
Submitted:
30 May 2025
Posted:
02 June 2025
You are already at the latest version
Abstract
Myxoma virus (MYXV), a rabbit specific poxvirus and non-pathogenic to humans and mice, is an excellent candidate oncolytic virus for cancer therapy. MYXV also has immunotherapeutic benefits. In ovarian cancer (OC), immunosuppressive tumor associated macrophages (TAMs) are key to inhibit antitumor immunity while hindering therapeutic benefit by chemotherapy and dendritic cell (DC) vaccine. Because MYXV favors binding/entry of macropahges/monocytes, we examined the therapeutic potential of MYXV against TAMs. We found previously that a replication-defective MYXV with targeted deletion of an essential gene, M062R, designated M062R MYXV, activated both host DNA sensing pathway and the SAMD9 pathway. Treatment with M062R leads to therapeutic benefit as monotherapy comparable to wildtype replicating MYXV in preclinical models. Here we found that M062R MYXV, when integrated with cisplatin and DC immunotherapy, further improved treatment benefit likely through promoting tumor antigen-specific T cell function. Moreover, we also tested M062R MYXV in targeting human immunosuppressive TAMs from OC patient ascites in a co-culture system. We found that M062R treatment subverted the immunosuppressive properties of TAMs and elevated the avidity of cytokine production in tumor antigen-specific CD4+ T cells. Overall, M062R presents a promising immunotherapeutic platform and is as a beneficial adjuvant to chemotherapy and DC vaccine.
Keywords:
oncolytic virus
; myxoma virus
; immunoregulatory
; immunotherapy platform
; host range factor
; type 1 interferon
; type 1 interferon stimulated genes
; proinflammatory
1. Introduction
Cancer cells can cultivate and maintain the tumor environment to escape from host immune detection and elimination [1]. Once the tumor environment is established, cancer cells often evade targeted treatment regimes such as chemotherapy and even develop resistance and tolerance to repeated chemotherapy treatments, which eventually leads to irreversible progression in cancer cell growth and lethality in patients [2]. This is often the case for ovarian cancer (OC) patients [3]. Ovarian cancer is a leading cause of cancer death in women with over 200,000 death annually worldwide [4]. While platinum-based chemotherapy remains the standard care for OC patients and most patients initially responded, chemo-resistance arises frequently in patients [5,6] and tumor progression is especially shielded by an immunosuppressive tumor environment [3]. High grade serous OC (HGSOC) is the most common form of OC in women (over 70%) [3], and parameters indicating an immunosuppressive tumor environment suggest a poor prognosis in patients [7,8]. Targeting key elements of the OC immunosuppressive tumor environment, such as tumor associated macrophages (TAMs) [9] and Tregs [10], leads to development of novel therapeutics, for instance through the use of dendritic cell (DC) vaccines [11,12]. In this study, we considered the promise of DC vaccines in treating immunosuppressive HGSOC and aim to develop combinatorial treatment strategy to further advance the therapeutic benefit.
Myxoma virus (MYXV) belongs to the poxvirus family and has a unique safety profile for therapeutic applications in humans and in preclinical mouse models [13]. These viruses also have a large genome capacity to allow extensive genetic engineering for therapeutic molecules, such as IL-12 [14], IL-15 [15], and TNF [16], to edit regional and systemic immune networks. Because WT MYXV has oncolytic properties by specifically infecting and killing cancer cells [17] and engineered mutant MYXVs can also activate host innate immune signaling [16,18,19], this virotherapy vector represents a novel approach to eliminate tumor cells while reprogramming the tumor environment for customized therapy. Besides utility as monotherapy, MYXV has its versatility as a complement for other therapeutics, e.g., chemotherapy (cisplatin [20] and gemcitabine [21]). In this study, we further explore the potential application of mutant MYXV for immunotherapy and as an adjuvant to chemotherapy and DC vaccination.
Although replicating oncolytic viruses can amplify killing of tumor cells, antiviral innate immunity often rapidly eliminates their presence in the tumor environment quickly [22]. However, despite their removal, their immunological impact of oncolytic viruses may continue to provide therapeutic benefit [23]. The debate on whether replication-competency determines the longevity and effectieveness of therapeutic immune responses continues. Our study provides another argument on the therapeutic benefit of replication-defective virus.
In this study, we tested MYXV virotherapy combined with cisplatin chemotherapy and a Th17—inducing DC vaccine that has recently shown promise for treatment of advanced stage ovarian cancer patients [24]. We show that combinatorial treatments with MYXV improve survival in a mouse model of OC when compared with cisplatin or DC monotherapy. We further show that MYXV can alleviate immune suppression by patient-derived ovarian tumor-associated CD14+ macrophages (TAMs) in coculture with tumor antigen-specific CD4+ T cells.
2. Materials and Methods
Human Subjects
Deidentified ovarian cancer patient samples were provided by the Women’s Oncology clinic, Winthrop P. Rockefeller Cancer Institute, University of Arkansas for Medical Sciences (UAMS) and the Mayo Clinic Ovarian Cancer Research Program Biospecimens Bank under IRB-approved protocols. Ovarian tumor ascites samples were recovered at the time of surgery.
Cell Culture and Virus Stock
Patient ovarian cancer cells (KAJ343, OvCa-37, and OvCa-43)[20] were cultured in RPMI complete growth medium supplemented with glutamine, penicillin/streptomycin and 10% fetal bovine serum. KAJ343 and OvCa-43 were high-grade serous ovarian carcinomas. OvCa-37 underwent neoadjuvant chemotherapy, and diagnosis was based on paracentesis ascites rather than surgical pathology, which was not defined. BSC40 (ATCC CRL-2761), ID8 TP53-/- (provided by Iain McNeish, Imperial College, London) [25], and ID8-SP17 cells (provided by Keith Knutson, Mayo Clinic, Jacksonville)[26] are cultured in DMEM complete medium. The complete growth medium (e.g., DMEM Lonza/BioWhittaker Catalog no 12-604Q) was supplemented with 10% FBS (Atlanta Biologicals, Minneapolis, MN), 2 mM glutamine (Corning Cellgro, Millipore Sigma, St. Louis, MO), and 100 μg per ml of Pen/Strep (Corning Cellgro, Millipore Sigma, St. Louis, MO); for RPMI1640 complete culture medium, in addition to FBS, glutamine, and Pen/Strep, 2-mercaptoethanol (MP biomedicals, Solon, OH) was supplemented to a final concentration of 0.05 mM.
The viruses used were originally derived from Lausanne strain of myxoma virus (MYXV) (GenBank Accession NC_001132.2/AF170726.2). The MYXV M062R deletion mutant (ΔM062R) and WT MYXV have been described previously [27]. Myxoma virus stocks were prepared on BSC-40 cells and purified with sucrose step gradient through ultracentrifugation as previously described [28].
RT Realtime (RT2) PCR
Human primary patient cancer cells KAJ434, OvCa-37 and OvCa-43 or murine ID8 derivatives ID8-TP53-/- are seeded the day before. The next day cells are mock treated, or infected at an moi of 5 for either WT or ΔM062R MYXV [27]. At 24 hrs post-infection, cells were harvested with Direct-zol RNA Mini Prep kit (catalog # R2052, Zymo, Irvine, CA, USA) according to manufacturer standard protocol. RNA quality is examined by running on the RNA gel to check 28S and 18S integrity and photospectrometer to estimate concentration. Equal amounts of total RNA in a maximal volume of 6 μL are used for cDNA synthesis using NEB PhotoScript® First Strand cDNA Synthesis Kit (Catalog # E6300L, NEB Inc, Ipswitch, MA) as instructed in the manufacturer standard protocol. using NEB ProtoScript First Strand cDNA Synthesis Kit (Catalog # E6300L, NEB Inc, Ipswitch, MA) as instructed in the manufacturer standard protocol. Realtime PCR is conducted following manufacturer standard protocol (Luna Universal qPCR Master Mix, NEB Inc). Sybr green RT-PCR primers used in this study is listed in Table 1.
Tumor Model Establishment and Treatment
The animal study was approved by the UAMS Institutional Animal Care and Usage Committee (IACUC) at the University of Arkansas for Medical Sciences (UAMS). ID8-TP53-/- and ID8-SP17 cells are used for mouse model studies. The tumor model establishment is similar to what we previously described for the intraperitoneal carcinomatosis model. Briefly, 4 studies were performed. (a) The “WT MYXV+ cisplatin+ DC vaccine” study. In this study, ID8-TP53-/- tumor cells were injected at 3x106 cells per mouse through intraperitoneal injection (IP), followed by WT MYXV treatment started on day 7 post-tumor establishment at 108 pfu per injection with a total of 4 injections at 2 days apart. In the same study, for combination treatment, the cisplatin is at 3mg/Kg of dose with a regimen of every 3 days starting on day 16 post-tumor establishment for a total of 3 treatments. The DC vaccine started at 30 days post-tumor establishment for a total of 4 treatments at a frequency of every 7 days. (b) The “late Cisplatin+ ΔM062R +DC vaccine” study with the ID8-TP53-/- model. At 16 days post-tumor establishment at the same dose as in (a), cisplatin was given via IP route at 3 mg/Kg at the same frequency as in (a). On 28 days post-tumor establishment, ΔM062R treatment was provided at 108 pfu per mouse via IP route for every 2 days for a total of 4 treatments. On 44 days post-tumor establishment, DC vaccine was given at the same frequency as in (a). The 3rd study (c) is called “early Cisplatin+ΔM062R +DC vaccine” study. The same doses and frequencies of cisplatin, ΔM062R, and DC vaccine as in (b) were used, but the treatment started at 7 days post-tumor establishment. For the ID8-SP17 HGSOC model (d), we tested targeted immunotherapy against tumor antigen Sp17. The tumor cells were injected at 106 cells per mouse via IP route, and cisplatin was at 1 mg/Kg of dose with a regimen of one injection every 3 days; the ΔM062R treatment was first performed 6 days after the last cisplatin treatment and followed by DC vaccine 2 days afterwards, and the ΔM062R -DC vaccine mini-regimen was repeated for 4 times every 7 days.
Human Ovarian Cancer Ascites CD14+ Macrophages and CD4+ T Cell Co-Culture System
CD14+ TAMs were recovered from primary ovarian tumor ascites by CD14+ microbead and magnetic column purification (Miltenyi). Human CD4+ T cells specific for ovarian tumor peptide antigens were derived through stimulation with TADG14v peptide-loaded Th17-inducing dendritic cells, as previously described [33,34,35]. CD14+ ascites TAMs were either uninfected or infected with WT or ΔM062R MYXV (at a moi of 10, pre-incubated on ice for 1 hr, then washed one time with PBS). Ascites CD14+ TAMs (5 x105/well) were then co-cultured with ovarian tumor antigen--specific CD4+ T cells (5 x 105/well) in 0.7 ml RPMI plus antibiotics, glutamine 5 x 10-5 M 2-mercatopethanol and 10% human AB serum (RPMI 10Hu). After 48 hr, tumor antigen peptide-loaded and irradiated (75 gray) autologous lymphoblastoid cells (2.5 x 105 cells/well) were added in 0.2 ml RPMI/10Hu, and the cocultures incubated o/n in the presence of Brefeldin A. Where indicated, cocultures were treated with a STING inhibitor (1 μM H-151) or a TBK-1 inhibitor (2 μM BX-795). Following coculture, CD4+ T cells were stained with PE-anti-CD4 antibody, fixed and permeabilized with fixation/permeabilization buffer (Affymetrix eBioscience) and stained with APC anti-TNF antibody. Samples were analyzed with a FACSCalibur flow cytometer and CellQuest software.
