Submitted:
06 June 2025
Posted:
06 June 2025
You are already at the latest version
Abstract
The expression of recombinant proteins in heterologous hosts is a common strategy to obtain larger quantities of the "protein of interest" (POI) for scientific, therapeutic or commercial purposes. However, the experimental success of such an approach critically depends on the choice of an appropriate host system to obtain biologically active forms of the POI. The correct folding of the molecule, mediated by disulfide bond formation, is one of the most critical steps in that process. Here we describe the recombinant expression of hirudin, a leech-derived anticoagulant and thrombin inhibitor, in the yeast Komagataella phaffii (formerly known and mentioned throughout the manuscript as Pichia pastoris) and in two different strains of Escherichia coli, one of them being especially designed for improved disulfide bond formation. Cultivation of the heterologous hosts and expression of hirudin were performed at different temperatures, ranging from 22 to 42 °C for the bacterial strains and from 20 to 30 °C for the yeast strain, respectively. The thrombin inhibitory potencies of all hirudin preparations were determined using the thrombin time coagulation assay. To our surprise, the hirudin preparations of P. pastoris were considerably less potent as thrombin inhibitors compared to the respective preparations of both E. coli strains, indicating that an eukaryotic background is not per se a better choice for the expression of a biologically active eukaryotic protein. The hirudin preparations of both E. coli strains exhibited comparable high thrombin-inhibitory potencies when the strains were cultivated at their respective optimal temperatures, whereas lower or higher cultivation temperatures reduced the inhibitory potencies.
Keywords:
hirudin
; hirudin‐like factor
; Escherichia coli
; Komagataella phaffii
; recombinant protein expression
1. Introduction
Hirudin of the medicinal leech, Hirudo medicinalis Linne (1758) is a small molecule of about 7 kDa in size (Dodt et al. 1984). It is a very strong natural inhibitor of thrombin and hence a very effective anticoagulant (Markwardt 1970). In fact, it is so far the one and only leech-derived bioactive factor that found its way from “bench to bedside” (Markwardt 1989; Johnson 1994; Greinacher and Warkentin 2008). The structure of hirudin is characterized by the presence of six conserved cysteine residues that form three intra-molecular disulfide bonds and thereby stabilize the so-called central globular domain (Dodt et al. 1985; Rydel et al. 1990). The thrombin-inhibitory effect of hirudin depends on both the short N-terminus and the elongated C-terminal tail of the molecule. The N-terminal five amino acid residues of hirudin penetrate the active-site cleft and block the catalytic centre of thrombin, whereas the C-terminal tail binds to exosite 1, the fibrinogen-binding site of thrombin (Rydel et al. 1991). Hirudin is hence a bivalent thrombin inhibitor (Warkentin 2004). However, the thrombin-inhibitory effect of hirudin is also strongly influenced by the central globular domain, very likely through the correct spacing, positioning and orientation of both termini and by stabilizing the binding to thrombin (Müller et al. 2020a). Several variants of hirudin were described in H. medicinalis including variant HV1 (VV-hirudin), variant HV2 (IT-hirudin) and two forms of variant HV3 (PA-hirudin) (Dodt et al. 1984; Harvey et al. 1986; Dodt et al. 1986). All variants exhibit about the same thrombin-inhibitory potencies (Müller et al. 2016).
Hirudin-like factors (HLF) represent an additional group of leech-derived bioactive factors. They exhibit certain shared features with hirudins including six cysteine residues within the central globular domain and a conserved gene structure that is comprised of four exons and three introns (Müller et al. 2016). However, beside common features, hirudins and HLFs significantly deviate in other aspects, such as molecular masses and isoelectric points (pI values) (Müller et al. 2017). While certain HLFs (including HLF1V) display thrombin-inhibitory potencies equivalent to those of hirudins, others demonstrate either minimal or no thrombin inhibition (Müller et al. 2016, 2020b; Ben Ahmed et al. 2024).
Living medicinal leeches of several genera are used for bloodletting and the treatment of circulatory disorders since ancient times (Abdualkader et al. 2013; Sig et al. 2017). In addition, dried leeches (Shui Zhi) are an ingredient of the Traditional Chinese Medicine (TCM) and are routinely orally administered to cure blood stasis and to stimulate menstruation discharge (Dong et al. 2016; Qiu et al. 2019). In modern Western medicine, hirudin is clinically used as an alternative to heparin in patients that suffer from heparin-induced thrombocytopenia (HIT), a potentially lethal complication of a long-term heparin administration (Greinacher et al. 1999; Lubenow and Greinacher 2002). Both yeast-based and bacterial expression systems are commonly used for the production of recombinant hirudin (Harvey et al. 1986; Riehl-Bellon et al. 1989; Rosenfeld et al. 1996; Kochanowski et al. 2006) and both alternatives lead to hirudin preparations that are almost equally effective as natural hirudin (Nowak 2002). However, only recently the cell-free synthesis of hirudin HV1 was shown to be a convincing alternative to the recombinant expression, yielding a form of hirudin HV1 that exhibited an up to 100 times higher thrombin-inhibitory potency compared to bacterially expressed hirudin HV1 (Wüstenhagen et al. 2020). This observation highlights the necessity to further evaluate and improve the “classical” expression strategies for hirudin. Here we present an investigation that compares a yeast-based eukaryotic expression system and two bacterial expression systems in terms of the thrombin-inhibitory potencies of the respective recombinant hirudin preparations.
2. Materials and Methods
2.1. Expression of Recombinant Hirudin HV1 and HLF1V in E. coli
For expression, pQE30Xa-clones containing cDNAs encoding hirudin HV1 or HLF1V of H. medicinalis were transformed into either E. coli DH5α cells (Hanahan 1985) or SHuffle® T7 cells (Lobstein et al. 2012) using chemically competent cells and the standard procedure described by Green and Sambrook (2012). Cells were plated out on LB (lysogeny broth) agar plates containing 100 μg/mL ampicillin for selection and incubated overnight for 16–20 h at 37 °C. A few colonies were used to inoculate an overnight culture of a final volume of 11 mL LB medium containing 100 μg/mL ampicillin and shaking at either 37 °C (strain DH5α) or 30 °C (strain SHuffle® T7). The next morning, two flasks, each containing 500 mL LB medium with ampicillin, were inoculated with 5 mL of the overnight culture and incubated under constant shaking at 30 °C (experiment 1) or at 22 °C, 30 °C, 37 °C and 42 °C, respectively (experiment 2). Optical densities were determined in a regular manner using a DU530 spectrophotometer (Beckman Coulter, Krefeld, Germany). At an OD600 = 0.5, the expression of recombinant hirudins was induced by adding isopropyl-β-D-thiogalactopyranoside (IPTG) to a final concentration of 1 mmol/L.
After 3 h of incubation, cells were harvested (centrifugation for 20 min at 4 °C and 3,900 g; Labofuge 400R, Thermo Scientific, Schwerte, Germany) , the pellets were carefully resuspended in 25 mL binding buffer (20 mmol/L Tris/HCl, 500 mmol/L NaCl, 5 mmol/L imidazole, pH 7.9) and the cells were sonicated using a Sonoplus homogenizer (Bandelin, Berlin, Germany) by applying a total of 10 sonication cycles with the following parameters: 75 impulses with a duration of 0.8 s and a power of 65–68 % each. To avoid heating, the samples were placed on ice during sonication and cooled on ice for 2 min before the start of the next cycle. After sonication, the samples were centrifuged for 45 min at 4 °C and 3,900 g (Labofuge 400R, Thermo Scientific, Schwerte, Germany). The supernatant was carefully removed and centrifuged for additional 10 min under the same conditions.
2.2. Expression of Recombinant Hirudin HV1 in P. pastoris
For the expression of recombinant hirudin HV1 in P. pastoris, a respective vector based on pPIC9K (InvitrogenTM, Thermo Scientific, Schwerte, Germany) was constructed using the gene synthesis service of Biocat (Biocat GmbH, Heidelberg, Germany). The vector confers resistance to kanamycin in bacteria and G-418 (geneticinTM) in eukaryotic cells, secures the stable integration into the genome of P. pastoris and allows for the expression of recombinant proteins under the control of the methanol-inducible alcohol oxidase 1 (AOX1) promoter. In addition, it contains an alpha factor secretion signal to guide the expressed recombinant protein to the growth medium. The cDNA sequence of hirudin HV1 that was used to construct the yeast expression vector was the same as it was used for the expression in bacterial cells (see above) and hence contained the coding regions for both a His-tag and a factor Xa protease cleavage site, located 5` to the coding region of hirudin HV1.
SacI-linearized plasmids were introduced into competent cells of P. pastoris strain GS115 (InvitrogenTM, Thermo Scientific, Schwerte, Germany) by electroporation using the following settings: 400 V/cm, 10 μF, low resistance. Transformed cells were selected on minimal dextrose medium (MD) plates. Final clones were selected by a two-step-process: 1) transferring single colonies from MD to YPD plates with increasing concentrations of geneticin (1 mg/mL, 1.5 mg/mL and 1.75 mg/mL, respectively) and 2) transferring single colonies from YPD plates to minimal methanol (MM) plates. Two clones (2 and 29) were selected for the following experiments.
