central Nervous System Disorders. J. Adv. Res. 2025. [Google Scholar] [CrossRef]
- Xu, X.-H.; Yuan, T.-J.; Dad, H. A.; Shi, M.-Y.; Huang, Y.-Y.; Jiang, Z.-H.; Peng, L.-H. Plant Exosomes As Novel Nanoplatforms for MicroRNA Transfer Stimulate Neural Differentiation of Stem Cells In Vitro and In Vivo. Nano Lett. 2021, 21(19), 8151–8159. [Google Scholar] [CrossRef]
- Zhao, W.-J.; Bian, Y.-P.; Wang, Q.-H.; Yin, F.; Yin, L.; Zhang, Y.-L.; Liu, J.-H. Blueberry-Derived Exosomes-like Nanoparticles Ameliorate Nonalcoholic Fatty Liver Disease by Attenuating Mitochondrial Oxidative Stress. Acta Pharmacol. Sin. 2022, 43(3), 645–658. [Google Scholar] [CrossRef]
- Kim, D. K.; Rhee, W. J. Antioxidative Effects of Carrot-Derived Nanovesicles in Cardiomyoblast and Neuroblastoma Cells. Pharmaceutics 2021, 13(8), 1203. [Google Scholar] [CrossRef]
- Liu, B.; Li, X.; Yu, H.; Shi, X.; Zhou, Y.; Alvarez, S.; Naldrett, M. J.; Kachman, S. D.; Ro, S.-H.; Sun, X.; Chung, S.; Jing, L.; Yu, J. Therapeutic Potential of Garlic Chive-Derived Vesicle-like Nanoparticles in NLRP3 Inflammasome-Mediated Inflammatory Diseases. Theranostics 2021, 11(19), 9311–9330. [Google Scholar] [CrossRef]
- He, X.-Y.; Gao, Z.-Y.; Liang, W.; Sun, Y.-C. Ameliorative Effect of Ginsenoside Rg1 on Dextran Sulfate Sodium-Induced Colitis: Involvement of Intestinal Barrier Remodeling in Mice. Ann. Transl. Med. 2022, 10(24), 1328. [Google Scholar] [CrossRef]
- Lei, C.; Teng, Y.; He, L.; Sayed, M.; Mu, J.; Xu, F.; Zhang, X.; Kumar, A.; Sundaram, K.; Sriwastva, M. K.; Zhang, L.; Chen, S.-Y.; Feng, W.; Zhang, S.; Yan, J.; Park, J. W.; Merchant, M. L.; Zhang, X.; Zhang, H.-G. Lemon Exosome-like Nanoparticles Enhance Stress Survival of Gut Bacteria by RNase P-Mediated Specific tRNA Decay. iScience 2021, 24(6), 102511. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Luo, T.; Wang, D.; Zhao, Y.; Jin, Y.; Yang, G.; Zhang, X. Therapeutic Application and Potential Mechanism of Plant-Derived Extracellular Vesicles in Inflammatory Bowel Disease. J. Adv. Res. 2025, 68, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Leng, Y.; Yang, L.; Zhu, H.; Li, D.; Pan, S.; Yuan, F. Stability of Blueberry Extracellular Vesicles and Their Gene Regulation Effects in Intestinal Caco-2 Cells. Biomolecules 2023, 13(9), 1412. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Chen, X.; Luo, Q.; Liu, X.; Wang, X.; Cui, Z.; He, A.; He, S.; Jiang, Z.; Wu, N.; Chen, P.; Yu, K.; Zhuang, J. Retinoblastoma Cell-Derived Exosomes Promote Angiogenesis of Human Vesicle Endothelial Cells through microRNA-92a-3p. Cell Death Dis. 2021, 12(7), 695. [Google Scholar] [CrossRef]
- Sundaram, K.; Miller, D. P.; Kumar, A.; Teng, Y.; Sayed, M.; Mu, J.; Lei, C.; Sriwastva, M. K.; Zhang, L.; Yan, J.; Merchant, M. L.; He, L.; Fang, Y.; Zhang, S.; Zhang, X.; Park, J. W.; Lamont, R. J.; Zhang, H.-G. Plant-Derived Exosomal Nanoparticles Inhibit Pathogenicity of Porphyromonas Gingivalis. iScience 2019, 21, 308–327. [Google Scholar] [CrossRef]
- Bui, F. Q.; Almeida-da-Silva, C. L. C.; Huynh, B.; Trinh, A.; Liu, J.; Woodward, J.; Asadi, H.; Ojcius, D. M. Association between Periodontal Pathogens and Systemic Disease. Biomed. J. 2019, 42(1), 27–35. [Google Scholar] [CrossRef]
- Rosa, C. D. D. R. D.; Gomes, J. M. de L.; Moraes, S. L. D. de; Lemos, C. A. A.; da Fonte, T. P.; Limirio, J. P. J. de O.; Pellizzer, E. P. Use of Chlorhexidine Chip after Scaling and Root Planning on Periodontal Disease: A Systematic Review and Meta-Analysis. Saudi Dent. J. 2021, 33(1), 1–10. [Google Scholar] [CrossRef]
- Kim, M.; Park, J. H. Isolation of Aloe Saponaria-Derived Extracellular Vesicles and Investigation of Their Potential for Chronic Wound Healing. Pharmaceutics 2022, 14(9), 1905. [Google Scholar] [CrossRef]
- Kim, M.; Jang, H.; Park, J. H. Balloon Flower Root-Derived Extracellular Vesicles: In Vitro Assessment of Anti-Inflammatory, Proliferative, and Antioxidant Effects for Chronic Wound Healing. Antioxidants 2023, 12(6), 1146. [Google Scholar] [CrossRef]
- Savcı, Y.; Kırbaş, O. K.; Bozkurt, B. T.; Abdik, E. A.; Taşlı, P. N.; Şahin, F.; Abdik, H. Grapefruit-Derived Extracellular Vesicles as a Promising Cell-Free Therapeutic Tool for Wound Healing. Food Funct. 2021, 12(11), 5144–5156. [Google Scholar] [CrossRef]
