Submitted:
19 May 2025
Posted:
19 May 2025
You are already at the latest version
Abstract
This study aimed to evaluate clinical signs, visceral changes, and weight development in intermediate hosts infected with Cystoisospora ohioensis-like oocysts from naturally infected dogs. Using 135 albino mice divided into three groups—INFECTED (inoculated with oocysts), PAIR-FED (given the same food as infected mice), and CONTROL (fed ad libitum)—researchers monitored weight and pathological changes over 35 days. Mice were euthanized at various intervals post-inoculation (1–35 days), with organs from some INFECTED mice fed to dogs on day 60 for a biological assay. The INFECTED group showed higher body weight and weight gain than the CONTROL group, though organ weights were greater, leading to lower carcass yield. Tissue cysts with hypnozoites were found in the intestines (1st day), Peyer’s patches (up to 5th day), and lymph nodes/spleen (up to 35th day). Dogs inoculated with hypnozoites (from infected mice) had a shorter prepatent period, longer patent period, and higher oocyst shedding compared to those given sporulated oocysts. The findings indicate that C. ohioensis-like species affect mouse weight, persist in visceral tissues for up to 60 days, and exhibit biological differences in infectivity between sporozoites and hypnozoites in dogs.
Keywords:
cystoisosporosis
; oocysts
; hypnozoites
; host infectivity
1. Introduction
The genus Cystoisospora was established by Frenkel [1] to accommodate certain species formerly classified under Isospora, characterized by sporulated oocysts containing two sporocysts with four sporozoites each, the absence of Stieda bodies in the sporocysts, and the ability to form monzoic tissue cysts (hypnozoites) in intermediate hosts. Smith [2], who recognized the involvement of an intermediate host, proposed placing Cystoisospora within the subfamily Cystoisosporinae under the family Sarcocystidae. Molecular studies subsequently confirmed the classification of mammalian Cystoisospora species within the Sarcocystidae family [3,4,5,6].
Within the subfamily Cystoisosporinae, the genus Cystoisospora comprises four species for which the dog is the definitive host: C. canis, C. ohioensis, C. burrowsi, and C. rivolta. Among these, C. canis is distinct because of its large, ovoid oocysts (28–34 µm), whereas the latter three, with intermediate-sized oocysts (17–23 µm), are considered C. ohioensis-like owing to their overlapping size ranges [7,8]. The third group, consisting of small oocysts (10–13 µm) shed in the feces of infected dogs, includes two members of the subfamily Toxoplasmatinae: Neospora caninum, which is clinically significant, and Heydornia heydorni (syn. Hammondia heydornii), which is nonpathogenic [9]. The developmental stages of these parasites are primarily found in the digestive tracts of young animals, which become infected by ingesting sporulated oocysts from the environment or monzoic cysts present in the viscera of previously infected intermediate hosts [10]. Cystoisospora ohioensis has been characterized as pathogenic when experimentally inoculated into puppies, which subsequently develop diarrhea as a clinical manifestation [11,12]. Additionally, diarrhea associated with C. ohioensis has been reported in a naturally infected three-month-old puppy in South Korea, with the diagnosis confirmed by PCR [13].
Cystoisospora ohioensis is widely distributed and fairly common in epithelial cells throughout the small intestine as well as in the cecum and colon of dogs, coyotes, red foxes, raccoons, and possibly dingoes [14]. Dogs fed tissues from camels [15,16], sheep [15], pigs, donkeys, and buffaloes [17] naturally shed C. ohioensis oocysts in their feces; however, there has been no microscopic confirmation of monzoic tissue cysts. Experimentally, intermediate hosts include mice, rats, golden hamsters, cats, and dogs, in which monzoic cysts have been found in viscera, mainly mesenteric lymph nodes, liver, and spleen [14]. Frenkel and Dubey [18], in describing mice as intermediate hosts of Cystoisospora species in felines, characterized the formation of merozoites through endodyogeny as an asexual reproductive process occurring in the mucosa of the small intestine of intermediate hosts. Subsequent studies led Dubey and Lindsay [8] to suggest that the two zoites likely enter the same host cell rather than resulting from the division of a single zoite. The monzoic tissue cysts of C. felis, C. rivolta, and C. ohioensis increase in size over time but do not undergo division [10,18]. Thus, the formation of monzoic cysts is considered one of the distinguishing characteristics of Cystoisospora species [1], a feature also observed in the asexual stages of Cystoisospora spp. in mouse tissues [19].
There is limited information regarding the economic significance of species within this genus, although the presence of the parasite has been reported in the tissues of naturally infected livestock [15,17] as well as in experimentally infected livestock [20,21,22,23]. Pathologies associated with monzoic cysts have been reported for the genus Cystoisospora in humans, particularly in cases involving Cystoisospora belli, the causative agent of human cystoisosporosis [24]. This organism has been identified as an opportunistic etiological agent in patients with acquired immunodeficiency syndrome (AIDS) and has been associated with chronic diarrhea, acalculous cholecystitis, and cholangiopathy [25,26]. Velásquez et al. [27] reported that in this patient group, infection by C. belli may lead to disseminated cystoisosporosis, with the presence of monzoic tissue cysts in the intestinal lamina propria, lymph nodes, liver, and spleen [28,29,30,31,32].
To diagnose coccidiosis in dogs, it is necessary to detect oocysts through microscopic examination [14] or identify the developmental stages of the parasite in mucosal scrapings during the acute phase of infection [33,34]. Lee et al. [13] examined the feces of a three-month-old female dog with persistent diarrhea using real-time multiplex polymerase chain reaction (PCR), which targeted 23 pathogens known to cause diarrheal syndromes. Sequence analysis was performed using nested PCR amplification of 18S ribosomal RNA. Oocysts were identified in the fecal smear. Although real-time multiplex PCR was positive for Cyclospora cayetanensis, the final diagnosis was C. ohioensis infection, which was confirmed through phylogenetic analysis of the 18S rRNA gene. Another relevant aspect is the difficulty in isolating oocysts of C. canis [1,35] and C. ohioensis [1,36], as these species are most often shed in the feces of dogs along with other coccidia, such as C. burrowsi [37] Rommel & Zielasko, 1981 [38], C. rivolta [39] Frenkel, 1977 [1], as well as small oocysts of Heydornia heydorni (syn. Hammondia heydorni) [40], Neospora caninum [41], and some species of the genus Sarcocystis.
Thus, from the diagnosis of the shedding of subspherical, disporic, tetrazoic coccidian oocysts of medium size by six dogs from the same litter—morphologically and morphometrically characterized as belonging to the C. ohioensis-like group—the objective of this research was to experimentally infect mice, describe possible clinical signs of cystoisosporosis in these animals, and evaluate whether there are alterations in the viscera of the infected animals and whether cystoisosporosis influences the weight development of the intermediate host using an experimental mouse model.
2. Materials and Methods
This study is part of a doctoral thesis [42] and was conducted at the Protozoology Laboratory of the Experimental Station for Parasitological Research under Dr. Wilhelm Otto Neitz, the Department of Animal Parasitology, the Institute of Veterinary Medicine, the Federal Rural University of Rio de Janeiro (UFRRJ), Seropédica Campus, RJ, Brazil.
2.1. Origin of the Oocysts
Cystoisospora ohioensis-like oocysts were obtained from the feces of six eight-week-old puppies from the same litter. The animals were naturally infected with Cystoisospora spp. and lived freely in peridomestic urban areas of the city of Seropédica, State of Rio de Janeiro, Brazil. They were maintained in accordance with Law No. 6,638 of May 8, 1979, repealed by Law No. 11,794/2008, which regulates the licensing of activities related to the production, maintenance, or use of animals for education or scientific research in Brazil [43], and in accordance with CFMV Resolution No. 714/2002, which establishes authorization requirements for animal experimentation and requires that the method chosen must depend on the species involved, the available means for animal restraint, the technical ability of the practitioner, and the number of animals [44]. The animals were fed a commercial diet formulated for puppies provided ad libitum. Pens were cleaned daily and sanitized weekly using a flame broom. For 15 days, the feces were examined daily for oocyst detection and collection, according to Oliveira et al. [43].
2.2. Oocyst Sporulation
The collected fecal material was diluted at a ratio of 1:2 in a 2.5% potassium dichromate solution and aerated using a conventional aquarium pump (Alpha II, Vigor Ar, São Paulo) at room temperature. After 72 hours of aeration, a drop of the suspension was placed between a microscope slide and coverslip and examined under a light microscope. Once the sporulation index reached 80% or higher, the fecal suspensions were transferred to 50 mL Falcon-type centrifuge tubes and centrifuged at 200 × g for 10 minutes. The sporulated oocysts were recovered from the supernatant using flotation in a saturated sugar solution.
2.3. Oocyst Concentration
The sporulated oocysts were concentrated by resuspension in a saturated sugar solution followed by additional centrifugation (200 × g for 5 minutes). The resulting supernatant was poured into a 14 cm diameter Petri dish, over which the bottom of a 15 cm Petri dish was placed, creating an interface between the lid (the base of the 15 cm Petri dish) and the convergent meniscus of the sugar solution surface. At 15-minute intervals over one hour, the upper dish was removed so that its bottom could be rinsed with phosphate-buffered saline (PBS), pH 7.2, using a 10 mL syringe fitted with a hypodermic needle. The suspension of sporulated oocysts obtained from washing with PBS was collected using a funnel placed in a glass beaker. These oocysts were subjected to another round of centrifugation, followed by two consecutive resuspensions in PBS to eliminate potassium dichromate residue and excess sucrose. The final sediment, containing a high concentration of oocysts, was resuspended in a small volume of PBS and transferred to a new sterile 50 mL Falcon tube. The total volume of feces excreted by the dogs was concentrated to a final volume of 50 mL and stored at 4 °C until use.
2.4. Oocyst Counting and Morphometry
An aliquot of purified oocyst suspension was placed in a Neubauer chamber for counting. The average number from five counts of oocysts quantified in the four outer reticulated areas—the area used for leukocyte quantification—was multiplied by 2.5, which is the chamber height correction factor, corresponding to the number present in 1 mm³ of the material. To convert the number of oocysts per mL of the concentrated suspension, the number of oocysts found in 1 mm³ was multiplied by 1,000. Thus, the total number of oocysts recovered from all the samples collected from all the animals was equal to the number of oocysts quantified in one mL multiplied by the total volume of the concentrated suspension. The major diameter (MD) and minor diameter (md) of sporulated oocysts were measured one hundred times via a K-15X micrometer eyepiece (PZO, Poland) coupled with a Leitz Westler microscope (H.M. Lux, Germany). The morphometric indices (MIs) were obtained by the ratio of MD to md and were calculated for both sporulated oocysts and the sporocysts within each oocyst.
2.5. Standardization of the Inoculum and Infection of the Mice
For the concentrated sample, the total number of oocysts in the sample was previously evaluated, and the inoculum was standardized for each mouse to 10⁵ sporulated oocysts of the C. ohioensis-like group in 0.5 mL of PBS.
A total of 135 albino mice from the Central Animal Facility of Fundação Instituto Oswaldo Cruz (FIOCRUZ) were distributed into 27 polypropylene cages with five animals per cage. These animals were divided into three groups with nine cages and 45 mice per group. In one of the groups, the mice received the standardized inoculum orally using a gavage needle. This group was named INFECTED, and the animals were fed species-specific feed ad libitum. The other two groups, with a total of 90 mice, received 0.5 mL of PBS as the inoculum and served as experimental controls. One of these groups with 45 mice was fed ad libitum and called CONTROL, whereas the other 45 mice received, throughout the entire experiment, the same amount of feed consumed by their counterparts in the INFECTED group and was named the pair-fed group (PAIR FED). All 135 mice in the INFECTED, CONTROL and PAIR FED groups received water ad libitum throughout the experiment and were maintained in compliance with Law No. 6,638 of May 8, 1979 (revoked by Law No. 11,794/2008 – [44]), in accordance with CFMV Resolution No. 714/2002 [45].
2.6. Evaluation of Body Weight Development
The relative weights of the mice, whether infected or not, were proportionally assessed through daily weight measurements of each animal over 35 days. The influence of food intake on weight was evaluated by comparing the mean body weight of the infected group with that of the control groups.