Statistical Analyses
GraphaPrism 10.1 was used for statistical analyses. Survival study was performed using Kaplan-Meier analyses followed by Long-rank comparison. Statistical significance is defined by *p<0.05.
3. Results
3.1. Myxoma Virus Infection in Primary Cells of Human Ovarian Cancer Environment Stimulated Pro-Inflammatory Gene Expression and Up-Regulation of Sp17
Antigen Sp17 is a signature protein found in OC cells [36,37]. Expression of Sp17, however, is not found in normal female cells; thus Sp17 antigen may be an excellent target for OC immunotherapy [37,38,39]. We included Sp17 in our RT-PCR array as an indicator for cancer cells. Two MYXV viruses, a replicating WT and replication defective mutant virus (ΔM062R), were used for the study. Primary OC patient ascites cells from 3 patients were mock treated or treated with either MYXV at an moi of 5-10 before examining the gene expression profile. MYXV treatment increased Sp17 expression (Figure 1A-C, red frame) in addition to up-regulation of IFNβ and other proinflammatory molecules (e.g., IL12b and CXCL10) (Figure 1). We performed similar tests using the murine OC cell line ID8-TP53-/- that was used for establishing a syngeneic OC model system as previously described [25]. We found that MYXV treatment increased expression of pro-inflammatory molecules (Figure 2A) similar to what we saw in human cells (Figure 1). Moreover, Sp17 expression is also up-regulated (Figure 2A and 2B, red frame) similarly as found in human cells from the cancer environment (Figure 1). Using an array panel to examine gene expression that shapes a pro-cancer tumor environment, we found that ΔM062R treatment moderately altered gene expression to a lesser degree than wildtype (WT) MYXV (Figure 2B). This is consistent with our previous observation that ΔM062R treatment stimulates pro-inflammatory gene expression at both RNA and protein levels compared with WT MYXV infected cells and mock treated primary cells [18].
3.2. Wildtype MYXV Treatment Proceeding Cisplatin Treatment Led to Moderate Therapeutic Benefit in a High Grade Serious Ovarian Cancer (HGSOC) Murine Syngeneic Model
The recent success of a DC vaccine trial in OC patients provided exciting and novel options targeting the unique ovarian tumor environment [24]. With both human and murine Sp17 expressed in OC cells, we proposed a proof-of-concept DC vaccination strategy of pulsing Sp17 peptide on DC cells for the in vivo study as previously described [26]. In our previous study, we found that WT MYXV treatment before cisplatin treatment provided better therapeutic benefit than either WT MYXV monotherapy or WT MYXV provided after cisplatin treatment [20]. The ability of replicating oncolytic WT MYXV to eliminate OC cells facilitated cisplatin control of tumor progression, hence the rationale of our combination treatment strategy. Thus, with a replicating oncolytic agent promoting the expression of tumor antigen, we tested whether DC vaccine may be a good option to prolong survival further. We indeed found that WT MYXV and DC provided a significant improvement of survival with and without cisplatin treatment (Figure 3A) with a median survival of 87 days for either regime. Mock treatment, cisplatin alone, WT MYXV alone, and WT MYXV plus cisplatin show a median survival of 50, 65, 72, and 75 days, respectively.
3.3. M062R-Null (ΔM062R) MYXV Treatment as Monotherapy or in Combination with DC Vaccine Targeting Sp17 Antigen Showed Significant Treatment Benefit in the HGSOC Murine Syngeneic Models.
Due to the association of Sp17 antigen in OC with tumorigenicity, tumor progression, chemoresistance, and the immunosuppressiveness, Sp17 becomes a promising therapeutic target for vaccine development in treating OC [37,38,40,41]. In our study, the DC vaccine is indeed designed to target the antigenic epitopes of Sp17 [42].
In our previous study [20], we examined the therapeutic potential of a replication defective MYXV called M062R-null MYXV (ΔM062R) as either monotherapy or in combination with chemotherapy. We found previously that ΔM062R treatment after cisplatin provided better survival [20]. Recently, we further examined the immunostimulatoary effect of ΔM062R and found this virus superior in inducing proinflammatory responses in monocytes/macrophages by triggering DNA-sensing stimulated type 1 interferon responses (IFN-I) [18]. The effect induced by ΔM062R in myeloid cells is different from the conventional DNA-sensing stimulated IFN-I responses and promotes a potent pro-inflammatory outcome [18,43]. indicating an additional mechanism governing the unique immunological state [43]. Thus, besides its being a potential adjuvant to cisplatin, we proposed to examine if ΔM062R has therapeutic potential for immunotherapy and may also be an adjuvant for DC vaccine. In the clinical trial of DC vaccine, all the patients were treated after platinum-based therapy and tumor debunking, thus we did not specifically examine DC vaccine as a monotherapy in our study. The ID8-TP53-/- syngeneic model system portrays the conditions of HGSOC in humans [44], and it presents an aggressive form of OC (Figure 3). We found that combined treatment with cisplatin, ΔM062R, and DC vaccine led to significant improvement in survival (p<0.0001, Long-rank Mantel-Cox test, with a median surivial of 106 days, Figure 3B) compared to mock treatment, cisplatin alone, and ΔM062R alone, with median survival of 50, 65, and 67 days, respectively. Moreover, while combination treatment of cisplatin-ΔM062R or ΔM062R-DC vaccine significantly improved survival compared to monotherapy with median survival of 85 and 72 days, respectively, cisplatin-ΔM062R-DC vaccine combinatorial regime is significantly better than the other two-regime treatments (p<0.0197 and p<0.0127, respectively, by Long-rank Mantel-Cox test). In this study, however, we did not evaluate the therapeutic effect of DC vaccine in facilitating cisplatin chemotherapy or ΔM062R virotherapy. We also did not examine whether ΔM062R therapeutic benefit is synergistic/additive based on the same mechanism of DC vaccine. We recognized that in this study the treatment of cisplatin or virotherapy was provided at a much later time, which were from days 16 or 28, respectively (see Figure 3B diagram on treatment schedule) than earlier studies of starting treatment at 7 days post-tumor establishment [20]. Although there is little evidence suggesting that a moderate delay in treatment would affect overall patient survival [45,46], we decided to investigate whether starting ΔM062R virotherapy or DC vaccine early in the combinatorial treatment offers any therapeutic benefit. We thus designed a follow-up study using the same ID8-TP53-/- syngeneic model system that will be discussed next.
3.4. M062R-Null (ΔM062R) MYXV Treatment in Combination with DC Vaccine Targeting Sp17 Antigen Significantly Improve the Treatment Outcome of Cisplatin.
In this second stage of study, we modified the treatment schedules of corresponding controls as shown in the diagram of Figure 4. We focused on the logistics of therapy design similar to a clinical setting and followed the following general principles, (1) to start the first treatment at 7 days post-tumor establishment, and (2) to start the DC vaccine at 8-9 days after the prior treatment for combination treatments. Although introducing ΔM062R treatment early as the first course of therapy (median survival of 59 days, p=0.0071, Kaplan-Meier analysis by Long-rank Mandel-Cox test) significantly improved the survival compared to mock treated group (median survival of 49 days), it did not provide a dramatic edge to prolong survival over either cisplatin alone (median 75, p=0.0019) or DC vaccine alone (median 75, p=0.0019). This is consistent with our previous finding on the utility and timing of ΔM062R [20]. We also found that as a monotherapy, early initiation of DC vaccine (median survival of 75 days) is as effective as cisplatin (median survival of 75 days) at the same time frame in this model (no statistically significant difference). Although two-regime and three-regime (all p<0.0001) combinatorial treatments significantly improved survival compared to mock, no significant difference among cisplatin plus ΔM062R (median survial 96 days), cisplatin plus DC (median survival 113 days), and cisplatin plus ΔM062R and DC vaccine (median survival 115 days) was observed, suggesting the importance of early cisplatin treatment in survival. Adding ΔM062R (p=0.0002) or DC vaccine (p<0.0001) to cisplatin significantly improved treatment outcome compared to cisplatin alone. Clustered ΔM062R treatment (four consecutive doses at every two days) at an early time (7 days post tumor establishment) followed by clustered DC vaccine treatment (four consecutive doses every week) showed no difference in survival to DC vaccine alone starting at 7 days. This prompted us to investigate utilizing individual ΔM062R and DC vaccine sequentially in a combination treatment to exploit the immunostimulatory properties of ΔM062R in facilitating the effects of DC vaccine that is shown in the next section.
3.5. Scheduling of ΔM062R MYXV Treatment Is Critical to Achieve the Optimal Immunotherapeutic Outcome.
To further investigate the immunotherapeutic potential of ΔM062R, we revised our experimental system to utilize OC ID8 cells engineered to overexpress Sp17 tumor antigen [26], designated ID8-Sp17. In this model system, we specifically investigate how ΔM062R impacts Sp17-targeted DC vaccine when Sp17 expression is elevated in tumor cells. We also reduced the dose of cisplatin from 3 mg/Kg to 1 mg/Kg, as it was shown that low dose cisplatin possessed immunotherapeutic benefit while maintaining its chemotherapeutic effect [47]. We tested if ΔM062R presents immunostimulatory effect directly to sequential DC vaccine as one integrated treatment course. As shown in Figure 5 top diagram, the three-regime combinatorial treatment, 6 days after the completion of cisplatin, ΔM062R virotherapy is followed by DC vaccine 2 days later, and this ΔM062R-DC vaccine sub-regime is repeated weekly 4 times. Besides the mock treatment group, single agent or dual agent regimes are scheduled in a fixed time-frame and frequency as shown in the Figure 5 top diagram for the corresponding agent such as cisplatin-ΔM062R-DC vaccine regime. We found that significant survival differences were observed (Kaplan-Meier survival and Long-rank Mandel-Cox test p<0.0001) (Figure 5 bottom, survival curve). Treatment with DC alone (median survival 110 days, p=0.0018) and ΔM062R alone (median survival 114 days, p=0.0197) significantly improved survival compared to mock treatment (median survival 76 days) and cisplatin only treatment (median survival 87 days) with p=0.0066 and p=0.0277 to DC alone and ΔM062R alone, respectively. Interestingly, we did not see significant differences in survival among ΔM062R plus DC (median survival 144 days), cisplatin plus ΔM062R (median survival 125 days), and the triple combinatorial regime (median survival undefined). However, these 3 treatments performed significantly better than cisplatin plus DC (median survival 100 days, ΔM062R plus DC p=0.0342, cisplatin plus ΔM062R p=0.0018, and triple-regime p=0.0018). Because of the relatively small sample size (n=5 per group), we did not see a significant difference between DC alone and cisplatin plus DC or ΔM062R plus DC, but the longer median survival times for the combination treatments reflected a trend of better survival. We thus concluded that to achieve a superior immunotherapeutic response, ΔM062R virotherapy should be placed prior to DC vaccine as a sub-regime. If low dose cisplatin is chosen for therapy, providing either ΔM062R or DC vaccine will lead to significantly improved survival, with ΔM062R (median survival 144 days) a better choice than DC vaccine (median survival 100 days). Overall, ΔM062R with DC drastically improved low dose cisplatin to achieve relatively long-term survival (median survival undefined).
