Submitted:
03 June 2025
Posted:
04 June 2025
You are already at the latest version
Abstract
The growing global prevalence of non-communicable diseases (NCDs) has gained increasing attention to the health-promoting potential of whole grain dietary fibers. Whole grains are rich sources of both soluble dietary fiber (SDF) and insoluble dietary fiber (IDF), contributing distinct physicochemical properties and playing vital roles in promoting human health. This review provides a comprehensive analysis of the dietary fiber composition of various whole grains including wheat, oats, barley, rye, corn, sorghum, and rice, highlighting their structural characteristics, physiochemical properties, and associated health benefits. The physicochemical properties of dietary fibers such as solubility, water and oil holding capacity, viscosity, swelling ability, and bile acid binding capacity contribute significantly to their technological applications and potential health benefits, particularly in the prevention of NCDs. Despite strong evidence supporting their health benefits, global whole grain intake remains below recommended levels. Therefore, promoting whole grain intake and developing fiber rich functional foods are essential for enhancing public health and preventing chronic diseases. Future research should focus on enhancing the bioavailability and functionality of whole grain dietary fibers, optimizing its extraction methods, and exploring its potential applications in the food and pharmaceutical industries.
Keywords:
Whole grains
; dietary fiber
; physicochemical properties
; non-communicable diseases
1. Introduction
The rising prevalence of non-communicable diseases (NCDs) has led to increasing global attention on healthy dietary choices. Previous studies have shown that inadequate dietary fiber intake is closely associated with a higher risk of NCDs, including cardiovascular diseases, cancers, gastrointestinal disorders, and type 2 diabetes. Dietary fiber is a type of edible, non-starch group of polysaccharides that cannot be digested and absorbed by endogenous enzymes in the human small intestine. However, these fibers are partially or completely fermented in the large intestine by beneficial microbiota, contributing to various health benefits [1].
Whole grain products are considered grains that contain the relative proportions of bran, germ, and endosperm similar to those of untreated grains, and they play a vital role in healthy, sustainable diets due to their rich content of plant-based dietary fibers. In contrast, refined grains undergo processing that removes the bran and the germ, leaving only the starchy endosperm, which significantly reduces their nutritional composition compared to the original whole grain products. Whole grain consumption is associated with numerous health benefits due to its higher dietary fiber content, including soluble and insoluble fibers. These health benefits are largely associated with the physicochemical properties of dietary fibers and their fermentation in the large intestine [2,3].
Epidemiological studies suggest that consuming at least 50 g of whole grains can lead to significant reductions in health risks, including a 25% lower risk of type 2 diabetes, a 20% reduction in cardiovascular mortality, a 12% decrease in cancer-related mortality, and a 15% reduction in overall mortality [4]. However, guidelines for the recommended intake of whole grain products vary across the world. According to the US food guide, individuals consuming a 2,000-calorie diet are advised to consume approximately 85 grams of whole grains per day. Within the European Union, Germany reports one of the highest whole grain intakes, with an average daily consumption of 127.4 g for males and 132.6 g for females. Denmark recommends a minimum daily intake of 75 g, while Norway suggests a daily intake of 80 to 90 grams [2,4]. Despite these recommendations, global whole grain consumption remains below the recommended levels in many countries, highlighting the need for increased awareness of their health benefits.
While previous studies have extensively focused on the health benefits of whole grains, there is a lack of comprehensive analysis of the different dietary fiber fractions in various whole grains and their physicochemical properties associated with health benefits. Therefore, this review aims to provide a thorough analysis of the dietary fiber composition of different whole grains, their physicochemical properties, and their functional roles in the prevention of various NCDs.
2. Dietary Fiber in Whole Grains
2.1. Structure of Whole Grains
According to the Cereals and Grains Association, whole grains consist of the intact, ground, cracked, flaked, or otherwise processed kernel, excluding inedible parts such as the hull and husk. In whole grains, all anatomical components, including the endosperm, germ, and bran must be present in the same relative proportions as in the intact kernel [5]. Examples of whole grains include wheat, rice, maize, oats, rye, barley, triticale, sorghum, and millet, all of which belong to the grass family Poaceae. Whole grains have a complex structure characterized by multiple cell layers. It contains a bark-like protective hull, the endosperm, bran, and germ. The germ contains the plant embryo, while the endosperm provides nourishment for the growing seedling. Surrounding both the germ and the endosperm is the bran, which serves as a protective outer layer for the developing embryo [6].
The endosperm accounts for 80–85% of the whole grain and is primarily composed of starch and protein. The bran and germ represent approximately 12–18% and 2–3% of the dry grain weight, respectively. The embryo is vital for the germination process, as it contains the embryonic axis and scutellum. The endosperm serves as an energy reserve, storing starch and proteins necessary for germination. The aleurone layer, which forms the outermost part of the endosperm, typically consists of 1 to 3 layers of cells, varying among different whole grains. A significant portion of this layer is often removed during roller milling and, therefore, absent in refined flour. The bran fraction consists of several distinct layers, including the outer pericarp, inner pericarp, testa, and nucellar epidermis (hyaline layer). The inner and outer pericarp is rich in polysaccharides, such as cellulose, lignin, and heteroxylan. The testa is a single cell layer in barley, oats, and rice, while in wheat and rye, it generally has two distinct layers [2].
2.2. Dietary Fiber Composition of Selected Whole Grains
Dietary fibers are defined as the edible parts of plants or analogous carbohydrates that resist digestion and absorption in the human small intestine but undergo complete or partial fermentation in the large intestine. Dietary fiber includes polysaccharides (cellulose, hemicellulose, pectin), oligosaccharides (inulin, fructo-oligosaccharides), lignin, and other associated plant compounds. It can be classified into two categories based on its water solubility: insoluble dietary fibers (IDF) and soluble dietary fibers (SDF). IDF, which includes cellulose, most of hemicellulose, and lignin, primarily serves as a structural component of plant cell walls. In contrast, SDF comprises various non-cellulosic polysaccharides and oligosaccharides, mainly arabinoxylans and beta-glucans (Figure 2) [7,8].
The total dietary fiber (TDF) content of wheat ranges from 9% to approximately 38% on a dry weight basis and includes both insoluble and soluble fractions. In wheat, the fiber in bran is primarily insoluble, whereas soluble is more prevalent in white flour. The endosperm tissues, including the aleurone and starchy endosperm, contain high proportions of arabinoxylans (60–70%), moderate amounts of (1,3:1,4) β-D-glucans (20–30%), small amounts of cellulose and glucomannan, and no lignin. In contrast, the outer pericarp tissue of wheat has a cell wall composition rich in cellulose (30%), lignin (12%), and xylans (60%), but contains only limited amounts of β-glucans [2].
In oats, the TDF content typically ranges from 60% to 80%, with approximately 30% comprising SDF, which is primarily located in tissues outside the aleurone layer. Oat bran, which accounts for approximately 30–50% of the whole grain, contributes significantly to the dietary fiber content. β-glucan with (1-3), (1-4)-β-D-glucoside linkages, is the primary SDF, followed by arabinoxylans. In contrast, lignin and cellulose constitute the predominant IDF fraction [9,10]. A previous study investigating 43 oat genotypes reported that dietary fiber content varies significantly by genotype, with some varieties differing by up to 20% in measured fiber content [11,12]. Notably, there are cultivated oat varieties known as hull-less oats, which contain similar quantity as other types, but the husk separates much more easily during harvest [13]. Therefore, selecting dietary fiber-rich oat varieties can offer significant benefits to both food producers and consumers, especially in terms of their physicochemical and antioxidant properties [14].
Barley contains approximately 20-30% of TDF, with β-glucans and arabinoxylans being the most important components [15]. SDF, primarily β-glucans, is abundantly found in the endosperm cells. However, mixed linkage β-glucans and arabinoxylans are also present in the aleurone cell walls. These two groups of compounds are distributed throughout the whole grain, although previous studies have indicated that in hull-less barley, arabinoxylans are primarily concentrated in the outer bran layer and aleurone cells. IDF fraction- including hemicellulose (in part), cellulose, and lignin- is predominantly located in the bran [2,15]. Cellulose has a linear structure composed of glucose units linked by β-1,4-glucosidic bonds, forming microfibrils. This microfibrils are typically associated with hemicellulose and lignin in plant cell walls. Barley hemicellulose is a heterogenic complex, composed a various monosaccharide, including xylose, arabinose, glucose, and mannose. Unlike cellulose, hemicellulose does not consist solely of β-1,4- linkages; instead, it features a range of linkages such as β-1,4- β-1,3, and β-1,6, resulting in a more branched and amorphous structure. This branched structure contributes to its greater solubility in water, in contrast to the insoluble nature of cellulose and lignin. The presence of arabinosyl substituents on the xylan backbone has been shown to hinder the enzymatic degradation of arabinoxylans by endoxylanase enzymes. This enzymatic resistance, particularly due to xylose substitution, may significantly influence the malting process of barley [16].
Rye is distinguished from other cereals due to the highest dietary fiber content, ranging from 15-22% on a dry matter basis [17,18]. IDF fraction accounts for approximately 10%, while low molecular weight SDF expressed as an indigestible oligomer with a degree of polymerisation (<10%) comprises about 5%. High molecular weight SDF makes up around 4% [19]. The main non-starch polysaccharides in rye include arabinoxylans, mixed-linkage glucans, arabinogalactans, cellulose, and lignin [17]. Arabinoxylan, a major component of the rye cell wall (approximately 1.3–1.4%), particularly its water-extractable fraction, plays a significant role in bread dough formation [20,21]. β-glucan is present in rye in lower quantities (1.3-2.2%) compared to oats (2.2-7.8%) and barley (3.5-11.3%) [21]. The ratio of cellotriosyl and cellotetraosyl units in rye and barley is higher than in oats, which may influence the functional properties of the SDF fraction [21].
Sorghum has been reported to contain approximately 19% total dietary fiber (TDF), providing about 6.7 grams of dietary fiber per 100 grams, which aids digestion by adding bulk to the diet and helping to prevent constipation [22,23]. These findings are consistent with a previous study that reported a dietary fiber content of 10.51% [23]. Of the TDF in sorghum, 35% to 48% is composed of IDF. Dietary fibers in sorghum are primarily located in the cell walls of the pericarp and endosperm. Arabinoxylans and β-glucans are the main types of dietary fiber present, accounting for approximately 55% and 40% of the TDF, respectively. Additionally, the cell wall components of sorghum have been reported to contain around 20% lignin [24,25].
Corn fiber has been reported to contain approximately 70% carbohydrates, including cellulose, xylan, residual starch, and lignin [26]. Its composition typically includes 25%–35% hemicellulose, 16%–20% cellulose, 25%–35% starch, 2%–3% lignin, and 8%–10% protein. However, these proportions can vary depending on the corn variety, maturity stage, and processing methods. A key component of hemicellulose in corn fiber is arabinoxylan, which accounts for about 30% of the hemicellulose content. Corn arabinoxylan is predominantly located in the outer layers of the kernel, such as the bran and pericarp. Arabinoxylans are primarily composed of D-xylose (48%–54%) and L-arabinose (33%–35%), with smaller amounts of glucose (7%–11%) and glucuronic acid (3%–6%). The arabinose-to-xylose ratio, typically around 1:4 or 1:5, plays a crucial role in determining the solubility, functionality, and interactions of arabinoxylan in various applications, including gel formation and fermentability by gut microbiota. Cellulose, a major structural component of the corn cell wall, along with its improved forms—such as microcrystalline cellulose and carboxymethyl cellulose—is widely utilized in the food industry to enhance the texture, stability, and mouthfeel of food products. [15].
