Submitted:
29 May 2025
Posted:
30 May 2025
You are already at the latest version
Abstract
Seafood (fish, crustacean, and mollusk) allergy represents a critical global health issue. Food processing offer a viable strategy for allergenicity mitigation, serves as a critical intervention for seafood allergy prevention. This paper reviews recent advances in seafood allergen research, with particular focus on molecular properties, epitopes, and structure-allergenicity relationships, which are foundations for designing processing technologies to mitigate allergenicity. Furthermore, an analysis of how various food processing techniques modulate allergen structures and epitopes, ultimately affecting their allergenicity was conducted. Current World Health Organization (WHO)/ International Union of Immunological Societies (IUIS) listings include 44 fish allergens and 60 shellfish allergens, with their characterization enabling targeted processing approaches for allergenicity elimination. The physical processing technique, including thermal and non-thermal treatment, can dramatically influence the conformational and linear epitopes by altering or destroying the structure of an allergen. The chemistry-based processing techniques (enzymatic-catalyzed cross-linking and glycation), which induce covalent/non-covalent interactions between allergens and various modifiers, can effectively masking epitopes through molecular complexation. Biological processing attenuates allergenicity by induces protein unfolding, polypeptide chain uncoiling, and enzymatic degradation. Nevertheless, the structure-activity relationship of seafood allergens remains insufficiently elucidated, despite its critical role in guiding processing technologies for allergenicity elimination and elucidating the fundamental mechanisms involved.

Keywords:
allergenic protein
; elimination processing
; epitopes
; protein structure
; seafood allergy
1. Introduction
Food allergy is an adverse reaction induced by ingestion of specific food and food products, the immune system recognizes certain food proteins as an allergen and generates an immune response that causes various allergies [1]. Food allergy affects nearly 10% of adults and 8% of children, and the prevalence has increased worldwide in recent decades [2]. More than 160 foods are known to cause food allergies, while a rather short list of foods account for most of the more serious disease burden, namely peanut, tree nuts, wheat, sesame, seeds, fish, shellfish (crustacean and mollusk), egg, and milk, among which fish and shellfish belong to seafood [3].
Seafoods, also called “blue foods”, is a general term for all edible aquatic organisms, of which fish and shellfish are mostly consumed (Figure 1) [4], they are healthy delicacies with high protein content, healthy fats, vitamins, and minerals [5]. However, seafood allergy becomes a serious global public health problem with increasing prevalence due to fast increased consumption [6]. A recent study updated prevalence estimates for food allergy in 2012-2021 in Europe showed the overall pooled estimates for fish and shellfish allergy of self-reported lifetime prevalence were 1.4% and 1.0%, respectively [7]. In certain parts of Europe shellfish has become the leading food allergen [6]. In the USA, shellfish and fin fish ranked the most common convincing allergenic foods, with an estimate of up to 2.9% and 0.9%, respectively. The prevalence rate of shellfish was even higher than that of peanut and tree nut among adults [8].
Seafood allergy is of particular importance in the Asian region, with up to 7.7% prevalence in some Asian countries [9]. IgE sensitization to shellfish was as high as 10.6% among Singaporean children, up to 13.1% of the Chinese and 10.3% of Indian populations have shrimp specific IgE that exceeded 0.70 kUA/L [10]. Shellfish is also the leading cause of food allergy in China, the shellfish prevalence is 7.3% among Taiwanese, and over 2.0% of the Hong Kong population was positive to shrimp by skin prick test [6,10]. The epidemiological survey of self-reported food allergy performed by Feng et al. showed that shrimp was the main allergenic food among university students, followed by shellfish [11]. Fish allergy is less common in Asia than shellfish allergy, but its prevalence is still considerably high, affected ~1.6% of Vietnamese and ~4.1% of Singaporean [12].
Unlike most other food allergies, seafood allergy is thought to persist for life in up to 90% of patients [13,14]. The avoidance of seafood-containing foods is the most effective measure to prevent seafood allergies, while this might result in various nutritional deficiency, there is an urgent to seek effective processing to reduce seafood allergenicity. The presence of allergens contributes to seafood allergy, therefore, processing methods have been explored aiming at modifying the allergens. In this review, we summarized current knowledge of seafood allergens and various processing methods to decrease allergenicity. It is conceivable that the information will provide rationales for novel strategies to prevent and manage allergy to seafood.
2. Seafood Allergens and Their Molecular Properties
During the past decades, several allergenic proteins and related isoforms have been identified in commonly consumed seafood species and record in the World Health Organization and International Union of Immunological Societies (WHO/IUIS) (http://www.allergen.org). This section provides an overview on the currently identified fish and shellfish allergens.
2.1. Fish Allergens
Up to now, 44 allergens from 22 species of fish, including 13 kinds of proteins, have been provided recognizable names by the WHO/IUIS, it can be found that an allergy to fish fillet is most prevalent, but allergic reactions have also been reported to fish roe, fish gelatin, and fish blood (Table 1). Parvalbumin is the primary fish fillet allergen, followed by the β-enolase and aldolase A, in recent years, tropomyosin and triosephosphate isomerase have been identified as novel fish allergens [15,16]. In addition, several minor fish allergens—such as creatine kinase, pyruvate kinase, lactate dehydrogenase, and glucose-6-phosphate isomerase—were also included in the WHO/IUIS list, the characterization of their biochemical properties from a food allergen perspective were also summarized (Table 2).
The most characterized allergenic fish parvalbumins were demonstrated to be responsible for over 70% of allergic reactions to fish and fish products [17]. Most individuals with fish allergies exhibit allergic reactions to multiple fish species due to the high cross-reactivity of parvalbumins, which share 88–100% sequence similarity with conserved regions [18]. Parvalbumin is an extremely thermostable allergen in fish muscles with a molecular weight of 10~13 kDa, it could be divided into α-type and β-type, the allergenicity of fish α-parvalbumin is generally considered to be lower than β-parvalbumin [19]. Parvalbumin is a member of the EF hand-containing Ca2+-binding protein superfamily, fish parvalbumins display the conserved spatial structure which consist of 6 α-helices, organized by the AB domain at N-terminal and CD-EF domain at C-terminal (Figure 2a) [20].Two Ca2+-binding sites in CD and EF domains can be found in the structure of parvalbumin, which are involved in its structural preservation. IgE-binding epitopes of various fish parvalbumins exist in a stereoscopic conformation maintained by Ca2+ binding, hence, depletion of Ca2+ leads to decreased IgE-binding capacity [19]. The epitopes of fish parvalbumins have been well documented, most of the IgE-binding epitopes located on the loop regions between α-helices [19,21]. Perez-Cordo et al. found that the amino acid (AA) 95~108 was recognized by all the fish allergic patients and can served as a severity marker of fish allergy [21], in a recent study by Huang et al. this region was also identified as an important IgE-binding epitope in β-parvalbumin [19]. Noteworthy, the three-dimensional structure showed this IgE-binding epitope located on the Ca2+-binding domain (Figure 2a, colored red).
Enolase is a glycolytic enzyme that catalyzes the reversible dehydration of 2-phosphoglycerate to phosphoenolpyruvate. In vertebrates, three enolase isotypes (α, β, and γ) have been identified, among which the β-isotype has been recognized as a fish allergen [22]. Kuehn et al. first characterized the thermolabile β-enolase as a novel allergen in 3 fish species, Gadus morhua, Thunnus albacares, and Salmo salar, demonstrating its reactivity in over 50% of fish-allergic patients. [23]. Meanwhile, in a recent study by Wai et al., IgE reactivity to β-enolase was observed in 17.8% of tested subjects [24]. While there is increasing evidence that β-enolase may play an important role in allergic disease, for it can sensitize individuals orally or via the respiratory tract, and even via skin, and the sequence conservation support its cross-reactivity [22]. β-Enolase form homodimers in muscle, and each monomer consists of 2 domains, the N-terminal domain folding into a two-layer sandwich composed of an anti-parallel β-sheet and an α-helical bundle, and a C-terminal domain folds into an α/β-barrel (Figure 2b). In addition, Mg2+ can be found in the α/β-barrel in each monomer [25]. However, the epitope of β-enolase remains to be characterized.
Aldolase A, another essential glycolytic enzyme with a molecular weight of 40 kDa, was first identified as an allergen in Pacific salmon in 2009 [26]. There are now 4 aldolase A registered as fish allergens in WHO/IUIS, like enolase, it is labile to thermal treatment, which can only be detected in raw fish extracts [23,27]. The structures of fish aldolase are yet to be determined, according to structures from other species, aldolase A seemed to be present as an oligomeric protein, each monomer folds into an α/β-barrel with 10 anti-parallel β-sheets bundled by 13 α-helices (Figure 2c) [28]. Although aldolase A has been shown to be heat-labile, its structure-allergenicity relationship remains unclear due to insufficient structural characterization and epitope mapping
Vertebrate tropomyosin has been reported to be nonallergenic protein until recent studies to challenge this belief. The tropomyosin from Oreochromis mossambicus was the first vertebrate tropomyosin been identified as an allergen by immunological analyses [29]. In 2020, Ruethers et al. expanded the allergen repertoire of Salmo salar and Pangasianodon hypophthalmus, including a tropomyosin from S. salar and 2 tropomyosin isoforms from P. hypophthalmus, thus these tropomyosins were registered in the WHO/IUIS [16]. In the recent research by Li et al., tropomyosin was also confirmed as an important heat-stable allergen in Lateolabrax japonicus [30]. Tropomyosin has a molecular mass of around 35~38 kDa, it belongs to the actin-binding protein family, with a conserved spatial structure that composed of 2 identically coiled subunits. Each of the tropomyosin subunit follows a repetitive “ABCDEF” heptamer pattern which contributes to the structure stability of the subunit (Figure 2d) [31,32]. The exceptionally stable α-helical coiled-coil secondary structure makes tropomyosin resist to food processing and retain high allergenic potential ability after heating and exposure to acidic conditions [16]. While Xu et al. revealed distinct digestive patterns between fish and shellfish tropomyosin, their study demonstrated that shrimp TM digestion products contained significantly more peptides matching known T/B cell epitopes compared to fish tropomyosin. This finding provides a plausible explanation for why tropomyosin serves as a major allergen in shellfish but only a minor allergen in fish species [33].
The α-Collagen was identified as a fish allergen in Japan in the early 2000s [34]. At present, α-collagens from Lates calcarifer and S. salar were registered at WHO/IUIS as Lat c 6 and Sal s 6, respectively, it has been proved to be a major fish allergen that could induce cross-reactivity by subsequent research [35]. Though limit cases of collagen-induced fish allergy have been reported in the past decades, Shimojo et al. found IgE reactivity to fish collagen was associated with symptom severity [36]. Different genes code for distinct collagen chains but all collagens share conserved structure called collagen triple helix which formed by association of 3 identical or different polypeptide chains, and each chain displays a conspicuous -Gly-X-Y- repetitive sequence (Figure 2e) [37,38]. Similar to tropomyosin, the triple-helical structure of collagen confers remarkable thermal stability. However, when subjected to heat treatment, collagen undergoes partial denaturation and becomes water-soluble, thereby retaining its potential to induce anaphylactic reactions [39]. Due to both the underreporting of collagen allergy cases and insufficient structural characterization, comprehensive analysis of the structure-allergenicity relationship in collagen remains limited.
In addition to allergens mentioned above, other minor allergens have been identified in fish, most of which are enzymes. The cytosolic protein pyruvate kinase was first noticed in a case which a child appeared allergic symptom after ingesting swordfish, pyruvate kinase was recognized by the IgE in patient serum, together with enolase, aldolase, and triosephosphate isomerase [40]. Triosephosphate isomerase, a key enzyme involved in glycolysis, is a known allergen from crustaceans and mollusks [41], has also been recognized as fish allergen in the research by Ruethers et al. [16]. The conserved structure that primarily composed of a (β/α)8-barrel motif prototype was considered contribute for its cross-reactivity [41]. The first case of allergy to creatine kinase was reported by Larco-Rojas et al. [42], the allergenicity and cross-reactivity of creatine kinase has subsequently been confirmed by Ruethers et al. and Yang et al [16,43]. Besides, lactate dehydrogenase, glucose-6-phosphate isomerase, glyceraldehyde-3-phosphate dehydrogenase were identified as fish allergens, allergen can also be found in fish roe, including lipovitellin and β’-component, which are sub-fragments of vitellogenin [16,44]. While the detailed information on their molecular properties and prevalence rate are to be further explored.
2.2. Shellfish Allergens
Hoffman et al. identified the IgE-binding tropomyosin in Penaeus aztecus for the first time in 1981 [45], then tropomyosin has been identified in various of species and subsequently considered to be the major allergen in shellfish that responsible for cross-reactivity of many invertebrate species [46,47]. As showed in Table 1~3, it is an allergen found in all 3 types of seafood of fish, crustaceans, and mollusks. The α-helical coiled-coil dimeric structure is highly conserved across invertebrates and vertebrates, while vertebrate tropomyosin shows closer helix-helix packing alanine clusters in which more enzymatic cleavage sites are accessible, leading to significantly weaker resistance to simulated gastric digestion than invertebrate tropomyosin [48]. Liu et al. found 11 heat/digested stable linear epitopes of tropomyosin, which are considered as the primary cause of allergenicity (Table 2, Figure 3a) [49]. In the original research on tropomyosin, linear epitopes were thought to be priority in tropomyosin because of its structural feature, while some recent studies revealed the existence and importance of its conformational epitopes [50,51]. Despite the limited structural data on conformational epitopes, current evidence suggests that both linear and conformational epitopes of tropomyosin appear to be evenly distributed across the protein.
Since Yu et al. identified arginine kinase as a novel shrimp allergen, it subsequently recognized as allergens in many other kinds of shellfish [17,52]. Until now, arginine kinase from 7 crustaceans and 1 mollusk were officially recognized by the WHO/IUIS as food allergens and have been proved to be responsible for cross-reaction among invertebrates [53] (Table 3). Arginine kinase and creatine kinase are members from phosphagen kinase family with the conserved spatial structure of a canonical fold composed of an α-helical N-terminal domain and an α-β C-terminal domain (Figure 3b) [53]. This structure is easily disrupted by the breakage of intramolecular disulfide bond between Cys201 and Cys271 [54]. Besides, arginine kinase is a thermal-sensitive protein which forms multimer in the temperature higher than 46 ℃ [55]. In arginine kinase, the conformational IgE epitopes are more predominant, gives reason for the decreasing IgE-binding activity of denatured arginine kinase [54,56]. While heat/digested stable linear epitopes were also found in arginine kinase, which are crucial for the capacity of inducing anaphylaxis (Table 2) [49].
Paramyosin has initially been identified as a major respiratory allergen, in the past decade, paramyosin has been identified as allergen in mollusks, Yu et al. identified paramyosin as a novel allergen in Rapana venosa which has been recognized by WHO/IUIS as Rap v 2 [57] (Table 3), and this allergen was proved to be a risk factor of mollusc-mite cross-reactivity [58]. Paramyosin belongs to the acting-binding protein family which sometimes referred to as “water-insoluble tropomyosin” or “tropomyosin-A” because of its conserved α-helical secondary structure. While it was predicted that the α-helical secondary structure folds into a globular spatial structure, which is different from the long chain structure of tropomyosin [59]. Besides, paramyosin is not so stable in heating treatment as tropomyosin, probably results from the structural property (Table 2). The Ca2+-binding sites also exist in the sequence of paramyosin (Table 2), while the structure-allergenicity relationship remains to explore.
Myosins are important functional muscle proteins of acting binding family, they are multimeric proteins, with each subunit composed of a heavy chain with a molecular mass of ~200 kDa and 2 light chains with a molecular mass of ~18-20 kDa (Table 2, Figure 3c) [60]. The heavy chain composed of a globular motor domain and a coiled-coil structure tail (Figure 3c, colored light grey); two light chains wrap around the neck region of each myosin heavy chain(Figure 3c, with myosin light chain 1 and 2 colored light blue and orange, respectively). Myosin heavy chain has been recognized as an allergen of fish and shrimp, which involved in the cross-reactivity between crustacean and mite [61,62,63]. Moreover, both the myosin light chain 1 and myosin light chain 2 have been reported as food allergens. Ayuso et al. identified myosin light chain 2 as a major allergen of Litopenaeus vannamei (Lit v 3) that recognized by 55% of shrimp allergic subjects and obtained a systematic name in WHO/IUIS [64] (Table 2). Subsequently, myosin light chain 2 from Homarus americanus (Hom a 3) and Penaeus monodon (Pen m 3) have also been recognized as allergens [65]. Besides, myosin light chain 1 from Artemia franciscana (Art f 5), Crangon crangon (Cra c 5), Scylla paramamosain (Scy p 3), and Procambarus clarkii (Pro c 5) have also been identified as allergens in crustaceans [66,67,68]. The myosin light chains are mainly composed of α-helixes and random coils and show canonical lobe with an N-terminal domain and a C-terminal domain (Figure 3c), the epitopes distribute in the α-helixes with highly conserved sequence [67,68]. Though Ca2+-binding sites can be found in the structures of myosin light chains, the influence of bound Ca2+ in it allergenicity remains unrevealed. Myosin light chain 1 was found to resistant to thermal treatment and extreme pH [69], the digestion product contains 5 IgE-reactive epitopes (Table 2, Figure 3c, colored red) [49], while information on the physicochemical properties of myosin light chain 2 is lacking.