3.6. M062R-Null MYXV Infection of Ovarian Cancer Patient Ascites CD14+ Cells Improved CD4+ T Cell Anti-Tumor Response in a Primary Cell Co-Culture System.
It is known that MYXV favors infecting monocytes/macrophages [48], providing a mechanistic rationale of targeting myeloid cells for immunotherapy with these viral agents. Based on our prior finding on the potent immunostimulatory effect by ΔM062R in monocytes and macrophages [18,43], we further examined whether ΔM062R immunotherapeutic benefit is due to its effects on immunosuppressive tumor-associated monocyte/macropahges or TAM population in the tumor environment, designated immunosuppressive CD14+ tumor-associated macrophages (TAMs). We thus utilized a highly innovative co-culture system to test how OC patient CD4+ T cell functions are regulated in OC tumor environment. It is known that in OC CD14+ TAMs suppress the anti-tumor function of CD4+ T cells [33]. The key question is whether MYXV influences or even subverts immunosuppression by CD14+ TAMs to enable a more effective anti-tumor CD4+ T cell response. In this co-culture system, patient CD4+ T cells were stimulated with tumor antigen (TADG14v)[33] pulsed DC in the presence of TAMs with or without prior treatments. We then examined the status of CD4+ T cell activation reflected by TNF production via flow cytometry [33]. As shown in Figure 6A, in the absence of CD14+ TAMs, human CD4+ T cells responded to tumor antigen with TNF production (Figure 6A first column). In the presence of TAMs, however, upon the same stimulation of OC tumor antigen, CD4+ T cells presented significant reduction of TNF production (Figure 6A second column). Both WT and ΔM062R treatment of TAMs reversed the immunosuppressive effect by TAMs but with ΔM062R treatment leading to promotion of T cell activation. Shown in Figure 6A (top panel) is a representative result from an allogeneic co-culture system with CD4+ T cells and CD14+ TAMs from different donors. We also performed similar tests using autologous specimens for the co-culture system and achieved similar results (shown in Figure 6A bottom panel).
Our previous study showed that ΔM062R activated CD14+ monocytes/macrophages to promote proinflammatory responses regulated by both DNA-sensing via cGAS/STING/TBK1/IRF3 axis and SAMD9 pathway [18]. Using a STING antagonist, H151, and a TBK1 inhibitor, BX-795, we found that reversal of TAM immunosuppression by ΔM062R was not affected (Figure 6B). We thus conclude that therapeutic effect of ΔM062R against immunosuppressive TAMs is independent from cGAS/STING/TBK1/IRF3 axis and is likely determined by SAMD9 pathway. This is consistent with our observation that ΔM062R treatment induced a unique transcriptional landscape and cellular translation profile independent from DNA-sensing pathway [43].
4. Discussion
MYXV was first formulated as oncolytic agent almost 2 decades ago [49] and have been examined for their therapeutic potential against multiple cancer types [50,51,52,53]. The WT MYXV and replication competent MYXVs armed with therapeutic molecules have been developed to purge metastatic [16,51] and hematological malignant cells [54,55,56] as well as being loaded on vehicle cells for a trojan-horse like delivery [19,57]. Furthermore, MYXV can be used in coordination with chemotherapeutic agents to achieve synergistic effect for improved treatment outcome in preclinical models [20,21,58,59]. Our previous study found that replication-competent WT MYXV provided better therapeutic benefit in an OC carcinomatosis model when it is used before cisplatin treatment [20]. Surprisingly, a replication-defective ΔM062R MYXV achieved superior therapeutic benefit as a monotherapy and as an adjuvant with cisplatin in combatting OC carcinomatosis [20]. In contrast with WT MYXV, the optimal schedule for ΔM062R MYXV is to be used after cisplatin treatment [20]. Infection by ΔM062R does not lead to progeny viral production [27,60], thus, treatment with ΔM062R does not cause amplifying oncolysis seen with WT MYXV. Our recent work demonstrated that ΔM062R infection triggered proinflammatory cytokine/chemokine production in monocytes/macrophages [18]. Such proinflammatory response is triggered in part through DNA-sensing pathway leading to IFN-I production [18]. However, upon transcriptomic analyses on ΔM062R effect to monocytes/macrophages, a distinct feature from that of cGAS/STING/TBK1/IRF3 axis of DNA sensing pathway was observed in monocytes/macrophages by ΔM062R [43]. This unique response is likely due to the activation of another host immunoregulatory factor, SAMD9 that is the host target of viral M062 protein [27,61]. Activating the SAMD9 pathway by ΔM062R is associated with the immunotherapeutic potential of this replication-defective viral agent. Although the concept of immunotherapeutic potential of oncolytic viruses have been recognized [62], the therapeutic promise of a replication defective viral vector is not yet broadly recognized. In this study, we presented strong evidence that such a replication-defective ΔM062R MYXV can enhance immunotherapy for OC and alleviate ovarian TAM immunosuppression of DC-stimulated tumor antigen-specific human CD4+ T cell responses. The MYXV ΔM062R platform is an excellent option for immunotherapy in OC and can be utilized as an adjuvant to existing chemotherapy and DC vaccine.
The utility of ΔM062R in immunotherapy is due to its immunostimulatory property by activating DNA-sensing associated IFN-I induction and the SAMD9 pathway [18,43]. However, the timing and design of incorporating ΔM062R treatment is important to achieve therapeutic benefit. We found that providing ΔM062R early before the establishment of tumor environment (1 week after tumor cell inoculation) did not render significant benefit as monotherapy nor boosted the benefit of DC vaccine (Figure 4B and 4E). On the other hand, when ΔM062R treatment is provided much later during disease progression (a month after tumor cell inoculation), as monotherapy it outperformed cisplatin monotherapy that started 2 weeks before ΔM062R treatment (Figure 3B). More importantly, adding ΔM062R treatment after chemotherapy in either clustered regime preceding DC vaccine (Figure 3B and Figure 4E) or individually administrated preceding DC vaccine (Figure 5) provided superior treatment advantage over chemotherapy alone, making ΔM062R regimens logical choices for therapeutic design in the clinical setting for OC patients, especially after tumor debunking and the initial chemotherapy. Among human leukocytes, MYXV presented preferred binding and entry in myeloid cells such as monocytes and macrophages [48], which establishes the rationale of using this viral vector against TAMs. To investigate possible mechanisms of ΔM062R-DC vaccine combinatorial treatment, we examined how ΔM062R influenced monocytes/macrophages to activate innate immune signaling events [18,43]. In OC TAMs, ΔM062R likely silences the expression of immunosuppressive cytokines/chemokines [20] or even promotes them to produce proinflammatory molecules. Alleviation of immunosuppression by CD14+ TAMs allows stronger CD4+ T cell responses to tumor antigen stimulation (Figure 6A). Besides being an informative predictor for prognosis [63], the immunosuppressive properties of the OC tumor environment also affect the efficacy of immunotherapy agents including DC vaccine [64] and CAR-T therapy [65,66]. The immunotherapeutic benefit of ΔM062R may not be restricted to monocytes/macrophages, as therapeutic benefit to influence T cell responses by MYXV has also been reported [67]. MYXV hijacks cells with short circuited IFN-I responses, often malignant cells or cells cultured in the tumor environment [68], but does not affect the functionality of normal cells such as hematopoietic stem cells [54,69], progenitor cells [54,69], and normal T cells [67,70], further rendering a safety profile of ΔM062R for clinical applications in the future.
5. Patents
A US patent resulting from the work reported in this manuscript, US 12,128,099 B2, was awarded to J.L. and M.J.C.
Author Contributions
Conceptualization, M.J.C and J.L.; methodology, M.J.C and J.L.; software, M.J.C. and J.L.; formal analysis, M.J.C and J.L.; investigation, M.J.C and J.L.; resources, J.L.; data curation, J.L.; writing—original draft preparation, J.L.; writing—review and editing, M.J.C and J.L.; visualization, J.L.; project administration, M.J.C. and J.L.; funding acquisition, J.L. All authors have read and agreed to the published version of the manuscript.
Funding
The study was supported by NIH R01AI139106 to J.L., a start-up by UAMS to J.L., the UAMS VCRI pioneer award to J.L., River Valley Ovarian Cancer Alliance Research Award to J.L., and supported in part by the Center for Microbial Pathogenesis and Host Inflammatory Responses grant P20GM103625 through the NIH National Institute of General Medical Sciences Centers of Biomedical Research Excellence.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the IRB of UAMS and the Mayo Clinic. The animal study protocol was approved by the IACUC of UAMS.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| MYXV | Myxoma virus |
| OC | Ovarian cancer |
| DC | Dendritic cell |
| TAM | Tumor-associated macrophage |
| HGSOC. | High grade serous ovarian cancer |
References
- Lengyel E 2010 Ovarian cancer development and metastasis Am J Pathol 177 1053–64. [CrossRef]
- Ozols R F 2006 Challenges for chemotherapy in ovarian cancer Ann Oncol 17 Suppl 5 v181-7. [CrossRef]
- Cummings M, Freer C and Orsi N M 2021 Targeting the tumour microenvironment in platinum-resistant ovarian cancer Semin. Cancer Biol. 77 3–28. [CrossRef]
- Sung H, Ferlay J, Siegel R L, Laversanne M, Soerjomataram I, Jemal A and Bray F 2021 Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries CA. Cancer J. Clin. 71 209–49. [CrossRef]
- Richardson D L, Eskander R N and O’Malley D M 2023 Advances in Ovarian Cancer Care and Unmet Treatment Needs for Patients With Platinum Resistance: A Narrative Review JAMA Oncol. 9 851–9. [CrossRef]
- Coleman M P, Forman D, Bryant H, Butler J, Rachet B, Maringe C, Nur U, Tracey E, Coory M, Hatcher J, McGahan C E, Turner D, Marrett L, Gjerstorff M L, Johannesen T B, Adolfsson J, Lambe M, Lawrence G, Meechan D, Morris E J, Middleton R, Steward J, Richards M A and Group I M 1 W 2011 Cancer survival in Australia, Canada, Denmark, Norway, Sweden, and the UK, 1995-2007 (the International Cancer Benchmarking Partnership): an analysis of population-based cancer registry data Lancet 377 127–38. [CrossRef]
- Baert T, Timmerman D, Vergote I and Coosemans A 2015 Immunological parameters as a new lead in the diagnosis of ovarian cancer Facts Views Vis Obgyn 7 67–72.