Five mice from each group were euthanized in a CO₂ chamber at 1, 3, 5, 9, 14, 21, 28, 35, and 60 days after infection (DAI), in accordance with the subsequently established Law No. 11,794/2008 (which revoked Law No. 6,638 of May 8, 1979; [44]) and CFMV Resolution No. 714/2002 [45]. Animals euthanized between 1 and 35 DAI underwent necropsy and evisceration. Their organs—stomach, small intestine, large intestine, Peyer's patches, mesenteric lymph nodes, spleen, liver, lungs, and heart—were collected, identified, and weighed using a precision balance (Mettler PE 360, USA). For each subject, the proportional weight of the viscera relative to the live body weight was determined. Similarly, the carcass weight postevisceration was comparatively assessed. The organs of the mice euthanized at 60 DAI were collected for subsequent hypnozoite recovery.
2.7. Recovery of Hypnozoites from Organs
For each of the five mice euthanized at 60 DPI, their organs were collected, weighed, and individually minced into small fragments using a mortar and pestle. A 0.9% saline solution was then added to achieve a final homogenate volume of 5 mL, which was transferred to a 50 mL plastic centrifuge tube. Cystoisospora ohioensis-like hypnozoites were recovered according to the methodology described by Dubey [46], with slight modifications as proposed by Oliveira et al. [47].
2.8. Infectivity of Sporozoites and Hypnozoites
To determine the viability of inocula containing 10⁵ sporulated oocysts of the C. ohioensis-like group and hypnozoites present in the viscera of mice from the INFECTED group, a pregnant bitch (in her final trimester) from Rio de Janeiro was selected and maintained at the UFRRJ experimental station. The bitch received daily oral treatment with trimethoprim (90 mg) and sulfadiazine (410 mg) (Triglobe®, Astra Química e Farmacêutica Ltda., Tamboré, São Paulo, Brazil) for 10 days and served as a puppy donor. Six puppies of both sexes born to this bitch were dewormed at 25 days of age (Basken® Plus, König do Brasil Ltda; single dose of 120 mg/kg body weight), weaned at 35 days, and housed in individual cages. Throughout the experiment, the animals received commercial puppy feed and water ad libitum. Two puppies were inoculated with 0.5 mL of a concentrated suspension containing 10⁵ sporulated oocysts. Another two puppies were fed organs from INFECTED group mice (euthanized at 60 DAI). The remaining two puppies served as controls: one received 0.5 mL of PBS orally, while the other was fed homogenized organs from CONTROL and PAIR-FED group mice.
2.9. Recovery of Oocysts from the Biological Assay
Oocysts recovered from the feces of two dogs inoculated with sporulated oocysts and two dogs fed viscera from mice in the INFECTED group were quantified daily using a modified Sheather's sucrose flotation technique, with slight adaptations as follows: the total fecal matter excreted daily by the dogs was collected, weighed, and homogenized. A 1-gram aliquot was placed in a 125 mL Erlenmeyer flask containing 100 mL of distilled water. The material was mixed to obtain a homogeneous suspension, from which 10 mL was withdrawn and transferred to a 15 mL Falcon centrifuge tube. The sample was subsequently centrifuged at 200 × g for five minutes. The supernatant was discarded, and the sediment was resuspended in saturated sugar solution before being centrifuged again under the same conditions (200 × g for 5 minutes). A disposable Pasteur pipette was used to add saturated sugar solution until a meniscus formed. A coverslip was placed on top for five minutes and then carefully transferred to a standard glass slide. The total number of oocysts present between the slide and coverslip was counted and multiplied by 10 to obtain the number of oocysts per gram of feces (OoPG). The total daily oocyst output was calculated by multiplying the OoPG value by the total weight (in grams) of feces collected each day. The remaining fecal material containing oocysts was further processed for concentration, sporulation, and morphological/morphometric analysis of the oocysts, following the same protocol used for oocysts collected from naturally infected dogs and initial donor animals in this study.
2.10. Statistical Analysis
The daily mean OoPG excreted and total daily output, along with the mean measurements and morphometric indices of oocysts and sporocysts from both naturally infected dogs and those inoculated with hypnozoites or oocysts, were subjected to descriptive analysis, and their means were compared using Student's t test. The mean values of live weight, weight gain, carcass weight, carcass yield, total visceral weight, percentage weight of each organ, and mean percentage organ weight relative to carcass weight from mice in the INFECTED, PAIR-FED and CONTROL groups were compared by Tukey's test. Additionally, the mean values of MD, md and MI of oocysts and sporocysts excreted by naturally infected dogs, dogs experimentally infected with sporulated oocysts, and dogs fed viscera (hypnozoites) from mice infected with sporulated oocysts were calculated and compared by Tukey's test. The number and percentage of hypnozoites in the viscera of the mice that were fed sporulated oocysts were also calculated. For statistical analysis of the data, GraphPad InStat software was used (https://graphpad-instat.software.informer.com/).
3. Results
A total of 26.73 × 10⁹ oocysts were recovered from the six naturally infected dogs over 15 days of shedding. None of the puppies presented clinical manifestations, and the texture of the collected feces was consistent in terms of their characteristic color and odor. The daily shedding behavior of the six puppies during the collection period can be observed in Figure 1a.
The sporulated C. ohioensis-like oocysts had a subspherical to spherical shape, with an oocyst wall consisting of two membranes averaging 0.29 µm in thickness and lacking a micropyle. The sporocysts were ellipsoidal in shape, with a smooth and thin wall enclosing four sporozoites inside, and no Stieda, Substieda, or Parastieda bodies were observed, nor were polar granules or oocyst residues. Sporocyst residues appeared as fine granular material that formed small clumps that aggregated into larger, denser granules (Figure 2). The mean dimensions and standard deviations of the MD, md and MI, as well as the highest observed values for oocysts and sporocysts, are shown in Table 1.
The average live weights (ALWs) of the mice in the INFECTED group were consistently lower than those in the CONTROL group, whereas compared with those in the PAIR-FED group, their average live weights followed a similar trend throughout the experiment (Figure 3a). When these parameters were assessed on the days when five mice from each group were euthanized, this observation was less evident. The ALW values of the statistically compared groups are shown in Table 2. The mean weight of the CONTROL group at 15 DAI was significantly greater (p≤0.05) than that of the INFECTED and PAIR-FED groups, whereas at the same time point, the INFECTED and PAIR-FED groups did not significantly differ (p≥0.05). The ALW of the CONTROL group was greater than that of the PAIR-FED group at 35 DAI, with statistically significant differences (p≤0.05) at 15, 16, 17, 18, 26, 27, and 28 DAI. The ALW of the PAIR-FED group did not differ significantly (p≥0.05) from that of the INFECTED group (Table 2).
The average weight gain (AWG) of the mice in the CONTROL group was lower than that of the INFECTED group until the 9th DAI, after which it increased and remained high until the end of the experiment, except at the 16th DAI (Table 3 and Figure 3c). There was no significant difference (p≥0.05) throughout the experiment, except at the 1st DAI, when the AWG of the CONTROL group was significantly lower (p≤0.05) than that of the INFECTED group (Table 3). As observed for ALW, when the AWG parameters were assessed on the days when five mice from each group were euthanized, this trend was less evident (Figure 3d). The PAIR-FED group had AWG values similar to those of the INFECTED group until the 5th DAI, after which its performance became inferior, with statistically significant differences (p≤0.05) on the 8th, 9th, 15th, 16th, 17th, and 27th DAI (Figures 3c and d). Similarly, the performance of the CONTROL group was inferior to that of the PAIR-FED group, with statistically significant differences (p≤0.05) on the 15th, 16th, 17th, 19th, and 27th DAI (Table 3).
Compared with that of the PAIR-FED group, the average carcass weight (ACW) of the mice in the INFECTED group was significantly lower (p≤0.05) on the 21st DAI, whereas on the 28th DAI, the ACW of the PAIR-FED group was significantly lower (p≤0.05) than that of the CONTROL group. At both time points (21st and 28th DAI), the ACW of the INFECTED group was lower than that of the CONTROL group, although the difference was not statistically significant (p≥0.05) (Table 4).
Considering the live weight of each individual, the average percentage carcass weight (ACW%) was established. In this case, the ACW% of the CONTROL group was consistently greater than that of the INFECTED group, except on the 14th and 28th DAI. However, these differences were not statistically significant (p≥0.05). On the 21st DAI, the ACW% of the INFECTED group [n=5 (X̄=59.73±2.04) 95% CI] was significantly lower (p≤0.05) than that of the PAIR-FED group [n=5 (X̄=63.39±1.76) 95% CI]. On the same day, the ACW% of the CONTROL group [n=5 (X̄=61.13±1.93) 95% CI] was greater than that of the INFECTED group and less than that of the PAIR-FED group, but the difference was not statistically significant (p≥0.05) (Figure 3e).
To determine whether organ (viscera) weight influences the carcass yield of infected animals, the live weight of each animal was subtracted from its carcass weight (LW-CW). As shown in Figure 3e and comparing it with Figure 3f, an inverse relationship was observed between the average percentage carcass yield (CY%) and viscera weight (LW-CW) in the INFECTED group. This pattern was not observed in the PAIR-FED and CONTROL groups.
No differences were found among animals in the INFECTED, PAIR-FED, and CONTROL groups (p≥0.05) regarding the average percentage weights of the stomach (SW%) and lungs (LW%) at any of the DAIs when the mice were euthanized throughout the experiment. With respect to the average heart weight (HW%), animals in the CONTROL group [n=5 (X̄=0.85±0.11) 95% CI] presented significantly greater HW% (p≤0.05) than did those in the INFECTED [n=5 (X̄=0.65±0.11) 95% CI] and PAIR-FED [n=5 (X̄=0.65±0.09) 95% CI] groups, but only on the 9th DAI. On the same day, these latter two groups did not significantly differ (p≥0.05).
On the 21st DAI, the average percentage weight of the small intestine (AWSI%) relative to the carcass weight (Figure 4a) in the CONTROL group [n=5 (X̄=4.43±1.40) 95% CI] was significantly lower (p≤0.05) than that in the PAIR-FED group [n=4 (X̄=9.39±1.21) 95% CI]. During the same period, the AWSI% of the INFECTED group [n=5 (X̄=6.35±2.8) 95% CI] was greater than that of both the CONTROL and PAIR-FED groups; however, these differences were not statistically significant (p≥0.05). When the average percentage weight of the large intestine (AWLI%) relative to the carcass weight was analyzed (Figure 4b), a significant difference was observed on the 1st DAI. The AWLI% of the CONTROL group [n=5 (X̄=10.47±1.88) 95% CI] was significantly greater (p≤0.05) than that of the PAIR-FED group [n=5 (X̄=7.57±1.35) 95% CI] but was not significantly different (p≥0.05) from that of the INFECTED group [n=5 (X̄=9.11±1.28) 95% CI].
Owing to the small size of Peyer's patches in the mice from all three groups, the average percentage weight of Peyer's patches (PPW%) could not be determined on the 1st DAI (Figure 4c). A significantly greater weight (p≤0.05) of PPW% was observed in the INFECTED group [n=5 (X̄=0.36±0.08) 95% CI] than in the CONTROL group [n=5 (X̄=0.23±0.09) 95% CI] on the 5th DAI (Figure 4c). On the 3rd and 5th DAI, the PPW% of the PAIR-FED group was higher than that of the INFECTED group, although not significantly (p≥0.05), and subsequently became lower (also not significantly, p≥0.05) on the 9th, 14th, and 21st DAI. By the 35th DAI, the PPW% values of all three groups were practically identical (Figure 4c).
Changes in the average percentage weight of mesenteric lymph nodes (MLNW%) were more clearly observed from the 9th DAI onward. A significantly greater MLNW% (p≤0.05) was found between the PAIR-FED [n=5 (X̄=9.39±1.21) 95% CI] and CONTROL [n=5 (X̄=4.42±1.40) 95% CI] groups on the 21st DAI, with values returning to similar levels by the 28th DAI (Figure 4d). On the 28th DAI, mice in the PAIR-FED group presented significantly greater MLNW% (p≤0.05) values than those in the INFECTED group did [n=5 (X̄=3.75±0.35) 95% CI], whereas no significant difference was observed compared with the CONTROL group [n=5 (X̄=4.32±0.82) 95% CI].
