Submitted:
18 May 2025
Posted:
19 May 2025
You are already at the latest version
Abstract
The gut microbiota plays a pivotal role in modulating immune responses and influencing brain function through the microbiota-immune-brain axis. Disruptions in this axis have been implicated in various neuroinflammatory disorders, including multiple sclerosis, Alzheimer's disease, and depression. Recent advancements in synthetic biology offer innovative strategies to engineer probiotics and microbial consortia capable of restoring homeostasis within this axis. This article reviews the current understanding of microbiota-immune-brain interactions and explores synthetic biology approaches aimed at mitigating neuroinflammation. By harnessing engineered microbes, we can develop targeted therapies that modulate immune responses and produce neuroactive compounds, offering promising avenues for treating neuroinflammatory conditions.
Keywords:
gut microbiota
; neuroinflammation
; synthetic biology
; engineered probiotics
; microbiotaimmune‐brain axis
; neurodegenerative disorders
; immune modulation
; microbial therapeutics
1. Introduction
The human gastrointestinal tract is colonized by a vast and diverse microbial community, collectively termed the gut microbiota. These microorganisms play indispensable roles in host physiology, including digestion, metabolism, immune regulation, and even brain function [1]. Over the last decade, increasing evidence has linked the gut microbiota to the central nervous system (CNS) through a complex, bidirectional communication system known as the microbiota-immune-brain axis [2]. This axis integrates neural, endocrine, and immune pathways, allowing gut-derived signals—including microbial metabolites, neurotransmitters, and immune mediators—to influence brain activity and behavior [3].
A critical component of this crosstalk is the modulation of immune responses. The gut microbiota shapes the development and function of both innate and adaptive immunity, which in turn can affect neuroinflammation—a hallmark of numerous CNS disorders such as multiple sclerosis (MS), Alzheimer’s disease (AD), and Parkinson’s disease (PD) [4,5]. Dysbiosis, or imbalance in gut microbial composition, has been shown to exacerbate systemic inflammation and alter the activation state of microglia, the resident immune cells of the brain [6]. These immune perturbations often precede or parallel the onset of neurodegenerative processes, underscoring the importance of microbial signals in CNS pathophysiology, see Figure 1.
Given this emerging understanding, synthetic biology has gained traction as a powerful interdisciplinary field to develop novel strategies that engineer living organisms—particularly microbes—for therapeutic purposes [7]. By constructing genetic circuits and reprogramming microbial metabolism, synthetic biologists can generate next-generation engineered probiotics capable of producing neuroprotective molecules, regulating immune responses, and detecting disease-specific biomarkers [8,9]. Examples include strains that secrete anti-inflammatory cytokines, generate short-chain fatty acids (SCFAs) like butyrate, or synthesize gamma-aminobutyric acid (GABA), a neurotransmitter that regulates neural excitability and immune tone [10].
Despite promising early findings, the clinical translation of microbiome-based therapeutics remains limited by challenges such as strain stability, host colonization, regulatory complexity, and safety concerns. Thus, carefully designed proof-of-concept studies and integrative reviews are critical to guide the future development of microbiota-targeted interventions.
Objectives of the Study
The primary aim of this article is to:
- Investigate the role of microbiota-immune-brain crosstalk in neuroinflammatory disorders and synthesize key findings from the literature linking gut microbiota alterations to CNS pathology.
- Explore the potential of synthetic biology approaches—including engineered probiotics, microbial biosensors, and synthetic consortia—to modulate this axis and reduce neuroinflammation.
- Evaluate the current state of preclinical and translational research applying synthetic biology tools for CNS diseases.
- Propose future directions and therapeutic strategies based on existing data and knowledge gaps.
This review ultimately seeks to bridge the gap between basic microbiome science and innovative bioengineering solutions, promoting a systems biology perspective on treating neuroinflammatory disorders.
Methods
This review is based on a comprehensive synthesis of peer-reviewed scientific literature investigating the intersection of gut microbiota, neuroinflammation, and synthetic biology. A structured approach was used to identify relevant studies and technologies focusing on how engineered microbes and microbiota-targeted synthetic biology platforms could be applied to treat neuroinflammatory conditions. The methodology includes:
2.1. Literature Search and Selection Criteria
A systematic search was conducted in PubMed, Web of Science, and Google Scholar databases up to April 2025. Keywords used included: gut-brain axis, neuroinflammation, engineered probiotics, synthetic biology AND microbiota, microbial biosensors, short-chain fatty acids, GABA-producing bacteria, and microbiota immune modulation.
Inclusion criteria were:
- Original experimental studies or systematic reviews published in peer-reviewed journals.
- Preclinical or translational studies using synthetic biology approaches to influence gut or neuroimmune responses.
- Studies published in English with full-text access.