Both clones were first inoculated in 5 mL of buffered glycerol-complex (BMGY) medium and incubated overnight at 30 °C and constant shaking. 1 mL of the overnight cultures was used to inoculate 50 mL of fresh BMGY medium. The cultures were incubated at 30 °C under constant shaking and the optical densities were measured in a regular manner. At an OD600 = 6, the cells were harvested, resuspended in 10 mL of buffered methanol-complex (BMMY) medium, and 1 mL of the resuspended cell solution was used to inoculate 500 mL of fresh BMMY medium. The cultures were incubated for additional four days at 30 °C (experiment 1) or 20 °C (experiment 2) under constant shaking, and each day 2.5 mL of methanol were added.
At the end of cultivation, the cells were harvested by centrifugation (Labofuge 400R, 45 min at 4 °C and 3,900 g) and the supernatants containing the secreted His-tagged hirudin HV1 were transferred to 50 mL tubes (Sarstedt, Nümbrecht, Germany). To each tube containing 45 mL of supernatant the following components were added: 5 mL of 200 mmol/L Tris/HCI pH 7.9 (final concentration 20 mmol/L), 1.461 g of NaCl (final concentration 500 mmol/L) and 0.5 mL of a 500 mmol/L imidazole solution (final concentration 5 mmol/L). In addition, the cell pellets were resuspended in binding buffer and sonicated as described above.
2.3. Purification of Recombinant Hirudins HV1 and HLF1V
The supernatant after sonication of bacterial cells or the supernatant after cultivation of P. pastoris were loaded on a self-packed column containing Ni-NTA His-Bind® resin (Merck, Darmstadt, Germany). Washing and elution steps were performed as recommended by the manufacturer of the resin. In detail, a stepwise washing and elution protocol was used with buffers (20 mmol/L Tris/HCl, 500 mmol/L NaCl, pH 7.9) containing increasing concentrations of imidazole (20 mmol/L, 50 mmol/L, 100 mmol/L, 200 mmol/L and 500 mmol/L, respectively). A final strip buffer (100 mmol/L EDTA) was applied to completely remove the Ni ions from the column. All elution fractions were collected and equal volumes of every fraction were analyzed by SDS-PAGE on 20 % acrylamide gels. Prior to factor Xa protease treatment, fractions of interest were dialyzed (regenerated cellulose dialysis membrane Spectra/Por® 7, MWCO 3.5 kDa; Carl Roth GmbH, Karlsruhe, Germany) two times for 24 h at 4 °C against a 100-fold excess of reaction buffer (20 mmol/L Tris/HCl, 50 mmol/L NaCl, 1 mmol/L CaCl2, pH 8.0).
Treatment of fusion proteins containing the factor Xa protease recognition sequence consisted of two steps: (1) factor Xa protease cleavage and (2) removal of factor Xa protease. Both steps were performed as recommended by the manufacturer (New England Biolabs, Frankfurt a. M., Germany). 1.7 mL of each hirudin or HLF1V preparation were incubated with 3 µg of factor Xa (3 μl of a 1 mg/mL stock solution) for 72 h at 22 °C in reaction buffer. The success of factor Xa treatment was controlled by SDS-PAGE analysis. Factor Xa protease was removed by adding an appropriate volume of equilibrated XarrestTM Xa protease removal resin (Merck, Darmstadt, Germany) to the reaction mixtures, incubation for 10 min at room temperature under constant shaking and collection of the supernatant following a centrifugation step for 5 min at 1000 g.
Molar concentrations of recombinant hirudin HV1 and HLF1V solutions were calculated by measuring the absorbance at λ = 280 nm and by dividing the obtained values by the molar absorption coefficient according to the equation ε = (nW × 5500) + (nY × 1490) + (nC × 125) (Gill and von Hippel 1989; Pace et al. 1995).
2.4. Blood Coagulation Assays
To verify the thrombin-inhibitory potencies of all recombinant hirudin HV1 and HLF1V preparations we applied the thrombin time test (TT; reference range 16.8-21.4 s) using a BFT II analyzer (Siemens Healthcare, Erlangen, Germany). All steps were carried out according to the instructions of the manufacturer. Protein samples were diluted with reaction buffer to reach final concentrations of 3.2 µmol/L, 0.32 µmol/L or 0.032 µmol/L in the reaction assays. The desired amount of substrate was directly transferred into the test cuvette immediately before the plasma was added. Dade® Ci-Trol® 1 (Siemens Healthcare, Erlangen, Germany) was used as standardised human plasma. The incubation of reaction mixtures was carried out at 37.4 °C. Measurements that exceeded 300 s were stopped and considered as complete inhibition of clot formation.
3. Results
3.1. Expression of Recombinant Hirudin HV1 and HLF1V in Escherichia coli Strains
Hirudin HV1 and HLF1V were expressed in the E. coli strains DH5α and SHuffle® T7 at 30 °C and were present in the soluble cytosolic fraction. Both factors could be successfully purified by His-tag affinity chromatography. SDS-PAGE analysis revealed that hirudin HV1 was eluted over a broad range of imidazole concentrations between 20 mmol/L and 500 mmol/L with almost equal amounts of recombinant protein present in the 50 mmol/L and 100 mmol/L imidazole elution fractions, whereas HLF1V was eluted over a more narrow range of imidazole concentrations between 50 mmol/L and 200 mmol/L with a sharp peak in the 100 mmol/L imidazole fraction (Figure 1).
In all cases the 100 mmol/L imidazole elution fractions contained only little amounts of unspecific contaminations and were selected for further processing.
After dialysis, all selected hirudin HV1 and HLF1V fractions were subject to a factor Xa treatment for 60 hours to remove the N-terminal His-tag. The completeness of His-tag removal was confirmed by subsequent SDS-PAGE analyses (Figure 2).
Please note that untagged hirudin HV1 and HLF1V are very poorly stained by Coomassie brilliant blue stain due to the low number (hirudin HV1) or even absence (HLF1V) of basic amino acid residues in the molecules, whereas the His-tagged proteins are properly stained due to the His-tag that not only six histidine residues, but also two additional basic amino acid residues.
After the removal of factor Xa protease (see Material and Methods), the protein concentrations in all treated samples were determined. The values differed only slightly and are in a range of about 130–190 µmol/L or 840–1340 µg/mL (see Table 1 for details).
3.2. Functional Characterization of Hirudin HV1 and HLF1V Expressed in E. coli Strains
All samples were subject to a functional characterization by the Thrombin Time Test (TT-Test), a coagulation assay that specifically addresses the activity of thrombin. The final concentrations of hirudin HV1 and HLF1V were adjusted to 3.2 µmol/L and 0.32 µmol/L, respectively, the values thereby followed our previous investigations. All samples displayed a strong thrombin inhibitory potency, but to our surprise, only the preparations of hirudin HV1 and HLF1V out of E. coli strain SHuffle® T7 reached the cut-off value of 300 s in the higher final concentration of 3.2 µmol/L. The preparations of both factors out of E. coli strain DH5α were noticeable less potent as thrombin inhibitors (see Figure 3).
The optimal cultivation temperature of E. coli strain DH5α is 37 °C, whereas the optimal cultivation temperature of E. coli strain SHuffle® T7 is 30 °C. To evaluate whether or not the cultivation temperature might influence the thrombin inhibitory potencies of hirudin HV1 preparations, bacteria of both strains were cultivated and the proteins expressed at the following different temperatures: 22, 30, 37 and 42 °C, respectively. The experimental routines were the same as described above. All factors were purified, and the final concentrations were in about the same range as for the previous preparations (see Table S1). All factors were functionally characterized, and the results clearly show that a maximal inhibitory potency of hirudin HV1 was only achieved when the respective strains were cultivated at their optimal temperatures (37 °C for DH5α and 30 °C for SHuffle® T7) whereas lower or higher cultivation temperatures led to hirudin HV1 preparations of reduced inhibitory potencies.
3.3. Expression of Recombinant Hirudin HV1 in P. pastoris GS115
As described in the Materials and Methods section, the recombinant hirudin HV1 that was expressed in P. pastoris GS115 contained a factor alpha secretion signal followed by the His-tag and the mature hirudin protein. We applied this strategy to achieve comparability to the His-tag purification process after bacterial protein expression (see above). Two clones, namely clone 2 and clone 29, were selected and cultivated as already described. Protein expression experiments in P. pastoris are routinely performed at temperatures up to 30 °C, but lower temperatures may reduce cell stress and enhance productivity (Zhong et al. 2014). We decided to cultivate both clones at 20 °C and at 30 °C.
