
Submitted:
18 May 2025
Posted:
19 May 2025
You are already at the latest version
Abstract
Droughts are predicted to intensify with climate change, posing a serious threat to global crop production. Increasing drought tolerance in plants requires understanding of underlying mechanisms. This study measured physiological, phytohormonal and transcriptomic responses to drought in two apple rootstocks to identify drought ‘biomarkers’ and investigated whether root application of acibenzolar-S-methyl (ASM) could enhance drought tolerance. Two potted-plant trials were conducted on dwarfing (M9) and semi-dwarfing (CG202) apple rootstocks. In both trials, response patterns in roots and leaves were compared in irrigated and non-irrigated plants over a 14-day period. In trial 2, ASM was applied 14-days before and immediately before with-drawing irrigation. Drought induced significant decreases in transpiration, photosynthesis and stomatal conductance in both trials. This was accompanied by accumulation of abscisic acid (ABA) metabolites and upregulation of ABA pathway transcripts (CYP707A1/A2 & NCED3), a decrease in 12-oxophytodienoic acid (cis-OPDA) and downregulation of ABA-receptor genes (PYL4). Responses to drought were greater in roots than leaves, broadly similar across both rootstocks, but differed in strength and timing between the rootstocks. Root application of ASM did not significantly affect responsiveness to drought in either rootstock. The identified phytohormonal and transcriptomic biomarkers require further validation across a broader range of genotypes.
Keywords:
ABA metabolism
; abiotic stress
; Malus domestica
; plant hormones
; water stress
1. Introduction
Drought is one of the most impactful abiotic factors affecting yield in agricultural and horticultural systems [1,2,3,4]. For example, Kim et al. [5] analysed historical drought impact and global production data and showed an average yield reduction for annual crops in a single drought period of 8% for wheat, 7% for maize, 3% for rice and 7% for soy. Yield losses tend to be greater in slower growing perennials, e.g., 26% loss was observed in apple in a single season [6], and can be further exacerbated because water deficit can adversely affect flower initiation, shoot length and branching, leading to decreased yield in subsequent growing seasons [7]. With climate change, the frequency, intensity, duration and geographic spread of drought periods are predicted to increase globally [5]. Furthermore, agriculture and horticulture are the biggest consumers of freshwater [3] and global demands for food (hence water) are increasing, placing even more pressure on a limited resource. Future crop yields and worldwide food security are at risk if we cannot address this issue.
Plants respond to drought by physical and chemical changes mediated by their transcriptome. Gene responses to drought have been intensely studied [3,4,8,9], with numerous reviews showing that key components include genes of the abscisic acid (ABA) pathway [10,11,12]. ABA is the primary phytohormone involved in responses to abiotic stresses such as water extremes (drought, flooding), temperature stress (both hot and cold), and excessive salinity [10,11,13,14]. ABA is a key mitigator of the plant drought response because it quickly stimulates short-term physiological changes, such as stomatal closure, thus regulating water loss through transpiration. In addition, ABA also regulates longer term responses, such as dormancy and growth inhibition via regulation of signaling peptides and various drought-responsive genes, e.g., genes encoding for membrane and protein stabilization, antioxidant activity, and biosynthesis of osmolytes. [11,12,15]. While considerable progress has been made in understanding phytohormone roles in the plant response to abiotic stress, there are still gaps in our knowledge, including a clear understanding of the complex effects of hormonal crosstalk and multiple stresses, understanding the potential trade-off between induced stress responses and yield, as well as the effects of innate vs induced tolerance in different plant varieties. Moreover, the rationale behind most of these cited studies is to provide an understanding of molecular responses to water stress, with a view to breeding new plant varieties with improved drought resilience [3]. In contrast, there are considerably fewer studies, particularly in perennials, that use combined measurements of phytohormonal and transcriptional biomarkers to understand how treatments can be applied to established cultivars to induce greater tolerance to subsequent drought stress in existing plantings, in a process known as priming [16,17,18].
Treatments that can be used to condition plants to improve their stress tolerance include gradual exposure to a stress factor, e.g., controlled water deprivation [19], and/or the exogenous application of compounds (priming agents) such as phytohormones and plant metabolites [20]. Experimental applications of phytohormones, including salicylic acid (SA) [21,22], cytokinins (CK) [23], and ABA [24], have demonstrated potential benefit in improving drought resilience. However, there are few commercialized priming agents to mitigate the impacts of drought stress, and these are largely restricted to use in annuals, rather than woody perennials because of the additional biological complexities and time constraints of slower growth cycles associated with the latter. The impact of severe drought on existing plantings of perennials has more durable and deleterious effects than on annuals, with the time taken to reach full production after re-planting requiring years, hence the use of priming agents in perennials merits further research. One suitable candidate for such research is acibenzolar-S-methyl (ASM), a commercially available SA analogue that is often used to induce biotic stress resistance. Exogenous application of SA activated stress response molecular pathways and alleviated water stress in watermelon, rice, wheat, and barley [25,26,27,28]. SA also triggered the accumulation of ABA under both normal and salinity stressed tomato plants, which enhanced osmotic adaptation and improved photosynthetic pigments and growth [29]. Furthermore, application of ASM has been shown to enhance drought tolerance in creeping bentgrass [30], induce stomatal closure in Japanese radish [31], and stimulate ABA accumulation in kiwifruit [32], which might improve tolerance to osmotic stress.
The exemplar crop chosen for this study was apple. Drought stress can affect apple quality and yield in both the current and subsequent seasons [33,34], and may cause tree death. Grafted apple trees take at least seven years to reach full production, and breeding of new cultivars can take up to 15 years [35,36], leaving the industry with a significant recovery period should existing plantings suffer severe drought. Commercial apple plantings comprise scions grafted onto rootstocks, with the rootstocks performing a key role in controlling tree growth and drought tolerance characteristics [37]. Rootstock genotype has been shown to have a major influence on the response of the scion to water stress [38]. Malling 9 rootstock (M9) is a commonly used strong-dwarfing rootstock thought to confer some drought resistance via ABA-mediated control of stomata [39]. Cornell-Geneva 202 (CG202), a semi-dwarfing rootstock, appears to have reduced drought tolerance relative to M9 [40], but the reverse result has also been observed [41]. Results from most studies on apple rootstocks are based on empirical physiological data without any matching data on biochemical and genetic biomarkers [39,40,41,42,43,44].
To address gaps in our knowledge, the aims of this study were:
- To identify robust phytohormonal and transcriptomic biomarkers that align with physiological drought responses in two apple rootstocks
- To determine the effect of exogeneous ASM application on these biomarkers and compare induced responses in the two rootstocks
The underlying hypotheses are that phytohormonal and transcriptional markers can be used to study drought responses in rootstocks from a metabolic angle, and that the SA analogue, ASM, can be used to induce/prime for drought resistance in a woody perennial (apple).
The study reported here provides the first results from a research programme that adopts a novel approach by the combined use of selected phytohormones and genetic targets, previously associated with stress response and signaling in apple and annual model plants. This provides more understanding into drought resilience mechanisms at a systems level than use of either group of biomarkers on its own and could be used in the future to examine the relationship between the timing and the amplitude of the biochemical and transcriptional responses.
2. Results
2.1. Gas Exchange and Water Loss
Two trials were carried out, with Trial 2 undertaken to validate the findings from Trial 1 and to further build on the learnings by using a priming agent (ASM). Pot weight was used to monitor the relative reduction in soil-water content in non-irrigated (Dry) pots over the duration of each experiment. Pot weight in the Irrigated plants was maintained throughout the course of the experiment for both trials. In comparison the Dry plants lost weight throughout the trial period, because of water loss, resulting in increasing drought stress over the 14 days (Table 1 and Table 2). Loss of pot weight in the Dry plants was similar between rootstocks and trials, suggesting similar water stress levels applied. However, the average temperature over the 14-day period differed between Trial 1 (21.7°C) and Trial Year 2 (18.1°C) (Supplementary Figure S1). The difference was particularly evident during days 1-5, with maximum temperature up to 8°C higher in Trial 1 and average temperature of 22.6 °C in Trial 1 compared with 17.1 °C in Trial 2.
Water deprivation induced a reduction in transpiration in both CG202 and M9 rootstocks with the reduction occurring earlier in the former genotype (Figure 1, A, B). In Trial 1, transpiration in CG202 was significantly lower in the Dry (non-irrigated) plants than in the Irrigated control plants on days 9 (p = 0.010) and 14 (p < 0.001), whereas in M9 the difference was significant only at day 14 (p < 0.01). Similarly, in Trial 2, transpiration in CG202 was significantly lower on days 5 (p = 0.047), 9 (p < 0.001) and 14 (p < 0.001) in Dry plants, compared with the Irrigated control plants whereas in M9 the difference was significant only at day 14 (p < 0.001), but the magnitude of the transpiration changes was smaller in Trial 2 vs Trial 1. (Figure 1 A, B)
Photosynthesis measurements show an effect of the Dry treatment over time for both rootstocks in both trials. In Trial 1, plants in the Dry treatment showed significantly reduced photosynthesis rate on day 14 compared to the Irrigated treatment (p<0.001 for both rootstocks). In Trial 2, photosynthesis was reduced after 9 days treatment compared to the Irrigated treatment in the CG202 rootstock (p = 0.064). In the M9 rootstock there was a reduction after 5 (p = 0.053) and 14 (p = 0.059) days in the Dry treatment. (Figure 1 C, D)
The effect of Dry treatment on stomatal conductance for both rootstocks in both trials followed the same pattern as the transpiration rate. In Trial 1, the Dry treatment reduced stomatal conductance compared to the Irrigated treatment for each rootstock. In the CG202 genotype transpiration was significantly reduced in days 9 (p = 0.024) and 14 (p = 0.001) in the Dry treatment compared to the Irrigated treatment. In the M9 rootstock there was a significant difference only at day 14 (p = 0.032). In Trial 2, a similar response was observed with a significant reduction occurring in CG202 at days 5, 9, and 14 (p = 0.021, < 0.01, < 0.01, respectively) and in M9 on day 14 (p = 0.022) (Figure 1 E, F)
There was no significant effect of the ASM treatment (only tested in Trial 2, data not shown).
2.2. Gene Expression Data – Selection of Suitable Markers
The expression of 53 genes of interest (GoI) and six reference genes (RG) were measured during the onset of drought stress in leaf and root tissues. Selection of GoIs was based on mining a list of candidate genes identified from both apple and model plant literature on ABA metabolic synthesis and catabolic pathways, and functional analysis of genes involved in drought resistance, whilst RGs were stably expressed apple genes identified from literature (Supplementary Table S1). Gene expression data presented here are limited to those that showed statistically significant differential expression and/or consistent trends in both Trial 1 and Trial 2 (for those genes measured in both trials). Statistically significant expression was usually less than 0.5-fold for downregulated genes and greater than 2-fold for upregulated genes (Figure 2 and Figure 3). All other gene expression results can be found in the supplementary section (Supplementary Figures S2 – S9).
Heat maps show fold change expression (log2 transformed count data that has been backtransformed) relative to Irrigated control plants at each time point for Trials 1 and 2 respectively (Figure 2 and Figure 3). In Trial 1, water deficit induced significant upregulation of NCED3 (ABA-synthesis), CYP707A1/A2 (encode cytochrome P450 monooxygenases involved in ABA catabolism), and RD29B (ABA-inducible) gene families in both rootstock genotypes (Figure 2). Gene upregulation depended on isoform and was generally greater in roots than leaves and greater in CG202 than M9. For example, after 9 days without water, NCED3_iii increased by 31-fold in roots and 9-fold in leaves of CG202 plants compared with 12-fold and 4-fold increases respectively, in M9 plants (Figure 2). The expression of CYP707A2, which catabolizes ABA, was upregulated in roots only and, after 14 days without water, was 59-fold higher in CG202 and 30-fold higher in M9 relative to irrigated plants. The most highly upregulated gene in the roots of both rootstocks, RD29B_i, was upregulated by over 80-fold in CG202 and over 50-fold in M9 after 9 days. This compares with more moderate upregulation in leaves where RD29B_i expression increased by approximately 9-fold and 2-fold in CG202 and M9 plants, respectively, after 14 days without water (Figure 2). In contrast, water deficit resulted in significant downregulation of four members of the PYL4 family of ABA receptors (that play a key role in ABA signalling and the drought response). All four isoforms were downregulated, with 11- to 50-fold decreases after 9 days without water in roots of both GC202 and M9 (Figure 2). PYL4 fold decreases occurred earlier in leaf tissue of GC202 vs M9, with 14-to 100-fold decreases occurring by 9 days in non-irrigated GG202 leaves vs 14 days to reach similar fold decreases in M9 leaves (Figure 2).
The patterns of gene expression for the upregulated gene families (NCED3, CYP707A1/A2, and RD29B) and downregulated family l(PYL4) that were seen in Trial 1 were also observed in Trial 2 (Figure 3). Whilst the patterns of gene expression were consistent between the two trials, the amplitude of the response was much greater in Trial 1 than in Trial 2. For example, for the most highly upregulated gene, RD29B_i, the maximum fold change in the root tissue was 83-fold in CG202 and 57-fold in M9 after 9 and 14 days without water respectively in Trial 1 (Figure 2). In contrast, maximum fold increases in Trial 2 for RD29B_i in root tissue were14-fold in CG202 and 5-fold in M9 after 14 days without water (Figure 3). Similarly, the two most down-regulated isoforms, PYL4_i and PYL4_ii, had decreased by 100-fold in GC202 and M9 leaf tissue after 14 days without water in Trial 1, versus PYL4_i and PYL4_ii-averaged decreases of 25-fold and 5-fold for CG202 and M9 leaves, respectively, after 14 days of drought in Trial 2 (Figure 2 and Figure 3).
Analysis of the 42 GoI in Trial 1 showed that transcript abundances of 11 genes (PYL9_i, PYL9_ii, PYL9_iii, ERD15_i, RAP2.4_ii, FKBP_ii, VHA-B_i, VHA-B_ii, SAT1, HY5, and SERRATE) from the original gene set did not differ significantly in expression between treatments (Figure S2). These genes were replaced by 11 new candidates mined from the literature (OST1/SnRK 2.6, SnRK 2.3, SnRK 2.8, ATG18_i, MADS-box AGL16_i, MADS-box AGL16_ii, SDH1, BES1/BZR2, MdbHLH130, MdNAC1, MdNAC143) in the revised gene set used for Trial 2. Of these newly added genes, only two, MdbHLH130 and SDH1, showed significant upregulation (Figure 3 and Figures S3-S4). MdbHLH130, a transcription factor involved in regulating stomatal closure and reactive oxygen species homeostasis, increased up to 3-fold and 2-fold in CG202 and M9 roots respectively after 14 days of drought, but there was no significant increase in the leaves (Figure 3 and Figure S4). SDH1, involved in sugar metabolism, increased up to 6-fold and 3-fold in roots of CG202 and M9 respectively in roots after 14 days of drought, and up to 3-fold in GC202 leaves but without a significant increase in M9 leaves after 14 days without water (Figure 3).
In Trial 2, application of the SA-analogue, ASM, was tested to determine its potential to prime rootstocks for greater drought resilience. Treatment with ASM did not have a significant effect on gene expression relative to the untreated Irrigated and Dry controls (Figure 3). Moreover, the expression profiles for Dry and ASM+Dry treatments were the same, indicating no significant effect of ASM on genes associated with the Dry treatment.
Transcript counts for selected genes are presented to provide information on basal/innate levels of gene families that is not evident from heatmaps. The expression of CYP707A1/2 genes was differentially expressed in the roots and leaves of both rootstock genotypes. In Trial 1, basal level of CYP707A1_i at T0 (baseline) was greater in roots (500-600 transcripts) than in leaves (< 5) whilst the opposite was true for CYP707A1_ii and CYP707A2_i which were more abundant in leaves (400-1200) than in roots (<5) (Figure 4). There was also a differential response to drought in roots with CYP707A1_ii and CYP707A2_i transcripts increasing by 10-fold and 50-fold, respectively, after 14 days whilst levels of CYP707A1_i reduced by over half. The expression of CYP707A genes in leaves fluctuated over time but were not statistically different between Irrigated and Dry treatments after 14 days. Similar expression patterns were observed for CYP707A genes in Trial 2; however, basal transcript levels were higher and the fold-differences in counts between Irrigated and Dry treatments were lower (Figure S5).
PYL4 family genes were strongly expressed in the day 0 baseline measurement for both rootstock genotypes (Figure 5). PYL4_ii was the most abundant isoform in roots at T0, in both genotypes with > 2500 transcripts compared with approximately 1700, 1000 and 700, respectively for PYL4_i, iii and iv. Respective transcript levels in leaves at T0 were lower with 800-1000 copies of PLY4_i and ii, and 70-200 copies of isoforms iii and iv. The abundance of each PYL4 isoform reduced significantly in the Dry treatment after 14 days with PYL4_i & ii transcript levels falling by 100-fold in leaves and by between 14- and 50-fold in roots (Figure 5). PYL4_iii & iv transcript levels reduced by 11- to 20-fold in leaves and by 25-to 50-fold in roots. The response patterns of PYL4 genes in in the Dry treatment in Trial 2 mirrored those in Trial 1; however, as with CYP genes, basal transcript levels were generally higher and fold changes lower in Trial 2 than in Trial 1 (Figure S6).
NCED3 isoforms i-iii were expressed at low levels (<50 copies) in roots and leaves of CG202 and M9 irrigated control plants in Trial 1 (Figure S7). In droughted plants, NCED3_i and NCED3_iii copy number increased to between 300 and 500 in roots, irrespective of rootstock genotype, whilst NCED3 ii levels remained relatively low (Figure S7). The same trends, albeit to lesser degree, were observed for NCED3 isoforms in Irrigated and Dry treatments in Trial 2 (Figure S8). RD29B_i, like NCED3 i-iii, was expressed at low levels (< 50 copies) in the roots of irrigated controls but was one of the most strongly upregulated genes in droughted plants with copy number increasing over 3000 in CG202 and over 1800 in M9 after 14 days in Trial 1 (Figure S9).
2.3. Phytohormone Analysis
Compounds associated with SA, JA and ABA metabolism in addition to the plant growth promotor IAA were targeted for analysis from root and leaf tissues of the two apple rootstock genotypes. Fifteen compounds were identified in Trial 1 and these included bioactive forms of stress-related phytohormones (ABA, SA, JA, JA-Ile) and IAA, hydroxylated catabolites (12-OH-JA, 7-OH-ABA, PA, DPA), glucose conjugates (ABA-GE, SAG), precursors for SA (benzoic acid) and JA (cis-OPDA, OPC-4) biosynthesis and DHJA. Of these compounds, benzoic acid, 7-OH-ABA and ABA-GE were detected only in root tissue and 12 metabolites were common to both plant tissues. The phytohormone compositions of these common metabolites were primarily influenced by tissue type with roots accumulating higher concentrations of jasmonates and IAA (Figure S10), whereas ABA and SA-derived metabolites were predominant in leaves of both rootstock genotypes.