The average percentage spleen weight (SW%) of the animals in the INFECTED group was greater throughout most of the experiment than that of the PAIR-FED and CONTROL groups. Specifically, the SW% of the INFECTED group [n=5 (X̄=1.33±0.29) 95% CI] was significantly greater (p≤0.05) than that of the CONTROL group [n=5 (X̄=0.94±0.09) 95% CI] but not significantly different (p≥0.05) from that of the PAIR-FED group [n=5 (X̄=1.23±0.21) 95% CI] at the end of the experiment on the 35th DAI (Figure 4e).
The average percentage liver weight (LW%) of the mice in the INFECTED group increased from the 3rd DAI onward. A significantly greater LW% (p≤0.05) was observed in the INFECTED group [n=5 (X̄=12.16±2.10) 95% CI] than in the PAIR-FED group [n=5 (X̄=9.26±0.77) 95% CI] on the 9th DAI, whereas no significant difference (p≥0.05) was found versus the CONTROL group [n=5 (X̄=9.60±1.40) 95% CI] on that day. Compared with those of the other groups, the LW% of the INFECTED group subsequently decreased but remained nonsignificant (p≥0.05), ultimately reaching similar values among all three groups by the 35th DAI (Figure 4f).
In all the mice from the CONTROL and PAIR-FED groups, no C. ohioensis-like hypnozoites were observed in the organs after peptic digestion, nor were the hypnozoites recovered from the stomach, heart, lungs, or liver of the INFECTED group. From this group, a total of 2.4 × 10³ hypnozoites were recovered exclusively from the small and large intestines on the 1st DAI, from Peyer's patches between the 1st and 9th DAI, and from mesenteric lymph nodes and the spleen across all 35 evaluated DAIs (Table 5). However, by the 35th DAI, a significant reduction in the absolute number of isolates was observed in mesenteric lymph nodes and the spleen, which was accompanied by a relative decrease in isolation from mesenteric lymph nodes (4.42%) and a relative increase in isolation from the spleen (95.58%). The number of hypnozoites and their percentage relative to the total number of hypnozoites recovered daily from necropsies of various viscera are detailed in Table 5.
The viability of sporocysts in C. ohioensis oocysts inoculated into two puppies (designated the SPOROZOITE group) was confirmed when these animals began shedding a total of 9 × 10⁵ oocysts over 24 days starting from the 5th DAI (Figure 1b). Similarly, the viability of hypnozoites present in monozoic tissue cysts from the viscera of mice infected with 1 × 10⁵ sporulated oocysts was demonstrated when the other two puppies (designated the HYPNOZOITE group) shed a total of 16.2 × 10⁵ oocysts over 27 days starting from the 4th DAI (Figure 1b). Neither of the two puppies inoculated orally with the same volume of inoculum (0.5 mL of PBS) containing organs from the CONTROL or PAIR-FED group mice shed oocysts in their feces. All six puppies maintained a healthy appearance without signs of infection or notable gastrointestinal symptoms. The mean dimensions and standard deviations of MD, md and MI, along with the maximum and minimum measurements of oocysts and sporocysts recovered from the HYPNOZOITE and SPOROZOITE groups, are shown in Table 1. The MD measurements of oocysts from the HYPNOZOITE group were significantly greater (P ≤ 0.05) than those from the SPOROZOITE group, whereas the md and MI measurements were not significantly different (P ≥ 0.05) between the groups. The daily oocyst shedding patterns were similar between the HYPNOZOITE and SPOROZOITE groups until the 21st DAI, when the HYPNOZOITE group presented a renewed increase in oocyst excretion (Figure 1b).
4. Discussion
Coccidiosis is a common condition in dogs, and the most frequent clinical aspects are depression, weakness, loss of appetite, diarrhea, and dehydration [12,48,49,50,51,52,53,54]. Clinical coccidiosis was confirmed in a 10-week-old dog, which exhibited weight loss and yellowish and diarrheic feces, with the condition progressing to death despite therapeutic assistance with sulfaguanidine. Severe enteritis is associated with the asexual and sexual stages of organisms resembling C. ohioensis-like organisms [55]. There are few reports of C. canis causing coccidiosis in dogs; however, Mitchell et al. [56], upon experimentally infecting dogs with 1 × 10⁵ oocysts, reported weight loss, diarrhea, vomiting, and lethargy. Since other enteric etiological agents are responsible for similar clinical signs, a positive response to anticoccidial therapy has helped determine possible diagnoses [53]. However, a favorable response to anticoccidial therapy as support for the diagnosis of clinical coccidiosis is considered misleading by Kirkpatrick and Dubey [57], as most anticoccidial drugs also possess antibacterial properties. He et al. [58] did not observe any signs or symptoms of coccidiosis in 20 dogs that had sporulated oocysts of C. ohioensis-like, where at least one of these dogs was molecularly confirmed to have C. ohioensis. More recently, Barrera et al. [59], upon examining 524 fecal samples from dogs in a kennel over one year, diagnosed 46 (8.8%) animals as positive for Cystoisospora spp. Of these, the positivity rate among mothers was 10.6% (32/300), and that among litters was 6.2% (14/224). The Cystoisospora species identified were C. canis in 91.3% (42/46) of the samples and C. ohioensis-like species in 8.7% (4/46). Throughout the research period, none of the animals exhibited signs of coccidiosis. Similarly, in the present study, none of the dogs—whether naturally infected, infected with 1 × 10⁵ sporulated oocysts, or fed viscera containing hypnozoites from mice previously infected with 1 × 10⁵ sporulated oocysts—exhibited any signs or symptoms related to coccidiosis. These results may be related to the distinction of endogenous developmental stages, with C. ohioensis, according to Dubey [60], being confined to the superficial intestinal epithelium, which differs from C. rivolta and C. burrowsi, whose endogenous stages occur predominantly in the lamina propria. Thus, we cannot infer that the naturally infected dogs and oocyst donors in this study were or had been infected by C. ohioensis but rather by the group of medium-sized coccidia referred to as the C. ohioensis-like complex.
Oocysts were shed in significant numbers by all six naturally infected puppies, as well as by the two puppies fed hypnozoites contained in the viscera of previously infected mice and by the two puppies orally infected with sporozoites contained in sporulated oocysts (Figure 1a and 1b). The number of oocysts shed daily by the dogs in the groups inoculated with hypnozoites and sporozoites was similar until the 21st DAI (days post infection), when the HYPNOZOITES group presented a new increase in the number of oocysts shed (Figure 1b). The animals in this group reached peak daily shedding on the 25th DAI and continued shedding oocysts until the 27th DAI, differing from the animals in the SPOROZOITE group, which peaked on the 5th DAI and shed oocysts until the 24th DAI (Figure 1b).
The prepatent period of C. ohioensis can range from 13 to 23 days [36,61], and the prepatent period observed by Rocha & Lopes [62] in dogs inoculated with sporulated oocysts was 3 to 7 days. In induced infections, the observation of gametocytes in the intestinal cells of dogs at 96 and 114 hours postinoculation with hypnozoites and sporozoites, respectively, suggests a biological distinction between the two parasitic forms [63]. Dubey and Lindsay [8], in their review of canine coccidiosis, reported that the asexual and sexual stages in cycles induced by sporozoites or hypnozoites are structurally similar in size and location, but the hypnozoite-driven cycle develops 24 hours faster. This was evident in our research results, where dogs inoculated with hypnozoites had a shorter prepatent period, shed a greater number of oocysts, and exhibited a longer patent period, reinforcing the hypothesis that the two parasitic stages are biologically distinct, as suggested by Dubey [63] and Dubey & Lindsay [8]. The oocyst shedding pattern in naturally infected dogs was similar to that in dogs fed viscera from previously infected mice (hypnozoites), suggesting that the six naturally infected dogs had ingested hypnozoites through contaminated food. In 1995, Dubey reported that dogs infected with 1 × 10⁵ oocysts of C. canis or fed tissues from mice previously infected with 1 × 10⁵ oocysts produced similar numbers of oocysts and had comparable prepatent periods [64]. However, in our experiment, dogs inoculated with 1 × 10⁵ sporulated oocysts had a longer prepatent period than those fed viscera from mice previously inoculated with 1 × 10⁵ sporulated oocysts (Figure 1b).
The subspherical appearance of the oocysts and ellipsoid shape of the sporocysts shed by the six naturally infected dogs, as well as the measurements of the major and minor diameters of the oocysts and sporocysts, were compatible with those observed by Rocha & Lopes [62] and Dubey [65] for C. rivolta and with those characterized by Lindsay et al. [7] as intermediate-sized oocysts from dogs with dimensions ranging from 17 to 23 µm, grouped as C. ohioensis-like (Figure 1a). In the present study, the measurements of MD, md and MI of oocysts and sporocysts from the six naturally infected dogs were smaller (Table 1) than those of oocysts and sporocysts shed by the two puppies inoculated with sporulated oocysts and the two puppies fed mouse viscera containing hypnozoites. These differences in measurements may be related to varying proportions of medium-sized oocysts being shed, corresponding to the species C. ohioensis, C. burrowsi and C. rivolta, which cannot be distinguished morphologically [7].
The lower ALW of the animals in the INFECTED group throughout the experiment, which became statistically significant from the 15th DAI compared with that of the CONTROL group (Table 2), and the AWG of the PAIR FED group, which was consistently lower than that of the INFECTED group (Table 3), support the hypothesis that the reduced performance of intermediate hosts in terms of daily live weight and weight gain is related to the development of infective forms for the subsequent establishment of extraintestinal hypnozoites, where an increase in visceral weight (Figure 3f) when parasitized masks the wasting effect of C. ohioensis-like species. These observations were also detected in mice inoculated with sporulated oocysts of C. rivolta (=C. ohioensis-like) by Brösike et al. [66] and with C. felis by Loss and Lopes [67]; Freire and Lopes [68] and Costa and Lopes [69]. When Costa and Lopes [69] inoculated rabbits with sporulated oocysts of C. felis, they reported that the animals lost weight from the first week after infection until the end of the experiment on the 29th DAI. The differences in ALW and AWG between the INFECTED and CONTROL groups are likely related to the anorexia that follows infection, as described by Loss and Lopes [67].
The consistently lower ALW of the animals in the INFECTED group throughout the experiment, which became statistically significant from the 15th DAI compared with that of the CONTROL group (Table 2), along with the AWG of the PAIR-FED group being consistently lower than that of the INFECTED group (Table 3), supports the hypothesis that the reduced performance of intermediate hosts in terms of daily body weight and weight gain is related to the development of infection stages, leading to the establishment of extraintestinal hypnozoites. In this scenario, the increased visceral weight (Figure 3f) when parasitized masks the cachectic effect of C. ohioensis-like infection. Similar observations were reported in mice inoculated with sporulated oocysts of C. rivolta (=C. ohioensis-like) by Brösike et al. [66] and with C. felis by Loss and Lopes [67], Freire and Lopes [68], and Costa and Lopes [69]. When Costa and Lopes [69] inoculated rabbits with sporulated oocysts of C. felis, they observed weight loss starting from the first week postinfection until the end of the experiment on the 29th DAI. The differences in ALW and AWG between the INFECTED and CONTROL groups are likely related to postinfection anorexia, as described by Loss and Lopes [67].
Studies with cattle [70,71]; buffalo, sheep, and swine [17,21,23]; rabbits [20]; and broiler chickens [22] have indicated that production animals can harbor varying numbers of Cystoisospora genus hypnozoites in their viscera. However, whether parasitic action in these organs influences weight gain in these animals or causes significant lesions remains unknown. Nevertheless, recurrent clinical disease is common in both immunocompetent and immunosuppressed humans infected with C. belli, which is believed to be due to reactivation of the hypnozoites present in tissues and their migration to the human intestinal tract [24,31]. Immunosuppressed patients frequently experience clinical disease relapse after discontinuation of C. belli treatment [69], although Frenkel et al. [71] failed to infect suckling calves and chicks with 1 × 10⁶ and 3 × 10⁴ sporulated oocysts of C. belli, respectively. Ultimately, the interference of this parasitism on weight development in humans and animals remains unknown, whereas economic production of animal protein requires increasingly greater financial investments to meet the demand for meat, viscera, or both. Our research suggests that animals may have impaired zootechnical performance, as occurred with mice in the INFECTED group during the establishment of hypnozoites in target organs or due to anorexia induced during this process. This hypothesis is reinforced by the fact that mice in the PAIR-FED group of our experiment showed lower ALW and AWG performance than the INFECTED group, resulting from feed control that increased stress levels and the lack of nutrients necessary for the development of healthy animals.