The culture medium (that should contain the secreted His-tagged hirudin HV1 protein), the cell lysates after sonication and the separated supernatants as well as the resuspended pellets were analyzed by SDS-PAGE to verify the success of protein expression and to identify the fractions that contain the expressed hirudin HV1. However, in none of the fractions a protein of the expected size could be identified (Supplementary material Figure S1). The culture media of all four cultivations (about 450 mL each) were then loaded onto the Ni-agarose column and the elution fractions were analyzed by SDS-PAGE. As can be seen in Figure 5, hirudin HV1 could be isolated in various fractions of all four expression experiments, but in comparably low amounts. Almost no contaminations were present in the elution fractions.
For each clone and temperature, the fraction with the highest amount of hirudin HV1 was selected for further analyses. Dialysis, factor Xa protease treatment and removal of factor Xa protease were performed as described for the bacterial expression system. The completeness of His-tag removal was confirmed by subsequent SDS-PAGE analyses (see Supplementary Material Figure S2). The protein concentrations were determined and the results are listed in Table 2.
3.4. Functional Characterization of Hirudin HV1 Expressed in P. pastoris GS115
As for hirudin HV1 and HLF1 expressed in E. coli strains SHuffle T7® and DH5α (see above), the purified hirudin HV1 preparations that were expressed in P. pastoris GS115 clones 2 and 29 at either 20 °C or 30 °C of cultivation temperature were functionally tested for their thrombin-inhibitory potencies using the TT-test. The results are summarized in Figure 6.
All yeast-based hirudin HV1 preparations displayed thrombin-inhibitory potencies, but even at the high final concentration of 3.2 µmol/L the clotting times were markedly less prolonged compared to the hirudin HV1 preparations upon bacterial expression (see Figure 3 and Figure 4). In addition, no clear effect of different cultivation temperatures could be observed.
4. Discussion
Recombinant proteins are expressed in heterologous hosts like bacteria, yeast, insect or mammalian cells to obtain larger quantities of the “protein of interest” for scientific, therapeutic or commercial purposes (di Leandro et al. 2023; Jayakrishnan et al. 2024). However, many potential pitfalls exists and the final success critically depends on the choice of the right host expression system (Fernandez and Vega, 216; Gomes et al. 2016; Schütz et al. 2023). The correct disulfide bond formation is one of the most critical steps (de Marco 2009; Berkmen 2012; Pouresmaeil and Azizi-Dargahlou 2023). For the heterologous expression in bacteria, two strategies have been developed to enhance the correct formation of disulfide bonds in recombinant proteins: i) strains that carry knock outs of the (e.g., Origami of Novagen) or ii) strains that co-express proteins that catalyze oxidative protein folding (Saaranen and Ruddock, 2019; Ferrer-Miralles and Garcia-Fruitós, 2024). CyDisCo (cytoplasmic disulfide bond formation in E. coli) is an example of the second strategy (Hatahet et al. 2010; Nguyen et al. 2011; Gąciarz et al. 2017). The E. coli strain SHuffle® T7 carries the thioredoxin and glutathione reductases double knock-out, but additionally expresses a disulfide bond isomerase, DsbC (Lobstein et al.2012). A systematic comparison revealed that both strategies to enhance the correct formation of disulfide bonds in recombinant proteins have advantages and disadvantages (Castillo-Corujo et al. 2024).
The leech-derived thrombin inhibitor hirudin has not yet been expressed in one or the other of the improved E. coli strains. So far, all of our attempts to express recombinant hirudins or hirudin-like factors (HLFs) were performed in E. coli strain DH5α that was cultivated at 37 °C, and the preparations of recombinant hirudins displayed high thrombin inhibitory potencies (e.g., Müller et al. 2016; Müller et al. 2022). On the other hand, the E. coli strain SHuffle® T7 was already used to successfully express the recombinant human cysteine-rich proteins cathepsin B and L (Möller et al. 2024). Both hirudin HV1 and HLF1V of H. medicinalis were expressed in parallel in E. coli strains DH5α and SHuffle® T7 at 30 °C, the optimal cultivation temperature of strain SHuffle® T7. In all cases, the recombinant proteins were present within the soluble cytosolic fractions and all factors could be successfully purified by Ni-Agarose His-tag affinity chromatography (see Figure 1). After cleavage (see Figure 2) and removal of the His-tag, the final protein concentrations were determined and were in a range of appr. 1 mg/mL (see Table 1). The functional characterization using the thrombin-time test revealed that the preparations of both recombinant hirudin HV1 and HLF1V in E. coli SHuffle® T7 displayed high thrombin-inhibitory potencies (see Figure 3), comparable to previous preparations (Müller et al. 2020a). In contrast, the preparations in E. coli strain DH5α were considerably less inhibitory potent, not only compared to SHuffle® T7 (see Figure 3), but also to previous preparations in DH5a. This observation raised the question whether the effect was due to the temperature or due to the genetic background of the strain. In a second experiment we cultivated both E. coli strain DH5α and E. coli strain SHuffle® T7 at temperatures of 22, 30, 37 and 42 °C, respectively, spanning large parts of the total growth range of E. coli (Tuttle et al. 2021). Again, the expressed hirudin HV1 was present in the cytosolic fractions. The functional characterization of all preparations revealed clear optimum curves with either 37 °C (for E. coli strain DH5a) or 30 °C (for E. coli strain SHuffle® T7) being the optimal cultivation temperatures to achieve maximal inhibitory potencies of recombinant hirudin HV1 (see Figure 4A,B). The inhibitory potencies of hirudin HV1 preparations were higher when the protein was expressed in E. coli strain SHuffle® T7 cultivated at 30 °C compared to E. coli strain DH5α cultivated at 37 °C., indicating that both the optimal cultivation temperature and the genetic background of the respective E. coli strain influence the biological activity of an expressed recombinant protein. However, it is possible that further variations (e.g., media composition, IPTG concentration or duration of protein expression; Francis and Page 2010) may result in even higher inhibitory potencies of recombinant hirudin HV1 preparations.
It is very likely that the different inhibitory potencies of the different hirudin HV1 preparations are due to different amounts of correctly folded proteins, hence due to the correct formation of disulfide bonds. The presence of six cysteine residues within the amino acid sequence of hirudin HV1 results in a total of 76 different theoretically possible combinations of disulfide bond formation (calculated with the “Calculator of the number of possibilities for SS bridges in proteins”, Combet et al. 2000), and only one represents the correct form. Lower incubation temperatures generally favor the expression of soluble recombinant proteins in E. coli (Schein and Noteborn 1988; San-Miguel et al. 2013), but the influence of the cultivation temperature on the correct formation on disulfide bonds has not yet been thoroughly investigated.
Another promising strategy to enhance the output of a biologically active eukaryotic recombinant protein (e.g., hirudin HV1) is to switch from a bacterial to an eukaryotic expression system. The yeasts P. pastoris and S. cerevisiae represent such expression systems for hirudin (Rosenfeld et al. 1996; Huang et al. 2012; Sohn et al. 1991; Kim et al. 2001). Usually, recombinant hirudin expressed in yeast is secreted into the growth medium and eventually purified by gel filtration and anion exchange chromatography. We applied a slightly different strategy that, however, allowed for a direct comparison of the yeast expression system to the bacterial expression system. Hirudin HV1 was secreted into the medium with still the His-tag attached to the N-terminus. The subsequent protocol followed the purification (His-tag affinity chromatography) and processing (factor Xa protease treatment and removal) steps as outlined for the bacterial expression system. In general, the yield of hirudin HV1 was comparably low after yeast-based expression and did not substantially differ between the two selected clones (2 and 29) and the two incubation temperatures (20 °C and 30 °C, respectively; see Figure 5 and Table 2). But even more important, the thrombin-inhibitory potencies of all yeast-expressed hirudin HV1 preparations were much lower compared to the bacterially expressed preparations (compare Figure 3, Figure 4 and Figure 6). N-glycan hyper-mannosylation is a post-translationally modification that can occur in both S. cerevisiae and P. pastoris and may result in the expression of inactive recombinant proteins (Jayarai and Smooker 2009; Kastberg et al. 2022). Since we have not deglycosylated our eukaryotic hirudin HV1 preparations, an inhibitory effect of hyper-glycosylation hence cannot be ruled out. However, the expression of hirudin variant HV2 in P. pastoris by Rosenfeld et al. (1996) led to the purification of a recombinant hirudin that displayed a Ki value comparable to the Ki value of bacterially expressed recombinant hirudin. Interestingly, hirudin P6 of the Asian buffalo leech Hirudinaria manillensis Lesson (1842) is a naturally occurring Thr43-O-glycosylated and Tyr63-sulfated hirudin variant (Steiner et al. 1990; Steiner et al. 1992). Glycosylated forms of hirudin P6 displayed lower Ki values compared to the unmodified protein, but strikingly the sulfated and deglycosylated form displayed the lowest Ki value (Hsieh et al. 2014).