Sarcoplasmic calcium binding protein was initially identified as a minor allergen in saltwater shrimp [70]. In recent years, the allergenicity and cross-reactivity of sarcoplasmic calcium binding protein has further been demonstrated at the molecular level in other crustaceans and mollusks (Table 2, 3) [71,72]. Sarcoplasmic calcium binding protein belongs to the calcium binding family, there are 4 EF-hand motifs in its structure, 2 or 3 of which individually contain a Ca2+-binding site corresponding to the loop region of 12 amino acid residues (Figure 3d) [73]. Sarcoplasmic calcium binding protein is a heat-resistant allergen, Liu et al. identified 4 heat/digested peptides (AA 39-51, 77-82, 111-120, 139-149) that crucial for its allergenicity (Table 2, Figure 3d,colored red) [49], noteworthy, two of these peptides located in the Ca2+-binding sites (Figure 3d). Additionally, several linear and conformational epitopes also locate in the Ca2+-binding sites [71,73]. Despite of the heat-resistance of sarcoplasmic calcium binding protein, depletion of its Ca2+ leads to obvious conformation variation, which consequently reduce the immunoreactivity [71]. It can be speculated that the Ca2+ depletion might influent IgE-reactivity by either changing the accessible surface of IgE recognition region or disrupting the structure for conformational epitope formation.
Triosephosphate isomerase in C. crangon (Cra c 8) was the first identified allergenic crustacean triosephosphate isomerase, having approximately 20% sensitization frequency [66]. The three-dimensional structure of triosephosphate isomerase is primarily composed of a (β/α)8-barrel motif prototype formed by 8 central β-strands surrounded by 8 α-helices joined by loops [41]. This structure is easily disrupted by heat treatment, accompanied with reduction of IgE-binding activity [74]. Four heat/digested peptides were found to distribute in either surface α-helices or intramolecular β-strands (Table 2, Figure 3e,colored red), meanwhile 2 conformational epitope regions located on the protruding surface areas [41,49,75]. Filamin C first got notice for its cross-reactivity with triosephosphate isomerase in P. clarkii [74], and now it has also been identified as an allergen in S. paramamosain [76]. Different from other acting-binding family members, filamin C found to thermal-sensitive and decreasing IgE-binding capacity can be observed in extreme pH [74]. The digested product of filamin C retains 50% IgE-binding activity [74], indicating the existence of digested resistant peptides. The overall spatial structure of filamin C is not available at present, nevertheless, He et al. determined the crystal structure of its allergic predominant region (AA 336-531), which displays a barrel structure composed of 16 β-strands and 2 α-helices (Figure 3f), a conformational epitope and 6 linear epitopes were found to locate in this region [77].
Other shellfish allergens are also identified, such as troponin C/I, hemocyanin, fatty-acid-binding protein, ovary development-related protein, and glycogen phosphorylase-like protein (Table 3). Troponin is composed of 3 subunits, suffixed C, I and T, with troponin C and I being registered as crustacean allergens [78]. Like other members in acting-binding protein family, troponin C is mainly composed of α-helixes and random coils, besides, EF-hand motifs which contain functional Ca2+-binding loops can be found in the structure (Figure 3g) [79]. While there is no evidence of the influence on its allergenicity of Ca2+ binding. Allergenic hemocyanin can be found in shrimp cephalothorax or crab roe, it has been recommended as a tool for shrimp and crab allergy diagnostics that distinguished from allergic to muscle [80,81]. Hemocyanin is also an inhaled allergen, which is reported to inducing cross-reactivity between crustaceans and insects [80,82]. Ovary development-related protein has been registered in WHO/IUIS as a crab allergen (Eri s 2), however, neither physicochemical and digestion properties nor epitope and structural information was reported further [83]. Recently, Múnera et al. identified the 15 kDa fatty-acid-binding protein as a novel shrimp allergen and named as Lit v 13. Fatty-acid-binding protein, together with the house dust mite allergen Der p 13, are belong to the lipid binding protein superfamily, cross-reactivity can be observed between Lit v 13 and Der p 13, with the highly conserved AA 54-72 as the most involved region in the cross-reactivity [84]. Besides, Wai et al., identified glycogen phosphorylase-like protein in P. monodon (Pen m 14) as a novel allergen in addition to the fatty-acid-binding protein (Pen m 13) [85].
The above consolidates the documented seafood allergens to date, provides an analysis of their molecular structures, immunodominant epitopes, and key physicochemical properties. Analysis demonstrates that the majority of characterized seafood allergens are functionally restricted to three principal protein categories: calcium-binding proteins, actin-binding proteins, and enzymatic proteins, with the first two categories exhibit superior heat resistance (Table 2). Notably, allergens within each category exhibit similar physicochemical properties. Allergenic calcium-binding proteins, including parvalbumins and sarcoplasmic calcium binding protein, preserve evolutionarily conserved EF-hand domains that form the structural basis for Ca²⁺ chelation and likely contribute to IgE-reactivity. Actin-binding proteins, particularly tropomyosin and paramyosin, maintain highly conserved coiled-coil α-helical domains. These structurally stable motifs, characterized by their 7-residue (heptad) repeat pattern (Figure 2), confer remarkable thermal stability, enabling these allergens to maintain their immunoreactivity even after extensive heat processing. Enzymatic protein allergens, including enolase, aldolase, and arginine kinase, demonstrate marked thermolability, typically undergoing structural denaturation and loss of IgE-binding capacity during thermal treatment. The allergen category presented in this study guides food processors in matching treatment methods to specific protein characteristics. Moreover, mapped epitope data supports the design of processing strategies that specifically modify immunodominant regions
3. Effect of Processing Techniques on Seafood Allergens
Though avoidance of seafood and seafood products is the basic and common measure in managing seafood allergies, a long-term strict seafood-free diet might lead to a diet imbalance. Therefore, the food processing techniques that reduce or eliminate the allergenicity of seafood attracted research studies. There are many reports about the processing basing on physics, chemistry, and biology technique, or the combination of different techniques. In general, processing may destroy existing epitopes on an allergen or may generate new ones because of changes involving protein conformation. In some circumstance, the structure changing of an allergen during processing accompanied with more sites that is accessible to proteolytic enzymes, which producing hypoallergenic hydrolysates unable to elicit immune responses. This section reviews current knowledge of how food processing modifies the allergenic potential of seafood.
3.1. Processing Techniques Based on Physics
The physical processing technique including thermal treatment (e. g. boiling, steaming, baking, frying, microwave) and non-thermal treatment (e. g. high-pressure processing, ultrasound, irradiation, cold plasma). For seafood processing, traditional heating methods (boiling, steaming, frying, etc.) and ultrasound are commonly applied treatments.
3.1.1. Thermal Treatment
The effect of thermal processing on food allergens is of major interest for allergic patients who need to avoid all food containing active allergens. Thermal treatments of food proteins induce modifications including hydrolysis of peptide bonds, aggregation by disulfide and non-covalent bonds, denaturation, and reactions with other food components, such as carbohydrates and lipids [86]. For seafood allergens, the degradation, molecular aggregation, and denaturation of proteins are the primary factors altering allergenicity (Figure 4). The effect of thermal processing on fish allergenicity has been shown to be species and allergens dependent. Fish allergens such as aldolase, enolase, and creatine kinase have been described to be thermolabile (Figure 4a). Enolases from Gadus morhua and Thunnus albacore lost the IgE binding activity after heating at 90 °C for 1 minute, this treatment condition can sharply reduce the IgE-binding activity of aldolases at the same time [23]. In addition, creatine kinase from Thunnus tonggol was found to be undetected after boiled for 15 minutes [87]. Kubota et al. focused on the allergenicity of Pacific mackerel parvalbumin influenced by heat treatment, it was found that more than 50% reduction of allergenicity by heating at 80 °C for 10 minutes and a complete loss of IgE reactivity of parvalbumin heated at 140 °C [88]. By detecting the binding activity of heated parvalbumin to the monoclonal antibody PARV-19, which recognizes parvalbumin in a Ca2+ dependent manner, the authors revealed the mechanism of allergenicity decreasing as conformational changes owing to Ca2+ depletion with the heat [88]. A recent investigation by Liang et al. also demonstrated that parvalbumin from Scombriformes, Scorpaeniformes and Tetraodontiformes species showed weak or no immunoreactivity, because of losing the Ca2+ dependent epitope [89]. While in some species such as Cyprinus carpio and the order of Perciformes, parvalbumin showed increased immunoreactivity to antibodies, due to an enrichment of parvalbumin upon aggregation during heat treatment [90].
Heat treatment appears to exert a low impact on crustacean allergens because most of them are thermostable, except for triosephosphate isomerase, filamin C, and arginine kinase. The IgE-binding activity of triosephosphate isomerase was reduced with increasing temperature higher than 60 °C, accompanied with an irreversible denatured structure [74] (Figure 4b). Similar to triosephosphate isomerase, heat treatment at temperature higher than 60 °C resulted in structure disruption with a decreasing of IgE-binding activity of filamin C [74] (Figure 4b). Arginine kinase is also a thermal sensitive allergen which aggregates in a temperature higher than 44 °C, while increased IgE-binding activity of AK can be observed in 44-70 °C, continuous heat up lead to gradually IgE-binding activity decreasing and ultimately eliminated when temperatures were 80 °C or higher [55]. Arginine kinase contains free sulfhydryl in the molecule, it can be speculated that the free sulfhydryl participates in aggregation by disulfide, and arginine kinase are enriched upon aggregation, thus increased the IgE-binding activity (Figure 4c).
It is worthy to note that all the studies came to an understanding that crustacean tropomyosin retains a great allergenicity even after heat treatment. For instance, in the study by Chen et al., tropomyosin from 8 randomly selected fish species showed the most stable existence and immunoreactivity among all the muscle proteins [91]. Tropomyosin from Crassostrea gigas showed a significantly higher IgE reactivity after roasting and boiling processing, with roasted protein having slightly more IgE reactivity than the boiled one [92].The author then explored the mechanism from the aspect of structure and found that the two-stranded α-helical coiled-coil structure of tropomyosin was destroyed and gradually converted to a single-stranded α-helix, which leaded to an altering hydrophobic face that might contain IgE-binding epitopes (Figure 4d). Moreover, the IgE-binding of tropomyosin from the extracts of different processed shrimp, including boiling, steaming, baking, frying, and microwave was also higher than that of the raw sample [93]. While another thermostable allergen sarcoplasmic calcium binding protein showed virtually invisible immunoblot band intensity after heat treatment by the same methods, despite of the detectable band in SDS-PAGE [94]. It seemed that the IgE-binding activity of sarcoplasmic calcium binding protein can be variety in different thermal treatment condition. Sarcoplasmic calcium binding protein retained immunological binding capacity in the water bath at 30-100 °C despite of the structure changing and polymer formation [72]. Besides, myosin light chain 2 has also shown high resistance to boiling [64], while further condition optimization and the in-depth mechanism are to be fulfilled.
Microwave heating is a novel thermal treatment in which the interaction of microwaves and the medium by volumetric dissipation of electromagnetic energy in form of heat. Microwaves travel through the lossy medium producing an increase of the medium temperature [95]. Thus, microwaves are able to affect the kinetics of conformational changes of allergens and to accelerate their denaturation. Despite some attempts have been done in using microwaves to reduce the allergenicity of food [93], techniques that combine microwave heating with other processing method is necessary for eliminating the antigenicity of proteins.
3.1.2. Non-Thermal Treatment
Thermal treatment was found to be inefficient for altering the IgE-binding ability of thermostable allergens, and even has been shown to markedly increase the IgE reactivity of tropomyosin. For minimizing the negative effects of the thermal processing techniques, innovative non-thermal techniques are sought to reduce or eliminate the allergenicity of seafood. High-pressure processing involves pressure 100 MPa to 1000 MPa for a shorter duration of time which gives rise to structural distortion on a protein by changing of the non-covalent bonds [96] (Figure 5a). In the study by Jin et al. [97], tropomyosin from Todarodes pacificus was treated with high hydrostatic pressure of 200, 400 and 600 MPa at 20 °C, the structure of tropomyosin denatured by a conversion of α-helix to β-sheet and random coils and a decreasing of free sulfhydryl group. This structure distortion resulted in a decreasing of digestibility of tropomyosin and thus, reduced its allergenicity [97]. Electrophoresis analysis revealed that high-pressure processing decreased the SDS-PAGE band intensity of fish allergens including enolase, creatine kinase, aldolase, and triosephosphate isomerase when the pressure reached 430 MPa, while whether IgE-binding fragment exists has not been expounded [98]. Noteworthy, intensity of parvalbumin was found to increase at this pressure. Recently, the application of high hydrostatic pressure treatment of 200-400 MPa was found to successfully change the structure of parvalbumin, suggested the potential application of high hydrostatic pressure treatment in reducing the allergenicity of parvalbumin [99].
High-intensity ultrasound uses high-energy mechanical waves (20-100 kHz), which can cause the breaking of chemical bonds and the occurrence of the redox reaction of free radical, leading to covalent bonds breaking and molecular degradation [100] (Figure 5b). By treating with high-intensity ultrasound (20 kHz, 100-800 w) for 15 minutes, the allergenicity of Exopalaemon modestus tropomyosin was significantly reduced [101]. It was demonstrated that the ultrasound influenced the primary, secondary, and tertiary structure of tropomyosin: in the amino acid level, ultrasound treatment oxidized the cystine, methionine and lysine in tropomyosin and degraded it to generate protein fragments; in the secondary structure, a conversion of α-helix to β-sheet, β-turn and random coil occurred, resulted in a loosen tertiary structure [101] (Figure 5b). Li et al. reported that the ultrasound (30 kHz, 800 W, 1.5 h) at 0 °C and 50 °C slightly reduced the allergenicity raw shrimps, while allergenicity reduction by ultrasound treatment to boiled shrimp samples was more effective than raw samples [102]. It is interesting to note that the most reduced allergenicity for tropomyosin was found in the samples treated by ultrasound (30 kHz, 800 W, 1.5 h) at 0 °C [102]. It seemed that there were several allergens in the muscle extract that participate in the allergenicity, indicating that it is more comprehensive to optimize processing condition for extract than a purified allergen.
Irradiation has been regarded as a cold pasteurization practice for preserving food with minimal alteration in nutritive and sensorial characteristics of foods [103]. During irradiation, the energy from γ-rays, X-rays, or electron beams is directly absorbed, leading to changes in the spatial structure and conformation of the allergenic proteins, thereby reducing their allergenicity [104]. It was found that under the condition of irradiated with varying doses (1-13 kGy) by electron beam irradiation at 10 MeV, IgE-binding activity of turbot parvalbumin decreased markedly upon increasing the irradiation dose [105]. The author speculated that the decreasing IgE-binding activity was resulted from the carbonylation of amino acids in epitopes, according to the observation that protein carbonyls contents have positive relation with IgE binding capacity reduction (Figure 5c). The electron beam irradiation treatment is also efficiency in eliminating allergenicity of shrimp tropomyosin. Guan et al. demonstrated that the irradiation treatment broken the hydrogen bonds in tropomyosin, which brought a more extended structure of protein accompanied by epitope disruption, thus reduced the binding activity of tropomyosin with specific antibodies [106]. Besides, paramyosin and myosin heavy chain contents of the myofibrillar protein from Tegillarca granosa were partially degraded by irradiation treatment [107], indicating the capacity of irradiation treatment to eliminate allergenicity of thermostable allergens and the allergens from actin-binding family.