To explore the relationships between multiple explanatory variables (genotype, treatment, sampling day) and phytohormone metabolites as response variables in each plant organ separately, redundancy analysis (RDA) was performed. Permutation testing of the respective models was significant (both p < 0.01) for roots (F = 14.3) and leaves (F = 13.6) with genotype explaining the greatest significant (both p < 0.001) variation in phytohormone composition (Froots = 21.4, Fleaves = 32.3), followed by treatment for roots (F = 13.8) and sampling day for leaves (F = 10.7).
As a type of constrained principal component analysis, RDA experimental factors that explain the most variation in the response data were visualised as sample plots (Figure S10) and loading plots (Figure 6). The F-values were significant (all p < 0.003) for RDA1 (F = 38.7) - RDA5 (F = 3.3) and RDA1 (F = 34) - RDA4 (F = 5.2) in roots and leaves, respectively, indicating that the observed differences between groups are unlikely due to chance. For roots, the sample plots show that variation between genotypes separated on the second component and variation in response to treatment on the first component (Figure S11A). Phytohormone responses to drought were already apparent from day 9 with sampling day 14 of Dry plants showing most prominent separation in both rootstock genotypes. These trends are summarised in Figure 6A, which highlight the relationships of response variables to each other and to the principal RDA components as vectors. The length of each vector resembles the relative weight of the variable in the component and its angle indicates the correlation with other variables. This demonstrates that variation in phytohormone profiles between genotypes was primarily driven by higher concentrations of JA and cis-OPDA in CG202. Responses to the Dry treatment were positively associated with IAA, ABA, ABA-GE and PA. In leaves, samples separated by genotype on the first component (Figure S11B) and by treatment on the second component with responses to drought apparent from day 14. Variable loadings (Figure 6B) confirmed that higher JA and JA-Ile concentrations as characteristic for CG202 also in addition to DPA, while M9 showed increased contents of SA, SAG and DHJA as a minor metabolite. Responses to drought were comparable to those observed in roots and confirmed a raise in IAA, ABA and PA, while cis-OPDA showed the opposite trend.
To validate the selection of drought-responsive phytohormones and gain an understanding of magnitude, fold-change (FC) analysis at a 2-FC cut-off was performed between Dry and Irrigated samples (Figure 7). In both genotypes, increased metabolite concentrations were observed for ABA across the entire time course in roots (Figure 7A) and from day 9 in leaves (Figure 7B). PA increased at least 2-fold from day 9 in roots and day 14 in leaves and IAA from day 14 in roots but only for M9 in leaves. Cis-OPDA concentrations decreased 2-fold in roots in response to drought from day 5 in M9 and from day 14 in CG202 and 9-fold in leaves from day 14. In both rootstocks, ABA-GE also increased in roots from day 14, while SAG was 2-fold higher in M9 roots and SA in M9 leaves at this timepoint. CG202 responded to drought with increased 7-OH-ABA concentrations in roots from day 9 and decreased content of bioactive jasmonates at day 5. In conclusion, ABA and PA were identified as positive and cis-OPDA as negative tissue-independent biomarkers of drought stress and IAA and ABA-GE as positive biomarkers of drought stress in roots.
In Trial 2, the use of additional standards and a MS-detector with higher sensitivity resulted in the quantification of five additional metabolites across all samples, namely bioactive methyl jasmonate (MeJA), the ABA catabolite neophaseic acid (NeoPA) and SA associated compounds (cinnamic acid, 2.3-DHB, 3.4-DHB). Moreover, ABA-GE and benzoic acid were reliably quantified from leaves and roots, while 7-OH-ABA and 2.5-DHB were detected only in roots, resulting in 19 metabolites common to both plant tissues. In line with Trial 1, the strongest variation between these phytohormones and associated metabolites was observed between tissue types (Figure S12), with jasmonates and IAA accumulating in roots, and abscisates and salicylates in leaves of both rootstock genotypes. When metabolite compositions were compared for each plant tissue separately (Figure 8), samples clustered in two primary clusters with two subclusters each. For each tissue type the primary cluster grouped genotypes, reflecting higher contents of jasmonates and abscisates in CG202, and salicylates in M9 (Figure 8). Differences between Dry and Irrigated treatments for each genotype were apparent from the subclusters. Although the absolute concentrations of phytohormones and their metabolites differed between genotypes, the response to drought was consistent between apple rootstocks. These trends were not affected by ASM treatment and treated samples clustered directly with their respective untreated samples (Figure 8). Pairwise comparisons between the respective ASM-treated and untreated samples only highlighted a genotype-dependent significant increase for MeJA in M9 (Figure 8A), which was consistent for both, Dry and Irrigated roots, and higher concentrations of NeoPA, a minor catabolite, in Dry M9 roots. Thus, phytohormone profiles appeared to be primarily affected by tissue-type, genotype and drought-treatment, but not ASM-priming.
Since the effect of ASM on phytohormone responses was considered negligible, only non-primed samples were taken forward for RDA. The models significantly (p < 0.01) explained metabolite variations for roots (F = 8.2) and leaves (F = 6.4), which were primarily (p < 0.001) driven by genotype (Froots = 20.5, Fleaves = 18.4,) and treatment (Froots = 10.5, Fleaves = 3.1) as explanatory variables. For both tissues RDA1 and RDA2 were significant (p < 0.001) with FRDA1 > 19.7 and FRDA2 > 7.5. Sampling plots for Trial 2 (Figure S13) reflected the overall trends observed for Trial 1, with samples separating by genotype (on component 1 for both tissues) and in a time-dependent manner by treatment (on component 2 for both tissues). However, drought-induced changes in root metabolite composition became pronounced at a later timepoint (day 14) compared with Trial 1 (day 9). Loading plots (Figure 9) indicate that drought responses in roots and leaves from both genotypes were positively associated with ABA, ABA-GE and PA concentrations at day 14, while IAA and NeoPA were characteristic for roots only. Fold-change analysis (Figure S14), however, was only significant for ABA and PA at day 14 across genotypes and tissue types and for IAA in roots with higher metabolite concentrations measured from Dry plants (Figure 10). In contrast to Trial 1, ABA-GE did not show a 2FC in drought-treated roots. Cis-OPDA concentrations were only positively associated with irrigated roots across genotypes and in leaves for ‘M9’ at day 14 (Figure S14). In leaves, from CG202 cis-OPDA concentrations showed a lower trend trended with a 30% decrease at day 14 in Trial 2.
3. Discussion
This research aimed to establish if transcriptomic and phytohormonal markers could assess the response to drought in dwarfing (M9) and semi-dwarfing (CG202) apple rootstocks, and whether pre-treatment with the SA-analogue, ASM, could induce drought resistance. Drought resulted in a significant decrease in transpiration, photosynthesis and stomatal conductance in both rootstocks. The physiological responses occurred earlier in CG202 than M9 in both trials, suggesting a quicker response to water stress in CG202. Moreover, there was faster and greater accumulation of ABA metabolites (especially in Trial 2), and stronger upregulation of ABA pathway genes (in both trials) in CG202 than in M9. Biochemical and genetic reactions were more intense in the root than in the leaf tissue, with the roots being the primary site of water stress perception. Application of ASM had no discernable effect on the physiological, transcriptomic and biochemical parameters measured. Taken together, these data suggest that statistically significant, differentially accumulated biochemical compounds (ABA, PA, cisOPDA, and IAA) and gene families (CYP707A1/A2, NCED3, RD29B, PYL4, MdbHLH130 and SDH1) showing consistent trends in this study were suitable biomarkers of the drought response and that ASM was not an effective primer – at least within the constraints of this study.
Gas exchange measurements showed that the Dry treatment significantly affected both rootstocks in both trials leading to a significant reduction in the transpiration rate, photosynthesis, and stomatal conductance. Transpiration and stomatal responses generally occurred earlier in CG202 (after 5-9 days without water) than in M9 (after 14 days without water). It is not possible to conclude if the earlier responses in CG202 reflect greater sensitivity or greater resistance and/or resilience to drought. Drought resistance can be described as the ability to maintain growth during a drought and resilience as the ability to recover growth after a drought [45]. The gas exchange data indicated that water stress imposed by day 14 was possibly too severe, as both rootstocks had low photosynthetic rates, transpiration rates and stomatal conductance. The plants were showing severe physical symptoms of water stress. which in turn impaired health equally in both rootstocks, complicating any discussion of drought resistance. Whilst not a direct measurement of stomatal aperture, stomatal conductance measures the rate of gaseous exchange (e.g., CO2 exchange) through the stomata, with a drop indicating stomatal closure [46]. ABA-mediated stomatal closure is a common physiological response to drought [47], which aims to conserve water/limit water loss, as measured by transpiration rate, but which also limits gaseous exchange necessary for photosynthesis. In both trials, 14 days without water resulted in a 50% loss in pot weight and photosynthetic rates were significantly decreased to the same extent in both rootstocks. Visually, CG202 and M9 exhibited similar physical appearances indicative of severe water stress, i.e., leaf wilting/chlorosis, particularly in the top 25% of each plant. For future studies, especially on potted plants, we would recommend a less severe water stress, for example shorter periods of drought deprivation followed by rewatering cycles [40], and/or using water stress treatments that consist of percentage reductions of the maximum water content at pot capacity [44]. Recovery data after a period of re-watering were not collected in this study, so it is also not possible to comment on drought resilience. Recovery following dehydration stress is perhaps of greater significance in perennial crops than annuals and will be considered in our future studies. Information in the literature about the relative drought resistance and/or resilience of CG202 and M9 is conflicting, with Xu and Ediger [40] suggesting that M9 is more drought resistant than CG202, which they defined as more efficient CO2 assimilation, higher net photosynthesis, and smaller declines in stomatal conductance, i.e., more stringent stomatal control, and reduced water use/lower transpiration rates during stress. In contrast, Choi et al. [41] focused on measuring soil and water potential, water use efficiency (WUE), and vegetative growth and dry matter, both during drought stress and recovery periods, and found CG202 to be more drought resilient than M9. They observed that although M9 had better WUE than CG202 at the height of water stress, CG202 exhibited a lower leaf:fine root ratio than M9, which is thought to lessen the impact of xylem embolism (air-filled tracheids and/or vessels) on impairing stem water transport, thus contributing to better recovery post drought stress. The anomalous findings between these studies may also be associated with differences between resistance and resilience definitions as well as experimental differences including the age of the rootstock-scion systems used (1-3 years), the scions used for grafting (‘Ambrosia’ vs ‘Fuji’), the duration and frequency of the drought regimes imposed as well as highly variable environmental conditions and soil characteristics in field trials. Moreover, these results are not directly comparable to the current study on rootstocks alone because the published studies were on grafted rootstock-scions. However, whilst scion will affect results, generally the rootstock is considered to play the more important role in determining drought resilience in grafted apple [38,48]. Interestingly, Xu and Ediger [40] found that rootstock choice influenced both stomatal size and density on a common scion, Ambrosia™, with higher density and smaller sized stomata found on ‘Ambrosia’ leaves grafted onto M9 versus CG202. This may in part explain the more stringent stomatal control that they observed in the ‘Ambrosia’/M9 grafted plants.
To select suitable (transcriptional) gene markers, potential candidates needed to show convincing responses to transcriptional regulation/mRNA expression associated with the drought response, i.e., statistically significant differential expression, consistent patterns over the two trials (for those genes common to both gene sets) and responses that mirrored physiological and biochemical changes. Gene families meeting these requirements were the PYL4, CYP707A1/A2, NCED3, RD29B, MdbHLH130, and SDH1 gene families. Apart from the first gene family, these all started with low basal levels of expression (less than 50 copies) but were significantly upregulated with drought, with a faster and more elevated response occurring in the root tissue of CG202 than M9. The same trends were apparent in leaf tissue of both rootstocks, but overall, upregulation was less than in the root tissue, although expression within some gene families was sometimes tissue- and isoform-specific. The PYL4 gene family was expressed at high basal levels in irrigated plants in roots (up to 5000 copies in both rootstocks) and leaves (up to 1000 copies in both CG202 and M9) and decreased significantly with drought, with greater downregulation occurring in GC202 than M9. This pattern of downregulation was also found for this subfamily in Arabidopsis [49]. All these gene families, except SDH1, are directly or indirectly involved with ABA metabolism. As well as modulating stomatal closure, increased ABA concentrations are known to increase the root:shoot ratio and promote the growth of lateral roots in Arabidopsis [10]. This study did not analyse the transcriptional expression of genes where the predominant modes of regulation identified in the literature involved post-transcriptional, post-translational or epigenetic changes.
Intensity of the stress responses, e.g., differential expression of genes in the Dry treatment, was greater in Trial 1 than in Trial 2. This was reflected in the environmental data, where the temperature was hotter in Trial 1 than in Trial 2, especially during the first few days where the maximum temperature differed by approximately 8°C and the minimum temperature by 10°C degrees. High temperatures often accompany drought in nature and are likely to have had an additive effect [50] on the stress response seen in Trial 1. However, statistical comparisons between Trials 1 and 2 were not made because of the many variables that would complicate interpretation, e.g., different plants, seasonal factors, temperature differences.
CYP707A1/A2 genes were significantly upregulated by drought, especially in CG202 vs M9 roots. The CYP707A1/A2 enzyme groups are key ABA catabolic genes encoding ABA 8’-hydroxylases that convert biologically active ABA to an unstable intermediate 8’-hydroxy-ABA that is cyclized to the inert PA metabolite [51]. Endogenous concentrations of ABA are thought to be controlled by a balance between biosynthesis versus catabolism, transport to different parts of the plant, and cycling between inert glycosylated storage forms (ABA-glucose ester) and more biologically active aglycone pools [10]. However, the role of CYP707A family is so crucial to regulating internal ABA concentrations that mutants deficient in CYP707A activity accumulate more ABA than lines overexpressing ABA biosynthesis enzymes [10]. The different CYP707A enzyme groups are known to have different spatial and temporal patterns of expression that reflect slightly different physiological roles [10]. For instance, CYP707A1 is most important for ABA catabolism midway during seed development and is localized within the embryo whilst CYP707A2 regulates ABA levels during late stages of seed maturation/germination and is found in both the embryo and endosperm in Arabidopsis [52]. The transcript levels of all CYP707A groups were induced by dehydration in Arabidopsis, with CYP707A1 shown to play an important role in regulating ABA pools in the stomatal guard cells [53]. In the current study, tissue-differentiated gene expression patterns were also observed for the different CYP707A groups and isoforms within those groups. Basal expression levels of CYP707A1_ii and CYP707A2_i were up to 240-fold more abundant in leaves than roots and remained high in leaves throughout the experiment but increased significantly in roots in the Dry treatment. The reverse was true for CYP707A1_i, where baseline levels were up to 120-fold greater in roots than leaves, and transcript numbers decreased by up to 50% in roots in the Dry treatment. These observations may suggest a more fundamental role for CYP707A1_ii and CYP707A2_i in regulating ABA in the leaf guard cells, whereas reduced abundance of CYP707A1_i in the roots in the Dry treatment would lead to increased ABA concentrations, possibly promoting adventitious root formulation.
All isoforms in the NCED3 family were also significantly upregulated in the Dry treatment in roots, and NCED3_iii in leaves, with greater upregulation in CG202 than in M9. The NCED3 family is the key regulator of ABA synthesis in Arabidopsis, with increased ABA synthesis being a pivotal response to drought stress [11,12,13]. Upregulation of ABA synthesis was greater in the roots rather than leaves of both rootstocks, but especially in GC202, a result mirrored in a drought study in kiwifruit [54]. Although ABA is thought to be predominantly synthesised in the leaves where stomata are located, synthesis in the roots does occur [55]. Moreover, ABA synthesis in one plant part and transport to another is a well-established phenomenon, with Manzi et al. [56] showing that hormonal transport from aerial organs contributed to sustained to ABA accumulation in long-term drought-stressed tomato roots, and Hu et al. [55] data supporting transport of ABA in the reverse direction. In addition, Hu et al. [55] showed that the initial site of water stress governs the pattern of ABA synthesis, with synthesis occurring first in the roots when root tissue is directly stressed and vice versa when water stress was imposed directly on peanut leaves. In the current study, roots are the first part of the plant to sense the water shortages and therefore could be expected to respond strongly/rapidly in terms of ABA synthesis, with a stronger induced response in CG202 than M9. Further spatial expression profiles underlying ABA synthesis/catabolism in different tissue types could be an interesting avenue for further study. Overall, the significant increase in both NCED3-mediated ABA biosynthesis and CYP707A catabolysis suggests that drought appears to be stimulating ABA metabolism as a whole, with a stronger transcriptomic response occurring in CG202 (especially in the roots) than in M9.
Response to Dehydration 29B, isoform i (RD29B_i) was upregulated in both trials of the current study, with the strongest responses occurring in CG202 roots. The RD29B family are ABA-dependent genes involved in downstream parts of the ABA signaling cascade that have been shown to be upregulated in response to both drought and to application of an effective drought priming agent (a plant growth promoting rhizobacterium) in Arabidopsis [57]. The RD29B phenology tree in apple consists of two distinct branches, both of which were quite distant from RD29B in Arabidopsis, indicating evolutionary divergence. Only one of the family members, RD29B_i was significantly upregulated in response to drought in the current study and mainly in roots, which possibly points to the different tissue-specific roles played by members of the same gene family, with a strong drought-induced response in root tissue.
bHLH130, a TF, is thought to be involved in stomatal regulation, with bHLH TFs identified as contributing to improved drought tolerance in transgenic apple calli [3]. Dehydration-induced bHLH130 has been shown to regulate stomatal closure, increased the expression of ROS-scavenging and stress-related genes in tobacco leading to increased tolerance to drought [58] and belongs to the same clade as ABA-Responsive Kinase Substrate 1 (AKS1) from Arabidopsis. Takahashi et al. [59] showed that AKS1 is modulated by ABA which causes the inhibition of KAT1 expression by releasing AKS1 from interaction with the KAT1 protein, thereby limiting stomatal opening. The basis of this release is the monomerization of AKS1 through phosphorylation by ABA-responsive SnRK2 kinases. This stomatal response mediated by AKS1 is likely a late process and may, for example, reduce the reopening of stomata after severe drought episodes. In the current study, a significant upregulation of MdbHLH130 of more than 2-fold was observed only in roots of GC202, and in M9 to a lesser extent, which does not fit with the leaf stomatal response theory. However, baseline abundance of MdbHLH130 was up to 2.8-fold greater in leaves than in roots, suggesting that its main role in stomatal closure regulation in leaves is a basal rather than an induced response. This gene marker was only introduced into the second gene set used to probe Trial 2, so further trial data are required to verify this result.