Biological assays in cats have indicated that infectious stages of C. felis and C. rivolta can develop in gerbils (Meriones unguiculatus), guinea pigs (Cavia porcellus), rabbits (Oryctolagus cuniculus), and chickens (Gallus gallus domesticus) fed sporulated oocysts [74]. In addition to the results of this study, it can be inferred that infection of mice with C. ohioensis-like and potentially other intermediate hosts negatively affects carcass weight. The increased organ weight in the parasitized animals (Figure 3f) contributed to a lower carcass yield (Table 3 and Table 4), since parasitism also reduced the ALW (Figures 3a, b and Table 2) and AWG (Figures 3c, d and Table 3) of the infected mice.
Freire and Lopes [68] reported no differences in the mean weight percentages of the heart, lungs, kidneys, and brain in mice inoculated with sporulated oocysts of C. felis. Dubey [11] previously failed to demonstrate hypnozoites or extraintestinal stages of C. ohioensis in histopathological sections of the heart, lung, and brain, which led Freire and Lopes [66] to suggest that these organs, including the kidneys, are rarely parasitized by Cystoisospora species. Brösike [75] studied the frequency of C. rivolta (=C. ohioensis-like) hypnozoites in Swiss albino mice, and although this parasite is widely distributed throughout the host organism, it does not develop in the brain. In our research, no differences in ASW% (average stomach weight percentage) or AWH% (average weight heart percentage) were detected between the groups throughout the experiment. In fact, the AWSI% and AWLI% of the mice in the INFECTED group were greater, although not significantly so, over 35 days, likely because of the presence of transient parasites in these organs, as shown in Table 5. Considering the viscera where Cystoisospora spp. has been previously reported [66,67,68,69,75,76] and the ASW%, AWH%, AWPP% (percentage of the average weight of the Peyer's patch), AWSI%, and AWLI% observed in this experiment, these organs are not frequently parasitized by C. ohioensis-like organisms, as previously reported with other Cystoisospora species by these same researchers. All the aforementioned authors reported alterations in the AWPP%, AWML% (percentage of the average weight of the mesenteric lymph node), AWS% (percentage of the average weight of the spleen), and AWL% (percentage of the average weight of the liver) of animals inoculated with Cystoisospora spp., which is consistent with our observations for C. ohioensis-like organisms. Dubey [76] demonstrated that sporozoites released after the excystation of C. rivolta sporulated oocysts most frequently invade mouse mesenteric lymph nodes, which remain infected for at least 23 months, potentially explaining the increased AWML% returning to proportionality in our study (Figure 4d), as also verified by Freire and Lopes [68] between the 4th and 18th DAI, with this finding being attributed to the distribution of parasite hypnozoite forms to host organs. These researchers reported increased AWL% in infected mice from the 3rd to 18th DAI, whereas in our experiment, the AWL% increase occurred from the 3rd to 9th DAI (Figure 4f), followed by a decrease, with no differences among the INFECTED, PAIR-FED and CONTROL groups until the 35th DAI. These cyclic alterations, also observed in AWPP% and AWML%, may be due to spontaneous flow of parasite hypnozoite forms during their biological cycle development in the paratenic host, which is consistent with Dubey [76], who demonstrated that hypnozoites increased in size during the first 31 days and could be found in the spleen, liver, and skeletal muscle of mice infected with C. rivolta (=C. ohioensis-like). It has also been shown that the hypnozoites of C. felis, C. rivolta and C. ohioensis increase in size over time but do not divide [10,18]. Our results revealed that during parasite passage through organs, their weight percentages increased (Figures 4a,f), possibly due to elimination of some forms while others became encysted. Since parasite multiplication does not occur in the intermediate host, the organ weight percentages returned to normal during the chronic infection process, as observed in this experiment, with AWPP% (Figure 4c), AWML% (Figure 4d) and AWL% (Figure 4f) values. However, the spleen did not follow this pattern, with AWS% (Figure 4e) increasing throughout the entire process, including the chronic phase, suggesting that this organ best harbors and maintains C. ohioensis parasitic forms for the longest duration.
Parasite dissemination in rodent organs occurs similarly to that in other coccidia through phagocytes [10,77,78,79]. Freire and Lopes [68] suggested the existence of both hematogenous and lymphatic routes for extraintestinal dissemination of Cystoisospora species in intermediate hosts. Liver and spleen enlargement in the initial days postinoculation results from rapid parasite spread via the bloodstream, whereas slower dissemination likely occurs through the lymphatic system, sequentially increasing lymphoid organ size. According to Lindsay et al. [80], further investigations are needed to elucidate the complex life cycle and epidemiology of Cystoisospora spp., as developmental cycles were originally described in immunocompetent hosts, and the impact of immunosuppression on definitive host development remains unknown. Our research revealed temporary production losses in immunocompetent intermediate hosts, which are likely exacerbated in immunocompromised animals—a common scenario in meat and dairy production systems due to the confinement stress required for increased productivity.
Freire and Lopes [68] observed C. felis hypnozoites in the kidneys, lungs, and heart, attributing the presence of the parasite in these organs to accidental distribution rather than a normal parasitic access route. The absence of hypnozoites in the stomach, heart, and lungs in our study suggests that these organs are not preferential for C. ohioensis-like organisms in mice, a hypothesis supported by visceral weight evaluation findings. No hypnozoites were observed in the livers of INFECTED group mice in our research. Considering that the liver is a known site for hypnozoite observation in intermediate hosts infected with C. rivolta [66,75] and C. felis [68,69], we cannot conclusively state that the liver was free from C. ohioensis-like parasitism in our study, as a significant cellular mass remained after the peptic digestion of liver tissue, requiring substantial sample dilution to improve hypnozoite visualization. Furthermore, hypnozoite recovery in the spleen throughout the experiment, particularly at 1st DAI (Table 5), supports the hematogenous dissemination hypothesis [68] previously discussed in visceral weight analysis. Therefore, a more comprehensive evaluation of the presence of C. ohioensis-like hypnozoites is needed to definitively exclude liver parasitism in experimental animals.
In the spleen, a progressive increase in both the absolute and relative numbers of recovered hypnozoites was observed, indicating that this organ is the primary target of C. ohioensis-like organisms in mice. Brösigke [75] noted a strong predilection of C. rivolta for mesenteric lymph nodes at the 2nd, 4th, 84th, and 168th DAI, whereas the liver showed a preference only at the 3rd and 7th DAI. Freire and Lopes [68] reported that C. felis prefers the spleen and liver, with considerable hypnozoite recovery in Peyer's patches and mesenteric lymph nodes, suggesting that C. rivolta is more lymphotropic than C. felis is Velásquez et al. [27] reported the clinical manifestations, humoral responses, and histological findings of different stages during C. belli infection in an AIDS patient with liver disease. These findings led researchers to hypothesize that sporozoites and/or monozoic tissue cysts invade duodenal mucosal epithelial cells, penetrating the basement membrane and lamina propria. The authors further suggest that monozoic tissue cysts likely migrate to the liver via portal blood circulation, although lymphatic vessel migration cannot be ruled out [27]. The dynamics of C. ohioensis-like organisms in our study indicate a faster hematogenous route to the spleen between the 1st and 3rd DAI, as evidenced by increased hypnozoite recovery at these timepoints (Table 5), and a slower lymphatic migration route, as shown by increasing hypnozoite counts from the 9th DAI onward (Table 5). These results suggest that the C. ohioensis-like distribution is more lymphotropic, which aligns with Freire and Lopes' [66] observations for C. rivolta. Although the spleen was the target organ for C. ohioensis-like organisms in our study, decreased parasite recovery at the 35th DAI (Table 5) may reflect either parasite elimination during its biological cycle or survival bias of acid‒pepsin digestion-resistant stages [78]. These findings support the immunological regulation hypothesis, where C. rivolta-like species are more easily controlled than C. felis are [70,81].
5. Conclusions
Studies analyzing the deleterious effects of Cystoisospora spp. are scarce, and no research has addressed the pathogenic, biological, and nutrient-depleting aspects related to C. ohioensis-like organisms in intermediate hosts. However, studies on other coccidia, such as those of the genera Toxoplasma, Eimeria, Sarcocystis, and, more recently, on Neospora and Cystoisospora belli in humans, have been conducted, highlighting the importance of these coccidia in animal production and One Health. The role of coccidia as opportunistic agents is well known, and this aspect has gained significance with the emergence of immunosuppressive diseases such as AIDS and the stresses of modern life. The medications used in chemotherapy and organ transplant rejection control, along with the indiscriminate use of corticosteroids in veterinary medicine, render both humans and animals more vulnerable to pathogens once they are considered nonpathogenic. In livestock, the stress of confinement systems aimed at achieving high productivity in certain farming operations is a key factor in opportunistic infections. In the intermediate host, C. ohioensis-like species invade extraintestinal organs, being transported via the lymphatic system and—although suggested but not confirmed in this study—potentially through the bloodstream as well. The parasitized organs increase in size, masking the body weight and weight gain of the animals and ultimately impairing carcass yield. These findings demonstrate the nutrient-depleting and nutrient-wasting effects of C. ohioensis-like parasitism in intermediate hosts. Most transport hosts are optional for parasites and are considered "carriers" without true parasitism—meaning a harmonious parasite‒host relationship. However, on the basis of the results of this study, nondefinitive hosts of C. ohioensis-like species should be classified as intermediate hosts rather than transport or paratenic hosts, as the latter do not support parasite development or exhibit any host effects.
Author Contributions
“Conceptualization, F.C.R.O., S.S.M.G. and C.W.G.L.; methodology, F.C.R.O., S.S.M.G. and C.W.G.L.; software, F.C.R.O.; validation, F.C.R.O., S.S.M.G. and C.W.G.L.; formal analysis, S.S.M.G. and C.W.G.L.; investigation, F.C.R.O.; resources, C.W.G.L.; data curation, F.C.R.O., S.S.M.G.; writing—original draft preparation, F.C.R.O.; writing—review and editing, S.S.M.G. and C.W.G.L.; visualization, S.S.M.G.; supervision, C.W.G.L.; project administration, F.C.R.O. and C.W.G.L.. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Frenkel, J.K. Besnoitia wallace in cats and rodents: with a reclassification of other cyst-forming isosporoid coccidia. J. Parasitol. 1977, 63, 611–628. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.D. The Sarcocystidae: Sarcocystis, Frenkelia, Toxoplasma, Hammondia, and Cystoisospora. J. Parasitol. 1981, 28, 262–266. [Google Scholar] [CrossRef]
- Carreno, R.A.; Schnitzler, B.E.; Jeffries, A.C.; Tenter, A.M.; Johnson, A.M.; Barta, J.R. Phylogenetic analysis of coccidia based on 18S rDNA sequence comparison indicates that Isospora is most closely related to Toxoplasma and Neospora. J. Eukaryot. Microbiol. 1998, 45, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Franzen, C.; Müller, A.; Bialek, R.; Diehl, V.; Salzberger, B.; Fätkenheuer, G. Taxonomic position of the human intestinal protozoan parasite Isospora belli as based on ribosomal RNA sequences. Parasitol. Res. 2000, 86, 669–676. [Google Scholar] [CrossRef]
- Barta, J.R.; Martin, D.S.; Carreno, R.A.; Siddall, M.E.; Profousj-Uchelka, H.; Hozza, H.; Powles, M.A.; Sundermann, C. Molecular phylogeny of the other tissue coccidia: Lankesterella and Caryospora. J. Parasitol. 2001, 87, 121–127. [Google Scholar] [CrossRef]
- Barta, J.R.; Schrenzel, M.D.; Carreno, R.; Rideout, B.A. The genus Atoxoplasma (Garnham 1950) as a junior objective synonym of the genus Isospora (Schneider 1881) species infecting birds and resurrection of Cystoisospora (Frenkel 1977) as the correct genus for Isospora species infecting mammals. J. Parasitol. 2005, 91, 726–727. [Google Scholar] [CrossRef]
- Lindsay, D.S.; Dubey, J.P.; Blackburn, B.L. Biology of Isospora spp. from humans, nonhuman primates and domestic animals. Clin. Microbiol. Rev. 1997a, 10, 19–34. [CrossRef]
- Dubey, J.P; Lindsay, D.S. Coccidiosis in dogs—100 years of progress. Vet. Parasitol. 2019, 266, 34–55. [Google Scholar] [CrossRef]
- Dubey, J.P. Toxoplasma, Neospora, Sarcocystis and other tissue cyst forming coccidia of humans and animals. In: Parasitic Protozoa, Kreier, J.P. (ed). Academic Press, New York, 1993, pp. 5–57.