- Coosemans A, Decoene J, Baert T, Laenen A, Kasran A, Verschuere T, Seys S and Vergote I 2015 Immunosuppressive parameters in serum of ovarian cancer patients change during the disease course Oncoimmunology 5 e1111505. [CrossRef]
- Krishnan V, Schaar B, Tallapragada S and Dorigo O 2018 Tumor associated macrophages in gynecologic cancers Gynecol. Oncol. 149 205–13. [CrossRef]
- Blanc-Durand F, Clemence Wei Xian L and Tan D S P 2023 Targeting the immune microenvironment for ovarian cancer therapy Front. Immunol. 14 1328651. [CrossRef]
- Goyne H E and Cannon M J 2013 Dendritic cell vaccination, immune regulation, and clinical outcomes in ovarian cancer Front Immunol 4 382. [CrossRef]
- Cannon M J, Goyne H, Stone P J B and Chiriva-Internati M 2011 Dendritic cell vaccination against ovarian cancer--tipping the Treg/TH17 balance to therapeutic advantage? Expert Opin. Biol. Ther. 11 441–5. [CrossRef]
- Rahman M M and McFadden G 2020 Oncolytic Virotherapy with Myxoma Virus J. Clin. Med. 9 171. [CrossRef]
- Stanford M M, Barrett J W, Gilbert P A, Bankert R and McFadden G 2007 Myxoma virus expressing human interleukin-12 does not induce myxomatosis in European rabbits J Virol 81 12704–8. [CrossRef]
- Tosic V, Thomas D L, Kranz D M, Liu J, McFadden G, Shisler J L, MacNeill A L and Roy E J 2014 Myxoma virus expressing a fusion protein of interleukin-15 (IL15) and IL15 receptor alpha has enhanced antitumor activity PLoS One 9 e109801. [CrossRef]
- Christie J D, Appel N, Canter H, Achi J G, Elliott N M, Matos A L de, Franco L, Kilbourne J, Lowe K, Rahman M M, Villa N Y, Carmen J, Luna E, Blattman J and McFadden G 2021 Systemic delivery of TNF-armed myxoma virus plus immune checkpoint inhibitor eliminates lung metastatic mouse osteosarcoma Mol. Ther. - Oncolytics 22 539–54. [CrossRef]
- Chan W M, Rahman M M and McFadden G 2013 Oncolytic myxoma virus: The path to clinic Vaccine. [CrossRef]
- Conrad S J, Raza T, Peterson E A, Liem J, Connor R, Nounamo B, Cannon M and Liu J 2022 Myxoma virus lacking the host range determinant M062 stimulates cGAS-dependent type 1 interferon response and unique transcriptomic changes in human monocytes/macrophages PLoS Pathog. 18 e1010316. [CrossRef]
- Zheng N, Fang J, Xue G, Wang Z, Li X, Zhou M, Jin G, Rahman M M, McFadden G and Lu Y 2022 Induction of tumor cell autosis by myxoma virus-infected CAR-T and TCR-T cells to overcome primary and acquired resistance Cancer Cell 40 973-985.e7. [CrossRef]
- Nounamo B, Liem J, Cannon M and Liu J 2017 Myxoma virus optimizes cisplatin for the treatment of ovarian cancer in vitro and in a syngeneic murine dissemination model Mol. Ther.-Oncolytics 6 90–9. [CrossRef]
- Wennier S T, Liu J, Li S, Rahman M M, Mona M and McFadden G 2012 Myxoma virus sensitizes cancer cells to gemcitabine and is an effective oncolytic virotherapeutic in models of disseminated pancreatic cancer Mol Ther 20 759–68. [CrossRef]
- Jung M-Y, Offord C P, Ennis M K, Kemler I, Neuhauser C and Dingli D 2018 In vivo estimation of oncolytic virus populations within tumors Cancer Res. 78 5992–6000. [CrossRef]
- Lin D, Shen Y and Liang T 2023 Oncolytic virotherapy: basic principles, recent advances and future directions Signal Transduct. Target. Ther. 8 1–29. [CrossRef]
- Block M S, Dietz A B, Gustafson M P, Kalli K R, Erskine C L, Youssef B, Vijay G V, Allred J B, Pavelko K D, Strausbauch M A, Lin Y, Grudem M E, Jatoi A, Klampe C M, Wahner-Hendrickson A E, Weroha S J, Glaser G E, Kumar A, Langstraat C L, Solseth M L, Deeds M C, Knutson K L and Cannon M J 2020 Th17-inducing autologous dendritic cell vaccination promotes antigen-specific cellular and humoral immunity in ovarian cancer patients Nat. Commun. 11 5173. [CrossRef]
- Walton J, Blagih J, Ennis D, Leung E, Dowson S, Farquharson M, Tookman L A, Orange C, Athineos D, Mason S, Stevenson D, Blyth K, Strathdee D, Balkwill F R, Vousden K, Lockley M and McNeish I A 2016 CRISPR/Cas9-Mediated Trp53 and Brca2 Knockout to Generate Improved Murine Models of Ovarian High-Grade Serous Carcinoma Cancer Res 76 6118–29. [CrossRef]
- Luo Y, Shreeder B, Jenkins J W, Shi H, Lamichhane P, Zhou K, Bahr D A, Kurian S, Jones K A, Daum J I, Dutta N, Necela B M, Cannon M J, Block M S and Knutson K L 2023 Th17-inducing dendritic cell vaccines stimulate effective CD4 T cell-dependent antitumor immunity in ovarian cancer that overcomes resistance to immune checkpoint blockade J. Immunother. Cancer 11 e007661. [CrossRef]
- Liu J, Wennier S, Zhang L and McFadden G 2011 M062 is a host range factor essential for myxoma virus pathogenesis and functions as an antagonist of host SAMD9 in human cells J Virol 85 3270–82. [CrossRef]
- Smallwood S E, Rahman M M, Smith D W and McFadden G 2010 Myxoma virus: propagation, purification, quantification, and storage Curr Protoc Microbiol Chapter 14 Unit 14A 1. [CrossRef]
- Lee P Y, Li Y, Kumagai Y, Xu Y, Weinstein J S, Kellner E S, Nacionales D C, Butfiloski E J, van Rooijen N, Akira S, Sobel E S, Satoh M and Reeves W H 2009 Type I Interferon Modulates Monocyte Recruitment and Maturation in Chronic Inflammation Am. J. Pathol. 175 2023–33. [CrossRef]
- Lee P Y, Weinstein J S, Nacionales D C, Scumpia P O, Li Y, Butfiloski E, van Rooijen N, Moldawer L, Satoh M and Reeves W H 2008 A Novel Type I IFN-Producing Cell Subset in Murine Lupus1 J. Immunol. 180 5101–8. [CrossRef]
- Lee P Y, Kumagai Y, Li Y, Takeuchi O, Yoshida H, Weinstein J, Kellner E S, Nacionales D, Barker T, Kelly-Scumpia K, van Rooijen N, Kumar H, Kawai T, Satoh M, Akira S and Reeves W H 2008 TLR7-dependent and FcγR-independent production of type I interferon in experimental mouse lupus J. Exp. Med. 205 2995–3006. [CrossRef]
- Lee P Y, Kumagai Y, Xu Y, Li Y, Barker T, Liu C, Sobel E S, Takeuchi O, Akira S, Satoh M and Reeves WestleyH 2011 Interleukin-1 alpha modulates neutrophil recruitment in chronic inflammation induced by hydrocarbon oil J. Immunol. Baltim. Md 1950 186 1747–54. [CrossRef]
- Goyne H E, Stone P J, Burnett A F and Cannon M J 2014 Ovarian tumor ascites CD14+ cells suppress dendritic cell-activated CD4+ T-cell responses through IL-10 secretion and indoleamine 2,3-dioxygenase J Immunother 37 163–9. [CrossRef]
- Cannon M J, Goyne H E, Stone P J, Macdonald L J, James L E, Cobos E and Chiriva-Internati M 2013 Modulation of p38 MAPK signaling enhances dendritic cell activation of human CD4+ Th17 responses to ovarian tumor antigen Cancer Immunol Immunother 62 839–49. [CrossRef]
- Cannon M J, Ghosh D and Gujja S 2015 Signaling Circuits and Regulation of Immune Suppression by Ovarian Tumor-Associated Macrophages Vaccines 3 448–66. [CrossRef]
- Straughn J M Jr, Shaw D R, Guerrero A, Bhoola S M, Racelis A, Wang Z, Chiriva-Internati M, Grizzle W E, Alvarez R D, Lim S H and Strong T V 2004 Expression of sperm protein 17 (Sp17) in ovarian cancer Int J Cancer 108 805–11. [CrossRef]
- Chiriva-Internati M, Grizzi F, Weidanz J A, Ferrari R, Yuefei Y, Velez B, Shearer M H, Lowe D B, Frezza E E, Cobos E, Kast W M and Kennedy R C 2007 A NOD/SCID tumor model for human ovarian cancer that allows tracking of tumor progression through the biomarker Sp17 J Immunol Methods 321 86–93. [CrossRef]
- Song J X, Cao W L, Li F Q, Shi L N and Jia X 2012 Anti-Sp17 monoclonal antibody with antibody-dependent cell-mediated cytotoxicity and complement-dependent cytotoxicity activities against human ovarian cancer cells Med Oncol 29 2923–31. [CrossRef]
- Xiang S D, Gao Q, Wilson K L, Heyerick A and Plebanski M 2015 A Nanoparticle Based Sp17 Peptide Vaccine Exposes New Immuno-Dominant and Species Cross-reactive B Cell Epitopes Vaccines Basel 3 875–93. [CrossRef]
- Song J X, Li F Q, Cao W L, Jia X, Shi L N, Lu J F, Ma C F and Kong Q Q 2014 Anti-Sp17 monoclonal antibody-doxorubicin conjugates as molecularly targeted chemotherapy for ovarian carcinoma Target Oncol 9 263–72. [CrossRef]
- Ait-Tahar K, Anderson A P, Barnardo M, Collins G P, Hatton C S R, Banham A H and Pulford K 2017 Sp17 Protein Expression and Major Histocompatibility Class I and II Epitope Presentation in Diffuse Large B Cell Lymphoma Patients Adv Hematol 2017 6527306. [CrossRef]