- Liu, B.; Lu, Y.; Chen, X.; Muthuraj, P. G.; Li, X.; Pattabiraman, M.; Zempleni, J.; Kachman, S. D.; Natarajan, S. K.; Yu, J. Protective Role of Shiitake Mushroom-Derived Exosome-Like Nanoparticles in D-Galactosamine and Lipopolysaccharide-Induced Acute Liver Injury in Mice. Nutrients 2020, 12(2), 477. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, X.; Deng, Z.-B.; Mu, J.; Zhang, L.; Yan, J.; Miller, D.; Feng, W.; McClain, C. J.; Zhang, H.-G. Ginger-Derived Nanoparticles Protect against Alcohol-Induced Liver Damage. J. Extracell. Vesicles 2015, 4, 28713. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.-S.; Kim, H.-W.; Hwang, J.-H.; Eom, J.-Y.; Kim, D.-H.; Park, J.; Tae, H.-J.; Lee, S.; Yoo, J.-G.; Kim, J.-I.; Lim, J.-H.; Kwun, I.-S.; Baek, M.-C.; Cho, Y.-E.; Kim, D.-K. Plum-Derived Exosome-like Nanovesicles Induce Differentiation of Osteoblasts and Reduction of Osteoclast Activation. Nutrients 2023, 15(9), 2107. [Google Scholar] [CrossRef] [PubMed]
- Seo, K.; Yoo, J. H.; Kim, J.; Min, S. J.; Heo, D. N.; Kwon, I. K.; Moon, H.-J. Ginseng-Derived Exosome-like Nanovesicles Extracted by Sucrose Gradient Ultracentrifugation to Inhibit Osteoclast Differentiation. Nanoscale 2023, 15(12), 5798–5808. [Google Scholar] [CrossRef]
- Wang, B.; Zhuang, X.; Deng, Z.-B.; Jiang, H.; Mu, J.; Wang, Q.; Xiang, X.; Guo, H.; Zhang, L.; Dryden, G.; Yan, J.; Miller, D.; Zhang, H.-G. Targeted Drug Delivery to Intestinal Macrophages by Bioactive Nanovesicles Released from Grapefruit. Mol. Ther. J. Am. Soc. Gene Ther. 2014, 22(3), 522–534. [Google Scholar] [CrossRef]
- Teng, Y.; Ren, Y.; Sayed, M.; Hu, X.; Lei, C.; Kumar, A.; Hutchins, E.; Mu, J.; Deng, Z.; Luo, C.; Sundaram, K.; Sriwastva, M. K.; Zhang, L.; Hsieh, M.; Reiman, R.; Haribabu, B.; Yan, J.; Jala, V. R.; Miller, D. M.; Van Keuren-Jensen, K.; Merchant, M. L.; McClain, C. J.; Park, J. W.; Egilmez, N. K.; Zhang, H.-G. Plant-Derived Exosomal MicroRNAs Shape the Gut Microbiota. Cell Host Microbe 2018, 24(5), 637–652.e8. [Google Scholar] [CrossRef]
- Sriwastva, M. K.; Deng, Z.-B.; Wang, B.; Teng, Y.; Kumar, A.; Sundaram, K.; Mu, J.; Lei, C.; Dryden, G. W.; Xu, F.; Zhang, L.; Yan, J.; Zhang, X.; Park, J. W.; Merchant, M. L.; Egilmez, N. K.; Zhang, H.-G. Exosome-like Nanoparticles from Mulberry Bark Prevent DSS-Induced Colitis via the AhR/COPS8 Pathway. EMBO Rep. 2022, 23(3), e53365. [Google Scholar] [CrossRef]
- Rahimi Ghiasi, M.; Rahimi, E.; Amirkhani, Z.; Salehi, R. Leucine-Rich Repeat-Containing G-Protein Coupled Receptor 5 Gene Overexpression of the Rat Small Intestinal Progenitor Cells in Response to Orally Administered Grape Exosome-like Nanovesicles. Adv. Biomed. Res. 2018, 7, 125. [Google Scholar] [CrossRef]
- Ju, S.; Mu, J.; Dokland, T.; Zhuang, X.; Wang, Q.; Jiang, H.; Xiang, X.; Deng, Z.-B.; Wang, B.; Zhang, L.; Roth, M.; Welti, R.; Mobley, J.; Jun, Y.; Miller, D.; Zhang, H.-G. Grape Exosome-like Nanoparticles Induce Intestinal Stem Cells and Protect Mice from DSS-Induced Colitis. Mol. Ther. J. Am. Soc. Gene Ther. 2013, 21(7), 1345–1357. [Google Scholar] [CrossRef]
- Zu, M.; Xie, D.; Canup, B. S. B.; Chen, N.; Wang, Y.; Sun, R.; Zhang, Z.; Fu, Y.; Dai, F.; Xiao, B. “Green” Nanotherapeutics from Tea Leaves for Orally Targeted Prevention and Alleviation of Colon Diseases. Biomaterials 2021, 279, 121178. [Google Scholar] [CrossRef]
- Deng, Z.; Rong, Y.; Teng, Y.; Mu, J.; Zhuang, X.; Tseng, M.; Samykutty, A.; Zhang, L.; Yan, J.; Miller, D.; Suttles, J.; Zhang, H.-G. Broccoli-Derived Nanoparticle Inhibits Mouse Colitis by Activating Dendritic Cell AMP-Activated Protein Kinase. Mol. Ther. J. Am. Soc. Gene Ther. 2017, 25(7), 1641–1654. [Google Scholar] [CrossRef]
- Gao, C.; Zhou, Y.; Chen, Z.; Li, H.; Xiao, Y.; Hao, W.; Zhu, Y.; Vong, C. T.; Farag, M. A.; Wang, Y.; Wang, S. Turmeric-Derived Nanovesicles as Novel Nanobiologics for Targeted Therapy of Ulcerative Colitis. Theranostics 2022, 12(12), 5596–5614. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-López, C. M.; Manzaneque-López, M. C.; Pérez-Bermúdez, P.; Soler, C.; Marcilla, A. Characterization and Bioactivity of Extracellular Vesicles Isolated from Pomegranate. Food Funct. 2022, 13(24), 12870–12882. [Google Scholar] [CrossRef]
- Garlic exosome-like nanoparticles reverse high-fat diet induced obesity via the gut/brain axis - PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/35154484/ (accessed on 27 March 2025).