Sorbitol dehydrogenase (SDH1) is involved in sugar metabolism converting sorbitol to fructose [60]. This marker was newly introduced in Trial 2 and was the most abundantly expressed of all the genes in that trial, with the greatest expression occurring in GG202 roots and leaves (basal levels of around 10,000 copies increased to around 30,000 copies with drought). High abundance of SDH1 in root and leaf tissues is consistent with the central role of sugar metabolism in plant growth and source/sink relationships. Potential dual roles of this enzyme in the drought response could include modulating cellular osmotic adaptation, which is a common response in water deficiency, and/or the fructose could act as an emergency store of energy that can be utilised after an initial increase in sorbitol to adapt to the water stress. Emergency energy stores become particularly important once photosynthesis is limited by reduced external gaseous exchange associated with stomatal closure.
The PYL4 family acted as a negative marker of drought response, with significant downregulation occurring with drought in both rootstocks and tissue types, but with a more extreme and rapid response occurring in CG202 than M9, especially in root tissue. A particular class of protein phosphatases (PP2Cs of the A clade) inhibit a key node of the ABA response by binding SnRK2 kinases, preventing these kinases from activating downstream ABA signaling responses [10]. These PP2Cs are kept quiescent and effectively sequestered by the PYR/PYL ABA receptor family in the presence of ABA. There are different classes of PYR/PYL ABA receptor proteins which include dimeric and monomeric classes as well as classes with different affinities to ABA and different expression profiles. The dimeric PYL classes (e.g., PYR1/PYL1-2) show lower affinity/higher dissociation constants for ABA (>50 µM) than monomeric PYLs (e.g., PYL4, PYL9, ~1 µM) but in the presence of their matching PP2C-A clade protein partners they form ternary (PP2C sequestering) complexes with much higher ABA affinities in the 30-60 nM range [61,62]. Dimeric PYR/PYLs are compromised in the surface that interacts with their PP2Cs partners and are therefore highly dependent upon ABA to adopt a PP2C binding/sequestering conformations. Monomeric receptors are able to interact to some degree with PP2Cs even in the absence of ABA. Monomeric PYL4 shows high expression across multiple tissues and therefore probably plays a critical role in sequestering PP2CAs [63]. In support of this theory, the four members of the PYL4 family tested in this study also showed high basal expression in roots (up to 5000 copies) and leaves (up to 3000 copies) of both rootstocks. In maize, PYL4-like genes showed differential expression patterns in leaves with some members of the PYL4 family showing decreases in response to ABA while others showed increases [64] whilst in roots, PYL4 showed dose-dependent ABA down-regulation. PYL4 is rapidly downregulated in response to drought in Arabidopsis [49], and drought-induced downregulation of the PYL4 family was also observed in roots and leaves of another dicot, kiwifruit, with the degree of the response being more extreme in roots than leaves [54]. Interestingly the PYL9 family is thought to play an important role in regulating lateral root growth, and coordinated signals sent through PYL9 are an important component of drought recovery [65], with over-expression of PYL9 also shown to provide tolerance to drought in both Arabidopsis and rice [66]. However, the PYL9 genes tested in Trial 1 did not show any significant differential response to drought and so were excluded from Trial 2. One possible explanation for this anomaly is that the complete family of PYL9-like genes in apple was not included in this study, and rather representative members chosen from each of the distinct branches in the phenological tree were used. It is therefore possible that we did not select the most responsive members of this gene family.
Four main classes of phytohormones and their metabolites were quantified in this study, and their compositions were affected by tissue type, rootstock genotype and drought treatment, with consistent trends across both trials. In both genotypes, ABA and SA-derivatives accumulated in leaves, while higher concentrations of jasmonates and IAA were quantified from roots. While these phytohormones can be produced in multiple cell types and organs, they are predominantly synthesized in chloroplasts (SA, JA) or leaf cells (ABA, IAA) and transported where needed [67]. Root tips also synthesise IAA as key regulator for root development in crosstalk with ethylene and jasmonates. While the most bioactive jasmonate form, JA-Ile, is mainly known for its roles in plant defenses against biotic stresses, it also promotes lateral root formation by inhibiting primary and adventitious roots in crosstalk with auxin [68]. In this study young/developing root tissue was sampled in contrast to mature leaves, which may explain the elevated concentrations of IAA and JA-Ile in actively growing root samples. The genotype comparison showed that CG202 produced constitutively higher concentrations of abscisates and jasmonates in both roots and leaves, while M9 had higher SA content, especially in leaves. A recent study highlighted increased pest-resistance of CG202 [69], which is likely instigated by higher basal levels of JA-Ile and demonstrates the multifunctional role of this phytohormone. SA is also commonly associated with its role in mitigating biotic stresses, however, it is also known to induce stomata closure and consequently reduce photosynthetic activity in a variety of crop plants [70]. Higher basal concentrations of SA might compensate for lower ABA in regulating stomata movement in M9 when compared to CG202.
ABA is the primary phytohormone associated with drought stress and stomata closure [10] in plants and in this study its concentration increased rapidly in roots and after 9-days in leaves of both rootstock genotypes in response to water limitation. This was also observed for its catabolite PA, which forms spontaneously from 8’-hydroxy-ABA (8-OHABA), and might exhibit some residual physiological activity to extend ABA-regulated functions [71]. Both phytohormones were recognised as positive chemical markers for drought responses in roots and leaves. Cytochrome P450 monoxidases catalyse the hydroxylation of ABA to 8-OHABA and are encoded by the CYP707A gene family. CYP707A2 was identified as its most drought-responsive member in this study, with transcript levels magnitudes higher in CG202 than M9. Together with increased expression of the ABA biosynthesis gene NCED3_iii in droughted CG202, these findings indicate that ABA metabolism is genetically upregulated in this rootstock genotype.
In roots only, IAA was highlighted as positive chemical marker for drought stress as it increased significantly after 14-days without irrigation in both genotypes. This aligns with observations in tobacco, where water deficit induced IAA accumulation in roots but not in leaves [72]. Moreover, drought-induced IAA was shown to increase the formation of first and second order lateral roots, as a likely strategy to increase water uptake and a similar mechanism is possible for apple rootstocks.
In contrast, cis-OPDA was identified as a negative chemical marker to water limitation. While concentrations were higher in roots, they decreased in response to extended drought in both leaves and roots of both genotypes. This drought-induced downregulation of cis-OPDA in apple rootstocks is difficult to explain and requires further investigation, as it contrasts with observations from studies using annual plants, which commonly report an upregulation of this oxylipin in response to stress [73]. While cis-OPDA is the biosynthetic precursor of JA, it is known act as a signaling molecule independent of JA responses. Besides acting as transcriptional activator for stress-related genes [74], it was shown to accumulate in guard cells and partake in regulating stomatal closure [75], thus mitigating drought stress.
In recent years, there has been increasing interest in the use of phytohormones as priming agents to mitigate negative effects of drought in plants [76]. ASM (a functional analogue of SA) was an excellent candidate to assess for potential drought priming in our apple system because SA has been previously shown to ameliorate the negative effects of drought through improving the photosynthetic performance, stimulating SA-mediated defence responses and enhancing the activity of antioxidant enzymes [26,77]. ASM application also reduced transpirational water loss through inducing stomatal closure in the monocot, creeping bentgrass [30,78]. However, ASM application did not alter any of the physiological factors measured in the current study, in contrast to results shown in watermelon, rice, wheat, barley and tomatoes [25,27,29]. Differences between the current study and ours might be due to rate and frequency of ASM application, and/or timing of biomarker measurement relative to ASM application. However, one lasting response post-application was genotype-specific and resulted in increased MeJA concentrations in M9 roots in both Irrigated and Dry plants. As a precursor to JA-Ile production, MeJA might promote lateral root formation in ASM-treated M9 rootstocks but whether it mitigates drought-stress responses is unlikely because no other physiological, chemical or genetic markers were altered in response to its application
This study identified candidate phytohormonal and transcriptomic ‘drought’ biomarkers that correlated either positively or negatively with physiological responses to drought in M9 and CG202 rootstock plants. The transcriptomic biomarkers encompassed a wide range of functional processes associated with the main stress-response hormone, ABA, including biosynthesis (NCED3), catabolism (CYP707A1/2), and the signaling response (PYL4, RD29B). The biochemical markers also represent three phytohormone pathways associated with abiotic stress responses, i.e., ABA, PA (ABA pathway); cisOPDA (JA pathway); and IAA (Auxins). The development of robust drought biomarkers is essential to facilitate the development of stress mitigation strategies that rely on increased plant tolerance to drought. This applies both to breeding programmes and to selection of chemical and biological agents that may prime or condition plants for greater drought resilience, with the latter enabling protection in existing plantings. The characterisation of responses to moderate and severe drought across different apple rootstock genotypes can aid identification of genetic factors that are critical for drought tolerance. This would include integration between plant physiological, transcriptomic and phytohormonal data and environmental modelling to enable the selection of genotypes more suited to the different drought scenarios.
4. Materials and Methods
4.1. Plant Material and Treatment Application
4.1.1. Trial 1
A set of 55 bare rooted apple rootstocks each for M9 and CG202 were potted into 10 L plastic pots containing Daltons™ GB mix (Daltons, Matamata, New Zealand) and placed on a concrete floor, in a polytunnel on dripper lines. For each pot, there were two drippers that delivered a total volume of 1.8 L water per day (900 mL delivered over a 30 min period at 4 am and 4 pm). Following budburst, each plant was reduced to two shoots.
This trial started on 9 February 2022 (day 0), 10 weeks after potting. There were two treatments; ‘Irrigated’ where the pot continued to be watered and ‘Dry’ where the two drippers were removed. A Hydrosense™ soil moisture probe (Campbell Scientific Australia) found that the moisture content of 35-40 % in an Irrigated pot dropped to 3-5 % by day 14 after removal of the drippers.
There were five replicate pots for the baseline (day 0) and for each of the four sampling time points (day 2, 5, 9 and 14) at which the plant was destructively sampled for leaf and root material. The pots were arranged in a randomised design with the treatments and the two rootstock genotypes.
For the Dry treatment, the drippers were removed on day 0 from 25 pots and each pot was placed on an upturned tray to avoid accidental watering from any ground water leaking from neighbouring pots.
4.1.2. Trial 2
A set of 90 bare-rooted apple rootstocks each for M9 and CG202 were potted and placed in a polytunnel on dripper lines as for the previous trial and the pots were arranged in a randomised design.
This trial started on 16 November 2022, 10 weeks from potting. The plants were divided into two treatment groups; 1) untreated control and 2) Actigard®, a.i. acibenzolar-S-methyl (ASM)-treated (purchased from Syngenta NZ). ASM was applied at 10 mg a.i./plant (100 mL of a 0.2 g/ L solution of Actigard, which contains 50% a.i.) to the soil around each plant, both at 14 days prior to day 0 and again on day 0 (16 November 2022). Each treatment group was then further subdivided with half receiving normal watering (Irrigated) and the other half with drippers removed (Dry). This created four treatment groups; 1) Irrigated control, 2) Irrigated+ASM, 3) Dry control, and 4) Dry+ASM.
There were five replicate pots for each of three time points (day 5, 9 and 14) at which the plant was destructively sampled for leaf and root material. For the Irrigated and Irrigated+ASM treatments, there was an additional time point for baseline measurements on day 0.
4.2. Gas Exchange Measurements and Water Loss
Five plants for each treatment were used for repeat measurements of pot weight, plant photosynthesis rate, stomatal conductance and transpiration rate over the trial period for both trials. Measurements were carried out on days 0, 2 (Trial 1 only), 5, 9 and 14. Gas exchange measurements were carried out using a CIRAS-3 portable photosynthesis system (PP Systems, USA). Soil water content of the irrigated and dry treatments was measured as loss in weight over time attributed to water loss and presented as percent reduction in soil water content relative to day 0.
All gas exchange measurements were carried out on the same leaves for each date, leaves were selected as the second or third fully expanded leaf below the growing apex. Day 0 measurements were carried out before the dry treatment was applied so consisted of only the irrigated plants. All measurements were carried out between 10.30 am and 12.30pm. Leaves were remeasured after the initial measurements to check for any photoinhibition in the later measurements. A random treatment layout ensured that all measurements were spread across the treatments during the measurement period.
4.3. Tissue Sampling for Phytohormone and Gene Expression Analysis
Leaves and roots were sampled at each of the time points between 10 am and noon. The leaf samples consisted of two fully expanded leaves, one from each shoot collected approximately 15-20 cm from the shoot tip. The midrib was removed before placing in vials and snap frozen in liquid nitrogen. Root tissue was collected after removing the plant from the pot, then washing the roots under running water. Young root tissue (approximately 1 g, located up to 5 cm from the growing tip was removed, then blotted dry and snap frozen. For each of the two apple rootstock cultivars in Trial 1, there were 45 leaf and 45 root samples, making a total of 180 samples each for phytohormone and gene expression analyses. Samples were stored at -80°C until processing. In Trial 2, there were 70 leaf and 70 root samples, making a total of 280 samples for phytohormone and gene expression analyses. Due to much greater sample numbers, only three of the five replicates were processed for gene expression in Trial 2.
For processing, tissue samples were ground into a fine powder using a mortar and pestle with liquid nitrogen. Subsamples of ground tissue were weighed for RNA extraction and for freeze drying for phytohormone analyses. Samples were freeze dried for Trial 1 to normalise for changes in tissue weight that could potentially occur due to drought-associated water loss, but fresh weight samples were used in Trial 2.
4.4. RNA Extraction
Total RNA was extracted from 100 mg of frozen, ground tissue using the Spectrum Plant Total RNA kit (Merck, Auckland, New Zealand) according to the manufacturer’s instructions. RNA samples were quantified (with ≥ 20 ng/µL considered acceptable), and purity assessed (with a 260/280 absorbance ratio of ~ 2.0 accepted as “pure”) using a NanoDrop 2000c spectrophotometer (Thermo Scientific, Waltham, MA, USA).
4.5. Gene Selection
Candidate genes were mined from a list of genes identified from both apple and model plant literature on ABA metabolic synthesis and modification pathways, plant responses to drought and functional analysis of genes involved in drought resistance (see details in Supplementary Table 1). Two sequentially selected gene sets (one for each trial) consisting of the same six reference genes (RG) that were chosen based upon stability of expression [79,80], and 42 genes of interest (GoI) that were selected by a process of prioritisation (similar to that outlined by Wurms et al. [54]) based on the strength of evidence of their involvement in drought responses in plants. Selection of GoI were from a broad cross-section of the responses including both upregulated and downregulated components. In cases of larger gene families with multiple members, candidates were selected to represent distinct branches in the family phylogenetic tree. The selected genes used in both sets are given in Supplementary Table 1, where the key literature supporting the gene candidacy is also identified for each chosen gene. The genes in set 2 used in Trial 2 were an iteration of the genes in set 1, with non-responding genes from gene set 1 used in Trial 1 replaced by new candidates mined from the literature (Table S1).
4.6. Measurement of Gene Expression by PlexSet Nanostring
Measurement of the direct counts of genes being expressed was carried out on a NanoString nCounter® platform using PlexSet® chemistry, following the steps of probe hybridisation, titration to determine optimal RNA input, sample immobilization onto a cartridge, and digital counting as described by Wurms et al. [54]. The only differences were that RNA input was optimised at 280 ng per sample in Trial 1 and 320 ng in Trial 2. Results were normalised against the in-situ system positive controls and the most stably expressed RGs, which comprised CKB4, FYPP3 and GPAT1 in Trial 1, and CKB4, FYPP3, GPAT1, LTL1 and Protein GRIP in Trial 2.
4.7. Phytohormone Extraction and Analysis
Phytohormones in Trial 1 were extracted and quantified by liquid chromatography mass spectrometry (LCMS) on a 5500 QTrap triple quadrupole/linear ion trap (QqLIT) mass spectrometer equipped with a Turbo V™ ion source ESI probe (AB Sciex, Concord, ON, Canada) coupled to a Shimadzu Exion UHPLC (Shimadzu, Tokyo, Japan) as described in Wurms et al. [54], with modifications to the sample weight extracted (Trial 1, 100 mg dry weight, DM).
For Trial 2, the methodology was modified to expand the scope of the targeted analytes to include neophaseic acid (neoPA) and to expand the scope of the labelled internal standards added at extraction ([2H3]-DPA 5 ng, [2H3]-PA 2 ng, [2H3]-neoPA 2 ng, [2H4]-7-OH-ABA 5 ng, [2H5]-ABA-GE 5 ng). For Trial 2 LCMS experiments were carried out on a 7500 QTrap QqLIT mass spectrometer equipped with a Turbo V™ ion source ESI probe (AB Sciex, Concord, ON, Canada) coupled to a Shimadzu Nexera LC40 UHPLC (Shimadzu, Tokyo, Japan), using material extracted from 100 mg fresh weight, FW.
4.8. Statistical Analysis and Data Visualisation
Phytohormone and gene expression data from Trials 1 and 2 were log-transformed (to stabilise variance) and analysed with multi-factor analysis of variance. The factors in Trial 1 were rootstock genotype, Irrigated/ Dry treatment and sampling day, plus interactions. The factors in Trial 2 were rootstock genotype, Irrigated/ Dry treatment, ASM treatment and day, plus interactions. As the treatments did not apply on day 0, treatment and day were not completely orthogonal, and so effects were tested adjusted for other factors.
The means were summarised as fold changes, either compared to the control treatment on the same rootstock on the same sample day, or between the two rootstocks on the same treatment on the same day. The least significant differences (p = 0.05) from the models corresponded to fold changes of between 1.5 and 3-fold, so a 2-fold change (positive or negative) was taken as the threshold for reporting.
This analysis was done using Genstat, version 22, (VSNi Ltd., UK, 2022).
For the gas exchange data, analysis of variance was carried out separately for the response’s photosynthesis, transpiration and stomatal conductance for the effects of Rootstock (CG202 and M9), assessment time (days), ASM treatment (ASM and none; Trial 2 only) and water treatment (watered vs drought conditions). Pairwise linear contrasts were used to assess significant differences between the means for watered and droughted treatments for each combination of rootstock and assessment day, for α = 0.05 level of significance. Analyses were carried out in Genstat 24th Edition (VSN International 2025).
Phytohormone data were analysed and visualised using R version 4.4.2. A small offset was used for concentrations below the limit of detection (LOD), which was replaced with 0.2* LOD. Data were log10 transformed and standardised by autoscaling (mean-centered and feature scaled) using the ‘mdatools’ package [81]. Heatmaps were computed based on Euclidean distances of the means and clustered using Ward’s method as implemented in the ‘gplots’ package [82]. Redundancy analysis (RDA) was performed using ‘vegan’ [83] to analyse the relationships between phytohormones and rootstock genotype, treatment and sampling day as explanatory variables. The ‘permutest’-function was used to assess the significance of the model with 9999 permutations and the significance of constraints was computed using ANOVA- like permutation testing by ‘term’ (explanatory variables) and ‘axis’ (component) at α=0.05. The proportion of variance explained by each RDA axis was computed based on their eigenvalues and ordination scores ($site) and visualized as RDA sample plots. Data ellipses were computed at a confidence level of 95%. Biplot visualisation was used for graphing the ordination of explanatory variables and response variables ($species) as loading vectors.