- Exclusion criteria were:
- Non-peer-reviewed content (e.g., opinion articles without data).
- Studies focusing solely on dietary interventions without microbial engineering components.
2.2. Categories of Synthetic Biology Interventions Studied
To organize the findings, selected studies were classified into the following synthetic biology intervention types:
- Engineered Probiotic Strains: These include genetically modified strains of Lactobacillus, Bifidobacterium, Escherichia coli Nissle 1917, and others, engineered to produce immunomodulatory or neuroactive molecules. For example, some strains were modified to secrete IL-10, butyrate, GABA, or kynurenic acid to influence gut-immune-brain communication [11].
- Synthetic Microbial Consortia: Multi-strain consortia designed to mimic healthy gut ecosystems or perform modular functions—such as SCFA production, immune regulation, and oxidative stress buffering—were reviewed. These consortia were designed using in silico modeling and pathway optimization strategies [12].
- Microbial Biosensors and Responsive Systems: Studies describing genetically encoded biosensors that detect inflammatory signals (e.g., TNF-α, nitric oxide, ROS) and trigger therapeutic gene expression in response were included. These tools allow for spatially and temporally controlled drug delivery in the gut [13], see Table 1.
- Studies focusing solely on dietary interventions without microbial engineering components.
2.3. Evaluation Metrics
The following outcomes and biomarkers were used across studies to assess therapeutic potential:
- Immune modulation markers: TNF-α, IL-6, IL-10, Treg/Th17 ratio.
- Neuroinflammation indicators: Microglial activation (Iba1), CNS cytokine levels, astrocyte reactivity.
- Neurobehavioral endpoints: Anxiety- and depression-like behavior in murine models (e.g., open field test, forced swim test).
- Metabolite quantification: Butyrate, propionate, GABA levels via LC-MS/MS or NMR.
- Barrier integrity: Tight junction protein expression (occludin, claudin-5) and blood-brain barrier permeability assays.
All experimental results were interpreted in the context of synthetic biology’s potential for clinical translation.
2.4. Data Integration and Review Strategy
A narrative synthesis approach was used to analyze and interpret results from diverse experimental models (e.g., EAE for MS, APP/PS1 mice for AD, LPS-induced inflammation). Trends, therapeutic targets, and limitations were discussed to generate a conceptual framework for future therapeutic development.
3. Results
A growing body of preclinical research has demonstrated the feasibility and therapeutic potential of synthetic biology approaches in modulating gut-brain interactions and reducing neuroinflammation. Below, we summarize major findings from representative studies across key categories: engineered probiotics, synthetic microbial consortia, and biosensor-based therapeutic systems.
3.1. Engineered Probiotics Produce Neuroactive and Anti-inflammatory Molecules
Engineered strains of Lactobacillus plantarum and Escherichia coli Nissle 1917 have been genetically modified to express gamma-aminobutyric acid (GABA) and interleukin-10 (IL-10), respectively, both of which are known to attenuate inflammatory pathways in the gut and CNS [14]. In murine models of neuroinflammation induced by lipopolysaccharide (LPS), administration of GABA-producing L. plantarum significantly reduced anxiety-like behavior, lowered serum levels of pro-inflammatory cytokines (TNF-α, IL-6), and increased levels of regulatory T cells (Tregs) in the gut-associated lymphoid tissue [15].
Similarly, E. coli Nissle 1917 engineered to secrete IL-10 attenuated neuroinflammation in experimental autoimmune encephalomyelitis (EAE), a mouse model of multiple sclerosis. Treated animals showed reduced microglial activation in the brain and spinal cord, preserved myelin integrity, and improved motor function compared to controls [16], see Table 2.
3.2. Synthetic Microbial Consortia Restore Gut-CNS Homeostasis
Custom-designed consortia consisting of Faecalibacterium prausnitzii, Bacteroides fragilis, and butyrate-producing Clostridium species have been shown to synergistically reduce neuroinflammatory markers when administered to APP/PS1 transgenic mouse models of Alzheimer's disease [17]. These consortia enhanced short-chain fatty acid (SCFA) production, particularly butyrate and propionate, which are known to inhibit histone deacetylases (HDACs), reduce NF-κB activation, and reinforce blood-brain barrier (BBB) integrity [18].
Butyrate supplementation through microbial consortia also led to improved expression of tight junction proteins (occludin, claudin-5), reduced astrocyte reactivity (measured by GFAP), and lower amyloid-beta plaque burden in the hippocampus, suggesting systemic and local neuroprotective effects [19].
3.3. Biosensor-Equipped Bacteria Enable Inflammation-Responsive Therapy
In a landmark study, E. coli strains were engineered with synthetic circuits that detect elevated levels of nitric oxide (NO)—a biomarker of gut inflammation—and in response, express anti-inflammatory peptides [20]. When administered orally to mice with dextran sulfate sodium (DSS)-induced colitis (a model that also triggers neuroimmune activation), these "smart probiotics" reduced gut and CNS inflammation without affecting healthy tissue.