Rice is unofficially categorized into white, brown, and black varieties based on the content and composition of bioactive substances. Brown and black rice varieties contain higher levels of dietary fiber, lipids, and bioactive phytochemicals, including phytosterol-enriched lipids, dark pigments such as anthocyanins, and phenolic compounds with strong antioxidant activity. [21,26]. In rice, dietary fiber is primarily located in the bran. Its content ranges from approximately 0.7–2.7% in white rice, 2.9–4.4% in brown rice, and around 6% in unpolished black rice.[27,28]. Recent studies have shown that the dietary fiber content, particularly the IDF fraction, is in the following order: black rice > brown rice > white rice. During the milling process, the bran is largely removed, reducing the health benefits of rice while generating food by-products [29]. It is worth noting that bran, as a main by-product of rice milling, is composed of 20.5-33.9% dietary fiber, and therefore the most extracted fraction is water insoluble, which consists of cellulose, hemicellulose, and lignin with small amounts of SDF. [30,31]. Consequently, numerous studies have analyzed the composition and physical properties of dietary fiber in rice bran [30,32,33]. White and glutinous rice varieties are reported to have a higher proportion of the SDF fraction. The highest SDF content has been found in white rice (16.39%), whereas the highest IDF content was observed in brown rice (16.51%) [34]. The IDF fraction is predominantly composed of the following saccharides: xylose (42–45%), glucose (26–32%), arabinose (8–13%), and galactose (10–11%), which reflects the presence of cellulose and arabinoxylan. In contrast, the SDF fraction is rich in xylose (37–42%), glucose (15–33%), galactose (17–19%), and arabinose (9–14%), due to the high concentrations of arabinoxylan and arabinose-rich pectic substances.
3. Physicochemical Properties of Dietary Fiber
Dietary fibers from whole grains exhibit a complex structure that significantly influences their various functions in the gastrointestinal tract. These functions are largely determined by their physiochemical properties. Processing methods applied to whole grains can alter both the composition and physicochemical characteristics of dietary fibers. These properties are influenced by various factors, including extraction methods, food sources, molecular structure, and particle size [35,36]. Several researchers have reported a range of physicochemical properties of dietary fiber, including water solubility, water-holding capacity, swelling ability, viscosity, gel-forming ability, bulking capacity, and binding ability [35,36].
3.1. Solubility
The solubility of dietary fibers plays a critical role in determining their technological functionality in food systems. Based on their solubility in water, dietary fibers are primarily classified as SDF and IDF. Solubility is influenced by several factors, including molecular branching, the presence of ionic groups, and specific positional bonding within the fiber structure. Compared to insoluble fibers, soluble fibers are known for their ability to increase viscosity and enhance emulsifying properties, making them particularly valuable in food processing applications [36]. Several studies have highlighted variations in the solubility of dietary fibers from different whole grains. In one study, the solubility of dietary fiber from oats and wheat was reported to be approximately 3.4% and 4.2%, respectively [37]. Solubility has also been observed to increase during processes such as germination. In whole wheat, for example, solubility has been reported to increase by 18.45 ± 0.23 % after germination [14]. The grinding process can alter the solubility of dietary fiber and is often employed as an effective method to enhance the functional properties of food products. In general, increasing the degree of fiber fragmentation tends to raise the content of SDF while reducing the proportion of IDF [38].
3.2. Water-Holding Capacity (WHC)
WHC refers to the amount of water retained by a known weight of dry fiber under defined conditions. The WHC of dietary fibers depends on several factors, including the molecular structure and the nature of water-binding sites within the fiber matrix [8]. The WHC of dietary fiber is crucial in various food applications, as it significantly influences the shelf life and texture of food products. According to Ahmad et al., β-glucan extracted using hot water treatment exhibited the highest WHC value (3.79 g/g), followed by β-glucan extracted through alkali and acid treatments [39]. In another study, quinoa bran exhibited higher WHC compared to oat and rice hulls [40]. Processing methods such as grinding can also affect WHC. For example, the WHC of oats increases from 3.53 to 5.23 g/g with superfine grinding. This improvement is attributed to the increased specific surface area of the sample, which enhances its contact with water. [41]. Moreover, the germination process in whole wheat increased the WHC from 2.12 ± 0.19% to 2.75 ± 0.23%, which is probably related to the increase of SDF fraction in TDF [37].
3.3. Oil Holding Capacity (OHC)
OHC refers to the amount of oil retained by fibers after mixing and centrifugation. This property is important for minimizing fat loss during cooking and may also aid in the removal of excess fat from the body. [8]. Numerous studies have demonstrated the OHC of whole grain fibers such as quinoa, wheat, oats, and rice bran [42,43]. A previous study found that SDF from quinoa bran SDF exhibited a higher OHC (1.85 ± 0.16 g/g to 2.95 ± 0.23 g/g) compared to the orange peel fiber and oat hull [40]. Another study reported that wheat fiber had the highest oil retention capacity (4.98 mL/g), followed by oat fiber (3.27 mL/g) [44]. Additionally, Dhillon et al. have found that OHC of whole wheat flour increased with germination, and reported that OHC of whole wheat flour 2.32 ± 0.21 g/g rose to 3.78 ± 0.43 g/g after germination [37]. The research done by other authors further demonstrates that dietary fibers derived from oats, whole wheat flour, millet bran, and rice bran possess a high WHC and a low OHC, suggesting their potential as a dietary resource for the development of functional food products that address water synergism in formulated foods and act as emulsifiers for high-fat foods [37,42,43].
3.4. Viscosity and Gel Formation
Viscosity (η) refers to a fluid's resistance to flow and is mathematically defined as the ratio of shear stress (τ) to shear rate (γ̇). Viscosity plays a significant role in increasing enteric viscosity, which can slow intestinal transit, delay gastric emptying, and reduce the absorption of glucose and sterols in the intestine. These effects contribute to lower serum cholesterol, postprandial blood glucose, and insulin levels. These health benefits are primarily associated with viscous SDF [45,46]. In whole grains, soluble dietary fiber (SDF) plays a crucial role in enhancing the viscosity of solutions. In contrast, insoluble fibers, such as cellulose found in rice bran and wheat bran, generally contribute less to viscosity due to their inability to dissolve in water and form gel-like structures [46].
A previous study found that oat dietary fiber, particularly β-glucan, has the ability to lower glycemic responses, primarily due to its role in increasing viscosity within the upper digestive tract. The viscosity of β-glucan samples ranged from 34.30 to 52.82 cP, depending on the extraction method. The highest viscosity (52.82 cP) was observed in enzymatically extracted β-glucan, while the lowest viscosity was found in alkali-extracted β-glucan. Additionally, factors such as temperature and pH of the solution also significantly influence the viscosity of β-glucan solutions. [39]. Soluble fibers such as β-glucan form gels that increase the viscosity of gastrointestinal contents. For example, long-chain polymers like guar gum and tragacanth gum retain large amounts of water and exhibit high viscosity, whereas highly soluble fibers like gum arabic display low viscosity despite their solubility [46].
3.5. Bile Acid Binding Capacity
Cholesterol serves as a precursor for bile acid synthesis in the liver. Dietary fibers can bind to bile acids, promoting their excretion from the body. This loss stimulates the conversion of more cholesterol into bile acids, thereby lowering blood cholesterol levels and reducing the risk of heart disease [28]. The gel formed by SDF in the presence of Ca²⁺ can encapsulate bile acids. Various water-soluble polysaccharides contribute to bile salt binding, primarily due to the presence of functional groups such as –COH, –C=O, –O–, –COOH, and –OH, which exhibit high affinity for cholesterol and bile salt structures. Additionally, lignin- a highly branched component of IDF- also plays a significant role in this mechanism, particularly due to its abundance of hydroxyl and carbonyl groups [47].
A previous study reported that the bile acid binding capacity of dietary fiber from the rice cultivars Mushq Budij and SR-4 was 18.10 μmol/g and 23.24 μmol/g, respectively [28]. A study by Liu et al. explored the bile acid-binding capacity of soluble dietary fiber extracted from various colored quinoa varieties [40]. Bile acids such as cholic acid (CA) and chenodeoxycholic acid (CDCA) play a vital role in lipid metabolism. The study found no significant differences in CA adsorption capacity among the different SDF samples extracted from quinoa. However, the adsorption capacity for CDCA varied significantly among the samples [40].
3.6. Swelling Ability
The swelling ability of dietary fiber refers to its capacity to expand or gelatinize when mixed with water and is expressed as the ratio of the fiber's volume after soaking in water to its actual weight. This ability varies depending on the solubility, chemical structure, and source of the dietary fiber, as well as additional factors such as temperature and ionic charge [48]. According to a previous study, the swelling ability of wheat bran dietary fiber increased from 2.14 to 2.60 mL/g after grinding. This improvement may be attributed to structural changes in the dietary fiber induced by the grinding process [49]. In another study, the swelling ability of oat bran dietary fiber increased significantly (p < .05) with extended grinding times. This enhancement is likely due to the superfine grinding process, which improves the specific surface area of the sample and increases its contact with water. [41]. Other authors have reported that extraction methods using alkaline and acidic treatments significantly affect the water swelling ability of dietary fiber. In both cases, the fibers extracted using alkaline treatment—both SDF and IDF—exhibited the highest swelling values. This is attributed, in the case of SDF, to a loose microstructure, reduced particle size, and consequently, a larger surface area for interaction with water molecules. For IDF, the strong alkali treatment breaks down cellulose chains into shorter polymers and disrupts the ether linkages between hemicellulose and lignin, enhancing water absorption capacity [48]. In food processing, enhanced swelling ability improves functional performance and can better meet the growing public demand for health-oriented food products. Therefore, in the context of whole grain-based foods, the physical properties of dietary fiber are critical in supporting overall health benefits.
4. Health Benefits of Whole Grain Dietary Fiber
4.1. Prevention of Cardiovascular Disease
Elevated levels of low-density lipoprotein cholesterol (LDL-C) and postprandial hyperglycemia are key risk factors contributing to the development of atherosclerotic cardiovascular disease (CVD). While synthetic medications such as statins are effective in managing CVD, their long-term use can be associated with high costs and potential side effects, particularly when consumed in large doses over extended periods [50]. Consequently, dietary fibers are receiving growing attention for their potential health benefits, especially in the prevention and management of CVD. A substantial body of research has examined the protective effects of dietary fibers derived from whole grain cereals against CVD, highlighting their role in modulating lipid profiles, reducing blood glucose levels, and improving overall cardiovascular health [51,52,53]. Compared to IDF, SDF is more frequently associated with reductions in serum low-density lipoprotein cholesterol (LDL-C) and is often linked to a lower risk of cardiovascular disease (CVD). SDF in whole grains can form viscous gels within the gastrointestinal tract, increasing gut viscosity. This elevated viscosity impedes the reabsorption of bile acids, thereby promoting their excretion and reducing the enterohepatic circulation of cholesterol. As a result, hepatic cholesterol is redirected for bile acid synthesis, contributing to a decline in circulating cholesterol levels [54].
A cohort study involving 7,469 participants with CVD demonstrated that increased dietary fiber intake was associated with significant reductions in both total cholesterol and LDL. Specifically, total cholesterol decreased by 0.42 mmol/L (95% CI: −0.78 to −0.05), while LDL cholesterol decreased by 0.47 mmol/L (95% CI: −0.85 to −0.10) [55]. Moreover, several studies have demonstrated the potential of oats and oat-based products to improve blood lipid profiles and lower blood pressure by modulating insulin metabolism in individuals with mild hypercholesterolemia [56,57,58]. β-glucans present in oats have been shown to reduce cholesterol absorption in the small intestine by forming a viscous gel that binds to cholesterol molecules. Additionally, a previous study reported that dietary fiber supplementation with oat bran was associated with a reduction in heart rate among patients with hypertension [57]. Experimental studies on rats have demonstrated that enzyme-treated wheat bran-rich in arabinoxylans, cellulose, and lignin- significantly reduced body weight and hepatic triglyceride content when administered to five-week-old mice [59].