- Chiriva-Internati M, Yu Y, Mirandola L, Jenkins M R, Chapman C, Cannon M, Cobos E and Kast W M 2010 Cancer testis antigen vaccination affords long-term protection in a murine model of ovarian cancer PLoS One 5 e10471. [CrossRef]
- Raza T, Perterson E, Liem J and Liu J 2024 Antagonizing the SAMD9 pathway is key to myxoma virus host shut-off and immune evasion BioRxiv Prepr. Serv. Biol. 2024.02.01.578447. [CrossRef]
- Ahmed A A, Etemadmoghadam D, Temple J, Lynch A G, Riad M, Sharma R, Stewart C, Fereday S, Caldas C, Defazio A, Bowtell D and Brenton J D 2010 Driver mutations in TP53 are ubiquitous in high grade serous carcinoma of the ovary J Pathol 221 49–56. [CrossRef]
- Zhao J, Chen R, Zhang Y, Wang Y and Zhu H 2024 Impact of Treatment Delay on the Prognosis of Patients with Ovarian Cancer: A Population-based Study Using the Surveillance, Epidemiology, and End Results Database J. Cancer 15 473–83. [CrossRef]
- Nagel C, Backes F, Donner J, Bussewitz E, Hade E, Cohn D, Eisenhauer E, O’Malley D, Fowler J, Copeland L and Salani R 2012 Effect of chemotherapy delays and dose reductions on progression free and overall survival in the treatment of epithelial ovarian cancer Gynecol. Oncol. 124 221–4. [CrossRef]
- Hopkins D, Sanchez H, Berwin B and Wilkinson-Ryan I 2021 Cisplatin increases immune activity of monocytes and cytotoxic T-cells in a murine model of epithelial ovarian cancer Transl. Oncol. 14 101217. [CrossRef]
- Chan W M, Bartee E C, Moreb J S, Dower K, Connor J H and McFadden G 2013 Myxoma and vaccinia viruses bind differentially to human leukocytes J. Virol. 87 4445–60. [CrossRef]
- Lun X, Yang W, Alain T, Shi Z Q, Muzik H, Barrett J W, McFadden G, Bell J, Hamilton M G, Senger D L and Forsyth P A 2005 Myxoma virus is a novel oncolytic virus with significant antitumor activity against experimental human gliomas Cancer Res 65 9982–90. [CrossRef]
- Rahman M M and McFadden G 2020 Oncolytic Virotherapy with Myxoma Virus J. Clin. Med. 9 171. [CrossRef]
- Stanford M M, Shaban M, Barrett J W, Werden S J, Gilbert P A, Bondy-Denomy J, Mackenzie L, Graham K C, Chambers A F and McFadden G 2008 Myxoma virus oncolysis of primary and metastatic B16F10 mouse tumors in vivo Mol Ther 16 52–9. [CrossRef]
- Stanford M M, Barrett J W, Nazarian S H, Werden S and McFadden G 2007 Oncolytic virotherapy synergism with signaling inhibitors: Rapamycin increases myxoma virus tropism for human tumor cells J Virol 81 1251–60. [CrossRef]
- Tong J G, Valdes Y R, Barrett J W, Bell J C, Stojdl D, McFadden G, McCart J A, DiMattia G E and Shepherd T G 2015 Evidence for differential viral oncolytic efficacy in an in vitro model of epithelial ovarian cancer metastasis Mol Ther Oncolytics 2 15013. [CrossRef]
- Bartee E, Chan W M, Moreb J S, Cogle C R and McFadden G 2012 Selective purging of human multiple myeloma cells from autologous stem cell transplantation grafts using oncolytic myxoma virus Biol Blood Marrow Transpl. 18 1540–51. [CrossRef]
- Bartee M Y, Dunlap K M and Bartee E 2016 Myxoma Virus Induces Ligand Independent Extrinsic Apoptosis in Human Myeloma Cells Clin Lymphoma Myeloma Leuk 16 203–12. [CrossRef]
- Madlambayan G J, Bartee E, Kim M, Rahman M M, Meacham A, Scott E W, McFadden G and Cogle C R 2012 Acute myeloid leukemia targeting by myxoma virus in vivo depends on cell binding but not permissiveness to infection in vitro Leuk Res 36 619–24. [CrossRef]
- Jazowiecka-Rakus J, Pogoda-Mieszczak K, Rahman M M, McFadden G and Sochanik A 2024 Adipose-Derived Stem Cells as Carrier of Pro-Apoptotic Oncolytic Myxoma Virus: To Cross the Blood-Brain Barrier and Treat Murine Glioma Int. J. Mol. Sci. 25 11225. [CrossRef]
- Thomas D L, Doty R, Tosic V, Liu J, Kranz D M, McFadden G, Macneill A L and Roy E J 2011 Myxoma virus combined with rapamycin treatment enhances adoptive T cell therapy for murine melanoma brain tumors Cancer Immunol Immunother 60 1461–72. [CrossRef]
- Zemp F J, Lun X, McKenzie B A, Zhou H, Maxwell L, Sun B, Kelly J J, Stechishin O, Luchman A, Weiss S, Cairncross J G, Hamilton M G, Rabinovich B A, Rahman M M, Mohamed M R, Smallwood S, Senger D L, Bell J, McFadden G and Forsyth P A 2013 Treating brain tumor-initiating cells using a combination of myxoma virus and rapamycin Neuro Oncol 15 904–20. [CrossRef]
- Nounamo B, Li Y, O’Byrne P, Kearney A M, Khan A and Liu J 2017 An interaction domain in human SAMD9 is essential for myxoma virus host-range determinant M062 antagonism of host anti-viral function Virology 503 94–102. [CrossRef]
- Liu J and McFadden G 2015 SAMD9 is an innate antiviral host factor with stress response properties that can be antagonized by poxviruses J. Virol. 89 1925–31. [CrossRef]
- Liu J, Wennier S and McFadden G 2010 The immunoregulatory properties of oncolytic myxoma virus and their implications in therapeutics Microbes Infect 12 1144–52. [CrossRef]
- Kroeger D R, Milne K and Nelson B H 2016 Tumor-Infiltrating Plasma Cells Are Associated with Tertiary Lymphoid Structures, Cytolytic T-Cell Responses, and Superior Prognosis in Ovarian Cancer Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 22 3005–15. [CrossRef]
- Lee K-W, Yam J W P and Mao X 2023 Dendritic Cell Vaccines: A Shift from Conventional Approach to New Generations Cells 12 2147. [CrossRef]
- Zhang X-W, Wu Y-S, Xu T-M and Cui M-H 2023 CAR-T Cells in the Treatment of Ovarian Cancer: A Promising Cell Therapy Biomolecules 13 465. [CrossRef]
- Kang C, Jeong S-Y, Song S Y and Choi E K 2020 The emerging role of myeloid-derived suppressor cells in radiotherapy Radiat. Oncol. J. 38 1–10. [CrossRef]
- Villa N Y, Rahman M M, Mamola J, Sharik M E, de Matos A L, Kilbourne J, Lowe K, Daggett-Vondras J, D’Isabella J, Goras E, Chesi M, Bergsagel P L and McFadden G 2022 Transplantation of autologous bone marrow pre-loaded ex vivo with oncolytic myxoma virus is efficacious against drug-resistant Vk*MYC mouse myeloma Oncotarget 13 490–504. [CrossRef]
- Bartee E, Chan W S, Moreb J S, Cogle C R and McFadden G 2012 Selective purging of human multiple myeloma cells from autologous stem cell transplant grafts using oncolytic myxoma virus Biol. Blood Marrow Transplant. J. Am. Soc. Blood Marrow Transplant. 18 1540–51. [CrossRef]
- Villa N Y, Bais S, Chan W M, Meacham A M, Wise E, Rahman M M, Moreb J S, Rosenau E H, Wingard J R, McFadden G and Cogle C R 2016 Ex vivo virotherapy with myxoma virus does not impair hematopoietic stem and progenitor cells Cytotherapy 18 465–80. [CrossRef]
- Villa N Y, Rahman M M, McFadden G and Cogle C R 2016 Therapeutics for Graft-versus-Host Disease: From Conventional Therapies to Novel Virotherapeutic Strategies Viruses 8 85. [CrossRef]
Figure 1.
Myxoma virus treatments of primary human ovarian cancer ascites cells induce proinflammatory molecule expression and upregulation of Sp17. Ascites cells from OC patients, (A) OvCa37, (B) KAJ343, and (C) OvCa43, were mock treated or infected with MYXV, WT or ΔM062R, at an MOI of 5 for 18 h before cells were harvested for RNA extraction and RT2-PCR to assay target gene expression levels for semi-quantitative comparison. Red frame marks the ovarian cancer cell specific antigen, Sp17, expression levels.
Figure 1.
Myxoma virus treatments of primary human ovarian cancer ascites cells induce proinflammatory molecule expression and upregulation of Sp17. Ascites cells from OC patients, (A) OvCa37, (B) KAJ343, and (C) OvCa43, were mock treated or infected with MYXV, WT or ΔM062R, at an MOI of 5 for 18 h before cells were harvested for RNA extraction and RT2-PCR to assay target gene expression levels for semi-quantitative comparison. Red frame marks the ovarian cancer cell specific antigen, Sp17, expression levels.
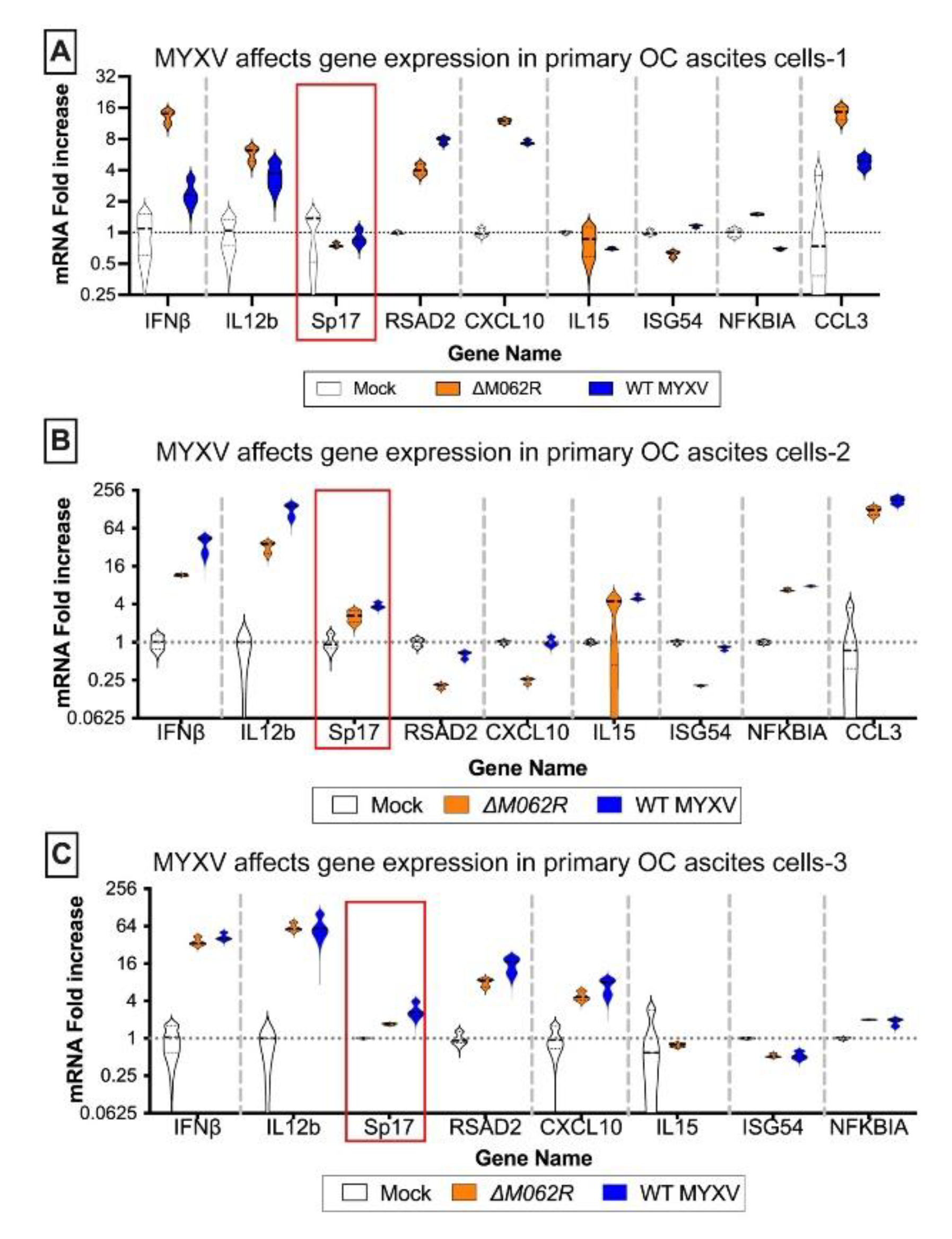
Figure 2.
Myxoma virus treatments of murine ID8 TP53-/- induce inflammatory gene expression and alter gene expression affecting tumor environment. Murine OC cells were mock treated or infected with MYXV at an MOI of 5 for 18 h before RNA extraction and RT2-PCR. Genes related to inflammatory responses (A) and contributing to tumor environment maintenance (B) were examined for their expression affected by MYXV treatments. In either array, OC tumor marker, murine Sp17, was also examined.