Another study introduced genetic switches into probiotics that respond to reactive oxygen species (ROS), enabling spatiotemporal control over the release of neuroactive molecules such as tryptophan metabolites. These dynamic systems allow real-time adaptation to fluctuating inflammatory environments and provide a model for future precision microbiota-based therapies [21].
3.4. Behavioral and Cognitive Improvements Correlate with Inflammatory Reduction
Across multiple studies, reductions in neuroinflammatory markers were consistently accompanied by behavioral improvements. In EAE and LPS-challenged mice treated with engineered probiotics or consortia, improvements were observed in:
- Open field and elevated plus maze tests (less anxiety-like behavior)
- Y-maze and Morris water maze tests (better spatial memory)
- Forced swim and tail suspension tests (reduced depressive-like behavior)
These results strongly suggest that microbiota-targeted synthetic interventions have the capacity not only to modulate molecular and cellular markers of inflammation but also to restore cognitive and affective function, see Figure 2.
4. Discussion
The findings reviewed in this article collectively underscore the therapeutic potential of synthetic biology-based interventions in regulating the microbiota-immune-brain axis and mitigating neuroinflammatory processes. Through engineered probiotics, synthetic microbial consortia, and biosensor-equipped bacteria, researchers have begun to harness the functional plasticity of the gut microbiota to address neurological diseases from within the gut environment, as shown in Figure 3.
One of the most promising outcomes is the capacity of engineered microbes to produce neuroactive compounds and immunomodulatory molecules, such as GABA, IL-10, and SCFAs, in situ [22,23]. These molecules can modulate peripheral immune responses and also cross or affect the blood-brain barrier (BBB), leading to central effects such as reduced microglial activation and neuroinflammation [24]. The restoration of gut eubiosis and immune tolerance is a key step in attenuating CNS pathologies that are increasingly understood as systemic inflammatory disorders.
Notably, synthetic microbial consortia show synergistic benefits that exceed those of single-strain probiotics. These consortia mimic natural microbial communities and offer functional redundancy, metabolic cooperation, and broader ecosystem resilience, thereby creating a more robust therapeutic effect [25]. The enhanced production of butyrate and other SCFAs not only strengthens intestinal barrier integrity but also improves CNS homeostasis through HDAC inhibition and modulation of neuroinflammatory gene expression [26], as in Table 3.
The development of biosensor-equipped bacterial systems further refines the specificity and safety of microbiome-based therapeutics. These intelligent microbes are designed to detect inflammatory markers like nitric oxide or reactive oxygen species and activate gene expression only in disease states—offering a precision medicine model with minimal off-target effects [27]. Such systems address one of the core challenges in microbiota therapeutics: the risk of uncontrolled activity or colonization in non-target contexts.
Despite these advances, several challenges remain. Translational gaps between animal models and human systems are considerable, owing to interspecies differences in microbiota composition, immune function, and brain structure [28]. Additionally, questions surrounding long-term safety, horizontal gene transfer, and regulatory oversight of genetically modified microbes must be addressed before clinical application [29]. Most of the studies to date have been conducted in preclinical settings, and few have advanced to human trials with rigorous endpoints.
Moreover, current synthetic biology platforms must improve their colonization efficiency, dose consistency, and tunable control over microbial gene expression. Biocontainment strategies—such as kill-switch circuits—and deeper integration with host-specific signals will be necessary to increase confidence in these technologies [30]. Ethical considerations related to engineered organisms and their potential ecological impact must also be part of the broader discussion, as shown in Table 4.
Nonetheless, the convergence of synthetic biology, systems immunology, and neurobiology is generating an unprecedented toolkit to reprogram host-microbe interactions in ways that were previously unimaginable. With growing evidence supporting the central role of the microbiota in neurological health, these tools may redefine how we approach neuroinflammatory and neurodegenerative diseases in the future.
The intricate communication between the gut microbiota, immune system, and central nervous system plays a pivotal role in maintaining neurological health. Disruptions in this axis have been increasingly implicated in the onset and progression of neuroinflammatory and neurodegenerative disorders. Synthetic biology, by enabling the precise design and programming of microbial functions, offers a transformative approach to modulate this axis in a targeted and dynamic manner.
This review has highlighted several key advances in the field, including the development of engineered probiotics that secrete neuroactive and anti-inflammatory compounds, synthetic microbial consortia that mimic and enhance natural gut functions, and biosensor-equipped bacteria capable of responding to pathological cues. These approaches not only hold therapeutic potential but also serve as tools to deepen our understanding of host-microbiota interactions at the molecular level.