Univariate, pairwise comparisons were performed on log10-transformed data using the ‘tukey-HSD’ function with false-discovery rate correction for multiple comparisons at α=0.05 as implemented in ‘rstatix’ [84]. Linegraphs of raw data were prepared using ‘ggplot2’ [85] with the ‘geom-smooth’ function and default confidence level of 95%.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Temperature data and respective sample time points (arrows) in Trial 1 and Trial 2. Tissue samples were collected, and physiology measurements were recorded between 10:00 am and noon on each day. Figure S2: Heat map presenting fold-change gene expression data in A) root, and B) leaf tissue, induced by drought stress conditions in Malus domestica CG202 and M9 rootstocks after 2, 5, 9, and 14 days without water, in Trial 1. Control irrigated plants received a total volume of 1.8 L water/day via drippers. There were five replicate plants/sample time/treatment/rootstock. Gene expression was quantified by PlexSet® NanoString using CKB4, FYPP3 and GPAT1 as reference genes. Genes of interest are listed in the left-hand column. Numeric values give the fold changes relative to irrigated plants, sampled at the same time periods. Red colouration indicates a fold-increase, blue a fold-decrease, and white is no fold change relative to the control. Colour intensity is indicative of the degree of change, but all fold increases greater than 10 have the same red colour intensity. Statistically significant differences from the control plants at each timepoint, as determined by AVOVA (p ≤ 0.05) analysis, are indicated in bold typeface. Figure S3: Heat map presenting fold-change in gene expression data in leaves, induced by drought stress conditions and acibenzolar-S-methyl (ASM) application in Malus domestica GG202 and M9 rootstocks after 5, 9, and 14 days without water, in Trial 2. ASM (10 mg a.i./plant) was applied as a root drench, both 14 days before the start of experiment and again on day 0, immediately before dripper removal. Control irrigated plants received a total volume of 1.8 L water/day. There were three replicate plants/sample time/treatment/rootstock. Gene expression was quantified by PlexSet® NanoString using CKB4, FYPP3, GPAT1, LTL1, and Protein GRIP as reference genes. All fold changes (numeric values in the figure) are relative to irrigated plants at the same time. Red colouration indicates a fold-increase, blue a fold-decrease, and white is no fold change relative to the control. Statistically significant differences from the control at each timepoint, as determined by ANOVA (p ≤ 0.05) analysis, are indicated in bold typeface. Figure S4: Heat map presenting fold-change in gene expression data in leaves, induced by drought stress conditions and acibenzolar-S-methyl (ASM) application in Malus domestica GG202 and M9 rootstocks after 5, 9, and 14 days without water, in Trial 2. ASM (10 mg a.i./plant) was applied as a root drench, both 14 days before the start of experiment and again on day 0, immediately before dripper removal. Control irrigated plants received a total volume of 1.8 L water/day. There were three replicate plants/sample time/treatment/rootstock. Gene expression was quantified by PlexSet® NanoString using CKB4, FYPP3, GPAT1, LTL1, and Protein GRIP as reference genes. All fold changes (numeric values in the figure) are relative to irrigated plants at the same time. Red colouration indicates a fold-increase, blue a fold-decrease, and white is no fold change relative to the control. Statistically significant differences from the control at each timepoint, as determined by ANOVA (p ≤ 0.05) analysis, are indicated in bold typeface. Figure S5: Transcript counts (backtransformed log2 gene counts) as measured by PlexSet® NanoString in roots (A-C) and leaves (D-F) in CG202 and M9 Malus domestica rootstocks after 5, 9 and 14 days without water, in Trial 2. Control irrigated plants received a total volume of 1.8 L water/day via drippers. Acetyl-S-methyl (ASM), at 10 mg a.i./plant, was applied as a root drench, both 14 days before the start of experiment and again on day 0, immediately before dripper removal. There were three replicate plants/sample time/treatment/rootstock used for NanoString. CKB4, FYPP3, GPAT1, LTL1, and Protein GRIP were used as reference genes. The genes presented are A&D) CYP707A1_i; B&E) CYP707A1_ii C&F) CYP707A2_i. Different lettering over bars indicates statistically significant differences, as shown by Fisher’s Least Significant Difference (LSD), p ≤ 0.05, for each gene. Figure S6: Transcript counts (backtransformed log2 gene counts) as measured by PlexSet® NanoString in roots (A-D) and leaves (E-H) in CG202 and M9 Malus domestica rootstocks after 5, 9 and 14 days in irrigated and Dry plants, in Trial 2. Control irrigated plants received a total volume of 1.8 L water/day via drippers. Acibenzolar-S-methyl (ASM), at 10 mg a.i./plant, was applied as a root drench, both 14 days before the start of experiment and again on day 0, immediately before dripper removal. There were three replicate plants/sample time/treatment/rootstock. CKB4, FYPP3, GPAT1, LTL1, and Protein GRIP were used as reference genes. The genes presented are A&E) PYL4_i; B&F) PYL4_ii C&G) PYL4_iii; D&H) PYL4_iv. Different lettering over bars indicates statistically significant differences, as shown by Fisher’s Least Significant Difference (LSD), p ≤ 0.05, for each gene. Figure S7: Transcript counts (backtransformed log2 gene counts) as measured by PlexSet® NanoString in roots (A-C) and leaves (D-F) in CG202 and M9 Malus domestica rootstocks after 0, 2, 5, 9 and 14 days in irrigated and Dry plants, in Trial 1. Control irrigated plants received a total volume of 1.8 L water/day via drippers. There were five replicate plants/sample time/treatment/rootstock. CKB4, FYPP3 and GPAT1 were used as reference genes. The genes presented are A&D) NCED3_i; B&E) NCED3_ii; C&F) NCED3_iii. Different lettering over bars indicates statistically significant differences, as shown by Fisher’s Least Significant Difference (LSD), p ≤ 0.05, for each gene. Figure S8: Transcript counts (backtransformed log2 gene counts) as measured by PlexSet® NanoString in roots (A-C) and leaves (D-F) in CG202 and M9 Malus domestica rootstocks after 0, 5, 9 and 14 days in irrigated and Dry plants, in Trial 2. Control irrigated plants received a total volume of 1.8 L water/day via drippers. Acibenzolar-S-methyl (ASM), at 10 mg a.i./plant, was applied as a root drench, both 14 days before the start of experiment and again on day 0, immediately before dripper removal. There were three replicate plants/sample time/treatment/rootstock. CKB4, FYPP3, GPAT1, LTL1, and Protein GRIP were used as reference genes. The genes presented are A&D) NCED3_i; B&E) NCED3_ii; C&F) NCED3_iii. Different lettering over bars indicates statistically significant differences, as shown by Fisher’s Least Significant Difference (LSD), p ≤ 0.05, for each gene. Figure S9: Transcript counts (backtransformed log2 gene counts) for RD29B_i as measured by PlexSet® NanoString in roots (A&C) and leaves (B&D) in CG202 and M9 Malus domestica rootstocks. Measurements were recorded after 0, 2, 5, 9 and 14 days in irrigated and Dry plants in Trial1 (A&C) and after 5, 9 and 14 days in Trial 2 (B&D). Control irrigated plants received a total volume of 1.8 L water/day via drippers. In Trial 2, acibenzolar-S-methyl (ASM), at 10 mg a.i./plant, was applied as a root drench, at 14 days before the start of experiment and again on day 0, immediately before dripper removal. There were five replicate plants/sample time/treatment/rootstock in Trial 1 and three in Trial 2. CKB4, FYPP3 and GPAT1 were used as reference genes in Trial 1, and CKB4, FYPP3, GPAT1, LTL1, and Protein GRIP in Trial 2. Different lettering over bars indicates statistically significant differences, as shown by Fisher’s Least Significant Difference (LSD), p ≤ 0.05. Figure S10: Clustered Heatmap of phytohormone concentrations common to leaves and roots of two apple rootstock genotypes (CG202, M9) across common sampling timepoints (day 0, 2, 5, 9, 14) in Trial 1. Dry: drought-treated, Control: Irrigated. Data are scaled by row with yellow hues indicating positive and blue hues negative standard (Z-) scores, i.e., relative concentrations. SA: salicylic acid; SAG: salicylic acid glucoside; IAA: indole-3-acetic acid; ABA: abscisic acid; PA: phaseic acid; DPA: dihydrophaseic acid; JA: jasmonic acid; JA-Ile: jasmonic acid-isoleucine; X12.OH.JA: 12-hydroxyjasmonic acid; DHJA: dihydrojasmonic acid; OPC-4: 4-(3-oxo-2-(pent-2-en-1-yl)cyclopentyl)octanoic acid; cis-OPDA: 12-oxo-phytodienoic acid. Figure S11: Redundancy analysis (RDA) sample plots for A) roots and B) leaves from two apple rootstock genotypes (CG202, M9), with genotype, treatment (Dry vs Irrigated) and sampling day (0, 2, 5, 9, 14 days) as response variables, in Trial 1. The asterisks indicate that explained variance is significant by the respective component at *α=0.05, ** α=0.01, *** α=0.001. Figure S12: Clustered Heatmap of phytohormone concentrations common to leaves and roots of two apple rootstock genotypes (CG202, M9) across common sampling timepoints (day 5, 9, 14) in Trial 2. Dry: drought-treated, Control: Irrigated, ASM: Actiguard-treated. Data are scaled by row with purple hues indicating positive and green hues negative standard (Z-) scores, i.e., relative concentrations. SA: salicylic acid; SAG: salicylic acid glucoside; DHB: dihydroxy benzoic acid; IAA: indole-3-acetic acid; ABA: abscisic acid; PA: phaseic acid; DPA: dihydrophaseic acid; ABA.GE: abscisic acid glucoside; NeoPA: Neophaseic acid; JA: jasmonic acid; JA-Ile: jasmonic acid-isoleucine; X12.OH.JA: 12-hydroxyjasmonic acid; DHJA: dihydrojasmonic acid; OPC-4: 4-(3-oxo-2-(pent-2-en-1-yl)cyclopentyl)octanoic acid; cis-OPDA: 12-oxo-phytodienoic acid; MeJA: Methyl Jasmonate. Figure S13: Redundancy analysis (RDA) sample plots for A) roots and B) leaves from two apple rootstock genotypes (CG202, M9), with genotype, treatment (dry vs irrigated) and sampling day (5, 9, 14 days) as response variables, in Trial 2. The asterisks indicate that explained variance is significant by the respective component at *α=0.05, ** α=0.01, *** α=0.001. Figure S14: Heat map presenting fold-change data of metabolite concentrations in A) leaves, and B) roots of Malus domestica GG202 and M9 rootstocks after 0, 2, 5, 9, and 14 days without water. All fold changes are relative to irrigated plants at the same time point. Red colouration indicates a fold-increase, blue a fold-decrease, and white is no change relative to the irrigated control. A 2-fold change cutoff was applied for variable selection and bolded data indicate samples with min 2-fold change difference. DHB: dihydroxy benzoic acid; IAA: indole-3-acetic acid; ABA: abscisic acid; PA: phaseic acid; DPA: dihydrophaseic acid; 7-OH-ABA: 7’-hydroxy-abscisic acid; ABA-GE: abscisic acid glucoside; NeoPA: Neo phaseic acid; JA: jasmonic acid; JA-Ile: jasmonic acid-isoleucine; 12-OH-JA: 12-hydroxyjasmonic acid; DHJA: dihydrojasmonic acid; cis-OPDA: 12-oxo-phytodienoic acid; MeJA: Methyl Jasmonate. Table S1: Identification of reference genes (RGs) and genes of interest (GoI) used for gene expression analysis by PlexSet® NanoString of apple rootstock responses to drought and acibenzolar-S-methyl (ASM) application in this study.
Author Contributions
Conceptualization, T.R., E.H.A.R., N.G. and K.V.W; methodology, T.R., A.A.C., C.B.F., P.B., K.V.W., N.G., J.M.C. and D.J.A.J; validation, C.S.G., and D.H.; formal analysis, D.H.; writing—original draft preparation, K.V.W., T.R., C.S.G; E.H.A.R., N.G., J.M.C., D.H. and A.A.C writing—review and editing, K.V.W., T.R., C.S.G; E.H.A.R., N.G., J.M.C. and D.H.; visualisation T.R., A.A.C., C.S.G., J.M.C., N.G. and K.V.W; project administration, T.R. and E.H.A.R.; funding acquisition, T.R. and E.H.A.R. All authors have read and agreed to the published version of the manuscript.”.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable for studies not involving humans or animals.
Informed Consent Statement
Not applicable for studies not involving humans.
Data Availability Statement
Please refer to suggested Data Availability Statements in section “MDPI Research Data Policies” at https://www.mdpi.com/ethics.
Acknowledgments
We gratefully acknowledge the extra technical contributions from Magan Schipper, Sergio Marshall, Rachelle Anderson and Emma Todd. The authors also wish to thank Catherine McKenzie for carrying out additional statistical analyses, Drs Sean Bulley and Andrew Dare for reviewing this manuscript, and Tony Corbett for producing some of the high-resolution figures.
Conflicts of Interest
All authors were employed by the New Zealand Institute for Plant & Food Research Ltd. All authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest
References
- Ji, X.L.; Li, H.L.; Qiao, Z.W.; Zhang, J.C.; Sun, W.J.; Wang, C.K.; Yang, K.; You, C.X.; Hao, Y.J. The BTB-TAZ protein MdBT2 negatively regulates the drought stress response by interacting with the transcription factor MdNAC143 in apple. Plant Sci. 2020, 301, 10. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Mao, K.; Wang, P.; Wang, Y.; Jia, X.; Huo, L.; Sun, X.; Che, R.; Gong, X.; Ma, F. Overexpression of <i>MdATG8i</i> improves water use efficiency in transgenic apple by modulating photosynthesis, osmotic balance, and autophagic activity under moderate water deficit. Hortic. Res.-England 2021, 8, 15. [Google Scholar] [CrossRef]
- Li, X.; Ma, Z.; Song, Y.; Shen, W.; Yue, Q.; Khan, A.; Tahir, M.; Wang, X.; Malnoy, M.; Ma, F. , et al. Insights into the molecular mechanisms underlying responses of apple trees to abiotic stresses. Hortic. Res.-England 2023, 10, 14. [Google Scholar] [CrossRef]
- Singh, D.; Laxmi, A. Transcriptional regulation of drought response: a tortuous network of transcriptional factors. Front. Plant Sci. 2015, 6, 11. [Google Scholar] [CrossRef]
- Kim, W.; Iizumi, T.; Nishimori, M. Global Patterns of Crop Production Losses Associated with Droughts from 1983 to 2009. J. Appl. Meteorol. Climatol. 2019, 58, 1233–1244. [Google Scholar] [CrossRef]
- Espinoza-Meza, S.; Ortega-Farias, S.; López-Olivari, R.; Araya-Alman, M.; Carrasco-Benavides, M. Response of fruit yield, fruit quality, and water productivity to different irrigation levels for a microsprinkler-irrigated apple orchard (cv. Fuji) growing under Mediterranean conditions. Eur. J. Agron. 2023, 145, 8. [Google Scholar] [CrossRef]
- Suran, P.; Pravcova, G. Drought stress affects productivity and fruit size of new apple cultivars. Acta Horticulturae 2023, 1372. [Google Scholar] [CrossRef]
- Urano, K.; Maruyama, K.; Jikumaru, Y.; Kamiya, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Analysis of plant hormone profiles in response to moderate dehydration stress. Plant J. 2017, 90, 17–36. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Sengupta, S.; Fritschi, F.B.; Azad, R.K.; Nechushtai, R.; Mittler, R. The impact of multifactorial stress combination on plant growth and survival. New Phytol. 2021, 230, 1034–1048. [Google Scholar] [CrossRef]
- Finkelstein, R. Abscisic acid synthesis and response. The Arabidopsis Book 2013, 2013. [Google Scholar] [CrossRef]
- Hewage, K.A.H.; Yang, J.-F.; Wang, D.; Hao, G.-F.; Yang, G.-F.; Zhu, J.-K. Chemical Manipulation of Abscisic Acid Signaling: A New Approach to Abiotic and Biotic Stress Management in Agriculture. Advanced Science 2020, 7, 2001265. [Google Scholar] [CrossRef] [PubMed]
- Todaka, D.; Takahashi, F.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Seo, M.; Marion-Poll, A. Chapter three - ABA-responsive gene expression in response to drought stress: cellular regulation and long-distance signaling. In Advances in Botanical Research, Academic Press: 2019; Vol. 92, pp 83-113.
- Ali, S.; Hayat, K.; Iqbal, A.; Xie, L.N. Implications of abscisic acid in the drought stress tolerance of plants. Agronomy-Basel 2020, 10, 28. [Google Scholar] [CrossRef]
- Shu, K.; Zhou, W.G.; Chen, F.; Luo, X.F.; Yang, W.Y. Abscisic Acid and Gibberellins Antagonistically Mediate Plant Development and Abiotic Stress Responses. Front. Plant Sci. 2018, 9, 8. [Google Scholar] [CrossRef]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress responses. Bmc Plant Biology 2016, 16. [Google Scholar] [CrossRef]
- Conrath, U.; Beckers, G.J.; Langenbach, C.J.; Jaskiewicz, M.R. Priming for enhanced defense. Annual review of phytopathology 2015, 53, 97–119. [Google Scholar] [CrossRef]
- Mauch-Mani, B.; Baccelli, I.; Luna, E.; Flors, V. Defense Priming: An Adaptive Part of Induced Resistance. In Annual Review of Plant Biology, Vol 68, Merchant, S.S., Ed. Annual Reviews: Palo Alto, 2017; Vol. 68, pp 485-512.