The pulsed light and cold plasma are two novel processing technique that are able to decrease the seafood allergenicity by alter the conformation of allergens (Figure 5d, 5e). Pulsed light delivers wavelengths from 200 nm to 1000 nm is formed by short and intensive pulses of white light with a broad spectrum and is made up of 54% ultraviolet light, 20% infrared, and 26% visible light radiation [108]. Such efficient pulsed ultraviolet light can ionize proteins by absorbing photons, such as the absorption and recombination of aromatic amino acids. Pulsed ultraviolet light induces allergenic proteins cross-linking with some heat-sensitive proteins and further leads to allergenicity reduction [32] (Figure 5d). Cold plasma is in a gaseous state generated at 30-60 °C under an atmospheric or vacuum, it changes the protein conformation presumably by the cleavage of peptide bonds, oxidization of amino acid side residuals, and formation of the protein-protein cross-linkages, which further induce the effectively lower IgE-binding activity of allergens [109] (Figure 5e). The cold plasma has been applied for the eliminating of shrimp tropomyosin, surface hydrophobicity increased, and free sulfhydryl decreased was observed in tropomyosin with a longer time of treatment [110]. The current studies show a promising performance of pulsed light and cold plasma in reducing allergenicity, follow-up studies are now focusing on better setup parameters and combining these methods with other technologies.
3.2. Processing Techniques Based on Chemistry
In recent years, enzymatic-catalyzed cross-linking and glycation modification (Maillard reaction and enzymatic-catalyzed glycation) are preferred to process hypoallergenic food as effective, selective, energy-saving, and eco-friendly techniques.
3.2.1. Enzymatic-Catalyzed Cross-Linking
Cross-linking of proteins has been exploited in the food industry for cereal, dairy, meat, and fish processing as means to stabilize food structure/texture and to improve food functionality [111]. In addition to its textural-modifying effects, the cross-linking of food proteins can effectively conceal IgE-binding epitopes, consequently reducing the allergenic potential of food products [112,113]. Fei et al. reported that the polymerization with tyrosinase of crab arginine kinase affected the action of the gastrointestinal enzymes, and it presented lower antigenic properties than those of the untreated protein [114]. The IgE-binding activity of crab tropomyosin was proved to be mitigated by 63.5% and 34.5% through cross-linking by tyrosinase and horseradish peroxidase, respectively [115]. Noteworthy, polymerization with horseradish peroxidase of tropomyosin facilitated the action of the gastrointestinal enzymes, and its cross-linking product not only has potential of reducing the allergenicity but also has the capacity of inducing the oral tolerance in mice [115]. Allergenicity elimination of tropomyosin can also be found in tropomyosin cross-linking by tyrosinase and laccase in a dose-dependent manner, due to the changes in the structural integrity of the allergenic protein [113,116]. Tian et al. reported that the polymerization with tyrosinase of fish parvalbumin facilitated the action of the gastrointestinal enzymes, and its presented lower antigenic properties than those of the untreated protein [117]. Tyrosinase has also been performed to cross-link crab sarcoplasmic calcium binding protein for allergenicity eliminating by Hu et al. [72].
It can be found from the above examples that the ultimate reactions of enzymatic cross-linking are achieved depending on the enzyme used, the availability of the target reactive groups and the applied process conditions. Tyrosinase is an oxidase that catalyzes the oxidation of tyrosine and results in oxidative cross-linking of tyrosine side chains [118]. Tyrosine in an allergen can be catalyzed by tyrosinase to form new bonds between tyrosine residues and reactive benzoquinone intermediates in proteins, which may introduce both intra- and intermolecular covalent cross-links, thus change and mask the allergic epitopes (Figure 6a). Laccase is another general used oxidase that catalyzes the oxidation of wide-ranging phenolics and related compounds, which results in the formation of unstable aromatic radicals that react with proteins, leading to cross-linked products [113] (Figure 6b). With the help of hydroxyl functional group provide by caffeic acid, the cross-linking efficacy of oxidative enzymes can be augmented [113,114]. While the cross-linking of tropomyosin by tyrosinase was not so efficiency in allergenicity eliminating as peroxidase, because of the different structure of cross-linked tropomyosin: cross-linked by peroxidase resulted in product with higher hydrophilicity than that of tyrosinase, which is easier for pepsin to find cleavage sites [115]. Transglutaminase is responsible for acyl transfer, deamidation, and crosslinking of intra- or inter-chain glutamine peptide moiety, which is the acyl donor and lysine peptide moiety, which is the acyl acceptor [119] (Figure 6c). The modifications of the allergenic proteins using enzyme-catalyzed protein cross-linking offer mild and safe approaches to mitigate the allergenicity of seafood, while more investigation in allergen structure and epitope is necessary for making full use of this technique.
3.2.2. Glycation Modification
Glycation modification is considered a method to change food allergenicity with mild reaction conditions, low cost, and improved functional properties of proteins. The non-enzymatic glycation, also referred to as Maillard reaction, involves a non-enzymatic browning reaction between the amino acid residue of protein and reducing sugar [120]. By introducing sugar chains to the free amino groups on proteins, usually lysine, arginine, and N-terminal amino acid, the Maillard reaction can modify, destroy, or mask sequential epitopes, result in changes in conformational epitopes, or even cause damage to the structure of an allergen (Figure 7a). Previous studies have demonstrated that the impact of Maillard reaction on food allergenicity is varied and relies on the structural properties of allergens, the types and amounts of reducing sugars, and treatment conditions (e. g. temperature, pH, and moisture).
Most of the works in allergenicity elimination through Maillard reaction were targeted at shellfish tropomyosin. According to Han et al., crab tropomyosin glycated by galactose, glucose, and arabinose modified the arginine and lysine in the epitope as well as variated the protein structure, resulted in the decreasing of allergenicity [121]. Similar result was supported by the work of Zhang et al., where shrimp tropomyosin reacted with either glucose, maltose, or maltotriose, after been glycated, lysine in shrimp tropomyosin, especially in the epitopes were modified, and the products showed lower IgE-binding capacity and induced weaker allergy symptoms in vivo [122]. The utilization of distinct reducing sugars in the Maillard reaction induces differential modifications of amino acid residues, with the specific chemical alterations being contingent upon the structural characteristics of both the saccharide and amino acid involved. Lv et al. incubated shrimp tropomyosin with ribose leaded to modification occurred in phenylalanin, isoleucine, and methionine residues of epitopes, the IgE-binding capacity of tropomyosin was decreased for the masked or destroyed epitopes [123]. In a recent work by Zhao et al., Maillard reaction between shrimp tropomyosin and different kinds of reducing sugar were found to sharply decrease the IgG/IgE-binding activity of allergen with different mechanism [124]. The glucose, ribose, and lactose modified the lysine and arginine in the epitopes, while the rehalose and chitosan molecules can form non-covalent interaction with tropomyosin.
Maillard reaction was also found to be efficiency in reducing the allergenicity of tropomyosin from mollusks. The IgE-binding activity of tropomyosin from Octopus fangsiao can be reduced by approximately 60% compared with untreated protein after a 24 h reaction with glucose, it was speculated that the glucose added to lysine and arginine residues of tropomyosin altered the cut sites of trypsin and chymotrypsin [125]. Under different reaction conditions, xylose decreased immunobinding activity of tropomyosin from both Chlamys nobilis and Haliotis discus hannai in different ways. The Maillard reaction between scallop tropomyosin and xylose in dry-state leaded to the modification of lysine and arginine and epitopes were destroyed and masked by saccharide residues on the tropomyosin surface [126]. While in the wet-state Maillard reaction, the xylose tended to modify methionine and asparagine in linear epitopes of abalone tropomyosin, resulted to IgE-binding capacity decreased by approximately 70% [127]. There are some other allergens in seafood where Maillard reaction resulted in allergenicity changes. Arginine kinase from S. paramamosain reacted with arabinose was found to present lower allergenicity both in vitro and in vivo, by the modification of arginine and lysine in epitopes, accompanied with the variation of spatial structure [72,128].
Recently, significant attention has been paid to enzymatic-catalyzed glycation that can site-specifically link the sugar at lower temperatures due to its strict substrate specificity. Transglutaminase can induce an acyl-transfer reaction between the γ-carboxyamide of glutaminyl residues and primary amines, thus, amino sugars can be regarded as a substrate for the cross-linking of protein and sugar [129] (Figure 7b). It was demonstrated that glutamine, lysine, and arginine contain free amino groups were involved in enzymatic cross-linking reactions, which affected the structure of tropomyosin and altered the allergenicity [127,130,131]. These investigations revealed the glutamine and lysine in epitopes were modified by glucosamine, and further glycation might also lead to structure variation of the allergen, thus reduced the IgE-binding activity. As compared with other methods, such as heating or Maillard reaction, enzymatic-catalyzed glycation can induce site-specific modification under mild conditions, transglutaminase-catalyzed glycation between allergen and glucosamine appears to be useful for reducing the allergic hazards of seafood in food industry.
3.3. Processing Techniques Based on Biology
Several studies highlighted the use of biological food processing technologies as a potential allergen mitigation strategy. In terms of reducing food allergenicity, the biological processing is mainly divided into enzymatic hydrolysis and fermentation treatment [132].
3.3.1. Enzymatic Hydrolysis
Enzymatic hydrolysis via proteolytic enzymes is a traditional and effective way to modify the allergenicity of allergens [112,113]. The enzymatic hydrolysis in low degree can partially cleave the protein and leads to the disrupture of conformational and linear epitopes, greater degree of hydrolysis results in the degradation of amino acid sequence which cuts the allergens into small fragments [112]. The proteinases utilized for the preparation of protein hydrolysates can be found in different sources, such as gastric enzymes (e.g., pepsin, trypsin, and chymotrypsin), plant-derived enzymes (e.g., papain and bromelain), and microbial-derived enzymes (e.g., alcalase and substilisin) [133].
Untersmayr et al. reported that under pepsin digestion at pH < 2.5, all proteins of codfish extract including parvalbumin were degraded to small fragments within 1 minute and lost their IgE-binding capability [134]. With the increased gastric enzyme/protein ratio, crustacean tropomyosin can also be degraded into small fragments and underwent notable decreasing in IgE-binding sites [135,136]. The findings by Keshavarz et al. demonstrated similar digestion characteristics of fish tropomyosin to that of crustacean tropomyosin, allergenicity of tropomyosin was gradually decreased as a function of pepsin digestion time. However, allergen digested in gastrointestinal tract is not directly equivalent to an elimination of allergenicity [137]. However, a number of seafood allergens such as arginine kinase, triosephosphate isomerase, filamin C, myosin light chain, and sarcoplasmic calcium binding protein can be digested in gastrointestinal tract while the digestion products retained IgE binding activity [49]. García-Moreno et al. evaluated the impact of different enzymatic treatments (subtilisin, trypsin, and combination of both) on allergenicity of parvalbumin from Micromesistius poutassou and found that subtilisin hydrolysates present the lowest allergenicity [138]. It has been reported that enzymes used singly or in combination could be effective to develop hypoallergenic milk and oyster products [133,139], indicating that modifications of the allergenic proteins using proteolytic hydrolysis may offer new approaches to mitigate the allergenicity of seafood allergens.
3.3.2. Fermentation Treatment
Fermentation It takes place when microorganisms act on food substrates, it plays a crucial role in improving food quality and microbiological stability in the food industry. During fermentation, microbial enzymatic activity modifies or disrupts allergenic epitopes in food substrates, leading to reduced allergen immunoreactivity. Current evidence suggests that fermentation mitigates food allergenicity predominantly by 3 routes: 1) structures and epitopes were destroyed due to proteolysis and acid-induced denaturation; 2) epitopes were masked by glycosylation and Maillard reactions that occurred during fermentation; 3) some allergens were released into the water [140].
Previous research has shown that among various industrial processing methods, fermentation is one of the most effective methods for reducing IgE-binding capacity of fish products [141]. Zou et al observed the changes in allergenicity of Atlantic cod treated by fermentation with Lactobacillus helveticus in the salt concentration of 2% at 30 °C for 60 h. The study reported that the immunoreactivity of parvalbumin decreased after the fermentation, due to protein hydrolysis which leading to the destroying of epitope [142]. Similar results have also been documented by Zhu et al., IgG/IgE-binding activity of Mylopharyngodon piceus muscle can be decreased by fermentation process [143]. For shellfish, fermentation processing has also been demonstrated to effectively mitigate the allergenicity. The manufacturing process of fermented shrimp paste induces gradual protein degradation, particularly affecting tropomyosin, which consequently diminishes the IgE-binding activity of the product [144]. It can be concluded that fermentation processing can be used to reduce seafood allergenicity. Moreover, fermentation, compared to other processing techniques, is able to mitigate food allergenicity without being limited by safety reason (e.g., irradiation) and high cost (e.g., high pressure). Nevertheless, given the complexity of fermentation parameters, clarifying the precise mechanisms underlying allergenicity reduction proves particularly challenging.
3.4. Combination Processing
Combined treatments probably show a better performance in reducing allergenicity. Studies applied the combination of physical and chemical processing methods to lower seafood allergenicity, normally including combined high temperature with high pressure, combined high temperature with enzymatic-catalyzed cross-linking, and combined non-thermal treatment with glycation.
Tropomyosin maintains its activity even after being boiled in water, Long et al. treated shrimp tropomyosin with high hydrostatic pressure (500 MPa and 55 °C for 10 min), the product exhibited significant loss of allergenicity, by combining the high hydrostatic pressure with thermal treatment (55-75 °C), the allergenicity of tropomyosin was thoroughly eliminated [145]. By combining Maillard reaction (with galactose) with high temperature-pressure (0.08 MPa, 115 °C), the IgE-binding activity of the shrimp meat can also be significantly decreased [146]. Moreover, the lysine and arginine in the epitopes of sarcoplasmic calcium binding protein were modified by galactose during Maillard reaction, and the high temperature-pressure can further alter the thermostable allergens, for instance, by breaking the disulfide bonds [146]. Fei et al. found that the singly processed crab arginine kinase was relatively stable without significant alteration of IgE-binding activity after heating (50 °C, 30 min) and cross-linking (tyrosinase, 37 °C, 8 h) [114]. However, the combined treatment of heating (50 °C, 30 min) and cross-linking (tyrosinase, 37 °C, 8 h) significantly reduced the intensity of the allergenicity of arginine kinase as determined inhibition ELISA analysis, and in vivo studies, respectively [114].
Microwave cooking-assisted Maillard reaction (with glucose) was found to be a better way to reduce the immunogenicity and immunoreactivity of the parvalbumin than that by traditional heating method. The microwave heating can speed up chemical synthesis reaction and shorten reaction time from several days to several minutes. The lysine, threonine, and aspartic acid in epitopes of glycated parvalbumin were modified by glucose, meanwhile Ca2+-binding sites were destroyed in the microwave-assisted Maillard reaction, which affected the parvalbumin allergenicity [147]. In addition, the combined Maillard reaction and non-thermal processing techniques to allergen immunogenicity are also reported. Zou et al. reported that cold plasma combined with Maillard reaction (with ribose) is efficiency in altering IgE-binding capacity of shrimp tropomyosin [148]. The reduction was found to associated with the combined effects: modification induced by cold plasma destroyed the core helical structure of tropomyosin and occupied the potential glycation sites, leading to sequent glycation on conserved areas of tropomyosin [148]. Additionally, the destroyed structure exposed increasing number of cleavage sites of trypsin, which may result in lower digestibility and reduced IgE-binding capacity of digestion products.
Compared to the single treatments, the combination of different processing leads to a more significant effect in decreasing seafood allergenicity, because the reduction of allergenicity benefits from different mechanisms of various processing methods. However, Studies investigating combined treatment methods for seafood processing are still insufficient. Information on the epitope and structural property of allergens is the basis for choosing and optimization of the appropriate combined processing methods, research involving combined processing techniques should take the property of allergens into consideration when optimizing the processing condition by orthogonal designed experiments.
4. Challenge and Future Directions
Allergic reactions triggered by seafood consumption represent a worldwide public health issue. Food processing has been widely recognized as a practical and efficient method for reducing allergenicity by inducing structural alterations or epitope modifications in allergens. However, despite its demonstrated efficacy, significant challenges remain in fully characterizing processing-induced structural modifications and elucidating the fundamental mechanisms involved.
Firstly, the molecular foundation of allergic responses lies in the specific interaction between IgE antibodies and their cognate epitopes [149]. Detailed information on the epitope provides crucial insights for elucidating the molecular mechanisms underlying allergenicity reduction through food processing. Chemical-based and biological- based processing methods - enzymatic glycosylation, Maillard reaction, and enzymatic hydrolysis - all function by selectively modifying or cleaving specific amino acid residues within allergen molecules, thereby altering the structure, epitopes, and allergenicity of allergens. The amino acid profile of epitopes can serve as a selection criterion for optimal enzymes/reducing sugars in these processing techniques, consequently enhancing allergen mitigation efficacy. Processing technologies targeting epitope modification might become an important research direction, while the number of characterized epitopes remains limited compared to the actual epitope repertoire, with conformational epitopes being particularly underrepresented. Furthermore, the structural reorganization and conformational shifts occurring during food processing of some seafood allergens can effectively diminish their allergenic potential. Under these conditions, comprehensive structural characterization of allergens is required to establish robust structure-allergenicity relationships and advance mechanistic understanding. Given the structural homology among allergens within the same protein family [150], processing strategies targeting their shared structural motifs could potentially mitigate the allergenicity of multiple allergens concurrently. These findings highlight the critical need for further structural investigations of seafood allergens to advance our understanding. Moreover, in most previous studies, purified allergens have been subjected to food processing that exclude the potential impact of the food matrix. While growing evidence suggests that food processing can induce physicochemical interactions between food matrix components and allergens [151]. These interactions may have dual effects: processing may promote the aggregation of allergens with matrix components, thereby hindering the degradation of allergenic epitopes, alternatively, it can facilitate chemical reactions between allergens and matrix constituents, potentially leading to the formation of novel allergenic compounds [152]. consequently, the complexity of food matrices should be considered in practical production processes. In addition, current investigations into processed seafood allergenicity primarily rely on preclinical data obtained through serological assays, cellular models, and murine systems, critical clinical evidence derived from seafood-allergic patients remains insufficient.