- Dubey, J.P.; Mehlhorn, H. Extraintestinal stages of Isospora ohioensis from dogs in mice. J. Parasitol. 1978, 64, 689–685. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P. Life cycle of Isospora ohioensis in dogs. Parasitol. 1978, 77, 1–11. [Google Scholar] [CrossRef]
- Buehl, I.E.; Prosl, H.; Mundt, H.C.; Tichy, A.G.; Joachim, A. Canine isosporosis -epidemiology of field and experimental infections. J. Vet. Med. 2006, 53, 482–487. [Google Scholar] [CrossRef]
- Lee, S.; Kim, J.; Cheon, D.S.; Moon, E.A.; Seo, D.J.; Jung, S.; Shin, H.; Choi, C. Identification of Cystoisospora ohioensis in a Diarrheal Dog in Korea. Korean J. Parasitol. 2018, 56, 371–374. [Google Scholar] [CrossRef] [PubMed]
- Levine, N.D. Veterinary Protozoology; Iowa State University Press, Ames, 1985, 414 pp.
- Hilali, M.; Nassar, A.M.; El-Ghaysh, A. Camel (Camelus dromedarius) and sheep (Ovis aries) meat as a source of dog infection with some coccidian parasites. Vet. Parasitol. 1992, 43, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Hilali, M.; Fatani, A.; Al-Atiya, S. Isolation of tissue cysts of Toxoplasma, Isospora, Hammondia and Sarcocystis from camel (Camelus dromedarius) meat in Saudi Arabia. Vet. Parasitol. [CrossRef]
- Zayed, A.A.; El-Ghaysh, A. Pig, donkey and buffalo meat as a source of some coccidian parasites infecting dogs. Vet. Parasitol. 1998, 78, 161–168. [Google Scholar] [CrossRef]
- Frenkel, J.K.; Dubey, J.P. Rodents as vectors for feline coccidia Isospora felis and Isospora rivolta. J. Infect. Dis. 1972, 125, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Frenkel, J.K.; Smith, D.D. Determination of the genera of cystforming coccidia. Parasitol. Res. 2003, 91, 384–389. [Google Scholar] [CrossRef]
- Costa, P.S.; Lopes, C.W.G. Avaliação do parasitismo por Cystoisospora felis (Wenyon, 1923) Frenkel, 1977 (Apicomplexa: Cystoisosporinae) em coelhos tipo carne. Braz. J. Vet. Parasitol. 1998, 7, 15–19. [Google Scholar] [CrossRef]
- Carvalho Filho, P.R.; Melo, P.S.; Massad, F.V.; Lopes, C.W.G. Determinação da infecção de suínos por Cystoisospora felis (Wenyon, 1923) Frenkel, 1977 (Apicomplexa: Cystoisosporinae) através de prova biológica em felinos livres de coccídios. Rev. Bras. Parasitol. Vet. 2003, 12, 37–42. [Google Scholar]
- Massad, F.V.; Oliveira, F.C.R.; Albuquerque, G.R.; Lopes, C.W.G. Hipnozoítas de Cystoisospora ohioensis (Dubey, 1975) Frenkel, 1977 (Apicomplexa: Cystoisosporinae) em frangos. Rev. Bras. Ciênc. Vet. 2003, 10, 57–58. [Google Scholar] [CrossRef]
- Melo, P.S.; Carvalho Filho, P.R.; Lopes, C.W.G.; Flausino, W.; Oliveira, F.C.R. Hypnozoites of Cystoisospora felis (Wenyon, 1923) Frenkel, 1977 (Apicomplexa: Cystoisosporinae) isolated from piglets experimentally infected. Rev. Bras. Parasitol. Vet. 2003, 12, 57–59. [Google Scholar]
- Lindsay, D.S.; Dubey, J.P.; Toivio-Kinnuncan, M.A. , Michiels, J.F.; Blagburn, B.L. Examination of extraintestinal tissue cysts of Isospora belli. J. Parasitol. 83, 620–625. [CrossRef] [PubMed]
- Benator, D.A.; French, A.L.; Beaudet, L.M.; Levy, C.S.; Orenstein, J.M. Isospora belli infection associated with acalculous cholecystitis in a patient with AIDS. Ann. Intern. Med. 1994, 121, 663–664. [Google Scholar] [CrossRef] [PubMed]
- Walther, Z.; Topazian, M.D. Isospora cholangiopathy: case study with histologic characterization and molecular confrmation. Hum. Pathol. 2009, 40, 1342–1346. [Google Scholar] [CrossRef]
- Velásquez, J.N.; Etchart, C.B.; Astudillo, O.G.; Chertcoff, A.V.; Pantano, M.L.; Carnevale, S. Cystoisospora belli, liver disease and hypothesis on the life cycle. Parasitol. Res. 2022, 121, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Restrepo, C.; Macher, A.M; Radany, E.H. Disseminated extraintestinal isosporiasis in a patient with acquired immune deficiency syndrome. Am. J. Clin. Pathol. 1987, 87, 536–542. [Google Scholar] [CrossRef] [PubMed]
- Michiels, J.F.; Hofman, P.; Bernard, E.; Saint Paul, M.C.; Boissy, C.; Mondain, V.; Lefichoux, Y.; Loubiere, R. Intestinal and extraintestinal Isospora belli infection in an AIDS patient. A second case report. Pathol. Res. Pract. 1994, 190, 1089–1094. [Google Scholar] [CrossRef]
- Comin, C.E.; Santucci, M. Submicroscopic profile of Isospora belli enteritis in a patient with acquired immune deficiency syndrome. Ultrastruct. Pathol. 1994, 18, 473–482. [Google Scholar] [CrossRef]
- Velásquez, J.N.; Carnevale, S.; Mariano, M.; Kuo, L.H.; Caballero, A.; Chertcoff, A.; Ibáñez, C.; Bozzini, J.P. Isosporosis and unizoite tissue cysts in patients with acquired immunodefciency syndrome. Hum. Pathol. 2001, 32, 500–505. [Google Scholar] [CrossRef]
- Frenkel, J.K.; Silva, M.B.; Saldanha, J.C. ; De Silva-Vergara. M.L.; Correia, D.; Barata, C.H.; Silva, E.L.; Ramirez, L.E.; Prata, A. Presença extra-intestinal de cistos unizóicos de Isospora belli em paciente com SIDA: relato de caso. Rev. Soc. Bras. Med. Trop. 2003a, 36, 409–412. [Google Scholar] [CrossRef]
- Loss, Z.G.; Lopes, C.W.G. . Alguns aspectos clínicos na infecção experimental por Cystoisospora felis (Wenyon, 1926) Frenkel, 1976 (Apicomplexa: Cystoisosporinae) em gatos. Arq. UFRRJ 1992a, 15, 79–84. [Google Scholar]
- Loss, Z.G.; Lopes, C.W.G. Aspecto patológicos da infecção experimental por Cystoisospora felis (Wenyon, 1926) Frenkel, 1976 (Apicomplexa: Cystoisosporinae) em gatos. Arq. UFRRJ 1992b, 15, 113–119. [Google Scholar]
- Nemeséri, L. Beiträge Zur Ätiologie der Coccidiose der hunde. 1. Isospora canis sp. n. Acta Vet. Hung. 1959, 10, 95–99. [Google Scholar]
- Dubey, J.P. Isospora ohioensis sp. n. proposed for I. rivolta of the dog. J. Parasitol. 61, 462–465. [CrossRef] [PubMed]
- Trayser, C.V.; Todd, K.S. Life cycle of Isospora burrowsi n sp (Protozoa: Eimeriidae) from the dog Canis familiaris. Am. J. Vet. Res. 1978, 39, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Rommel, M.; Zielasko, B. Untersuchungen uber den Lebenszyklus von Isospora burrowsi (Trayser and Todd, 1978) aus dem Hund. Berl. tierarztl. Wschr. 1981, 94, 87–90. [Google Scholar] [PubMed]
- Dubey, J.P.; Mahrt, J.L. Isospora neorivolta sp. n. from the domestic dog. J. Parasitol. 1978, 64, 1067–1073. [Google Scholar] [CrossRef] [PubMed]
- Tadros, W.; Laarman, J.J. Sarcocystis and related coccidium parasites: a brief general review, together with a discussion on some biological aspects of their life cycles and a new proposal for their classification. Act. Leid. 1976, 44, 1–107. [Google Scholar] [PubMed]
- Dubey, J.P.; Capenter, J.L.; Speer, C.A.; Topper, M.J.; Uggla, A. Newly recognized fatal protozoan disease of dogs. J. Am. Vet. Med. Assoc. 1988, 192, 1269–1285. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, F.C.R. Avaliação da infecção experimental em camundongos albinos com oocistos esporulados de Cystoisospora ohioensis (Dubey, 1975) Frenkel, 1977 (Apicomplexa: Cystoisosporinae) e sua transmissão ao cão doméstico. Tese (Medicina Veterinária, Parasitologia Veterinária), Instituto de Veterinária, Universidade Federal Rural do Rio de Janeiro, Seropédica, 172 pp., 2001.
- Oliveira, F.C.R.; Albuquerque, G.R.; Munhoz, A.D.; Lopes, C.W.G. . Oocysts of Cystoisospora ohioensis (Dubey, 1975) Frenkel, 1977 (Apicomplexa: Cystoisosporinae) in naturally infected puppies from a litter. Rev. Univ. Rural 2000, 22, 107–111. [Google Scholar]
- Brazil. Law No. 11,794, of October 8, 2008. It regulates subsection VII of §1 of Article 225 of the Federal Constitution, establishing procedures for the scientific use of animals; repeals Law No. 6,638, of May 8, 1979; and makes other provisions.
- CFMV. Resolution No. 714, of June 20, 2002. It establishes procedures and methods for animal euthanasia and provides other measures. Available at: http://ts.cfmv.gov.br/manual/arquivos/resolucao/714.pdf.
- Dubey, J.P. Refinement of pepsin digestion method for isolation of Toxoplasma gondii from infected tissues. Vet. Parasitol. 1997, 1, 75–77. [Google Scholar] [CrossRef]
- Oliveira, F.C.R.; Albuquerque, G.R.; Munhoz, A.D.; Lopes, C.W.G.; Massad, F.V. Hipnozoítas de Cystoisospora ohioensis (Dubey, 1975) Frenkel, 1977 (Apicomplexa: Cystoisosporinae) recuperados de órgãos de camundongos através da digestão péptica. Rev. Bras. Parasitol. 2001, 10, 29–35. [Google Scholar]
- Oduye, O.O.; Bobade, P.A. Studies on an outbreak of intestinal coccidiosis in the dog. J Small Anim Pract. 1979, 20, 181–184. [Google Scholar] [CrossRef] [PubMed]
- Correa, W.M.; Correa, C.N.M.; Langoni, H.; Volpato, O.A.; Tsunoda, K. Canine isosporosis. Canine Pract., 1983, 10: 44-46.
