Submitted:
16 May 2025
Posted:
16 May 2025
You are already at the latest version
Abstract
Yellow stunt of root rot of Astragalus adsurgens causes premature degradation of its A.adsurgens grasslands in China. However, the etiological factors underlying livestock poisoning subsequent to the ingestion of diseased plants remain elusive. The present study aimed to comprehensively characterize the alterations in toxic substances such as swainsonine and trace element profiles after infection with Alternaria gansuense in A.adsurgens, thereby elucidating the underlying mechanisms of livestock toxicity. Using ELISA and regression analysis, we found that diseased plants had higher selenium levels than healthy ones, with varietal differences. Selenium in the Zahua variety was higher in healthy plants, while the Henan variety had the highest levels in diseased ones. Moreover, diseased plants demonstrated decreased levels of iron, zinc, sodium, and magnesium, while manganese and calcium concentrations remained unchanged. Swainsonine was detected in both healthy and infected specimens of Zhongsha No.1 and Henan varieties, with a marked post - infection increase. In conclusion, swainsonine are the primary toxins causing livestock poisoning, and soil-accumulated selenium is unlikely to poison animals. However, potential correlations might exist among the contents of selenium, sodium, and swainsonine. We recommend cautious use of diseased A.adsurgens as livestock feed.
Keywords:
astragalus adsurgens
; alternaria gansuense
; element differences
; swainsonine
1. Introduction
Astragalus adsurgens (Pall) is an important perennial legume grass of the Astragalus genus in China. Due to its good yield, high protein content, and high nutritional value, it is used as feed in China[1,2]. It is widely grown in temperate regions of Russia, Mongolia, Japan, South Korea, and North America[1]. It also has excellent characteristics such as drought resistance and cold resistance, and is usually used as a windbreak and sand fixing plant, playing an important role in protecting the ecological environment. Other countries such as the United States believe that A.adsurgens is a poisonous herb, also known as locoweed, which can cause poisoning in livestock after consumption. However, research in China has found that it is a low toxicity plant and safe for ruminant animals to consume[3,4,5].
The yellow stunt and root rot caused by Alternaria gansuense is a serious systemic disease found on A.adsurgens in China. It is also an important factor contributing to the premature degradation of A.adsurgens grasslands in northern China[6,7].This fungus was initially named Embellisia astragali, but later, according to Woudenberg’s new taxonomic system, the taxonomic status of this fungus was revised, and it was reclassified and named A.gansuense[8,9,10,11].
The symptoms of the disease include discoloration of stems, wrinkling of leaves, dwarfing and proliferation of branches, decay of roots, and death in severe cases[12]. Previous studies have found that the main factor causing poisoning in livestock after consuming madder is the production of toxic coumarin by endophytic fungi in the locoweed. The pathogenic bacterium of yellow stunt and root rot, A.gansuense, is closely related to endophytic fungi in locoweed. After making pellet feed from healthy and infected A.adsurgens for consumption by mice, the weight gain of mice fed with infected A.adsurgens was smaller, the content of lactate dehydrogenase and alanine aminotransferase was higher, and the feed utilization rate was lower. Therefore, the toxicity of A.adsurgens was enhanced after infection[8,10,13]. And there is a yeast amino acid reductase gene related to the production of coumarin in A.gansuense[8]. The pathogen can also synthesize swainsonine, which makes infected A.adsurgens toxic to animals that ingest it, causing serious effects on animal growth and development. Low doses of swainsonine may lead to weight loss and biochemical damage, while high doses may cause chronic diseases in livestock after prolonged consumption[14,15].