Temperature is another factor that might influence the expression of recombinant proteins in yeast, and sub-physiological temperatures (20 °C instead of 30 °C) did improve expression and assembly of difficult-to-express heterologous proteins in S. cerevisiae (So et al. 2023). However, the cultivation of P. pastoris at 20 °C did neither improve the yield nor the inhibitory potencies of the respective hirudin HV1 preparations (see Figure 6 and Table 2).
5. Summary
We have expressed recombinant hirudin HV1 and HLF1V in two different bacterial (both factors) and an eukaryotic (hirudin HV1 only) expression systems and subsequently purified, processed and functionally characterized the expressed recombinant proteins in terms of their thrombin-inhibitory potencies. For the bacterial strains, the choice of an E. coli strain that was particularly designed for the enhanced formation of disulfide bonds in cysteine-containing proteins (namely E. coli strain SHuffle® T7) resulted in the expression of a hirudin HV1 preparation with a moderately higher thrombin-inhibitory potency compared to the expression in a standard strain (namely E. coli strain DH5α) when both strains were cultivated at their optimal cultivation temperature of 30 °C (SHuffle® T7) or 37 °C (DH5α), respectively. However, the expression of hirudin HV1 in both E. coli strains at different temperatures ranging from 22 °C–42 °C led to preparations of the recombinant protein that in both cases exhibited a clear temperature-dependent optimum-curve of the inhibitory potencies: highest at 37 °C for E. coli DH5α and 30 °C for E. coli SHuffle® T7, respectively, but lower at cultivation temperatures above the optimum and lowest at 22 °C. The correct formation of disulfide bonds is hence a function of the optimal cultivation temperature of the bacterial host, at least for E. coli strains DH5α and SHuffle® T7 in combination with the expression of recombinant hirudin HV1. The expression of hirudin HV1 in the yeast P. pastoris, however, did result in preparations that exhibited only comparably low thrombin-inhibitory potencies. Hyper-glycosylation might be an explanation for that observation. Taken together, both E. coli strains DH5α and SHuffle® T7 represents appropriate expression systems for hirudin variants (and very likely for other cysteine-rich leech-derived factors like decorsins and ornatins, too) as long as the strains are cultivated at their optimal temperatures.
Author Contributions
C.M. and D.B. conceived the ideas and designed the methodology; Z.W., D.B. and C.M. performed the experiments; Z.W. and C.M. analyzed the data and drafted the manuscript; C.M. and U.T.B. supervised the experimental work. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
We declare that the investigations described in this paper comply with the current laws in Germany.
References
- Abdualkader AM, Ghawi AM, Alaama M, Awang M, Merzouk A (2013) Leech therapeutic applications. Indian J Pharm Sci 75(2):127–137.
- Ben Ahmed R, Abilov A, Müller C (2024) Diversity of hirudin and hirudin-like factor genes in the North-African medicinal leech, Hirudo troctina. Parasitol Res 14;123(11):382. [CrossRef]
- Berkmen M (2012) Production of disulfide-bonded proteins in Escherichia coli. Protein Expr Purif 82(1):240-251. [CrossRef]
- Castillo-Corujo A, Uchida Y, Saaranen MJ, Ruddock LW (2024) Escherichia coli cytoplasmic expression of disulfide-bonded proteins: Side-by-side comparison between two competing strategies. J Microbiol Biotechnol 34(5):1126-1134. [CrossRef]
- Combet C, Blanchet C, Geourjon C, Deléage G (2000) NPS@: network protein sequence analysis. Trends Biochem Sci 25(3):147-150. [CrossRef]
- de Marco A (2009) Strategies for successful recombinant expression of disulfide bond-dependent proteins in Escherichia coli. Microb Cell Fact 8:26. [CrossRef]
- di Leandro L, Colasante M, Pitari G, Ippoliti R (2023) Hosts and heterologous expression strategies of recombinant toxins for therapeutic purposes. Toxins (Basel) 15(12):699. [CrossRef]
- Dodt J, Müller H-P, Seemüller U, Chang J-Y (1984) The complete amino acid sequence of hirudin, a thrombin specific inhibitor: Application of colour carboxymethylation. FEBS Lett 165 2(9):180-184 . [CrossRef]
- Dodt J, Seemüller U, Maschler R, Fritz H (1985) The complete covalent structure of hirudin. Localization of the disulfide bonds. Biol Chem Hoppe Seyler 366(4):379-385. [CrossRef]
- Dodt J, Machleidt W, Seemüller U, Maschler R, Fritz H (1986) Isolation and characterization of hirudin isoinhibitors and sequence analysis of hirudin PA. Biol Chem Hoppe Seyler 367(8):803–811. [CrossRef]
- Fernández FJ, Vega MC (2016). Choose a suitable expression host: A survey of available protein production platforms. In: Vega, M. (eds) Advanced technologies for protein complex production and characterization. Advances in experimental medicine and biology, vol 896. Springer, Cham. [CrossRef]
- Ferrer-Miralles N, Garcia-Fruitós E (2024) Heterologous expression of difficult to produce proteins in bacterial systems. Int J Mol Sci 25(2):822. [CrossRef]
- Francis DM, Page R (2010) Strategies to optimize protein expression in E. coli. Curr Protoc Protein Sci 61(1):5.24.1–5.24.29. doi: 10.1002/0471140864.ps0524s61.
- Gąciarz A, Veijola J, Uchida Y, Saaranen MJ, Wang C, Hörkkö S, Ruddock LW (2016) Systematic screening of soluble expression of antibody fragments in the cytoplasm of E. coli. Microb Cell Fact 15:22. https://doi. org/10.1186/s12934-016-0419-5.
- Gill SC, von Hippel PH (1989) Calculation of protein extinction coefficients from amino acid sequence data. Anal Biochem 182:319–326. https://doi. org/10. 1016/ 0003- 2697(89) 90602-7.
- Gomes AR, Byregowda SM, Veeregowda BM, Balamurugan V (2016) An overview of heterologous expression host systems for the production of recombinant proteins. Adv Anim Vet Sci 4:346-356. [CrossRef]
- Green M, Sambrook J (2012) Molecular Cloning: A Laboratory Manual. 4th Edition, Vol. II, Cold Spring Harbor Laboratory Press, New York.
- Greinacher A, Völpel H, Janssens U, Hach-Wunderle V, Kemkes-Matthes B, Eichler P, Mueller-Velten HG, Pötzsch B (1999) Recombinant hirudin (lepirudin) provides safe and effective anticoagulation in patients with heparin-induced thrombocytopenia: a prospective study. Circulation 99(1):73-80. [CrossRef]
- Greinacher A, Warkentin TE (2008) The direct thrombin inhibitor hirudin. Thromb Haemost 99(05): 819-829. [CrossRef]
- Hanahan D (1985) Techniques for transformation of E. coli. In: Glover D M, editor. DNA cloning: a practical approach. Vol. 1. Oxford, United Kingdom: IRL Press; 1985.