- Savvides, A.; Ali, S.; Tester, M.; Fotopoulos, V. Chemical Priming of Plants Against Multiple Abiotic Stresses: Mission Possible? Trends Plant Sci. 2016, 21, 329–340. [Google Scholar] [CrossRef]
- Yang, B.; Fu, P.; Lu, J.; Ma, F.; Sun, X.; Fang, Y. Regulated deficit irrigation: an effective way to solve the shortage of agricultural water for horticulture. Stress Biol. 2022, 2, 19. [Google Scholar] [CrossRef]
- Godoy, F.; Olivos-Hernández, K.; Stange, C.; Handford, M. Abiotic Stress in Crop Species: Improving Tolerance by Applying Plant Metabolites. Plants-Basel 2021, 10, 19. [Google Scholar] [CrossRef]
- Antonic, D.; Milosevic, S.; Cingel, A.; Lojic, M.; Trifunovic-Momcilov, M.; Petric, M.; Subotic, A.; Simonovic, A. Effects of exogenous salicylic acid on Impatiens wallerianaL. grown in vitro under polyethylene glycol-imposed drought. South African Journal of Botany 2016, 105, 226–233. [Google Scholar] [CrossRef]
- Rao, S.R.; Qayyum, A.; Razzaq, A.; Ahmad, M.; Mahmood, I.; Sher, A. Role of foliar application of salicylic acid and l-tryptophan in drought tolerance of maize. Journal of Animal and Plant Sciences 2012, 22, 768–772. [Google Scholar]
- Aremu, A.; Fawole, O.; Makunga, N.; Masondo, N.; Moyo, M.; Buthelezi, N.; Amoo, S.; Spíchal, L.; Dolezal, K. Applications of Cytokinins in Horticultural Fruit Crops: Trends and Future Prospects. Biomolecules 2020, 10, 68. [Google Scholar] [CrossRef] [PubMed]
- Sah, S.; Reddy, K.; Li, J. Abscisic Acid and Abiotic Stress Tolerance in Crop Plants. Front. Plant Sci. 2016, 7, 26. [Google Scholar] [CrossRef]
- Khan, M.; Fatma, M.; Per, T.; Anjum, N.; Khan, N. Salicylic acid-induced abiotic stress tolerance and underlying mechanisms in plants. Front. Plant Sci. 2015, 6, 17. [Google Scholar] [CrossRef]
- Rai, G.; Magotra, I.; Khanday, D.; Choudhary, S.; Bhatt, A.; Gupta, V.; Rai, P.; Kumar, P. Boosting Drought Tolerance in Tomatoes through Stimulatory Action of Salicylic Acid Imparted Antioxidant Defense Mechanisms. Agronomy-Basel 2024, 14, 13. [Google Scholar] [CrossRef]
- Silva, J.; da Silva, G.J.; Bonifácio, A.; Dutra, A.; Prado, R.; Neto, F.; Zuffo, A.; Melo, R.; Pereira, T.; de Sousa, R. Exogenous salicylic acid alleviates water stress in watermelon plants. Ann. Appl. Biol. 2023, 182, 121–130. [Google Scholar] [CrossRef]
- Song, W.; Shao, H.; Zheng, A.; Zhao, L.; Xu, Y. Advances in roles of salicylic acid in plant tolerance responses to biotic and abiotic stresses. Plants 2023, 12. [Google Scholar] [CrossRef]
- Szepesi, A.; Csiszár, J.; Gémes, K.; Horváth, E.; Horváth, F.; Simon, M.; Tari, I. Salicylic acid improves acclimation to salt stress by stimulating abscisic aldehyde oxidase activity and abscisic acid accumulation, and increases Na<SUP>+</SUP> content in leaves without toxicity symptoms in <i>Solanum lycopersicum</i> L. J. Plant Physiol. 2009, 166, 914–925. [Google Scholar] [CrossRef]
- Jespersen, D.; Yu, J.; Huang, B. Metabolic Effects of Acibenzolar-S-Methyl for Improving Heat or Drought Stress in Creeping Bentgrass. Front. Plant Sci. 2017, 8, 15. [Google Scholar] [CrossRef]
- Sakata, N.; Ishiga, T.; Taniguchi, S.; Ishiga, Y. Acibenzolar-S-Methyl Activates Stomatal-Based Defense Systemically in Japanese Radish. Front. Plant Sci. 2020, 11, 9. [Google Scholar] [CrossRef]
- Reglinski, T.; Wurms, K.V.; Vanneste, J.L.; Chee, A.A.; Schipper, M.; Cornish, D.; Yu, J.E.; McAlinden, J.; Hedderley, D. Kiwifruit Resistance to <i>Sclerotinia sclerotiorum</i> and <i>Pseudomonas syringae</i> pv. <i>actinidiae</i> and Defence Induction by Acibenzolar-S-methyl and Methyl Jasmonate Are Cultivar Dependent. Int. J. Mol. Sci. 2023, 24, 22. [Google Scholar] [CrossRef]
- Ebel, R.; Proebsting, E.; Evans, R. Apple tree and fruit responses to early termination of irrigation in a semi-arid environment. Hortscience 2001, 36, 1197–1201. [Google Scholar] [CrossRef]
- Lopez, G.; Boini, A.; Manfrini, L.; Torres-Ruiz, J.; Pierpaoli, E.; Zibordi, M.; Losciale, P.; Morandi, B.; Corelli-Grappadelli, L. Effect of shading and water stress on light interception, physiology and yield of apple trees. Agric. Water Manage. 2018, 210, 140–148. [Google Scholar] [CrossRef]
- Volz, R. Streamlining the breeding process. Available online: https://www.sciencelearn.org.nz/videos/428-streamlining-the-breeding-process#more (accessed on day month year).
- Fenning, T.; Gershenzon, J. Where will the wood come from? Plantation forests and the role of biotechnology. Trends Biotechnol. 2002, 20, 291–296. [Google Scholar] [CrossRef]
- Jones, H. How do rootstocks control shoot water relations? New Phytol. 2012, 194, 301–303. [Google Scholar] [CrossRef]
- Valverdi, N.; Kalcsits, L. Apple rootstock genotype affects scion responses to water limitations under field conditions. Acta Physiol. Plant. 2021, 43, 14. [Google Scholar] [CrossRef]
- Tworkoski, T.; Fazio, G.; Glenn, D. Apple rootstock resistance to drought. Sci. Hortic. 2016, 204, 70–78. [Google Scholar] [CrossRef]
- Xu, H.; Ediger, D. Rootstocks with Different Vigor Influenced Scion-Water Relations and Stress Responses in Ambrosia<SUP>TM</SUP> Apple Trees (<i>Malus Domestica</i> var. Ambrosia). Plants-Basel 2021, 10, 19. [Google Scholar] [CrossRef]
- Choi, B.; Bhusal, N.; Jeong, W.; Park, I.; Han, S.; Yoon, T. Drought Tolerance of ‘Fuji’ Apple Trees Grafted onto G, CG, or M Series Rootstocks: Growth and Physiology. Hortic. Sci. Technol. 2020, 38, 583–594. [Google Scholar] [CrossRef]
- Shi, C.; Liu, L.; Li, Q.; Wei, Z.; Gao, D. Comparison of drought resistance of rootstocks ‘M9-T337’ and ‘M26’ grafted with ‘Huashuo’ apple. Hortic. Environ. Biotechnol. 2022, 63, 299–310. [Google Scholar] [CrossRef]
- Sun, P.; Tahir, M.; Lu, X.; Liu, Z.; Zhang, X.; Zuo, X.; Shao, Y.; Xiao, X.; An, N.; Wang, C. , et al. Comparison of leaf morphological, anatomical, and photosynthetic responses to drought stress among eight apple rootstocks. Fruit Res. 2022, 2, 13. [Google Scholar] [CrossRef]
- Wright, D.; Cline, J.; Earl, H. Physiological responses of four apple (<i>Malus</i> x <i>domestica</i> Borkh.) rootstock genotypes to soil water deficits. Can. J. Plant Sci. 2019, 99, 510–524. [Google Scholar] [CrossRef]
- Chen, J.; Liu, Z.; Mao, J.; Zhao, T.; Tu, T.; Cheng, L.; Dong, C. Co-regulation of water and energy in the spatial heterogeneity of drought resistance and resilience. Environmental Research Letters 2023, 18. [Google Scholar] [CrossRef]
- Farquhar, G.; Sharkey, T. Stomatal conductance and photosynthesis. Annual Review of Plant Physiology and Plant Molecular Biology 1982, 33, 317–345. [Google Scholar] [CrossRef]
- Liang, X.; Wang, D.; Ye, Q.; Zhang, J.; Liu, M.; Liu, H.; Yu, K.; Wang, Y.; Hou, E.; Zhong, B. , et al. Stomatal responses of terrestrial plants to global change. Nature Communications 2023, 14. [Google Scholar] [CrossRef]
- Fernandez, R.; Perry, R.; Flore, J. Drought response of young apple trees on three rootstocks: Growth and development. J. Am. Soc. Hortic. Sci. 1997, 122, 14–19. [Google Scholar] [CrossRef]
- Goda, H.; Sasaki, E.; Akiyama, K.; Maruyama-Nakashita, A.; Nakabayashi, K.; Li, W.; Ogawa, M.; Yamauchi, Y.; Preston, J.; Aoki, K. , et al. The AtGenExpress hormone and chemical treatment data set: experimental design, data evaluation, model data analysis and data access. Plant J. 2008, 55, 526–542. [Google Scholar] [CrossRef]
- Prasad, P.V.V.; Pisipati, S.R.; Momcilovic, I.; Ristic, Z. Independent and Combined Effects of High Temperature and Drought Stress During Grain Filling on Plant Yield and Chloroplast EF-Tu Expression in Spring Wheat. Journal of Agronomy and Crop Science 2011, 197, 430–441. [Google Scholar] [CrossRef]
- Marion-Poll, A.; Leung, J. Abscisic acid synthesis, metabolism and signal transduction. Annual Plant Reviews Online 2018, 24. [Google Scholar] [CrossRef]
- Okamoto, M.; Kuwahara, A.; Seo, M.; Kushiro, T.; Asami, T.; Hirai, N.; Kamiya, Y.; Koshiba, T.; Nambara, E. CYP707A1 and CYP707A2, which encode abscisic acid 8′-hydroxylases, are indispensable for proper control of seed dormancy and germination in Arabidopsis. Plant Physiology 2006, 141, 97–107. [Google Scholar] [CrossRef]
- Okamoto, M.; Tanaka, Y.; Abrams, S.R.; Kamiya, Y.; Seki, M.; Nambara, E. High Humidity Induces Abscisic Acid 8′-Hydroxylase in Stomata and Vasculature to Regulate Local and Systemic Abscisic Acid Responses in Arabidopsis. Plant Physiology 2009, 149, 825–834. [Google Scholar] [CrossRef]
- Wurms, K.V.; Reglinski, T.; Buissink, P.; Chee, A.A.; Fehlmann, C.; McDonald, S.; Cooney, J.; Jensen, D.; Hedderley, D.; McKenzie, C. , et al. Effects of Drought and Flooding on Phytohormones and Abscisic Acid Gene Expression in Kiwifruit. Int. J. Mol. Sci. 2023, 24. [Google Scholar] [CrossRef]
- Hu, B.; Cao, J.; Ge, K.; Li, L. The site of water stress governs the pattern of ABA synthesis and transport in peanut. Scientific Reports 2016, 6. [Google Scholar] [CrossRef]
- Manzi, M.; Lado, J.; Rodrigo, M.; Zacarías, L.; Arbona, V.; Gómez-Cadenas, A. Root ABA Accumulation in Long-Term Water-Stressed Plants is Sustained by Hormone Transport from Aerial Organs. Plant Cell Physiol. 2015, 56, 2457–2466. [Google Scholar] [CrossRef]
- Liu, W.; Thapa, P.; Park, S.-W. <i>RD29A</i> and<i> RD29B</i> rearrange genetic and epigenetic markers in priming systemic defense responses against drought and salinity. Plant Sci. 2023, 337. [Google Scholar] [CrossRef]
- Zhao, Q.; Fan, Z.; Qiu, L.; Che, Q.; Wang, T.; Li, Y.; Wang, Y. <i>MdbHLH130</i>, an Apple bHLH Transcription Factor, Confers Water Stress Resistance by Regulating Stomatal Closure and ROS Homeostasis in Transgenic Tobacco. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- Takahashi, Y.; Ebisu, Y.; Shimazaki, K.-i. Reconstitution of Abscisic Acid Signaling from the Receptor to DNA via bHLH Transcription Factors. Plant Physiology 2017, 174, 815–822. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, J.; Li, C.; Zhang, Z.; Ma, F.; Li, M. Response of sugar metabolism in apple leaves subjected to short-term drought stress. Plant Physiology and Biochemistry 2019, 141, 164–171. [Google Scholar] [CrossRef]
- Dupeux, F.; Santiago, J.; Betz, K.; Twycross, J.; Park, S.-Y.; Rodriguez, L.; Gonzalez-Guzman, M.; Jensen, M.R.; Krasnogor, N.; Blackledge, M. , et al. A thermodynamic switch modulates abscisic acid receptor sensitivity. Embo Journal 2011, 30, 4171–4184. [Google Scholar] [CrossRef]
- Szostkiewicz, I.; Richter, K.; Kepka, M.; Demmel, S.; Ma, Y.; Korte, A.; Assaad, F.F.; Christmann, A.; Grill, E. Closely related receptor complexes differ in their ABA selectivity and sensitivity. Plant J. 2010, 61, 25–35. [Google Scholar] [CrossRef]
- Pizzio, G.A.; Rodriguez, L.; Antoni, R.; Gonzalez-Guzman, M.; Yunta, C.; Merilo, E.; Kollist, H.; Albert, A.; Rodriguez, P.L. The PYL4 A194T Mutant Uncovers a Key Role of PYR1-LIKE4/PROTEIN PHOSPHATASE 2CA Interaction for Abscisic Acid Signaling and Plant Drought Resistance. Plant Physiology 2013, 163, 441–455. [Google Scholar] [CrossRef]
- Fan, W.; Zhao, M.; Li, S.; Bai, X.; Li, J.; Meng, H.; Mu, Z. Contrasting transcriptional responses of PYR1/PYL/RCAR ABA receptors to ABA or dehydration stress between maize seedling leaves and roots. Bmc Plant Biology 2016, 16. [Google Scholar] [CrossRef] [PubMed]
- Xing, L.; Zhao, Y.; Gao, J.; Xiang, C.; Zhu, J.-K. The ABA receptor PYL9 together with PYL8 plays an important role in regulating lateral root growth. Scientific Reports 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Chan, Z.; Gao, J.; Xing, L.; Cao, M.; Yu, C.; Hu, Y.; You, J.; Shi, H.; Zhu, Y. , et al. ABA receptor PYL9 promotes drought resistance and leaf senescence. Proceedings of the National Academy of Sciences of the United States of America 2016, 113, 1949–1954. [Google Scholar] [CrossRef]
- Bittner, A.; Ciesla, A.; Gruden, K.; Lukan, T.; Mahmud, S.; Teige, M.; Vothknecht, U.C.; Wurzinger, B. Organelles and phytohormones: a network of interactions in plant stress responses. Journal of Experimental Botany 2022, 73, 7165–7181. [Google Scholar] [CrossRef]
- Xu, P.; Zhao, P.X.; Cai, X.T.; Mao, J.L.; Miao, Z.Q.; Xiang, C.B. Integration of Jasmonic Acid and Ethylene Into Auxin Signaling in Root Development. Front Plant Sci 2020, 11, 271. [Google Scholar] [CrossRef]
- Dallabetta, N. Innovative rootstocks for apple crop. Available online: https://apal.org.au/wp-content/uploads/2019/08/Innovative-Rootstocks-for-Apple-crop.pdf (accessed on day month year).
- Akter, F.; Munemasa, S.; Nakamura, T.; Nakamura, Y.; Murata, Y. Negative regulation of salicylic acid-induced stomatal closure by glutathione in Arabidopsis thaliana. Bioscience Biotechnology and Biochemistry 2022, 86, 1378–1382. [Google Scholar] [CrossRef]
- Weng, J.-K.; Ye, M.; Li, B.; Noel, J.P. Co-evolution of Hormone Metabolism and Signaling Networks Expands Plant Adaptive Plasticity. Cell 2016, 166, 881–893. [Google Scholar] [CrossRef]
- Wang, C.; Zhao, Y.; Gu, P.; Zou, F.; Meng, L.; Song, W.; Yang, Y.; Wang, S.; Zhang, Y. Auxin is Involved in Lateral Root Formation Induced by Drought Stress in Tobacco Seedlings. Journal of Plant Growth Regulation 2018, 37, 539–549. [Google Scholar] [CrossRef]
- de Ollas, C.; Dodd, I.C. Physiological impacts of ABA-JA interactions under water-limitation. Plant Molecular Biology 2016, 91, 641–650. [Google Scholar] [CrossRef]
- Boettcher, C.; Pollmann, S. Plant oxylipins: Plant responses to 12-oxo-phytodienoic acid are governed by its specific structural and functional properties. Febs Journal 2009, 276, 4693–4704. [Google Scholar] [CrossRef]
- Savchenko, T.; Dehesh, K. Drought stress modulates oxylipin signature by eliciting 12-OPDA as a potent regulator of stomatal aperture. Plant signaling & behavior 2014, 9, e28304–e28304. [Google Scholar] [CrossRef]
- Iqbal, S.; Wang, X.; Mubeen, I.; Kamran, M.; Kanwal, I.; Diaz, G.A.; Abbas, A.; Parveen, A.; Atiq, M.N.; Alshaya, H. , et al. Phytohormones Trigger Drought Tolerance in Crop Plants: Outlook and Future Perspectives. Front. Plant Sci. 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Khalil, N.; Elhady, S.; Diri, R.; Fekry, M.; Bishr, M.; Salama, O.; El-Zalabani, S. Salicylic Acid Spraying Affects Secondary Metabolites and Radical Scavenging Capacity of Drought-Stressed <i>Eriocephalus africanus</i> L. Agronomy-Basel 2022, 12, 12. [Google Scholar] [CrossRef]
- Shekoofa, A.; Rosas-Anderson, P.; Carley, D.S.; Sinclair, T.R.; Rufty, T.W. Limited transpiration under high vapor pressure deficits of creeping bentgrass by application of Daconil-Action®. Planta 2016, 243, 421–427. [Google Scholar] [CrossRef]
- Bowen, J.; Ireland, H.; Crowhurst, R.; Luo, Z.; Watson, A.; Foster, T.; Gapper, N.; Giovanonni, J.; Mattheis, J.; Watkins, C. , et al. Selection of low-variance expressed <i>Malus x domestica</i> (apple) genes for use as quantitative PCR reference genes (housekeepers). Tree Genet. Genomes 2014, 10, 751–759. [Google Scholar] [CrossRef]
- Zhou, Z.; Cong, P.; Tian, Y.; Zhu, Y. Using RNA-seq data to select reference genes for normalizing gene expression in apple roots. PLoS One 2017, 12, 17. [Google Scholar] [CrossRef]
- Kucheryayskiy, S. mdatools - R package for chemometrics. Chemometrics Intell. Lab. Syst. 2020, 198, 10. [Google Scholar] [CrossRef]
- Warnes, G.R. gplots: Various R programming tools for plotting data. Available online: https://CRAN.R-project.org/package=gplots (accessed on day month year).
- Oksanen, J.; Simpson, G.; Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.; Solymos, P.; Stevens, M.; Szoecs, E., et al. vegan: Community Ecology Package. R package version 2.6-8. Available online: https://CRAN.R-project.org/package=vegan (Accessed on.
- Kassambara, A. rstatix: Pipe-friendly framework for basic statistical tests. R package version 0.7.2. Available online: https://CRAN.R-project.org/package=rstatix (Accessed on: 21/03/2025).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis. . Sringer-Verlag: New York, 2016.
Figure 1.
Transpiration rate (A, B), photosynthesis rate (C, D), and stomatal conductance (E, F), in CG202 and M9 Malus domestica rootstocks after 2 (Trial 1 only), 5, 9 and 14 days without water (Dry). Data for Trial 1 given in A, C, E and from Trial 2 in B, D, F. Irrigated control plants received a total volume of 1.8 L water/day via drippers. There were five replicate plants per sampling time, treatment and rootstock. Mean ± standard error bars from ANOVA shown.