- Cai, H.; Huang, L.-Y.; Hong, R.; Song, J.-X.; Guo, X.-J.; Zhou, W.; Hu, Z.-L.; Wang, W.; Wang, Y.-L.; Shen, J.-G.; Qi, S.-H. Momordica Charantia Exosome-Like Nanoparticles Exert Neuroprotective Effects Against Ischemic Brain Injury via Inhibiting Matrix Metalloproteinase 9 and Activating the AKT/GSK3β Signaling Pathway. Front. Pharmacol. 2022, 13, 908830. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.-W.; Ye, C.; Wang, K.-X.; Yang, X.; Zhu, P.-Y.; Hu, K.; Lan, T.; Huang, L.-Y.; Wang, W.; Gu, B.; Yan, C.; Ma, P.; Qi, S.-H.; Luo, L. Momordica. Charantia-Derived Extracellular Vesicles-Like Nanovesicles Protect Cardiomyocytes Against Radiation Injury via Attenuating DNA Damage and Mitochondria Dysfunction. Front. Cardiovasc. Med. 2022, 9, 864188. [Google Scholar] [CrossRef]
- Raimondo, S.; Nikolic, D.; Conigliaro, A.; Giavaresi, G.; Lo Sasso, B.; Giglio, R. V.; Chianetta, R.; Manno, M.; Raccosta, S.; Corleone, V.; Ferrante, G.; Citarrella, R.; Rizzo, M.; De Leo, G.; Ciaccio, M.; Montalto, G.; Alessandro, R. Preliminary Results of CitraVesTM Effects on Low Density Lipoprotein Cholesterol and Waist Circumference in Healthy Subjects after 12 Weeks: A Pilot Open-Label Study. Metabolites 2021, 11(5), 276. [Google Scholar] [CrossRef]
- Li, L.; Zhong, D.; Wang, S.; Zhou, M. Plant-Derived Materials for Biomedical Applications. Nanoscale 2025, 17(2), 722–739. [Google Scholar] [CrossRef]
- Wang, Q.; Zhuang, X.; Mu, J.; Deng, Z.-B.; Jiang, H.; Zhang, L.; Xiang, X.; Wang, B.; Yan, J.; Miller, D.; Zhang, H.-G. Delivery of Therapeutic Agents by Nanoparticles Made of Grapefruit-Derived Lipids. Nat. Commun. 2013, 4, 1867. [Google Scholar] [CrossRef]
- Doyle, L. M.; Wang, M. Z. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8(7), 727. [Google Scholar] [CrossRef]
- Umezu, T.; Takanashi, M.; Murakami, Y.; Ohno, S.-I.; Kanekura, K.; Sudo, K.; Nagamine, K.; Takeuchi, S.; Ochiya, T.; Kuroda, M. Acerola Exosome-like Nanovesicles to Systemically Deliver Nucleic Acid Medicine via Oral Administration. Mol. Ther. Methods Clin. Dev. 2021, 21, 199–208. [Google Scholar] [CrossRef]
- Seow, S. L. S.; Hong, S. L.; Lee, G. S.; Malek, S. N. A.; Sabaratnam, V. 6-Shogaol, a Neuroactive Compound of Ginger (Jahe Gajah) Induced Neuritogenic Activity via NGF Responsive Pathways in PC-12 Cells. BMC Complement. Altern. Med. 2017, 17(1), 334. [Google Scholar] [CrossRef] [PubMed]
- Lozano, O.; Lázaro-Alfaro, A.; Silva-Platas, C.; Oropeza-Almazán, Y.; Torres-Quintanilla, A.; Bernal-Ramírez, J.; Alves-Figueiredo, H.; García-Rivas, G. Nanoencapsulated Quercetin Improves Cardioprotection during Hypoxia-Reoxygenation Injury through Preservation of Mitochondrial Function. Oxid. Med. Cell. Longev. 2019, 2019, 7683051. [Google Scholar] [CrossRef] [PubMed]
- Dad, H. A.; Gu, T.-W.; Zhu, A.-Q.; Huang, L.-Q.; Peng, L.-H. Plant Exosome-like Nanovesicles: Emerging Therapeutics and Drug Delivery Nanoplatforms. Mol. Ther. J. Am. Soc. Gene Ther. 2021, 29(1), 13–31. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Lin, H.; Jiang, X.; Li, W.; Gao, Y.; Li, M.; Yu, Y.; Chen, N.; Gao, J. Exosome-like Nanoparticles Derived from Fruits, Vegetables, and Herbs: Innovative Strategies of Therapeutic and Drug Delivery. Theranostics 2024, 14(12), 4598–4621. [Google Scholar] [CrossRef]
- Yi, C.; Lu, L.; Li, Z.; Guo, Q.; Ou, L.; Wang, R.; Tian, X. Plant-Derived Exosome-like Nanoparticles for microRNA Delivery in Cancer Treatment. Drug Deliv. Transl. Res. 2025, 15(1), 84–101. [Google Scholar] [CrossRef]
- Lozano, O.; Lázaro-Alfaro, A.; Silva-Platas, C.; Oropeza-Almazán, Y.; Torres-Quintanilla, A.; Bernal-Ramírez, J.; Alves-Figueiredo, H.; García-Rivas, G. Nanoencapsulated Quercetin Improves Cardioprotection during Hypoxia-Reoxygenation Injury through Preservation of Mitochondrial Function. Oxid. Med. Cell. Longev. 2019, 2019, 7683051. [Google Scholar] [CrossRef]
- Tian, J.; Han, Z.; Song, D.; Peng, Y.; Xiong, M.; Chen, Z.; Duan, S.; Zhang, L. Engineered Exosome for Drug Delivery: Recent Development and Clinical Applications. Int. J. Nanomedicine 2023, 18, 7923–7940. [Google Scholar] [CrossRef]
Figure 1.