Figure 2.
Myxoma virus treatments of murine ID8 TP53-/- induce inflammatory gene expression and alter gene expression affecting tumor environment. Murine OC cells were mock treated or infected with MYXV at an MOI of 5 for 18 h before RNA extraction and RT2-PCR. Genes related to inflammatory responses (A) and contributing to tumor environment maintenance (B) were examined for their expression affected by MYXV treatments. In either array, OC tumor marker, murine Sp17, was also examined.
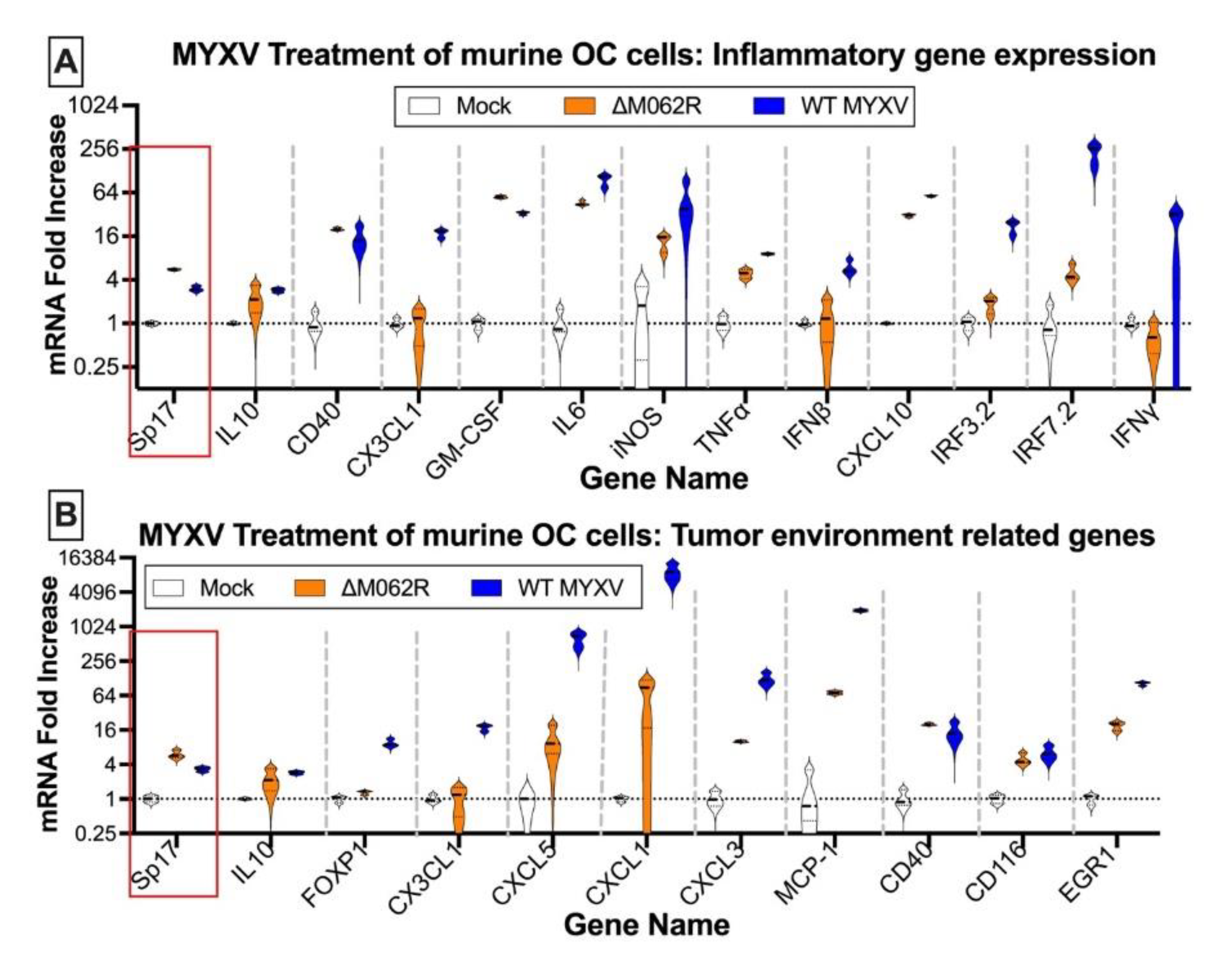
Figure 3.
Myxoma virus treatments facilitate survival in the combinatorial treatment with cisplatin and DC vaccine. C57/BL6 mice were injected through intraperitoneal route (i.p.) with 3x106 ID8-TP53-/- cells on day 0 and randomly grouped to 5 mice per treatment. Cisplatin was injected at 3 mg/kg i.p. on days 16, 19 and 22. WT-MYXV (108 pfu, i.p.) (A) was injected on days 7, 9, 11 and 13 (before cisplatin treatment), while ΔM062R MYXV (108 pfu i.p.) (B) was injected on days 28, 30, 32 and 34 (following cisplatin treatment). Sp17 peptide-loaded DC (106) were injected subcutaneously (s.c.) on days 30, 37, 44 and 51 for WT-MYXV recipients and days 44, 51, 58 and 65 for ΔM062R MYXV recipients. The log-rank (Mantel-Cox) test was conducted for Kaplan-Meier survival comparison. * p<0.05, ** p<0.001, n.s.: not significant.
Figure 3.
Myxoma virus treatments facilitate survival in the combinatorial treatment with cisplatin and DC vaccine. C57/BL6 mice were injected through intraperitoneal route (i.p.) with 3x106 ID8-TP53-/- cells on day 0 and randomly grouped to 5 mice per treatment. Cisplatin was injected at 3 mg/kg i.p. on days 16, 19 and 22. WT-MYXV (108 pfu, i.p.) (A) was injected on days 7, 9, 11 and 13 (before cisplatin treatment), while ΔM062R MYXV (108 pfu i.p.) (B) was injected on days 28, 30, 32 and 34 (following cisplatin treatment). Sp17 peptide-loaded DC (106) were injected subcutaneously (s.c.) on days 30, 37, 44 and 51 for WT-MYXV recipients and days 44, 51, 58 and 65 for ΔM062R MYXV recipients. The log-rank (Mantel-Cox) test was conducted for Kaplan-Meier survival comparison. * p<0.05, ** p<0.001, n.s.: not significant.
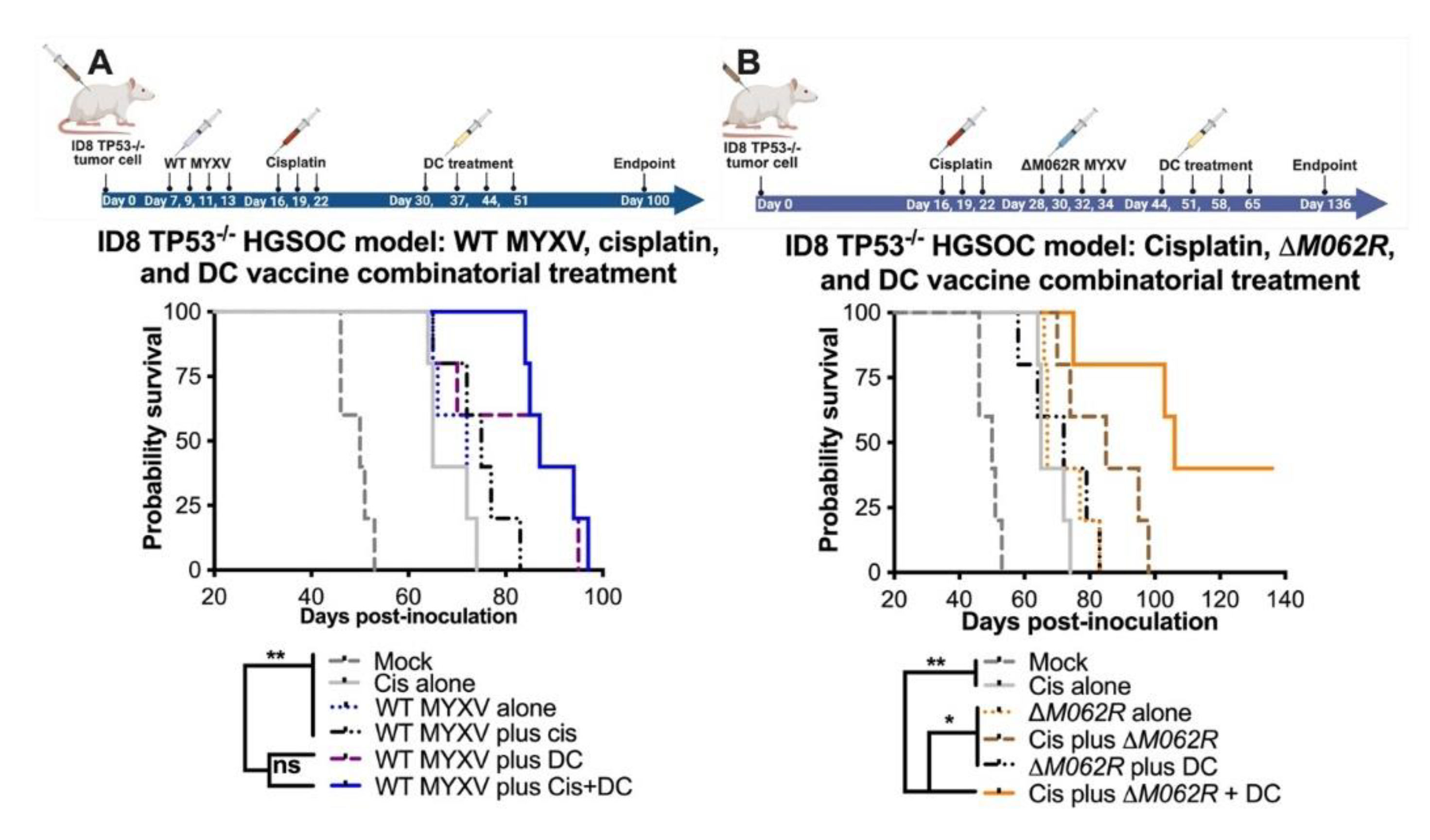
Figure 4.
Treatment of M062R further enhances therapeutic effect by DC vaccine and cisplatin in a combinatorial treatment. C57/BL6 mice were injected i.p. with 3x106 ID8-TP53-/- cells on day 0. Five mice were randomly assigned per group for mock and the individual treatment controls, and 10 mice were assigned per group for double or triple combinatorial treatments. Regime design is shown in A for cisplatin plus ΔM062R and the triple combinatorial treatment. B. Treatment design for ΔM062R alone or combination of ΔM062R and DC vaccine. Treatment schedule of DC vaccine alone is shown in C. Treatment schedule of cisplatin alone or cisplatin plus DC vaccine schedule is shown in D. In general, treatment starts at 7 days after tumor establishment. Treatment with ΔM062R is at 108 pfu, i.p. every 2 days for a total of 4 injections. Cisplatin was injected i.p. at 3 mg/Kg every 3 days for a total of 3 treatments. DC vaccine is injected s.c. 8-9 days after the previous treatment (if it is a combination treatment) every 7 days for a total of 4 vaccinations. E. Kaplan-Meier survival curve is shown and ΔM062R plus DC provided superior therapeutic outcome with or without cisplatin. The log-rank (Mantel-Cox) test was conducted with statistical significance defined as the following, * p<0.05, ** p<0.001, **** p<0.0001, and n.s.: not significant.