Figure 1.
Transpiration rate (A, B), photosynthesis rate (C, D), and stomatal conductance (E, F), in CG202 and M9 Malus domestica rootstocks after 2 (Trial 1 only), 5, 9 and 14 days without water (Dry). Data for Trial 1 given in A, C, E and from Trial 2 in B, D, F. Irrigated control plants received a total volume of 1.8 L water/day via drippers. There were five replicate plants per sampling time, treatment and rootstock. Mean ± standard error bars from ANOVA shown.
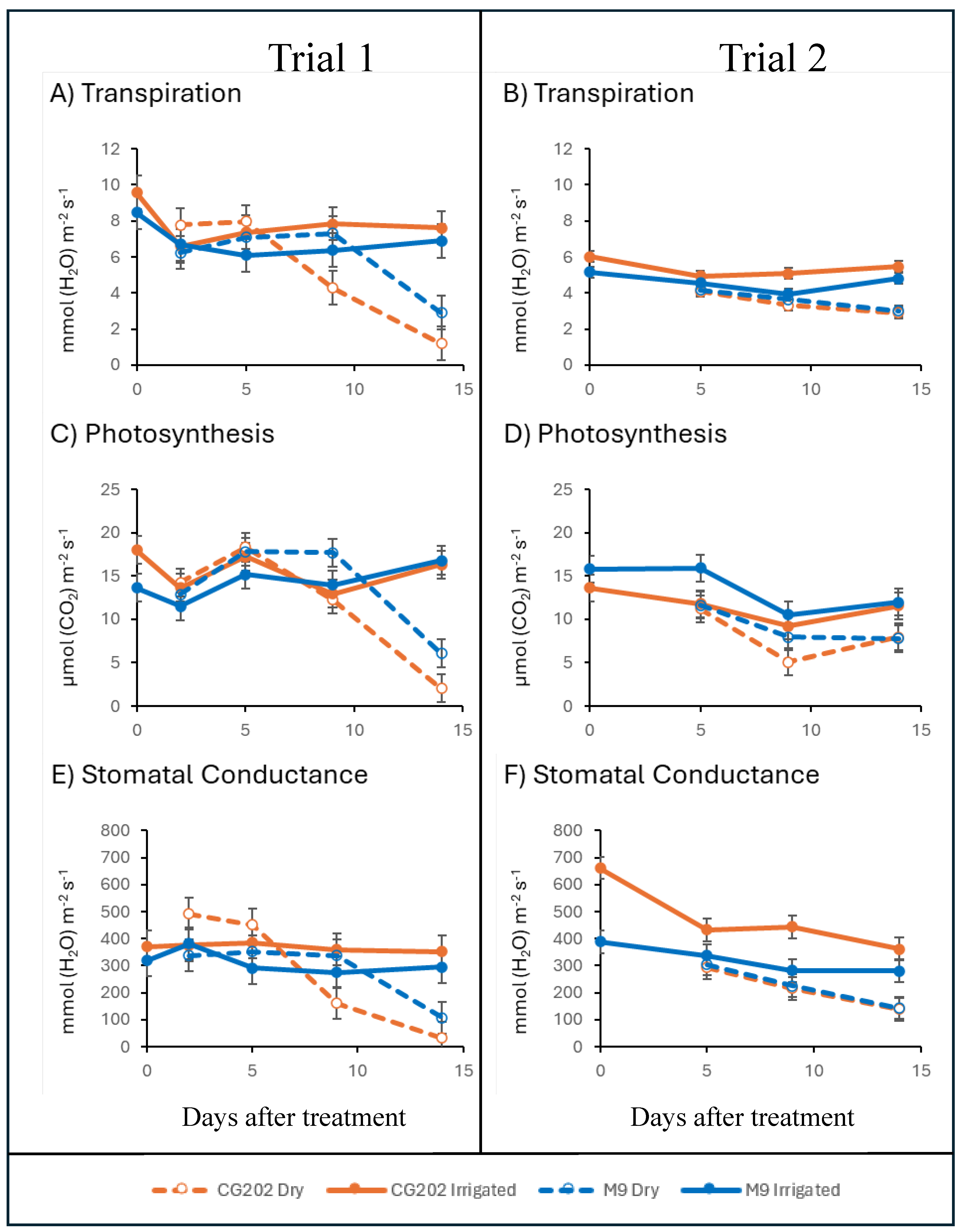
Figure 2.
Heat map presenting fold-change gene expression data in A) root tissue, and B) leaf tissue, induced by drought stress conditions in Malus domestica GG202 and M9 rootstocks after 2, 5, 9, and 14 days without water, in Trial 1. Control irrigated plants received a total volume of 1.8 L water/day via drippers. There were five replicate plants/sample time/treatment/rootstock. Gene expression was quantified by PlexSet® NanoString using CKB4, FYPP3 and GPAT1 as reference genes. Genes of interest are listed in the left-hand column. Numeric values give the fold changes relative to irrigated plants, sampled at the same time periods. Red colouration indicates a fold-increase, blue a fold-decrease, and white is no fold change relative to the control. Colour intensity is indicative of the degree of change, but all fold increases greater than 10 have the same red colour intensity. Statistically significant differences from the control plants at each timepoint, as determined by AVOVA (p ≤ 0.05) analysis, are indicated in bold typeface.
Figure 2.
Heat map presenting fold-change gene expression data in A) root tissue, and B) leaf tissue, induced by drought stress conditions in Malus domestica GG202 and M9 rootstocks after 2, 5, 9, and 14 days without water, in Trial 1. Control irrigated plants received a total volume of 1.8 L water/day via drippers. There were five replicate plants/sample time/treatment/rootstock. Gene expression was quantified by PlexSet® NanoString using CKB4, FYPP3 and GPAT1 as reference genes. Genes of interest are listed in the left-hand column. Numeric values give the fold changes relative to irrigated plants, sampled at the same time periods. Red colouration indicates a fold-increase, blue a fold-decrease, and white is no fold change relative to the control. Colour intensity is indicative of the degree of change, but all fold increases greater than 10 have the same red colour intensity. Statistically significant differences from the control plants at each timepoint, as determined by AVOVA (p ≤ 0.05) analysis, are indicated in bold typeface.
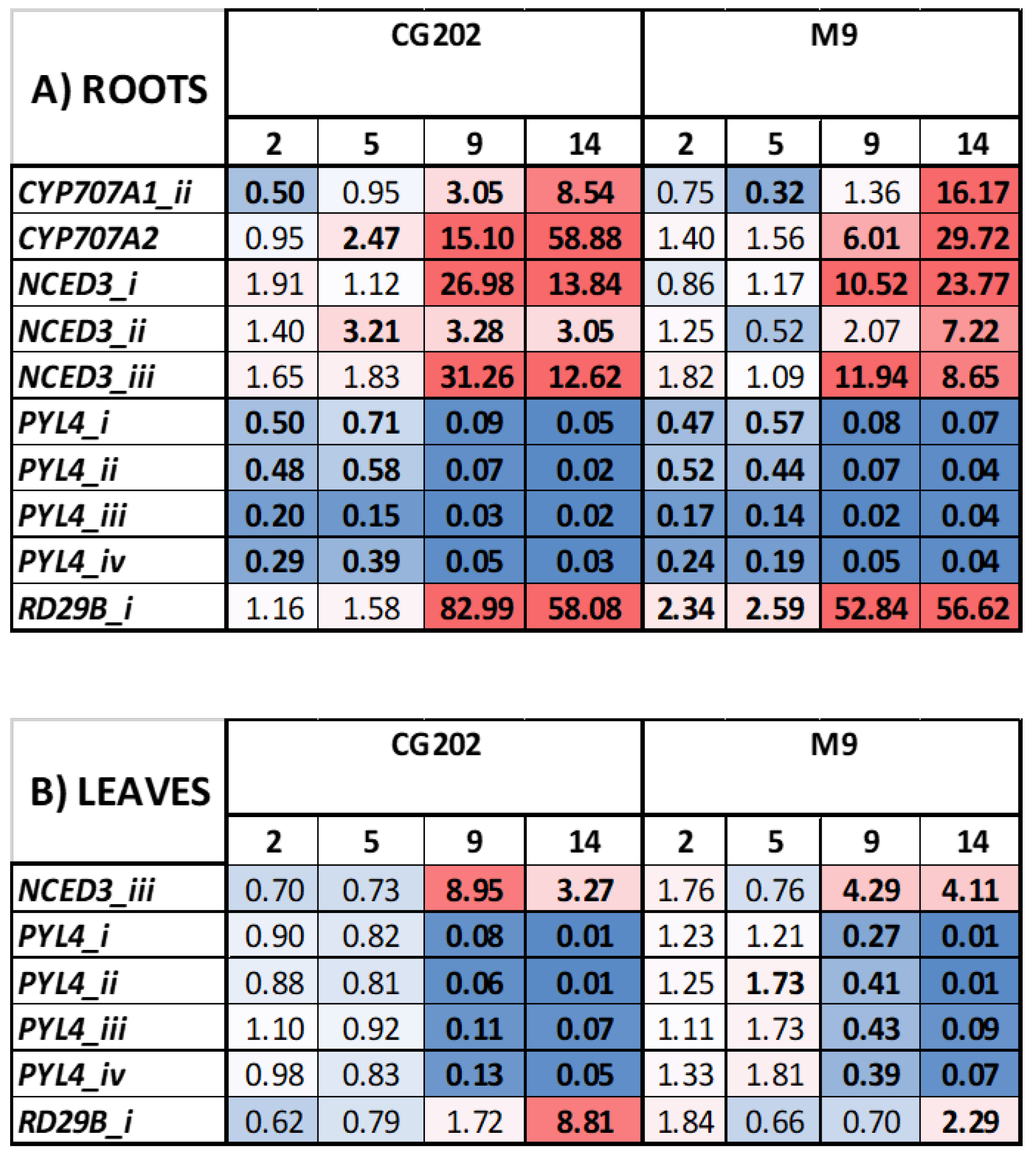
Figure 3.
Heat map presenting fold-change in gene expression data in A) roots, and B) leaves, induced by drought stress conditions and acibenzolar-S-methyl (ASM) application in Malus domestica ‘GG202’ and ‘M9’ rootstocks after 5, 9, and 14 days without water, in Trial 2. ASM (10 mg a.i./plant) was applied as a root drench, both 14 days before the start of experiment and again on day 0, immediately before dripper removal. Control irrigated plants received a total volume of 1.8 L water/day. There were three replicate plants/sample time/treatment/rootstock. Gene expression was quantified by PlexSet® NanoString using CKB4, FYPP3, GPAT1, LTL1, and Protein GRIP as reference genes. All fold changes (numeric values in the figure) are relative to irrigated plants at the same time. Red colouration indicates a fold-increase, blue a fold-decrease, and white is no fold change relative to the control. Statistically significant differences from the control as each timepoint, as determined by AVOVA (p ≤ 0.05) analysis, are indicated in bold typeface.
Figure 3.
Heat map presenting fold-change in gene expression data in A) roots, and B) leaves, induced by drought stress conditions and acibenzolar-S-methyl (ASM) application in Malus domestica ‘GG202’ and ‘M9’ rootstocks after 5, 9, and 14 days without water, in Trial 2. ASM (10 mg a.i./plant) was applied as a root drench, both 14 days before the start of experiment and again on day 0, immediately before dripper removal. Control irrigated plants received a total volume of 1.8 L water/day. There were three replicate plants/sample time/treatment/rootstock. Gene expression was quantified by PlexSet® NanoString using CKB4, FYPP3, GPAT1, LTL1, and Protein GRIP as reference genes. All fold changes (numeric values in the figure) are relative to irrigated plants at the same time. Red colouration indicates a fold-increase, blue a fold-decrease, and white is no fold change relative to the control. Statistically significant differences from the control as each timepoint, as determined by AVOVA (p ≤ 0.05) analysis, are indicated in bold typeface.
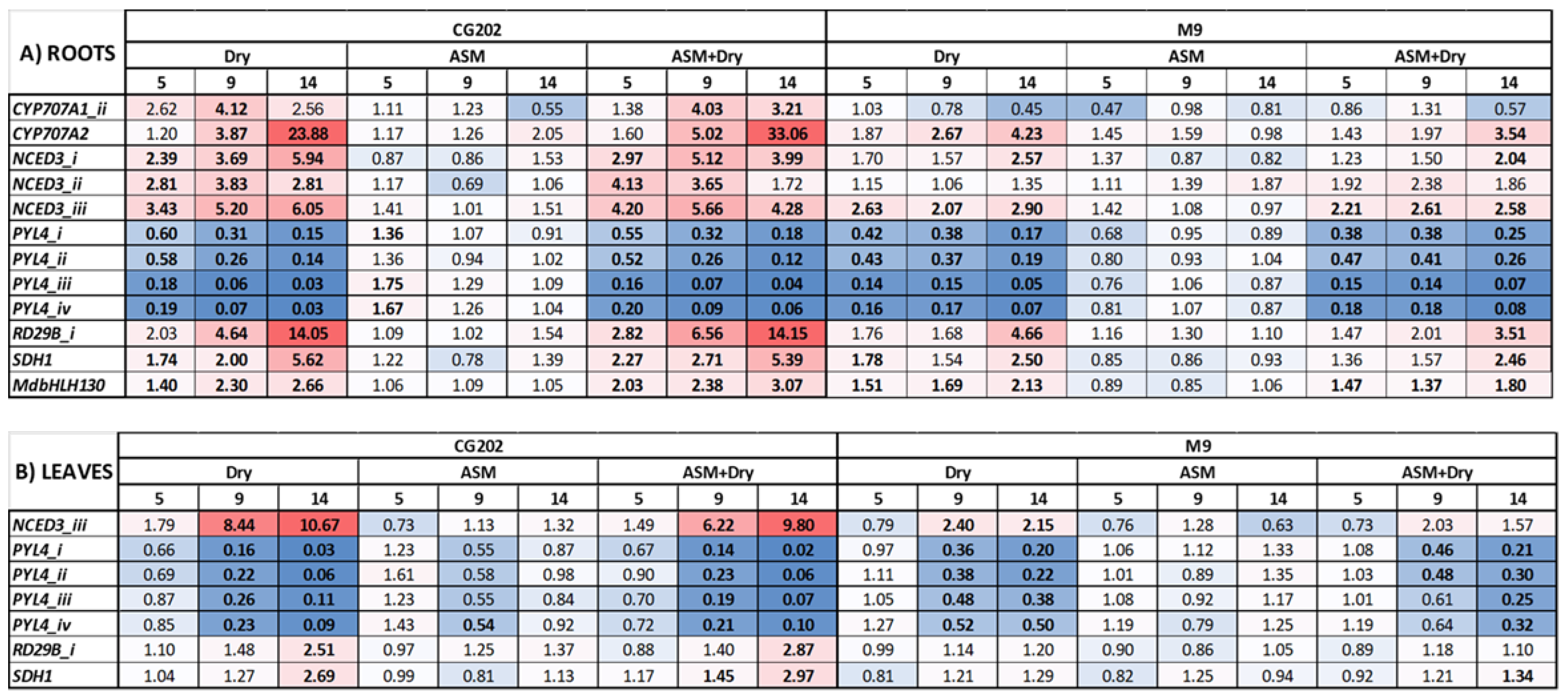
Figure 4.
Transcript counts (backtransformed log2 gene counts) as measured by PlexSet® NanoString in roots (A-C) and leaves (D-E) in CG202 and M9 Malus domestica rootstocks after 2, 5, 9 and 14 days without water, in Trial 1. Control irrigated plants received a total volume of 1.8 L water/day via drippers. Measurements taken on day 0 represent the baseline level of gene expression for plants from both treatment groups. There were five replicate plants/sample time/treatment/rootstock. CKB4, FYPP3 and GPAT1 were used as reference genes. The genes presented are A&D) CYP707A1_i; B&E) CYP707A1_ii C&F) CYP707A2_i. Different lettering over bars indicates statistically significant differences, as shown by Fisher’s Least Significant Difference (LSD), p ≤ 0.05, for each gene.
Figure 4.
Transcript counts (backtransformed log2 gene counts) as measured by PlexSet® NanoString in roots (A-C) and leaves (D-E) in CG202 and M9 Malus domestica rootstocks after 2, 5, 9 and 14 days without water, in Trial 1. Control irrigated plants received a total volume of 1.8 L water/day via drippers. Measurements taken on day 0 represent the baseline level of gene expression for plants from both treatment groups. There were five replicate plants/sample time/treatment/rootstock. CKB4, FYPP3 and GPAT1 were used as reference genes. The genes presented are A&D) CYP707A1_i; B&E) CYP707A1_ii C&F) CYP707A2_i. Different lettering over bars indicates statistically significant differences, as shown by Fisher’s Least Significant Difference (LSD), p ≤ 0.05, for each gene.
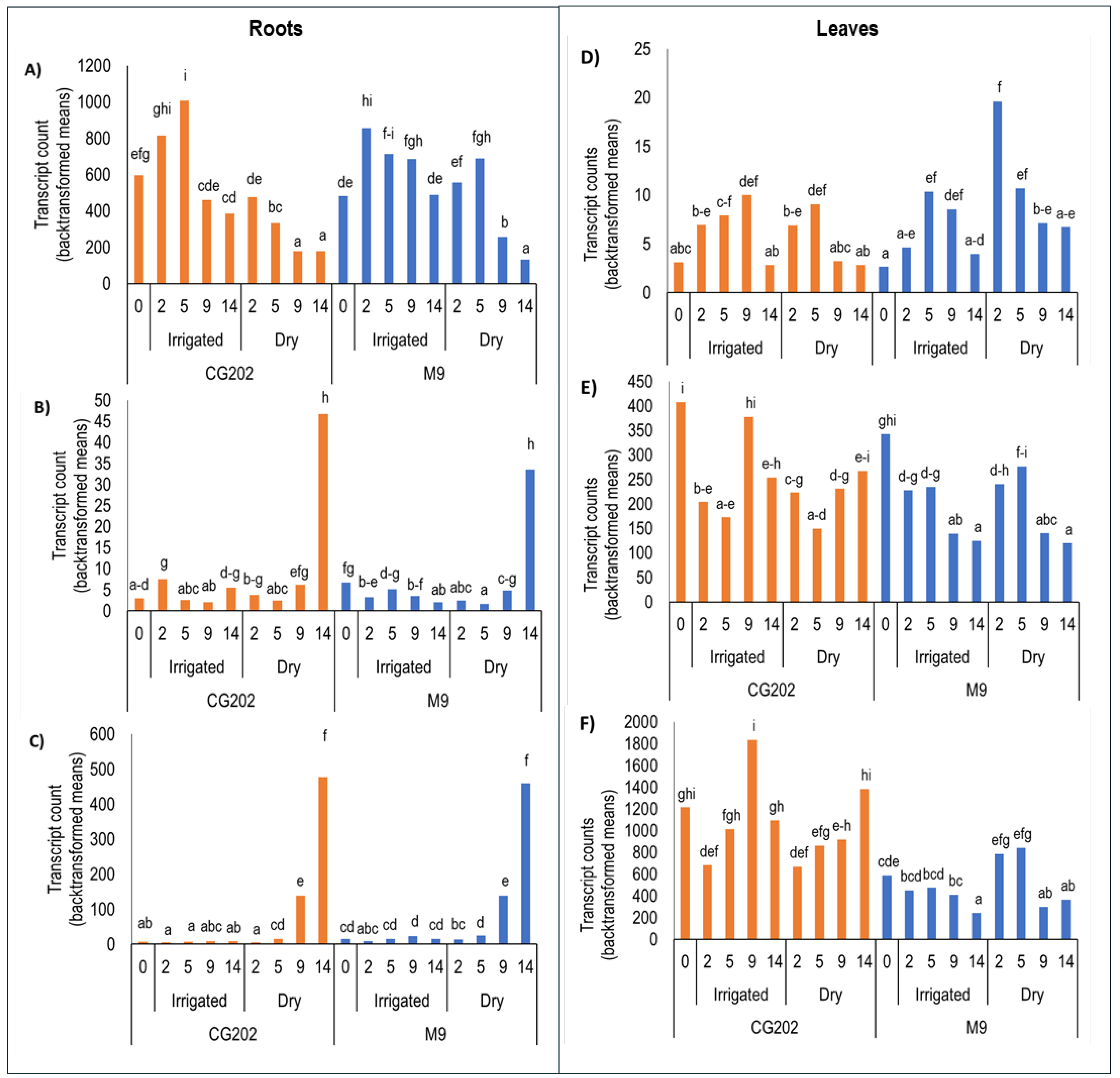
Figure 5.