- Harvey RP, Degryse E, Stefani L, Schamber F, Cazenave JP, Courtney M, Tolstoshev P, Lecocq JP (1986) Cloning and expression of a cDNA coding for the anticoagulant hirudin from the bloodsucking leech, Hirudo medicinalis. Proc Nat Acad Sci USA 83(4):1084–1088. [CrossRef]
- Hatahet F, Nguyen VD, Salo KEH, Ruddock LW (2010) Disruption of reducing pathways is not essential for efficient disulfide bond formation in the cytoplasm of E. coli. Microb Cell Fact 9:67. [CrossRef]
- Hsieh YSY, Wijeyewickrema LC, Wilkinson BL, Pike RN, Payne RJ (2014) Total synthesis of homogeneous variants of hirudin P6: a post-translationally modified anti-thrombotic leech-derived protein. Angew Chem Int Ed Engl 53(15):3947-3951. [CrossRef]
- Huang Y, Zhang Y, Wu Y, Wang J, Liu X, Dai L, Wang L, Yu M, Mo W (2012) Expression, purification, and mass spectrometric analysis of 15N, 13C-labeled RGD-hirudin, expressed in Pichia pastoris, for NMR studies. PLoS One 7(8):e42207. [CrossRef]
- Jayakrishnan A, Rosli WRW; Tahir ARM, Razak FSA, Kee PE, Ng HS, Chew Y-L, Lee S-K, Ramasamy M, Tan CS, Liew KB (2024) Evolving paradigms of recombinant protein production in pharmaceutical industry: A rigorous review. Sci 6, 9. [CrossRef]
- Jayaraj R, Smooker PM (2009) So you need a protein—A guide to the production of recombinant proteins. Open Vet Sci J 3:28-34. [CrossRef]
- Johnson PH (1994) Hirudin: clinical potential of a thrombin inhibitor. Annu Rev Med 45:165-177. [CrossRef]
- Kim MD, Rhee SK, Seo JH (2001) Enhanced production of anticoagulant hirudin in recombinant Saccharomyces cerevisiae by chromosomal delta-integration. J Biotechnol 85(1):41-48. [CrossRef]
- Kochanowski R, Kotłowski R, Szweda P (2006) Novel method of expression and purification of hirudin based on pBAD TOPO, pTYB12 vectors and gene synthesis. Protein Expr Purif 50(1):25-30. [CrossRef]
- Lobstein J, Emrich CA, Jeans C, Faulkner M, Riggs R, Berkmen M (2012) SHuffle, a novel Escherichia coli protein expression strain capable of correctly folding disulfide bonded proteins in its cytoplasm. Microb Cell Fact 11:56. [CrossRef]
- Markwardt F (1970) Hirudin as an inhibitor of thrombin. Method Enzym 19: 924-932. [CrossRef]
- Markwardt F (1989) Development of hirudin as an antithrombotic agent. Semin Thromb Hemost 15(3):269-282. [CrossRef]
- Möller C, Rimkus N, Skala FFO, Merouze M, Böttcher D, Dörr M, Bornscheuer UT (2024) Improved recombinant expression of soluble cathepsin B and L in Escherichia coli. Appl Microbiol Biotechnol 108(1):536. [CrossRef]
- Müller C, Mescke K, Liebig S, Mahfoud H, Lemke S, Hildebrandt J-P (2016) More than just one: multiplicity of Hirudins and Hirudin-like Factors in the Medicinal Leech, Hirudo medicinalis. Mol Genet Genomics 291(1):227-240. [CrossRef]
- Müller C, Haase M, Lemke S, Hildebrandt J-P (2017) Hirudins and hirudin-like factors in Hirudinidae: implications for function and phylogenetic relationships. Parasitol Res 116(1):313-325. [CrossRef]
- Müller C, Lukas P, Böhmert M, Hildebrandt J-P (2020a) Hirudin or hirudin-like factor—that is the question: insights from the analyses of natural and synthetic HLF variants. FEBS Lett 594(5):841-850. [CrossRef]
- Müller C, Lukas P, Sponholz D, Hildebrandt J-P (2020b) The hirudin-like factors HLF3 and HLF4-hidden hirudins of European medicinal leeches. Parasitol Res 119(6):1767-1775. [CrossRef]
- Müller C, Wang Z, Hamann M, Sponholz D, Hildebrandt J-P (2022) Life without blood: Molecular and functional analysis of hirudins and hirudin-like factors of the Asian non-hematophagous leech Whitmania pigra. J Thromb Haemost 20(8):1808-1817. [CrossRef]
- Nguyen VD, Hatahet F, Salo KEH, Enlund E, Zhang C, Ruddock LW (2011) Pre-expression of a sulfhydryl oxidase significantly increases the yields of eukaryotic disulfide bond containing proteins expressed in the cytoplasm of E.coli. Microb Cell Fact 10:1. [CrossRef]
- Nowak G (2002) Pharmacology of recombinant hirudin. Semin Thromb Hemost 28(5):415-424. [CrossRef]
- Pace CN, Vajdos F, Fee L, Grimsley G, Gray T (1995) How to measure and predict the molar absorption coefficient of a protein. Protein Sci 4:2411–2423. [CrossRef]
- Pouresmaeil M, Azizi-Dargahlou S (2023) Factors involved in heterologous expression of proteins in E. coli host. Arch Microbiol 205(5):212. [CrossRef]
- Qiu J, Lingna W, Jinghong H, Yongqing Z (2019) Oral administration of leeches (Shuizhi): A review of the mechanisms of action on antiplatelet aggregation. J Ethnopharmacol 232:103-109. [CrossRef]
- Riehl-Bellon N, Carvallo D, Acker M, Van Dorsselaer A, Marquet M, Loison G, Lemoine Y, Brown SW, Courtney M, Roitsch C (1989) Purification and biochemical characterization of recombinant hirudin produced by Saccharomyces cerevisiae. Biochemistry 28(7):2941-2949. [CrossRef]
- Rydel TJ, Ravichandran KG, Tulinsky A, Bode W, Huber R, Roitsch C, Fenton 2nd JW (1990) The structure of a complex of recombinant hirudin and human alpha-thrombin. Science 249(4966):277-280. [CrossRef]
- Rydel TJ, Tulinsky A, Bode W, Huber R (1991) Refined structure of the hirudin-thrombin complex. J Mol Biol 221(2):583-601. [CrossRef]
- Saaranen MJ, Ruddock LW (2019) Applications of catalyzed cytoplasmic disulfide bond formation. Biochem Soc Trans 47(5):1223-1231. [CrossRef]
- San-Miguel T, Pérez-Bermúdez P, Gavidia I (2013) Production of soluble eukaryotic recombinant proteins in E. coli is favoured in early log-phase cultures induced at low temperature. Springerplus 2:89. doi: 10.1186/2193-1801-2-89.
- Schein C and Noteborn M (1988) Formation of soluble recombinant proteins in Escherichia coli is favored by lower growth temperature. Nat Biotechnol 6:291–294. doi: 10.1038/nbt0388-291.
- Schütz A, Bernhard F, Berrow N, Buyel JF, Ferreira-da-Silva F, Haustraete J, van den Heuvel J, Hoffmann J-E, de Marco A, Peleg Y, Suppmann S, Unger T, Vanhoucke M, Witt S, Remans K (2023) A concise guide to choosing suitable gene expression systems for recombinant protein production. STAR Protoc 4(4):102572. [CrossRef]
- Sig AK, Guney M, Guclu AU, Ozmen E (2017) Medicinal leech therapy—an overall perspective. Integr Med Res 6(4):337–343. [CrossRef]
- So K-K, Le NMT, Nguyen N-L, Kim D-H (2023) Improving expression and assembly of difficult-to-express heterologous proteins in Saccharomyces cerevisiae by culturing at a sub-physiological temperature. Microb Cell Fact 22(1):55. [CrossRef]
- Sohn J-H, Lee S-K, Choi E-S, Rhee S-K (1991) Gene expression and secretion of the anticoagulant hirudin in Saccharomyces cerevisiae. J Microbiol Biotechnol 1(4):266-273.
- Steiner V, Knecht R, Gruetter M, Raschdorf F, Gassmann E, Maschler R (1990) Isolation and purification of novel hirudins from the leech Hirudinaria manillensis by high-performance liquid chromatography. J Chromatogr 530(2):273-282. [CrossRef]
- Steiner V, Knecht R, Börnsen KO, Gassmann E, Stone SR, Raschdorf F, Schlaeppi JM, Maschler R (1992) Primary structure and function of novel O-glycosylated hirudins from the leech Hirudinaria manillensis. Biochemistry 31(8):2294-2298. [CrossRef]
- Tuttle AR, Trahan ND, Son MS (2021) Growth and maintenance of Escherichia coli laboratory strains. Curr Protoc 1, e20. [CrossRef]
- Warkentin TE (2004) Bivalent direct thrombin inhibitors: hirudin and bivalirudin. Best Pract Res Clin Haematol 17(1):105-125. [CrossRef]
- Wüstenhagen DA, Lukas P, Müller C, Aubele SA, Hildebrandt J-P, Kubick S (2020) Cell-free synthesis of the hirudin variant 1 of the blood-sucking leech Hirudo medicinalis. Sci Rep 10(1):19818. [CrossRef]
Figure 1.
SDS-PAGE analysis of elution fractions of hirudin HV1 (A, C) and HLF1V (B, D) after expression in E. coli strains DH5α (A, B) and SHuffle® T7 (C, D). The red arrows indicate the respective factors. The numbers on the left side indicate the molecular mass in kDa of the respective molecular weight marker (MWM) bands. The numbers 5–500 indicate the imidazole concentrations in mmol/L in the respective elution fractions.
Figure 1.
SDS-PAGE analysis of elution fractions of hirudin HV1 (A, C) and HLF1V (B, D) after expression in E. coli strains DH5α (A, B) and SHuffle® T7 (C, D). The red arrows indicate the respective factors. The numbers on the left side indicate the molecular mass in kDa of the respective molecular weight marker (MWM) bands. The numbers 5–500 indicate the imidazole concentrations in mmol/L in the respective elution fractions.
