Submitted:
20 May 2025
Posted:
21 May 2025
You are already at the latest version
Abstract
Galectin-3 is a β-galactoside-binding lectin that mediates diverse signaling events in multiple cell types, including immune cells. It is also a biomarker for multiple clinically important disorders, including cardiovascular disease. Galectin-3 binds to cell surface glycans to form lattices that modulate surface receptor signaling and internalization. However, the tissue-specific regulation of galectin-3 surface expression remains poorly understood. Here, we review evidence for the involvement of galectin-3 in cell surface signaling, intranuclear events, and intracellular trafficking. Our focus will be on the O-GlcNAc modification as a regulator of galectin-3 biosynthesis, non-canonical secretion, and recycling. We argue that the nutrient-driven cytoplasmic hexosamine biosynthetic pathway (HBP) and endomembrane transport mechanisms generate unique pools of nucleotide sugars. The differing levels of nucleotide sugars in the cytosol, endoplasmic reticulum (ER), and Golgi apparatus generate differential thresholds for responsiveness of O-GlcNAc cycling, N- and O- linked glycan synthesis/branching, and glycolipid synthesis. By regulating galectin-3 synthesis and non-canonical secretion, O-GlcNAc cycling may serve as a nexus constraining galectin-3 cell surface expression and lattice formation. This homeostatic feedback mechanism would be critical under conditions where extensive glycan synthesis and branching in the endomembrane system and cell surface are maintained by elevated hexosamine synthesis. Thus, O-GlcNAc cycling and galectin-3 synergize to regulate galectin-3 secretion and influence cellular signaling. In humans, galectin-3 serves as an early-stage diagnostic biomarker for heart disease, kidney disease, viral infection, autoimmune disease, and neurodegenerative disorders. Since O-GlcNAc cycling has also been linked to these pathologic states, exploring the interconnections between O-GlcNAc cycling and galectin-3 expression and synthesis is likely to emerge as an exciting area of research.
Keywords:
HBP
; UDP-GlcNAc
; N-Glycan
; O-GlcNAcylation
; Galectin-3
1. Overview of Galectin-3; A Unique β-Galactoside Binding Lectin
Lectins are classified into families which include the galectins, an ancient and particularly interesting family. All members of the galectin family were numbered by order of discovery. Galectins are defined by evolutionarily conserved amino acid sequences, and by their ability to recognize β-galactoside structures [1]. Galectins are omnipresent in sponges, fungi, nematodes, insects, vertebrates including mammals, and viral galectins have even been identified [1]. Their homologs are also present in plants, but not in yeast. To date, 16 mammalian galectins have been identified, each featuring a conserved carbohydrate-recognition-binding domain (CRD) comprised of about 130 amino acids [2]. Galectins are classified into three subgroups based on the number and structural organization of their carbohydrate recognition domains (CRDs), in addition to their functional characteristics. The first subgroup includes the non-covalently associated homodimer proto-type galectins (Gal-1, -2, -5, -7, -10, -11, -13, -14, -15 and -16), which contain a single CRD that forms non-covalent homodimers (Figure 1A) [2]. The second subgroup includes the tandem-repeat galectins (galectin-4, -6, -8, -9, and -12), and these heterodimers carry two distinct, but homologous, CRD motifs separated by a peptide linker sequence of up to 70 amino acids (Figure 1B) [2]. Lastly, the third subgroup is the tri-modular chimera containing a single canonical CRD connected to an unusually long, intrinsically disordered, N-terminal proline- and glycine-rich domain. This third subgroup is uniquely represented by galectin-3 (Gal-3) (Figure 1C) [2]. Galectin-3, initially named CBP35, is the only galectin discovered in vertebrates that can pentamerize (Figure 1C). It is also among the most intensively studied galectins [3]. It exerts distinct pleiotropic effects, contributing critically to a wide range of physiological and pathological processes [4]. Galectins are soluble proteins with no transmembrane domains. Secreted galectins remain tethered with the cell surface through interactions with cell-surface glycoproteins or glycolipids. Remarkably, these secreted galectins, particularly the multivalent galectin-3, can mediate their functions by forming ordered interactions with multivalent glycan ligand structures, often termed as galectin lattices (Figure 1D) [5]. The formation of these lattices is thought to be a major contributor to cellular signaling [5,6].
2. Tissue Distribution of Galectin-3 and Associations with Human Disease
Galectin-3 recognizes the distinct galactose β1-4 N-acetylglucosamine (N-acetyllactosamine) linkage present in both N- and O-glycans, glycolipids, and blood group antigens (Figure 2A to 2D). Galectin-3 is encoded by the LGALS3 gene, and its expression has been detected in many different cell types. These cell types include small intestinal epithelial cells, corneal and conjunctival epithelia, olfactory epithelia, colonic epithelia, epithelial cells of the kidney, lung, breast, thymus, and prostate, ductal cells of the salivary glands, kidney, eye, and pancreas, intrahepatic bile ducts, fibroblasts, chondrocytes, osteoclasts, osteoblasts, keratinocytes, Schwann cells, gastric mucosa, and endothelial cells from various tissues and organs. LGALS3 is also expressed in cell types involved in the immune response, such as neutrophils, eosinophils, basophils and mast cells, Langerhans cells, dendritic cells, as well as monocytes and macrophages from different tissues, but not in lymph nodes or spleen (Figure 3) [2,4]. During embryogenesis galectin-3 expression is regulated in a tissue-specific and temporally dynamic manner. During the early stages of embryogenesis, galectin-3 expression is more restricted, and primarily localized in the epithelial tissues, the developing kidney, chondrocytes, and liver [2,7]. Intriguingly, galectin-3-knockout mice are viable, and show no apparent abnormalities, except for premature senescence [8,9].
In terms of intracellular distribution, although galectin-3 is predominantly located in the cytoplasm, it can shuttle between the cytoplasm and nucleus, a pathway involving import and export signals at the CRD and N-terminal tail (NT) [10,11] (See discussion below). Ligands available for galectin-3 are expressed ubiquitously throughout the cell and can be subclassified based on their localization [4]. Nuclear ligands include: Gemin-4, Gemin-2, Gemin-3, ALG-2 linked protein x (Alix) or ALG-2 interacting protein-1 (AIP-1), and survival of motor neuron (SMN) protein. These ligands regulate gene transcription and promote pre-RNA splicing [12,13] [7], but galectin-3 does not interact with RNA directly [14]. Intracellular ligands for galectin-3 include Cytokeratins, CBP70, Chrp, Gemin-4, Alix/AIP-I, and Bcl-2. These ligands play a key role in cell survival. Their interaction with galectin-3 is a protein-protein interaction rather than a lectin-conjugation interaction [12,13,15,16]. Extracellular galectin-3 ligands include laminin, fibronectin, tenascin, chondroitin sulfate, Mac-2 binding protein, and some cell adhesion small molecules like integrins. These ligands modulate cell-cell interactions, cell differentiation, inflammation, fibrogenesis, and the host defense [7,17,18,19,20,21].
The multifunctional protein galectin-3 is expressed in many different tissues, engages in multiple biological events, and has been suggested to play a pivotal role in numerous diseases and clinical conditions. On average, more than 500 publications are published each year on galectin-3, and the numbers are constantly increasing [https://pubmed.ncbi.nlm.nih.gov/?term=galectin-3&filter=datesearch.y_1]. This large number of publications, many of which in the most recent year discuss the role of galectin-3 in clinical conditions and diseases, attest to its growing potential relevance to human pathology. Galectin-3 is a widely used biomarker whose protein levels have been utilized as a marker for many diseases. These disorders include asthma, atherosclerosis, atopic dermatitis, cerebral infarction, and chronic obstructive pulmonary disease (COPD). In addition, galectin-3 has been used as a biomarker for the following: aortic stenosis, endometriosis, enteric nervous system, encephalitis, fibrosis, gastritis, HIV infection, inflammation, interstitial lung diseases, juvenile idiopathic arthritis, liver fibrosis, non-alcoholic fatty liver disease (NASH), obesity, pneumonia, pulmonary hypertension, plaque psoriasis, Q fever, rheumatoid arthritis, sepsis, systematic sclerosis, urinary tract infection, venomous thrombosis, and especially heart failure, renal diseases, diabetes mellitus and cancers [4,7]. As a biomarker, it is worth noting that galectin-3 has not been causally linked to any of these disorders, but can serve as a measurable indicator of disease progression.
Given that galectin-3 is a biomarker for fibrosis, inflammation, and immunity, it has been extensively investigated for potential involvement in fibrotic diseases such as heart disease and kidney disease. Galectin-3 is a possible therapeutic approach to slow the progression of these fibrotic diseases, which may advance overtime into more severe heart failure (HF) and renal failure [7]. While not recommended on its own as a prognostic biomarker for individuals with HF, galectin-3 does show prognostic value in combination with other already established HF biomarkers [22]. Notably, elevated galectin-3 levels have been found in nearly all types of cardiovascular disease, not just HF. Elevated serum galectin-3 levels were associated with cardiovascular mortality and a greater incidence of cardiovascular events in patients with coronary heart disease [23,24]. Moreover, galectin-3 is also associated with new-onset atrial fibrillation, myocardial infarction (MI) size, and left ventricular remodeling in patients with a history of MI [25,26,27]. In addition to its role in cardiovascular disease, galectin-3 is also implicated in renal failure and associated complications [28]. Increased galectin-3 expression was identified in both the serum and tumor tissues of renal cell carcinoma patients, as well as in patients with chronic kidney disease [29,30,31,32]. Furthermore, galectin-3 mRNA expression increased in folic-acid induced acute renal failure in a rat model [28].
3. Galectin-3 Structure, Conservation, and Function
The N-terminal domain (NTD) in galectin-3 is composed of 110 to 130 amino acids depending on the species. This relatively flexible structure contains multiple repeats, each of which includes a consensus sequence Pro–Gly–Ala–Tyr–Pro–Gly, followed by three additional amino acids [5]. The NTD is highly conserved among galectin-3 molecules isolated from different species. The initial 12 amino acid N-terminal peptide sequence preceding the proline/glycine-rich repetitive domain, also called the small N-terminal domain, is highly conserved in all mammalian galectin-3. In the case of human galectin-3, the 21-amino-acid-long NTD with its two sites for serine phosphorylation are followed by nine non-triple-helical collagen-like Pro/Gly-rich repeats (I-IX), which harbor cleavage sites for diverse proteases [19]. The NTD and the nine repeats form the N-terminal tail (NT). At least two functional characteristics are ascribed to this NT of galectin-3: deletion of these first 11 amino acids (following the first methionine) blocks secretion of galectin-3 [32], while mutation of the conserved Ser6 affects galectin-3 anti-apoptotic signaling activity [33]. The NTD has also been implicated in secretion of galectin-3 outside of cells [34]. In full-length galectin-3, the NTD is connected to the C-terminal carbohydrate recognition domain (CRD), composed of about 130 amino acids, forming a globular structure responsible for the lectin activity of galectin-3 [35,36]. It is evident from nuclear magnetic resonance (NMR) experiments that these three sections, i.e., NTD, collagen-like repeats, and CRD, likely cooperate in a not yet clearly defined manner to account for their special role within the galectin network [37].
4. Galectin-3 Nuclear Import and Export
Intracellular trafficking of galectin-3 is complex, and some reports of its cell biology are contradictory. Analysis by in vitro nuclear import assays using digitonin-permeabilized cells revealed that the galectin-3 protein could be imported into the nucleus by either passive diffusion, N-terminal region-dependent active transport, or importin-α/β complex-dependent transport [11,33]. Site directed mutagenesis and truncation of full-length galectin-3 suggested the CRD of the galectin-3 polypeptide is essential for nuclear localization [10,34]. Transfections of serine-to-alanine (S6A) and serine-to-glutamic acid (S6E) galectin-3 mutants in the BT-549 human breast carcinoma cell line revealed that Serine6 phosphorylation by casein kinase 1 (CK1) in the NTD favors export from the nucleus [35,36]. Secretion to the extracellular environment proceeds via an endoplasmic reticulum (ER)-Golgi complex independent, non-classical, pathway which involves the NTD and segments of the collagen-like repeat region [36,37,38,39]. Galectin-3 is transported to the early/recycling endosomes and is then partitioned into two routes: recycling back to the plasma membrane or targeting to the late endosomes/lysosomes. Systematic comparison among various N- and C-terminal truncated forms of galectin-3 with the full-length protein showed that the CRD of galectin-3, along with the NTD and collagen repeating units, are required for galectin-3 binding and endocytosis. Neither the CRD nor the N-terminal half of the protein, which comprises the NTD and the collagen-like internal repeating domain, can mediate binding and endocytosis alone. Although the collagen-like domain is largely irrelevant to galectin-3 trafficking to the early/recycling endosomes, it is required for targeting galectin-3 to the late endosomes/lysosomes [40].
The biology of galectin-3, its application as an omnipotent biomarker, and the fact that it is the ‘alarm protein’ in the hierarchy of immune responses, has already been well studied and reviewed [4,7,41,42,43,44,45,46,47,48,49]. In this review, we’ll focus on the regulation of the galectin-3 transcriptional and secretory mechanism by hexosamine biosynthetic flux, and therefore protein or lipid glycosylation.