Biological pathways and structures of PDVLNs. Three pathways generate PDVLNs: (1) EXPOs merge with the PM, releasing vesicles to the cell wall. (2) MVB matures to form ILVs, aided by (2a) fungal infection, (2b) plant–microbe symbiosis, or (2c) direct fusion with the PM, releasing ILVs as PDVLNs. (3) MVBs fuse with vacuoles, which then fuse with the PM, releasing ILVs.
Figure 1.
Biological pathways and structures of PDVLNs. Three pathways generate PDVLNs: (1) EXPOs merge with the PM, releasing vesicles to the cell wall. (2) MVB matures to form ILVs, aided by (2a) fungal infection, (2b) plant–microbe symbiosis, or (2c) direct fusion with the PM, releasing ILVs as PDVLNs. (3) MVBs fuse with vacuoles, which then fuse with the PM, releasing ILVs.
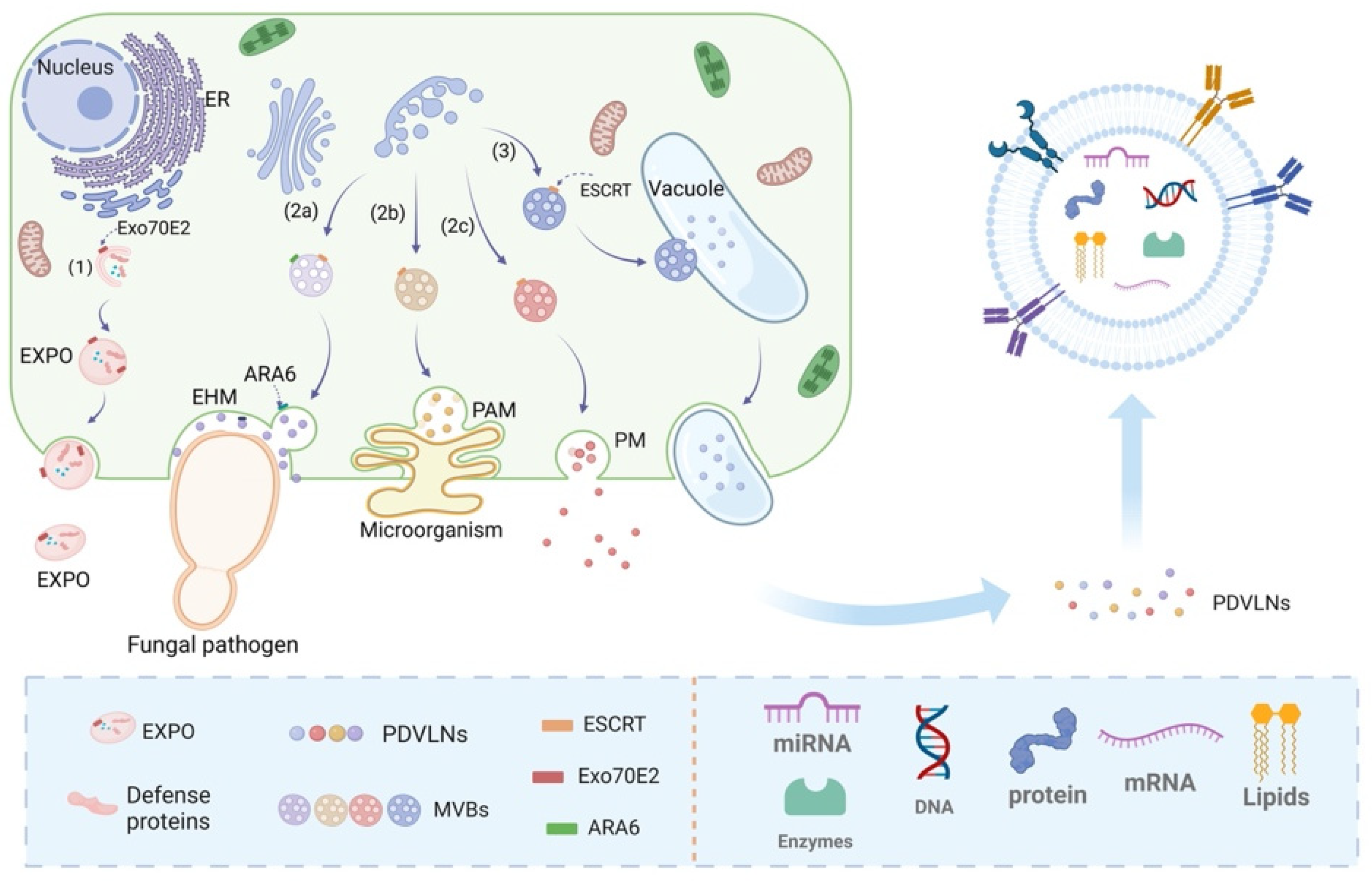
Figure 2.
Different processes for obtaining PDVLNs. The preparation liquid is created via a blender, homogenizer, or tissue permeate along with traditional Chinese medicine decoctions. A employs ultraspeed differential centrifugation, often with density gradient centrifugation (B), for PDVLN separation and purification. C uses ultrafiltration to filter out organelles and dead cells, resulting in PDVLNs. D employs SEC to separate PDVLNs on the basis of flow rates in a chromatographic column. E utilizes immunoprecipitation with specific antibodies to isolate PDVLNs efficiently.
Figure 2.
Different processes for obtaining PDVLNs. The preparation liquid is created via a blender, homogenizer, or tissue permeate along with traditional Chinese medicine decoctions. A employs ultraspeed differential centrifugation, often with density gradient centrifugation (B), for PDVLN separation and purification. C uses ultrafiltration to filter out organelles and dead cells, resulting in PDVLNs. D employs SEC to separate PDVLNs on the basis of flow rates in a chromatographic column. E utilizes immunoprecipitation with specific antibodies to isolate PDVLNs efficiently.