Transcript counts (backtransformed log2 gene counts) as measured by PlexSet® NanoString in roots (A-D) and leaves (E-H) in CG202 and M9 Malus domestica rootstocks after 2, 5, 9, and 14 days without water, in Trial 1. Control irrigated plants received a total volume of 1.8 L water/day via drippers. Measurements taken on day 0 represent the baseline level of gene expression for plants from both treatment groups. There were five replicate plants/sample time/treatment/rootstock. CKB4, FYPP3 and GPAT1 were used as reference genes. The genes presented are A&E) PYL4_i; B&F) PYL4_ii; C&G) PYL4_iii; and D&H) PYL4_iv. Different lettering over bars indicates statistically significant differences, as shown by Fisher’s Least Significant Difference (LSD), p ≤ 0.05, for each gene.
Figure 5.
Transcript counts (backtransformed log2 gene counts) as measured by PlexSet® NanoString in roots (A-D) and leaves (E-H) in CG202 and M9 Malus domestica rootstocks after 2, 5, 9, and 14 days without water, in Trial 1. Control irrigated plants received a total volume of 1.8 L water/day via drippers. Measurements taken on day 0 represent the baseline level of gene expression for plants from both treatment groups. There were five replicate plants/sample time/treatment/rootstock. CKB4, FYPP3 and GPAT1 were used as reference genes. The genes presented are A&E) PYL4_i; B&F) PYL4_ii; C&G) PYL4_iii; and D&H) PYL4_iv. Different lettering over bars indicates statistically significant differences, as shown by Fisher’s Least Significant Difference (LSD), p ≤ 0.05, for each gene.
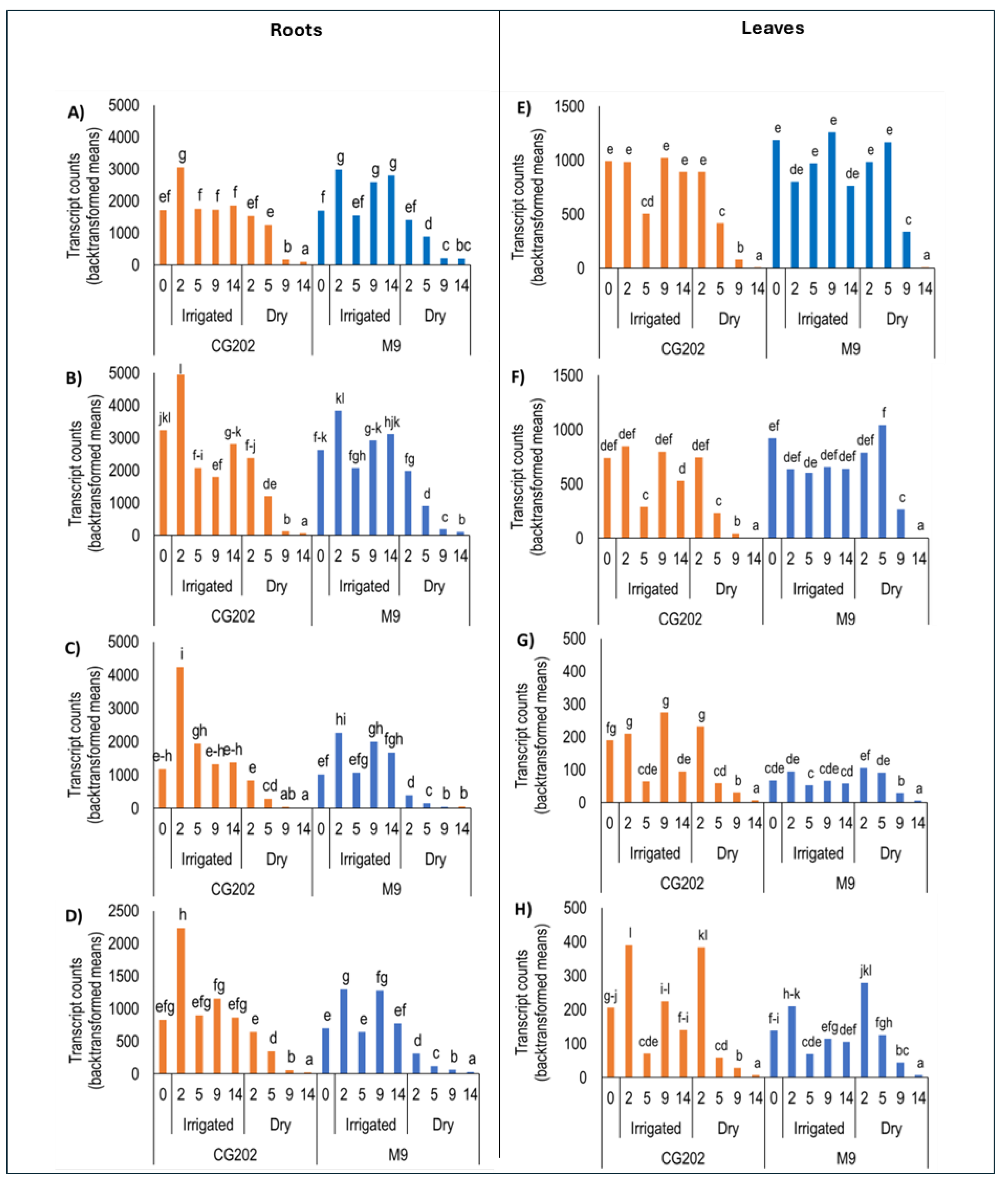
Figure 6.
Redundancy analysis (RDA) biplots for A) roots and B) leaves from two apple rootstock genotypes (CG202, M9) with treatment (Dry vs Irrigated), genotype and sampling day as response variables in Trial 1. The vectors visualise variable loadings and sample distributions by group are highlighted in brown (B) and green (A) writing for roots leaves, respectively. The asterisks indicate that explained variance is significant by the respective component at *α=0.05, ** α=0.01, *** α=0.001. SA: salicylic acid; SAG: salicylic acid glucoside; IAA: indole-3-acetic acid; ABA: abscisic acid; PA: phaseic acid; DPA: dihydrophaseic acid; X7.OH.ABA: 7’-hydroxy-abscisic acid; ABA-GE: abscisic acid glucoside; JA: jasmonic acid; JA-Ile: jasmonic acid-isoleucine; X12OH.JA: 12-hydroxyjasmonic acid; DHJA: dihydrojasmonic acid; OPC.4: 4-(3-oxo-2-(pent-2-en-1-yl)cyclopentyl)octanoic acid; cis.OPDA: 12-oxo-phytodienoic acid.
Figure 6.
Redundancy analysis (RDA) biplots for A) roots and B) leaves from two apple rootstock genotypes (CG202, M9) with treatment (Dry vs Irrigated), genotype and sampling day as response variables in Trial 1. The vectors visualise variable loadings and sample distributions by group are highlighted in brown (B) and green (A) writing for roots leaves, respectively. The asterisks indicate that explained variance is significant by the respective component at *α=0.05, ** α=0.01, *** α=0.001. SA: salicylic acid; SAG: salicylic acid glucoside; IAA: indole-3-acetic acid; ABA: abscisic acid; PA: phaseic acid; DPA: dihydrophaseic acid; X7.OH.ABA: 7’-hydroxy-abscisic acid; ABA-GE: abscisic acid glucoside; JA: jasmonic acid; JA-Ile: jasmonic acid-isoleucine; X12OH.JA: 12-hydroxyjasmonic acid; DHJA: dihydrojasmonic acid; OPC.4: 4-(3-oxo-2-(pent-2-en-1-yl)cyclopentyl)octanoic acid; cis.OPDA: 12-oxo-phytodienoic acid.
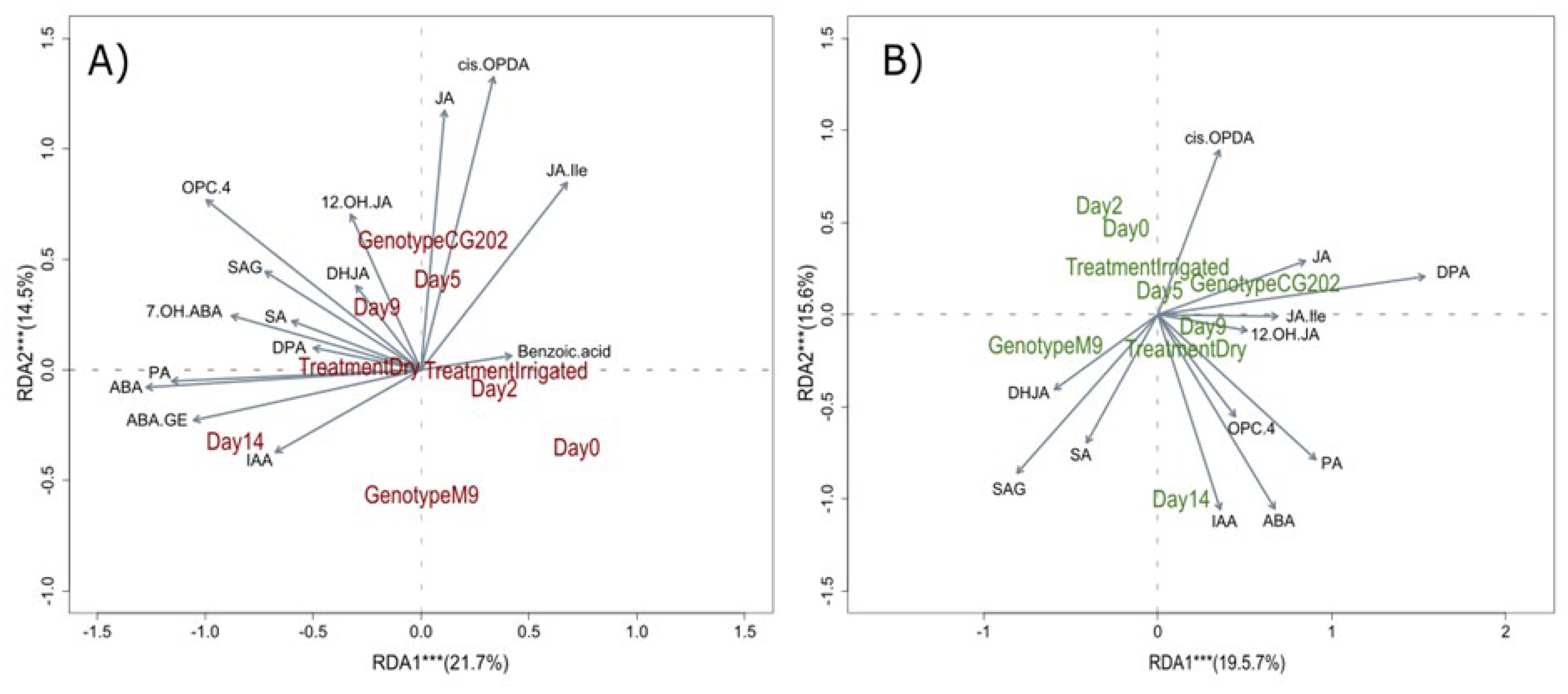
Figure 7.
Heat map presenting fold-change data of metabolite concentrations in A) roots and B) leaves, of Malus domestica GG202 and M9 rootstocks after 0, 2, 5, 9, and 14 days without water in Trial 1. All fold changes are relative to irrigated plants at the same time point. Red colouration indicates a fold-increase, blue a fold-decrease, and white is no change relative to the irrigated control. Colour intensity is indicative of the degree of change, but all fold increases greater than 10 have the same red colour intensity. A 2-fold change cutoff was applied for variable selection and bolded data indicate samples with min 2-fold change difference. SA: salicylic acid; SAG: salicylic acid glucoside; IAA: indole-3-acetic acid; ABA: abscisic acid; PA: phaseic acid; DPA: dihydrophaseic acid; 7-OH-ABA: 7’-hydroxy-abscisic acid; ABA-GE: abscisic acid glucoside; JA: jasmonic acid; JA-Ile: jasmonic acid-isoleucine; 12-OH-JA: 12-hydroxyjasmonic acid; DHJA: dihydrojasmonic acid; cis.OPDA: 12-oxo-phytodienoic acid.
Figure 7.
Heat map presenting fold-change data of metabolite concentrations in A) roots and B) leaves, of Malus domestica GG202 and M9 rootstocks after 0, 2, 5, 9, and 14 days without water in Trial 1. All fold changes are relative to irrigated plants at the same time point. Red colouration indicates a fold-increase, blue a fold-decrease, and white is no change relative to the irrigated control. Colour intensity is indicative of the degree of change, but all fold increases greater than 10 have the same red colour intensity. A 2-fold change cutoff was applied for variable selection and bolded data indicate samples with min 2-fold change difference. SA: salicylic acid; SAG: salicylic acid glucoside; IAA: indole-3-acetic acid; ABA: abscisic acid; PA: phaseic acid; DPA: dihydrophaseic acid; 7-OH-ABA: 7’-hydroxy-abscisic acid; ABA-GE: abscisic acid glucoside; JA: jasmonic acid; JA-Ile: jasmonic acid-isoleucine; 12-OH-JA: 12-hydroxyjasmonic acid; DHJA: dihydrojasmonic acid; cis.OPDA: 12-oxo-phytodienoic acid.
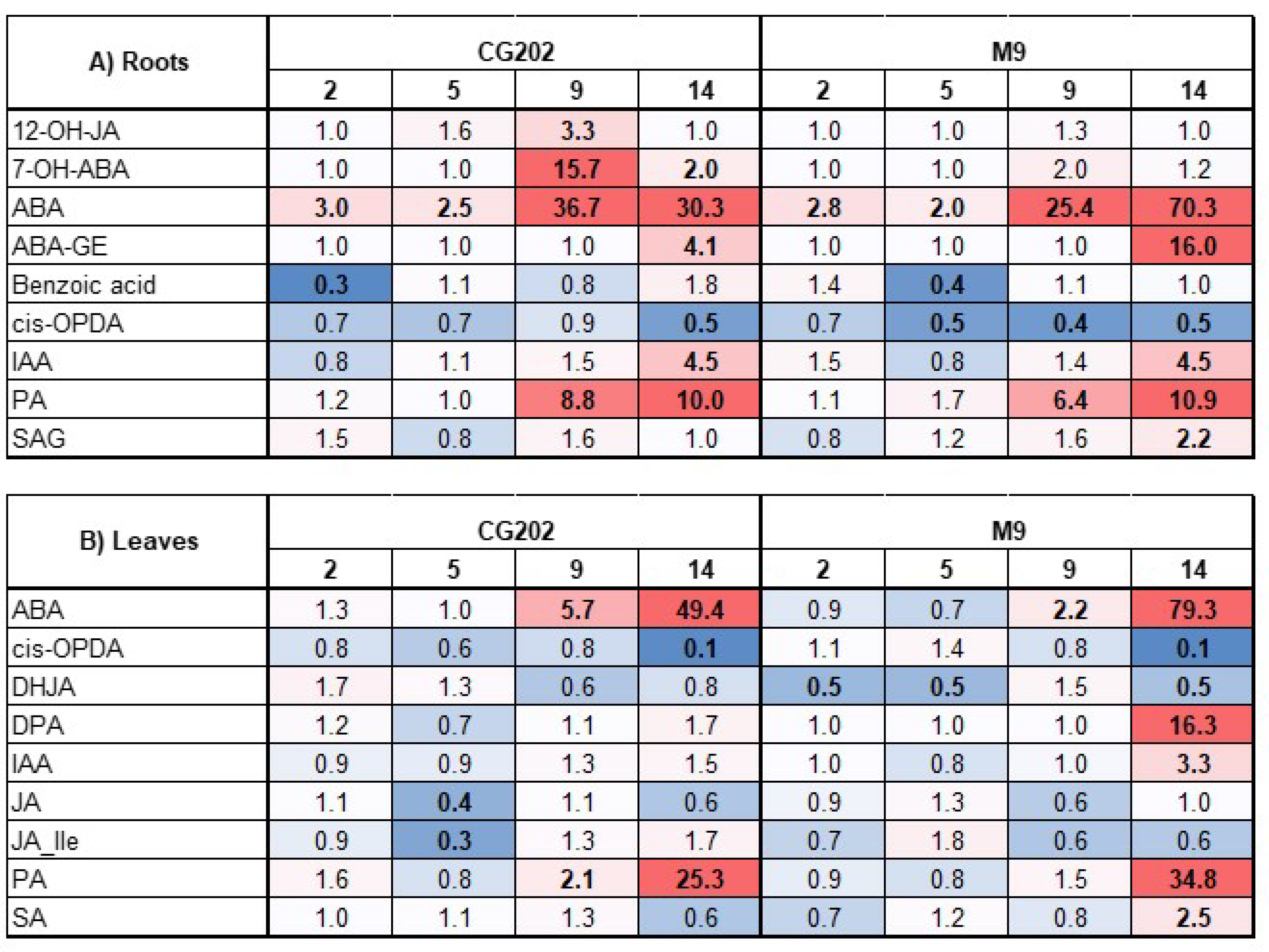
Figure 8.