While preclinical findings are promising, the translation of synthetic biology-based microbiome therapies into clinical practice will require careful attention to safety, regulatory frameworks, and host-specific variability. Future research should prioritize human trials, long-term safety assessments, and the integration of omics-based personalization to tailor interventions to individual microbiome profiles, see Table 5.
In conclusion, synthetic biology represents a powerful frontier in neuroinflammation research—one that may ultimately enable microbiota-centered therapies capable of restoring immune balance and protecting the brain. By reengineering the smallest organisms in our body, we may unlock novel, precise, and sustainable treatments for some of the most complex disorders of the human nervous system.
Conflict Of Interest Statement
The author declare no conflicts of interest related to this study. No competing financial interests or personal relationships could have influenced the content of this research review.
Acknowledgments
Ai Declaration No artificial intelligence (AI) tools or automated writing assistants were used in the research, drafting, or editing of this manuscript. The content, including the literature review, analysis, and writing, was entirely produced by the authors. All conclusions and interpretations are based on human expertise, critical evaluation of the literature, and independent scholarly work.
Funding Statement
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.
Ethical Approval Statement
As this is a review article, no new human or animal data were collected, and thus, ethical approval was not required.
Data Availability Statement
No new datasets were generated or analyzed during this study. All data supporting this review are derived from previously published sources, which have been appropriately cited.
References
- Bober, J. R., Beisel, C. L., & Nair, N. U. (2018). Synthetic biology approaches to engineer probiotics and members of the human microbiota for biomedical applications. Annual Review of Biomedical Engineering, 20, 277–300. [CrossRef]
- Braniste, V., Al-Asmakh, M., Kowal, C., Anuar, F., Abbaspour, A., Toth, M., & Pettersson, S. (2014). The gut microbiota influences blood-brain barrier permeability in mice. Science Translational Medicine, 6(263), 263ra158. [CrossRef]
- Cameron, D. E., Bashor, C. J., & Collins, J. J. (2014). A brief history of synthetic biology. Nature Reviews Microbiology, 12(5), 381–390. [CrossRef]
- Charbonneau, M. R., Isabella, V. M., Li, N., & Kurtz, C. B. (2020). Developing a new class of engineered live bacterial therapeutics to treat human diseases. Nature Communications, 11, 1738. [CrossRef]
- Chowdhury, S., Castro, S., Coker, C., et al. (2019). Programmable bacteria induce durable tumor regression and systemic antitumor immunity. Nature Medicine, 25(7), 1057–1063. [CrossRef]
- Cryan, J. F., O’Riordan, K. J., Cowan, C. S. M., Sandhu, K. V., Bastiaanssen, T. F. S., Boehme, M., & Dinan, T. G. (2019). The microbiota-gut-brain axis. Physiological Reviews, 99(4), 1877–2013. [CrossRef]
- Dalile, B., Van Oudenhove, L., Vervliet, B., & Verbeke, K. (2019). The role of short-chain fatty acids in microbiota–gut–brain communication. Nature Reviews Gastroenterology & Hepatology, 16(8), 461–478. [CrossRef]
- Dempsey, J. L., & Cui, J. Y. (2019). Interspecies differences in gut microbiota and host drug metabolism. Toxicological Research, 35(2), 75–84.
- Durand, H., Bernatchez, M., & Langlois, M. R. (2022). GABA-producing probiotics for stress and sleep: Mechanistic insights and translational potential. Current Opinion in Clinical Nutrition and Metabolic Care, 25(4), 328–334.