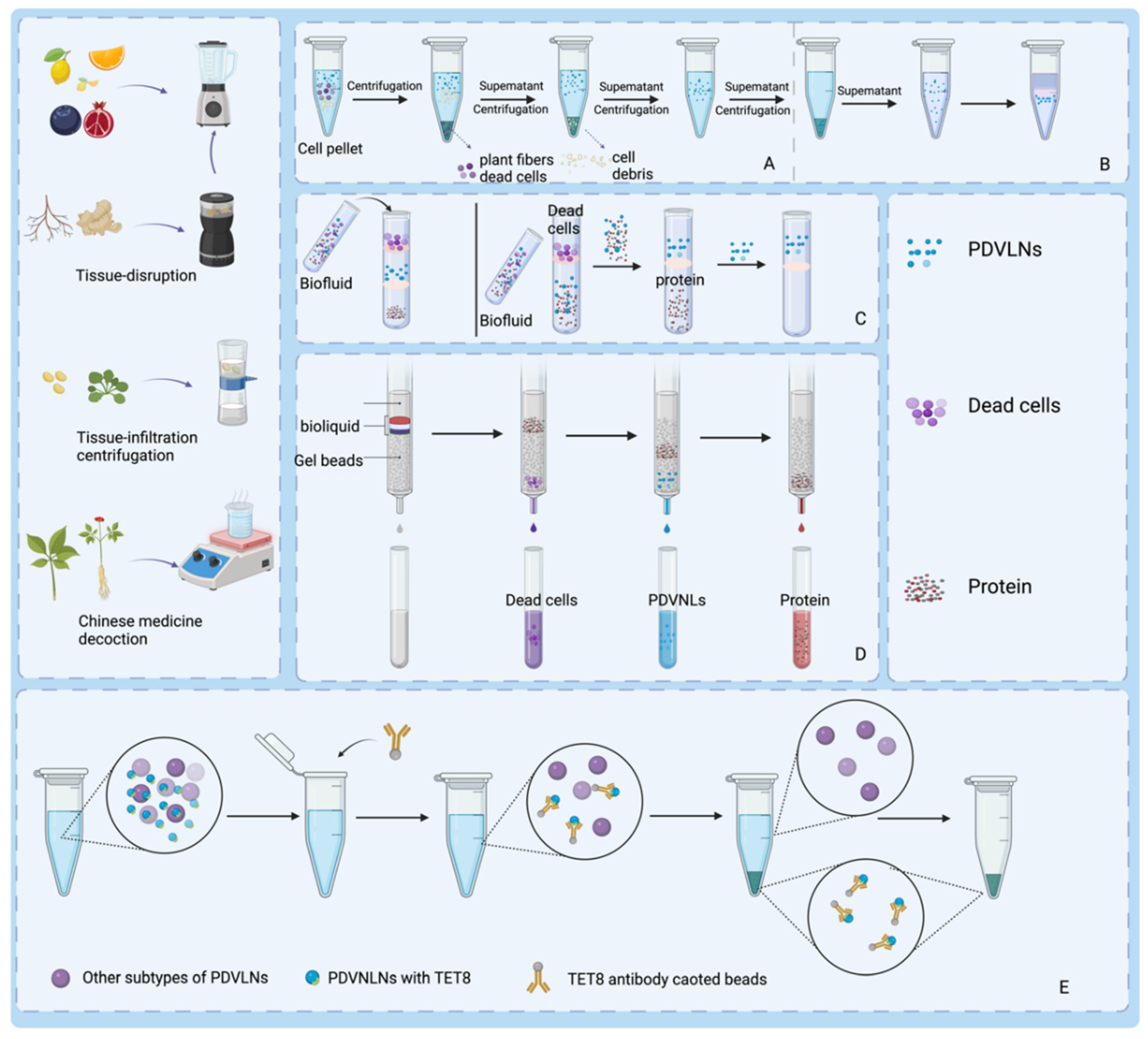
Figure 3.
Promotion of wound healing by PDVLNs.
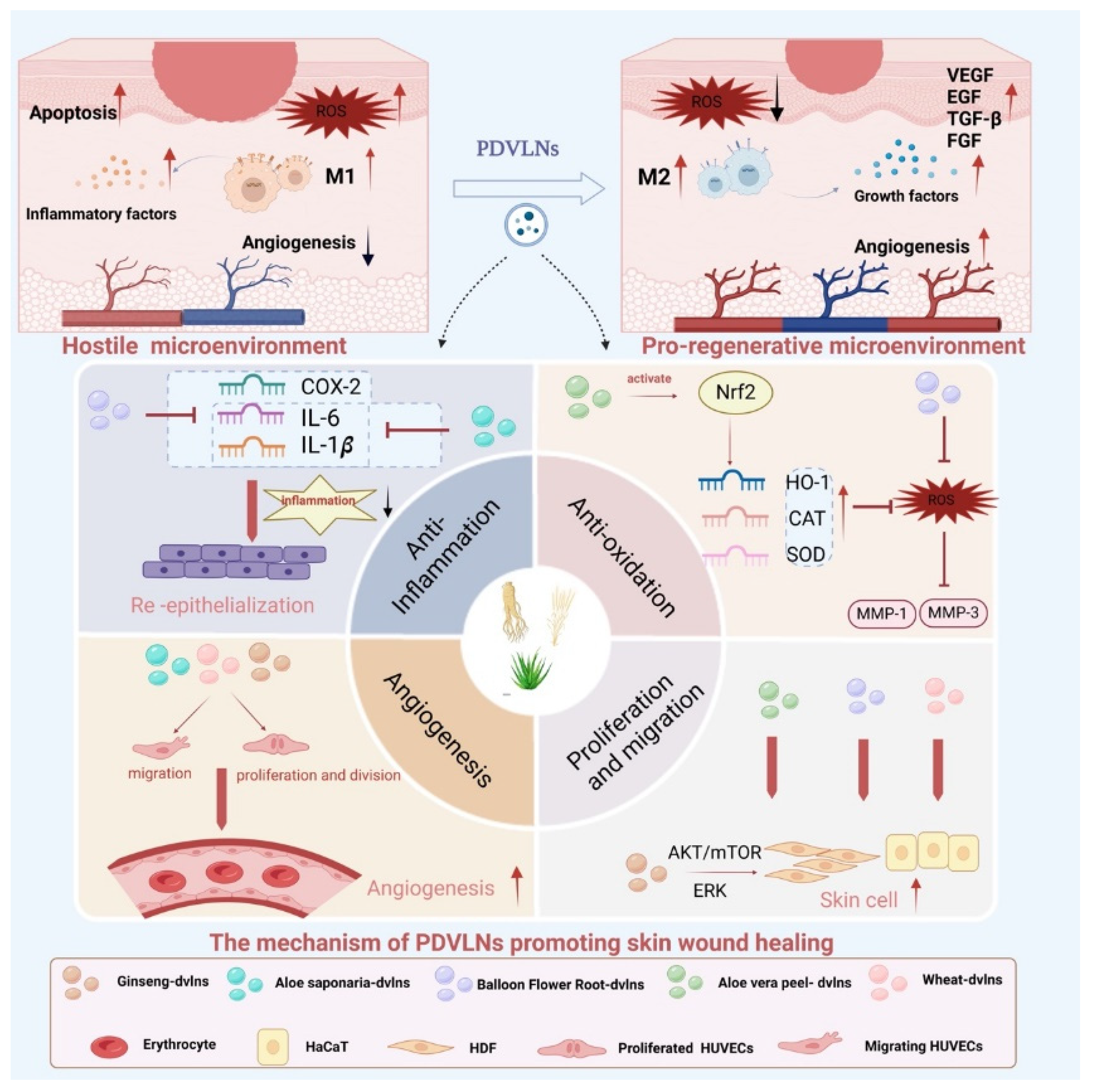
Figure 4.