In addition to the effects of the secondary metabolite swainsonine in infected plants, A.adsurgens is also a selenium-accumulating plant belonging to the genus Astragalus. Studies have shown that when the content of selenium and selenium compounds in plants is too high, feeding them to animals can also cause poisoning[16]. At present, it is generally believed that seventeen elements are essential elements, including nine macronutrients and night micronutrients[17]. Mineral elements play a crucial physiological role in plant growth. If a certain element is deficient, the plant cannot grow and develop normally, leading to corresponding physiological diseases. Moreover, insufficient or imbalanced supply of elements in plants can also influence the occurrence of infectious disease caused by fungi or bacteria[18,19]. The content of mineral elements can influence plant diseases, mainly reflected in two aspects. Firstly, as components of plants, mineral elements significantly affect the formation and changes in the morphological structure of plant cells, thereby influencing the infection process of pathogens on host plant cells. Secondly, mineral elements participate in and regulate various metabolic reactions in plants, which can affect their disease resistance of plants[20,21,22].
The occurrence of many diseases is directly related to the deficiency of elements. Applying nitrogen fertilizer to Lolium perenne can accelerate the expansion of Rhizoctonia solani, leading to worsening of the disease[23]. Potassium, like other elements, has a certain effect on improving plant disease resistance. It can coordinate carbon and nitrogen metabolism processes, promote protein synthesis, and therefore increase potassium fertilizer to reduce the content of amino acids in plants, which is beneficial for promoting the synthesis of phenolic compounds in plants and hindering the growth and development of fungal hyphae [24]. A study has found that the application of potassium fertilizer can increase the content of polyphenolic compounds in rapeseed and inhibit the germination of A.brassicae spores, leading to a decrease in the incidence of rapeseed black spot disease[25]. When the ratio of potassium and nitrogen is balanced, the proportion of lemon infected with gummosis decreases[26].
Calcium is an important element in plants’ resistance to fungal or bacterial infections. When pathogens destroy the cell wall of host plants by producing pectinase, which then infects plant cells, Ca2+ can form calcium galacturonate, enhancing cell wall stability and reducing the impact of pectinase on the cell wall. It can also enhance the activity of various enzymes such as catalase (CAT) and superoxide dismutase (SOD) in plants, thereby improving plant resistance to pathogens[27,28]. Magnesium can inhibit plant diseases such as Candidatus Liberibacter spp. and A.solani, and promote the occurrence of other plant diseases, such as alfalfa mosaic disease and cotton root rot disease (Phytotrichum omnivorum). Magnesium is also a component of plant cells involved in resistance to pathogens[29].
Trace elements can also be closely related to the occurrence and development of plant diseases by participating in metabolic reactions and affecting enzyme activity. Zinc is an essential element for plant sugar metabolism. The application of zinc fertilizer has no effect on stem rust and stripe rust in winter wheat but significantly reduces the occurrence of Puccinia reconditata. As a component of phenolase, copper can affect the synthesis of phytoalexin, and is also widely used as an effective ingredient in inorganic fungicides. It has been reported that the application of copper elements or the combined application of copper and iron elements will increase the incidence rate of onion white rot disease. On the other hand, the spraying of copper fertilizers can reduce the leaf blight of cucurbitaceous crops (Alternaria cucumerina)[30,31]. Boron can regulate the synthesis of various phenolic compounds in plants. In addition, there are reports that application of calcium chloride can alleviate powdery mildew, and the proportion of chlorine in the fertilizer determines its effectiveness against powdery mildew.[32,33].
The objectives of this paper is to elucidate the differences in mineral elements and changes in the content of toxic substance swainsonine between diseased plants and healthy plants among different varieties of A.adsurgens. In this study, we first collected samples of diseased and healthy plants, identified the materials through identification, and then ground them into grass powder. The corresponding mineral element content and swainsonine content were measured using relevant instruments. These results contribute to a deeper understanding of the differences in toxic substance content between yellow stunt and root rot and healthy plants among different varieties of A.adsurgens, providing data support for further understanding of the disease and preventing its further expansion.
2. Materials and Methods
2.1. Overview and Management of the Experimental Site
The tested plants of A.adsurgens were collected from the experimental site of Heping Campus of Gansu Agricultural Vocational and Technical College in Yuzhong County, Lanzhou City. The experimental site is located on the Loess Plateau, with an east longitude of 103.58°, north latitude of 36.00°, and an altitude of 1750 meters (Figure 1A). It belongs to a temperate continental climate, with an average annual precipitation of about 300 400 millimeters, and the soil is yellow loess. The plant was established in May 2012 using the method of hole sowing, and the field management method is to irrigate once a year, manually remove weeds, and no chemical fertilizer is applied.