Soluble fiber is suggested to reduce cholesterol synthesis by modulating serum concentrations of hormones and short-chain fatty acids (SCFAs), such as acetate, propionate, and butyrate, which influence lipid metabolism. Notably, butyrate has been shown to significantly inhibit de novo cholesterol synthesis in isolated rat hepatocytes [60]. These findings suggest that increasing dietary fiber intake, particularly from whole grain sources, may represent an effective nutritional strategy for the prevention and management of CVD.
4.2. Prevention of Type 2 Diabetes Mellitus
Diabetes mellitus is a non-communicable metabolic disorder characterized by the body's inability to produce or effectively utilize insulin. Type 2 diabetes mellitus (T2DM), which results from inadequate insulin production or insulin resistance, accounts for approximately 90% of diabetes cases in the adult population [2]. Whole grain dietary fibers play a critical role in the prevention of T2DM, largely due to their association with a low glycemic index, increased satiety, and reduced body weight. Regular consumption of at least 15.9 g of dietary fiber per day has been shown to improve glycemic control by lowering fasting blood glucose levels, postprandial insulin concentrations, and serum triglyceride levels [61].
Previous studies have demonstrated that SDF present in whole grains can increase the viscosity of gastric contents. This elevated viscosity significantly contributes to reduced glucose absorption by slowing gastric emptying and decreasing the rate of starch digestion, thereby delaying the postprandial rise in blood glucose levels [2,62]. The increased viscosity induced by SDF may also result in the formation of a ‘thick layer’ surrounding food particles, thereby limiting the accessibility of digestive enzymes to the inner components of the food matrix and delaying their interaction with the absorptive surfaces of the gastrointestinal tract [2]. SDF plays a vital role in the management of T2DM, whereas IDF does not directly impact postprandial glucose levels or insulin sensitivity in T2DM [62]. However, some studies suggest that IDF may also have significant potential to reduce the risk of T2DM [62,63]. The beneficial effects of IDF may be attributed to its fermentation by gut microbiota and the subsequent production of bioactive compounds. IDF also acts as a physical barrier, slowing the transit of digestive products across the enterocyte brush border and modulating the activity of digestive enzymes. Additionally, microbial fermentation releases phenolic compounds bound to the fiber matrix, thereby increasing the bioavailability of these phytochemicals. Polyphenols, recognized as potent bioactive compounds, are considered a promising alternative strategy for the prevention and management of T2DM [62].
A higher glycemic index (GI) has been linked to an increased risk of T2DM. Previous studies have reported average GI values for various whole grain products, including 42 for whole grain cold breakfast cereals, 55 for oatmeal and brown rice, and a range of 27 to 70 for whole grain breads. This variability in GI for whole grain breads depends largely on the main ingredients used, such as barley, buckwheat, oats, and rye [64]. Furthermore, a comprehensive systematic review of eleven prospective cohort studies, encompassing 463,282 participants and 37,249 type 2 diabetes cases, found that a higher daily intake of whole grains was associated with a reduced risk of developing T2DM [65]. Specifically, higher oat intake (more than 5.7 g/day) was significantly associated with lower risk of T2DM [66]. Oat-based products, such as oat flakes, oatmeal, and oat bran generally have a lower glycemic index compared to similar products made from wheat, barley, or corn. This effect is largely attributed to β-glucans, the primary component of oat fiber. β-glucan is a water-soluble, non-digestible polysaccharide that is not absorbed in the small intestine. By increasing the viscosity of the alimentary bolus in the upper gastrointestinal tract, β-glucan slows nutrient absorption and reduces postprandial glucose spikes [66]. Similarly, dietary fiber from millet bran has demonstrated the ability to reduce fasting blood glucose (FBG) levels in rat models, further highlighting the potential of whole grain fibers in glycemic control [67]. Wheat bran dietary fibers have also demonstrated significant benefits in diabetic models by increasing serum insulin content. Specifically, intake of modified wheat bran dietary fibers significantly elevated serum insulin levels and appeared to inhibit hepatic gluconeogenesis by suppressing the 1,2-DAG-PKCε signaling pathway. This suppression enhanced insulin receptor activity and insulin signal transduction, collectively resulting in reduced blood glucose levels in diabetic mice [68].
4.3. Control of Obesity
Obesity is a major global health concern, impacting both developed and developing countries, and significantly increasing the risk of CVD, T2DM, and certain types of cancer [69]. According to the World Health Organization (WHO), more than 1 billion people worldwide were living with obesity in 2023, including approximately 650 million adults and 39 million children [70]. The primary cause of obesity is an imbalance between energy intake and expenditure, driven by excessive calorie consumption from food and beverages combined with insufficient physical activity. Effective obesity management requires creating a negative energy balance, which reduces fat stores while preserving lean body mass. Treatment plans should be personalized, taking into account factors such as age, sex, degree of obesity, and individual health risks. Various approaches exist to tackle this global epidemic, with lifestyle modification playing a central role in all weight management strategies. Behavioral changes that promote self-control of daily energy balance through diet and physical activities are essential for long-term success and, importantly, for maintaining body weight [71,72].
Compared to refined grains, whole grains contain fewer starches and calories but are richer in micronutrients and phytochemicals, offering significant health benefits [73]. Previous studies have shown significant inverse associations between whole grain fiber intake and measures of body weight [74,75]. A previous meta-analysis found that higher whole grain intake was significantly associated with lower body mass index (BMI), showing a weighted slope of −0.0141 kg/m² per gram per day (95% CI: −0.0207 to −0.0077; r = −0.526, P = 0.0001) [76]. Similarly, another study reported a significant reduction in BMI and fat mass among overweight and obese adults after a whole-grain diet intervention over a defined period [77]. A 2019 meta-analysis of observational studies and randomized controlled trials found that higher whole-grain consumption is significantly associated with lower BMI values. Analysis of cross-sectional data from 12 studies, involving 136,834 participants, indicated a significant inverse correlation between whole-grain intake and BMI [78]. In another study involving 5,094 randomly selected Finnish adult men, a significant inverse association was observed between whole grain intake and BMI (P < 0.001), waist circumference (P < 0.001), and total cholesterol levels (P = 0.02) [79].
The naturally high fiber content in most whole-grain foods may help prevent weight gain by enhancing appetite control and slowing carbohydrate absorption. Additionally, the presence of multiple enzyme inhibitors within the whole grain fiber matrix may directly influence metabolic efficiency, providing another mechanism through which whole grains can positively impact body weight. Conversely, the high insulin concentrations often associated with consuming low-fiber refined grains may, over the long term, promote weight gain by shifting metabolic fuels from oxidation to storage [80,81].
4.4. Gastrointestinal Health
Whole grains are an important source of dietary fiber, which positively impacts digestive health by increasing stool bulk, accelerating intestinal transit, promoting the production of short-chain fatty acids (SCFAs), and modulating gut microbiota composition [3]. Recent studies have demonstrated that β-glucan consumption enhances the production of SCFA and positively modulates gut microbiota composition. For example, Thandapilly et al. reported that high molecular weight barley β-glucan significantly increased fecal SCFA concentrations in individuals with mild hypercholesterolemia [82]. Additionally, Pi et al. found that hydrolyzed β-glucan promoted the production of SCFAs and gases, also increasing the abundance of beneficial bacteria such as Bifidobacterium and Faecalibacterium, suggesting its potential role in improving gut health and modulating the microbiota [83]. Previous research has shown that whole grain oats, barley, and their β-glucans generally exhibit prebiotic properties by promoting an overall increase in colonic microbial populations and activity, particularly favoring Lactobacillus acidophilus and Bifidobacterium longum [84]. These studies also observed an increase in SCFAs production using porcine and mouse models [85,86]. In another study involving 31 adults with overweight and class-I obesity, supplementation with corn arabinoxylan was found to modulate gut microbiota composition by promoting the growth of Bifidobacterium longum, Blautia obeum, and Prevotella copri. Additionally, it increased fecal propionate concentrations, which are known for their potential metabolic benefits. Compared to arabinoxylans from other sources such as rice, corn, and wheat, corn arabinoxylan yielded the highest production of health-beneficial SCFA [15].
Bifidobacteria and Lactobacilli are key gut microorganisms associated with significant health benefits. Bifidobacterium has been shown to protect against diseases such as colorectal cancer, diarrhea, necrotizing enterocolitis, and inflammatory bowel disease (IBD). It also competitively inhibits pathogens from attaching to epithelial cell binding sites. Lactobacillus plays a protective role in maintaining intestinal barrier integrity by mitigating inflammation, chemical damage, and stress-induced permeability. Additionally, Lactobacillus produces lactate, which is further metabolized into SCFAs. Bacterial fermentation of dietary fibers in the colon generates SCFAs, including acetate, propionate, and butyrate. Bifidobacterium contributes to SCFA production by generating acetate and maintaining a symbiotic relationship with butyrate-producing bacteria. Consumption of dietary fibers, such as those from whole grains, enriches butyrate levels, which are vital for maintaining gut health [87]. Therefore, the consumption of whole grains positively influences the gut microbial community, particularly by promoting the growth of beneficial bacterial groups that promote gastrointestinal health.
Recent studies have confirmed the beneficial effects of whole grains on bowel health. A randomized controlled trial demonstrated that a diet rich in wholegrain fiber enhances fecal bulk and colonic fermentation, resulting in increased stool frequency and higher fecal water content- both indicators of improved bowel function [88]. Additionally, research on barley β-glucan consumption has shown increased fecal bile acid excretion and elevated production of SCFAs, including butyrate. Butyrate plays a crucial role in maintaining colonocyte health and promoting apoptosis in mutated colon cells, thereby supporting colon health and potentially reducing cancer risk [89].
Rye-based foods have been reported to be more effective than whole-wheat and low-fiber foods in increasing plasma enterolactone and fecal butyrate concentrations. The elevated enterolactone levels result from the beneficial microbial fermentation of mammalian lignans in the gut lumen, followed by absorption through the colon wall. Enterolactone is considered a potential biomarker for large bowel health [90]. Additionally, fecal ammonia and p-cresol potentially toxic compounds produced in the large bowel can negatively impact colon wall health. Whole grain dietary fibers have been shown to reduce the colon’s exposure to these harmful substances, thereby promoting improved large bowel health [91].
Dietary fiber, especially rich in phenolic compounds like ferulic acid, is abundant in the bran of the wheat kernel and is released through microbial fermentation in the colon. The presence of ferulic acid in plasma or feces serves as a biomarker for bacterial fermentation activity in the colon [92,93,94]. Additionally, a controlled trial reported the beneficial effects of consuming 70 g of wholegrain wheat (providing 8 g of fiber) daily for 8 weeks, compared to 60 g of refined wheat (providing 2.2 g of fiber). The study demonstrated significant increases in ferulic acid levels, with a 4-fold rise in plasma concentration and a 2-fold increase in fecal excretion in the wholegrain group compared to refined wheat consumers [93].