5. Conclusions
Many attempts have been made to identify seafood allergens and clarify the epitopes and structure-allergenicity relationship. Accumulating knowledge regarding allergen structures and epitope profiles has provided critical insights into the molecular basis of food processing-mediated allergenicity mitigation. From a structural-immunological perspective, food processing, particularly those mediated by physical methods, induces conformational modifications in allergenic proteins that ultimately diminish their immunoreactive potential. From an epitope perspective, chemical processing techniques can induce covalent or non-covalent interactions between allergens and various modifiers (enzymes, saccharides, or other allergens), effectively masking critical epitopes through molecular complexation. Biological processing induces substantial structural modifications in allergens, including protein unfolding, polypeptide chain uncoiling, and enzymatic degradation, which collectively alter conformational epitopes and significantly attenuate allergenic potential. Future investigations should prioritize elucidating structure-allergenicity relationships to clarify the fundamental mechanisms underlying allergenicity reduction. This mechanistic understanding will subsequently facilitate the development of innovative processing technologies for effective seafood allergen mitigation or elimination.
Author Contributions
Conceptualization, Yang Yang, Yehao Zhang and Shinong Gu; methodology, Yang Yang, Xinrong He and Fei Huan; software, Jinli Chen and Siyang He; validation, Meng Liu, and Guangming Liu; formal analysis, Yang Yang, Yehao Zhang, and Fei Huan; investigation, Yehao Zhang and Shinong Gu; resources, Yang Yang and shinong Gu; data curation, Jinli Chen; writing—original draft preparation, Yang Yang and Yehao Zhang; writing—review and editing, Guangming Liu; visualization, shinong Gu; supervision, Guangming Liu; project administration, Guangming Liu; funding acquisition, Yang Yang and Guangming Liu. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Natural Science Foundation of Xiamen Municipality, grant number 3502Z202373132, the Natural Science Foundation of Fujian Province, grant number 2023J011666, and the National Natural Science Foundation of China, grant number 31901811, 32072336.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| WHO/IUIS | World Health Organization and International Union of Immunological Societies |
| MW | Molecular weight |
| PDB | Protein data bank |
References
- Frischmeyer-Guerrerio, P. A.; Young, F. D.; Aktas, O. N.; Haque, T. Insights into the clinical, immunologic, and genetic underpinnings of food allergy. Immunol Rev 2024, 326, 162–172. [Google Scholar] [CrossRef] [PubMed]
- Bartha, I.; Almulhem, N.; Santos, A. F. Feast for thought: A comprehensive review of food allergy 2021-2023. J Allergy Clin Immun 2024, 153, 576–594. [Google Scholar] [CrossRef]
- FAO/WHO. Summary report of the Ad hoc Joint FAO/WHO Expert Consultation on Risk Assessment of Food Allergens. Part 1: Review and validation of Codex priority allergen list through risk assessment.; 2022. Available online: https://www.fao.org/3/cb4653en/cb4653en.pdf.
- Kamath, S. D.; Bublin, M.; Kitamura, K.; et al. Cross-reactive epitopes and their role in food allergy. J Allergy Clin Immun 2023, 151, 1178–1190. [Google Scholar] [CrossRef]
- Kvasnicka, J.; Stylianou, K. S.; Nguyen, V. K.; et al. Human Health Benefits from Fish Consumption vs. Risks from inhalation exposures associated with contaminated sediment remediation: Dredging of the Hudson river. Environ Health Perspect 2019, 127, 127004. [Google Scholar] [CrossRef] [PubMed]
- Wai, C. Y. Y.; Leung, N. Y. H.; Chu, K. H.; et al. Overcoming shellfish allergy: How far have we come? Int J Mol Sci 2020, 21, 2234. [Google Scholar] [CrossRef]
- Spolidoro, G. C. I.; Ali, M. M.; Amera, Y. T.; et al. Prevalence estimates of eight big food allergies in Europe: Updated systematic review and meta-analysis. Allergy 2023, 78, 2361–2417. [Google Scholar] [CrossRef]
- Gupta, R. S.; Warren, C. M.; Smith, B. M.; et al. Prevalence and severity of food allergies among US adults. JAMA Netw Open 2019, 2, e185630. [Google Scholar] [CrossRef]
- Davis, C. M.; Gupta, R. S.; Aktas, O. N.; et al. Clinical management of seafood allergy. J Allergy Cl Imm-Pract 2020, 8, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Wai, C. Y. Y.; Leung, N. Y. H.; Leung, A. S. Y.; et al. Seafood allergy in Asia: Geographical Specificity and beyond. Front Allergy 2021, 2, 676903. [Google Scholar] [CrossRef]
- Feng, H.; Zhou, J.; Lu, Y.; et al. Prevalence of self-reported food allergy among adults in Jiangxi, China. World Allergy Organ J 2023, 16, 100773. [Google Scholar] [CrossRef]
- Le, T. T. K.; Tran, T. T. B.; Ho, H. T. M.; et al. The predominance of seafood allergy in Vietnamese adults: Results from the first population-based questionnaire survey. World Allergy Organ J 2020, 13, 100102. [Google Scholar] [CrossRef] [PubMed]
- Leung, A. S. Y.; Wai, C. Y. Y.; Leung, N. Y. H.; et al. Real-world sensitization and tolerance pattern toseafood in fish-allergic individuals. J Allergy Cl Imm-Prac 2024, 12. [Google Scholar]
- Zotova, V.; Clarke, A. E.; Chan, E. S.; et al. Low resolution rates of seafood allergy. J Allergy Cl Imm-Prac 2019, 7, 690–692. [Google Scholar] [CrossRef]
- Sudharson, S.; Kalic, T.; Hafner, C.; Breiteneder, H. Newly defined allergens in the WHO/IUIS Allergen Nomenclature Database during 01/2019-03/2021. Allergy 2021, 76, 3359–3373. [Google Scholar] [CrossRef]
- Ruethers, T.; Taki, A. C.; Karnaneedi, S.; et al. Expanding the allergen repertoire of salmon and catfish. Allergy 2021, 76, 1443–1453. [Google Scholar] [CrossRef]
- Fu, L. L.; Wang, C.; Zhu, Y.; Wang, Y. Seafood allergy: Occurrence, mechanisms and measures. Trends Food Sci Tech 2019, 88, 80–92. [Google Scholar] [CrossRef]
- Wu, Y. T.; Lu, Y. Y.; Huang, Y. H.; et al. Insight analysis of the cross-sensitization of multiple fish parvalbumins via the Th1/Th2 immunological balance and cytokine release from the perspective of safe consumption of fish. Food Qual Saf-Oxford 2022, 6, fyac056. [Google Scholar] [CrossRef]
- Huang, Y. H.; Li, Z. X.; Wu, Y. T.; et al. Comparative analysis of allergenicity and predicted linear epitopes in α and β parvalbumin from turbot (Scophthalmus maximus). J Sci Food Agr 2023, 103, 2313–2324. [Google Scholar] [CrossRef]
- Yang, R. Q.; Chen, Y. L.; Chen, F.; et al. Purification, characterization, and crystal structure of parvalbumins, the major allergens in Mustelus griseus. J Agric Food Chem 2018, 66, 8150–8159. [Google Scholar] [CrossRef]
- Perez-Gordo, M.; Pastor-Vargas, C.; Lin, J.; et al. Epitope mapping of the major allergen from Atlantic cod in Spanish population reveals different IgE-binding patterns. Mol Nutr Food Res 2013, 57, 1283–1290. [Google Scholar] [CrossRef]
- Morales-Amparano, M. B.; Huerta-Ocampo, J. A.; Pastor-Palacios, G.; Teran, L. M. The Role of enolases in allergic disease. J Allergy Cl Imm-Prac 2021, 9, 3026–3032. [Google Scholar] [CrossRef] [PubMed]
- Kuehn, A.; Hilger, C.; Lehners-Weber, C.; et al. Identification of enolases and aldolases as important fish allergens in cod, salmon and tuna: component resolved diagnosis using parvalbumin and the new allergens. Clin Exp Allergy 2013, 43, 811–822. [Google Scholar] [CrossRef] [PubMed]
- Wai, C. Y. Y.; Leung, N. Y. H.; Leung, A. S. Y.; et al. Differential patterns of fish sensitization in Asian populations: Implication for precision diagnosis. Allergol Int 2023, 72, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Sariyer, E.; Yakarsonmez, S.; Danis, O.; et al. A study of Bos taurus muscle specific enolase; biochemical characterization, homology modelling and investigation of molecular interaction using molecular docking and dynamics simulations. Int J Biol Macromol 2018, 120 Pt B, 2346–2353. [Google Scholar] [CrossRef]
- Nakamura, R.; Satoh, R.; Nakajima, Y.; et al. Comparative study of GH-transgenic and non-transgenic amago salmon (Oncorhynchus masou ishikawae) allergenicity and proteomic analysis of amago salmon allergens. Regul Toxicol Pharmacol 2009, 55, 300–308. [Google Scholar] [CrossRef]
- Ruethers, T.; Taki, A. C.; Nugraha, R.; et al. Variability of allergens in commercial fish extracts for skin prick testing. Allergy 2019, 74, 1352–1363. [Google Scholar] [CrossRef]
- Boucher, L. E.; Bosch, J. Structure of Toxoplasma gondii fructose-1,6-bisphosphate aldolase. Acta Crystallogr F Struct Biol Commun 2014, 70 Pt 9, 1186–1192. [Google Scholar] [CrossRef]
- Liu, R.; Holck, A. L.; Yang, E.; et al. Tropomyosin from tilapia (Oreochromis mossambicus) as an allergen. Clin Exp Allergy 2013, 43, 365–377. [Google Scholar] [CrossRef]
- Li, M. S.; Xia, F.; Liu, Q. M.; et al. Identification and allergenicity analysis of tropomyosin: A heat-stable allergen in Lateolabrax japonicus. J Agric Food Chem 2024, 73, 822–834. [Google Scholar] [CrossRef]
- Whitby, F. G.; Phillips, G. N. Crystal structure of tropomyosin at 7 Angstroms resolution. Proteins 2000, 38, 49–59. [Google Scholar] [CrossRef]
- Cheng, J. H.; Wang, H.; Sun, D. W. An overview of tropomyosin as an important seafood allergen: Structure, cross-reactivity, epitopes, allergenicity, and processing modifications. Compr Rev Food Sci Food Saf 2022, 21, 127–147. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Zhang, X. M.; Wen, Y. Q.; et al. Comparison of tropomyosin released peptide and epitope mapping after in vitro digestion from fish (Larimichthys crocea), shrimp (Litopenaeus vannamei) and clam (Ruditapes philippinarum) through SWATH-MS based proteomics. Food Chem 2023, 403. [Google Scholar] [CrossRef] [PubMed]
- Hamada, Y.; Nagashima, Y.; Shiomi, K. Identification of collagen as a new fish allergen. Biosci Biotech Bioch 2001, 65, 285–291. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Akiyama, H.; Huge, J.; et al. Fish collagen is an important panallergen in the Japanese population. Allergy 2016, 71, 720–723. [Google Scholar] [CrossRef]
- Shimojo, N.; Yagami, A.; Ohno, F.; et al. Fish collagen as a potential indicator of severe allergic reactions among patients with fish allergies. Clin Exp Allerg 2022, 52, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Bella, J. Collagen structure: new tricks from a very old dog. Biochem J 2016, 473, 1001–1025. [Google Scholar] [CrossRef]
- Boudko, S. P.; Bachinger, H. P. Structural insight for chain selection and stagger control in collagen. Sci Rep-UK 2016, 6, 37831. [Google Scholar] [CrossRef]
- Kalic, T.; Kamath, S. D.; Ruethers, T.; et al. Collagen-an important fish allergen for improved diagnosis. J Allergy Cl Imm-Prac 2020, 8, 3084–3092 e3010. [Google Scholar] [CrossRef]
- Valverde-Monge, M.; Pastor-Vargas, C.; Rodriguez Del Rio, P.; et al. Anaphylaxis by exclusive allergy to swordfish and identification of a new fish allergen. Pediat Allerg Imm-UK 2018, 29, 563–565. [Google Scholar] [CrossRef]
- Xia, F.; Li, M. S.; Liu, Q. M.; et al. Crystal structure analysis and conformational epitope mutation of triosephosphate isomerase, a mud crab allergen. J Agric Food Chem 2019, 67, 12918–12926. [Google Scholar] [CrossRef]
- Larco-Rojas, X.; Gonzalez-Gutierrez, M. L.; Vazquez-Cortes, S.; et al. Occupational asthma and urticaria in a fishmonger due to creatine kinase, a cross-reactive fish allergen. J Invest Allerg Clin 2017, 27, 386–388. [Google Scholar] [CrossRef]
- Yang, Y.; He, X. R.; Huan, F.; et al. A Crystal structure of Pro c 2 provides insights into cross-reactivity of aquatic allergens from the phosphagen kinase family. J Agric Food Chem 2024, 72, 28400–28411. [Google Scholar] [CrossRef] [PubMed]
- Hanaoka, K.; Takahagi, S.; Ishii, K.; et al. Type-I-hypersensitivity to 15 kDa, 28 kDa and 54 kDa proteins in vitellogenin specific to Gadus chalcogrammus roe. Allergol Int 2020, 69, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, D. R.; Day, E. D., Jr.; Miller, J. S. The major heat stable allergen of shrimp. Ann Allergy 1981, 47, 17–22. [Google Scholar]
- Nugraha, R.; Kamath, S. D.; Johnston, E.; et al. Conservation analysis of B-cell allergen epitopes to predict clinical cross-reactivity between shellfish and inhalant invertebrate allergens. Front Immunol 2019, 10, 2676. [Google Scholar] [CrossRef] [PubMed]
- Chen, B. J.; He, H.; Wang, X.; et al. Research progress on shrimp allergens and allergenicity reduction methods. Foods 2025, 14, 895. [Google Scholar] [CrossRef]
- James, J. K.; Pike, D. H.; Khan, I. J.; et al. Structural and dynamic properties of allergen and non-allergen forms of tropomyosin. Structure 2018, 26, 997. [Google Scholar] [CrossRef]
- Liu, M.; Huan, F.; Li, M. S.; et al. Mapping and IgE-binding capacity analysis of heat/digested stable epitopes of mud crab allergens. Food Chem 2021, 344, 128735. [Google Scholar] [CrossRef]
- Lv, L. T.; Wei, F. L.; Liu, L.; et al. Study on the allergenicity of tropomyosin from different aquatic products based on conformational and linear epitopes analysis. J Agric Food Chem 2025, 73, 4936–4946. [Google Scholar] [CrossRef]
- Li, M. S.; Xia, F.; Liu, Q. M; et al. IgE Epitope analysis for Scy p 1 and Scy p 3, the heat-stable myofibrillar allergens in mud crab. J Agric Food Chem 2022, 12189–12202. [Google Scholar] [CrossRef]