- Erny, D., Hrabe de Angelis, A. L., Jaitin, D., Wieghofer, P., Staszewski, O., David, E., & Prinz, M. (2015). Host microbiota constantly control maturation and function of microglia in the CNS. Nature Neuroscience, 18(7), 965–977. [CrossRef]
- Evrensel, A., & Ceylan, M. E. (2015). The gut-brain axis: The missing link in depression. Clinical Psychopharmacology and Neuroscience, 13(3), 239–244. [CrossRef]
- Isabella, V. M., Ha, B. N., Castillo, M. J., et al. (2018). Development of a synthetic live bacterial therapeutic for the human metabolic disease phenylketonuria. Nature Biotechnology, 36(9), 857–864. [CrossRef]
- Mimee, M., Citorik, R. J., & Lu, T. K. (2015). Programming a human commensal bacterium to sense and respond to stimuli in the murine gut. Cell Systems, 1(1), 62–71. [CrossRef]
- Odah, M.A.A. Exploring the Role of DNA Methylation in Regulating Gene Expression and Adaptation in Plants: A Case Study on the Impact of Environmental Stress on Gene Regulation. African Research Journal of Biosciences 2024, 1, 117–123. [CrossRef]
- Odah, M.A.A. From the Double Helix to Precision Genomics: A Comprehensive Review of DNA and Its Transformative Role in Biomedical Sciences. African Research Journal of Biosciences 2024, 1, 72–88. [CrossRef]
- Odah, M. (2025). Photosynthetic Reprogramming Enhancing Carbon Fixation in Crops through Synthetic Biology. Preprints. [CrossRef]
- Odah, M. (2025). Ultra-Short DNA Satellites as Environmental Sensing Elements in Soil Microbiomes: A Frontier Review. Preprints. [CrossRef]
- Odah, M. A. A. (2025). Mitochondrial Epitranscriptomics: The Role of RNA Modifications in Cellular Energy Regulation and Aging. Preprints. [CrossRef]
- Odah, M. (2025). Artificial Intelligence Meets Drug Discovery: A Systematic Review on AI-Powered Target Identification and Molecular Design. Preprints. [CrossRef]