4.5. Prevention of Cancers
Several studies have suggested that whole grain fibers help to protect the body against various types of cancer, including gastric, colorectal, breast, and prostate cancers [95,96,97,98]. Moreover, higher whole grain intake is inversely associated with weight gain and the risk of T2DM, both of which are established risk factors for cancer development [99]. Moreover, the dietary fibers in whole grains increase fecal bulk and reduce intestinal transit time, which helps dilute carcinogens and limit their absorption by the intestinal epithelium. Resistant starches and oligosaccharides present in whole grains are fermented in the colon to produce SCFAs, including butyrate. These SCFAs serve as a preferred energy source for mucosal cells and exhibit proapoptotic and antineoplastic properties that help to inhibit tumor growth. Additionally, SCFAs lower the intestinal pH, reducing the solubility of free bile acids and thereby decreasing their availability for carcinogenic activity [100,101]. Therefore, dietary fibers in whole grains play a vital role in protecting the body against various types of cancer.
A previous study found that participants in the highest quintile of whole grain intake had a 22% lower risk of developing liver cancer [102]. Additionally, Liu et al. reported that whole grain fiber shows promising potential in breast cancer prevention. Another study demonstrated that barley exhibits anti-tumor activities in both rat mammary tumor models and MCF-7 breast cancer cell lines, through mechanisms including induction of cell cycle arrest, promotion of apoptosis, and inhibition of cell proliferation [102,103]. Numerous studies have documented the anticancer potential of oat β-glucans, demonstrating their effectiveness against various cancer cell lines, both in vitro and in vivo [104,105]. Another study suggested that oat β-glucans are a safe and effective option for managing early-stage colorectal cancer [105].
Dietary fibers in whole grains play a significant role in colorectal cancer prevention, largely due to their interactions with bile acids. Bile acids are amphiphilic molecules essential for lipid absorption in the small intestine and for the formation of micelles that solubilize cholesterol. In the large intestine, gut bacteria metabolize bile acids by deconjugating and dehydroxylating primary bile acids to form secondary bile acids—compounds that have been implicated in colonic carcinogenesis. Several studies have demonstrated that these bile acids are cytotoxic to colonocytes and can stimulate abnormal cell proliferation. Dietary fibers bind bile acids, thereby modifying the gut–liver axis, reducing circulating cholesterol levels, and consequently decreasing the risk of colorectal cancer development [106].
Figure 3.
Health benefits of whole grain dietary fibers.
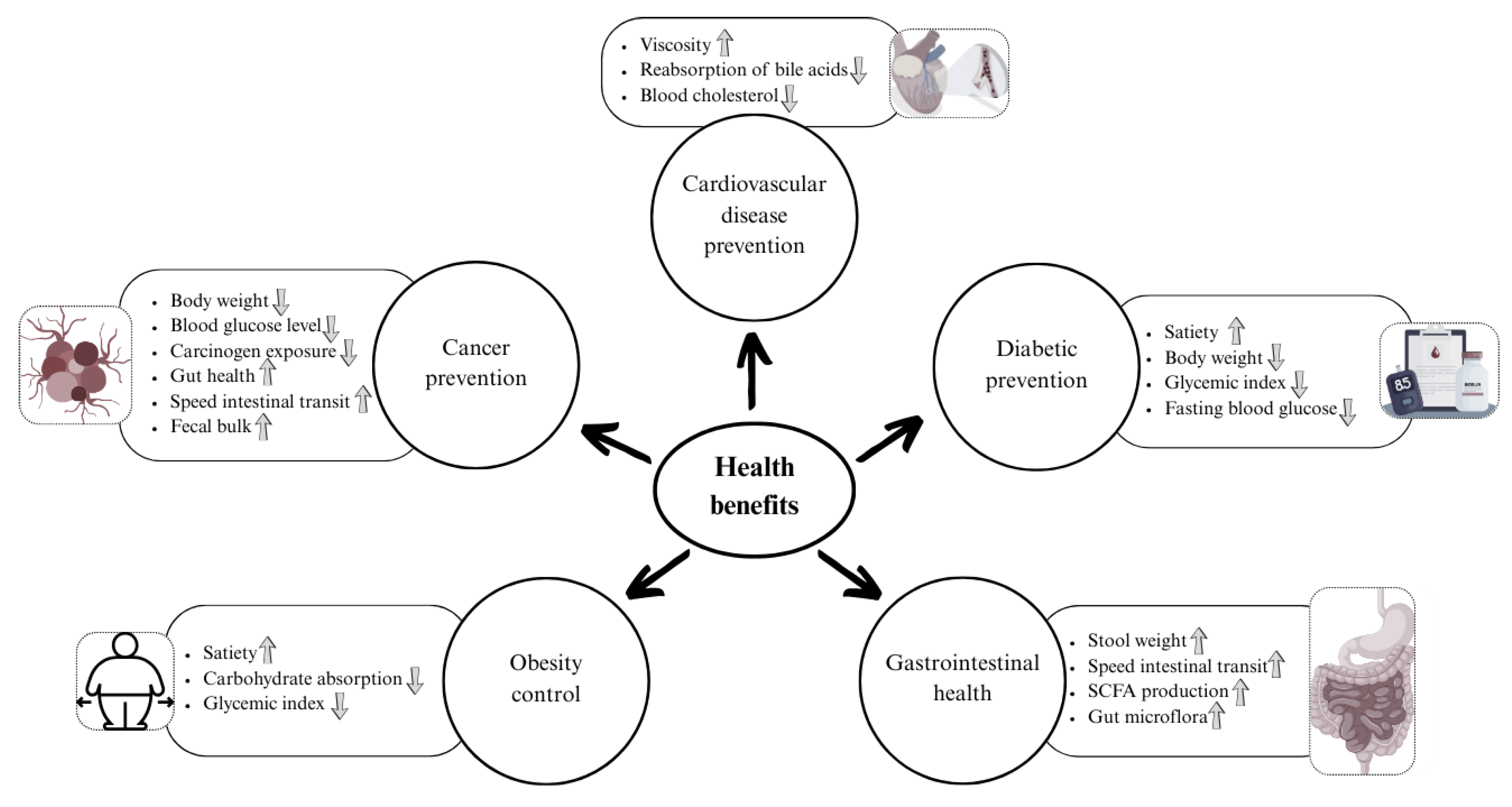

Table 1.
Health benefits of different whole grains.
| Dietary fiber source | Study type | Health benefit | Research findings | Reference |
|---|---|---|---|---|
| Rye | Human intervention | Cardiovascular disease prevention | Total and LDL cholesterols were lowered (-0.06 and -0.09 mmol/L, respectively; P < 0.05) after consumption of whole grain rye with lignan supplements after 4 weeks. | [107] |
| Diabetic control | Rye kernel bread decreased blood glucose (0-120 min, P = 0.001), serum insulin response (0-120 minutes, P<0.05), and fasting FFA concentrations (P<0.05). | [108] | ||
| Rye-based foods decreased postprandial glucose- and insulin responses. | [109] | |||
| Obesity control | Participants who consumed a rye-based diet for the 12-week period had lost 1.08 kg body weight and 0.54% body fat more than the refined wheat consumed group (95% confidence interval (CI): 0.36; 1.80, p < 0.01 and 0.05; 1.03, p = 0.03, respectively). | [110] | ||
| Gastrointestinal health | Induced some changes in gut microbiota composition, including increased abundance of the butyrate-producing Agathobacter. | [111] | ||
| Oat | Human intervention | Cardiovascular disease prevention | Significant reduction in office systolic blood pressure (oSBP; P < 0.001) and office diastolic blood pressure (oDBP; P < 0.028) in the oat bran consumed group (30 g/day of oat bran contains 8.9 g of dietary fiber) compared to the control group after 3-month period. | [58] |
| Consumption of oat dietary fiber reduces levels of systemic chronic inflammation after two weeks post-treatment. | [112] | |||
| Diabetic control | The intake of 5 g of oat β-glucan-enriched diet for 12 weeks can help improve glycemic control, increase the feeling of satiety, and promote changes in the gut microbiota profile. | [113] | ||
| The study demonstrated that a hypocaloric oat-based nutrition diet led to a significant reduction in total insulin dosage and HbA1c levels in insulin-treated outpatients with type 2 diabetes. | [114] | |||
| Obesity control | Oat β-glucan intervention increases the abundance of Lactobacillus and Bifidobacterium. These microbiota alterations contributed to an increase in 7-ketodeoxycholic acid and it enhances bile acid synthesis. | [115] | ||
| Ageing control | A decrease in the Eotaxin-1 protein, an aging-related chemokine, independent of a person’s gender, body mass index, or age. | [112] | ||
| Wheat | Human intervention | Cardiovascular disease prevention | Total and LDL cholesterol were lowered by -0.09 mmol/L at (P < 0.05) after consumption of a wheat-based diet in 40 men with a metabolic syndrome risk profile after 4 weeks. | [107] |
| Obesity control | The study found that consumption of resistant starch-enriched wheat rolls significantly increased fasting and peak concentrations of peptide YY3-36 (PYY3-36), a hormone associated with satiety while decreasing peak concentrations and iAUC of glucose-dependent insulinotropic peptide (GIP), which is involved in hunger regulation. | [116] | ||
| Gastrointestinal health | The study has found that intake of 15 g/day of wheat bran extract increases fecal Bifidobacterium and softens stool consistency without major effects on energy metabolism in healthy humans with a slow GI transit. | [117] | ||
| In vivo study | Gastrointestinal health | The study has found that high amylose wheat (HAW) consumption led to an increase in fecal bacterial load and gastrointestinal health in mice. | [118] | |
| Corn | Human intervention | Cardiovascular disease prevention | Whole grain corn flour significantly decreased LDL cholesterols over time (-10.4 ± 3.6 mg/dL, P = 0.005) and marginally decreased total cholesterol (-9.2 ± 3.9 mg/dL, P = 0.072) over time. | [119] |
| Brown rice | Human intervention | Diabetic control | Improved endothelial function, without changes in HbA1c levels | [120] |
| Whole grains | Human intervention | Diabetic control | Higher intake of whole grain fiber was positively associated with better β-cell function, insulin sensitivity, and postprandial glycemic control. | [121] |
| A systematic review found that increasing whole grain fiber intake improves glycemic control and reduces cardiometabolic risk factors in individuals with prediabetes, type 1, or type 2 diabetes. The study suggests increasing daily fiber intake by 15 g or to a total of 35 g per day to lower the risk of premature mortality and enhance diabetes management. | [122] |
5. Conclusions
Whole grain dietary fibers exhibit a wide range of physicochemical properties including solubility, water and oil holding capacity, swelling ability, viscosity, gel formation, and bile acid binding which contribute not only to their technological utility but also to their role in preventing non-communicable diseases. The dietary fiber content and composition vary significantly among different whole grains such as wheat, oats, barley, rye, corn, sorghum, and rice. The evidence from in vitro, in vivo, and clinical studies highlights the beneficial effects of whole grain dietary fibers against various chronic diseases, including cardiovascular diseases, type 2 diabetes mellitus, obesity, gastrointestinal disorders, and certain cancers through different mechanisms. Despite the strong evidence of the importance of whole grain dietary fibers, global intake of whole grains remains below recommended levels. Therefore, promoting whole grain intake and developing fiber-rich functional foods are essential for enhancing public health and preventing chronic diseases. Future research should focus on enhancing the bioavailability and functionality of whole grain dietary fibers, optimizing their extraction methods, and exploring their potential applications in the food and pharmaceutical industries.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Dietary fiber composition of different whole grains.