5. Galectin-3 Trafficking and Secretion
Mechanisms for the cellular export of galectins are not clearly understood. Galectin-3 circumvents the classical “endoplasmic reticulum-Golgi apparatus” pathway [39], and is secreted via a non-canonical mechanism (Figure 4). Research on the galectin-3 secretory mechanism has pointed to different routes for secretion, but it is possible that secretion can employ more than one pathway [50]. To date, four distinct non-conventional transport pathway mechanisms have been described as potential explanations of how these unconventionally secreted proteins leave the cell [51]. The first one is direct translocation of the protein across the plasma membrane, as is best studied in the fibroblast growth factor 2 (FGF-2). The second pathway for unconventional protein secretion involves the sequestration of a soluble cytoplasmic factor by secretory lysosomes, as is shown for interleukin 1b (IL1b). Vesicle-mediated secretion by micro vesicles or exosomes cover the third unconventional protein secretion pathway [50]. Transmembrane proteins which are inserted in the ER membrane, and reach the plasma membrane without passing through the Golgi, typically constitute the fourth type of pathway [52]. Findings from Mehul and Hughes indicate the third pathway’s implication for galectin-3, as they describe that an acylated galectin-3/chloramphenicol acyltransferase (CAT) chimera is released by microvesicle shedding in COS-7 cells [38]. Additionally, several proteomic approaches have led to the identification of galectin-3 in exosomes derived from different cell types [53,54,55,56]. Thus, it seems that galectin-3 uses distinct pathways for unconventional secretion, but this may differ for each individual cell type. However, the exact secretion mechanism of galectin-3 has not been identified for any of these pathways. Nevertheless, the Hughes lab was able to demonstrate that a short octapeptide in the N-terminus of galectin-3 was essential, but not sufficient, for unconventional secretion of an acylated fusion protein of the chloramphenicol acyltransferase (CAT) with N-terminal residues 1–120 of hamster galectin-3 [39]. Many factors are known to influence galectin-3 secretion, e.g., heat shock, calcium ionophores, acylation, phosphorylation and O-GlcNAcylation [37,38,57,58]. Intracellularly, phosphorylation and importin-mediated mechanisms appear to be involved in nucleo-cytoplasmic shuttling of galectin-3 [35,59,60,61], while synexin-mediated mechanisms are indicated in galectin-3 translocation to the mitochondria [62]. It has been reported that deletion of the first twelve residues in the NTD of galectin-3 blocks the export of galectin- 3 into the cell supernatant [36]. Delacour et al. propose that it is not the NTD itself, but rather oligomerization, that is necessary for galectin-3 secretion, as the first twelve residues of the NTD in galectin-3 are responsible for oligomerization of galectin-3 [63].
6. Galectin-3 and Cell Surface Signaling via Lattice Formation
One of the important functions galectins play extracellularly is in the organizing of the cell membrane via the formation of macroscale interaction networks, often referred to as galectin lattices [4]. Galectin-3 is especially important in organizing the galectin lattice, as the multivalence of its pentameric form allows large networks of galectins to form [3]. In solution, galectin-3 is monomeric unless high enough concentrations are reached, or bi- to multivalent ligands are present that serve as a core for aggregation [35,36,42,67,68,69,70,71,72,73,74,75]. The mode of galectin-3 functional oligomerization is a conundrum. Depending on which domain is involved, the interactions are categorized as type-N (6, 9, 12–14) or type-C (15–18) for the NTD- or CRD-mediated self-associations. Both the NT and the CRD play a functional role in this intermolecular assembly to varying extents [76,77,78,79]. Galectin and glycan interactions have been shown to play important roles in clathrin-independent endocytosis (CIE) [4]. Unlike clathrin-mediated endocytosis, which is well characterized with extensive intracellular machinery identified [80], CIE is still poorly understood, with little known about what cytoplasmic components regulate it [81,82,83,84]. CIE cargo, like most cell membrane bound proteins, are glycosylated [85]. Attempts have been made to identify both intracellular regulators of CIE, as well as plausible modulation on the extracellular side of the membrane via well-established galectin-glycan interactions. The effect of galectin-glycan interactions have been observed to have both a stimulatory role as well as an inhibitory role on CIE. The stimulatory role was shown by the Johannes group, and is due to initiation of direct interactions between galectin-3 and gangliosides (GSLs), which induce membrane bending on protein-free giant unilamellar vesicles (GUVs) and raft-dependent endocytosis or pit formation through clathrin-independent carriers (CLICs) in cells [86,87]. The pentameric conformation of galectin-3, which is induced by high-density glycolipid ligands, might have geometric features that promote GSL clustering, membrane bending, and raft-dependent endocytosis [86]. Galectin-3 was shown to promote the formation of endocytic pits resulting in stimulated internalization of the hyaluronan receptor (CD44) [86,87,88]. Galectin 3 was also reported to stimulate the internalization of β-1 integrin by CIE [89]. In contrast, glycan-galectin interactions have also been demonstrated to impede internalization via galectin lattice–mediated cell-surface sequestration in the CIE of the epidermal growth factor receptor [90,91,92,93,94,95]. CD59 and MHC-I like CIE cargo are sensitive to changes in galectin-glycan profiles, and interactions modulated by metabolic flux and nutrient availability [96].
These two modes of action act in opposing directions. There have been hints that these two functions may not be mutually exclusive, and could rather represent two ends of a spectrum [64], as illustrated in Figure 4. Galectin-3 induced effects occur on a continuum from stimulatory to inhibitory, with distinct CIE cargo proteins having unique responses. In addition, different cell types may exist at different positions on this continuum [65]. In polarized epithelial Madin-Darby canine kidney (MDCK) cells, galectin-3 was found to be endocytosed at the apical membrane, following activation of a non-raft-dependent pathway, to recycling endosomes in a lactose and pH dependent manner [66,67]. Further interactions of the internalized galectin-3 with glycosylated cargo in apical recycling endosomes sort glycoproteins and perturbate them to the apical membrane [66,68]. While export via exosomes has been proposed as a means of galectin-3 secretion [69], a recent genome-wide CRISPR screen for galectin-3 secretory machinery demonstrated that secreted galectin-3 was primarily free, and not exosome-bound, suggesting that exosomal export was not the primary secretory pathway [70].
A relatively new model termed the ‘desialylation glycoswitch’ has been introduced by Johannes and coworkers [71]. These findings propose an epidermal growth factor receptor (EGFR)-controlled reversible sialic acid (Sia) glycan-based regulatory circuit, which through EGF-induced desialylation and Gal3-dependent retrograde trafficking of plasma membrane glycoproteins, provides access to the Golgi apparatus to reset glycoprotein sialylation state. In this model, EGF-induced CIE of galectin-3 occurs through the triggered removal of terminal sialic acid residues from N-glycan cargo client proteins, and extracellular cues are relayed to galectin-driven endocytic processes and transduced into pathophysiological responses through the acute remodeling of cell surface glycans. They have established that EGFR, and its tyrosine kinase activity required for EGF-induced galectin-3 binding, are directly dependent on the presence of sialic acid residues and their removal by membrane sialidases (neuraminidases, Neu). Following EGF-induced desialylation through extracellular acidification of Neu1 and Neu3 by sodium-proton antiporter NHE1, integrins are transported via the retrograde route to the Golgi apparatus. Here the glycan makeup of the integrins are reset by resialylation, and their functions are repurposed by polarized secretion to the leading edge to support invasive cell migration. Galectin-3 trafficking, therefore, comprises non-canonical secretion from the cytoplasm as well as glycan-dependent raft endocytosis to recycling endosomes, where it plays significant roles in post-Golgi sorting.
7. Hexosamine Biosynthetic Pathway (HBP) and Galectin-3
The hexosamine biosynthetic pathway (HBP) is a glucose metabolic pathway that branches from the main glycolytic pathway, and it is essential for amino sugar synthesis. The HBP integrates the key metabolic pathways of glucose, amino acids, nucleosides, and lipids, thereby serving as a nutrient sensor. Cells monitor nutrient availability via HBP flux under physiological and pathological conditions, even though the HBP only utilizes about 2% to 5% of the glucose taken up by cultured cells. The end-product of the HBP is UDP-GlcNAc, a high-energy nucleotide sugar donor substrate used for the biosynthesis of glycoproteins (both O-Glycans and N-glycans), glycolipids, proteoglycans, and glycosaminoglycans. UDP-GlcNAc cellular dynamics are strictly regulated by mechanisms involved in multistep enzyme catalysis. The HBP is not only an essential pathway for UDP-GlcNAc synthesis, but also an essential conduit regulating the biosynthesis of other nucleotide sugars. The end-product of the HBP, UDP-GlcNAc, is further epimerized by GALE to generate UDP-GalNAc which serves as a donor substrate for biosynthesis of mucin-type O-glycoproteins, complex glycolipids, and proteoglycans [72]. Additionally, UDP-GlcNAc is metabolized by GNE to produce ManNAc, and hence sialic acid, mannose, and fucose. Their corresponding nucleotide sugars: CMP-sialic acid, GDP-mannose, and GDP-fucose respectively, are essential donor substrates for biosynthesis of N-glycans, glycolipids, proteoglycans, and glycosaminoglycans. The enzymatic steps involved in nucleotide sugar biosynthesis are summarized in Figure 5. An altered glucose or GlcNAc flux alters the UDP-GlcNAc concentration through the HBP, resulting in alteration in glycoprotein conformation, oligomerization, trafficking, localization, turnover, and stability.
8. N-Glycan Branching and Galectin-3
Galectin-3 is evolutionarily conserved and is ubiquitously expressed with a distinct binding partner galactose β1-4 N-acetylglucosamine (N-acetyllactosamine) linkage [1]. This N-acetyllactosamine linkage is a recurring sequence in branched N-glycans and the major family of galectin ligands at the cell surface [6,73,74]. In vertebrates, the secretory pathway translates ∼30% of the transcriptome, and on the luminal side of the membrane, most of the proteins are modified by the N-glycosylation pathway. Above 97% of N-glycans are attached at the NXS/T (X ≠ Pro) motif within mammalian glycoproteins [75], and they are transferred from the glycolipid Glc3Man9GlcNAc2-pyrophosphate-dolichol to an asparagine (Asn) residue of the nascently synthesized protein by the oligosaccharyltransferase complex (OST) [76]. This N-glycan is a ligand for the ER chaperone proteins like calnexin and calreticulin, which promote protein folding efficiency, protein quality control, and secretion or degradation of misfolded proteins [77,78]. After initial folding in the ER, where the three glucose (Glc) residues are consecutively trimmed by glucosidases, most glycoproteins transition to the Golgi apparatus. Here, trimming of the mannose (Man) residues by mannosidase I and II (Man I and Man II) takes place successively, forming the[77,78 Man5GlcNAc2–Asn core structure [76,79]. This Man5GlcNAc2–Asn structure is further remodeled by branching N-acetylglucosaminyltransferases, which form an ordered enzyme cascade that comprises Mgat1, Mgat2, Mgat4a/b/c and Mgat5 [80]. Each Mgat enzyme transfers one GlcNAc unit in a specific linkage to N-glycans which is followed by the consecutive addition of a β-linked galactose. Therefore, each branch is a potential ligand for galectin (Galactose β1-4 N-acetylglucosamine i.e., N-acetyllactosamine i.e LacNAc). Although the affinity of the N-glycans for galectins increases with branching and poly-N-acetyllactosamine extensions (repeating Galβ1,4GlcNAcβ-units) [81], lattice formation is a complex process that is not simply a function of binding affinities between galectins and N-glycans. A single glycoprotein with three N-glycans and nine N-acetyllactosamine (LacNAc) units shows a 1000-fold range of galectin-3 binding constants [82]. In addition, galectin-3 oligomerizes through its N-terminal domain and forms a lattice that has been visualized on the cell surface, and that varies with cell state [83].
Galectins bind to glycan motifs found on many glycoconjugates at the cell surface, and in this regard, they may appear to lack specificity for individual effectors. Indeed, proteomic analyses of galectin-3-interacting partners like Mgat5-dependent raft association, and LPHA-binding proteins (the higher affinity galectin ligands), indicate binding of many glycoproteins in parallel [64,84,85]. The fact that the number of N-glycans and the degree of branching are sensitive to UDP-GlcNAc flux and its galectin-3 affinity to regulate cell proliferation and differentiation was well studied by Lau et. al. [73]. Their systematic computational results and experimental data with cell lines fit this model linked the evolutionary origins of the N-glycan multiplicity in the growth factor receptor (GFRs) glycoproteins regulate with metabolic flux through the hexosamine pathway that regulates N-glycan branching in the Golgi apparatus. They showed that variations in the hexosamine flux through exogenous addition of GlcNAc, especially at hyper physiological concentrations, affect the residence time of receptors on the cell surface by modulating the interactions of branched N-glycans with the galectin-3 lattice. The relative affinity of transmembrane glycoproteins for the galectin lattice is proportional to their number of Asn-X-Ser/Thr (NXS/T, X ≠ Pro) N-glycan sites, as well as modification through the Golgi N-glycan branching pathway. Glycoproteins with high numbers of N-glycans have higher avidities for the galectin-3 lattice compared to glycoproteins with fewer N-glycans. Glycoforms generated in the Golgi complex for stable association with the galectin-3 lattice are above and below affinity threshold respectively. The Golgi pathway is ultrasensitive to hexosamine flux, and the pathway produces tri- and tetra-antennary N-glycans in glycoproteins with high numbers of N-glycans. These N-glycans bind to the galectin-3 lattice, restricting glycoprotein endocytosis, and hindering the downregulation of signaling [73].