2.2. Collection and Preservation of Plant Materials
In October 2015, mature branches of A. adsurgens (Zahua, Zhongsha No.1, Henan, and Liaoning Fuxin cultivars) were collected. Whole plants were harvested using large flat-bladed shears, leaving a 5 cm basal stubble (Figure 1B and C). From each sampled community, 50 healthy branches and 50 branches with yellow stunt and root rot symptoms (characterized by complete reddish-brown discoloration and confirmed by microbiological testing) were randomly selected. All branches were individually labeled, sealed in kraft paper envelopes, and transported to the laboratory for further analysis. Upon arrival, samples were oven-dried at 60 ℃ for 48 hours. Dried branches were pulverized using a mechanical grinder, and resulting grass powder was transferred into sterile resealable plastic bags. Between samples, the grinder was thoroughly cleaned to prevent cross-contamination. Powdered samples were stored at -20 ℃ until subsequent analysis.
2.3. Digestion of Grass Samples
All glassware was pre-treated by immersion in 50% nitric acid for a 48-hour period, followed by thorough rinsing with ultrapure water and oven-drying at 105°C until use. For each sample, 1.0 g of powdered herb was accurately weighed into digestion tube. A total of 10 mL acid mixture (8 mL concentrated nitric acid, 1 mL perchloric acid, and 1 mL concentrated sulfuric acid) was sequentially added. Blank controls containing only the acid mixture were prepared identically. The digestion system was preheated to 420°C and equilibrated for at least 15 minutes prior to use. With the fume hood activated, digestion tubes were placed in the heating block, capped, and digested at 420°C for 30 minutes. Post-digestion, tubes were allowed to cool to room temperature, rinsed with ultrapure water, and the contents quantitatively transferred to a 50 mL volumetric flask. Final volume adjustments were made with ultrapure water. Digested solutions were filtered through a 0.44 μm cellulose acetate membrane using a 50 mL syringe, collecting the filtrate into pre-cleaned, dried amber glass vials. All samples were stored at 4°C pending analysis, with four technical replicates performed per sample.
2.4. Isolation and Cultivation of A.gansuense
Symptomatic branches of A.adsurgens displaying complete reddish-brown discoloration were selected. A 4-5 cm segment was excised, rinsed under running tap water, and transferred to a sterile Petri dish on a laminar flow hood (ESCO, Airstream 1360). Surface sterilization was performed via sequential immersion in 70% ethanol for 30 seconds (ethanol discarded into a designated waste container), followed by 1% sodium hypochlorite solution for 1 minute, and triple rinsing with sterile deionized water. Excess moisture was blotted using autoclaved filter paper. The sterilized segment was longitudinally sectioned with a sterile scalpel, and medullary tissue was aseptically transferred to potato dextrose agar (PDA) plates. Plates were incubated in a low-temperature incubator (Panasonic MIR-554-PC, Japan) at 22°C in darkness for 4 days. Pure cultures (strain LYZ0001) were obtained by hyphal tip isolation from colony margins on fresh PDA plates.
For A.gansuense sporulation, cultures were maintained on PDA for 4 weeks. Single-spore isolation was conducted under a stereomicroscope (Olympus SZ51, Japan) using flame-sterilized forceps. Fruiting bodies were located microscopically, transferred to fresh PDA plates, and gently streaked to release spores. Ten replicate plates were prepared. After 24 hours of incubation, germinated spores were individually picked with a fine sterile inoculation needle and transferred to separate PDA plates (10 replicates). After 14 days of dark incubation at 22°C, a pure colony (strain LYZ0001D) was selected for subculture. For biomass production, 3-4 week-old LYZ0001D mycelia were transferred to 2 mL sterile water, homogenized, and inoculated into 250 mL Erlenmeyer flasks containing 100 mL potato dextrose broth (PDB). Cultures were incubated in the dark at 22°C on an orbital shaker (Shanghai Sukun NSKY-2102C) at 150 rpm for 20 days. Harvested mycelia were filtered, blotted dry with autoclaved filter paper, and stored at -80°C in sterile microcentrifuge tubes.