Author Contributions
Conceptualization, P.A. and K.D.; methodology, P.A. and K.D.; software, P.M.; resources, P.A., K.D., and J.W.; data curation, P.A., and K.D.; writing—original draft preparation, P.A.; writing—review and editing, K.D. and J.W.; visualization, P.A. and K.D.; supervision, K.D.; project administration, K.D. and J.W.; funding acquisition, J.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
No new data were created or analyzed in this study.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| NCDs | Non-Communicable Diseases |
| IDF | Insoluble Dietary Fibers |
| SDF | Soluble Dietary Fibers |
| TDF | Total Dietary Fibe |
| WHC | Water-Holding Capacity |
| OHC | Oil Holding Capacity |
| CA | Cholic Acid |
| CDCA | Chenodeoxycholic Acid |
| LDL-C | Low-Density Lipoprotein Cholesterol |
| CVD | Cardiovascular Disease |
| SCFA | Short-Chain Fatty Acids |
| T2DM | Type 2 Diabetes Mellitus |
| GI | Glycemic Index |
| WHO | World Health Organization |
| BMI | Body Mass Index |
| IBD | Inflammatory Bowel Disease |
References
- Zhang, S.; Xu, X.; Cao, X.; Liu, T. The Structural Characteristics of Dietary Fibers from Tremella Fuciformis and Their Hypolipidemic Effects in Mice. Food Science and Human Wellness 2023, 12, 503–511. [CrossRef]
- P., N.P. V.; Joye, I.J. Dietary Fibre from Whole Grains and Their Benefits on Metabolic Health. Nutrients 2020, 12, 3045. [CrossRef]
- Fu, J.; Zheng, Y.; Gao, Y.; Xu, W. Dietary Fiber Intake and Gut Microbiota in Human Health. Microorganisms 2022, 10, 2507. [CrossRef]
- European Commission Supporting Policy with Scientific Evidence, Whole Grain Intake across European Countries.
- van der Kamp, J.-W.; Jones, J.M.; Miller, K.B.; Ross, A.B.; Seal, C.J.; Tan, B.; Beck, E.J. Consensus, Global Definitions of Whole Grain as a Food Ingredient and of Whole-Grain Foods Presented on Behalf of the Whole Grain Initiative. Nutrients 2021, 14, 138. [CrossRef]
- Khalid, A.; Hameed, A.; Tahir, M.F. Wheat Quality: A Review on Chemical Composition, Nutritional Attributes, Grain Anatomy, Types, Classification, and Function of Seed Storage Proteins in Bread Making Quality. Front Nutr 2023, 10. [CrossRef]
- Cheng, W.; Sun, Y.; Fan, M.; Li, Y.; Wang, L.; Qian, H. Wheat Bran, as the Resource of Dietary Fiber: A Review. Crit Rev Food Sci Nutr 2022, 62, 7269–7281. [CrossRef]
- Liu, T.; Zhen, X.; Lei, H.; Li, J.; Wang, Y.; Gou, D.; Zhao, J. Investigating the Physicochemical Characteristics and Importance of Insoluble Dietary Fiber Extracted from Legumes: An in-Depth Study on Its Biological Functions. Food Chem X 2024, 22, 101424. [CrossRef]
- Zhu, L.; Wang, L.F.; Bialobzyski, S.; Zijlstra, R.T.; Beltranena, E. Energy and Nutrient Digestibility and Effect of Increasing the Dietary Inclusion of Hull-Less Oats Replacing Wheat Grain on Growth Performance of Weanling Pigs. Anim Feed Sci Technol 2024, 318, 116139. [CrossRef]
- Kang, Z.; Meng, N.; Liu, M.; Liu, Y.; Jiang, P.; Qiao, C.-C.; Tan, B. Enhancement of Physicochemical, in Vitro Hypoglycemic, Hypolipidemic, Antioxidant and Prebiotic Properties of Oat Bran Dietary Fiber: Dynamic High Pressure Microfluidization. Food Biosci 2024, 61, 104983. [CrossRef]
- kaur, S.; Bhardwaj, R.D.; Kapoor, R.; Grewal, S.K. Biochemical Characterization of Oat (Avena Sativa L.) Genotypes with High Nutritional Potential. LWT 2019, 110, 32–39. [CrossRef]
- Ibrahim, M.S.; Nadeem, M.; Sultan, M.; Sajjad, U.; Hamid, K.; Qureshi, T.M.; Javaria, S. Techno-Functional Characteristics, Mineral Composition and Antioxidant Potential of Dietary Fiber Extracted by Sonication from Different Oat Cultivars (Avena Sativa). Future Foods 2024, 9, 100349. [CrossRef]
- Zhu, L.; Wang, L.F.; Bialobzyski, S.; Zijlstra, R.T.; Beltranena, E. Energy and Nutrient Digestibility and Effect of Increasing the Dietary Inclusion of Hull-Less Oats Replacing Wheat Grain on Growth Performance of Weanling Pigs. Anim Feed Sci Technol 2024, 318, 116139. [CrossRef]
- Ibrahim, M.S.; Nadeem, M.; Sultan, M.; Sajjad, U.; Hamid, K.; Qureshi, T.M.; Javaria, S. Techno-Functional Characteristics, Mineral Composition and Antioxidant Potential of Dietary Fiber Extracted by Sonication from Different Oat Cultivars (Avena Sativa). Future Foods 2024, 9, 100349. [CrossRef]
- Boukid, F. Corn (Zea Mays L.) Arabinoxylan to Expand the Portfolio of Dietary Fibers. Food Biosci 2023, 56, 103181. [CrossRef]
- Boukid, F. Comprehensive Review of Barley Dietary Fibers with Emphasis on Arabinoxylans. Bioactive Carbohydrates and Dietary Fibre 2024, 31, 100410. [CrossRef]
- Kozlova, L. V.; Nazipova, A.R.; Gorshkov, O. V.; Gilmullina, L.F.; Sautkina, O. V.; Petrova, N. V.; Trofimova, O.I.; Ponomarev, S.N.; Ponomareva, M.L.; Gorshkova, T.A. Identification of Genes Involved in the Formation of Soluble Dietary Fiber in Winter Rye Grain and Their Expression in Cultivars with Different Viscosities of Wholemeal Water Extract. Crop J 2022, 10, 532–549. [CrossRef]
- Tagliasco, M.; Font, G.; Renzetti, S.; Capuano, E.; Pellegrini, N. Role of Particle Size in Modulating Starch Digestibility and Textural Properties in a Rye Bread Model System. Food Research International 2024, 190, 114565. [CrossRef]
- Pferdmenges, L.E.; Lohmayer, R.; Frommherz, L.; Brühl, L.; Hüsken, A.; Mayer-Miebach, E.; Meinhardt, A.-K.; Ostermeyer, U.; Sciurba, E.; Zentgraf, H.; et al. Overview of the Nutrient Composition of Selected Milling Products from Rye, Spelt and Wheat in Germany. Journal of Food Composition and Analysis 2025, 140, 107275. [CrossRef]
- Rakha, A.; Saulnier, L.; Åman, P.; Andersson, R. Enzymatic Fingerprinting of Arabinoxylan and β-Glucan in Triticale, Barley and Tritordeum Grains. Carbohydr Polym 2012, 90, 1226–1234. [CrossRef]
- Li, W.; Xu, R.; Qin, S.; Song, Q.; Guo, B.; Li, M.; Zhang, Y.; Zhang, B. Cereal Dietary Fiber Regulates the Quality of Whole Grain Products: Interaction between Composition, Modification and Processing Adaptability. Int J Biol Macromol 2024, 274, 133223. [CrossRef]
- Tanwar, R.; Panghal, A.; Chaudhary, G.; Kumari, A.; Chhikara, N. Nutritional, Phytochemical and Functional Potential of Sorghum: A Review. Food Chemistry Advances 2023, 3, 100501. [CrossRef]
- Lee, S.; Choi, Y.-M.; Shin, M.-J.; Yoon, H.; Wang, X.; Lee, Y.; Yi, J.; Jeon, Y.; Desta, K.T. Exploring the Potentials of Sorghum Genotypes: A Comprehensive Study on Nutritional Qualities, Functional Metabolites, and Antioxidant Capacities. Front Nutr 2023, 10. [CrossRef]
- Espitia-Hernández, P., C.G.M., A.-V.J., D.-M.D., F.-N.A., S.T., R.-C.X. and S.-T.L., Sorghum (Sorghum Bicolor L. Moench): Chemical Composition and Its Health Benefits; Asociación Mexicana de Ciencia de los Alimentos, México, 2022; Vol. 7;
- Kanwar, P.; Yadav, R.B.; Yadav, B.S. Cross-Linking, Carboxymethylation and Hydroxypropylation Treatment to Sorghum Dietary Fiber: Effect on Physicochemical, Micro Structural and Thermal Properties. Int J Biol Macromol 2023, 233, 123638. [CrossRef]
- Guo, Y.; Liu, G.; Ning, Y.; Li, X.; Hu, S.; Zhao, J.; Qu, Y. Production of Cellulosic Ethanol and Value-Added Products from Corn Fiber. Bioresour Bioprocess 2022, 9, 81. [CrossRef]
- Colasanto, A.; Travaglia, F.; Bordiga, M.; Coïsson, J.D.; Arlorio, M.; Locatelli, M. Impact of Traditional and Innovative Cooking Techniques on Italian Black Rice (Oryza Sativa L., Artemide Cv) Composition. Food Research International 2024, 194, 114906. [CrossRef]
- Qadir, N.; Wani, I.A. Physicochemical and Functional Characterization of Dietary Fibres from Four Indian Temperate Rice Cultivars. Bioactive Carbohydrates and Dietary Fibre 2022, 28, 100336. [CrossRef]
- Zhang, S.; Ma, Q.; Deng, M.; Jia, X.; Huang, F.; Dong, L.; Zhang, R.; Sun, Z.; Zhang, M. Composition, Structural, Physicochemical and Functional Properties of Dietary Fiber from Different Milling Fractions of Black Rice Bran. LWT 2024, 195, 115743. [CrossRef]
- Ma, Q.; Wang, X.; Zhang, R.; Huang, F.; Jia, X.; Dong, L.; Liu, D.; Zhang, M. Structural, Physicochemical and Functional Properties of Dietary Fiber from Black Rice Bran Treated by Different Processing Methods. Food Biosci 2025, 65, 106025. [CrossRef]
- Liu, Y.; Zhang, H.; Yi, C.; Quan, K.; Lin, B. Chemical Composition, Structure, Physicochemical and Functional Properties of Rice Bran Dietary Fiber Modified by Cellulase Treatment. Food Chem 2021, 342, 128352. [CrossRef]
- Liu, X.; Fu, Y.; Ma, Q.; Yi, J.; Cai, S. Anti-Diabetic Effects of Different Phenolic-Rich Fractions from Rhus Chinensis Mill. Fruits in Vitro. eFood 2021, 2, 37–46. [CrossRef]
- Zheng, B.; Zhao, X.; Ao, T.; Chen, Y.; Xie, J.; Gao, X.; Liu, L.; Hu, X.; Yu, Q. The Role of Bound Polyphenols in the Anti-Obesity Effects of Defatted Rice Bran Insoluble Dietary Fiber: An Insight from Multi-Omics. Food Chem 2024, 459, 140345. [CrossRef]
- 1Thomas, R., 1*Rajeev B. and 2Kuang, Y.T. Composition of Amino Acids, Fatty Acids, Minerals and Dietary Fiber in Some of the Local and Import Rice Varieties of Malaysia. Int Food Res J 2015, 22, 1148–1155.