- Yu, C. J.; Lin, Y. F.; Chiang, B. L.; et al. Proteomics and immunological analysis of a novel shrimp allergen, Pen m 2. J Immunol 2003, 170, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liu, G. Y.; Yang, H.; et al. Crystal structure determination of Scylla paramamosain arginine kinase, an allergen that may cause cross-reactivity among invertebrates. Food Chem 2019, 271, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Cao, M. J.; Alcocer, M.; et al. Mapping and characterization of antigenic epitopes of arginine kinase of Scylla paramamosain. Mol Immunol 2015, 65, 310–320. [Google Scholar] [CrossRef] [PubMed]
- Chen, H. L.; Mao, H. Y.; Cao, M. J.; et al. Purification, physicochemical and immunological characterization of arginine kinase, an allergen of crayfish (Procambarus clarkii). Food Chem Toxicol 2013, 62, 475–484. [Google Scholar] [CrossRef]
- Mao, H. Y.; Cao, M. J.; Maleki, S. J.; et al. Structural characterization and IgE epitope analysis of arginine kinase from Scylla paramamosain. Mol Immunol 2013, 56, 463–470. [Google Scholar] [CrossRef]
- Yu, C.; Gao, X.; Lin, H.; et al. Identification and amino acid analysis of allergenic epitopes of a novel allergen paramyosin (Rap v 2) from Rapana venosa. J Agric Food Chem 2021, 69, 5381–5391. [Google Scholar] [CrossRef]
- Yu, C.; Ding, X.; Gao, X.; et al. Immunological cross-reactivity involving mollusc species and mite-mollusc and cross-reactive allergen PM are risk factors of mollusc allergy. J Agric Food Chem 2022, 70, 360–372. [Google Scholar] [CrossRef]
- Yu, C.; Gao, X.; Lin, H.; et al. Purification, characterization, and three-dimensional structure prediction of paramyosin, a novel allergen of Rapana venosa. J Agric Food Chem 2020, 68, 14632–14642. [Google Scholar] [CrossRef]
- Yang, Y. T.; Gourinath, S.; Kovacs, M.; et al. Rigor-like structures from muscle myosins reveal key mechanical elements in the transduction pathways of this allosteric motor. Structure 2007, 15, 553–564. [Google Scholar] [CrossRef]
- Conti, A.; Alqassir, N.; Breda, D.; et al. Serological proteome analysis identifies crustacean myosin heavy chain type 1 protein and house dust mite Der p 14 as cross-reacting allergens. Adv Clin Exp Med 2023, 32, 107–112. [Google Scholar] [CrossRef]
- Saenz de San Pedro, B.; Lopez Guerrero, A.; Navarrete del Pino, M. A.; et al. Myosin heavy chain: An allergen involved in anaphylaxis to shrimp head. J Invest Allerg Clin 2023, 33, 66–68. [Google Scholar]
- Shibata, Y.; Satoshi, S.; Oishi, T.; et al. Myosin heavy chain, a novel allergen for fish allergy in patients with atopic dermatitis. J Invest Dermatol 2019, 139, S223–S223. [Google Scholar] [CrossRef]
- Ayuso, R.; Grishina, G.; Bardina, L.; et al. Myosin light chain is a novel shrimp allergen, Lit v 3. J Allergy Clin Immun 2008, 122, 795–802. [Google Scholar] [CrossRef]
- Rahman, A. M. A.; Kamath, S.; Lopata, A. L.; et al. Analysis of the allergenic proteins in black tiger prawn (Penaeus monodon) and characterization of the major allergen tropomyosin using mass spectrometry. Rapid Commun Mass Sp 2010, 24, 2462–2470. [Google Scholar] [CrossRef]
- Bauermeister, K.; Wangorsch, A.; Garoffo, L. P.; et al. Generation of a comprehensive panel of crustacean allergens from the North Sea Shrimp Crangon crangon. Mol Immunol 2011, 48, 1983–1992. [Google Scholar] [CrossRef]
- Li, M. S.; Xia, F.; Liu, M.; et al. Cloning, expression, and epitope identification of myosin light chain 1: An allergen in mud crab. J Agric Food Chem 2019, 67, 10458–10469. [Google Scholar] [CrossRef]
- Yang, Y.; Yan, H. F.; Zhang, Y. X.; et al. Expression and epitope identification of myosin light chain isoform 1, an allergen in Procambarus clarkii. Food Chem 2020, 317, 126422. [Google Scholar] [CrossRef]
- Zhang, Y. X.; Chen, H. L.; Maleki, S. J.; et al. Purification, characterization, and analysis of the allergenic properties of myosin light chain in Procambarus clarkii. J Agric Food Chem 2015, 63, 6271–6282. [Google Scholar] [CrossRef] [PubMed]
- Ayuso, R.; Grishina, G.; Dolores Ibanez, M.; et al. Sarcoplasmic calcium-binding protein is an EF-hand-type protein identified as a new shrimp allergen. J Allergy Clin Immun 2009, 124, 114–120. [Google Scholar] [CrossRef]
- Han, T. J.; Huan, F.; Liu, M.; et al. IgE epitope analysis of sarcoplasmic-calcium-binding protein, a heat-resistant allergen in Crassostrea angulata. Food Funct 2021, 12, 8570–8582. [Google Scholar] [CrossRef]
- Hu, M. J.; Liu, G. Y.; Yang, Y.; et al. Cloning, expression, and the effects of processing on sarcoplasmic calcium-binding protein: an important allergen in mud crab. J Agric Food Chem 2017, 65, 6247–6257. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y. Y.; Jin, T.; Li, M. S.; et al. Crystal structure analysis of sarcoplasmic-calcium-binding protein: an allergen in Scylla paramamosain. J Agric Food Chem 2023, 71, 1214–1223. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhang, Y. X.; Liu, M.; et al. Triosephosphate isomerase and filamin C share common epitopes as novel allergens of Procambarus clarkii. J Agric Food Chem 2017, 65, 950–963. [Google Scholar] [CrossRef]
- Xia, F.; Li, M. S.; Liu, Q. M.; et al. Allergenicity and linear epitope analysis of Scy p 8, an allergen from mud crab. J Agric Food Chem 2024, 72, 13402–13414. [Google Scholar] [CrossRef]
- He, X. R.; Yang, Y.; Chen, Y. X.; et al. Immunoglobulin E epitope mapping and structure-allergenicity relationship analysis of crab allergen Scy p 9. J Agric Food Chem 2023, 71, 17379–17390. [Google Scholar] [CrossRef]
- He, X. R.; Yang, Y.; Kang, S.; et al. Crystal structure analysis and ige epitope mapping of allergic predominant region in Scylla paramamosain filamin C, Scy p 9. J Agric Food Chem 2022, 70, 1282–1292. [Google Scholar] [CrossRef] [PubMed]
- Karnaneedi, S.; Huerlimann, R.; Johnston, E. B.; et al. Novel allergen discovery through comprehensive de novo transcriptomic analyses of five shrimp species. Int J Mol Sci 2021, 22, 32. [Google Scholar] [CrossRef]
- Kato, Y. S.; Yumoto, F.; Tanaka, H.; et al. Structure of the Ca2+-saturated C-terminal domain of scallop troponin C in complex with a troponin I fragment. Biological Chemistry 2013, 394, 55–68. [Google Scholar] [CrossRef]
- Giuffrida, M. G.; Villalta, D.; Mistrello, G.; et al. Shrimp allergy beyond tropomyosin in Italy: clinical relevance of arginine kinase, sarcoplasmic calcium binding protein and hemocyanin. Eur Ann Allergy Clin 2014, 46, 172–177. [Google Scholar]
- Lu, J.; Luan, H.; Wang, C.; et al. Molecular and allergenic properties of natural hemocyanin from Chinese mitten crab (Eriocheir sinensis). Food Chem 2023, 424, 136422. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, Y.; Zhang, S.; et al. The effect of immunotherapy on cross-reactivity between house dust mite and other allergens in house dust mite -sensitized patients with allergic rhinitis. Expert Rev Clin Immu 2021, 17, 969–975. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; She, T.; Zhang, Y.; et al. Identification and characterization of ovary development-related protein EJO1 (Eri s 2) from the ovary of Eriocheir sinensis as a new food allergen. Mol Nutr Food Res 2016, 60, 2275–2287. [Google Scholar] [CrossRef]
- Munera, M.; Martinez, D.; Wortmann, J.; et al. Structural and allergenic properties of the fatty acid binding protein from shrimp Litopenaeus vannamei. Allergy 2022, 77, 1534–1544. [Google Scholar] [CrossRef] [PubMed]
- Wai, C. Y. Y.; Leung, N. Y. H.; Leung, A. S. Y.; et al. Comprehending the allergen repertoire of shrimp for precision molecular diagnosis of shrimp allergy. Allergy 2022, 77, 3041–3051. [Google Scholar] [CrossRef] [PubMed]
- Khan, M. U.; Ahmed, I.; Lin, H.; et al. Potential efficacy of processing technologies for mitigating crustacean allergenicity. Crit Rev Food Sci Nutr 2019, 59, 2807–2830. [Google Scholar] [CrossRef]
- Rosmilah, M.; Shahnaz, M.; Meinir, J.; et al. Identification of parvalbumin and two new thermolabile major allergens of Thunnus tonggol using a proteomics approach. Int Arch Allergy Imm 2013, 162, 299–309. [Google Scholar] [CrossRef]
- Kubota, H.; Kobayashi, A.; Kobayashi, Y.; et al. Reduction in IgE reactivity of Pacific mackerel parvalbumin by heat treatment. Food Chem 2016, 206, 78–84. [Google Scholar] [CrossRef]
- Liang, J.; Taylor, S. L.; Baumert, J.; et al. Effects of thermal treatment on the immunoreactivity and quantification of parvalbumin from Southern hemisphere fish species with two anti-parvalbumin antibodies. Food Control 2021, 121, 107675. [Google Scholar] [CrossRef]
- van der Ventel, M. L.; Nieuwenhuizen, N. E.; Kirstein, F.; et al. Differential responses to natural and recombinant allergens in a murine model of fish allergy. Mol Immunol 2011, 48, 637–646. [Google Scholar] [CrossRef]
- Chen, Y. T.; Hsieh, Y. H. P. Characterization of a 36 kDa antigenic protein of fish-specific monoclonal-antibody 8F5. Food Chem 2022, 379, 132149. [Google Scholar] [CrossRef]
- Fang, L.; Li, G. M.; Zhang, J. T.; et al. Identification and mutational analysis of continuous, immunodominant epitopes of the major oyster allergen Crag 1. Clin Immunol 2019, 201, 20–29. [Google Scholar] [CrossRef]
- Zhao, J. L.; Li, Y. H.; Xu, L. L.; et al. Thermal induced the structural alterations, increased IgG/IgE binding capacity and reduced immunodetection recovery of tropomyosin from shrimp (Litopenaeus vannamei). Food Chem 2022, 391, 133215. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J. L; Zhu, W. Y.; Zeng, J. H.; et al. Insight into the mechanism of allergenicity decreasing in recombinant sarcoplasmic calcium-binding protein from shrimp (Litopenaeus vannamei) with thermal processing via spectroscopy and molecular dynamics simulation techniques. Food Res Int 2022, 157. [Google Scholar] [CrossRef]
- Cabanillas, B.; Novak, N. Effects of daily food processing on allergenicity. Crit Rev Food Sci Nutr 2019, 59, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Ekezie, F. G. C.; Cheng, J. H.; Sun, D. W. Effects of nonthermal food processing technologies on food allergens: A review of recent research advances. Trends Food Sci Tech 2018, 74, 12–25. [Google Scholar] [CrossRef]
- Jin, Y. F.; Deng, Y.; Qian, B. J.; et al. Allergenic response to squid (Todarodes pacificus) tropomyosin Tod p1 structure modifications induced by high hydrostatic pressure. Food Chem Toxicol 2015, 76, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Carrera, M.; Fidalgo, L. G.; Saraiva, J. A.; et al. Effects of high-pressure treatment on the muscle proteome of hake by bottom-up proteomics. J Agric Food Chem 2018, 66, 4559–4570. [Google Scholar] [CrossRef]
- Zhang, H. E.; Liao, H. Q.; Lu, Y. B.; et al. Effects of high hydrostatic pressure on the structural characteristics of parvalbumin of cultured large yellow croaker (Larimichthys crocea). J Food Process Pres 2020, 44, e14911. [Google Scholar] [CrossRef]
- Chen, W. Q.; Ma, H. L.; Wang, Y. Y. Recent advances in modified food proteins by high intensity ultrasound for enhancing functionality: Potential mechanisms, combination with other methods, equipment innovations and future directions. Ultrason Sonochem 2022, 85, 105993. [Google Scholar] [CrossRef]
- Zhang, Z. Y.; Zhang, X. F.; Chen, W.; et al. Conformation stability, in vitro digestibility and allergenicity of tropomyosin from shrimp (Exopalaemon modestus) as affected by high intensity ultrasound. Food Chem 2018, 245, 997–1009. [Google Scholar] [CrossRef]
- Li, Z. X.; Lin, H.; Cao, L. M.; et al. Effect of high intensity ultrasound on the allergenicity of shrimp. J Zhejiang Univ Sci B 2006, 7, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Vanga, S. K.; Singh, A.; Raghavan, V. Review of conventional and novel food processing methods on food allergens. Crit Rev Food Sci Nutr 2017, 57, 2077–2094. [Google Scholar] [CrossRef] [PubMed]
- Pan, M. F.; Yang, J. Y.; Liu, K. X.; et al. Irradiation technology: An effective and promising strategy for eliminating food allergens. Food Res Int 2021, 148, 110578. [Google Scholar] [CrossRef]
- Li, Z. X.; Lu, Z. C.; Khan, M. N.; et al. Protein carbonylation during electron beam irradiation may be responsible for changes in IgE binding to turbot parvalbumin. Food Chem Toxicol 2014, 69, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Guan, A.; Mei, K.; Lv, M.; et al. The effect of electron beam irradiation on IgG binding capacity and conformation of tropomyosin in shrimp. Food Chemistry 2018, 264, 250–254. [Google Scholar] [CrossRef]
- Lv, M. C.; Mei, K. L.; Zhang, H.; et al. Effects of electron beam irradiation on the biochemical properties and structure of myofibrillar protein from Tegillarca granosa meat. Food Chem 2018, 254, 64–69. [Google Scholar] [CrossRef]
- Salazar, F.; Pizarro-Oteiza, S.; Kasahara, I.; et al. Effect of pulsed ultraviolet light on the degree of antigenicity and hydrolysis in cow's milk proteins. J Food Process Eng 2024, 47, e14629. [Google Scholar] [CrossRef]
- Kaur, S.; Kumar, Y.; Singh, V.; et al. Cold plasma technology: Reshaping food preservation and safety. Food Control 2024, 163, 110537. [Google Scholar] [CrossRef]
- Ekezie, F. G. C.; Sun, D. W.; Cheng, J. H. Altering the IgE binding capacity of king prawn (Litopenaeus vannamei) tropomyosin through conformational changes induced by cold argon-plasma jet. Food Chem 2019, 300, 125143. [Google Scholar] [CrossRef]
- Shan, C. Y.; Wu, G. Q.; Bu, Y.; et al. Effect of transglutaminase cross-linking on debittering of Alaska pollock frame hydrolysate. Int J Food Sci Tech 2024, 59, 438–449. [Google Scholar] [CrossRef]