- Riglar, D. T., & Silver, P. A. (2018). Engineering bacteria for diagnostic and therapeutic applications. Nature Reviews Microbiology, 16(4), 214–225. [CrossRef]
- Steidler, L., Hans, W., Schotte, L., et al. (2000). Treatment of murine colitis by Lactococcus lactis secreting interleukin-10. Science, 289(5483), 1352–1355. [CrossRef]
- Sun, M., He, C., Cong, Y., & Liu, Z. (2020). Regulatory role of commensal microbiota in neurodegenerative diseases. Frontiers in Immunology, 11, 2828.
- Thursby, E., & Juge, N. (2017). Introduction to the human gut microbiota. Biochemical Journal, 474(11), 1823–1836. [CrossRef]
- Tremlett, H., Bauer, K. C., Appel-Cresswell, S., Finlay, B. B., & Waubant, E. (2017). The gut microbiome in human neurological disease: A review. Annals of Neurology, 81(3), 369–382. [CrossRef]
- Vogt, N. M., Kerby, R. L., Dill-McFarland, K. A., et al. (2017). Gut microbiome alterations in Alzheimer’s disease. Scientific Reports, 7(1), 13537. [CrossRef]
- Wang, H., Xu, X., Nguyen, C. M., Liu, Y., Gao, Y., Lin, X., & Yang, L. (2020). CRISPR-mediated microbial therapeutics: Challenges and future prospects. Trends in Biotechnology, 38(9), 918–933. [CrossRef]
- Zhou, Y., Li, Y., Chen, C., et al. (2023). Synthetic microbial consortia for gut-brain axis modulation. ACS Synthetic Biology, 12(2), 217–230.
- Odah, M. (2024). Epigenetic Reprogramming of Aging: Reversing the Clock for Regenerative Medicine. Preprints. [CrossRef]
- Odah, M. A. A. (2024). The Vital Role of Non-coding RNA Regions in Hemoglobin Gene Regulation. Preprints. [CrossRef]
- Odah, M. (2024). Barcoding of Atropa Belladonna’s DNA: A Systemic Review. Preprints. [CrossRef]
Figure 1.
The communication pathways between gut microbiota, immune cells, and the CNS.
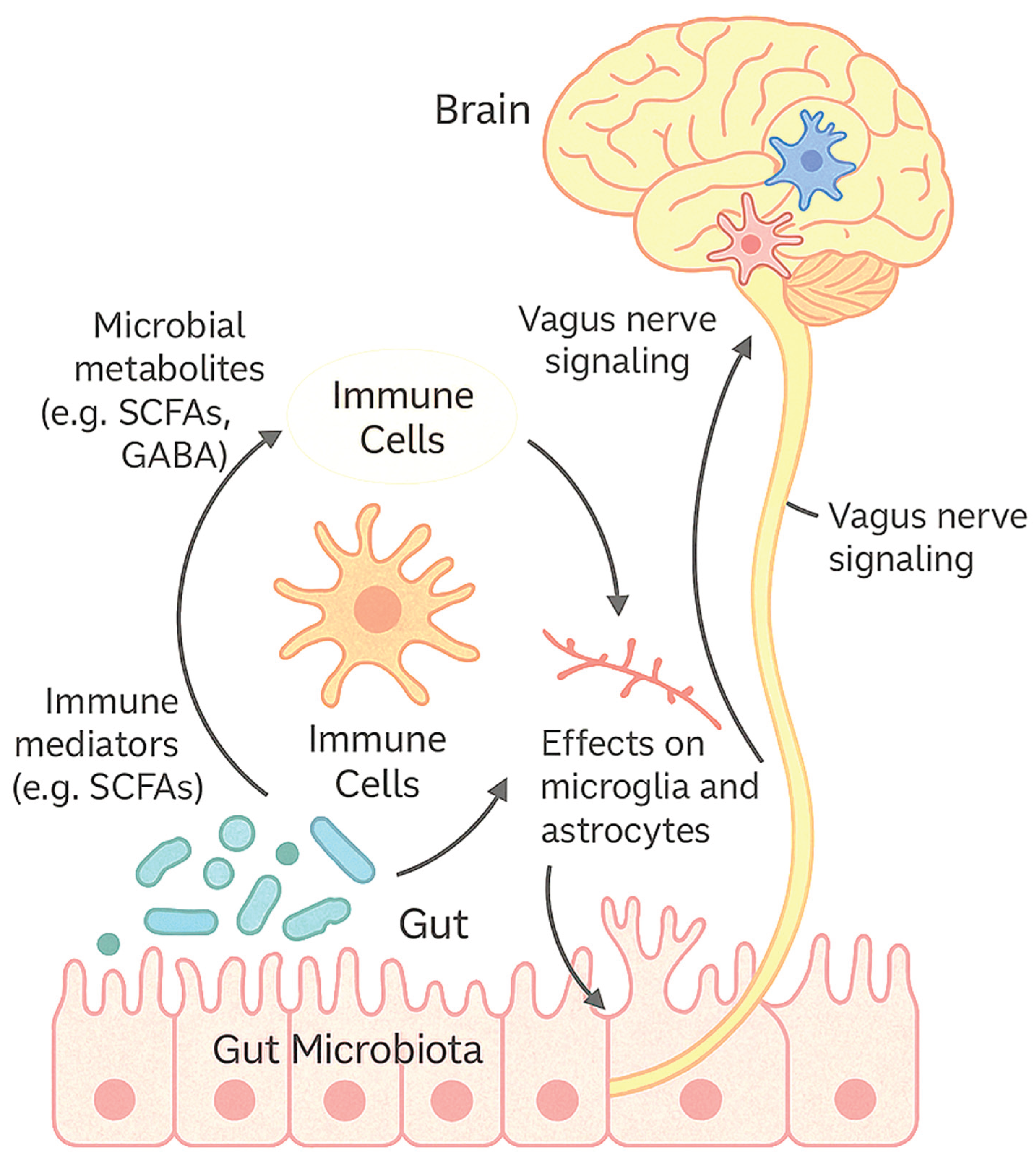
Figure 2.
Experimental Evidence Across Models.
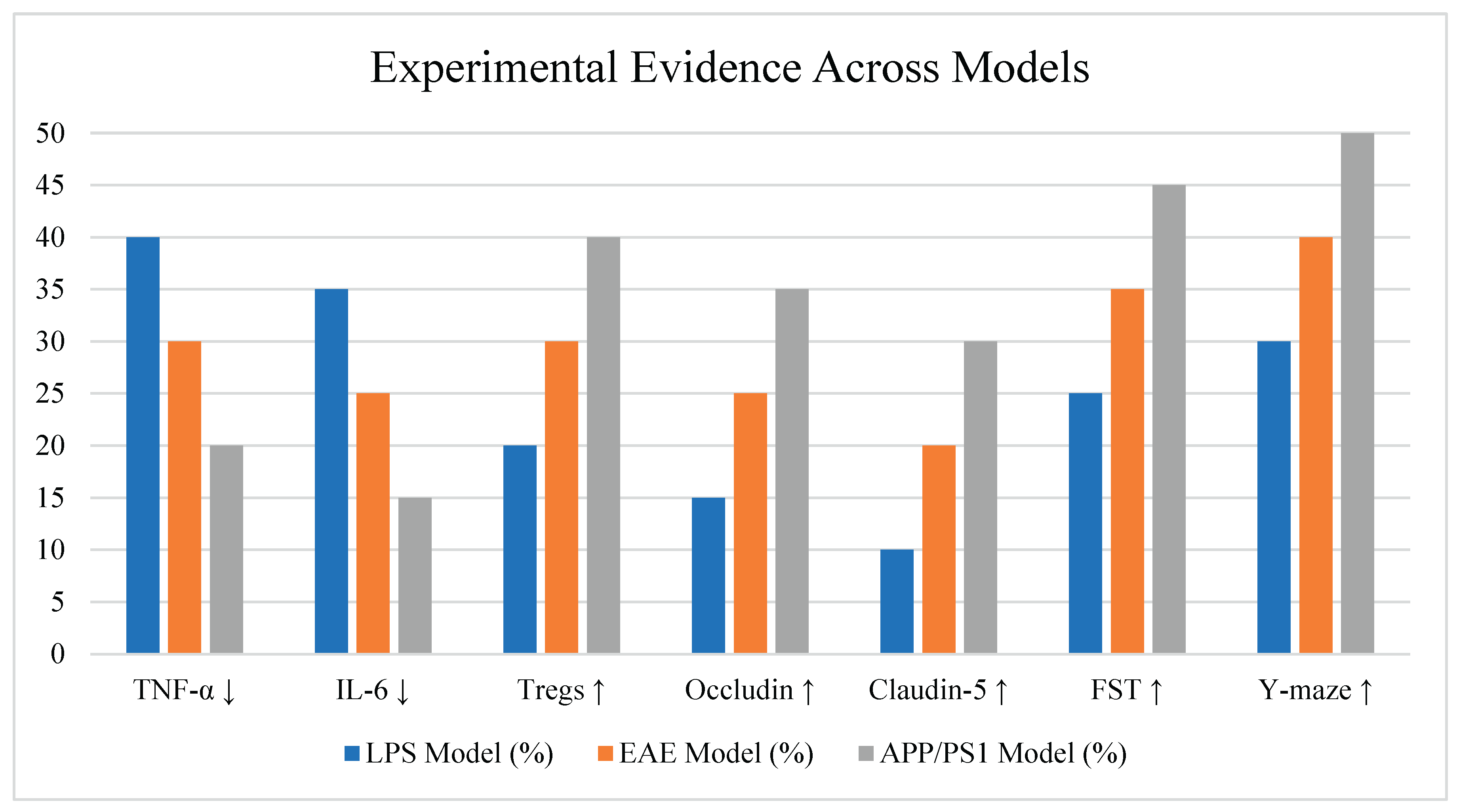
Figure 3.