- Bader Ul Ain, H.; Saeed, F.; Ahmed, A.; Asif Khan, M.; Niaz, B.; Tufail, T. Improving the Physicochemical Properties of Partially Enhanced Soluble Dietary Fiber through Innovative Techniques: A Coherent Review. J Food Process Preserv 2019, 43, e13917. [CrossRef]
- Zou, X.; Xu, X.; Chao, Z.; Jiang, X.; Zheng, L.; Jiang, B. Properties of Plant-Derived Soluble Dietary Fibers for Fiber-Enriched Foods: A Comparative Evaluation. Int J Biol Macromol 2022, 223, 1196–1207. [CrossRef]
- Dhillon, B.; Choudhary, G.; Sodhi, N.S. A Study on Physicochemical, Antioxidant and Microbial Properties of Germinated Wheat Flour and Its Utilization in Breads. J Food Sci Technol 2020, 57, 2800–2808. [CrossRef]
- Zhu, R.; Xu, T.; He, B.; Wang, Y.; Zhang, L.; Huang, L. Modification of Artichoke Dietary Fiber by Superfine Grinding and High-Pressure Homogenization and Its Protection against Cadmium Poisoning in Rats. Foods 2022, 11, 1716. [CrossRef]
- Ahmad, A.; Anjum, F.M.; Zahoor, T.; Nawaz, H.; Din, A. Physicochemical and Functional Properties of Barley Β-glucan as Affected by Different Extraction Procedures. Int J Food Sci Technol 2009, 44, 181–187. [CrossRef]
- Liu, J.; Wang, Z.; Wang, Z.; Hao, Y.; Wang, Y.; Yang, Z.; Li, W.; Wang, J. Physicochemical and Functional Properties of Soluble Dietary Fiber from Different Colored Quinoa Varieties (Chenopodium Quinoa Willd). J Cereal Sci 2020, 95, 103045. [CrossRef]
- Zhang, Y.; Zhang, M.; Guo, X.; Bai, X.; Zhang, J.; Huo, R.; Zhang, Y. Improving the Adsorption Characteristics and Antioxidant Activity of Oat Bran by Superfine Grinding. Food Sci Nutr 2023, 11, 216–227. [CrossRef]
- Zhao, G.; Zhang, R.; Dong, L.; Huang, F.; Tang, X.; Wei, Z.; Zhang, M. Particle Size of Insoluble Dietary Fiber from Rice Bran Affects Its Phenolic Profile, Bioaccessibility and Functional Properties. LWT 2018, 87, 450–456. [CrossRef]
- Wei, C.; Ge, Y.; Liu, D.; Zhao, S.; Wei, M.; Jiliu, J.; Hu, X.; Quan, Z.; Wu, Y.; Su, Y.; et al. Effects of High-Temperature, High-Pressure, and Ultrasonic Treatment on the Physicochemical Properties and Structure of Soluble Dietary Fibers of Millet Bran. Front Nutr 2022, 8. [CrossRef]
- Huang, S.C.; Tsai, Y.F.; Chen, C.M. Effects of Wheat Fiber, Oat Fiber, and Inulin on Sensory and Physico-Chemical Properties of Chinese-Style Sausages. Asian-Australas J Anim Sci 2011, 24, 875–880. [CrossRef]
- Giuntini, E.B.; Sardá, F.A.H.; de Menezes, E.W. The Effects of Soluble Dietary Fibers on Glycemic Response: An Overview and Futures Perspectives. Foods 2022, 11, 3934. [CrossRef]
- Lu, K.; Yu, T.; Cao, X.; Xia, H.; Wang, S.; Sun, G.; Chen, L.; Liao, W. Effect of Viscous Soluble Dietary Fiber on Glucose and Lipid Metabolism in Patients with Type 2 Diabetes Mellitus: A Systematic Review and Meta-Analysis on Randomized Clinical Trials. Front Nutr 2023, 10. [CrossRef]
- Khorasani, A.C.; Kouhfar, F.; Shojaosadati, S.A. Pectin/Lignocellulose Nanofibers/Chitin Nanofibers Bionanocomposite as an Efficient Biosorbent of Cholesterol and Bile Salts. Carbohydr Polym 2021, 261, 117883. [CrossRef]
- Zhao, Y.; Kong, X.; Xing, X.; Hu, X.; Sun, Y. Comparison of Different Technologies for Dietary Fiber Extraction from Cold-Pressed Corn Germ Meal: Changes in Structural Characteristics, Physicochemical Properties and Adsorption Capacity. J Cereal Sci 2025, 121, 104077. [CrossRef]
- Wang, Z.; Chen, Y.; Wang, W.; Huang, C.; Hu, Y.; Johnston, L.; Wang, F. Dietary Supplementation With Fine-Grinding Wheat Bran Improves Lipid Metabolism and Inflammatory Response via Modulating the Gut Microbiota Structure in Pregnant Sow. Front Microbiol 2022, 13. [CrossRef]
- Barkas, F.; Adamidis, P.; Koutsogianni, A.-D.; Liamis, G.; Liberopoulos, E. Statin-Associated Side Effects in Patients Attending a Lipid Clinic: Evidence from a 6-Year Study. Archives of Medical Science – Atherosclerotic Diseases 2021, 6, 182–187. [CrossRef]
- Kelly, S.A.; Hartley, L.; Loveman, E.; Colquitt, J.L.; Jones, H.M.; Al-Khudairy, L.; Clar, C.; Germanò, R.; Lunn, H.R.; Frost, G.; et al. Whole Grain Cereals for the Primary or Secondary Prevention of Cardiovascular Disease. Cochrane Database of Systematic Reviews 2017, 2021. [CrossRef]
- Yanai, H.; Adachi, H.; Hakoshima, M.; Katsuyama, H. Postprandial Hyperlipidemia: Its Pathophysiology, Diagnosis, Atherogenesis, and Treatments. Int J Mol Sci 2023, 24, 13942. [CrossRef]
- Mortensen, M.B.; Dzaye, O.; Bøtker, H.E.; Jensen, J.M.; Maeng, M.; Bentzon, J.F.; Kanstrup, H.; Sørensen, H.T.; Leipsic, J.; Blankstein, R.; et al. Low-Density Lipoprotein Cholesterol Is Predominantly Associated With Atherosclerotic Cardiovascular Disease Events in Patients With Evidence of Coronary Atherosclerosis: The Western Denmark Heart Registry. Circulation 2023, 147, 1053–1063. [CrossRef]
- Naumann, S.; Schweiggert-Weisz, U.; Eglmeier, J.; Haller, D.; Eisner, P. In Vitro Interactions of Dietary Fibre Enriched Food Ingredients with Primary and Secondary Bile Acids. Nutrients 2019, 11, 1424. [CrossRef]
- Reynolds, A.N.; Akerman, A.; Kumar, S.; Diep Pham, H.T.; Coffey, S.; Mann, J. Dietary Fibre in Hypertension and Cardiovascular Disease Management: Systematic Review and Meta-Analyses. BMC Med 2022, 20, 139. [CrossRef]
- Llanaj, E.; Dejanovic, G.M.; Valido, E.; Bano, A.; Gamba, M.; Kastrati, L.; Minder, B.; Stojic, S.; Voortman, T.; Marques-Vidal, P.; et al. Effect of Oat Supplementation Interventions on Cardiovascular Disease Risk Markers: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Eur J Nutr 2022, 61, 1749–1778. [CrossRef]
- Ju, Y.; Zhang, C.; Zhang, Z.; Zhu, H.; Liu, Y.; Liu, T.; Ojo, O.; Qiu, J.; Wang, X. Effect of Dietary Fiber (Oat Bran) Supplement in Heart Rate Lowering in Patients with Hypertension: A Randomized DASH-Diet-Controlled Clinical Trial. Nutrients 2022, 14, 3148. [CrossRef]
- Xue, Y.; Cui, L.; Qi, J.; Ojo, O.; Du, X.; Liu, Y.; Wang, X. The Effect of Dietary Fiber (Oat Bran) Supplement on Blood Pressure in Patients with Essential Hypertension: A Randomized Controlled Trial. Nutrition, Metabolism and Cardiovascular Diseases 2021, 31, 2458–2470. [CrossRef]
- Kieffer, D.A.; Piccolo, B.D.; Marco, M.L.; Kim, E.B.; Goodson, M.L.; Keenan, M.J.; Dunn, T.N.; Knudsen, K.E.B.; Adams, S.H.; Martin, R.J. Obese Mice Fed a Diet Supplemented with Enzyme-Treated Wheat Bran Display Marked Shifts in the Liver Metabolome Concurrent with Altered Gut Bacteria. Journal of Nutrition 2016, 146, 2445–2460. [CrossRef]
- Paudel, D.; Dhungana, B.; Caffe, M.; Krishnan, P. A Review of Health-Beneficial Properties of Oats. Foods 2021, 10, 2591. [CrossRef]
- Wu, W.; Qiu, J.; Wang, A.; Li, Z. Impact of Whole Cereals and Processing on Type 2 Diabetes Mellitus: A Review. Crit Rev Food Sci Nutr 2020, 60, 1447–1474. [CrossRef]
- Torres, N.; Avila-Nava, A.; Medina-Vera, I.; Tovar, A.R. Dietary Fiber and Diabetes. In; 2020; pp. 201–218.