With a change in UDP-GlcNAc concentration there is a subtle change in N-glycan branching. The enzymes involved in N-glycan branching show an intriguing pattern in their recognition of their substrates. The branching enzymes (Mgat1 to Mgat5) show a different binding affinity for their common substrate UDP-GlcNAc (∼300-fold). The Michaelis constant (Km) changes from 0.04 to 11 mM for Mgat1 and Mgat5 respectively. Mgat1 has a low affinity for the acceptor N-glycan at ~2 mM, and this relationship is reversed for Mgat4 and Mgat5 (Figure 5). With Mgat1 functioning near saturation and Mgat4 and Mgat5 operating well below their Km, this makes the synthesis of tri- and tetra-antennary glycans highly sensitive to the supply of this metabolite [73]. Computational modeling which considered branching as a ‘‘linear pathway’’ with no removal of the intermediate products, and an experimental setup with Mgat5 knockout cell lines under high GlcNAc concentrations, led to the identification that the sequential decreasing efficiency of Golgi enzyme reactions, and the removal of intermediate products from the Golgi compartment, are the two major conditions required for N-glycan branching ultra sensitivity. However, all the experiments were conducted with hyper physiological concentrations of GlcNAc, and hence UDP-GlcNAc, which is much higher than that required to fully activate O-GlcNAc. Estimates suggest that if the ER and Golgi concentrate UDP-GlcNAc from 10–30-fold, the concentration of UDP-GlcNAc in the cytoplasm, nucleus, and mitochondria is in the range of 2–30 μM, near the known Michaelis constants for O-GlcNAc transferase (OGT) and the UDP-GlcNAc transporters [86]. Thus, overexpression of UDP-GlcNAc transporter should lead the responses to occur at lower concentrations of exogenous GlcNAc, and this remains a subject for future investigations. Additionally, high UDP-GlcNAc flux will also result in hyper O-GlcNAcylation [87,88], altered glycosphingolipid synthesis [89], and augmented mucin type glycan formation [90], which in turn are also considerable players of the galectin-3 lattice. Additionally, glucose, glutamine, and acetyl-CoA supply the hexosamine pathway, regulating the biosynthesis of UDP-GlcNAc and N-glycan branching, and thereby regulating glycoprotein retention in the galectin lattice [91,92].
9. O-GlcNAc Regulation of Galectin-3
Like N-glycosylation, protein O-glycosylation (O-glycans) are structurally diverse. They can be further subclassified based on the sugar attachment, including O-GalNAcylation, O-GlcNAcylation, O-fucosylation, O-mannosylation, and O-glucosylation [93,94]. Among them, the two most abundant classes of protein O-glycosylations are the structurally diverse mucin type O-glycosylation (attachment of O-GalNAc to Ser or Thr) occurring in the Golgi apparatus, and O-GlcNAcylation, the installation of a single sugar residue O-GlcNAc which occurs in the nucleus, cytoplasm, and mitochondria [95,96,97].
O-GlcNAcylation is an abundant PTM in mammals [96]. More than 15,000 proteins from 43 different species have been identified with O-GlcNAc modifications [98]. O-GlcNAcylation modulates protein synthesis, subcellular localization, stability, and protein–protein interactions, thereby influencing diverse cellular processes at the molecular level. Interplay between O-GlcNAcylation and other post-translational modifications markedly amplifies the complexity of protein regulatory networks [97,99]. For instance, O-GlcNAcylation has been shown to modulate Ser/Thr phosphorylation by modifying kinase substrates at proximal sites, competing for shared residues, and altering kinase activity through direct modification [100,101,102].
A defining characteristic of O-GlcNAcylation, setting it apart from most other post-translational modifications, is its regulation by a single pair of antagonistic enzymes. O-GlcNAc transferase (OGT) and O-GlcNAcase (OGA) are responsible for installation and removal of the monosaccharide O-GlcNAc from protein substrates [103,104]. O-GlcNAcylation also appears to serve as a nutrient sensing mechanism in cells. Elevated cellular glucose levels enhance glucose flux through the hexosamine biosynthetic pathway. This pathway generates the precursor molecule UDP-GlcNAc, the nucleotide sugar donor substrate for O-GlcNAcylation. Consequently, elevated glucose concentrations lead to a boost in the supply of UDP-GlcNAc availability, thereby promoting increased protein O-GlcNAcylation [96]. The interplay between O-GlcNAcylation and phosphorylation, its dynamic regulation by just two enzymes, and its sensitivity to nutrient availability have collectively sparked significant interest in its roles in cellular signaling and diverse disease states. The nutrient-sensing function of O-GlcNAcylation is especially critical, as it allows cells to detect nutrient abundance and swiftly modulate cellular processes in response to changing metabolic conditions in order to rapidly adapt to fluctuations in nutrient availability [102,105]. This regulatory axis links metabolism to signaling, contributing to the fine-tuning of cellular functions and implicating O-GlcNAcylation in a broad spectrum of physiological and pathological contexts. Beyond its role in nutrient sensing, O-GlcNAcylation is involved in a broad range of biological processes, including gene transcription, epigenetic regulation, signal transduction, stress response, metabolic homeostasis, and immune function response [86,97,103,106].
Galectin-3 is a nucleocytoplasmic protein that is also secreted into the extracellular space. Since galectin-3 is synthesized in the cytoplasm, and translocates into nucleus, it is exposed to the enzymes of O-GlcNAc cycling. Once reaching the surface, galectin-3 binds to the terminal galactose residue of the LacNAc (Gal-β-1,4-GlcNAc) substrate. An emerging question is whether the pathways of O-GlcNAc cycling may be linked by galectin-3 secretion. O-GlcNAcylation is known to change the functional activities of proteins and govern the trafficking and localization of proteins in cells [99,105]. O-GlcNAcylation occurs directly on key subunits of the protein complexes involved in generating COPII, COPI, and clathrin-coated vesicles (CCVs) [99]. Furthermore, some proteins in autophagy and unconventional secretory pathways are reported to be regulated by O-GlcNAcylation [107,108]. There is growing evidence on the potential involvement of individual galectins in processes of cellular differentiation and to explore formal links to relevant O-GlcNAc cellular homeostasis [109].
9.1. Established Links Between O-GlcNAcylation and Galectin-3 Activity
Alteration of the O-GlcNAcylation cycle with the OGA inhibitor Thiamet G (TMG), or the OGT inhibitor Ac-5SGlcNAc (AC), changed RNA expression of the galectin-3 gene (LGALS3) in the adherent malignant breast cancer cell line MCF7, but not in the suspended acute promyelocytic leukemia cell line HL-60 [110]. Further investigation with HL-60 cells showed that neutrophilic differentiation of those cells induced by all-trans retinoic acid (ATRA) and 6-diazo-5-oxo-L-norleucine (DON, GFPT inhibitor) were associated with a significant drop in cellular O-GlcNAc levels in serum-contained and serum-free cell culture media. The amount of secreted galectin-3 (LGALS3) (measured by ELISA assay) was significantly upregulated by ATRA and DON, while intracellular galectin-3 protein expression was upregulated by both ATRA and DON in serum-contained media, and by all four treatments (ATRA, DON, TMG, and AC) in FBS containing media (measured by RT-qPCR followed by Tukey’s multiple comparison analysis). A much higher magnitude of galectin-3 protein expression was observed in ATRA induced serum free cell culture media [111]. High levels of O-GlcNAc in embryonic stem (ES) cells promote intracellular accumulation of galectins in stem cells, while low levels of O-GlcNAc in extraembryonic endoderm (XEN) cells promote galectin secretion and cell differentiation [112]. Immunoassays (western blot and ELISA) revealed that the XEN cells had lower concentrations of both intracellular and extracellular galectin-3 compared to the ES cells, and the relative secretion of galectin-3 was significantly increased as a result of O-GlcNAcylation occurring concomitantly with XEN differentiation. Label-free LC-MS/MS quantitative proteomic analysis of control and iOGA groups (O-GlcNAcylation dynamics disrupted with OGA inhibitor Thiamet G) identified altered galectin-3 expression in glioblastoma (GBM) secretome, predicting galectin-3 could be a substrate for O-GlcNAc addition and removal [113].
An interesting study with yellow-rumped warblers (Setophaga coronata) showed RNA expression of LGALS3 (galectin-3) significantly increased in flight muscles and hearts for the migratory flown (MF) cohort compared to the migratory unflown (MU) and winter unflown (WU) cohort, while protein expression of the O-GlcNAcylated protein pool (detected with RL2 antibody) in flight muscle was higher in the case of WU compared to MF and MU [114].
Using OGA WT and OGA KO Mouse Embryonic Fibroblast cells (MEFs), our group has shown that galectin-3 indeed is a substrate for O-GlcNAc transferase, and its secretion is tightly regulated by O-GlcNAcylation dynamics and O-GlcNAc levels [58]. Immunoprecipitation of GFP-tagged galectin-3, and western blotting, have shown a significant difference in O-GlcNAcylation status between cytoplasmic and secreted galectin-3. A dramatic alteration in galectin-3 secretion was also observed in response to nutrient conditions, which were dependent on dynamic O-GlcNAcylation. Although no O-GlcNAcylation site is predicted for galectin-3 itself [115], the YinOYang server predicted six (6) possible O-GlcNAcylation cites with a high degree of confidence. Five (5) of these predicted sites are clustered in the multimerization region of galectin-3, and all the mutants which have all five of these sites deleted or mutated were secreted at a significantly lower level than control galectin 3-GFP. The last one (Ser 243) is adjacent to the nuclear export region, and mutation of this site did not appear to alter galectin-3 secretion. O-GlcNAcylation of galectin-3 may play a role in its secretion, and the removal of O-GlcNAcylation is an important step for regulation of galectin-3 multimerization and secretion. O-GlcNAc-driven alterations in galectin-3 secretion also facilitated changes in CIE. This indicates that dynamic O-GlcNAcylation of galectin-3 plays a role in modulating its secretion and can tune its function of transducing nutrient sensing information coded in cell surface glycosylation into biological effects (Figure 6). Whether the effect of the O-GlcNAcylation cycle in the regulation of galectin-3 is only secretory, transcriptional, translational, or some combination, is yet to be determined.
10. Discussion
Galectin-3 is a widely studied protein with diverse binding targets mediating a wide variety of biological outputs. Galectin-3 is ubiquitously expressed in various cell types and tissues, and is detectable inside and outside of cells, as well as on the cell surface. Galectin-3 is implicated in cell and cell–extracellular matrix adhesion, cell growth and differentiation, the cell-cycle, cell signaling, apoptosis, and angiogenesis. Consequently, galectin-3 is involved in regulation of development, immune reactions, tumorigenesis, and tumor growth and metastasis. Important questions remain concerning how regulation of galectin-3 expression may occur in different cell types under different physiological and pathophysiological conditions. The biological roles of galectin-3 have been initially attributed to its carbohydrate-binding activity, but during the past decade a whole new spectrum of functions unrelated to lectin activity have been revealed. Molecular mechanisms that drive the trafficking of galectin-3 in cells, as well as its secretion, remain elusive. This includes mechanisms of unconventional secretion of galectin-3 since it lacks the typical ER-signal peptide required for the conventional pathway.
The evidence presented here suggests that galectin-3 secretion could serve as a nexus linking nutrient information to cell surface signaling events. Our model reflects recent findings demonstrating that O-GlcNAc may regulate the expression and secretion of many factors including galectin-3 (Figure 7). The model is predicated on the nucleotide sugar requirements for diverse surface glycan structures. In particular, the model treats UDP-GlcNAc concentrations in different cell organelles as a key variable. We also point out the importance of the Km values in glycan processing enzymes that utilize UDP-GlcNAc. Changes in UDP-GlcNAc levels linked to nutrient and stress conditions can occur in many cell types such as activated T-cells. In our model, we argue for threshold effects in response to these changes. At lower levels of UDP-GlcNAc, perturbation of the O-GlcNAcylation cycle may occur, triggering altered galectin-3 transcriptional regulation, and secretion. These changes can occur over the range of cytoplasmic concentrations which occur physiologically, and the levels are regulated by feedback inhibition of hexosamine synthesis. Owing to the presence of transporters, the ER and Golgi concentrations of UDP-GlcNAc are much higher as reflected in the thresholds shown for high occupancy N-glycan branching and mucin type O-glycan synthesis. Finally, at very high levels of UDP-GlcNAc, low occupancy N-glycans are synthesized. The threshold-driven changes we depict would suggest that as synthesis and branching of N-glycans occurs, galectin-3 synthesis and secretion would be limited to avoid hyper-activation of signaling pathways normally triggered by galectin-3. In this model, O-GlcNAc would serve as a homeostatic regulator of galectin-3 stimulated signaling. It should be noted that experimental thresholds for the synthesis of the various glycan forms have not been rigorously established. There is evidence for increased branching of glycans at high levels of hexosamine flux [73]. There is also evidence that this leads to an increase in galectin-3 binding and lattice formation. However, this model emphasizes that the different glycan species likely have different thresholds for assembling galactin-3 binding sites. The feedback mechanism illustrated is one means of self-limiting the signaling in response to galectin-3.
5. Conclusions
Although galectin-3 has been implicated in several debilitating disorders, its detailed mechanism of action has yet to be elucidated. To clarify the precise mechanisms of galectin-3 action, it is of the utmost importance that its physiologically functional ligands are identified. Moreover, the development of potent, low-molecular weight inhibitors of galectin-3 could provide powerful analytical tools for investigating functions of galectin-3 [116,117]. These galectin-3 inhibitors with high oral bioavailability, and low toxicity profiles, could lead to combatting progressive tissue fibrosis, and galectin-3-related HF. Alternatively, this O-GlcNAcylation regulation of galectin-3 could be targeted for altering galectin-3 secretion. Current understanding of the various binding partners of galectin-3 is also incomplete. Further characterization and visualization of the galectin-3 lattice utilizing advanced (optical) techniques is necessary to understand the exact mechanisms by which this regulatory protein influences various (extra)cellular processes.