2.5. Determination of Selenium Content
The prepared test solutions were transferred to 10 mL centrifuge tubes and arranged in sequence on a tube rack. Selenium standard solutions at concentrations of 5, 10, 25, 50, and 100 μg/L were prepared using 8% nitric acid as the matrix-matched diluent. All samples and standards were analyzed using a Thermo Scientific iCAP Q ICP-MS system. Instrumental parameters were optimized as follows: extraction voltage 120 V, radio frequency (RF) power 1400 W, carrier gas flow rate 1.05 L/min, and nebulizer temperature 2 ℃. Each sample was measured in triplicate with automatic internal standardization. Prior to analysis, the instrument was calibrated using a multi-point standard curve (R2 ≥ 0.999) and daily performance verification was conducted with certified reference materials.
2.6. Determination of Mineral Elements
Dilute 2.5 mL of standard solutions for Copper (Cu), Manganese (Mn), Iron (Fe), Zinc (Zn), Calcium (Ca), Sodium (Na), and Magnesium (Mg) elements, and make up to volume with a 50 mL volumetric flask. Dilute step by step according to the concentration gradient, inject the sample from low to high according to the measurement conditions of different elements, and obtain a qualified standard curve. When preparing magnesium element, first prepare a 50 mg/mL strontium chloride solution, add 5 mL of strontium chloride solution to the magnesium element solution, and then make a series of concentration gradient standard solutions by constant volume. When preparing the standard curve of magnesium, strontium chloride can reduce the interference effect of other elements. Place the prepared test solution in the corresponding element determination conditions in Table S1 in order of numbering, and then directly inject it into the atomic absorption spectrophotometer (Thermo, S SERIES, USA). Record the absorbance values of each sample and substitute them into the standard curve equation to calculate the content. Due to the high concentrations of calcium and magnesium in the sample solution, they were diluted 25 and 100 times respectively before measurement.
2.7. Preparation for Determination of Swainsonine
The plant materials consisted of the aforementioned prepared grass powder, including the disease-resistant cultivar Henan, the low-resistance cultivar Zhongsha No.1, and plants of an undetermined cultivar collected from severely diseased specimens. The microbial material comprised the previously obtained A.gansuense mycelium. The prepared instruments include weighing bottles, mortars, beakers, volumetric flasks, pipettes, 96 well ELISA plates, precision air drying ovens, Powerwave XS full wavelength enzyme-linked immunosorbent assay (ELISA) analyzer (USA), ultrasonic cleaning machine (Ningbo Xinzhi Biotechnology Co., Ltd., SB-5200D), and centrifuge (Thermo). The reagents include acetic acid, methanol, boric acid (analytical pure), swainsonine standard (provided by the College of Veterinary Medicine, Northwest A&F University), α-mannosidase, and p-nitrophenyl-α-D-mannosidase (SUPELCO).
2.8. Sample Processing
First, place the grass powder in a sterilized centrifuge tube and freeze dry it in a freeze dryer for 48 hours to remove moisture. Weigh 100 mg of dried grass powder, grind it thoroughly into fine powder using a grinder machine, and transfer it to a dry and sterilized centrifuge tube. Add 5 mL of acetic acid and sonicate at 60 ℃ for 30 minutes. Then, centrifuge at 5000 r/min for 15 minutes, collect the supernatant and place it in a clean small beaker. Add the same volume of acetic acid again and repeat the previous step: centrifuge at 5000 r/min for 15 minutes, collect the supernatant, and repeat this process twice. Place the supernatant obtained three times in the same small beaker, evaporate the extraction solution to obtain a residue, dissolve the residue obtained by evaporation in an appropriate amount of methanol, centrifuge at 12000 r/min for 10 minutes, discard any remaining solid residue, dilute to 10 mL with chromatography pure methanol, and store in a refrigerator at 4 ℃ for subsequent analysis.