- Kabthymer, R.H.; Karim, M.N.; Hodge, A.M.; de Courten, B. High Cereal Fibre but Not Total Fibre Is Associated with a Lower Risk of Type 2 Diabetes: Evidence from the Melbourne Collaborative Cohort Study. Diabetes Obes Metab 2023, 25, 1911–1921. [CrossRef]
- Hu, Y.; Ding, M.; Sampson, L.; Willett, W.C.; Manson, J.E.; Wang, M.; Rosner, B.; Hu, F.B.; Sun, Q. Intake of Whole Grain Foods and Risk of Type 2 Diabetes: Results from Three Prospective Cohort Studies. BMJ 2020, m2206. [CrossRef]
- Ghanbari-Gohari, F.; Mousavi, S.M.; Esmaillzadeh, A. Consumption of Whole Grains and Risk of Type 2 Diabetes: A Comprehensive Systematic Review and Dose–Response Meta-analysis of Prospective Cohort Studies. Food Sci Nutr 2022, 10, 1950–1960. [CrossRef]
- Wehrli, F.; Taneri, P.E.; Bano, A.; Bally, L.; Blekkenhorst, L.C.; Bussler, W.; Metzger, B.; Minder, B.; Glisic, M.; Muka, T.; et al. Oat Intake and Risk of Type 2 Diabetes, Cardiovascular Disease and All-Cause Mortality: A Systematic Review and Meta-Analysis. Nutrients 2021, 13, 2560. [CrossRef]
- Zhang, J.; Wang, W.; Guo, D.; Bai, B.; Bo, T.; Fan, S. Antidiabetic Effect of Millet Bran Polysaccharides Partially Mediated via Changes in Gut Microbiome. Foods 2022, 11, 3406. [CrossRef]
- Li, X.-X.; Zhang, X.-X.; Zhang, R.; Ni, Z.-J.; Elam, E.; Thakur, K.; Cespedes-Acuña, C.L.; Zhang, J.-G.; Wei, Z.-J. Gut Modulation Based Anti-Diabetic Effects of Carboxymethylated Wheat Bran Dietary Fiber in High-Fat Diet/Streptozotocin-Induced Diabetic Mice and Their Potential Mechanisms. Food and Chemical Toxicology 2021, 152, 112235. [CrossRef]
- Shah, B. Obesity in Modern Society: Analysis, Statistics, and Treatment Approaches. International Journal of Science and Research (IJSR) 2023, 12, 358–360. [CrossRef]
- 2023; FAO Food Loss and Food Waste; 2023;
- Spring, B.; Champion, K.E.; Acabchuk, R.; Hennessy, E.A. Self-Regulatory Behaviour Change Techniques in Interventions to Promote Healthy Eating, Physical Activity, or Weight Loss: A Meta-Review. Health Psychol Rev 2021, 15, 508–539. [CrossRef]
- Palmeira, A.L.; Marques, M.M.; Sánchez-Oliva, D.; Encantado, J.; Santos, I.; Duarte, C.; Matos, M.; Carneiro-Barrera, A.; Larsen, S.C.; Horgan, G.; et al. Are Motivational and Self-Regulation Factors Associated with 12 Months’ Weight Regain Prevention in the NoHoW Study? An Analysis of European Adults. International Journal of Behavioral Nutrition and Physical Activity 2023, 20, 128. [CrossRef]
- Khan, J.; Gul, P.; Liu, K. Grains in a Modern Time: A Comprehensive Review of Compositions and Understanding Their Role in Type 2 Diabetes and Cancer. Foods 2024, 13, 2112. [CrossRef]
- Tammi, R.; Männistö, S.; Maukonen, M.; Kaartinen, N.E. Whole Grain Intake, Diet Quality and Risk Factors of Chronic Diseases: Results from a Population-Based Study in Finnish Adults. Eur J Nutr 2024, 63, 397–408. [CrossRef]
- Sanders, L.M.; Zhu, Y.; Wilcox, M.L.; Koecher, K.; Maki, K.C. Effects of Whole Grain Intake, Compared with Refined Grain, on Appetite and Energy Intake: A Systematic Review and Meta-Analysis. Advances in Nutrition 2021, 12, 1177–1195. [CrossRef]
- Maki, K.C.; Palacios, O.M.; Koecher, K.; Sawicki, C.M.; Livingston, K.A.; Bell, M.; Nelson Cortes, H.; McKeown, N.M. The Relationship between Whole Grain Intake and Body Weight: Results of Meta-Analyses of Observational Studies and Randomized Controlled Trials. Nutrients 2019, 11, 1245. [CrossRef]
- Wang, Y.; Feng, J.; Liu, T.; Gong, Z.; Zhuo, Q. Association between Whole-Grain Intake and Obesity Defined by Different Anthropometric Indicators and Dose–Response Relationship Analysis among U.S. Adults: A Population-Based Study. Nutrients 2024, 16, 2373. [CrossRef]
- Maki, K.C.; Palacios, O.M.; Koecher, K.; Sawicki, C.M.; Livingston, K.A.; Bell, M.; Nelson Cortes, H.; McKeown, N.M. The Relationship between Whole Grain Intake and Body Weight: Results of Meta-Analyses of Observational Studies and Randomized Controlled Trials. Nutrients 2019, 11, 1245. [CrossRef]
- Tammi, R.; Männistö, S.; Maukonen, M.; Kaartinen, N.E. Whole Grain Intake, Diet Quality and Risk Factors of Chronic Diseases: Results from a Population-Based Study in Finnish Adults. Eur J Nutr 2024, 63, 397–408. [CrossRef]
- Hosseinpour-Niazi, S.; Bakhshi, B.; Mirmiran, P.; Gaeini, Z.; Hadaegh, F.; Azizi, F. Effect of Weight Change on the Association between Overall and Source of Carbohydrate Intake and Risk of Metabolic Syndrome: Tehran Lipid and Glucose Study. Nutr Metab (Lond) 2023, 20, 39. [CrossRef]
- Malin, S.K.; Kullman, E.L.; Scelsi, A.R.; Haus, J.M.; Filion, J.; Pagadala, M.R.; Godin, J.-P.; Kochhar, S.; Ross, A.B.; Kirwan, J.P. A Whole-Grain Diet Reduces Peripheral Insulin Resistance and Improves Glucose Kinetics in Obese Adults: A Randomized-Controlled Trial. Metabolism 2018, 82, 111–117. [CrossRef]
- Thandapilly, S.J.; Ndou, S.P.; Wang, Y.; Nyachoti, C.M.; Ames, N.P. Barley β-Glucan Increases Fecal Bile Acid Excretion and Short Chain Fatty Acid Levels in Mildly Hypercholesterolemic Individuals. Food Funct 2018, 9, 3092–3096. [CrossRef]
- Pi, X.; Yu, Z.; Yang, X.; Du, Z.; Liu, W. Effects of Zymosan on Short-Chain Fatty Acid and Gas Production in in Vitro Fermentation Models of the Human Intestinal Microbiota. Front Nutr 2022, 9. [CrossRef]
- Jiang, C.; Zeng, X.; Wei, X.; Liu, X.; Wang, J.; Zheng, X. Improvement of the Functional Properties of Insoluble Dietary Fiber from Corn Bran by Ultrasonic-Microwave Synergistic Modification. Ultrason Sonochem 2024, 104, 106817. [CrossRef]
- Tamura, K.; Hemsworth, G.R.; Déjean, G.; Rogers, T.E.; Pudlo, N.A.; Urs, K.; Jain, N.; Davies, G.J.; Martens, E.C.; Brumer, H. Molecular Mechanism by Which Prominent Human Gut Bacteroidetes Utilize Mixed-Linkage Beta-Glucans, Major Health-Promoting Cereal Polysaccharides. Cell Rep 2017, 21, 417–430. [CrossRef]
- Sebastià, C.; Folch, J.M.; Ballester, M.; Estellé, J.; Passols, M.; Muñoz, M.; García-Casco, J.M.; Fernández, A.I.; Castelló, A.; Sánchez, A.; et al. Interrelation between Gut Microbiota, SCFA, and Fatty Acid Composition in Pigs. mSystems 2024, 9. [CrossRef]
- Valido, E.; Stoyanov, J.; Bertolo, A.; Hertig-Godeschalk, A.; Zeh, R.M.; Flueck, J.L.; Minder, B.; Stojic, S.; Metzger, B.; Bussler, W.; et al. Systematic Review of the Effects of Oat Intake on Gastrointestinal Health. J Nutr 2021, 151, 3075–3090. [CrossRef]
- Procházková, N.; Venlet, N.; Hansen, M.L.; Lieberoth, C.B.; Dragsted, L.O.; Bahl, M.I.; Licht, T.R.; Kleerebezem, M.; Lauritzen, L.; Roager, H.M. Effects of a Wholegrain-Rich Diet on Markers of Colonic Fermentation and Bowel Function and Their Associations with the Gut Microbiome: A Randomised Controlled Cross-over Trial. Front Nutr 2023, 10. [CrossRef]
- Thandapilly, S.J.; Ndou, S.P.; Wang, Y.; Nyachoti, C.M.; Ames, N.P. Barley β-Glucan Increases Fecal Bile Acid Excretion and Short Chain Fatty Acid Levels in Mildly Hypercholesterolemic Individuals. Food Funct 2018, 9, 3092–3096. [CrossRef]
- Liu, Y.; Xue, K.; Iversen, K.N.; Qu, Z.; Dong, C.; Jin, T.; Hallmans, G.; Åman, P.; Johansson, A.; He, G.; et al. The Effects of Fermented Rye Products on Gut Microbiota and Their Association with Metabolic Factors in Chinese Adults – an Explorative Study. Food Funct 2021, 12, 9141–9150. [CrossRef]
- Faubel, N.; Blanco-Morales, V.; Barberá, R.; Garcia-Llatas, G. Impact of Colonic Fermentation of Plant Sterol-Enriched Rye Bread on Gut Microbiota and Metabolites. In Proceedings of the Foods 2023; MDPI: Basel Switzerland, October 13 2023; p. 87.
- Jefferson, A.; Adolphus, K. The Effects of Intact Cereal Grain Fibers, Including Wheat Bran on the Gut Microbiota Composition of Healthy Adults: A Systematic Review. Front Nutr 2019, 6.
- Vitaglione, P.; Mennella, I.; Ferracane, R.; Rivellese, A.A.; Giacco, R.; Ercolini, D.; Gibbons, S.M.; La Storia, A.; Gilbert, J.A.; Jonnalagadda, S.; et al. Whole-Grain Wheat Consumption Reduces Inflammation in a Randomized Controlled Trial on Overweight and Obese Subjects with Unhealthy Dietary and Lifestyle Behaviors: Role of Polyphenols Bound to Cereal Dietary Fiber. Am J Clin Nutr 2015, 101, 251–261. [CrossRef]