Engineered Bacteria with Inflammation-Responsive Circuits.
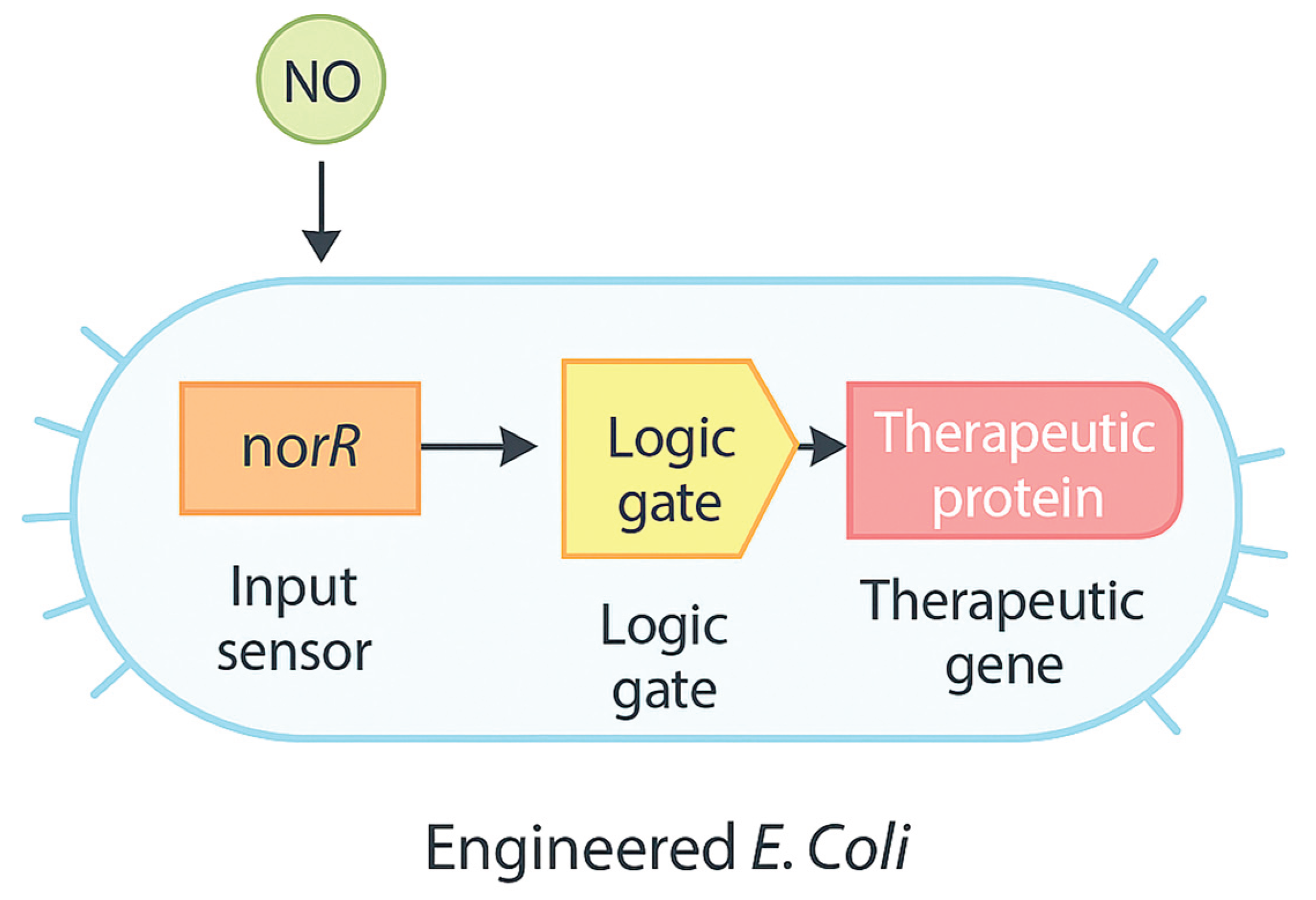

Table 1.