Author Contributions
Conceptualization, M.M.M. and J.A.H.; writing—original draft preparation, M.M.M., and J.A.H; writing—review and editing, M.M.M., D.B., and J.A.H; visualization, M.M.M and J.A.H..; supervision, J.A.H.; project administration, J.A.H.; funding acquisition, J.A.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research and the APC was funded by NIDDK Intramural research, grant number DK060101-18.
Institutional Review Board Statement
Not applicable for this literature review.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
M.M.M., D.B., and J.A.H. thank members of the Hanover lab for discussion, advice, and valuable suggestions. This work was conducted in the Laboratory of Cell and Molecular Biology (LCMB) section in NIDDK, National Institutes of Health (NIH), and supported by the NIDDK, National Institutes of Health, intramural research program grant (DK060101-18). The views, information or content, and conclusions presented do not necessarily represent the official position or policy of, nor should any official endorsement be inferred on the part of, the NIDDK, the NIH, or the Department of Health and Human Services (DHHS). During the preparation of this manuscript/study, the author(s) used biorender.com for the purposes of the preparation of figures. The authors have reviewed and edited the output and take full responsibility for the content of this publication.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study, in the collection, analyses, or interpretation of data, in the writing of the manuscript, or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| HBP | Hexosamine biosynthetic pathway |
| UDP | Uridine diphosphate |
| GlcNAc | N-Acetyl-D-glucosamine |
References
- Barondes, S.H.; Castronovo, V.; Cooper, D.N.W.; Cummings, R.D.; Drickamer, K.; Felzi, T.; Gitt, M.A.; Hirabayashi, J.; Hughes, C.; Kasai, K.-i.; et al. Galectins: A family of animal β-galactoside-binding lectins. Cell 1994, 76, 597–598. [Google Scholar] [CrossRef] [PubMed]
- Dumic, J.; Dabelic, S.; Flogel, M. Galectin-3: an open-ended story. Biochim. Biophys. Acta 2006, 1760, 616–635. [Google Scholar] [CrossRef] [PubMed]
- Roff, C.F.; Wang, J.L. Endogenous lectins from cultured cells. Isolation and characterization of carbohydrate-binding proteins from 3T3 fibroblasts. J. Biol. Chem. 1983, 258, 10657–10663. [Google Scholar] [CrossRef]
- Sciacchitano, S.; Lavra, L.; Morgante, A.; Ulivieri, A.; Magi, F.; De Francesco, G.P.; Bellotti, C.; Salehi, L.B.; Ricci, A. Galectin-3: One Molecule for an Alphabet of Diseases, from A to Z. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Rabinovich, G.A.; Toscano, M.A.; Jackson, S.S.; Vasta, G.R. Functions of cell surface galectin-glycoprotein lattices. Curr. Opin. Struct. Biol. 2007, 17, 513–520. [Google Scholar] [CrossRef]
- Nabi, I.R.; Shankar, J.; Dennis, J.W. The galectin lattice at a glance. J. Cell Sci. 2015, 128, 2213–2219. [Google Scholar] [CrossRef]
- Dong, R.; Zhang, M.; Hu, Q.; Zheng, S.; Soh, A.; Zheng, Y.; Yuan, H. Galectin-3 as a novel biomarker for disease diagnosis and a target for therapy (Review). Int. J. Mol. Med. 2018, 41, 599–614. [Google Scholar] [CrossRef]
- Colnot, C.; Fowlis, D.; Ripoche, M.-A.; Bouchaert, I.; Poirier, F. Embryonic implantation in galectin 1/galectin 3 double mutant mice. Dev. Dyn. 1998, 211, 306–313. [Google Scholar] [CrossRef]
- Kim, S.J.; Lee, H.W.; Gu Kang, H.; La, S.H.; Choi, I.J.; Ro, J.Y.; Bresalier, R.S.; Song, J.; Chun, K.H. Ablation of galectin-3 induces p27(KIP1)-dependent premature senescence without oncogenic stress. Cell Death Differ. 2014, 21, 1769–1779. [Google Scholar] [CrossRef]
- Davidson, P.J.; Li, S.Y.; Lohse, A.G.; Vandergaast, R.; Verde, E.; Pearson, A.; Patterson, R.J.; Wang, J.L.; Arnoys, E.J. Transport of galectin-3 between the nucleus and cytoplasm. I. Conditions and signals for nuclear import. Glycobiology 2006, 16, 602–611. [Google Scholar] [CrossRef]
- Nakahara, S.; Hogan, V.; Inohara, H.; Raz, A. Importin-mediated nuclear translocation of galectin-3. J. Biol. Chem. 2006, 281, 39649–39659. [Google Scholar] [CrossRef] [PubMed]
- Park, J.W.; Voss, P.G.; Grabski, S.; Wang, J.L.; Patterson, R.J. Association of galectin-1 and galectin-3 with Gemin4 in complexes containing the SMN protein. Nucleic Acids Res. 2001, 29, 3595–3602. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.-T.; Patterson, R.J.; Wang, J.L. Intracellular functions of galectins. Biochimica et Biophysica Acta (BBA)—General Subjects 2002, 1572, 263–273. [Google Scholar] [CrossRef]
- Peltan, E.L.; Riley, N.M.; Flynn, R.A.; Roberts, D.S.; Bertozzi, C.R. Galectin-3 does not interact with RNA directly. Glycobiology 2024, 34. [Google Scholar] [CrossRef]
- Goletz, S.; Hanisch, F.-G.; Karsten, U. Novel αGalNAc containing glycans on cytokeratins are recognized in vitro by galectins with type II carbohydrate recognition domains. J. Cell Sci. 1997, 110, 1585–1596. [Google Scholar] [CrossRef] [PubMed]
- Akahani, S.; Nangia-Makker, P.; Inohara, H.; Kim, H.-R. C.; Raz, A. Galectin-3: A Novel Antiapoptotic Molecule with A Functional BH1 (NWGR) Domain of Bcl-2 Family1. Cancer Res. 1997, 57, 5272–5276. [Google Scholar]
- Sato, S.; Hughes, R.C. Binding specificity of a baby hamster kidney lectin for H type I and II chains, polylactosamine glycans, and appropriately glycosylated forms of laminin and fibronectin. J. Biol. Chem. 1992, 267, 6983–6990. [Google Scholar] [CrossRef]
- Talaga, M.L.; Fan, N.; Fueri, A.L.; Brown, R.K.; Bandyopadhyay, P.; Dam, T.K. Multitasking Human Lectin Galectin-3 Interacts with Sulfated Glycosaminoglycans and Chondroitin Sulfate Proteoglycans. Biochemistry 2016, 55, 4541–4551. [Google Scholar] [CrossRef]
- Rosenberg, I.; Cherayil, B.J.; Isselbacher, K.J.; Pillai, S. Mac-2-binding glycoproteins. Putative ligands for a cytosolic beta-galactoside lectin. J. Biol. Chem. 1991, 266, 18731–18736. [Google Scholar] [CrossRef]
- Hughes, R.C. Mac-2: a versatile galactose-binding protein of mammalian tissues. Glycobiology 1994, 4, 5–12. [Google Scholar] [CrossRef]
- Colin Hughes, R. Galectins as modulators of cell adhesion. Biochimie 2001, 83, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Srivatsan, V.; George, M.; Shanmugam, E. Utility of galectin-3 as a prognostic biomarker in heart failure: where do we stand? Eur. J. Prev. Cardiol. 2015, 22, 1096–1110. [Google Scholar] [CrossRef] [PubMed]
- Maiolino, G.; Rossitto, G.; Pedon, L.; Cesari, M.; Frigo, A.C.; Azzolini, M.; Plebani, M.; Rossi, G.P. Galectin-3 Predicts Long-Term Cardiovascular Death in High-Risk Patients With Coronary Artery Disease. Arterioscler., Thromb., Vasc. Biol. 2015, 35, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Tuñón, J.; Blanco-Colio, L.; Cristóbal, C.; Tarín, N.; Higueras, J.; Huelmos, A.; Alonso, J.; Egido, J.; Asensio, D.; Lorenzo, Ó.; et al. Usefulness of a Combination of Monocyte Chemoattractant Protein-1, Galectin-3, and N-Terminal Probrain Natriuretic Peptide to Predict Cardiovascular Events in Patients With Coronary Artery Disease. Am. J. Cardiol. 2014, 113, 434–440. [Google Scholar] [CrossRef] [PubMed]
- Szadkowska, I.; Wlazeł, R.; Migała, M.; Szadkowski, K.; Zielińska, M.; Paradowski, M.; Pawlicki, L. The association between galectin-3 and clinical parameters in patients with fi rst acute myocardial infarction treated with primary percutaneous coronary angioplasty. Cardiology Journal 2013, 20, 577–582. [Google Scholar] [CrossRef]
- Tsai, T.-H.; Sung, P.-H.; Chang, L.-T.; Sun, C.-K.; Yeh, K.-H.; Chung, S.-Y.; Chua, S.; Chen, Y.-L.; Wu, C.-J.; Chang, H.-W.; et al. Value and Level of Galectin-3 in Acute Myocardial Infarction Patients Undergoing Primary Percutaneous Coronary Intervention. J. Atheroscler. Thromb. 2012, 19, 1073–1082. [Google Scholar] [CrossRef]
- Singsaas, E.G.; Manhenke, C.A.; Dickstein, K.; Orn, S. Circulating Galectin-3 Levels Are Increased in Patients with Ischemic Heart Disease, but Are Not Influenced by Acute Myocardial Infarction. Cardiology 2016, 134, 398–405. [Google Scholar] [CrossRef]
- Nishiyama, J.; Kobayashi, S.; Ishida, A.; Nakabayashi, I.; Tajima, O.; Miura, S.; Katayama, M.; Nogami, H. Up-Regulation of Galectin-3 in Acute Renal Failure of the Rat. Am. J. Pathol. 2000, 157, 815–823. [Google Scholar] [CrossRef]
- Sakaki, M.; Fukumori, T.; Fukawa, T.; Elsamman, E.; Shiirevnyamba, A.; Nakatsuji, H.; Kanayama, H.-o. Clinical significance of Galectin-3 in clear cell renal cell carcinoma. J. Med. Invest. 2010, 57, 152–157. [Google Scholar] [CrossRef]
- Kaneko, N.; Gotoh, A.; Okamura, N.; Matsuo, E.-i.; Terao, S.; Watanabe, M.; Yamada, Y.; Hamami, G.; Nakamura, T.; Ikekita, M.; et al. Potential tumor markers of renal cell carcinoma: α-Enolase for postoperative follow up, and galectin-1 and galectin-3 for primary detection. Int. J. Urol. 2013, 20, 530–535. [Google Scholar] [CrossRef]
- von Klot, C.-A.; Kramer, M.W.; Peters, I.; Hennenlotter, J.; Abbas, M.; Scherer, R.; Herrmann, T.R.W.; Stenzl, A.; Kuczyk, M.A.; Serth, J.; et al. Galectin-1 and Galectin-3 mRNA expression in renal cell carcinoma. BMC Clin. Pathol. 2014, 14, 15. [Google Scholar] [CrossRef] [PubMed]
- Ji, F.; Zhang, S.; Jiang, X.; Xu, Y.; Chen, Z.; Fan, Y.; Wang, W. Diagnostic and prognostic value of galectin-3, serum creatinine, and cystatin C in chronic kidney diseases. J. Clin. Lab. Anal. 2017, 31, e22074. [Google Scholar] [CrossRef]
- Nakahara, S.; Oka, N.; Wang, Y.; Hogan, V.; Inohara, H.; Raz, A. Characterization of the Nuclear Import Pathways of Galectin-3. Cancer Res. 2006, 66, 9995–10006. [Google Scholar] [CrossRef] [PubMed]
- Li, S.Y.; Davidson, P.J.; Lin, N.Y.; Patterson, R.J.; Wang, J.L.; Arnoys, E.J. Transport of galectin-3 between the nucleus and cytoplasm. II. Identification of the signal for nuclear export. Glycobiology 2006, 16, 612–622. [Google Scholar] [CrossRef]