- Zeng, Z.; Zhu, Y.; Li, Y.; Chang, X.; Lin, J.; Chen, L.; Lyu, Q.; Chen, X.; Ding, W. Examination of the Bioavailability and Bioconversion of Wheat Bran-Bound Ferulic Acid: Insights into Gastrointestinal Processing and Colonic Metabolites. J Agric Food Chem 2025, 73, 1331–1344. [CrossRef]
- Hu, J.; Wang, J.; Li, Y.; Xue, K.; Kan, J. Use of Dietary Fibers in Reducing the Risk of Several Cancer Types: An Umbrella Review. Nutrients 2023, 15, 2545. [CrossRef]
- Zhang, X.-F.; Wang, X.-K.; Tang, Y.-J.; Guan, X.-X.; Guo, Y.; Fan, J.-M.; Cui, L.-L. Association of Whole Grains Intake and the Risk of Digestive Tract Cancer: A Systematic Review and Meta-Analysis. Nutr J 2020, 19, 52. [CrossRef]
- Collatuzzo, G.; Cortez Lainez, J.; Pelucchi, C.; Negri, E.; Bonzi, R.; Palli, D.; Ferraroni, M.; Zhang, Z.-F.; Yu, G.-P.; Lunet, N.; et al. The Association between Dietary Fiber Intake and Gastric Cancer: A Pooled Analysis of 11 Case–Control Studies. Eur J Nutr 2024, 63, 1857–1865. [CrossRef]
- Hullings, A.G.; Sinha, R.; Liao, L.M.; Freedman, N.D.; Graubard, B.I.; Loftfield, E. Whole Grain and Dietary Fiber Intake and Risk of Colorectal Cancer in the NIH-AARP Diet and Health Study Cohort. Am J Clin Nutr 2020, 112, 603–612. [CrossRef]
- Kyrø, C.; Tjønneland, A.; Overvad, K.; Olsen, A.; Landberg, R. Higher Whole-Grain Intake Is Associated with Lower Risk of Type 2 Diabetes among Middle-Aged Men and Women: The Danish Diet, Cancer, and Health Cohort. J Nutr 2018, 148, 1434–1444. [CrossRef]
- Makarem, N.; Nicholson, J.M.; Bandera, E. V.; McKeown, N.M.; Parekh, N. Consumption of Whole Grains and Cereal Fiber in Relation to Cancer Risk: A Systematic Review of Longitudinal Studies. Nutr Rev 2016, 74, 353–373. [CrossRef]
- Procházková, N.; Venlet, N.; Hansen, M.L.; Lieberoth, C.B.; Dragsted, L.O.; Bahl, M.I.; Licht, T.R.; Kleerebezem, M.; Lauritzen, L.; Roager, H.M. Effects of a Wholegrain-Rich Diet on Markers of Colonic Fermentation and Bowel Function and Their Associations with the Gut Microbiome: A Randomised Controlled Cross-over Trial. Front Nutr 2023, 10. [CrossRef]
- Liu, X.; Yang, W.; Petrick, J.L.; Liao, L.M.; Wang, W.; He, N.; Campbell, P.T.; Zhang, Z.-F.; Giovannucci, E.; McGlynn, K.A.; et al. Higher Intake of Whole Grains and Dietary Fiber Are Associated with Lower Risk of Liver Cancer and Chronic Liver Disease Mortality. Nat Commun 2021, 12, 6388. [CrossRef]
- Kubatka, P.; Kello, M.; Kajo, K.; Kruzliak, P.; Výbohová, D.; Šmejkal, K.; Maršík, P.; Zulli, A.; Gönciová, G.; Mojžiš, J.; et al. Young Barley Indicates Antitumor Effects in Experimental Breast Cancer In Vivo and In Vitro. Nutr Cancer 2016, 68, 611–621. [CrossRef]
- Choromanska, A.; Kulbacka, J.; Harasym, J.; Oledzki, R.; Szewczyk, A.; Saczko, J. High- and Low-Molecular Weight Oat Beta-Glucan Reveals Antitumor Activity in Human Epithelial Lung Cancer. Pathology & Oncology Research 2018, 24, 583–592. [CrossRef]
- Harasym, J.; Dziendzikowska, K.; Kopiasz, Ł.; Wilczak, J.; Sapierzyński, R.; Gromadzka-Ostrowska, J. Consumption of Feed Supplemented with Oat Beta-Glucan as a Chemopreventive Agent against Colon Cancerogenesis in Rats. Nutrients 2024, 16, 1125. [CrossRef]
- Celiberto, F.; Aloisio, A.; Girardi, B.; Pricci, M.; Iannone, A.; Russo, F.; Riezzo, G.; D’Attoma, B.; Ierardi, E.; Losurdo, G.; et al. Fibres and Colorectal Cancer: Clinical and Molecular Evidence. Int J Mol Sci 2023, 24, 13501. [CrossRef]
- Eriksen, A.K.; Brunius, C.; Mazidi, M.; Hellström, P.M.; Risérus, U.; Iversen, K.N.; Fristedt, R.; Sun, L.; Huang, Y.; Nørskov, N.P.; et al. Effects of Whole-Grain Wheat, Rye, and Lignan Supplementation on Cardiometabolic Risk Factors in Men with Metabolic Syndrome: A Randomized Crossover Trial. Am J Clin Nutr 2020, 111, 864–876. [CrossRef]
- Sandberg, J.C.; Björck, I.M.E.; Nilsson, A.C. Rye-Based Evening Meals Favorably Affected Glucose Regulation and Appetite Variables at the Following Breakfast; A Randomized Controlled Study in Healthy Subjects. PLoS One 2016, 11, e0151985. [CrossRef]
- Iversen, K.N.; Jonsson, K.; Landberg, R. The Effect of Rye-Based Foods on Postprandial Plasma Insulin Concentration: The Rye Factor. Front Nutr 2022, 9. [CrossRef]
- Iversen, K.N.; Carlsson, F.; Andersson, A.; Michaëlsson, K.; Langton, M.; Risérus, U.; Hellström, P.M.; Landberg, R. A Hypocaloric Diet Rich in High Fiber Rye Foods Causes Greater Reduction in Body Weight and Body Fat than a Diet Rich in Refined Wheat: A Parallel Randomized Controlled Trial in Adults with Overweight and Obesity (the RyeWeight Study). Clin Nutr ESPEN 2021, 45, 155–169. [CrossRef]
- Iversen, K.N.; Dicksved, J.; Zoki, C.; Fristedt, R.; Pelve, E.A.; Langton, M.; Landberg, R. The Effects of High Fiber Rye, Compared to Refined Wheat, on Gut Microbiota Composition, Plasma Short Chain Fatty Acids, and Implications for Weight Loss and Metabolic Risk Factors (the RyeWeight Study). Nutrients 2022, 14, 1669. [CrossRef]
- Dioum, E.H.M.; Schneider, K.L.; Vigerust, D.J.; Cox, B.D.; Chu, Y.; Zachwieja, J.J.; Furman, D. Oats Lower Age-Related Systemic Chronic Inflammation (IAge) in Adults at Risk for Cardiovascular Disease. Nutrients 2022, 14, 4471. [CrossRef]
- Pino, J.L.; Mujica, V.; Arredondo, M. Effect of Dietary Supplementation with Oat β-Glucan for 3 months in Subjects with Type 2 Diabetes: A Randomized, Double-Blind, Controlled Clinical Trial. J Funct Foods 2021, 77, 104311. [CrossRef]
- Fiedler, M.; Müller, N.; Kloos, C.; Kramer, G.; Kellner, C.; Schmidt, S.; Wolf, G.; Kuniß, N. A Randomized Controlled Trial to Assess the Feasibility and Practicability of an Oatmeal Intervention in Individuals with Type 2 Diabetes: A Pilot Study in the Outpatient Sector. J Clin Med 2024, 13, 5126. [CrossRef]
- Huang, K.; Hong, C.; Huang, Y.; Liu, Y.; Yu, Z.; Li, S.; Guan, X.; Zhao, W. Oat β-Glucan Prevents High Fat Diet Induced Obesity by Targeting Ileal Farnesoid X Receptor-Fibroblast Growth Factor 15 Signaling. Int J Biol Macromol 2025, 306, 141543. [CrossRef]
- Hughes, R.L.; Horn, W.F.; Wen, A.; Rust, B.; Woodhouse, L.R.; Newman, J.W.; Keim, N.L. Resistant Starch Wheat Increases PYY and Decreases GIP but Has No Effect on Self-Reported Perceptions of Satiety. Appetite 2022, 168, 105802. [CrossRef]
- Müller, M.; Hermes, G.D.A.; Emanuel E., C.; Holst, J.J.; Zoetendal, E.G.; Smidt, H.; Troost, F.; Schaap, F.G.; Damink, S.O.; Jocken, J.W.E.; et al. Effect of Wheat Bran Derived Prebiotic Supplementation on Gastrointestinal Transit, Gut Microbiota, and Metabolic Health: A Randomized Controlled Trial in Healthy Adults with a Slow Gut Transit. Gut Microbes 2020, 12, 1704141. [CrossRef]
- Lim, S.M.; Choo, J.M.; Li, H.; O’Rielly, R.; Carragher, J.; Rogers, G.B.; Searle, I.; Robertson, S.A.; Page, A.J.; Muhlhausler, B. A High Amylose Wheat Diet Improves Gastrointestinal Health Parameters and Gut Microbiota in Male and Female Mice. Foods 2021, 10, 220. [CrossRef]
- Liedike, B.; Khatib, M.; Tabarsi, B.; Harris, M.; Wilson, S.L.; Ortega-Santos, C.P.; Mohr, A.E.; Vega-López, S.; Whisner, C.M. Evaluating the Effects of Corn Flour Product Consumption on Cardiometabolic Outcomes and the Gut Microbiota in Adults with Elevated Cholesterol: A Randomized Crossover. J Nutr 2024, 154, 2437–2447. [CrossRef]
- Kondo, K.; Morino, K.; Nishio, Y.; Ishikado, A.; Arima, H.; Nakao, K.; Nakagawa, F.; Nikami, F.; Sekine, O.; Nemoto, K.; et al. Fiber-Rich Diet with Brown Rice Improves Endothelial Function in Type 2 Diabetes Mellitus: A Randomized Controlled Trial. PLoS One 2017, 12, e0179869. [CrossRef]
- Liu, J.; An, Y.; Yang, N.; Xu, Y.; Wang, G. Longitudinal Associations of Dietary Fiber and Its Source with 48-Week Weight Loss Maintenance, Cardiometabolic Risk Factors and Glycemic Status under Metformin or Acarbose Treatment: A Secondary Analysis of the March Randomized Trial. Nutr Diabetes 2024, 14, 81. [CrossRef]
- Reynolds, A.N.; Akerman, A.P.; Mann, J. Dietary Fibre and Whole Grains in Diabetes Management: Systematic Review and Meta-Analyses. PLoS Med 2020, 17, e1003053. [CrossRef]
Figure 2.
Examples of different dietary fiber and their main sources.
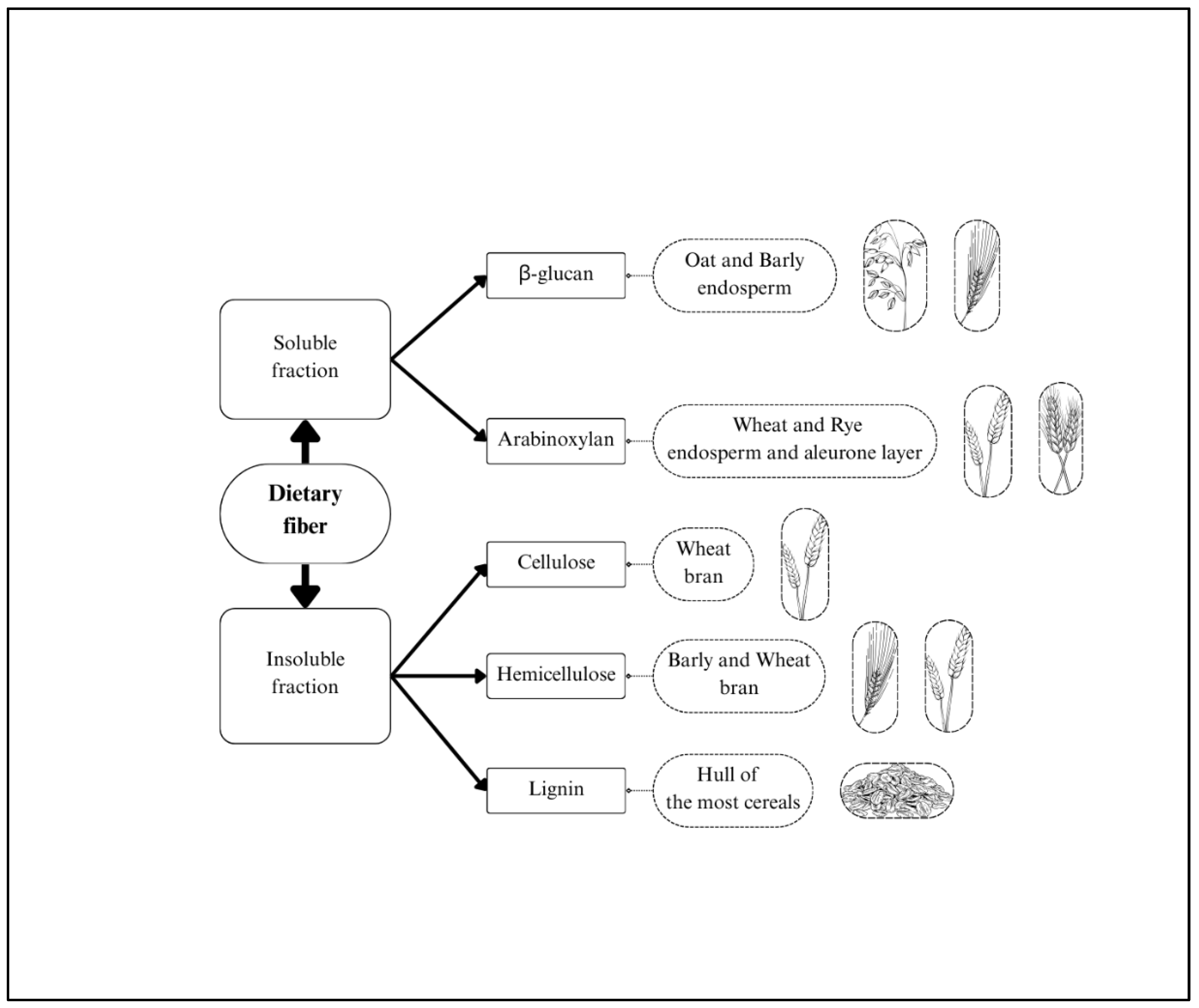
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