- Zeng, J.; Wang, Q.; Yi, H.; et al. Recent insights in cow's milk protein allergy: Clinical relevance, allergen features, and influences of food processing. Trends Food Sci Tech 2025, 156, 104830. [Google Scholar] [CrossRef]
- Ahmed, I.; Lin, H.; Xu, L.; et al. Immunomodulatory effect of laccase/caffeic acid and transglutaminase in alleviating shrimp tropomyosin (Met e 1) allergenicity. J Agric Food Chem 2020, 68, 7765–7778. [Google Scholar] [CrossRef] [PubMed]
- Fei, D. X.; Liu, Q. M.; Chen, F.; et al. Assessment of the sensitizing capacity and allergenicity of enzymatic cross-linked arginine kinase, the crab allergen. Mol Nutr Food Res 2016, 60, 1707–1718. [Google Scholar] [CrossRef]
- Liu, G. Y.; Hu, M. J.; Sun, L. C.; et al. Allergenicity and oral tolerance of enzymatic cross-linked tropomyosin evaluated using cell and mouse models. J Agric Food Chem 2017, 65, 2205–2213. [Google Scholar] [CrossRef]
- Ahmed, I.; Lin, H.; Li, Z. X.; et al. Tyrosinase/caffeic acid cross-linking alleviated shrimp (Metapenaeus ensis) tropomyosin-induced allergic responses by modulating the Th1/Th2 immunobalance. Food Chem 2021, 340, 127948. [Google Scholar] [CrossRef] [PubMed]
- Tian, S. L.; Ma, J. J.; Ahmed, I.; et al. Effect of tyrosinase-catalyzed crosslinking on the structure and allergenicity of turbot parvalbumin mediated by caffeic acid. J Sci Food Agri 2019, 99, 3501–3508. [Google Scholar] [CrossRef]
- Li, Z. Y.; Lu, F. P.; Liu, Y. H. A Review of the mechanism, properties, and applications of hydrogels prepared by enzymatic cross-linking. J Agric Food Chem 2023, 71, 10238–10249. [Google Scholar] [CrossRef]
- Vasic, K.; Knez, Z.; Leitgeb, M. Transglutaminase in foods and biotechnology. Int J Mol Sci 2023, 24, 12402. [Google Scholar] [CrossRef]
- Xu, Y.; Ahmed, I.; Zhao, Z. X.; et al. A comprehensive review on glycation and its potential application to reduce food allergenicity. Crit Rev Food Sci Nutr 2024, 64, 12184–12206. [Google Scholar] [CrossRef]
- Han, X. Y.; Yang, H.; Rao, S. T.; et al. The Maillard reaction reduced the sensitization of tropomyosin and arginine kinase from Scylla paramamosain, Simultaneously. J Agric Food Chem 2018, 66, 2934–2943. [Google Scholar] [CrossRef]
- Zhang, Z. Y.; Xiao, H.; Zhang, X. F.; et al. Insight into the effects of deglycosylation and glycation of shrimp tropomyosin on in vivo allergenicity and mast cell function. Food Funct 2019, 10, 3934–3941. [Google Scholar] [CrossRef]
- Lv, L. T.; Ahmed, I.; Qu, X.; et al. Effect of the structure and potential allergenicity of glycated tropomyosin, the shrimp allergen. Int J Food Sci Tech 2022, 57, 1782–1790. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, J.; Xu, L.; et al. Insights into the mechanism underlying the influence of glycation with different saccharides and temperatures on the IgG/IgE binding ability, immunodetection, in vitro digestibility of shrimp (Litopenaeus vannamei) tropomyosin. Foods 2023, 12, 3049. [Google Scholar] [CrossRef] [PubMed]
- Shen, H. W.; Liu, Y. Y.; Chen, F.; et al. Purification, characterization and immunoreactivity of tropomyosin, the allergen in Octopus fangsiao. Process Biochem 2014, 49, 1747–1756. [Google Scholar] [CrossRef]
- Bai, T. L.; Han, X. Y.; Li, M. S.; et al. Effects of the Maillard reaction on the epitopes and immunoreactivity of tropomyosin, a major allergen in Chlamys nobilis. Food Funct 2021, 12, 5096–5108. [Google Scholar] [CrossRef]
- Ji, N. R.; Yu, C. C.; Han, X. Y.; et al. Effects of three processing technologies on the structure and immunoreactivity of α-tropomyosin from Haliotis discus hannai. Food Chem 2023, 405, 134947. [Google Scholar] [CrossRef]
- Wu, Y. T.; Lu, Y. Y.; Huang, Y. H.; et al. Glycosylation reduces the allergenicity of turbot (Scophthalmus maximus) parvalbumin by regulating digestibility, cellular mediators release and Th1/Th2 immunobalance. Food Chem 2022, 382, 132574. [Google Scholar] [CrossRef]
- Ahmed, I.; Chen, H.; Li, J. L.; et al. Enzymatic crosslinking and food allergenicity: A comprehensive review. Compr Rev Food Sci Food Saf 2021, 20, 5856–5879. [Google Scholar] [CrossRef]
- Yuan, F. Z.; Lv, L. T.; Li, Z. X.; et al. Effect of transglutaminase-catalyzed glycosylation on the allergenicity and conformational structure of shrimp (Metapenaeus ensis) tropomyosin. Food Chem 2017, 219, 215–222. [Google Scholar] [CrossRef]
- Li, D. X.; He, X. R.; Li, F. J.; et al. Effect of transglutaminase-catalyzed glycosylation on the allergenicity of tropomyosin in the Perna viridis food matrix. Food Funct 2024, 15, 9136–9148. [Google Scholar] [CrossRef] [PubMed]
- Gunal-Koroglu, D.; Karabulut, G.; Ozkan, G.; et al. Allergenicity of alternative proteins: Reduction mechanisms and processing strategies. J Agric Food Chem 2025, 73, 7522–7546. [Google Scholar] [CrossRef]
- Abd El-Salam, M. H.; El-Shibiny, S. Reduction of Milk protein antigenicity by enzymatic hydrolysis and fermentation. A review. Food Rev Int 2021, 37, 276–295. [Google Scholar] [CrossRef]
- Untersmayr, E.; Poulsen, L. K.; Platzer, M. H.; et al. The effects of gastric digestion on codfish allergenicity. J Allergy Clin Immu 2005, 115, 377–382. [Google Scholar] [CrossRef]
- Liu, G. M.; Huang, Y. Y.; Cai, Q. F.; et al. Comparative study of in vitro digestibility of major allergen, tropomyosin and other proteins between Grass prawn (Penaeus monodon) and Pacific white shrimp (Litopenaeus vannamei). J Sci Food Agric 2011, 91, 163–170. [Google Scholar] [CrossRef]
- Gamez, C.; Paz Zafra, M.; Sanz, V.; et al. Simulated gastrointestinal digestion reduces the allergic reactivity of shrimp extract proteins and tropomyosin. Food Chem 2015, 173, 475–481. [Google Scholar] [CrossRef]
- Keshavarz, B.; Rao, Q. C.; Jiang, X. Y.; et al. Immunochemical analysis of pepsin-digested fish tropomyosin. Food Control 2020, 118, 107427. [Google Scholar] [CrossRef]
- Garcia-Moreno, P. J.; Perez-Galvez, R.; Javier Espejo-Carpio, F.; et al. Functional, bioactive and antigenicity properties of blue whiting protein hydrolysates: effect of enzymatic treatment and degree of hydrolysis. J Sci Food Agric 2017, 97, 299–308. [Google Scholar] [CrossRef]
- Wu, M. X.; Huan, F.; Gao, S.; et al. Preparation of the hypoallergenic enzymatic hydrolyzate of Cra a 4 with the potential to induce immune tolerance. J Agric Food Chem 2025, 73, 4299–4308. [Google Scholar] [CrossRef]
- Pi, X. W.; Yang, Y. L.; Sun, Y. X; et al. Recent advances in alleviating food allergenicity through fermentation. Crit Rev Food Sci Nutr 2022, 62, 7255–7268. [Google Scholar] [CrossRef]
- Mejrhit, N.; Azdad, O.; Aarab, L. Effect of industrial processing on the IgE reactivity of three commonly consumed Moroccan fish species in Fez region. Eur Ann Allergy Clin Immunol 2018, 50, 202–210. [Google Scholar] [CrossRef]
- Zou, H.; Zhao, J.; Qin, Z.; et al. Influence of fermentation by Lactobacillus helveticus on the immunoreactivity of Atlantic cod allergens. J Agric Food Chem 2023, 71, 10144–10154. [Google Scholar] [CrossRef]
- Zhu, Y. D.; Gao, L.; Xie, G. J.; et al. Effect of fermentation on immunological properties of allergens from black carp (Mylopharyngodon piceus) sausages. Int J Food Sci Tech 2020, 55, 3162–3172. [Google Scholar] [CrossRef]
- Amalia, U.; Shimizu, Y.; Saeki, H. Variation in shrimp tropomyosin allergenicity during the production of Terasi, an Indonesian fermented shrimp paste. Food Chem 2023, 398, 133876. [Google Scholar] [CrossRef]
- Long, F. Y.; Yang, X.; Wang, R. R.; et al. Effects of combined high pressure and thermal treatments on the allergenic potential of shrimp (Litopenaeus vannamei) tropomyosin in a mouse model of allergy. Innov Food Sci Emerg Tech 2015, 29, 119–124. [Google Scholar] [CrossRef]
- Liu, M.; Han, T. J.; Huan, F.; et al. Effects of thermal processing on the allergenicity, structure, and critical epitope amino acids of crab tropomyosin. Food Funct 2021, 12, 2032–2043. [Google Scholar] [CrossRef]
- Tai, J. J.; Qiao, D.; Huang, X.; et al. Structural property, immunoreactivity and gastric digestion characteristics of glycated parvalbumin from mandarin fish (Siniperca chuaisi) during microwave-assisted Maillard reaction. Foods 2023, 12, 52. [Google Scholar] [CrossRef]
- Zou, S.; Wang, F. Q.; Cheng, J. H.; et al. Mechanism of cold plasma combined with glycation in altering IgE-binding capacity and digestion stability of tropomyosin from shrimp. J Agric Food Chem 2023, 71, 15796–15808. [Google Scholar] [CrossRef]
- Crespo, J. F.; Cabanillas, B. Recent advances in cellular and molecular mechanisms of IgE-mediated food allergy. Food Chem 2023, 411, 135500. [Google Scholar] [CrossRef]
- Yang, Y.; He, X. R.; Li, F. J.; et al. Animal-derived food allergen: A review on the available crystal structure and new insights into structural epitope. Compr Rev Food Sci Food Saf 2024, 23, e13340. [Google Scholar] [CrossRef]
- Gou, J.; Liang, R.; Huang, H.; et al. Maillard reaction induced changes in allergenicity of food. Foods 2022, 11, 530. [Google Scholar] [CrossRef]
- Liu, Q. Z.; Lin, S. Y.; Sun, N. How does food matrix components affect food allergies, food allergens and the detection of food allergens? A systematic review. Trends Food Sci Tech 2022, 127, 280–290. [Google Scholar] [CrossRef]
Figure 1.
Classification of common consumed seafood species.

Figure 2.
Representative structures of fish allergens. (a) Structure of α-parvalbumins (PDB: 5ZGM) and β-parvalbumin (5ZH6), the protein contains 2 Ca2+-binding domains and the 2 bound Ca2+ were showed in cyan ball, the important IgE-binding epitope of β-parvalbumin (AA 95-108) was colored red; (b) Structure of β-enolase (PDB: 2XSX), the β-enolase homodimer is composed of 2 identical monomers, represented in light blue and light grey for clarity; the monomer of β-enolase is comprised of the N-terminal domain (the α-helixes represent in orange, β-sheet represent in yellow) and C-terminal α/β-barrel (the α-helixes represent in light blue, β-sheet represent in marine blue), a bound Mg2+ in the α/β-barrel was showed in cyan ball; (c) Structure of aldolase A (PDB: 4TU1), the aldolase A oligomer is composed of 4 identical monomers, represented in light blue, light grey, wheat, and pale green for clarity; the monomer of aldolase A folds into an α/β-barrel with 10 anti-parallel β-sheets (represent in marine blue) bundled by 13 α-helices (represent in light blue); (d) Structure of tropomyosin (PDB: 1C1G), the homodimer of tropomyosin is comprised of 2 identically coiled subunits, represented in wheat and light grey for clarity; (e) Structure of Collagen I (PDB: 5CVB), the collagen triple helix was formed by association of 3 identical or different polypeptide chains, represented in wheat, light blue, and light grey for clarity.
Figure 2.
Representative structures of fish allergens. (a) Structure of α-parvalbumins (PDB: 5ZGM) and β-parvalbumin (5ZH6), the protein contains 2 Ca2+-binding domains and the 2 bound Ca2+ were showed in cyan ball, the important IgE-binding epitope of β-parvalbumin (AA 95-108) was colored red; (b) Structure of β-enolase (PDB: 2XSX), the β-enolase homodimer is composed of 2 identical monomers, represented in light blue and light grey for clarity; the monomer of β-enolase is comprised of the N-terminal domain (the α-helixes represent in orange, β-sheet represent in yellow) and C-terminal α/β-barrel (the α-helixes represent in light blue, β-sheet represent in marine blue), a bound Mg2+ in the α/β-barrel was showed in cyan ball; (c) Structure of aldolase A (PDB: 4TU1), the aldolase A oligomer is composed of 4 identical monomers, represented in light blue, light grey, wheat, and pale green for clarity; the monomer of aldolase A folds into an α/β-barrel with 10 anti-parallel β-sheets (represent in marine blue) bundled by 13 α-helices (represent in light blue); (d) Structure of tropomyosin (PDB: 1C1G), the homodimer of tropomyosin is comprised of 2 identically coiled subunits, represented in wheat and light grey for clarity; (e) Structure of Collagen I (PDB: 5CVB), the collagen triple helix was formed by association of 3 identical or different polypeptide chains, represented in wheat, light blue, and light grey for clarity.
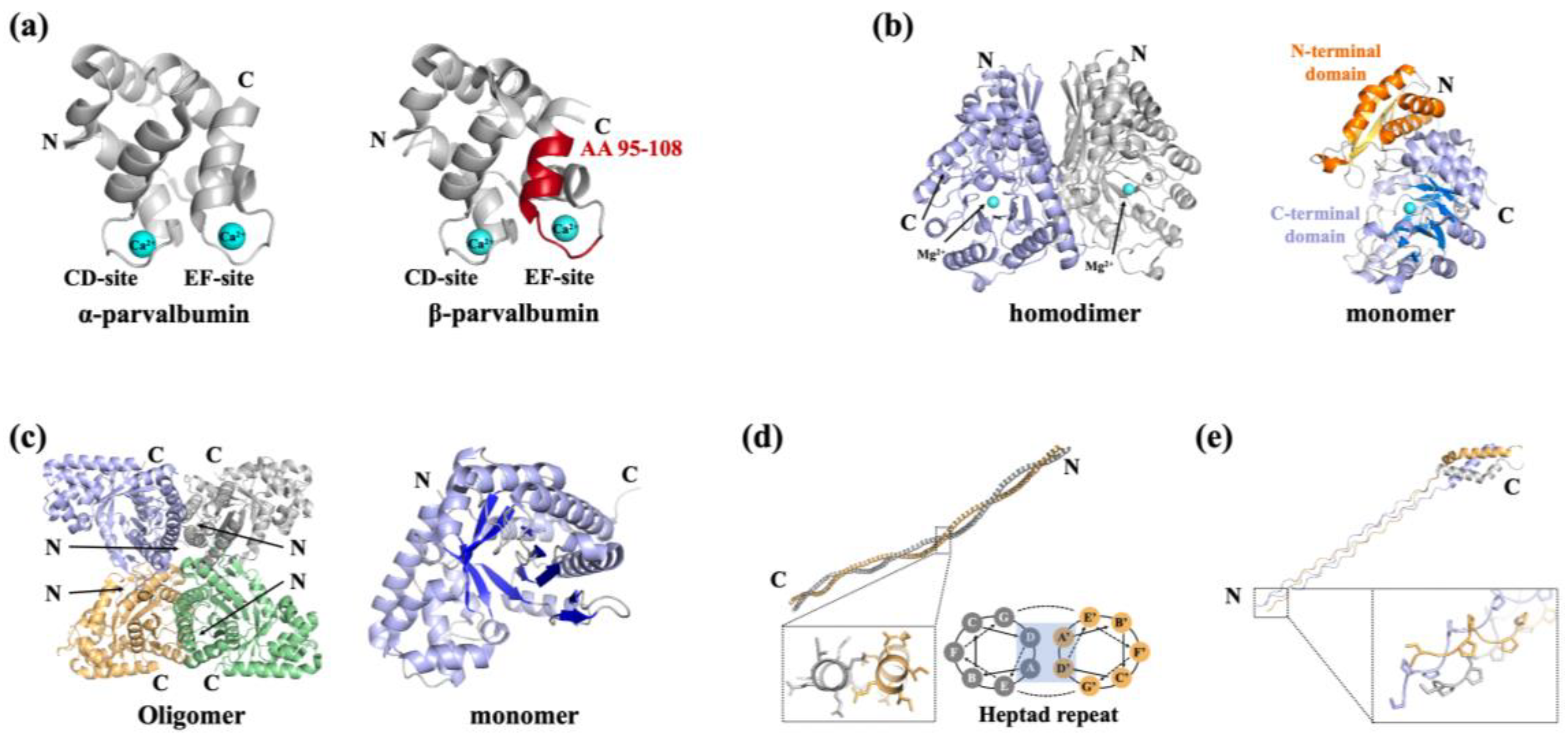
Figure 3.
Representative structures of shellfish allergens. (a) Structure of tropomyosin (Protein data bank, PDB: 1C1G) , the heat/digested stable epitopes of tropomyosin were colored red; (b) Structure of arginine kinase (PDB: 5ZHQ), the cysteines involved in the formation of disulfide bonds were colored red and the intramolecular disulfide bond was marked by indicated by the red dashed line; (c) Structure of myosins (PDB: 3I5F), the myosin is composed of a heavy chain and 2 light chains, represented in light grey, light blue, and orange, respectively; the heat/digested stable epitopes of myosin light chain 2 were colored red; (d) Structure of sarcoplasmic calcium binding protein (PDB: 7WBO), four bound Ca2+ were showed in cyan ball, the heat/digested stable epitopes of sarcoplasmic calcium binding protein were colored red; (e) Structure of triosephosphate isomerase (PDB: 5EYW), 8 central β-strands were colored light blue for clarity, the heat/digested stable epitopes of triosephosphate isomerase were colored red; (f) Structure of filamin C (partial, PDB: 7VZO), the β-sheets were colored light blue; (g) Structure of troponin C, a bound Ca2+ was showed in cyan ball.
Figure 3.