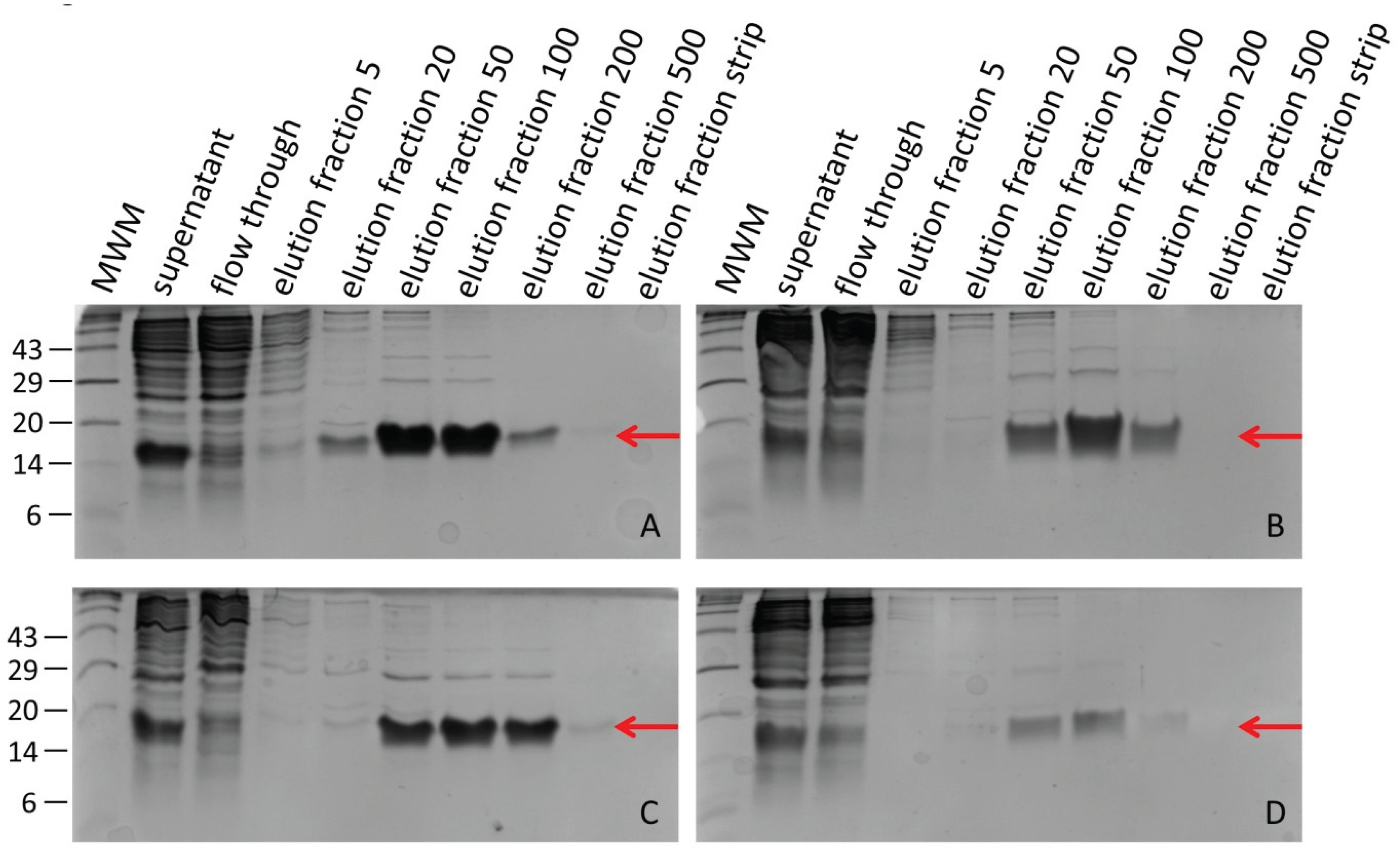
Figure 2.
Factor Xa protease treatment of selected elution fractions (ef) of hirudin HV1 and HLF1V expressed in E. coli strains SHuffle® T7 (A) or DH5α (B). The numbers on the left side indicate the molecular mass in kDa of the respective molecular weight marker (MWM) bands.
Figure 2.
Factor Xa protease treatment of selected elution fractions (ef) of hirudin HV1 and HLF1V expressed in E. coli strains SHuffle® T7 (A) or DH5α (B). The numbers on the left side indicate the molecular mass in kDa of the respective molecular weight marker (MWM) bands.
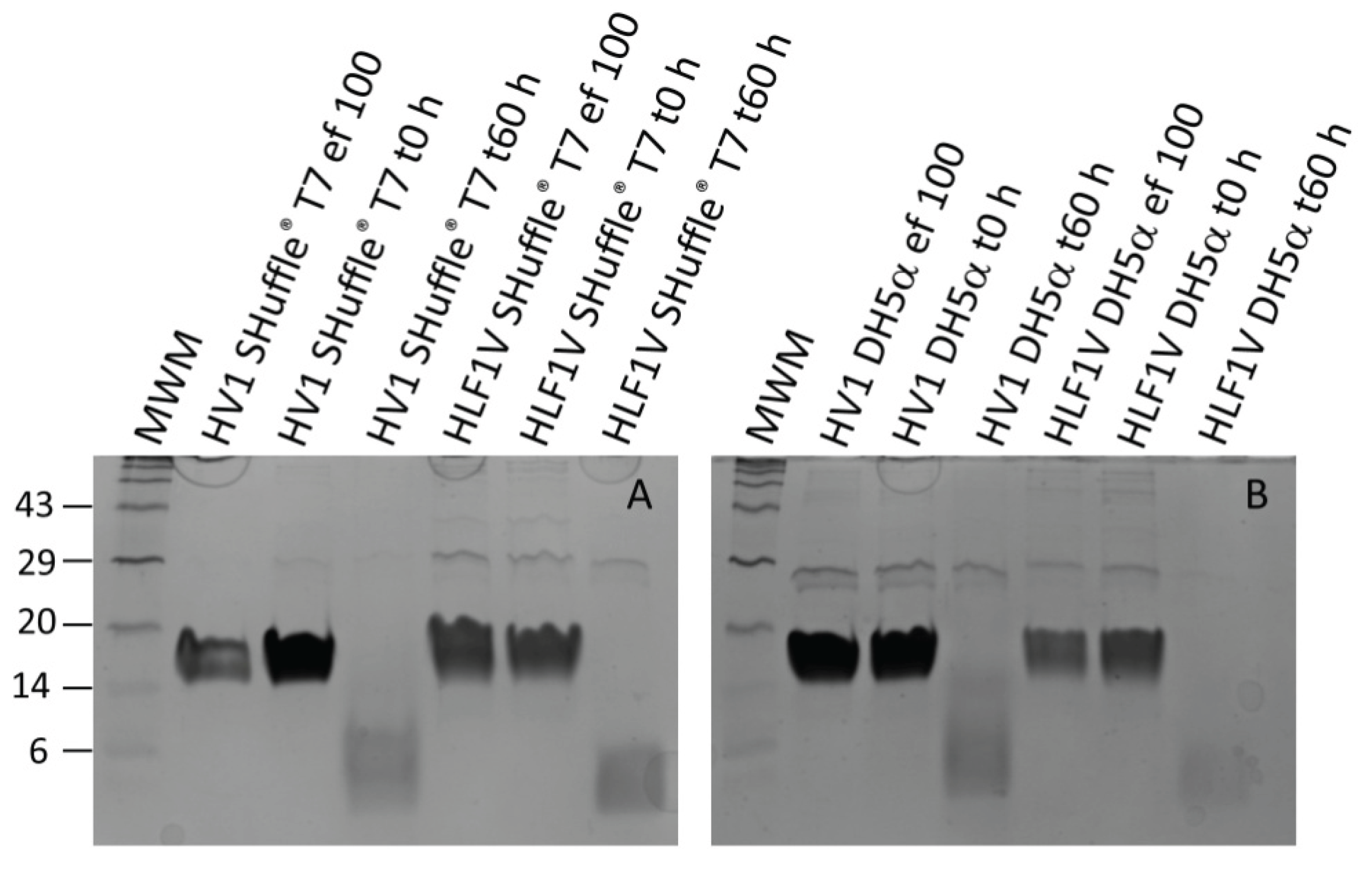
Figure 3.
Thrombin time coagulation assays of recombinant hirudin HV1 and HLF1V preparations after expression in E. coli strains SHuffle® T7 (SH) and DH5α. Results are means of three independent measurements.
Figure 3.
Thrombin time coagulation assays of recombinant hirudin HV1 and HLF1V preparations after expression in E. coli strains SHuffle® T7 (SH) and DH5α. Results are means of three independent measurements.