- Olson, M.E. Coccidiosis caused by Isospora ohioensis-like organisms in three dogs. Can. Vet. J. 1985, 26, 112–114. [Google Scholar] [PubMed]
- Penzhorn, B.L.; De Cramer, K.G.M.; Booth, L.M. Coccidial infection in German shepherd dog pups in a breeding unit. J. S. Afr. Vet. Assoc. 1992, 63, 27–29. [Google Scholar] [PubMed]
- Wu, D.H.; Jin, H.; Zhang, H.D.; Xu, H.K. A case of mixed infection of canine distemper and intestinal protozoa. Chin. J. Prev. Vet. Med. 1993, 19, 32–33. [Google Scholar] [CrossRef]
- Randhawa, S.S.; Juyal, P.D.; Kalra, I.S. Clinical Isosporiasis in a racer greyhound dog. Indian Vet. J. 1997, 14, 413–414. [Google Scholar]
- Altreuther, G.; Gasda, N.; Schroeder, I.; Joachim, A.; Settje, T.; Schimmel, A.; Hutchens, D.; Krieger, K.J. Efficacy of emodepside plus toltrazuril suspension (Procox R® oral suspension for dogs) against prepatent and patent infection with Isospora canis and Isospora ohioensis-complex in dogs. Parasitol. Res., 2011, 109 (Suppl 1): S9–S20. [CrossRef]
- Dubey, J.P.; Weisbrode, S.E.; Rogers, W.A. Canine coccidiosis attributed to an Isospora ohioensis-like organism: a case report. J. Am. Vet. Med. Assoc. [PubMed]
- Mitchell, S.M.; Zajac, A.M.; Charles, S.; Duncan, R.B.; Lindsay, D.S. Cystoisospora canis Nemeséri, 1959 (syn. Isospora canis), infections in dogs: clinical signs, pathogenesis, and reproducible clinical disease in beagle dogs fed oocysts. J. Parasitol. [CrossRef]
- Kirkpatrick, C.E.; Dubey, J.P. Enteric Coccidial Infections: Isospora, Sarcocystis, Cryptosporidium, Besnoitia, and Hammondia. Vet. Clin: Small Anim. Pract. [CrossRef]
- He, P.; Li, J.; Gong, P.; Huang, J.; Zhang, X. Cystoisospora spp. from dogs in China and phylogenetic analysis of its 18S and ITS1 gene. Vet. Parasitol. [CrossRef]
- Barrera, J.P.; Montoya, A.; Marino, V.; Sarquis, J.; Checa, R.; Miró, G. Cystoisospora spp. infection at a dog breeding facility in the Madrid region: Infection rate and clinical management based on toltrazuril metaphylaxis. Vet. Parasitol. 2024; 48. [Google Scholar] [CrossRef]
- Dubey, J.P. Re-evaluation of merogony of a Cystoisospora ohioensis-like coccidian and its distinction from gametogony in the intestine of a naturally infected dog. Parasitol. [CrossRef]
- Mahrt, J.L. Endogenous stages of the life cycle of Isospora rivolta in the dog. J. Parasitol., 1967, 14, 754–759. [Google Scholar] [CrossRef]
- Rocha, E.M. , Lopes C.W.G. (1971). Comportamento da Isospora canis, Isospora felis e Isospora rivolta em infecções experimentais em cães e gatos. Arq. UFRRJ 1971, 1, 81–96. [Google Scholar]
- Dubey, J.P. Toxoplasma, Hammondia, Besnoitia, Sarcocystis and other tissue cyst-forming coccida of man and animals. In: Protozoa, Kreier, J.P., Ed.; Academic Press: New York, 1977; Volume 3, pp. 102–219.
- Dubey, J.P. Experimental Isospora canis and Isospora felis infection in mice, cats, and dogs. J. Protozool. [CrossRef]
- Dubey, J.P. Toxoplasma, Hammondia, Besnoitia, Sarcocystis and other tissue cyst-forming coccidia of man and animals. In: Protozoa, Kreier, J.P., Ed.; Academic Press: New York, 1992; Volume 6, pp.120–128.
- Brösigke, S.; Heine, J.; Boch, J. Der Nachweis extraintestinallen entwicklingstadien (Dormozoiten) in extraintestinall mit Cystoisospora rivolta oozysten infizierten Mausen. Kleintierprax., 1982, 27, 25–34. [Google Scholar]
- Loss, Z.G.; Lopes, C.W.G. Efeito da infecção experimental por Cystoisospora felis (Apicomplexa: Cystoisosporinae) no ganho de peso de camundongos. Arq. UFRRJ 1992c, 15, 109–111. [Google Scholar]
- Freire, R.B.; Lopes, C.W.G. Distribuição de hipnozoítas de Cystoisospora felis (Weyon, 1923) Frenkel, 1977 (Apicomplexa: Sarcocystidae) em camundongos albinos experimentalmente infectados. Rev. Bras. Parasitol. Vet. 1996, 5, 23–28. [Google Scholar]
- Costa, P.S.; Lopes, C.W.G. Hipnozoítas de Cystoisospora felis (Apicomplexa: Cystoisosporinae). Rev. Bras. Ciênc. Vet. [CrossRef]
- Fayer, R. , Frenkel J.K. (1979). Comparative infectivity for calves of oocysts of feline coccidia: Besnotia, Hammondia, Cystoisospora, Sarcocystis and Toxoplasma. J. Protozool. [PubMed]
- Wolters, E.; Heydorn, A.O.; Laudahn, C. Cattle as intermediate host of Cystoisospora felis. [Das Rind als Zwischenwirt von Cystoisospora felis]. Berl. tierarztl. Wschr. 1980, 93, 207–210. [Google Scholar] [PubMed]
- Boyles, T.H.; Black, J.; Meintjes, G.; Mendelson, M. Failure to eradicate Isospora belli diarrhea despite immune reconstitution in adults with HIV—A case series. PLoS One. [CrossRef]
- Frenkel, J.K.; Silva, M.B.; Saldanha, J.; De Silva, M.L.; Correia Filho, V.D.; Barata, C.H.; Lages, E.; Ramirez, L.E.; Prata, A. Isospora belli infection: Observation of unicellular cysts in mesenteric lymphoid tissues of a Brazilian patient with AIDS and animal inoculation. J. Eukaryot. Microbiol. [CrossRef]
- Oliveira, F.C.R.; Stabenow, C.S.; Massad, F.V.; Lopes, C.W.G. Hypnozoites of Cystoisospora Frankel, 1977 (Apicomplexa: Cystoisosporinae) in Mongolian gerbil lymph nodes and their transmission to cats free of coccidia. Rev. Bras. Parasitol. Vet. [PubMed]
- Brösigke, S. Untersuchungen an extraintestinalen entwicklungsstadien (Dormozoiten) von Cystoisospora rivolta der Katze in der Maus. Dissertation zur Erlangung der tiermedizinnischen Doktor der Tierarztlichen, Muchen, 1981, 37 pp.
- Dubey, J.P. Life cycle of Isospora rivolta (Grassi 1879) in cats and mice. J. Protozool., 1979, 26, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Mehlhorn, H.; Markus, M.B. Electron microscopy of stages of Isospora felis of the cat in the mesenteric lymph nodes of the mouse. Z. Parasitenkd., 1976, 51, 25–29. [CrossRef]
- Boch, J.; Göbel, E.; Heine, J.; Erber, M. Isospora-Infektionen bei Hund und Katze. Berl Munch Tierarztl Wochenschr, 1981, 94, 384–391.
- Markus, M.B. The hypnozoite of Isospora canis. S. Afr. J. Sci. 1983, 79, 117. [Google Scholar]
- Lindsay, D.S.; Houk, A.E.; Mitchell, S.M.; Dubey, J.P. Developmental Biology of Cystoisospora (Apicomplexa: Sarcocystidae) Monozoic Tissue Cysts. J. Parasitol. [CrossRef]
- Long, P.L. The biology of the coccidia. Univ. Park Press: Baltimore, 1982, 502 pp.
Figure 1.
Cystoisospora ohioensis-like oocysts shed daily by dogs. a, Total oocysts excreted by six naturally infected 8-week-old puppies. b, Mean daily excretion of oocysts per gram of feces (OoPG) divided by 1,000 (OoPG/1,000) from two puppies inoculated with viscera from five mice previously infected with 10⁵ sporulated oocysts of C. ohioensis-like species.
Figure 1.
Cystoisospora ohioensis-like oocysts shed daily by dogs. a, Total oocysts excreted by six naturally infected 8-week-old puppies. b, Mean daily excretion of oocysts per gram of feces (OoPG) divided by 1,000 (OoPG/1,000) from two puppies inoculated with viscera from five mice previously infected with 10⁵ sporulated oocysts of C. ohioensis-like species.
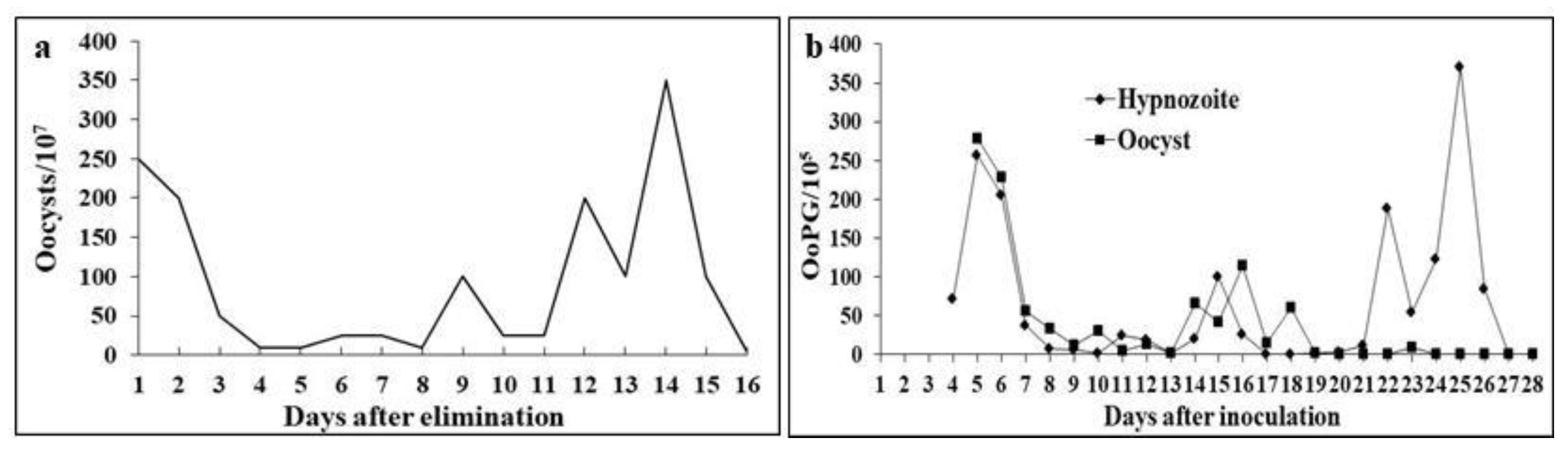
Figure 2.
Photomicrograph of sporulated oocysts of Cystoisospora ohioensis-like species excreted in feces by (a) naturally infected 8-week-old dogs and (b) dogs experimentally infected with 10⁵ sporulated oocysts at 35 days of age. Highlighted in (a) sporozoites (arrow) and (b) fine granular residual bodies (arrowhead). Scale bar: 5 µm.
Figure 2.
Photomicrograph of sporulated oocysts of Cystoisospora ohioensis-like species excreted in feces by (a) naturally infected 8-week-old dogs and (b) dogs experimentally infected with 10⁵ sporulated oocysts at 35 days of age. Highlighted in (a) sporozoites (arrow) and (b) fine granular residual bodies (arrowhead). Scale bar: 5 µm.