Antiaging Effects of PDVLNs.
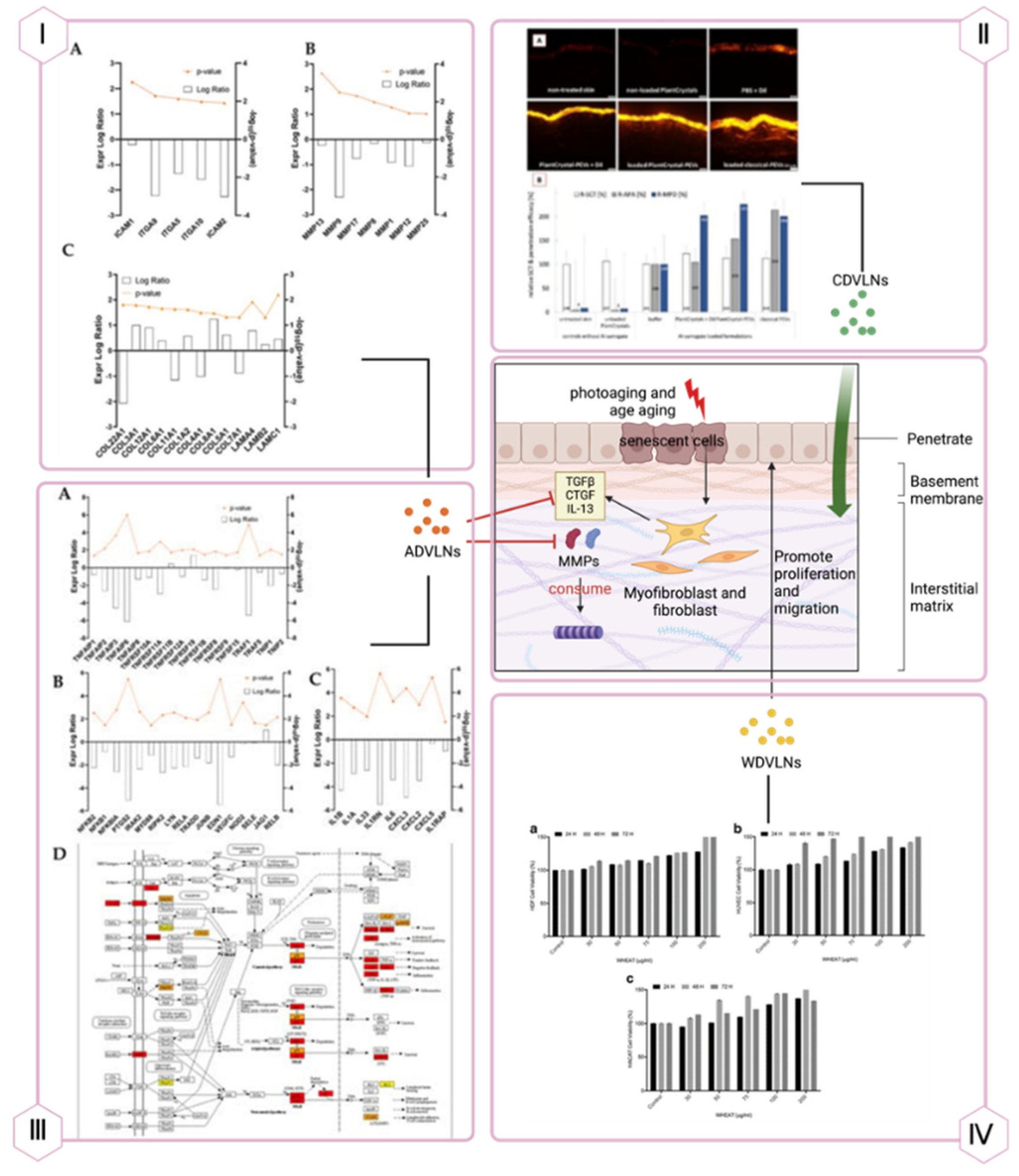
Figure 5.
Promotion of Bone Regeneration by PDVLNs.