- Takenaka, Y.; Fukumori, T.; Yoshii, T.; Oka, N.; Inohara, H.; Kim, H.R.; Bresalier, R.S.; Raz, A. Nuclear export of phosphorylated galectin-3 regulates its antiapoptotic activity in response to chemotherapeutic drugs. Mol. Cell. Biol. 2004, 24, 4395–4406. [Google Scholar] [CrossRef]
- Gong, H.C.; Honjo, Y.; Nangia-Makker, P.; Hogan, V.; Mazurak, N.; Bresalier, R.S.; Raz, A. The NH2 Terminus of Galectin-3 Governs Cellular Compartmentalization and Functions in Cancer Cells1. Cancer Res. 1999, 59, 6239–6245. [Google Scholar]
- Sato, S.; Burdett, I.; Hughes, R.C. Secretion of the Baby Hamster Kidney 30-kDa Galactose-Binding Lectin from Polarized and Nonpolarized Cells: A Pathway Independent of the Endoplasmic Reticulum-Golgi Complex. Exp. Cell Res. 1993, 207, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Mehul, B.; Hughes, R.C. Plasma membrane targetting, vesicular budding and release of galectin 3 from the cytoplasm of mammalian cells during secretion. J. Cell Sci. 1997, 110, 1169–1178. [Google Scholar] [CrossRef] [PubMed]
- Menon, R.P.; Hughes, R.C. Determinants in the N-terminal domains of galectin-3 for secretion by a novel pathway circumventing the endoplasmic reticulum–Golgi complex. Eur. J. Biochem. 1999, 264, 569–576. [Google Scholar] [CrossRef]
- Gao, X.; Liu, D.; Fan, Y.; Li, X.; Xue, H.; Ma, Y.; Zhou, Y.; Tai, G. The two endocytic pathways mediated by the carbohydrate recognition domain and regulated by the collagen-like domain of galectin-3 in vascular endothelial cells. PLoS One 2012, 7, e52430. [Google Scholar] [CrossRef]
- Hara, A.; Niwa, M.; Kanayama, T.; Noguchi, K.; Niwa, A.; Matsuo, M.; Kuroda, T.; Hatano, Y.; Okada, H.; Tomita, H. Galectin-3: A Potential Prognostic and Diagnostic Marker for Heart Disease and Detection of Early Stage Pathology. Biomolecules 2020, 10. [Google Scholar] [CrossRef]
- Hara, A.; Niwa, M.; Noguchi, K.; Kanayama, T.; Niwa, A.; Matsuo, M.; Hatano, Y.; Tomita, H. Galectin-3 as a Next-Generation Biomarker for Detecting Early Stage of Various Diseases. Biomolecules 2020, 10. [Google Scholar] [CrossRef]
- Cervantes-Alvarez, E.; la Rosa, N.L.; la Mora, M.S.; Valdez-Sandoval, P.; Palacios-Jimenez, M.; Rodriguez-Alvarez, F.; Vera-Maldonado, B.I.; Aguirre-Aguilar, E.; Escobar-Valderrama, J.M.; Alanis-Mendizabal, J.; et al. Galectin-3 as a potential prognostic biomarker of severe COVID-19 in SARS-CoV-2 infected patients. Sci. Rep. 2022, 12, 1856. [Google Scholar] [CrossRef]
- Liu, F.T.; Stowell, S.R. The role of galectins in immunity and infection. Nat. Rev. Immunol. 2023, 23, 479–494. [Google Scholar] [CrossRef]
- Kouo, T.; Huang, L.; Pucsek, A.B.; Cao, M.; Solt, S.; Armstrong, T.; Jaffee, E. Galectin-3 Shapes Antitumor Immune Responses by Suppressing CD8+ T Cells via LAG-3 and Inhibiting Expansion of Plasmacytoid Dendritic Cells. Cancer Immunol. Res. 2015, 3, 412–423. [Google Scholar] [CrossRef] [PubMed]
- Gajovic, N.; Markovic, S.S.; Jurisevic, M.; Jovanovic, M.; Arsenijevic, N.; Mijailovic, Z.; Jovanovic, M.; Jovanovic, I. Galectin-3 as an important prognostic marker for COVID-19 severity. Sci. Rep. 2023, 13, 1460. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Alvarez, L.; Ortega, E. The Many Roles of Galectin-3, a Multifaceted Molecule, in Innate Immune Responses against Pathogens. Mediators Inflamm. 2017, 2017, 9247574. [Google Scholar] [CrossRef]
- Ruvolo, P.P. Galectin 3 as a guardian of the tumor microenvironment. Biochim. Biophys. Acta. (BBA)-Mol. Cell Res. 2016, 1863, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Bouffette, S.; Botez, I.; De Ceuninck, F. Targeting galectin-3 in inflammatory and fibrotic diseases. Trends Pharmacol. Sci. 2023, 44, 519–531. [Google Scholar] [CrossRef]
- Honig, E.; Schneider, K.; Jacob, R. Recycling of galectin-3 in epithelial cells. Eur J Cell Biol 2015, 94, (7–9). [Google Scholar] [CrossRef]
- Nickel, W.; Rabouille, C. Mechanisms of regulated unconventional protein secretion. Nat. Rev. Mol. Cell Biol. 2009, 10, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Popa, S.J.; Stewart, S.E.; Moreau, K. Unconventional secretion of annexins and galectins. Semin. Cell Dev. Biol. 2018, 83, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, Y.; Kanai-Azuma, M.; Akimoto, Y.; Kawakami, H.; Yanoshita, R. Exosome-Like Vesicles with Dipeptidyl Peptidase IV in Human Saliva. Biol. Pharm. Bull. 2008, 31, 1059–1062. [Google Scholar] [CrossRef] [PubMed]
- Pisitkun, T.; Shen, R.-F.; Knepper, M.A. Identification and proteomic profiling of exosomes in human urine. Proc. Natl. Acad. Sci. U. S. A. 2004, 101, 13368–13373. [Google Scholar] [CrossRef]
- Welton, J.L.; Khanna, S.; Giles, P.J.; Brennan, P.; Brewis, I.A.; Staffurth, J.; Mason, M.D.; Clayton, A. Proteomics Analysis of Bladder Cancer Exosomes*. Mol. Cell. Proteomics 2010, 9, 1324–1338. [Google Scholar] [CrossRef]
- Théry, C.; Boussac, M.; Véron, P.; Ricciardi-Castagnoli, P.; Raposo, G. a.; Garin, J. m.; Amigorena, S. Proteomic Analysis of Dendritic Cell-Derived Exosomes: A Secreted Subcellular Compartment Distinct from Apoptotic Vesicles1. J. Immunol. 2001, 166, 7309–7318. [Google Scholar] [CrossRef]
- Menon, S.; Kang, C.-M.; Beningo, K.A. Galectin-3 secretion and tyrosine phosphorylation is dependent on the calpain small subunit, Calpain 4. Biochem. Biophys. Res. Commun. 2011, 410, 91–96. [Google Scholar] [CrossRef]
- Mathew, M.P.; Abramowitz, L.K.; Donaldson, J.G.; Hanover, J.A. Nutrient-responsive O-GlcNAcylation dynamically modulates the secretion of glycan-binding protein galectin 3. J. Biol. Chem. 2022, 298, 101743. [Google Scholar] [CrossRef]
- Hamann, K.K.; Cowles, E.A.; Wang, J.L.; Anderson, R.L. Expression of carbohydrate binding protein 35 in human fibroblasts: Variations in the levels of mRNA, protein, and isoelectric species as a function of replicative competence. Exp. Cell Res. 1991, 196, 82–91. [Google Scholar] [CrossRef]
- Funasaka, T.; Raz, A.; Nangia-Makker, P. Nuclear transport of galectin-3 and its therapeutic implications. Semin. Cancer Biol. 2014, 27, 30–38. [Google Scholar] [CrossRef]
- Funasaka, T.; Raz, A.; Nangia-Makker, P. Galectin-3 in angiogenesis and metastasis. Glycobiology 2014, 24, 886–891. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Finley, R.L.; Raz, A.; Kim, H.-R. C. Galectin-3 Translocates to the Perinuclear Membranes and Inhibits Cytochrome c Release from the Mitochondria: A ROLE FOR SYNEXIN IN GALECTIN-3 TRANSLOCATION*. J. Biol. Chem. 2002, 277, 15819–15827. [Google Scholar] [CrossRef] [PubMed]
- Delacour, D.; Koch, A.; Jacob, R. The Role of Galectins in Protein Trafficking. Traffic 2009, 10, 1405–1413. [Google Scholar] [CrossRef] [PubMed]
- Lakshminarayan, R.; Wunder, C.; Becken, U.; Howes, M.T.; Benzing, C.; Arumugam, S.; Sales, S.; Ariotti, N.; Chambon, V.; Lamaze, C.; et al. Galectin-3 drives glycosphingolipid-dependent biogenesis of clathrin-independent carriers. Nat. Cell Biol. 2014, 16, 595–606. [Google Scholar] [CrossRef]
- Mathew, M.P.; Donaldson, J.G. Distinct cargo-specific response landscapes underpin the complex and nuanced role of galectin-glycan interactions in clathrin-independent endocytosis. J. Biol. Chem. 2018, 293, 7222–7237. [Google Scholar] [CrossRef]
- Schneider, D.; Greb, C.; Koch, A.; Straube, T.; Elli, A.; Delacour, D.; Jacob, R. Trafficking of galectin-3 through endosomal organelles of polarized and non-polarized cells. Eur J Cell Biol 2010, 89, 788–798. [Google Scholar] [CrossRef]
- Straube, T.; von Mach, T.; Honig, E.; Greb, C.; Schneider, D.; Jacob, R. pH-dependent recycling of galectin-3 at the apical membrane of epithelial cells. Traffic 2013, 14, 1014–1027. [Google Scholar] [CrossRef]
- Delacour, D.; Greb, C.; Koch, A.; Salomonsson, E.; Leffler, H.; Le Bivic, A.; Jacob, R. Apical sorting by galectin-3-dependent glycoprotein clustering. Traffic 2007, 8, 379–388. [Google Scholar] [CrossRef]
- Banfer, S.; Schneider, D.; Dewes, J.; Strauss, M.T.; Freibert, S.A.; Heimerl, T.; Maier, U.G.; Elsasser, H.P.; Jungmann, R.; Jacob, R. Molecular mechanism to recruit galectin-3 into multivesicular bodies for polarized exosomal secretion. Proc. Natl. Acad. Sci. U. S. A. 2018, 115, E4396–E4405. [Google Scholar] [CrossRef]
- Stewart, S.E.; Menzies, S.A.; Popa, S.J.; Savinykh, N.; Petrunkina Harrison, A.; Lehner, P.J.; Moreau, K. A genome-wide CRISPR screen reconciles the role of N-linked glycosylation in galectin-3 transport to the cell surface. J. Cell Sci. 2017, 130, 3234–3247. [Google Scholar] [CrossRef]
- MacDonald, E.; Forrester, A.; Valades-Cruz, C.A.; Madsen, T.D.; Hetmanski, J.H.R.; Dransart, E.; Ng, Y.; Godbole, R.; Shp, A.A.; Leconte, L.; et al. Growth factor-triggered de-sialylation controls glycolipid-lectin-driven endocytosis. Nat. Cell Biol. 2025, 27, 449–463. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, M.M.; Bond, M.R.; Abramowitz, L.K.; Biesbrock, D.; Woodroofe, C.C.; Kim, E.J.; Swenson, R.E.; Hanover, J.A. Tools and tactics to define specificity of metabolic chemical reporters. Front. Mol. Biosci. 2023, 10, 1286690. [Google Scholar] [CrossRef] [PubMed]
- Lau, K.S.; Partridge, E.A.; Grigorian, A.; Silvescu, C.I.; Reinhold, V.N.; Demetriou, M.; Dennis, J.W. Complex N-glycan number and degree of branching cooperate to regulate cell proliferation and differentiation. Cell 2007, 129, 123–134. [Google Scholar] [CrossRef]
- Stanley, P. A method to the madness of N-glycan complexity? Cell 2007, 129, 27–29. [Google Scholar] [CrossRef] [PubMed]
- Zielinska, D.F.; Gnad, F.; Wisniewski, J.R.; Mann, M. Precision mapping of an in vivo N-glycoproteome reveals rigid topological and sequence constraints. Cell 2010, 141, 897–907. [Google Scholar] [CrossRef]
- Varki, A. Essentials of Glycobiology, Cold Spring Harbour Laboratory Press, 2009.