Clustered Heatmap of phytohormone concentrations from two apple rootstock genotypes (CG202, M9) across common sampling timepoints (day 5, 9, 14) in Trial 2. A) roots, B) leaves. Dry: drought-treated, Irrigated: Control, ASM: Actigard-treated. Data are scaled by row with orange (A) and green (B) hues indicating positive and blue (A) and yellow (B) hues negative standard (Z-) scores, i.e relative concentrations. Asterisks indicate statistical difference of ASM-treatment by sample based on pairwise comparison with α=0.05. SA: salicylic acid; SAG: salicylic acid glucoside; DHB: dihydroxy benzoic acid; IAA: indole-3-acetic acid; ABA: abscisic acid; PA: phaseic acid; DPA: dihydrophaseic acid; 7-OHABA: 7’-hydroxy-abscisic acid; ABA-GE: abscisic acid glucoside; NeoPA: neo phaseic acid; JA: jasmonic acid; JA-Ile: jasmonic acid-isoleucine; 12-OH-JA: 12-hydroxyjasmonic acid; DHJA: dihydrojasmonic acid; OPC-4: 4-(3-oxo-2-(pent-2-en-1-yl)cyclopentyl)octanoic acid; cis-OPDA: 12-oxo-phytodienoic acid; MeJA: methyl jasmonate.
Figure 8.
Clustered Heatmap of phytohormone concentrations from two apple rootstock genotypes (CG202, M9) across common sampling timepoints (day 5, 9, 14) in Trial 2. A) roots, B) leaves. Dry: drought-treated, Irrigated: Control, ASM: Actigard-treated. Data are scaled by row with orange (A) and green (B) hues indicating positive and blue (A) and yellow (B) hues negative standard (Z-) scores, i.e relative concentrations. Asterisks indicate statistical difference of ASM-treatment by sample based on pairwise comparison with α=0.05. SA: salicylic acid; SAG: salicylic acid glucoside; DHB: dihydroxy benzoic acid; IAA: indole-3-acetic acid; ABA: abscisic acid; PA: phaseic acid; DPA: dihydrophaseic acid; 7-OHABA: 7’-hydroxy-abscisic acid; ABA-GE: abscisic acid glucoside; NeoPA: neo phaseic acid; JA: jasmonic acid; JA-Ile: jasmonic acid-isoleucine; 12-OH-JA: 12-hydroxyjasmonic acid; DHJA: dihydrojasmonic acid; OPC-4: 4-(3-oxo-2-(pent-2-en-1-yl)cyclopentyl)octanoic acid; cis-OPDA: 12-oxo-phytodienoic acid; MeJA: methyl jasmonate.
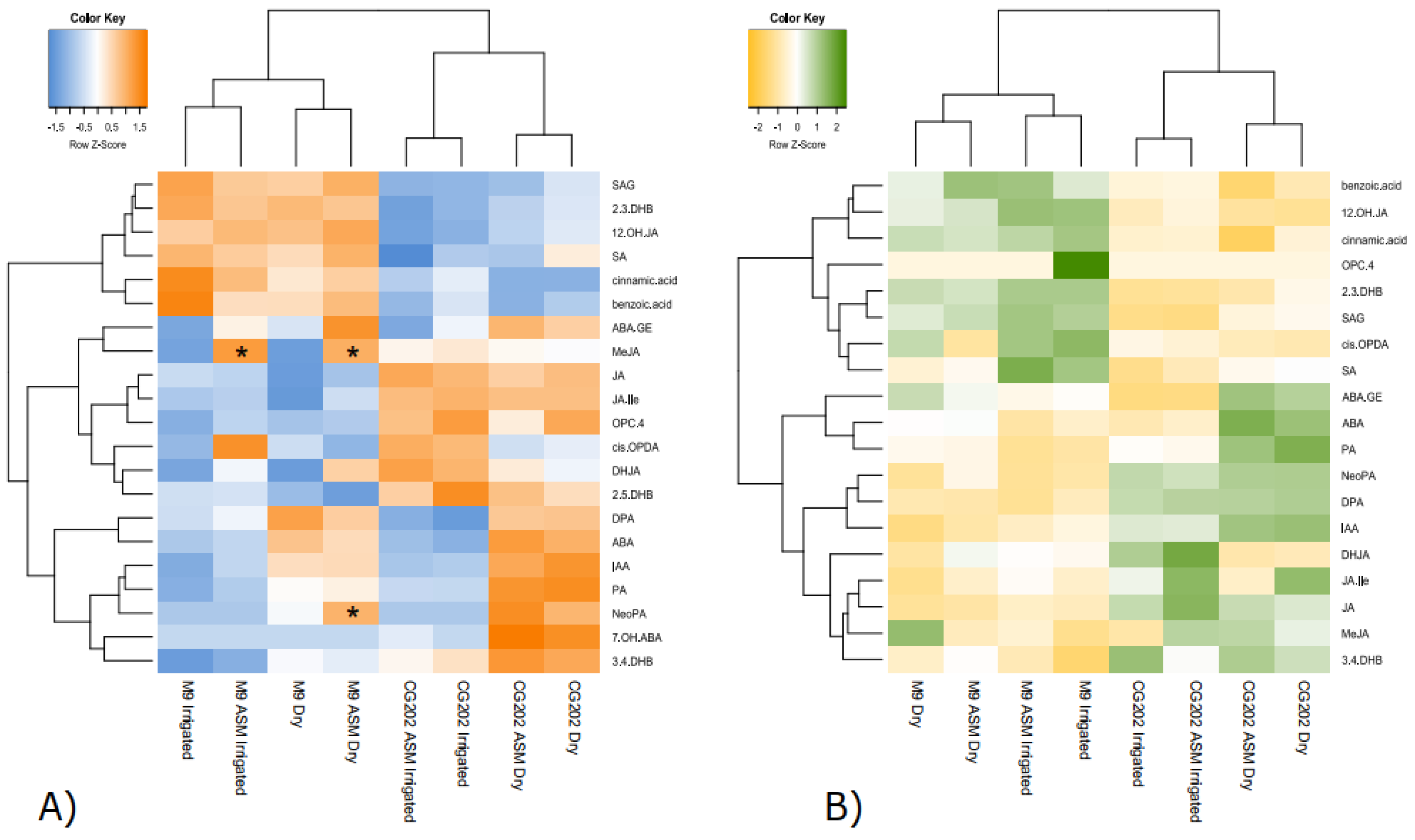
Figure 9.
Redundancy analysis (RDA) biplots for A) leaves and B) roots from two apple rootstock genotypes (CG202, M9) with treatment (Dry vs Irrigated), genotype and sampling day as response variables in Trial 2. The vectors visualise variable loadings and sample distributions by group are highlighted in green (A) and brown (B) text for leaves and roots, respectively. The asterisks indicate that explained variance is significant by the respective component at *α=0.05, ** α=0.01, *** α=0.001. SA: salicylic acid; SAG: salicylic acid glucoside; DHB: dihydroxy benzoic acid; IAA: indole-3-acetic acid; ABA: abscisic acid; PA: phaseic acid; DPA: dihydrophaseic acid; 7-OHABA: 7’-hydroxy-abscisic acid; ABA-GE: abscisic acid glucoside; NeoPA: neo phaseic acid; JA: jasmonic acid; JA-Ile: jasmonic acid-isoleucine; 12-OH-JA: 12-hydroxyjasmonic acid; DHJA: dihydrojasmonic acid; OPC-4: 4-(3-oxo-2-(pent-2-en-1-yl)cyclopentyl)octanoic acid; cis-OPDA: 12-oxo-phytodienoic acid; MeJA: methyl jasmonate.
Figure 9.
Redundancy analysis (RDA) biplots for A) leaves and B) roots from two apple rootstock genotypes (CG202, M9) with treatment (Dry vs Irrigated), genotype and sampling day as response variables in Trial 2. The vectors visualise variable loadings and sample distributions by group are highlighted in green (A) and brown (B) text for leaves and roots, respectively. The asterisks indicate that explained variance is significant by the respective component at *α=0.05, ** α=0.01, *** α=0.001. SA: salicylic acid; SAG: salicylic acid glucoside; DHB: dihydroxy benzoic acid; IAA: indole-3-acetic acid; ABA: abscisic acid; PA: phaseic acid; DPA: dihydrophaseic acid; 7-OHABA: 7’-hydroxy-abscisic acid; ABA-GE: abscisic acid glucoside; NeoPA: neo phaseic acid; JA: jasmonic acid; JA-Ile: jasmonic acid-isoleucine; 12-OH-JA: 12-hydroxyjasmonic acid; DHJA: dihydrojasmonic acid; OPC-4: 4-(3-oxo-2-(pent-2-en-1-yl)cyclopentyl)octanoic acid; cis-OPDA: 12-oxo-phytodienoic acid; MeJA: methyl jasmonate.
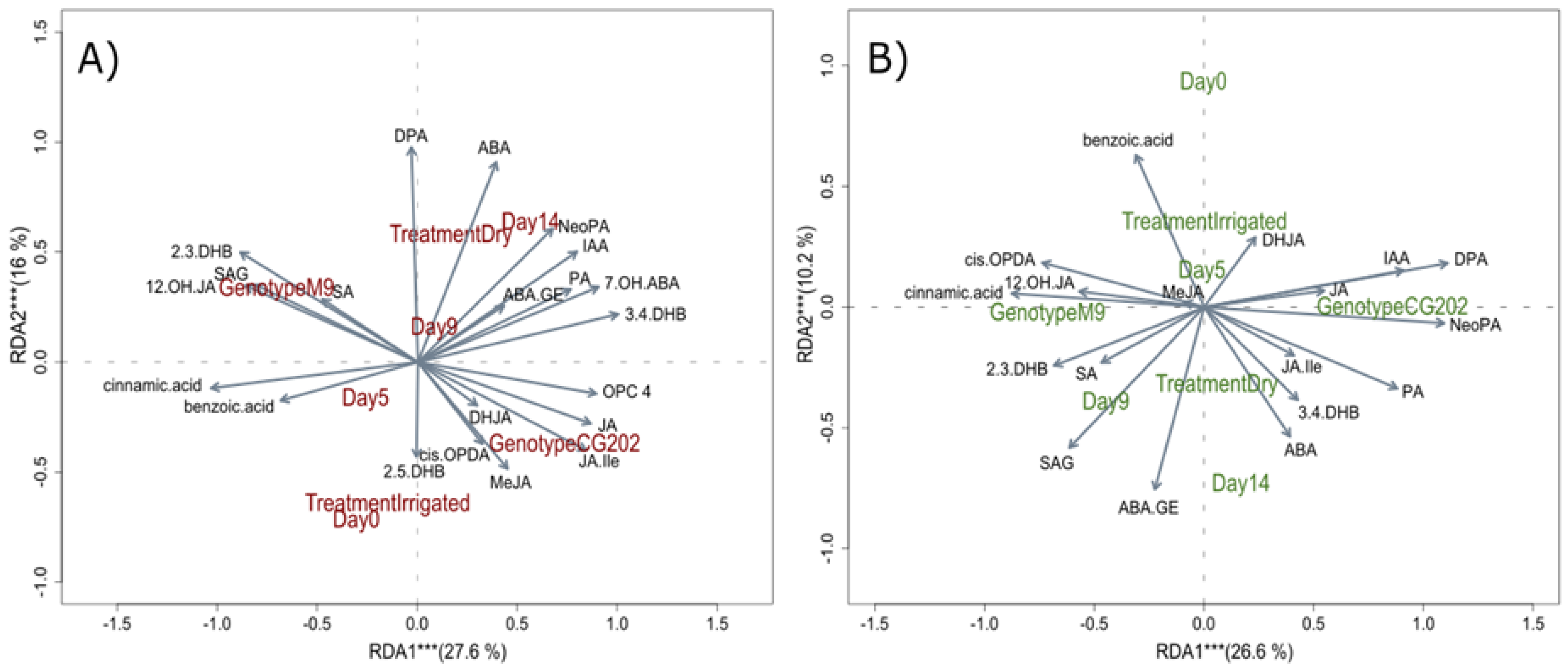
Figure 10.
Line graphs showing the mean concentrations of drought-responsive phytohormone metabolites from A) Roots and B) Leaves of two apple rootstock genotypes (CG202, M9) in Trial 2. Days indicate the time since watering for drought treated (Dry) plants, while control (Irrigated) plants were watered during the entire time course. Concentrations are given per tissue fresh weight and represent the mean of four biological replicates. 95% Confidence intervals are visualised by shading.
Figure 10.
Line graphs showing the mean concentrations of drought-responsive phytohormone metabolites from A) Roots and B) Leaves of two apple rootstock genotypes (CG202, M9) in Trial 2. Days indicate the time since watering for drought treated (Dry) plants, while control (Irrigated) plants were watered during the entire time course. Concentrations are given per tissue fresh weight and represent the mean of four biological replicates. 95% Confidence intervals are visualised by shading.
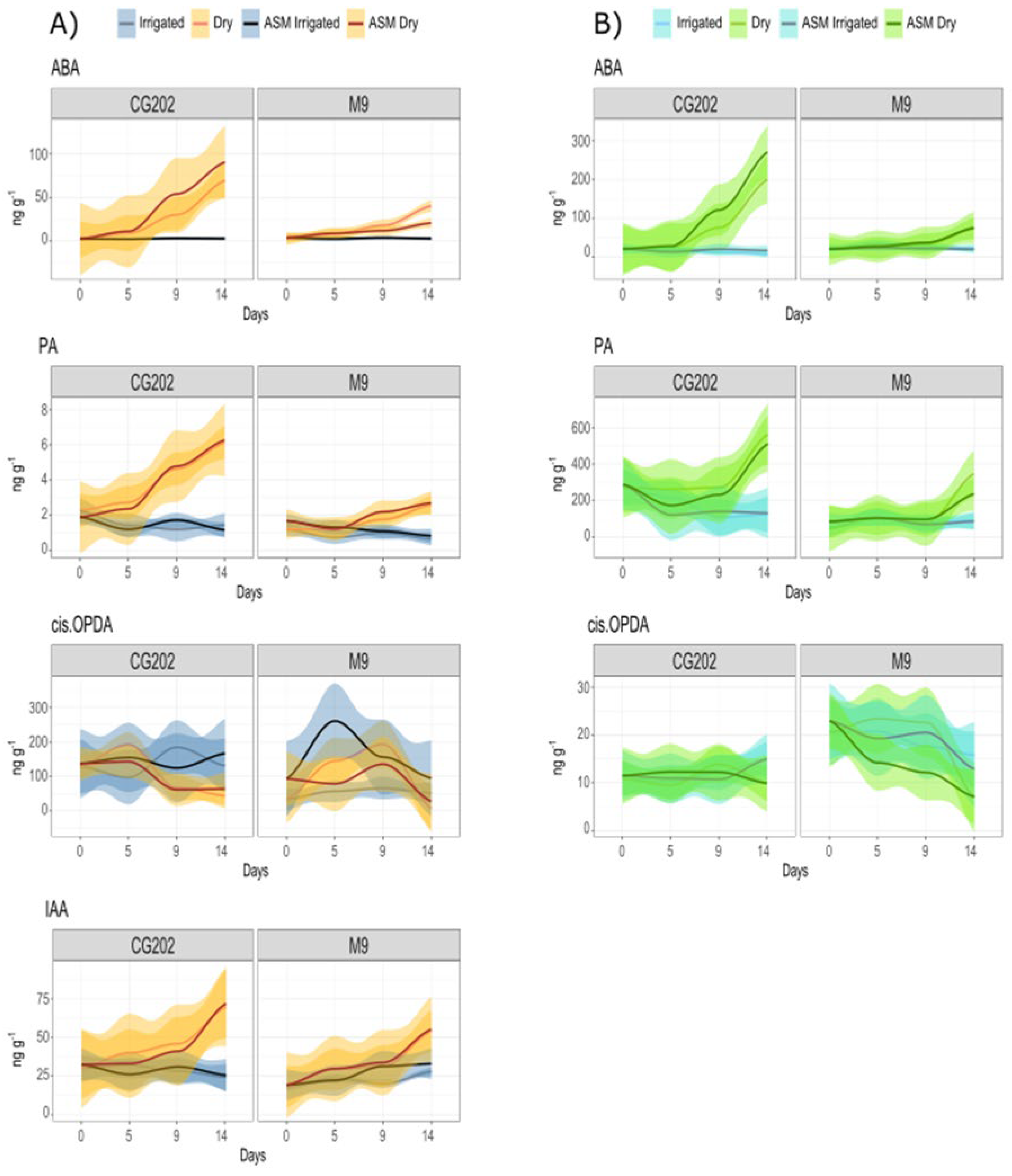

Table 1.
Change in pot weight (%) of droughted (Dry) plants over time relative to watered (Irrigated) plants at time 0 in two apple rootstocks, CG202 and M9, in Trial 1.
Table 1.
Change in pot weight (%) of droughted (Dry) plants over time relative to watered (Irrigated) plants at time 0 in two apple rootstocks, CG202 and M9, in Trial 1.
| Rootstock | ± Water | % weight relative to controls at T01 | ||||
| 0d | 2d | 5d | 9d | 14d | ||
| CG202 | Irrigated | - | 103 ±0.7 | 103 ±0.6 | 103 ±0.5 | 103 ±0.8 |
| Dry | 100 ±2.9 | 89 ± 2.5 | 79 ± 2.2 | 60 ±1.9 | 52 ±1.7 | |
| M9 | Irrigated | - | 100 ±1.3 | 100 ±1.3 | 99 ±1.0 | 103 ±1.7 |
| Dry | 100 ±1.3 | 91 ±1.4 | 82 ±1.5 | 65 ±1.8 | 54 ±1.1 | |
1Average weight (n = 5) at T0: CG202= 6.99 Kg, M9= 7.19 Kg. Least significant difference (LSD) when comparing between rootstocks = 5.0, LSD when comparing within rootstock = 1.8.
Table 2.
Change in pot weight (%) of droughted (Dry) plants over time relative to watered (Irrigated) plants at time 0 in two apple rootstocks Trial 2.
Table 2.
Change in pot weight (%) of droughted (Dry) plants over time relative to watered (Irrigated) plants at time 0 in two apple rootstocks Trial 2.
| Rootstock | ±Water | ±ASM | % weight relative to controls at T01 | ||
| 5d | 9d | 14d | |||
| CG202 | Irrigated | No ASM | 99 ±1.4 | 101 ±1.8 | 100 ±1.1 |
| ASM | 100 ±1.7 | 101 ±1.4 | 100 ±0.8 | ||
| Dry | No ASM | 77 ±2.4 | 63 ±1.6 | 56 ±1.7 | |
| ASM | 78 ±1.7 | 64 ±2.3 | 55 ±1.2 | ||
| M9 | Irrigated | No ASM | 100 ±1.2 | 98 ±1.5 | 99 ±1.4 |
| ASM | 100 ±1.6 | 98 ±2.0 | 99 ±1.7 | ||
| Dry | No ASM | 79 ±2.0 | 68 ±1.6 | 59 ±3.4 | |
| ASM | 82 ±1.7 | 69 ±0.4 | 61 ±0.9 | ||
1Average weight (n = 5) at T0: CG202= 6.38 Kg, M9= 6.35 Kg. LSD when comparing between rootstock = 4.8, LSD when comparing within rootstock = 4.2.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