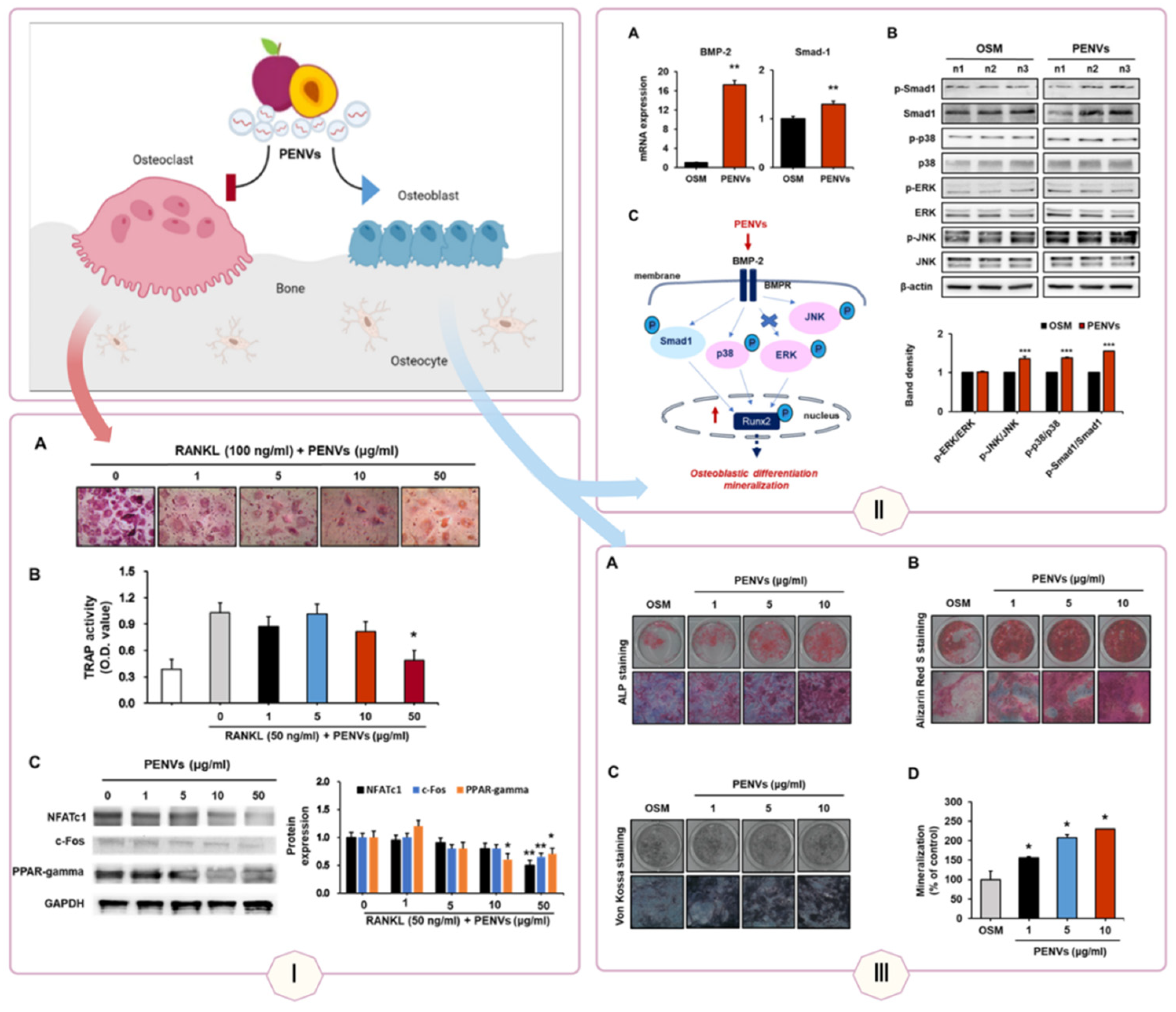
Figure 6.
PDVLNs in Gastrointestinal Repair.Gg-DVLNs suppress NLRP3 inflammasome activity and enhance LGG13A/IL-22 levels in IBD models, while Bb-DVLNs reduce IL-8β and IL-5 via NF-κB/TLR4 modulation in Caco-1 cells, promoting mucosal repair.
Figure 6.
PDVLNs in Gastrointestinal Repair.Gg-DVLNs suppress NLRP3 inflammasome activity and enhance LGG13A/IL-22 levels in IBD models, while Bb-DVLNs reduce IL-8β and IL-5 via NF-κB/TLR4 modulation in Caco-1 cells, promoting mucosal repair.