- Aebi, M.; Bernasconi, R.; Clerc, S.; Molinari, M. N-glycan structures: recognition and processing in the ER. Trends Biochem. Sci. 2010, 35, 74–82. [Google Scholar] [CrossRef]
- Deprez, P.; Gautschi, M.; Helenius, A. More Than One Glycan Is Needed for ER Glucosidase II to Allow Entry of Glycoproteins into the Calnexin/Calreticulin Cycle. Mol. Cell 2005, 19, 183–195. [Google Scholar] [CrossRef]
- Mukherjee, M.M.; Biesbrock, D.; Abramowitz, L.K.; Pavan, M.; Kumar, B.; Walter, P.J.; Azadi, P.; Jacobson, K.A.; Hanover, J.A. Selective bioorthogonal probe for N-glycan hybrid structures. Nat. Chem. Biol. 2025, 21, 681–692. [Google Scholar] [CrossRef]
- Schachter, H. Biosynthetic controls that determine the branching and microheterogeneity of protein-bound oligosaccharides. Biochem. Cell Biol. 1986, 64, 163–181. [Google Scholar] [CrossRef]
- Hirabayashi, J.; Hashidate, T.; Arata, Y.; Nishi, N.; Nakamura, T.; Hirashima, M.; Urashima, T.; Oka, T.; Futai, M.; Muller, W.E.G.; et al. Oligosaccharide specificity of galectins: a search by frontal affinity chromatography. Biochimica et Biophysica Acta (BBA)—General Subjects 2002, 1572, 232–254. [Google Scholar] [CrossRef]
- Dam, T.K.; Gabius, H.-J.; André, S.; Kaltner, H.; Lensch, M.; Brewer, C.F. Galectins Bind to the Multivalent Glycoprotein Asialofetuin with Enhanced Affinities and a Gradient of Decreasing Binding Constants. Biochemistry 2005, 44, 12564–12571. [Google Scholar] [CrossRef] [PubMed]
- Nieminen, J.; Kuno, A.; Hirabayashi, J.; Sato, S. Visualization of galectin-3 oligomerization on the surface of neutrophils and endothelial cells using fluorescence resonance energy transfer. J. Biol. Chem. 2007, 282, 1374–1383. [Google Scholar] [CrossRef] [PubMed]
- Abbott, K.L.; Aoki, K.; Lim, J.-M.; Porterfield, M.; Johnson, R.; O’Regan, R.M.; Wells, L.; Tiemeyer, M.; Pierce, M. Targeted Glycoproteomic Identification of Biomarkers for Human Breast Carcinoma. J. Proteome Res. 2008, 7, 1470–1480. [Google Scholar] [CrossRef]
- Boscher, C.; Zheng, Y.Z.; Lakshminarayan, R.; Johannes, L.; Dennis, J.W.; Foster, L.J.; Nabi, I.R. Galectin-3 protein regulates mobility of N-cadherin and GM1 ganglioside at cell-cell junctions of mammary carcinoma cells. J. Biol. Chem. 2012, 287, 32940–32952. [Google Scholar] [CrossRef]
- Bond, M.R.; Hanover, J.A. A little sugar goes a long way: the cell biology of O-GlcNAc. J. Cell Biol. 2015, 208, 869–880. [Google Scholar] [CrossRef]
- Ong, Q.; Han, W.; Yang, X. O-GlcNAc as an Integrator of Signaling Pathways. Front. Endocrinol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Le Minh, G.; Esquea, E.M.; Young, R.G.; Huang, J.; Reginato, M.J. On a sugar high: Role of O-GlcNAcylation in cancer. J. Biol. Chem. 2023, 299. [Google Scholar] [CrossRef]
- Mukherjee, M.M.; Watson, M.P.; Biesbrock, D.; Abramowitz, L.K.; Lee, J.; Hanover, J.A. Development of 9-Pr4ManNAlk phenanthrene Derivative as Fluorescent-Raman Bioorthogonal Probe to Specifically interrogate the Biosynthesis of Gangliosides. Glycobiology 2023, 33, 980–980. [Google Scholar]
- Jiang, Y.; Wu, J.; Guan, F.; Liang, L.; Wang, Y. O-GlcNAcylation determines the function of the key O-GalNAc glycosyltransferase C1GalT1 in bladder cancer. Acta Biochim Biophys Sin (Shanghai) 2024, 56, 1108–1117. [Google Scholar] [CrossRef]
- Abdel Rahman, A.M.; Ryczko, M.; Pawling, J.; Dennis, J.W. Probing the hexosamine biosynthetic pathway in human tumor cells by multitargeted tandem mass spectrometry. ACS Chem. Biol. 2013, 8, 2053–2062. [Google Scholar] [CrossRef]
- Grigorian, A.; Lee, S.U.; Tian, W.; Chen, I.J.; Gao, G.; Mendelsohn, R.; Dennis, J.W.; Demetriou, M. Control of T Cell-mediated autoimmunity by metabolite flux to N-glycan biosynthesis. J. Biol. Chem. 2007, 282, 20027–20035. [Google Scholar] [CrossRef] [PubMed]
- Peter-Katalinić, J. Methods in Enzymology: O-Glycosylation of Proteins. In Methods in Enzymology; Academic Press, 2005; Vol. 405, pp. 139–171. [Google Scholar]
- Haltiwanger Rs Fau—Wells, L.; Wells L Fau—Freeze, H.H.; Freeze Hh Fau—Jafar-Nejad, H.; Jafar-Nejad H Fau—Okajima, T.; Okajima T Fau—Stanley, P.; Stanley, P. Other Classes of Eukaryotic Glycans. BTI—Essentials of Glycobiology, 2022; 4th Ed, 155–164. [Google Scholar]
- Freeze, H.H.; Ng, B.G. Golgi glycosylation and human inherited diseases. Cold Spring Harb. Perspect. Biol. 2011, 3, (1943–0264. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Qian, K. Protein O-GlcNAcylation: emerging mechanisms and functions. Nat. Rev. Mol. Cell Biol. 2017, 18, 452–465. [Google Scholar] [CrossRef]
- Hanover, J.A. Glycan-dependent signaling: O-linked N-acetylglucosamine. The FASEB Journal 2001, 15, 1865–1876. [Google Scholar] [CrossRef]
- Mannino, M.P.; Hart, G.W. The Beginner’s Guide to O-GlcNAc: From Nutrient Sensitive Pathway Regulation to Its Impact on the Immune System. Front. Immunol. 2022, 13, 828648. [Google Scholar] [CrossRef] [PubMed]
- Ben Ahmed, A.; Lemaire, Q.; Scache, J.; Mariller, C.; Lefebvre, T.; Vercoutter-Edouart, A.-S. O-GlcNAc Dynamics: The Sweet Side of Protein Trafficking Regulation in Mammalian Cells. Cells 2023, 12. [Google Scholar] [CrossRef]
- Wang, Z.; Gucek, M.; Hart, G.W. Cross-talk between GlcNAcylation and phosphorylation: Site-specific phosphorylation dynamics in response to globally elevated O-GlcNAc. Proc. Natl. Acad. Sci. U. S. A. 2008, 105, 13793–13798. [Google Scholar] [CrossRef]
- Tarrant, M.K.; Rho, H.-S.; Xie, Z.; Jiang, Y.L.; Gross, C.; Culhane, J.C.; Yan, G.; Qian, J.; Ichikawa, Y.; Matsuoka, T.; et al. Regulation of CK2 by phosphorylation and O-GlcNAcylation revealed by semisynthesis. Nat. Chem. Biol. 2012, 8, 262–269. [Google Scholar] [CrossRef]
- Zeidan, Q.; Hart, G.W. The intersections between O-GlcNAcylation and phosphorylation: implications for multiple signaling pathways. J. Cell Sci. 2010, 123, 13–22. [Google Scholar] [CrossRef]
- Love, D.C.; Hanover, J.A. The Hexosamine Signaling Pathway: Deciphering the “O-GlcNAc Code”. Science’s STKE 2005, 2005, re13–re13. [Google Scholar] [CrossRef]
- Vocadlo, D.J. O-GlcNAc processing enzymes: catalytic mechanisms, substrate specificity, and enzyme regulation. Curr. Opin. Chem. Biol. 2012, 16, 488–497. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, Y. Emerging roles of O-GlcNAcylation in protein trafficking and secretion. J. Biol. Chem. 2024, 300, 105677. [Google Scholar] [CrossRef] [PubMed]
- Konzman, D.; Abramowitz, L.K.; Steenackers, A.; Mukherjee, M.M.; Na, H.J.; Hanover, J.A. O-GlcNAc: Regulator of Signaling and Epigenetics Linked to X-linked Intellectual Disability. Front. Genet. 2020, 11, 605263. [Google Scholar] [CrossRef]
- Guo, B.; Liang, Q.; Li, L.; Hu, Z.; Wu, F.; Zhang, P.; Ma, Y.; Zhao, B.; Kovacs, A.L.; Zhang, Z.; et al. O-GlcNAc-modification of SNAP-29 regulates autophagosome maturation. Nat. Cell Biol. 2014, 16, 1215–1226. [Google Scholar] [CrossRef]
- Jo, Y.K.; Park, N.Y.; Park, S.J.; Kim, B.-G.; Shin, J.H.; Jo, D.S.; Bae, D.-J.; Suh, Y.-A.; Chang, J.H.; Lee, E.K.; et al. O-GlcNAcylation of ATG4B positively regulates autophagy by increasing its hydroxylase activity. Oncotarget 2016, 7, 57186–57196. [Google Scholar] [CrossRef] [PubMed]
- Tazhitdinova, R.; Timoshenko, A.V. The Emerging Role of Galectins and O-GlcNAc Homeostasis in Processes of Cellular Differentiation. Cells 2020, 9, 1792. [Google Scholar] [CrossRef]
- Sherazi, A.A.; Jariwala, K.A.; Cybulski, A.N.; Lewis, J.W.; Karagiannis, J.I.M.; Cumming, R.C.; Timoshenko, A.V. Effects of Global <em>O</em>-GlcNAcylation on Galectin Gene-expression Profiles in Human Cancer Cell Lines. Anticancer Res. 2018, 38, 6691–6697. [Google Scholar] [CrossRef]
- McTague, A.; Tazhitdinova, R.; Timoshenko, A.V. O-GlcNAc-Mediated Regulation of Galectin Expression and Secretion in Human Promyelocytic HL-60 Cells Undergoing Neutrophilic Differentiation. Biomolecules 2022, 12. [Google Scholar] [CrossRef]
- Gatie, M.I.; Spice, D.M.; Garha, A.; McTague, A.; Ahmer, M.; Timoshenko, A.V.; Kelly, G.M. O-GlcNAcylation and Regulation of Galectin-3 in Extraembryonic Endoderm Differentiation. Biomolecules 2022, 12. [Google Scholar] [CrossRef]
- Oliveira-Nunes, M.C.; Juliao, G.; Menezes, A.; Mariath, F.; Hanover, J.A.; Evaristo, J.A.M.; Nogueira, F.C.S.; Dias, W.B.; de Abreu Pereira, D.; Carneiro, K. O-GlcNAcylation protein disruption by Thiamet G promotes changes on the GBM U87-MG cells secretome molecular signature. Clin. Proteomics 2021, 18, 14. [Google Scholar] [CrossRef]
- Bradley, S.S.; Dick, M.F.; Guglielmo, C.G.; Timoshenko, A.V. Seasonal and flight-related variation of galectin expression in heart, liver and flight muscles of yellow-rumped warblers (Setophaga coronata). Glycoconj. J. 2017, 34, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Wulff-Fuentes, E.; Berendt, R.R.; Massman, L.; Danner, L.; Malard, F.; Vora, J.; Kahsay, R.; Olivier-Van Stichelen, S. The human O-GlcNAcome database and meta-analysis. Sci. Data 2021, 8, 25. [Google Scholar] [CrossRef] [PubMed]
- Sorme, P.; Qian, Y.; Nyholm, P.G.; Leffler, H.; Nilsson, U.J. Low micromolar inhibitors of galectin-3 based on 3′-derivatization of N-acetyllactosamine. Chembiochem 2002, 3, (2–3). [Google Scholar] [CrossRef]
- Sörme, P.; Arnoux, P.; Kahl-Knutsson, B.; Leffler, H.; Rini, J.M.; Nilsson, U.J. Structural and Thermodynamic Studies on Cation−Π Interactions in Lectin−Ligand Complexes: High-Affinity Galectin-3 Inhibitors through Fine-Tuning of an Arginine−Arene Interaction. J. Am. Chem. Soc. 2005, 127, 1737–1743. [Google Scholar] [CrossRef]
Figure 1.
Schematic presentation of galectin family members and the. galectin-glycan lattice. Galectin members are divided into three types based on the organization of the galectin carbohydrate recognition domain (CRD). (A) Prototype galectins (Gal-1, -2, -5, -7, 10, -11, -13, -14, -15, and -16). (B) Tandem repeat type galectins (Gal-4, -6, -8, -9, and -12). (C) The unique chimeric type of galectin (Gal-3). (D) Schematic presentation of bivalent lattice formation of galectin-3 pentamer with glycans.
Figure 1.
Schematic presentation of galectin family members and the. galectin-glycan lattice. Galectin members are divided into three types based on the organization of the galectin carbohydrate recognition domain (CRD). (A) Prototype galectins (Gal-1, -2, -5, -7, 10, -11, -13, -14, -15, and -16). (B) Tandem repeat type galectins (Gal-4, -6, -8, -9, and -12). (C) The unique chimeric type of galectin (Gal-3). (D) Schematic presentation of bivalent lattice formation of galectin-3 pentamer with glycans.
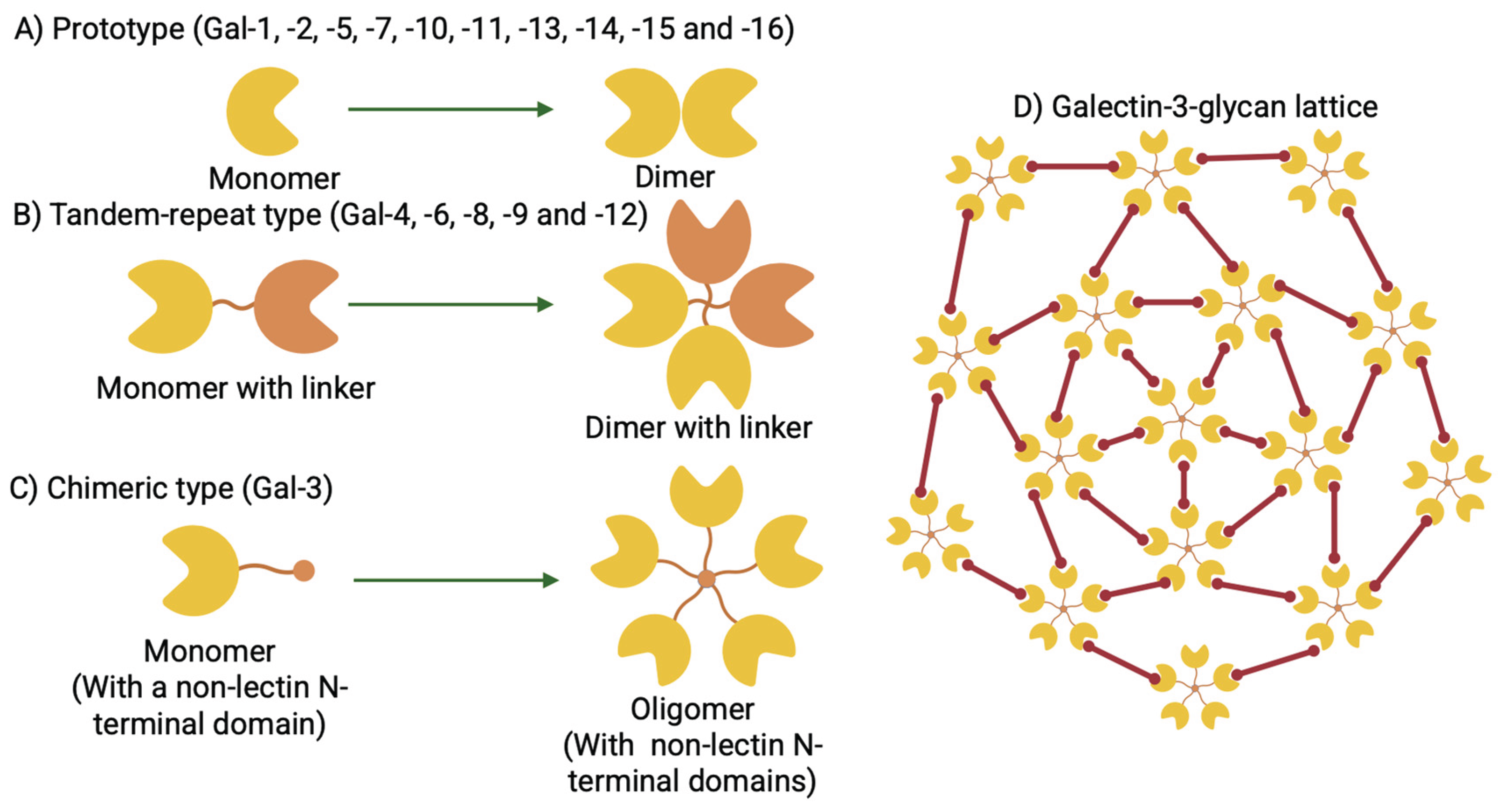
Figure 2.