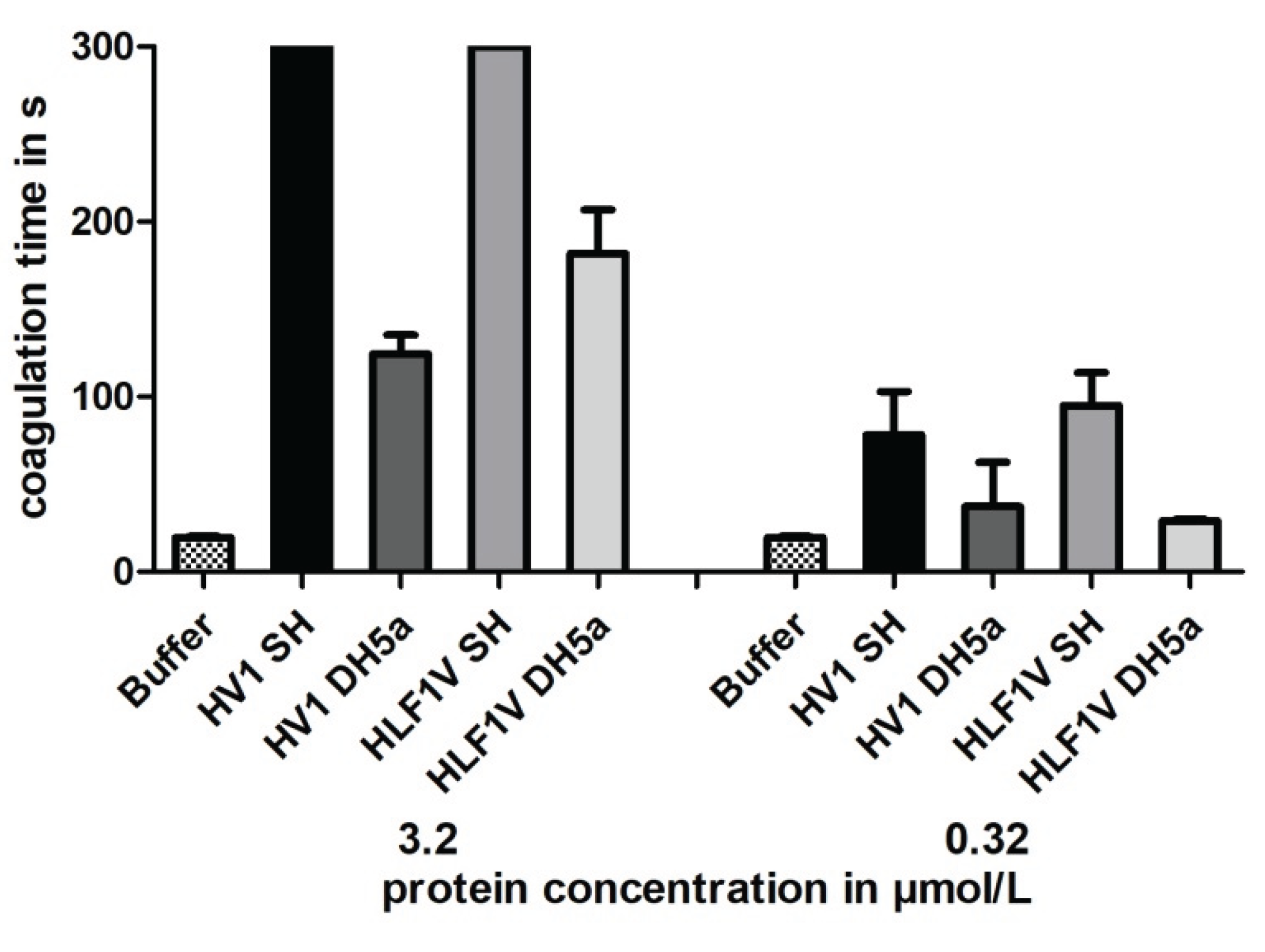
Figure 4.
Thrombin time coagulation assays of recombinant hirudin HV1 preparations after cultivation and expression in E. coli strains DH5α (A) or SHuffle® T7 (B) at 22, 30, 37 or 42 °C, respectively. Results are means of three to six independent measurements.
Figure 4.
Thrombin time coagulation assays of recombinant hirudin HV1 preparations after cultivation and expression in E. coli strains DH5α (A) or SHuffle® T7 (B) at 22, 30, 37 or 42 °C, respectively. Results are means of three to six independent measurements.
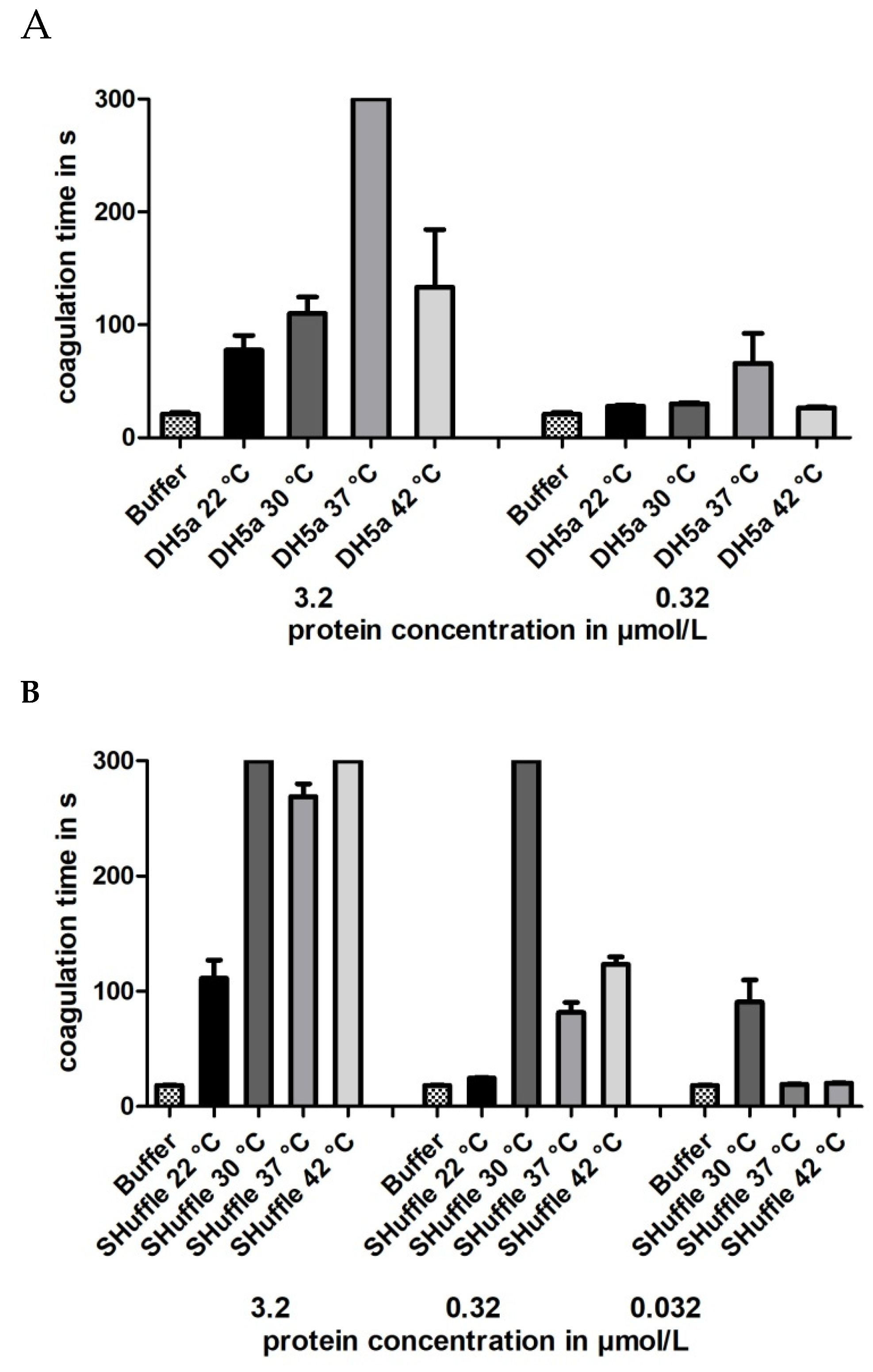
Figure 5.
SDS-PAGE analyses of elution fractions of hirudin HV1 after expression in P. pastoris GS115 clone 2 (A, B) and clone 29 (C, D) cultivated at either 20 °C (A, C) or 30 °C (B, D). HLF1V (B, D). The red arrow mark the HV1 protein. The numbers on the left side indicate the molecular mass in kDa of the respective molecular weight marker (MWM) bands. The numbers 5–500 indicate the imidazole concentrations in mmol/L in the respective elution fractions.
Figure 5.
SDS-PAGE analyses of elution fractions of hirudin HV1 after expression in P. pastoris GS115 clone 2 (A, B) and clone 29 (C, D) cultivated at either 20 °C (A, C) or 30 °C (B, D). HLF1V (B, D). The red arrow mark the HV1 protein. The numbers on the left side indicate the molecular mass in kDa of the respective molecular weight marker (MWM) bands. The numbers 5–500 indicate the imidazole concentrations in mmol/L in the respective elution fractions.