Representative structures of shellfish allergens. (a) Structure of tropomyosin (Protein data bank, PDB: 1C1G) , the heat/digested stable epitopes of tropomyosin were colored red; (b) Structure of arginine kinase (PDB: 5ZHQ), the cysteines involved in the formation of disulfide bonds were colored red and the intramolecular disulfide bond was marked by indicated by the red dashed line; (c) Structure of myosins (PDB: 3I5F), the myosin is composed of a heavy chain and 2 light chains, represented in light grey, light blue, and orange, respectively; the heat/digested stable epitopes of myosin light chain 2 were colored red; (d) Structure of sarcoplasmic calcium binding protein (PDB: 7WBO), four bound Ca2+ were showed in cyan ball, the heat/digested stable epitopes of sarcoplasmic calcium binding protein were colored red; (e) Structure of triosephosphate isomerase (PDB: 5EYW), 8 central β-strands were colored light blue for clarity, the heat/digested stable epitopes of triosephosphate isomerase were colored red; (f) Structure of filamin C (partial, PDB: 7VZO), the β-sheets were colored light blue; (g) Structure of troponin C, a bound Ca2+ was showed in cyan ball.
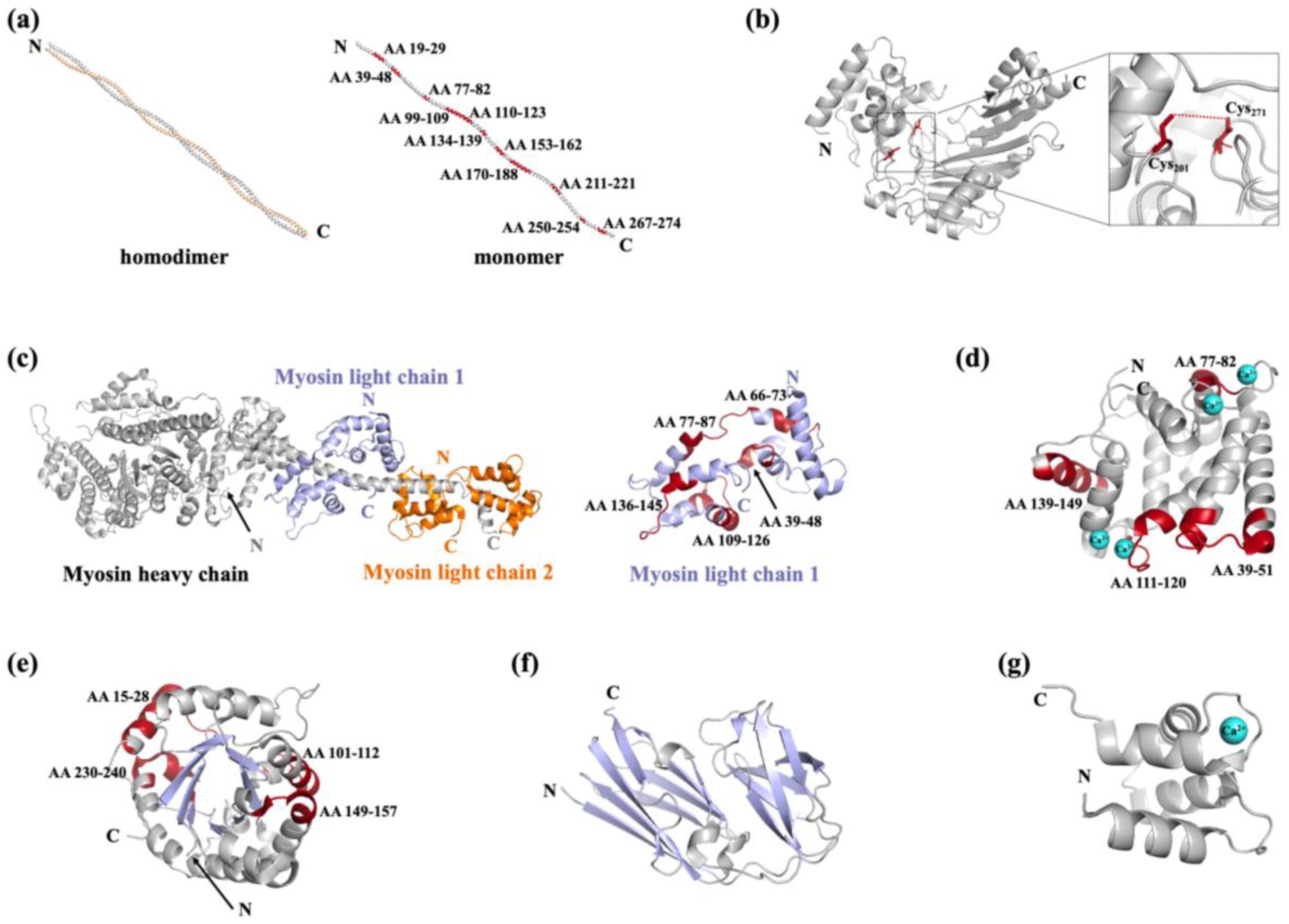
Figure 4.
Schematic representation of thermal processing methods on structure/epitope changes to seafood allergens. (a) Peptide bonds hydrolysis during thermal treatment; (b) heating-induced denaturation of allergen; (c) aggregation of allergens during heating; (d) the two-strand α-helical coiled-coil structure converted to a single-stranded α-helix during thermal processing. The epitopes were showed in dots with the destroyed/masked ones showed in grey.
Figure 4.
Schematic representation of thermal processing methods on structure/epitope changes to seafood allergens. (a) Peptide bonds hydrolysis during thermal treatment; (b) heating-induced denaturation of allergen; (c) aggregation of allergens during heating; (d) the two-strand α-helical coiled-coil structure converted to a single-stranded α-helix during thermal processing. The epitopes were showed in dots with the destroyed/masked ones showed in grey.
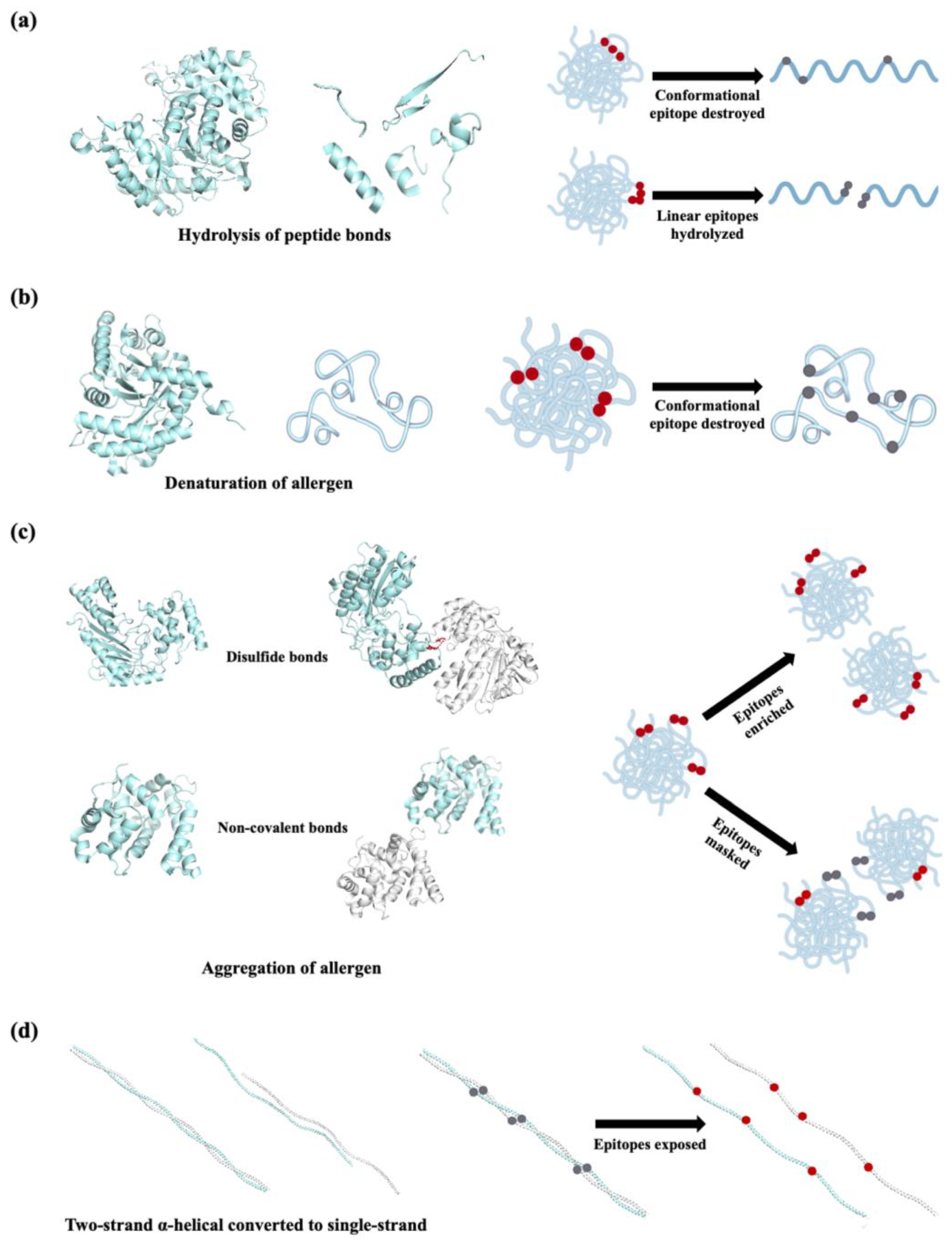
Figure 5.
Schematic representation of innovative non-thermal processing methods on structure changes to seafood allergens. (a) The secondary structure conversion during high pressure treatment; (b) the primary structure degradation and secondary structure conversion of allergen induced by high-intensity ultrasound treatment; (c) the carbonylation of amino acids in epitope during irradiation; (d) the allergenic proteins cross-linking with heat-sensitive proteins during pulsed ultraviolet light treatment; (e) the cleavage of allergen peptide bonds, oxidization of amino acid side residuals, and formation of the protein-protein cross-linkages during cold plasma treatment.
Figure 5.
Schematic representation of innovative non-thermal processing methods on structure changes to seafood allergens. (a) The secondary structure conversion during high pressure treatment; (b) the primary structure degradation and secondary structure conversion of allergen induced by high-intensity ultrasound treatment; (c) the carbonylation of amino acids in epitope during irradiation; (d) the allergenic proteins cross-linking with heat-sensitive proteins during pulsed ultraviolet light treatment; (e) the cleavage of allergen peptide bonds, oxidization of amino acid side residuals, and formation of the protein-protein cross-linkages during cold plasma treatment.
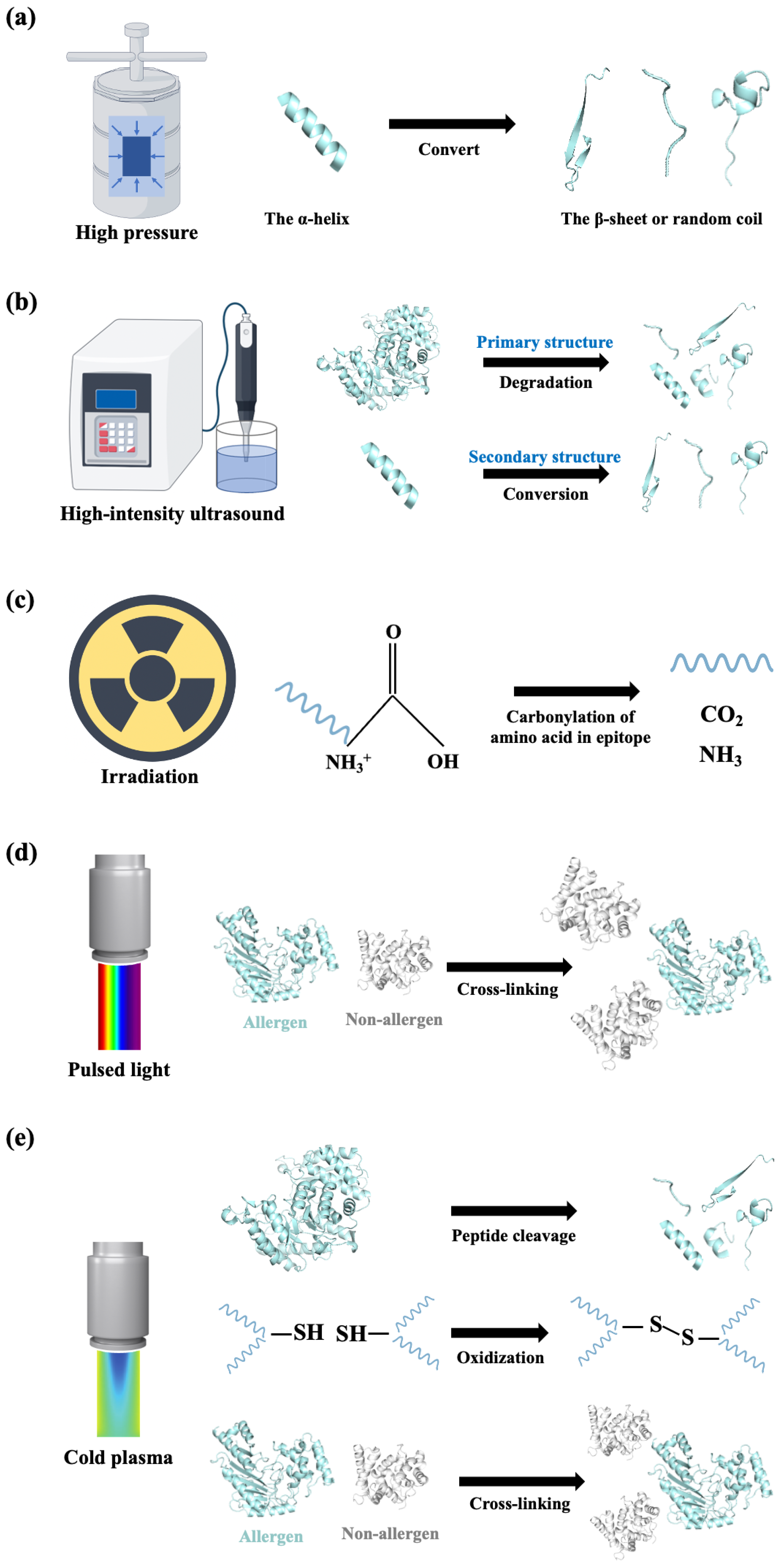
Figure 6.
Schematic diagram of the catalytic mechanism of tyrosinase (a), laccase (b), and transglutaminase (c). The different polypeptide chains were showed in wavy lines with different color.
Figure 6.
Schematic diagram of the catalytic mechanism of tyrosinase (a), laccase (b), and transglutaminase (c). The different polypeptide chains were showed in wavy lines with different color.
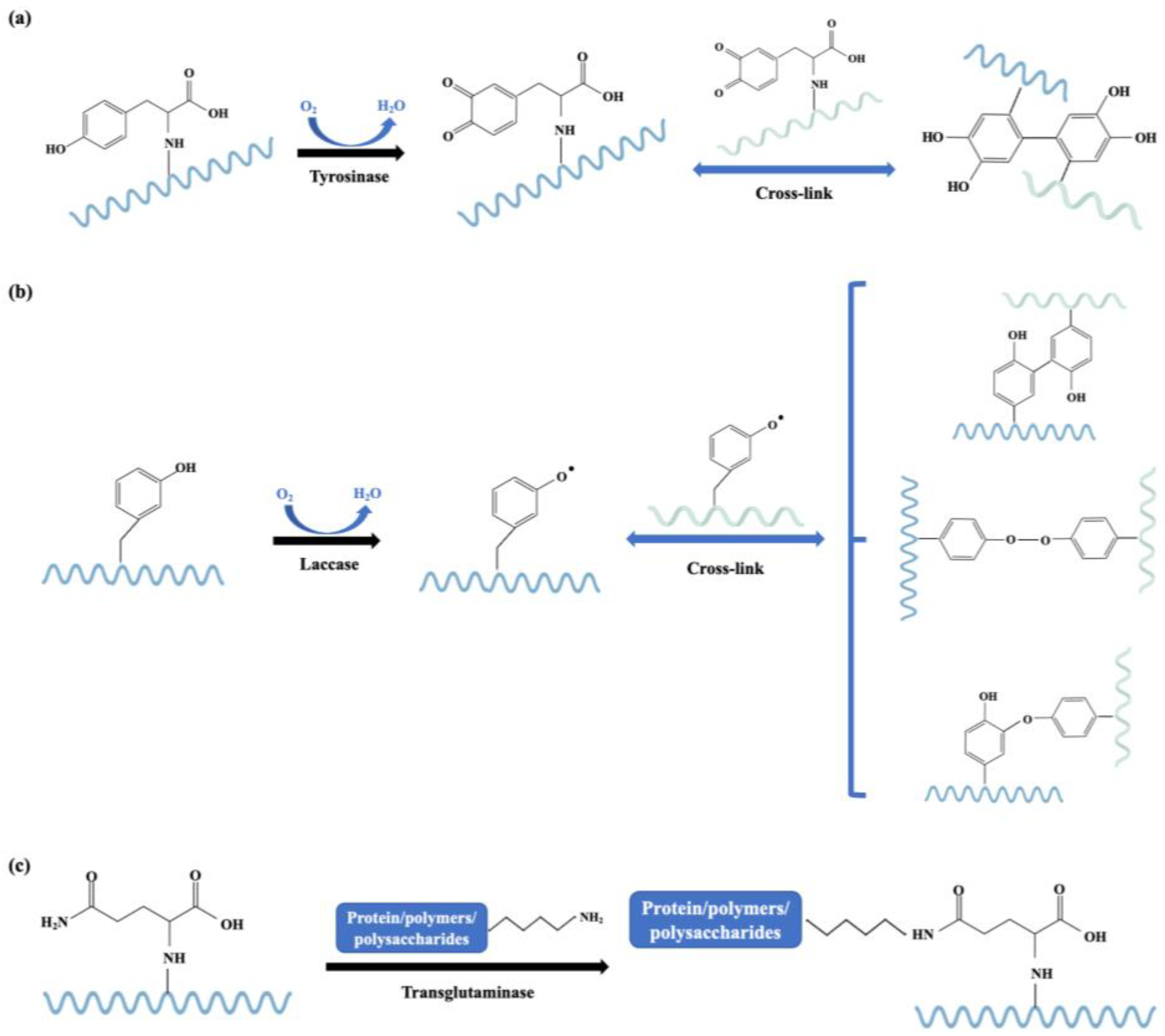
Figure 7.