2.9. Drawing and Measurement of the Standard Curve for Swainsonine
Preparation of swainsonine storage solution (1000 µg/mL), weigh 10 milligrams of swainsonine standard and dissolve it in 10 milliliters of chromatographically pure methanol. Prepare a series of swainsonine standard solution with 9 concentration gradients using the stock solution. Add 40 µL of each swainsonine standard solution at different concentrations into individual wells of a 96-well plate, ensuring three replicates for each concentration. Additionally, perform blank control and negative control simultaneously. Add 50 µL of substrate solution (20 mmol/L p-nitrophenyl-α-D-mannoside solution) and 50 µL of buffer solution (pH=4.5) to each well, gently shake the plate to ensure thorough mixing, and incubate in a 37 ℃ oven for 1 hour. After incubation, add 20 µL (0.025 μg/mL) of alpha glucosidase solution to each well, gently shake well, and incubate again at 37 ℃ oven for 1 hour. Following this, add 100 µL of reaction termination solution to each well, and gently shake to ensure homogeneity. Immediately measure the absorbance values at a wavelength of 405 nm using a microplate reader. Calculate the inhibition rate and plot a standard curve with the concentration of the swainsonine standard solution on the horizontal axis and the inhibition rate of swainsonine on α-mannosidase as the vertical axis, and obtain a regression equation for calculating the sample content.
Take 40 μL of the sample solution to be tested and add it to a 96 well plate. Repeat 3 times for each sample at the same time. Add 50 μL of buffer solution and substrate solution to each well, gently shake well, and incubate in a 37 ℃ oven for 1 hour. Add 20 μL of alpha mannosidase solution to each well, gently shake well, and incubate at 37 ℃ for 1 hour. Add 100 μL of termination solution to each well to complete the reaction, mix well, and measure the absorbance value of each well solution at a wavelength of 405 nm. According to the regression equation obtained in the previous step, substitute the calculated inhibition rate of α-mannosidase for each sample into it, calculate the concentration of swainsonine in the sample solution, and further convert it to obtain the swainsonine content in the grass sample.
3. Results
3.1. Determination Results of Selenium Content
After the occurrence of yellow stunt and root rot, the selenium content in the plant of A.adsurgens significantly increased compared to healthy plants (P<0.01) (Figure 2). Selenium content in healthy A.adsurgens plants from 192.42 to 366.61 μg/kg, the selenium content of diseased plants ranges from 747.78 to 1108.95 μg/kg. The selenium content in different varieties of A.adsurgens also varies. Before disease onset, the selenium content in the mixed flowers was significantly higher than that in Zhongsha No.1 (P<0.05); When the disease occurred, the selenium content in Henan was significantly higher than that in Zahua and Zhongsha No.1 varieties (P<0.05) (Table S2).
3.2. Standard Curves for Other Mineral Elements
The standard curve regression equations for the 7 tested mineral elements are shown in Table S3, and the linear correlation coefficients of all regression equations are greater than 0.995. Measure the absorbance values (Abs) of each element in the sample, input them into the regression equation of the standard curve, and obtain the concentrations of each mineral element in the sample solution. Based on the total volume of the solution and the mass of each grass sample taken during the experiment, calculate the content of various mineral elements in each sample of A.adsurgens.
3.3. The content of Elements
Six mineral elements were measured in diseased and healthy plants of different varieties of A.adsurgens, but no copper was detected. The copper content in A.adsurgens did not reach the minimum detection limit of the atomic absorption spectrophotometer. The following are the specific results (Table S3).
The calcium content in plants that have not been infected with yellow stunt and root rot was 3366.83−4925.83 mg/kg, while the content in diseased plants ranges from 3363.16−4213.55 mg/kg. For different varieties, the calcium content increased after the disease occurred in Liaoning Fuxin and Henan, while the content decreased after the disease occurred in Zahua and Zhongsha No.1. Moreover, in the comparison of calcium content between healthy and diseased plants of the same variety, the plants from Henan showed a significant increase in calcium content (P<0.05) (Figure 3A). The difference in calcium content between healthy and diseased varieties was not significant (Table S4).