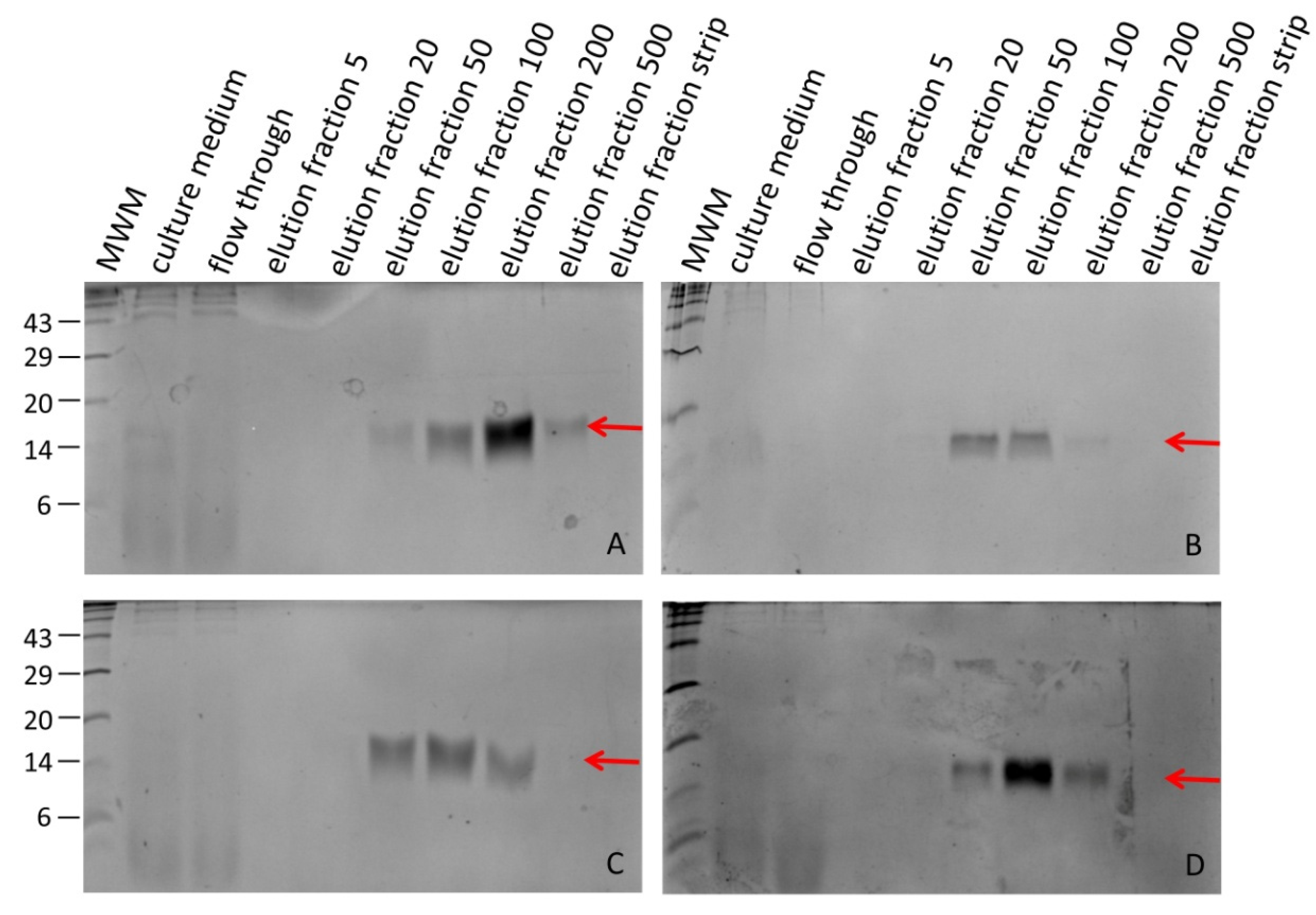
Figure 6.
Thrombin time coagulation assays of recombinant hirudin HV1 preparations after cultivation and expression in P. pastoris GS115 clones 2 and 29 at either 20 °C or 30 °C, respectively. Results are means of three to six independent measurements.
Figure 6.
Thrombin time coagulation assays of recombinant hirudin HV1 preparations after cultivation and expression in P. pastoris GS115 clones 2 and 29 at either 20 °C or 30 °C, respectively. Results are means of three to six independent measurements.
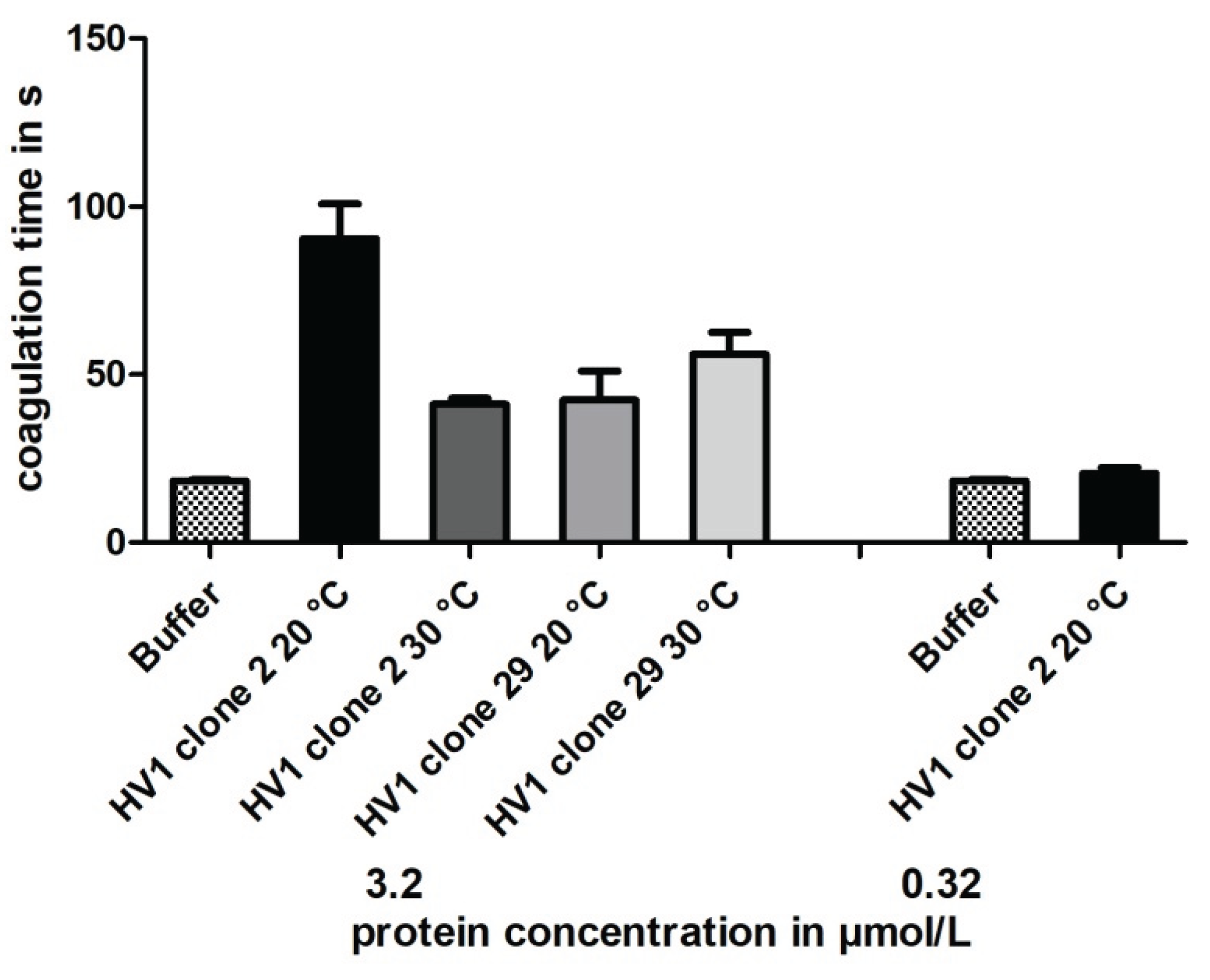

Table 1.
Concentrations of recombinant hirudin HV1 and HLF1V preparations after expression in E. coli strains SHuffle® T7 and DH5a.
Table 1.
Concentrations of recombinant hirudin HV1 and HLF1V preparations after expression in E. coli strains SHuffle® T7 and DH5a.
| Expression system | Hirudin HV1 | HLF1V |
| SHuffle® T7 | 132.4 µmol/L 931 µg/mL |
154.9 µmol/L 1011 µg/mL |
| DH5α | 190.9 µmol/L 1341 µg/mL |
128.2 µmol/L 842 µg/mL |
Table 2.
Concentrations of recombinant hirudin HV1 after expression in P. pastoris GS115 clone 2 and clone 29 at 20 °C or 30 °C, respectively.
Table 2.
Concentrations of recombinant hirudin HV1 after expression in P. pastoris GS115 clone 2 and clone 29 at 20 °C or 30 °C, respectively.
| Cultivation temperature | Clone 2 | Clone 29 |
| 20 °C | 59.3 µmol/L 416 µg/mL |
50.7 µmol/L 356 µg/mL |
| 30 °C | 11.3 µmol/L 79 µg/mL |
34.3 µmol/L 241 µg/mL |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