The reaction principle of glycation modification. (a) Overview of the key steps of Maillard reaction, the side chains of potential amino acids for Maillard reaction were marked red; (b) overview of the key steps of enzymatic-catalyzed glycation illustrated by the reaction of amino sugar with a glutaminyl residue that catalyzed by transglutaminase.
Figure 7.
The reaction principle of glycation modification. (a) Overview of the key steps of Maillard reaction, the side chains of potential amino acids for Maillard reaction were marked red; (b) overview of the key steps of enzymatic-catalyzed glycation illustrated by the reaction of amino sugar with a glutaminyl residue that catalyzed by transglutaminase.
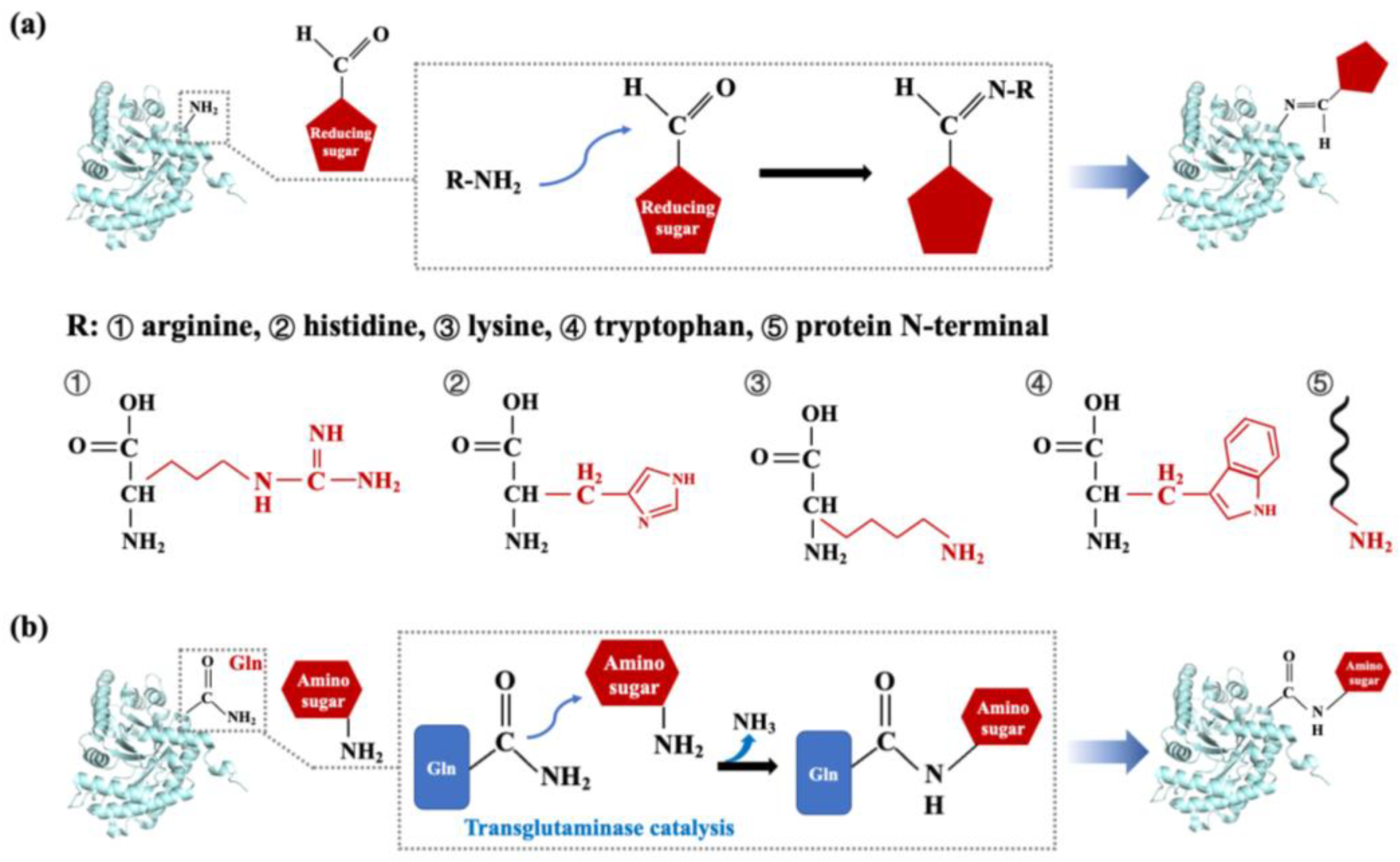

Table 1.
The allergens identified in fish (data from WHO/IUIS http://www.allergen.org).
Table 1.
The allergens identified in fish (data from WHO/IUIS http://www.allergen.org).
| Protein | Fish species | Allergen names in IUIS | MW (kDa) |
| β-Parvalbumin | Gadus callarias | Gad c 1 | 12 |
| Gadus morhua | Gad m 1 | 12 | |
| Crocodylus porosus | Cro p 1 | 11.6 | |
| Clupea harengus | Clu h 1 | 12 | |
| Ctenopharyngodon idella | Cten i 1 | 9 | |
| Cyprinus carpio | Cyp c 1 | 12 | |
| Lates calcarifer | Lat c 1 | 11.5 | |
| Lateolabrax maculatus | Late m 1 | 12 and 13 | |
| Lepidorhombus whiffiagonis | Lep w 1 | 11.5 | |
| Oncorhynchus mykiss | Onc m 1 | 12 | |
| Pangasianodon hypophthalmus | Pan h 1 | 11 | |
| Rastrelliger kanagurta | Ras k 1 | 11.3 | |
| Salmo salar | Sal s 1 | 12 | |
| Sardinops sagax | Sar sa 1 | 12 | |
| Scomber scombrus | Sco s 1 | 12 | |
| Sebastes marinus | Seb m 1 | 11 | |
| Solea solea | Sole s 1 | 11~14 | |
| Thunnus albacares | Thu a 1 | 11 | |
| Trichiurus lepturus | Ttic l 1 | 11 | |
| Xiphias gladius | Xip g 1 | 11.5 | |
| α-Parvalbumin | Crocodylus porosus | Cro p 2 | 13 |
| β-Enolase | Gadus morhua | Gad m 2 | 47.3 |
| Cyprinus carpio | Cyp c 2 | 47 | |
| Pangasianodon hypophthalmus | Pan h 2 | 50 | |
| Salmo salar | Sal s 2 | 47.3 | |
| Thunnus albacares | Thu a 2 | 50 | |
| Aldolase A | Gadus morhua | Gad m 3 | 40 |
| Pangasianodon hypophthalmus | Pan h 3 | 40 | |
| Salmo salar | Sal s 3 | 40 | |
| Thunnus albacares | Thu a 3 | 40 | |
| Tropomyosin | Salmo salar | Sal s 4 | 37 |
| Oreochromis mossambicus | Ore m 4 | 33 | |
| Pangasianodon hypophthalmus | Pan h 4 | 35 | |
| β-Prime-component of vitellogenin | Oncorhynchus keta | Onc k 5 | 18 |
| α-Collagen | Lates calcarifer | Lat c 6 | 130~1140 |
| Salmo salar | Sal s 6 | 130~140 | |
| Creatine kinase | Pangasianodon hypophthalmus | Pan h 7 | 43 |
| Salmo salar | Sal s 7 | 43 | |
| Triosephosphate isomerase | Pangasianodon hypophthalmus | Pan h 8 | 25 |
| Salmo salar | Sal s 8 | 25 | |
| Pyruvate kinase PKM-like | Pangasianodon hypophthalmus | Pan h 9 | 65 |
| L-lactate dehydrogenase | Pangasianodon hypophthalmus | Pan h 10 | 34 |
| Glucose 6-phosphate isomerase | Pangasianodon hypophthalmus | Pan h 11 | 60 |
| Glyceraldehyde-3-phosphate dehydrogenase | Pangasianodon hypophthalmus | Pan h 13 | 36 |
* The allergens were arranged in accordance with their allergen names in IUIS.
Table 2.
The allergens and their remarkable characteristics identified in fish and shellfish (data from WHO/IUIS http://www.allergen.org).
Table 2.
The allergens and their remarkable characteristics identified in fish and shellfish (data from WHO/IUIS http://www.allergen.org).
| Protein | MW (kDa) | Species | Biological function |
|
| β-Parvalbumin | 11~14 | Fish | Calcium-binding protein |
|
| α-Parvalbumin | ~13 | Fish | Calcium-binding protein |
|
| β-Enolase | 47~50 | Fish | Glycolytic enzyme |
|
| Aldolase A | 40 | Fish | Glycolytic enzyme |
|
| Tropomyosin | 33~40 | Fish Crustacean Mollusk |
Actin-binding protein |
|
| collagen | 130~140 | Fish | Structural protein |
|
| Pyruvate kinase PKM-like | 65 | Fish Crustacean |
A regulation enzyme involved in glycolysis |
|
| Triosephosphate isomerase | 25~28 | Fish Crustacean Mollusk |
A key enzyme involved in glycolysis |
|
| Creatine kinase | 43 | Fish | phosphoryl transfer enzymes |
|
| L-lactate dehydrogenase | 34 | Fish | An enzyme involved in glycolysis | - |
| Glucose 6-phosphate isomerase | 60 | Fish | A housekeeping enzyme of glycolysis and gluconeogenesis | - |
| Glyceraldehyde-3-phosphate dehydrogenase | 36 | Fish | - | - |
| β-Prime-component of vitellogenin | 18 | Fish | - | - |
| Arginine kinase | 38~45 | Crustacean Mollusk |
Phosphoryl transfer enzymes |
|
| Paramyosin | 99 | Mollusk | Actin-binding protein |
|
| Myosin light chain 2 | 18~23 | Crustacean | Actin-binding protein | - |
| Myosin light chain 1 | 17.5~18 | Crustacean |
Actin-binding protein |
|
| Sarcoplasmic calcium binding protein | 20~25 | Fish Crustacean |
Calcium-binding protein |
|
| Filamin C | ~90 | Crustacean | Actin-binding protein |
|
| Troponin C | 16.8~21 | Crustacean | Actin-binding protein |
|
| Troponin I | ~30 | Crustacean | Actin-binding protein | - |
| Hemocyanin | 76 | Crustacean | - |
|
| Ovary development-related protein | 28.2 | Crustacean | - | - |
| Cytoplasmic fatty acid binding protein | 15~20 | Crustacean | Lipid binding protein superfamily |
|
| Glycogen phosphorylase-like protein | 95 | Crustacean | - | - |
* The allergens were arranged in accordance with their mentioned order in the text.
Table 3.
The allergens identified in shellfish (data from WHO/IUIS http://www.allergen.org).
Table 3.
The allergens identified in shellfish (data from WHO/IUIS http://www.allergen.org).
| Protein | Shellfish species | Allergen names in IUIS | MW(kDa) |
| Tropomyosin | Charybdis feriatus | Cha f 1 | 34 |
| Crangon crangon | Cra c 1 | ~38 | |
| Crassostrea angulata | Cra a 1 | 38 | |
| Crassostrea gigas | Cra g 1 | 38 | |
| Exopalaemon modestus | Exo m 1 | 38 | |
| Haliotis laevigata x Haliotis rubra | Hal l 1 | 33.4 | |
| Helix aspersa | Hel as 1 | 36 | |
| Homarus americanus | Hom a 1 | 34 | |
| Litopenaeus vannamei | Lit v 1 | 36 | |
| Macrobrachium rosenbergii | Mac r 1 | 37 | |
| Melicertus latisulcatus | Mel l 1 | 38 | |
| Metapenaeus ensis | Met e 1 | 34 | |
| Pandalus borealis | Pan b 1 | 37 | |
| Panulirus stimpsoni | Pan s 1 | 34 | |
| Penaeus aztecus | Pen a 1 | 36 | |
| Penaeus indicus | Pen i 1 | 34 | |
| Penaeus monodon | Pen m 1 | 38 | |
| Portunus pelagicus | Por p 1 | 36 | |
| Procambarus clarkii | Pro c 1 | 36 | |
| Saccostrea glomerata | Sac g 1 | 38 | |
| Scylla paramamosain | Scy p 1 | 38 | |
| Todarodes pacificus | Tod p 1 | 15 | |
| Arginine kinase | Callinectes bellicosus | Cal b 2 | 40 |
| Crassostrea angulata | Cra a 2 | 38~41 | |
| Crangon crangon | Cra c 2 | ~45 | |
| Litopenaeus vannamei | Lit v 2 | 40 | |
| Macrobrachium rosenbergii | Mac r 2 | 40 | |
| Penaeus monodon | Pen m 2 | 40 | |
| Procambarus clarkii | Pro c 2 | 40 | |
| Scylla paramamosain | Scy p 2 | 40 | |
| Ovary development-related protein | Eriocheir sinensis | Eri s 2 | 28.2 |
| Paramyosin | Rapana venosa | Rap v 2 | 99 |
| Myosin light chain 2 | Homarus americanus | Hom a 3 | ~23 |
| Litopenaeus vannamei | Lit v 3 | 20 | |
| Penaeus monodon | Pen m 3 | 20 | |
| Scylla paramamosain | Scy p 3 | 18 | |
| Myosin light chain 1 | Artemia franciscana | Art fr 5 | ~17.5 |
| Crangon crangon | Cra c 5 | ~17.5 | |
| Procambarus clarkii | Pro c 5 | 18 | |
| Sarcoplasmic calcium binding protein | Crangon crangon | Cra c 4 | ~25 |
| Crassostrea angulata | Cra a 4 | 20 | |
| Litopenaeus vannamei | Lit v 4 | 20 | |
| Penaeus monodon | Pen m 4 | 20 | |
| Pontastacus leptodactylus | Pon l 4 | ~24 | |
| Portunus trituberculatus | Por t 4 | 22 | |
| Scylla paramamosain | Scy p 4 | 20 | |
| Troponin C | Crangon crangon | Cra c 6 | ~21 |
| Homarus americanus | Hom a 6 | ~20 | |
| Penaeus monodon | Pen m 6 | 16.8 | |
| Troponin I | Pontastacus leptodactylus | Pon l 7 | ~30 |
| Hemocyanin | Penaeus monodon | Pen m 7 | 76 |
| Triosephosphate isomerase | Archaeopotamobius sibiriensis | Arc s 8 | ~28 |
| Crangon crangon | Cra c 8 | ~28 | |
| Penaeus monodon | Pen m 8 | 27 | |
| Procambarus clarkii | Pro c 8 | 28 | |
| Scylla paramamosain | Scy p 8 | 28 | |
| Filamin C | Scylla paramamosain | Scy p 9 | 90 |
| Cytoplasmic fatty acid binding protein | Litopenaeus vannamei | Lit v 13 | 15 |
| Penaeus monodon | Pen m 13 | 20 | |
| Glycogen phosphorylase-like protein | Penaeus monodon | Pen m 14 | 95 |
* The allergens were arranged in accordance with their allergen names in IUIS.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