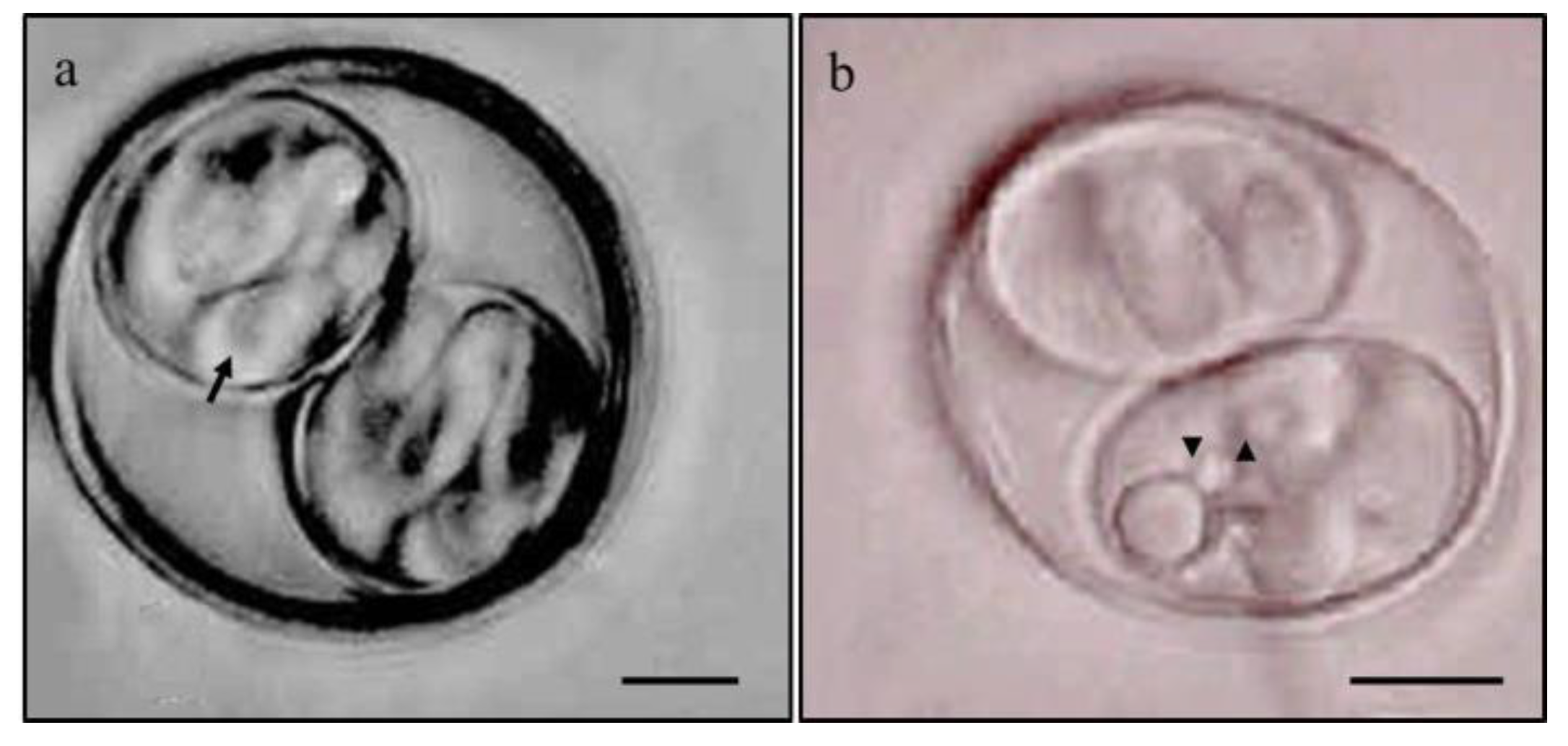
Figure 3.
Mean body weight development of mice inoculated with sporulated oocysts of Cystoisospora ohioensis-like species (Infected), pair-fed according to the mean daily consumption of their infected group counterparts (Pair-Fed), and fed ad libitum (Control): (a) Daily average live weight in grams over 35 experimental days (ALW/g); (b) average carcass weight on days when five animals from each group were euthanized (ACW/g); (c) daily average weight gain in grams over 35 experimental days (AWG/g); (d) average live weight gain on days when five animals from each group were euthanized; (e) average percentage carcass yield (CY%) on days when five animals from each group were euthanized; and (f) average live weight minus average carcass weight (Viscera weight) on days when five animals from each group were euthanized.
Figure 3.
Mean body weight development of mice inoculated with sporulated oocysts of Cystoisospora ohioensis-like species (Infected), pair-fed according to the mean daily consumption of their infected group counterparts (Pair-Fed), and fed ad libitum (Control): (a) Daily average live weight in grams over 35 experimental days (ALW/g); (b) average carcass weight on days when five animals from each group were euthanized (ACW/g); (c) daily average weight gain in grams over 35 experimental days (AWG/g); (d) average live weight gain on days when five animals from each group were euthanized; (e) average percentage carcass yield (CY%) on days when five animals from each group were euthanized; and (f) average live weight minus average carcass weight (Viscera weight) on days when five animals from each group were euthanized.
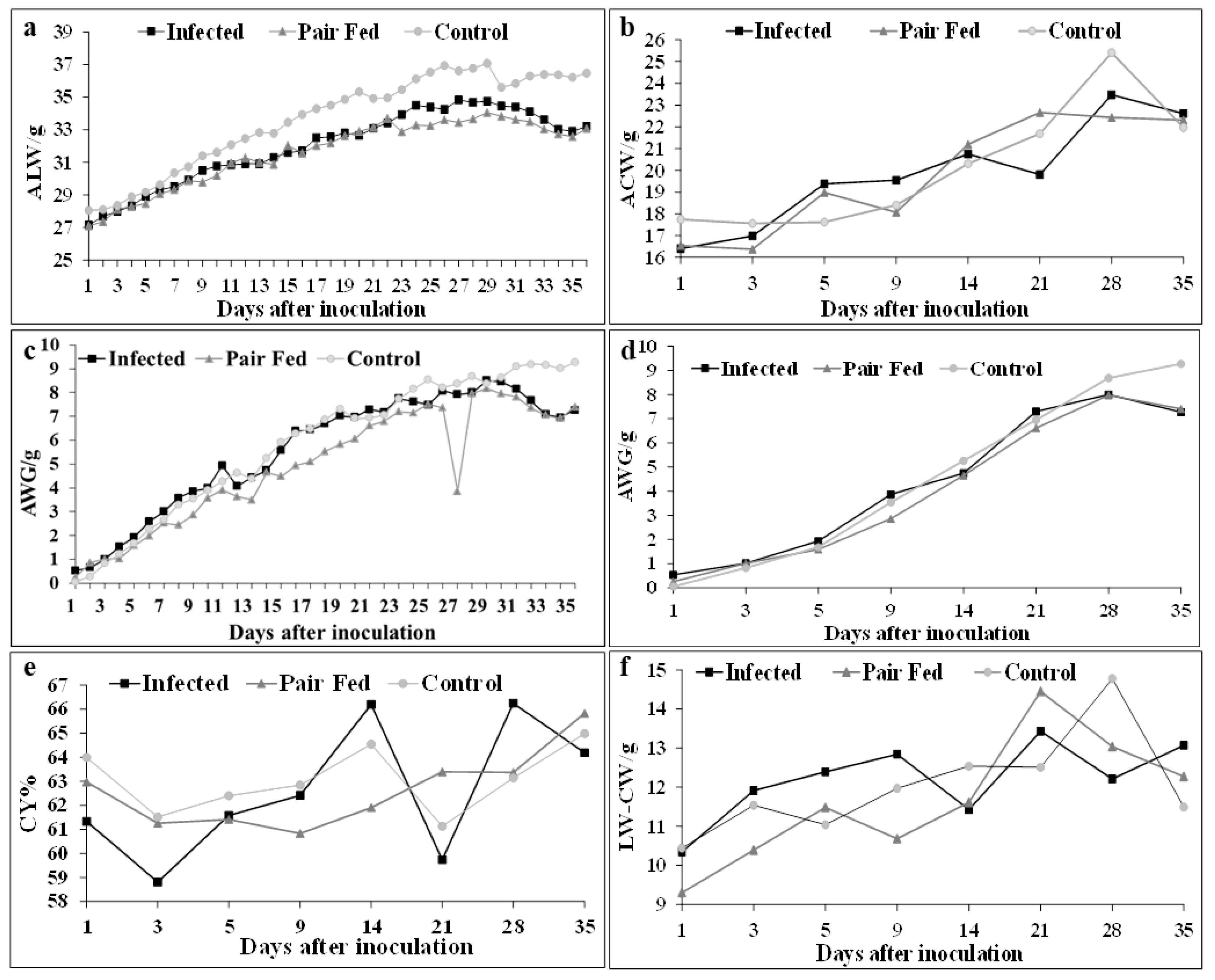
Figure 4.
Average percentage weights of the small intestine (AWSI%) (a), large intestine (AWLI%) (b), Peyer's patches (AWPP%) (c), mesenteric lymph nodes (AWML%) (d), spleen (AWS%) (e), and liver (AWL%) (f) relative to the carcasses of five mice inoculated orally with 105 sporulated oocysts of Cystoisospora ohioensis-like species (Infected), five fed the average daily intake of their counterparts in the infected group (Pair-Fed), and five fed ad libitum (Control) per evaluation day.
Figure 4.
Average percentage weights of the small intestine (AWSI%) (a), large intestine (AWLI%) (b), Peyer's patches (AWPP%) (c), mesenteric lymph nodes (AWML%) (d), spleen (AWS%) (e), and liver (AWL%) (f) relative to the carcasses of five mice inoculated orally with 105 sporulated oocysts of Cystoisospora ohioensis-like species (Infected), five fed the average daily intake of their counterparts in the infected group (Pair-Fed), and five fed ad libitum (Control) per evaluation day.
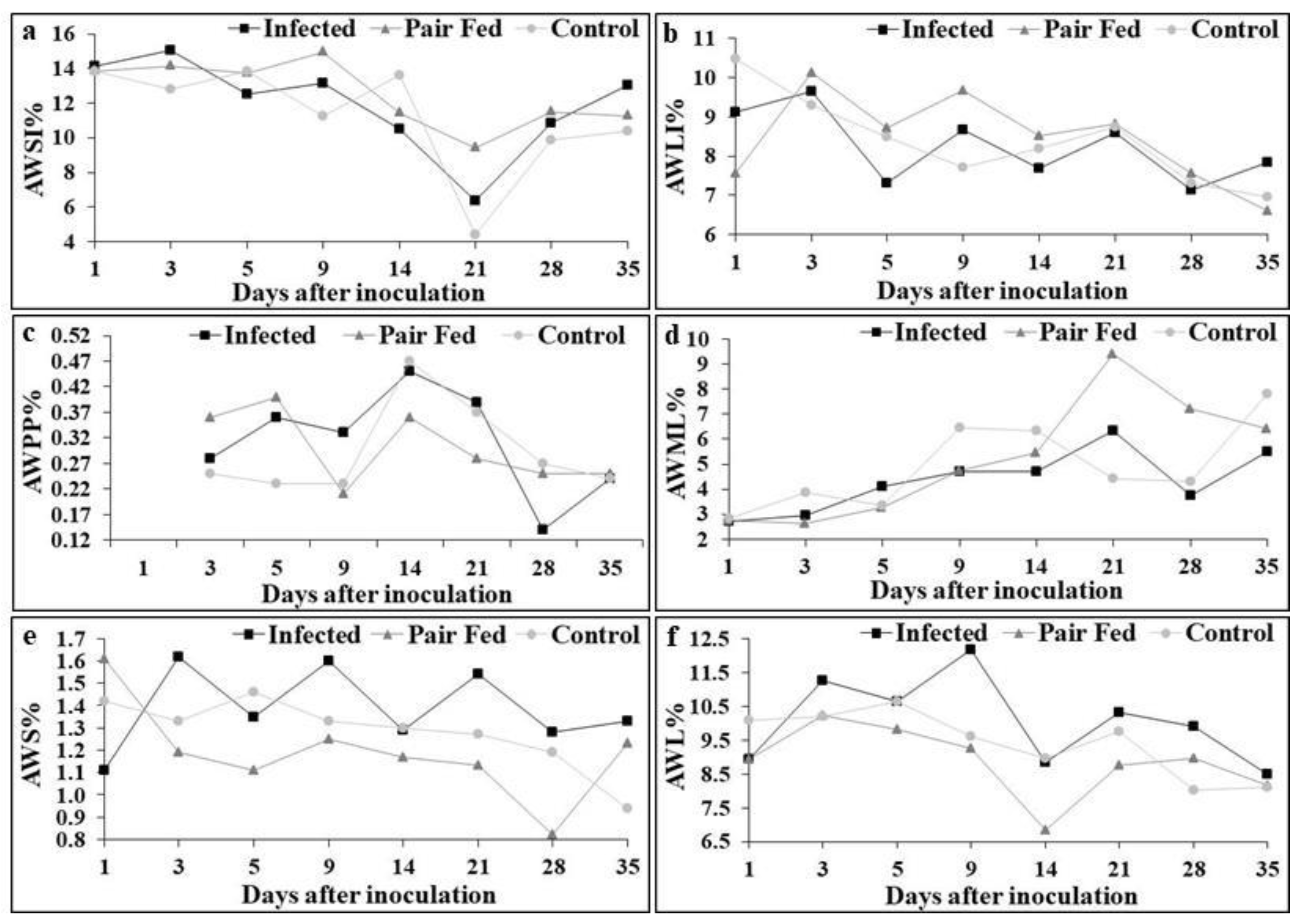

Table 1.