Figure 4.
Treatment of M062R further enhances therapeutic effect by DC vaccine and cisplatin in a combinatorial treatment. C57/BL6 mice were injected i.p. with 3x106 ID8-TP53-/- cells on day 0. Five mice were randomly assigned per group for mock and the individual treatment controls, and 10 mice were assigned per group for double or triple combinatorial treatments. Regime design is shown in A for cisplatin plus ΔM062R and the triple combinatorial treatment. B. Treatment design for ΔM062R alone or combination of ΔM062R and DC vaccine. Treatment schedule of DC vaccine alone is shown in C. Treatment schedule of cisplatin alone or cisplatin plus DC vaccine schedule is shown in D. In general, treatment starts at 7 days after tumor establishment. Treatment with ΔM062R is at 108 pfu, i.p. every 2 days for a total of 4 injections. Cisplatin was injected i.p. at 3 mg/Kg every 3 days for a total of 3 treatments. DC vaccine is injected s.c. 8-9 days after the previous treatment (if it is a combination treatment) every 7 days for a total of 4 vaccinations. E. Kaplan-Meier survival curve is shown and ΔM062R plus DC provided superior therapeutic outcome with or without cisplatin. The log-rank (Mantel-Cox) test was conducted with statistical significance defined as the following, * p<0.05, ** p<0.001, **** p<0.0001, and n.s.: not significant.
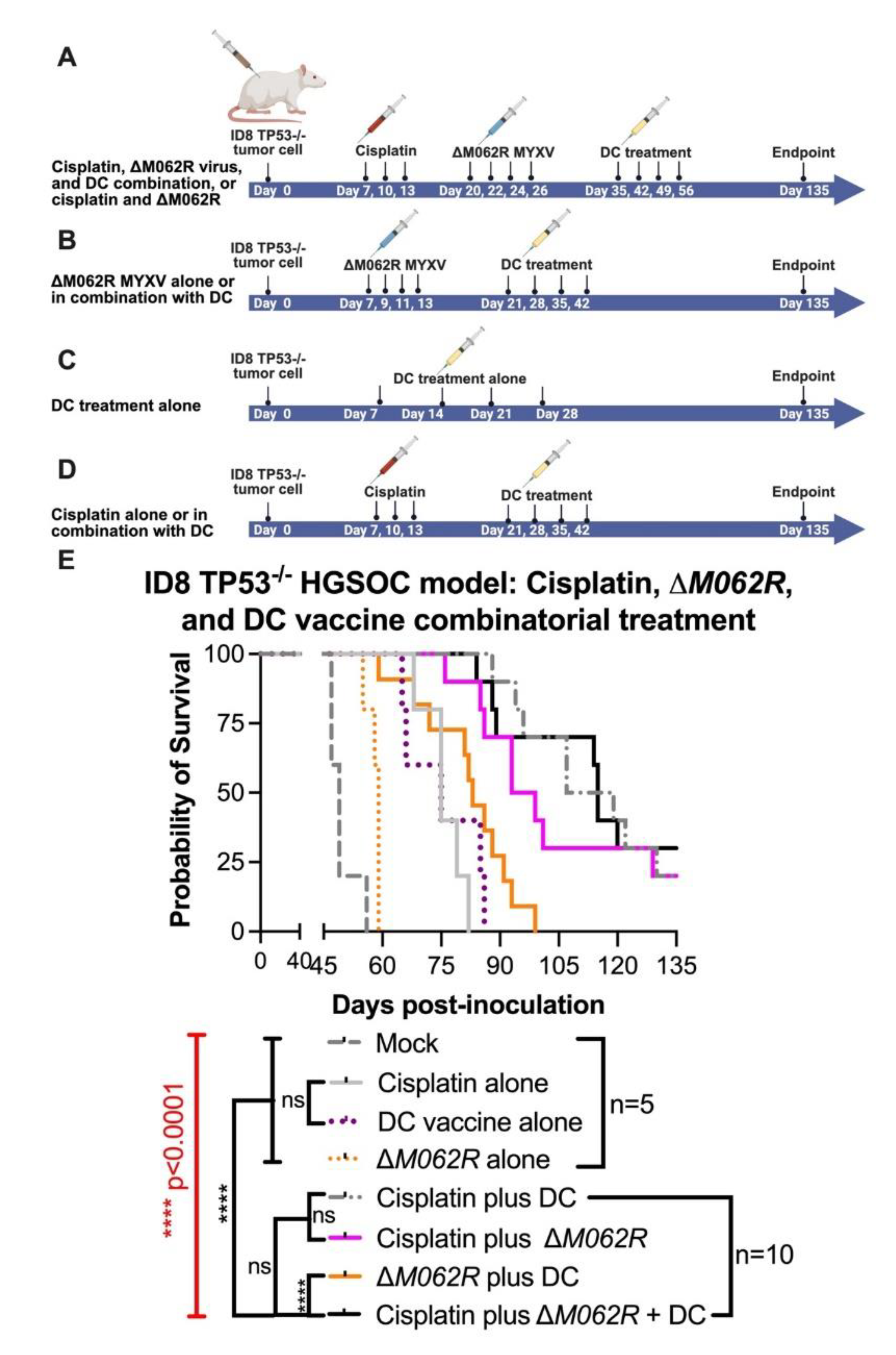
Figure 5.
Treatment with M062R provides survival advantage in HGSOC model with antigen targeted immunotherapy. A. Treatment design. C57/BL6 mice were injected i.p. with 1x106 ID8-Sp17 cells at day 0. Animals were randomly assigned to each treatment group with 4 mice per group. Cisplatin, in single or combinatorial treatment, was injected i.p. at 1 mg/Kg starting at 7 days post tumor establishment for a total of 3 treatments every 3 days. In the cisplatin-ΔM062R-DC or ΔM062R-DC combination treatments, ΔM062R-DC was administered 2 days before each DC vaccine. The ΔM062R-DC treatment block is repeated 4 times every 7 days. In treatments where ΔM062R or DC is not coincident, the ΔM062R or DC treatment was given on the set days shown in the diagram. B. Kaplan-Meier analysis of outcomes showed the therapeutic benefit of ΔM062R. The log-rank (Mantel-Cox) test was conducted with statistical significance defined as the following, * p<0.05, ** p<0.001, **** p<0.0001, and n.s.: not significant.
Figure 5.
Treatment with M062R provides survival advantage in HGSOC model with antigen targeted immunotherapy. A. Treatment design. C57/BL6 mice were injected i.p. with 1x106 ID8-Sp17 cells at day 0. Animals were randomly assigned to each treatment group with 4 mice per group. Cisplatin, in single or combinatorial treatment, was injected i.p. at 1 mg/Kg starting at 7 days post tumor establishment for a total of 3 treatments every 3 days. In the cisplatin-ΔM062R-DC or ΔM062R-DC combination treatments, ΔM062R-DC was administered 2 days before each DC vaccine. The ΔM062R-DC treatment block is repeated 4 times every 7 days. In treatments where ΔM062R or DC is not coincident, the ΔM062R or DC treatment was given on the set days shown in the diagram. B. Kaplan-Meier analysis of outcomes showed the therapeutic benefit of ΔM062R. The log-rank (Mantel-Cox) test was conducted with statistical significance defined as the following, * p<0.05, ** p<0.001, **** p<0.0001, and n.s.: not significant.
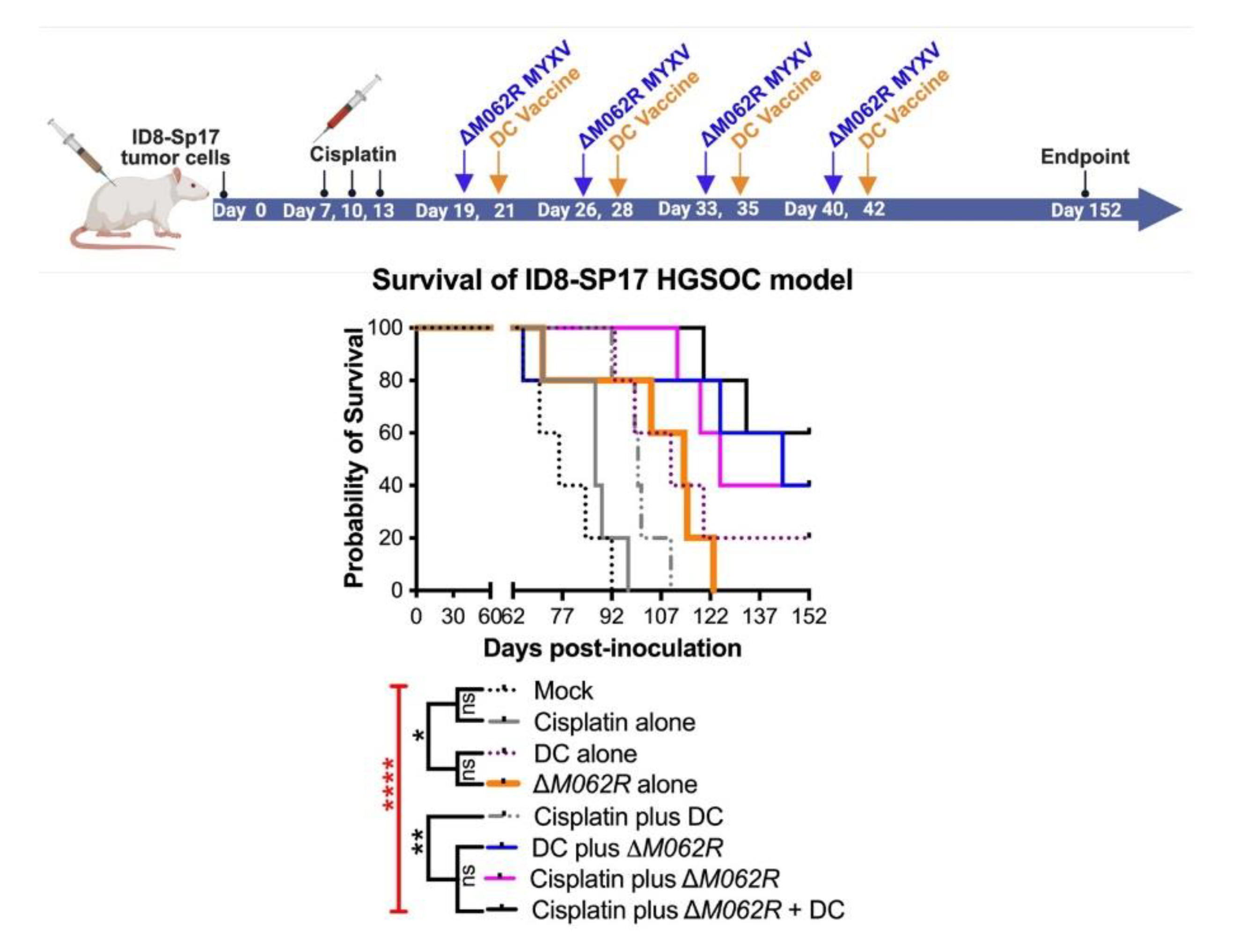
Figure 6.