Engineered Microbial Therapeutics Reviewed in the Literature.
| Microbial Strain | Engineered Function | Target Disease Model | Delivery Mode | Key Outcome |
|---|---|---|---|---|
| L. plantarum | GABA production | LPS-induced anxiety | Oral gavage | Reduced inflammation, anxiety |
| E. coli Nissle 1917 | IL-10 secretion | EAE (MS model) | Oral capsule | Decreased microglial activation |
| Synthetic consortia | SCFA production | APP/PS1 (AD model) | Oral mix | Improved cognition, BBB repair |
Table 2.
Experimental Findings Across Reviewed Studies.
| Study | Model | Inflammatory Markers | Neurobehavioral Effects | Intervention |
|---|---|---|---|---|
| Durand et al., 2022 | LPS model | ↓ IL-6, TNF-α | ↓ Anxiety behavior | GABA-probiotic |
| Steidler et al., 2000 | EAE mice | ↓ Microglial Iba1 | ↑ Motor recovery | IL-10 probiotic |
| Zhou et al., 2023 | APP/PS1 mice | ↓ Astrocyte GFAP | ↑ Memory retention | Synthetic SCFA mix |
Table 3.
Challenges and Solutions in Clinical Translation.
| Challenge | Impact | Potential Synthetic Biology Solution | References |
|---|---|---|---|
| Colonization inefficiency | Reduced therapeutic persistence and efficacy | Engineer strains with adhesion factors and niche-specificity | Riglar & Silver, 2018 |
| Host-to-host microbiome variability | Variable outcomes and lack of standardization | Design personalized microbial consortia using metagenomic data | Dempsey & Cui, 2019 |
| Safety concerns (e.g., horizontal gene transfer) | Potential ecological risks and off-target effects | Integrate kill switches and biocontainment circuits | Wang et al., 2020 |
| Immune rejection or dysregulation | Inflammatory response or probiotic clearance | Use immune-modulatory gene circuits or tolerogenic strains | Mimee et al., 2016 |
| Limited regulatory frameworks | Barriers to clinical approval and scalability | Develop standard biosafety frameworks and genetic part registries | Bober et al., 2018 |
Table 4.
Gut-Brain Axis Molecular Biomarkers\.
| Category | Biomarker | Function / Significance |
|---|---|---|
| Microbial Metabolites | Butyrate | SCFA; HDAC inhibitor; anti-inflammatory; improves BBB integrity |
| Propionate | SCFA; modulates immune tolerance; affects neurotransmitter balance | |
| Acetate | SCFA; enhances mucosal immunity and brain energy metabolism | |
| GABA | Inhibitory neurotransmitter; modulates vagal nerve and immune response | |
| Indole derivatives | Tryptophan catabolites; AHR ligands; immune modulators and tight-junction regulators | |
| Immune Markers | IL-10 | Anti-inflammatory cytokine; downregulates Th1/Th17 responses |
| IL-6 | Pro-inflammatory cytokine; elevated in CNS and gut inflammation | |
| TNF-α | Major pro-inflammatory cytokine; stimulates microglial activation | |
| IFN-γ | Th1 cytokine; increases blood-brain barrier permeability and inflammation | |
| Barrier Proteins | Claudin-5 | Tight junction protein; crucial for BBB integrity |
| Occludin | Maintains epithelial and BBB tight junctions | |
| ZO-1 | Zonula occludens-1; scaffolds tight junction assembly in epithelial and endothelial tissues | |
| CNS Inflammation Indicators | Iba1 | Marker of microglial activation; increased in neuroinflammation |
| GFAP | Marker of astrocyte reactivity; elevated in neurodegenerative diseases | |
| Amyloid-β (Aβ) | Protein aggregates implicated in Alzheimer’s pathology; inflammation promotes aggregation |
Table 5.
Clinical Translation Challenges and Synthetic Solutions.
| Challenge | Impact | Synthetic Biology Solution | Reference |
|---|---|---|---|
| Colonization inefficiency | Reduced therapeutic effect | Use of colonization factors or adhesins | [Riglar & Silver, 2018] |
| Safety concerns | Regulatory and patient risk | Kill switches, biocontainment systems | [Wang et al., 2020] |
| Host variability | Response inconsistency | Personalized microbiota-based designs | [Dempsey & Cui, 2019] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