Dimensions in micrometers of oocysts and sporocysts and morphometric indices of Cystoisospora ohioensis-like organisms, excreted by naturally infected dogs, experimentally infected with sporulated oocysts (sporozoites), and fed viscera (hypnozoites) from five mice infected with sporulated oocysts.
Table 1.
Dimensions in micrometers of oocysts and sporocysts and morphometric indices of Cystoisospora ohioensis-like organisms, excreted by naturally infected dogs, experimentally infected with sporulated oocysts (sporozoites), and fed viscera (hypnozoites) from five mice infected with sporulated oocysts.
| Route of Infection | na | Diameters | Morphometric | |
| Measured forms | Major | Minus | Indice | |
| Natural | ||||
| Oocyst | 100 | 21.39±1.71 (18.40-26.80) | 19.23±1.91 (14.40-25.00) | 1.11±0.08 (0.98-1.42) |
| Sporocyst | 100 | 15.48±1.42 (12.20-19.00) | 10.24±0.96 (8.40-13.00) | 1.51±0.13 (1.08-1.83) |
| Experimental using sporozoites | ||||
| Oocyst | 100 | 25.28±2.12 (19.84-29.76) | 23.32±2.18 (17.67-27.59) | 1.08±0.06 (1.00-1.34) |
| Sporocyst | 100 | 18.02±1.91 (13.02-21.70) | 11.59±1.57 (8.68-19.84) | 1.57±0.12 (1.00-1.97) |
| Experimental using hypnozoites | ||||
| Oocyst | 100 | 26.42±1.69 (22.32-30.07) | 24.47±1.36 (21.39-27.28) | 1.08±0.04 (1.00-1.20) |
| Sporocyst | 100 | 18.97±1.77 (11.47-22.63) | 12.00 (16.43-7.13) | 1.59±0.17 (1.13-2.07) |
aNumber of oocysts measured.
Table 2.
Average live weight in grams (ALW) of mice inoculated with sporulated oocysts of Cystoisospora ohioensis-like organisms (INFECTED), fed the average daily intake of their counterparts in the infected group (PAIR-FED), and fed ad libitum (CONTROL).
Table 2.
Average live weight in grams (ALW) of mice inoculated with sporulated oocysts of Cystoisospora ohioensis-like organisms (INFECTED), fed the average daily intake of their counterparts in the infected group (PAIR-FED), and fed ad libitum (CONTROL).
| DAIa | nb | GROUPSc | |||||||
| INFECTED | PAIR-FED | CONTROL | |||||||
| 0 | 45 | 27.22±2.74 | 27.11±2.22 | 28.04±2.80 | |||||
| 1 | 45 | 27.69±2.88 | 27.28±2.39 | 28.10±2.60 | |||||
| 3 | 40 | 28.33±3.18 | 28.30±2.01 | 28.88±2.77 | |||||
| 5 | 35 | 29.30±3.44 | 29.06±2.30 | 29.61±2.82 | |||||
| 9 | 30 | 30.77±2.86 | 30.19±2.56 | 31.28±3.22 | |||||
| 14 | 25 | 31.60±3.04 | 32.02±3.07 | 33.44±3.18 | |||||
| 15 | 20 | 31.70±2.78 | (K) | 31.57±2.44 | (K,Y) | 33.94±3.33 | (Z) | ||
| 16 | 20 | 32.66±2.70 | (KY) | 32.02±2.36 | (Y) | 34.30±3.35 | (K,Z) | ||
| 17 | 20 | 32.56±2.44 | (KY) | 32.18±2.36 | (Y) | 34.50±3.54 | (K,Z) | ||
| 18 | 20 | 32.80±2.25 | (KY) | 32.62±2.58 | (Y) | 34.86±3.60 | (K,Z) | ||
| 19 | 20 | 32.14±3.20 | (K) | 32.90±2.66 | (K,Y) | 35.32±3.64 | (Y,Z) | ||
| 21 | 20 | 33.42±2.21 | 33.75±3.05 | 34.95±3.93 | |||||
| 25 | 15 | 34.25±1.83 | (K) | 33.60±2.45 | (K,Y) | 36.93±4.50 | (Y,Z) | ||
| 26 | 15 | 34.83±2.02 | (KY) | 33.45±2.44 | (Y) | 36.61±4.31 | (K,Z) | ||
| 27 | 15 | 34.68±2.01 | (KY) | 33.66±2.59 | (Y) | 36.76±4.46 | (K,Z) | ||
| 28 | 15 | 34.75±2.03 | (K,Y) | 34.06±2.70 | (Y) | 37.07±4.34 | (K,Z) | ||
| 35 | 10 | 33.22±4.73 | 33.08±3.42 | 36.47±4.09 | |||||
aDays after inoculation. bNumber of samples in each group. cLetters, in parentheses, equal in the lines, ALW does not differ according to the Tukey Test (p ≥0.05) with a 95% confidence interval.
Table 3.
Average weight gain in grams (AWG) of mice inoculated with sporulated oocysts of Cystoisospora ohioensis-like organisms (INFECTED), fed the average daily intake of their counterparts in the infected group (PAIR-FED), and fed ad libitum (CONTROL).
Table 3.
Average weight gain in grams (AWG) of mice inoculated with sporulated oocysts of Cystoisospora ohioensis-like organisms (INFECTED), fed the average daily intake of their counterparts in the infected group (PAIR-FED), and fed ad libitum (CONTROL).
| DAIa | nb | GROUPSc | |||||||
| INFECTED | PAIR-FED | CONTROL | |||||||
| 1 | 45 | 0.53±0.61 | (K) | 0.25±0.75 | (K,Y) | 0.05±0.66 | (Y,Z) | ||
| 2 | 40 | 0.66±0.76 | (KY) | 0.87±0.98 | (Y) | 0.28±0.76 | (K,Z) | ||
| 3 | 40 | 1.01±0.98 | 1.01±0.98 | 0.82±1.11 | |||||
| 5 | 35 | 1.93±1.34 | 1.58±1.33 | 1.68±1.12 | |||||
| 8 | 30 | 3.58±1.45 | (K) | 2.45±1.13 | (Y,Z) | 3.30±1.66 | (K,Z) | ||
| 9 | 30 | 3.86±1.47 | (K) | 2.86±1.28 | (Y,Z) | 3.53±1.70 | (K,Z) | ||
| 14 | 25 | 4.74±2.03 | 4.65±1.30 | 5.26±1.54 | |||||
| 15 | 20 | 5.59±1.61 | (K,Y) | 4.48±1.61 | (Y) | 5.93±1.58 | (K,Z) | ||
| 16 | 20 | 6.39±1.53 | (K) | 4.94±1.69 | (Y) | 6.29±1.55 | (K,Z) | ||
| 17 | 20 | 6.44±1.85 | (K) | 5.10±1.73 | (Y) | 6.49±1.56 | (K,Z) | ||
| 19 | 20 | 7.03±1.91 | (K,Z) | 5.82±1.85 | (K,Y) | 7.32±1.71 | (Z) | ||
| 21 | 20 | 7.30±2.05 | 6.61±1.92 | 6.95±2.02 | |||||
| 27 | 15 | 7.93±1.93 | (K) | 3.84±1.33 | (Y) | 8.37±2.32 | (K,Z) | ||
| 28 | 15 | 8.00±2.24 | 7.97±2.22 | 8.68±2.26 | |||||
| 35 | 10 | 7.27±4.71 | 7.41±3.29 | 9.27±2.47 | |||||
aDays after inoculation. bNumber of samples in each group. cLetters, in parentheses, equal in the lines, AWG does not differ according to the Tukey Test (p ≥0.05) with a 95% confidence interval.
Table 4.
Average carcass weights in grams (ACW) relative to live weights of mice inoculated with sporulated oocysts of Cystoisospora ohioensis-like organisms (INFECTED), fed the average daily intake of their counterparts in the infected group (PAIR-FED), and fed ad libitum (CONTROL).
Table 4.
Average carcass weights in grams (ACW) relative to live weights of mice inoculated with sporulated oocysts of Cystoisospora ohioensis-like organisms (INFECTED), fed the average daily intake of their counterparts in the infected group (PAIR-FED), and fed ad libitum (CONTROL).
| DAIa | nb | GROUPSc | |||||||
| INFECTED | PAIR-FED | CONTROL | |||||||
| 1 | 5 | 16.40±0.65 | 16.54±1.12 | 17.75±2.75 | |||||
| 3 | 5 | 16.99±1.43 | 16.38±0.74 | 17.56±1.65 | |||||
| 5 | 5 | 19.38±1.35 | 18.98±0.36 | 17.62±1.91 | |||||
| 9 | 5 | 19.55±1.05 | 18.07±1.55 | 18.40±1.31 | |||||
| 14 | 5 | 20.75±3.59 | 21.19±3.11 | 20.30±1.65 | |||||
| 21 | 5 | 19.81±2.13 | (K,Z) | 22.66±0.41 | (Y) | 21.68±0.93 | (Y,Z) | ||
| 28 | 5 | 23.47±0.92 | (K,Y) | 22.43±1.30 | (Y) | 25.40±1.54 | (K,Z) | ||
| 35 | 5 | 22.61±1.00 | 22.31±2.41 | 21.96±1.30 | |||||
aDays after inoculation. bNumber of samples in each group. cLetters, in parentheses, equal in the lines, ACW does not differ according to the Tukey Test (p ≥0.05) with a 95% confidence interval.
Table 5.
Hypnozoites quantified by recovery from the viscera of five mice inoculated with sporulated oocysts of Cystoisospora ohioensis-like organisms.
Table 5.
Hypnozoites quantified by recovery from the viscera of five mice inoculated with sporulated oocysts of Cystoisospora ohioensis-like organisms.
| DAIa | HYPOZOITES/VISCERA | TOTAL | |||||||||||||||
| Small intestine | Large intestine | Peyer's patch | Mesenteric lymph node | Spleen | |||||||||||||
| nb | % | n | % | n | % | n | % | n | % | n | % | ||||||
| 1 | 102 | 17.62 | 147 | 25.39 | 135 | 23.31 | 86 | 14.85 | 109 | 18.83 | 579 | 100 | |||||
| 3 | - | - | - | - | 50 | 7.17 | 506 | 72.60 | 141 | 20.23 | 697 | 100 | |||||
| 5 | - | - | - | - | 28 | 25.00 | 44 | 39.29 | 40 | 35.71 | 112 | 100 | |||||
| 9 | - | - | - | - | 9 | 4.89 | 60 | 32.61 | 115 | 62.50 | 184 | 100 | |||||
| 14 | - | - | - | - | - | - | 38 | 18.45 | 168 | 81.55 | 206 | 100 | |||||
| 21 | - | - | - | - | - | - | 60 | 23.90 | 191 | 76.10 | 251 | 100 | |||||
| 28 | - | - | - | - | - | - | 35 | 12.96 | 235 | 87.04 | 270 | 100 | |||||
| 35 | - | - | - | - | - | - | 5 | 4.42 | 108 | 95.58 | 113 | 100 | |||||
| GRAND TOTAL | 102 | 4.23 | 147 | 6.09 | 222 | 9.20 | 834 | 34.58 | 1.107 | 45.90 | 2.412 | 100 | |||||
aDays after inoculation. bNumber of hypnozoites per viscera.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