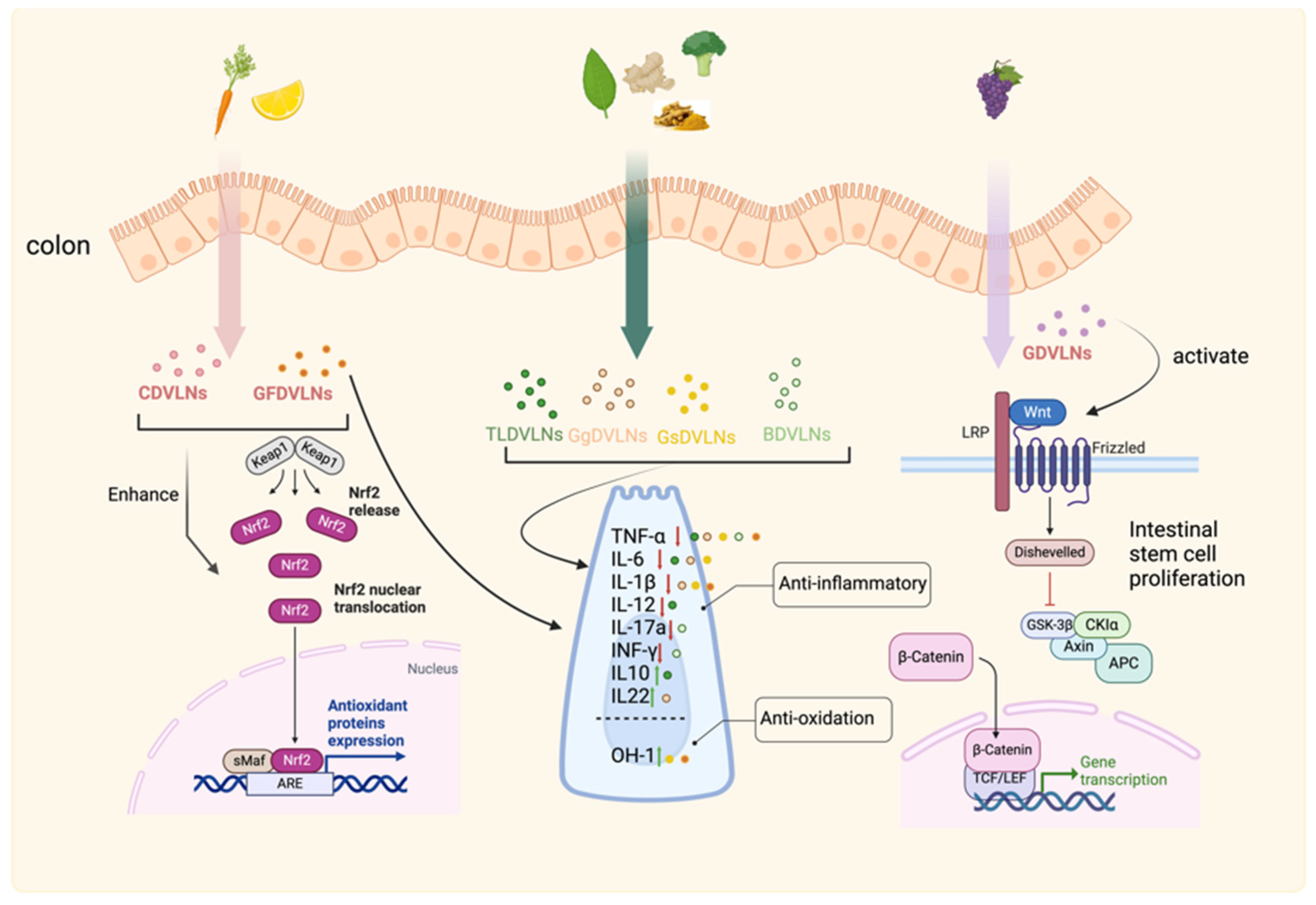
Figure 7.
PDVLNs in Periodontal Repair.
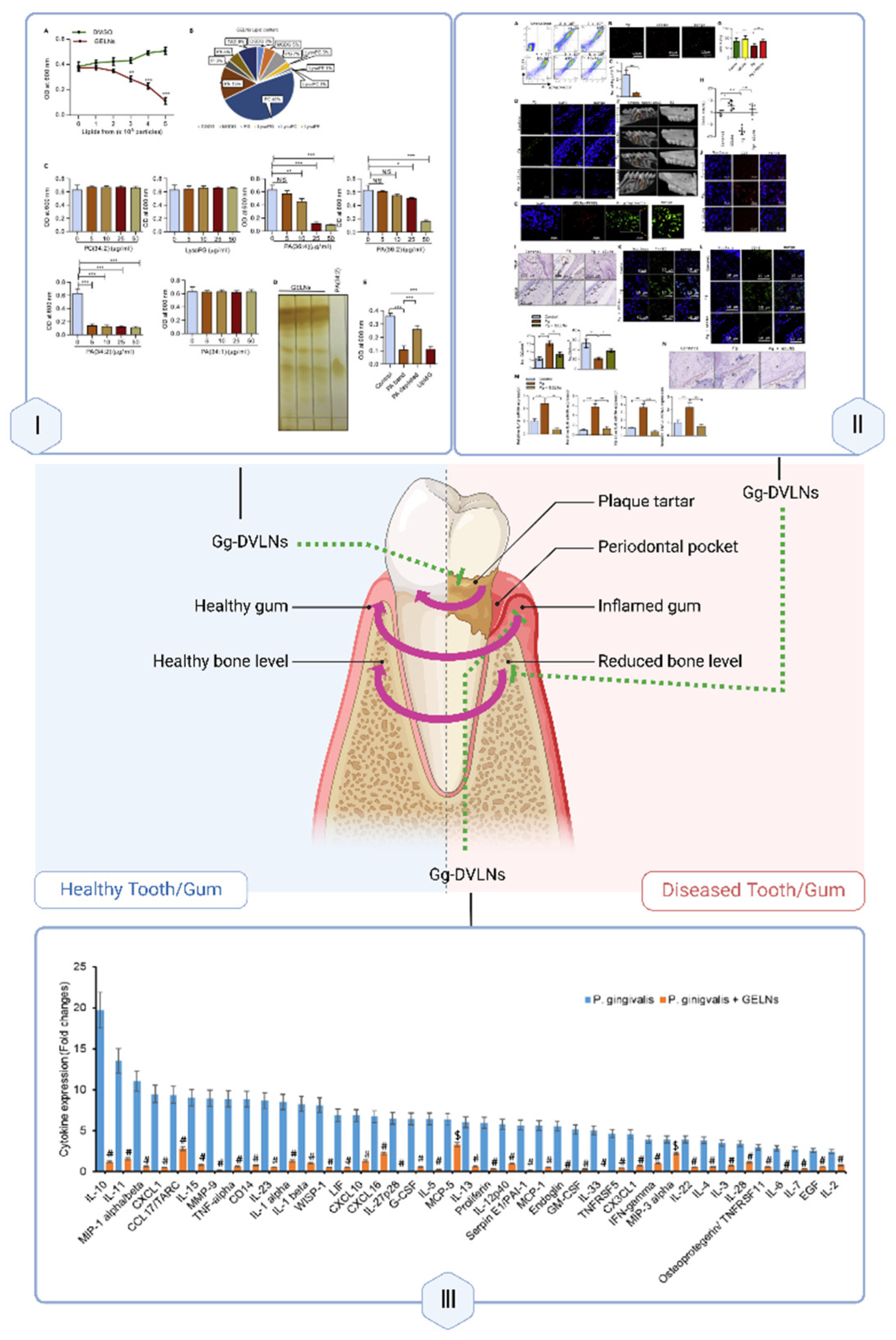
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