The iron content in healthy plants ranges from 63.57 to 198.88 mg/kg, while the content in diseased plants ranges from 53.75 to 135.26 mg/kg, showing an overall downward trend. However, the content in Liaoning Fuxin varieties has increased after the disease. The comparison within the same variety shows that the iron content significantly decreases in the diseased plants of Zahua and Zhongsha No.1(P<0.05) (Figure 3B). The comparison between varieties showed that the iron content in the plants of A.adsurgens without yellow stunt and root rot was significantly higher in Zhongsha No.1 than in Henan and Liaoning Fuxin varieties; In the case of disease, the iron content in Liaoning Fuxin was significantly higher than the other three varieties (P<0.05) (Table S4).
The magnesium content in healthy A.adsurgens plants ranges from 1902.82 to 2601.19 mg/kg, while the magnesium content in diseased plants ranges from 1523.38 to 2288.30 mg/kg. After being infected with the yellow stunt and root rot, the magnesium content in all four varieties of A.adsurgens decreased. The comparison within the same variety shows that the magnesium content in Zahua decreases significantly when it is diseased (P<0.05) (Figure 3C). There was no significant difference in magnesium content among different varieties when healthy. In the case of disease, there was a significant difference in magnesium element content among different varieties (P<0.05), with Henan having the highest content and Zhongsha No.1 having the lowest (Table S4).
The content of manganese in healthy plants was 17.24−23.73 mg/kg, and the content in diseased plants was in the range of 15.00−24.51 mg/kg. The comparison within the same variety shows that the manganese content in both Zahua and Zhongsha No.1 decreases significantly when they were diseased (P<0.05) (Figure 3D). In terms of varieties, there was no significant difference in the manganese content among different varieties when they are healthy. In the case of the occurrence of yellow stunt of root rot, the manganese content in the plants from Henan and Liaoning Fuxin was significantly higher than that in the mixed-flower variety and Zhongsha No. 1 varieties (P<0.05) (Table S4).
The sodium content in plants that had not been infected with yellow stunt and root rot was 624.14−846.43 mg/kg, while the sodium content in the affected plants ranges from 397.72−741.06 mg/kg. Except for Henan, the sodium content of the other three varieties sharply decreased after the occurrence of yellow stunt and root rot (P<0.05) (Figure 3E). In terms of variety, the sodium content of the healthy plant in Liaoning Fuxin was significantly higher than that of Zhongsha No.1 (P<0.05). In the case of illness, the content in Henan was significantly higher than the other three varieties (P<0.05) (Table S4).
The zinc content in plants that had not been infected with yellow stunt and root rot was 7.58−13.60 mg/kg, while the zinc content in diseased plants ranges from 8.16 to 11.41 mg/kg. The comparison within the same variety showed that the zinc content in both Henan and Zahua decreased significantly when they were diseased (P<0.05) (Figure 3F). In terms of variety, the zinc content in both healthy and susceptible plants was significantly lower than that in Henan and Zahua (P<0.05) (Table S4).
Overall, after being infected with the yellow stunt and root rot, there was little change in the magnesium content in A.adsurgens; The calcium content decreased, and the difference was not significant; The content of iron, zinc, and magnesium decreased significantly (P<0.05). The sodium content decreased significantly (P<0.01). The selenium content increased, and the difference was extremely significant (P<0.001) (Figure 4).
3.4. Results of Swainsonine Test
The regression equation of the standard curve fitted based on nine concentrations of standard solutions was y=0.3243x-0.2276, with a correlation coefficient R2=0.9984 (Figure S1). By enzyme-linked immunosorbent assay, swainsonine was detected in both infected and uninfected A.adsurgens, with a concentration ranging from 22.13 to 128 mg/kg. The content of swainsonine in healthy plants ranges from 22.13 to 27.91 mg/kg, while in diseased plants it ranges from 81.86 to 128.00 mg/kg. After infection with yellow stunt of root rot, the content of swainsonine in Zhongsha No.1 increased by 193%, and the swainsonine content in Henan increased by 329% (Figure 5). Furthermore, A.gansuense can also produce swainsonine, with a content of 55.02 mg/kg.