MYXV treatments to OC CD14+ TAMs promote CD4+ T cell activation against tumor antigen. A co-culture system with TADG14v peptide tumor antigen-specific CD4+ T cells and OC ascites CD14+ TAMs was used for this study. A. MYXV treatments of CD14+ TAMs reversed the immunosuppressive effect by TAMs. After incubating without any TAMs (mock), with untreated TAMs, with WT MYXV treated TAMs, or with ΔM062R treated TAMs, TNFa expression by allogeneic (top row) or autologous (bottom row) CD4+ T cells was determined following overnight stimulation with tumor peptide antigen-loaded autologous lymphoblastoid cells. A representative result from 3 individual patient TAM samples is shown. B. Reversal of TAM immunosuppression by ΔM062R is independent of the cGAS/STING/TBK1/IRF3 signaling axis. CD4+ T cell functionality was measured as described above in A. A STING antagonist, H-151 (1 μM), and TBK1 kinase inhibitor, BX-795 (2 μM), were used to inhibit different steps of the DNA sensing pathway in cultures of T cells alone (first row), T cells plus CD14+ TAMs (second row), or T cells plus ΔM062R MYXV treated TAMs (third row).
Figure 6.
MYXV treatments to OC CD14+ TAMs promote CD4+ T cell activation against tumor antigen. A co-culture system with TADG14v peptide tumor antigen-specific CD4+ T cells and OC ascites CD14+ TAMs was used for this study. A. MYXV treatments of CD14+ TAMs reversed the immunosuppressive effect by TAMs. After incubating without any TAMs (mock), with untreated TAMs, with WT MYXV treated TAMs, or with ΔM062R treated TAMs, TNFa expression by allogeneic (top row) or autologous (bottom row) CD4+ T cells was determined following overnight stimulation with tumor peptide antigen-loaded autologous lymphoblastoid cells. A representative result from 3 individual patient TAM samples is shown. B. Reversal of TAM immunosuppression by ΔM062R is independent of the cGAS/STING/TBK1/IRF3 signaling axis. CD4+ T cell functionality was measured as described above in A. A STING antagonist, H-151 (1 μM), and TBK1 kinase inhibitor, BX-795 (2 μM), were used to inhibit different steps of the DNA sensing pathway in cultures of T cells alone (first row), T cells plus CD14+ TAMs (second row), or T cells plus ΔM062R MYXV treated TAMs (third row).
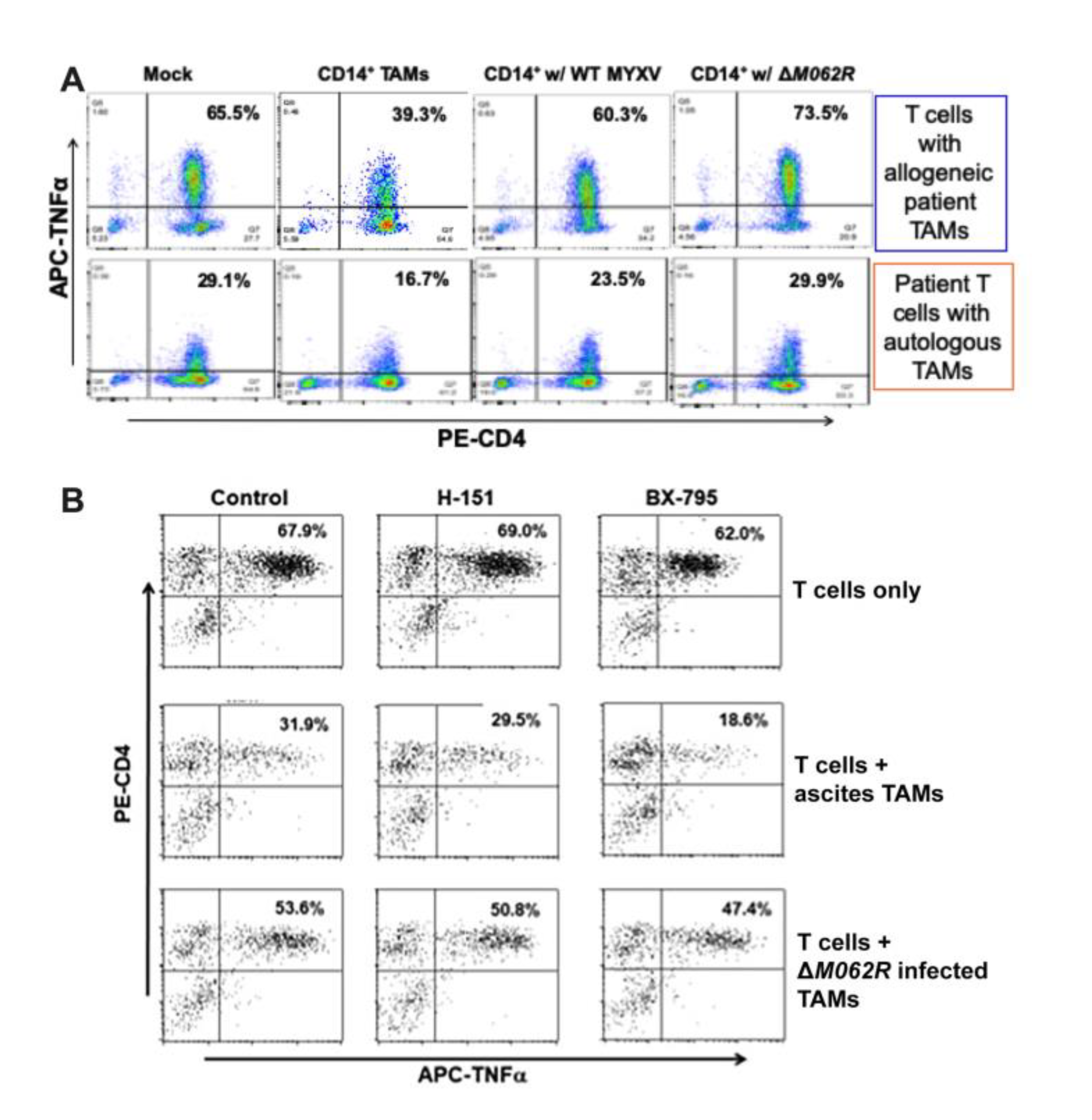

Table 1.
RT-PCR primers.
| Target Gene | Primer sequences |
|---|---|
|
Human IFNβ |
Fwd 5’- GCC ATC AGT CAC TTA AAC AGC -3’ |
| Rev 5’- GAA ACT GAA GAT CTC CTA GCC T -3’ | |
|
Human IL-12b |
Fwd 5’- CAAAGGAGGCGAGGTTCTAA-3’ |
| Rev 5’- GCAGGTGAAACGTCCAGAATA-3’ | |
|
Human Sp17 |
Fwd 5’- GGTTCCATAGGCAGTTCTTAC-3’ |
| Rev 5’- GGAAGGCAGCTTGGATTT-3’ | |
|
Human RSAD2 |
Fwd 5’- AGT GCA ACT ACA AAT GCG GC -3’ |
| Rev 5’- CTT GCC CAG GTA TTC TCC CC -3’ | |
|
Human CXCL-10 |
Fwd 5’- CTG TAC CTG CAT CAG CAT TAG TA -3’ |
| Rev 5’- GAC ATC TCT TCT CAC CCT TCT TT -3’ | |
|
Human IL15 |
Fwd 5’- AGCCAACTGGGTGAATGTAATA-3’ |
| Rev 5’- CATCTCCGGACTCAAGTGAAATA-3’ | |
|
Human ISG54 |
Fwd 5’- AGCGAAGGTGTGCTTTGAGA -3’ |
| Rev 5’- GAGGGTCAATGGCGTTCTGA -3’ | |
|
Human NFκB1A |
Fwd 5’- CCCTACACCTTGCCTGTGAG -3’ |
| Rev 5’- TGACATCAGCACCCAAGGAC -3’ | |
|
Human CCL3 |
Fwd 5’- CTCTCTGCAACCAGTTCTC-3’ |
| Rev 5’- CTGCTCGTCTCAAAGTAGTC-3’ | |
|
Murine Sp17 |
Fwd 5’- CTTTCTCCAACACCCACTAC-3’ |
| Rev 5’- CTTCATCTTCTTTACCTCTTCTCT-3’ | |
|
Murine IL-10 |
Fwd 5’- AGGCGCTGTCATCGATTTCT -3’ |
| Rev 5’- ATGGCCTTGTAGACACCTTGG-3’ | |
|
Murine CD40 |
Fwd 5’- GTAGGTCACCCCTGAGAACC-3’ |
| Rev 5’- ACAACCCGAACCATACACACAA-3’ | |
|
Murine CX3CL1 [29] |
Fwd 5’- GCTCCTAGCCCTGACCCATC-3’ |
| Rev 5’- AGCTGATAGCGGATGAGCAA-3’ | |
|
Murine GM-CSF |
Fwd 5’- CTGGCCCCATGTATAGCTGA-3’ |
| Rev 5’- ACAGTCCGTTTCCGGAGTTG-3’ | |
|
Murine IL-6 |
Fwd 5’- TCAATATTAGAGTCTCAACCCCCA-3’ |
| Rev 5’- GAAGGCGCTTGTGGAGAAGG-3’ | |
|
Murine iNOS [30] |
Fwd 5’- ATCGACCCGTCCACAGTATG-3’ |
| Rev 5’- GATGGACCCCAAGCAAGACT-3’ | |
|
Murine TNFα |
Fwd 5’-CCCTCACACTCACAAACCAC-3’ |
| Rev 5’- ACAAGGTACAACCCATCGGC-3’ | |
|
Murine IFNβ |
Fwd 5’- AGATCTCTGCTCGGACCACC-3’ |
| Rev 5’- CGTGGGAGATGTCCTCAACT-3’ | |
|
Murine CXCL10 |
Fwd 5’- ATGACGGGCCAGTGAGAATG -3’ |
| Rev 5’- TCGTGGCAATGATCTCAACAC-3’ | |
|
Murine IRF3.2 |
Fwd 5’- CACTCCCCACGCTACACTC-3’ |
| Rev 5’- TCCCATCCCCAGTAGCATGAG-3’ | |
|
Murine IRF7.2 [31] |
Fwd 5’- TGCTGTTTGGAGACTGGCTAT-3’ |
| Rev 5’- TCCAAGCTCCCGGCTAAGT-3’ | |
|
Murine IFNγ |
Fwd 5’- CGGCACAGTCATTGAAAGCC-3’ |
| Rev 5’- TGTCACCATCCTTTTGCCAGT-3’ | |
|
Murine CXCL1 [29] |
Fwd 5’- GCTGGGATTCACCTCAAGAA-3’ |
| Rev 5’- TCTCCGTTACTTGGGGACAC-3’ | |
|
Murine CXCL3 [32] |
Fwd 5’- CCACTCTCAAGGATGGTCAA -3’ |
| Rev 5’- GGATGGATCGCTTTTCTCTG -3’ | |
|
Murine MCP-1 [30] |
Fwd 5’- AGGTCCCTGTCATGCTTCTG-3’ |
| Rev 5’- GGATCATCTTGCTGGTGAAT-3’ | |
|
Murine EGR1 |
Fwd 5’- CACCTGACCGCAGAGTCTTTT-3’ |
| Rev 5’- GCGGCCAGTATAGGTGATGG-3’ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