Schematic presentation of galectin-3 binding glycans. Galectin-3 recognizes and binds to the galactose subunit of the galactose (β1-4) N-acetylglucosamine (N-acetyllactosamine) linkage present in (A) N-glycans, (B) O-glycans, (C) glycolipids, and (D) blood group antigens. Brackets represent galectin-3 binding N-acetyllactosamine (LacNAc) sites on the glycan species. Asn = Asparagine, ST6Gal1 = α (2-6) sialyltransferase 1, GCNT1 = N-acetylglucosaminyltransferase 1, GnT5 = N-acetylglucosaminyltransferase 5, ST3Gal1 = α (2-3) sialyltransferase 1.
Figure 2.
Schematic presentation of galectin-3 binding glycans. Galectin-3 recognizes and binds to the galactose subunit of the galactose (β1-4) N-acetylglucosamine (N-acetyllactosamine) linkage present in (A) N-glycans, (B) O-glycans, (C) glycolipids, and (D) blood group antigens. Brackets represent galectin-3 binding N-acetyllactosamine (LacNAc) sites on the glycan species. Asn = Asparagine, ST6Gal1 = α (2-6) sialyltransferase 1, GCNT1 = N-acetylglucosaminyltransferase 1, GnT5 = N-acetylglucosaminyltransferase 5, ST3Gal1 = α (2-3) sialyltransferase 1.
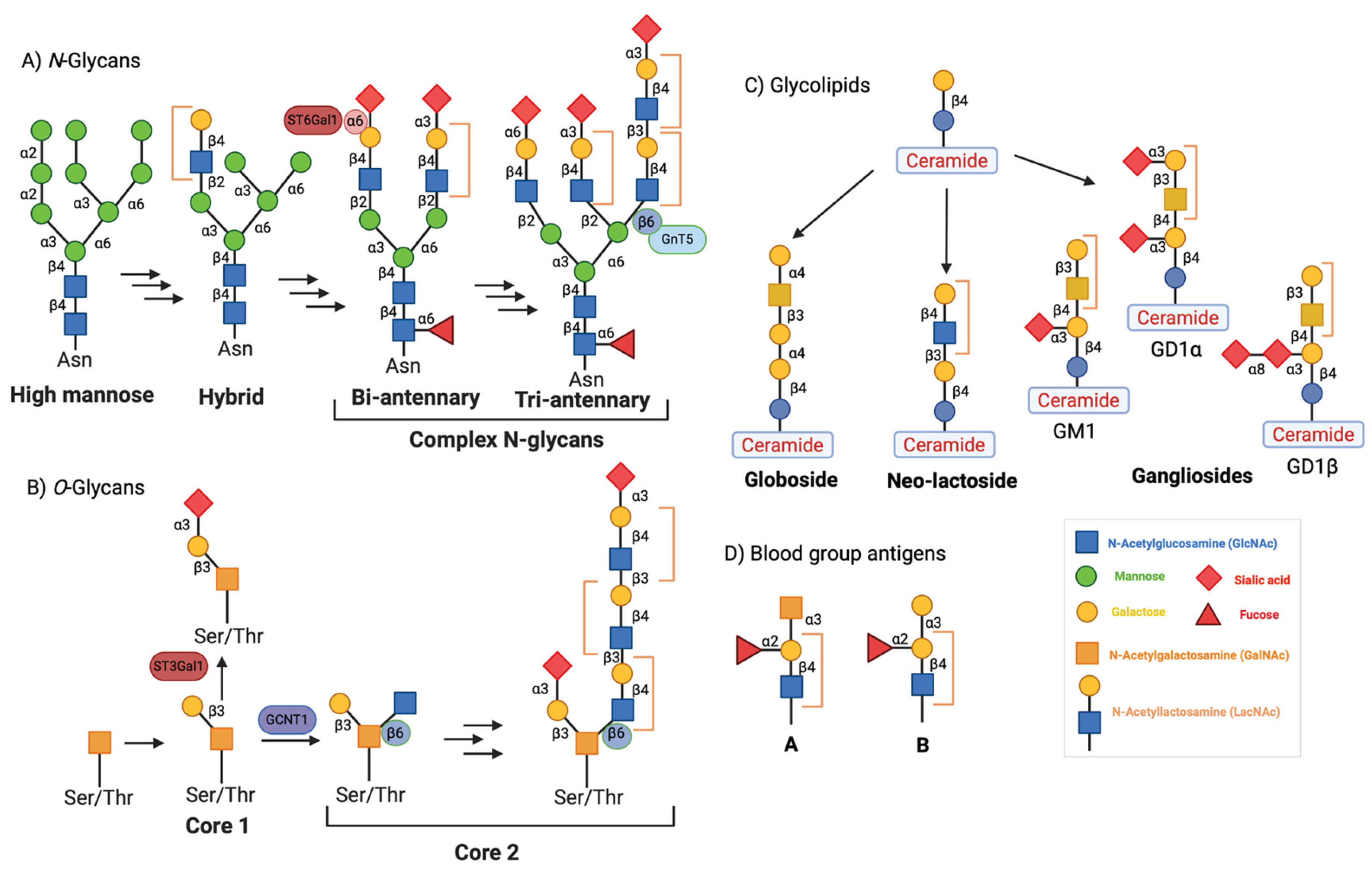
Figure 3.
Schematic presentation of galectin-3 expression and distribution in mammalian tissues. Galectin-3 is ubiquitously and differentially expressed throughout all tissues except the spleen and lymph nodes.
Figure 3.
Schematic presentation of galectin-3 expression and distribution in mammalian tissues. Galectin-3 is ubiquitously and differentially expressed throughout all tissues except the spleen and lymph nodes.
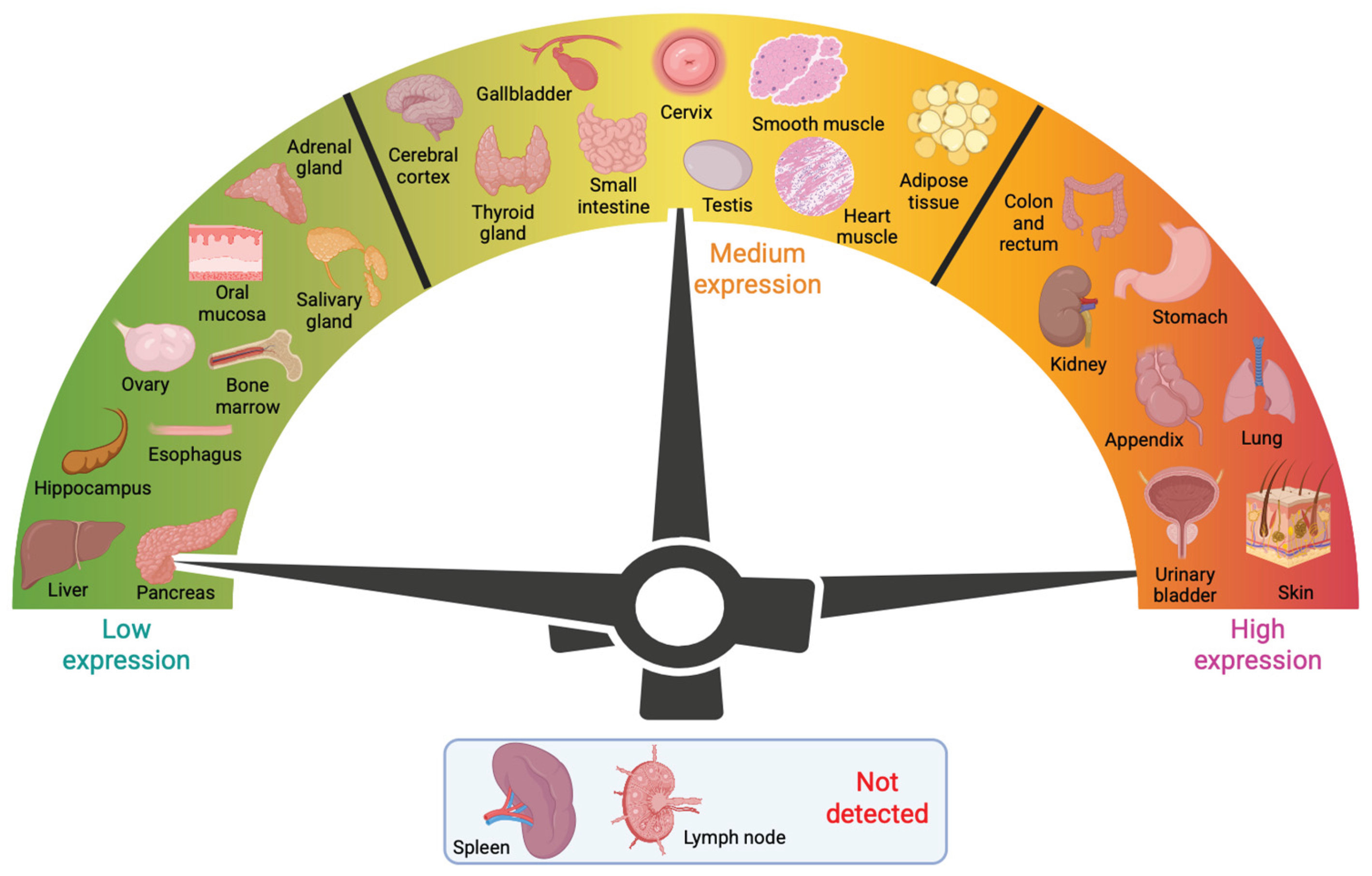
Figure 4.
Schematic presentation of galectin-3 trafficking showing its cytoplasmic synthesis, nuclear translocation, apical sorting, noncanonical secretion, glycan mediated clathrin independent endocytosis (CIE), and endosomal recycling.
Figure 4.
Schematic presentation of galectin-3 trafficking showing its cytoplasmic synthesis, nuclear translocation, apical sorting, noncanonical secretion, glycan mediated clathrin independent endocytosis (CIE), and endosomal recycling.
Figure 5.
The hexosamine biosynthetic pathway generates the nucleotide sugar donor UDP-GlcNAc. Oligosaccharyltransferase (OST) transfers the preassembled donor Glc3Man9GlcNAc2-pp-dolichol to NXS/T motifs on nascently synthesized proteins in the ER. Glycoproteins travel from the ER to the cis, medial, and trans Golgi en route to the cell surface. The Golgi resident N-acetylglucosaminyltransferase enzymes, entitled by their gene names (Mgat1, Mgat2, Mgat4, and Mgat5), generate branched N-glycans that display a range of affinities for galectins. The Km values for Mgat1, Mgat2, Mgat4, and Mgat5 are indicated for both UDP-GlcNAc and acceptor glycoproteins as measured in vitro.
Figure 5.
The hexosamine biosynthetic pathway generates the nucleotide sugar donor UDP-GlcNAc. Oligosaccharyltransferase (OST) transfers the preassembled donor Glc3Man9GlcNAc2-pp-dolichol to NXS/T motifs on nascently synthesized proteins in the ER. Glycoproteins travel from the ER to the cis, medial, and trans Golgi en route to the cell surface. The Golgi resident N-acetylglucosaminyltransferase enzymes, entitled by their gene names (Mgat1, Mgat2, Mgat4, and Mgat5), generate branched N-glycans that display a range of affinities for galectins. The Km values for Mgat1, Mgat2, Mgat4, and Mgat5 are indicated for both UDP-GlcNAc and acceptor glycoproteins as measured in vitro.
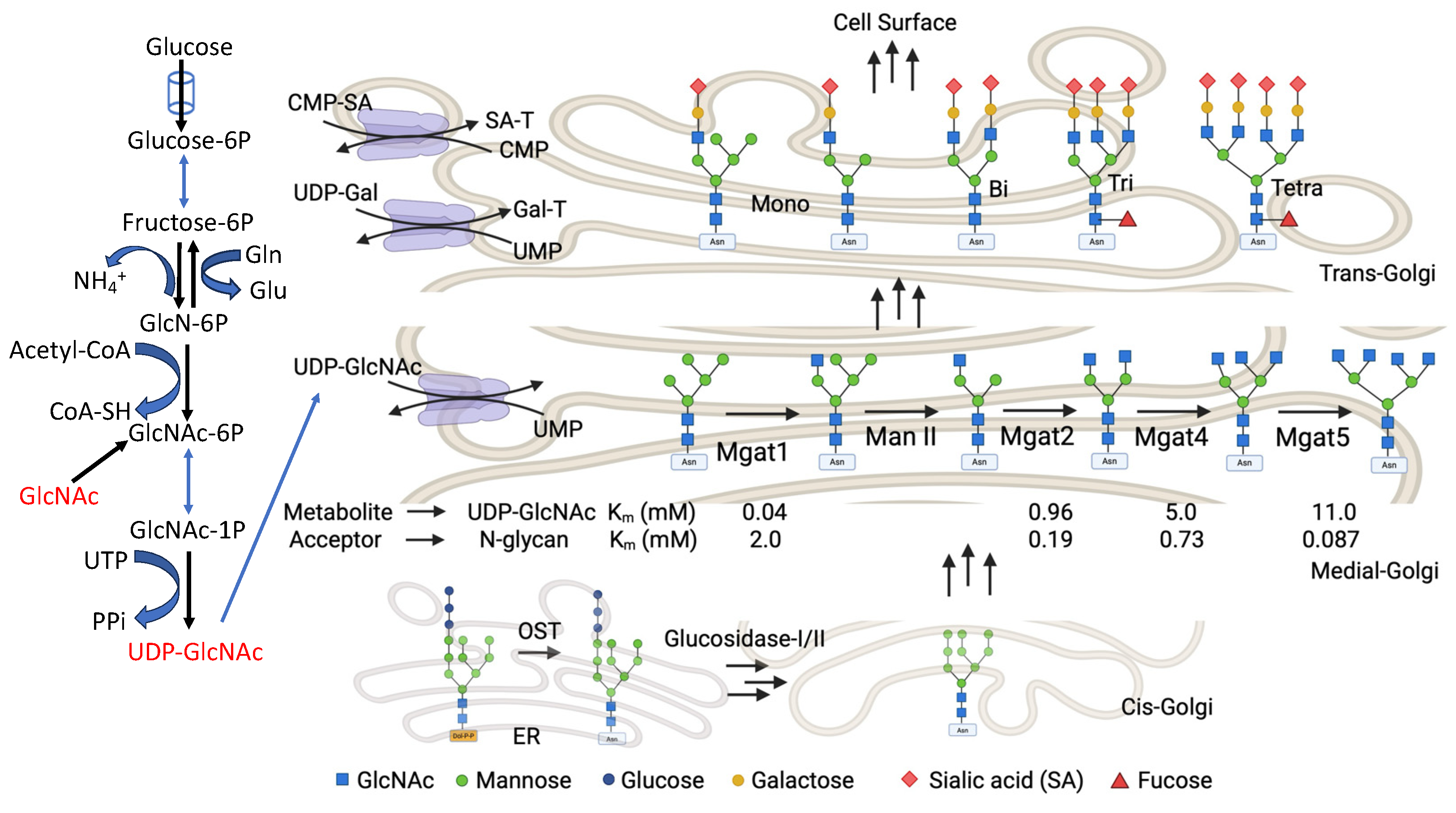
Figure 6.
Schematic presentation of the role of O-GlcNAcylation in galectin 3 secretion. Galectin 3 is synthesized in the cytoplasm and then secreted by noncanonical mechanisms. The cytoplasmic origin of galectin-3, and nuclear translocation, allows interaction with the O-GlcNAcylation machinery. This model indicates that galectin 3 necessitates O-GlcNAcylation and deglycosylation for proper secretion.
Figure 6.
Schematic presentation of the role of O-GlcNAcylation in galectin 3 secretion. Galectin 3 is synthesized in the cytoplasm and then secreted by noncanonical mechanisms. The cytoplasmic origin of galectin-3, and nuclear translocation, allows interaction with the O-GlcNAcylation machinery. This model indicates that galectin 3 necessitates O-GlcNAcylation and deglycosylation for proper secretion.
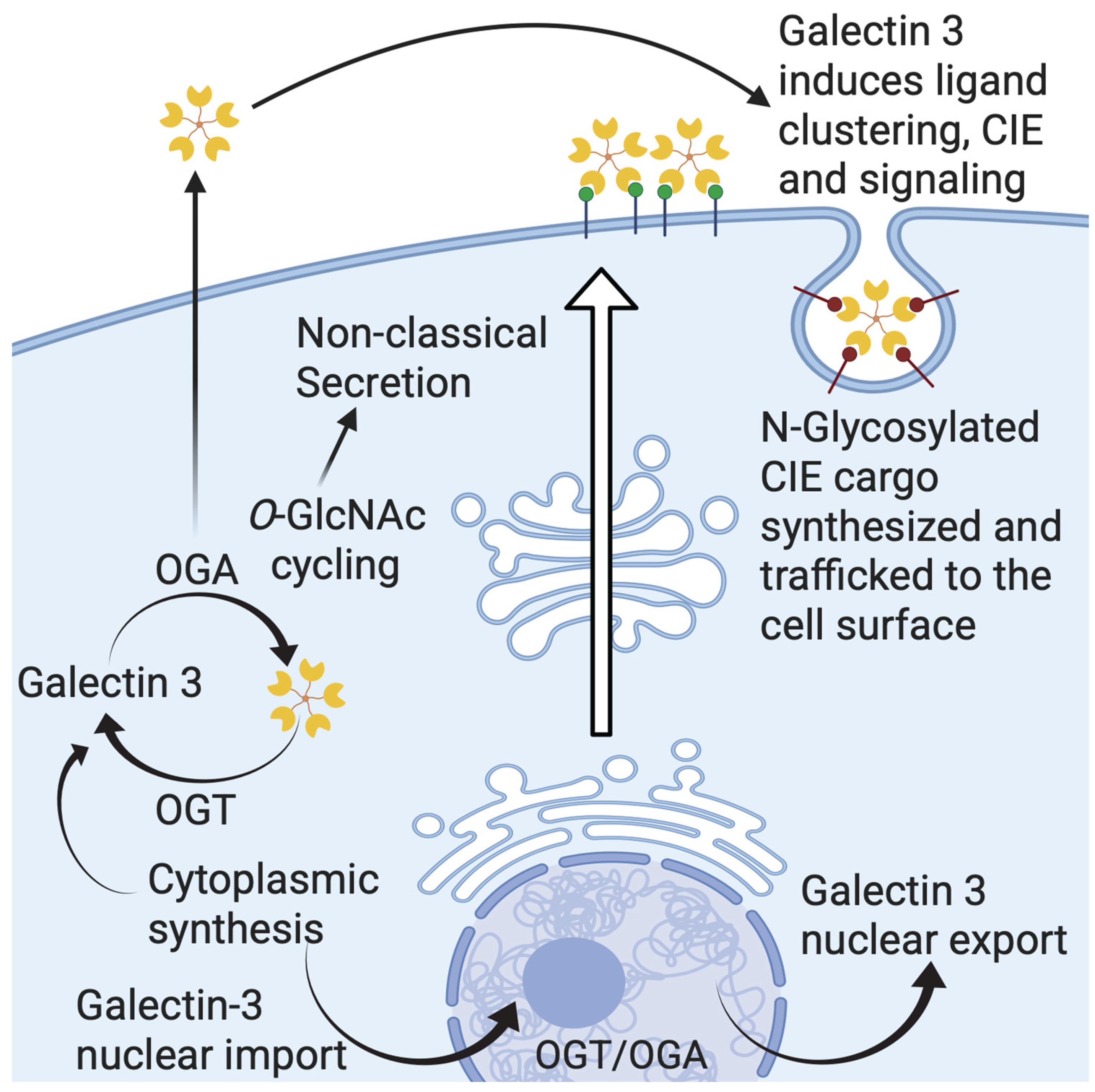
Figure 7.
Galectin-3 provides synergy between nutrient-driven O-GlcNAcylation and surface glycan signaling. At reduced intracellular concentrations of UDP-GlcNAc, dysregulation of the O-GlcNAcylation cycle may occur, leading to perturbations in galectin-3 transcriptional control and its subsequent secretion. These alterations can manifest across the physiologically relevant range of cytoplasmic concentrations, which are tightly regulated via feedback inhibition of the hexosamine biosynthetic pathway. Due to the activity of specific nucleotide sugar transporters, UDP-GlcNAc concentrations within the ER and Golgi apparatus are substantially elevated, as evidenced by the higher substrate thresholds required for extensive N-glycan branching and mucin-type O-glycan biosynthesis. At elevated UDP-GlcNAc concentrations, low-affinity N-glycosylation sites are modified, leading to the synthesis of low-occupancy N-glycans. The threshold-dependent changes illustrated suggest that as N-glycan synthesis and branching increase, galectin-3 expression and secretion are restrained to prevent excessive activation of galectin-3–mediated signaling pathways.
Figure 7.
Galectin-3 provides synergy between nutrient-driven O-GlcNAcylation and surface glycan signaling. At reduced intracellular concentrations of UDP-GlcNAc, dysregulation of the O-GlcNAcylation cycle may occur, leading to perturbations in galectin-3 transcriptional control and its subsequent secretion. These alterations can manifest across the physiologically relevant range of cytoplasmic concentrations, which are tightly regulated via feedback inhibition of the hexosamine biosynthetic pathway. Due to the activity of specific nucleotide sugar transporters, UDP-GlcNAc concentrations within the ER and Golgi apparatus are substantially elevated, as evidenced by the higher substrate thresholds required for extensive N-glycan branching and mucin-type O-glycan biosynthesis. At elevated UDP-GlcNAc concentrations, low-affinity N-glycosylation sites are modified, leading to the synthesis of low-occupancy N-glycans. The threshold-dependent changes illustrated suggest that as N-glycan synthesis and branching increase, galectin-3 expression and secretion are restrained to prevent excessive activation of galectin-3–mediated signaling pathways.
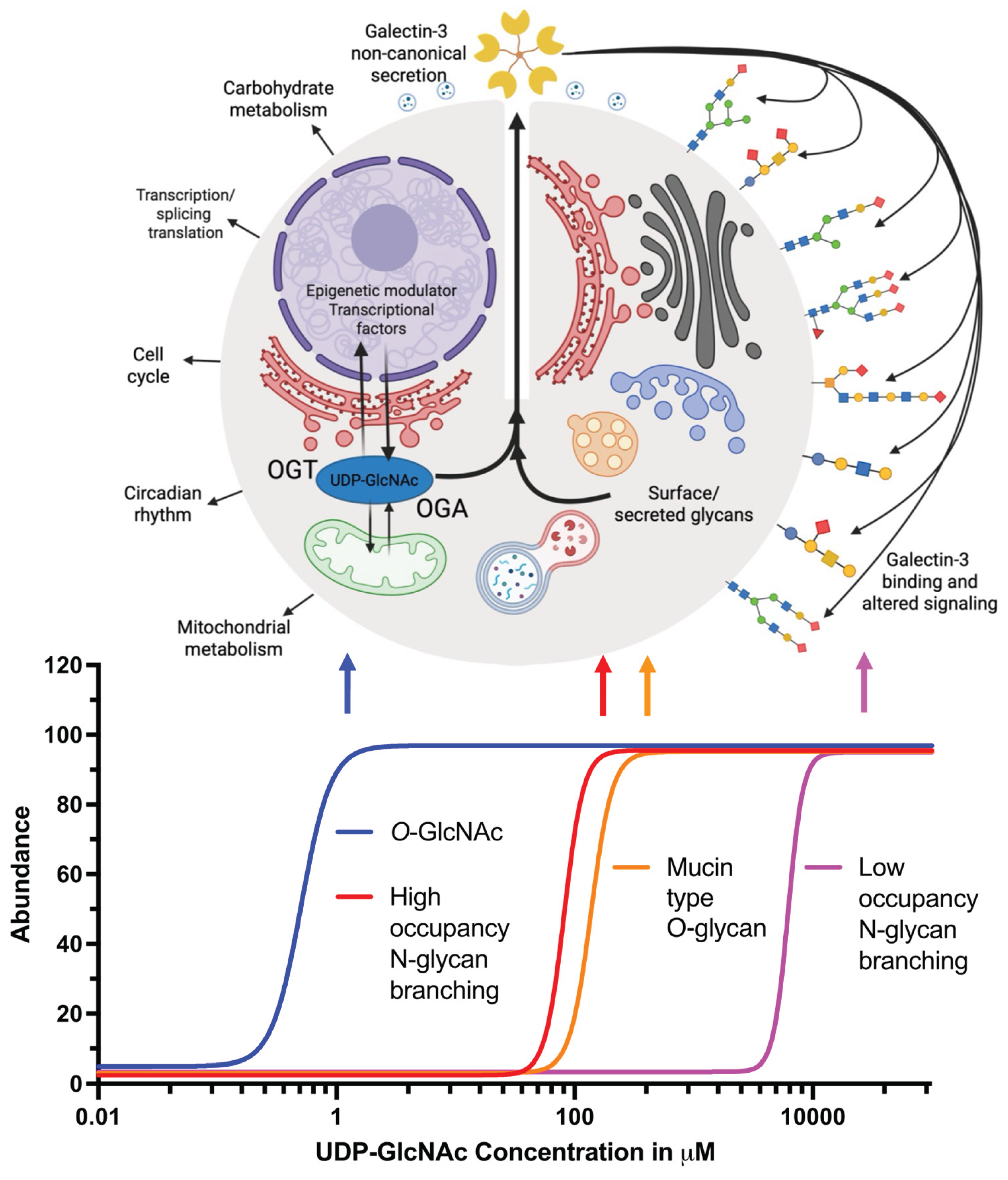
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