3.5. The correlation Between Swainsonine and Elements
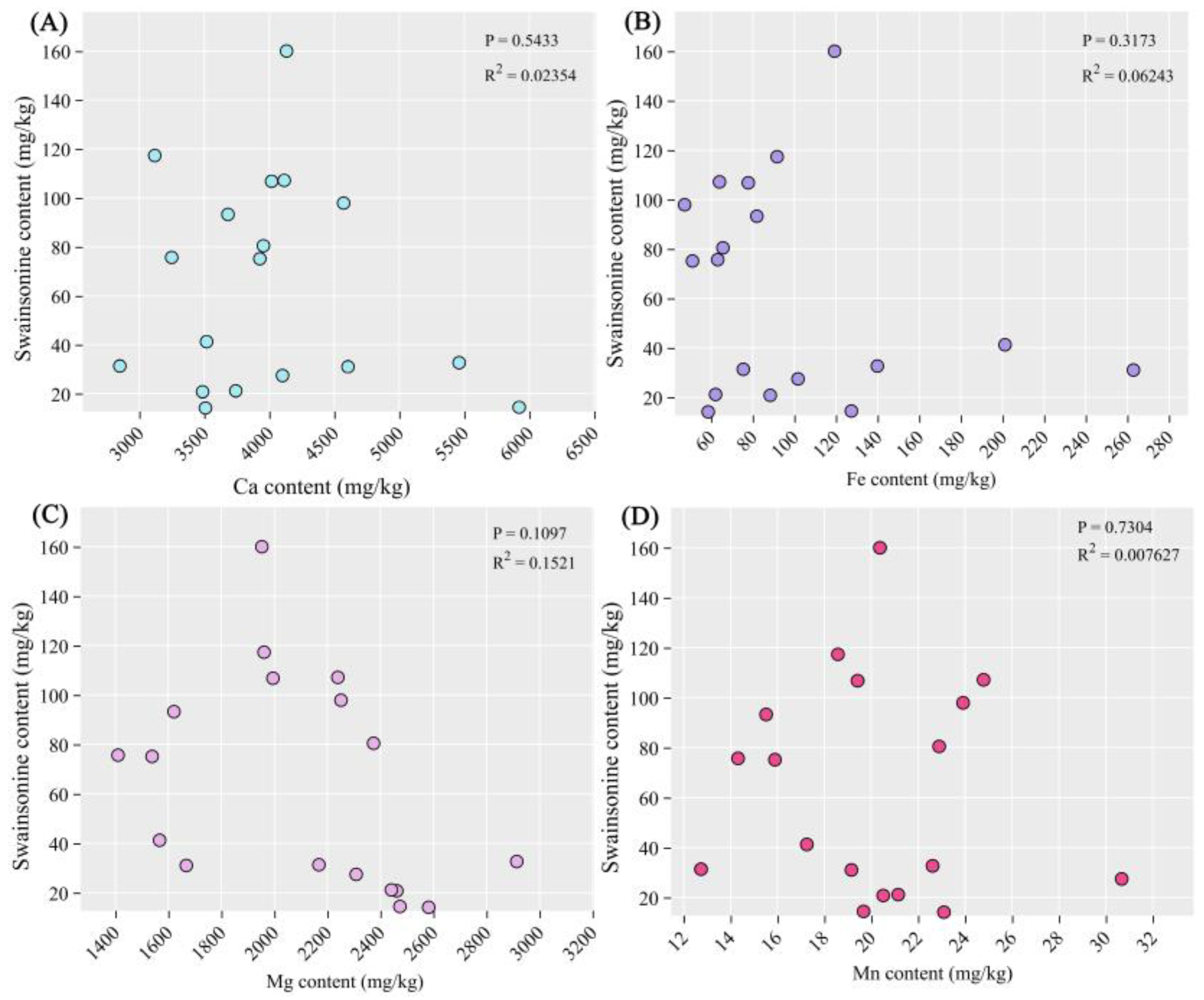
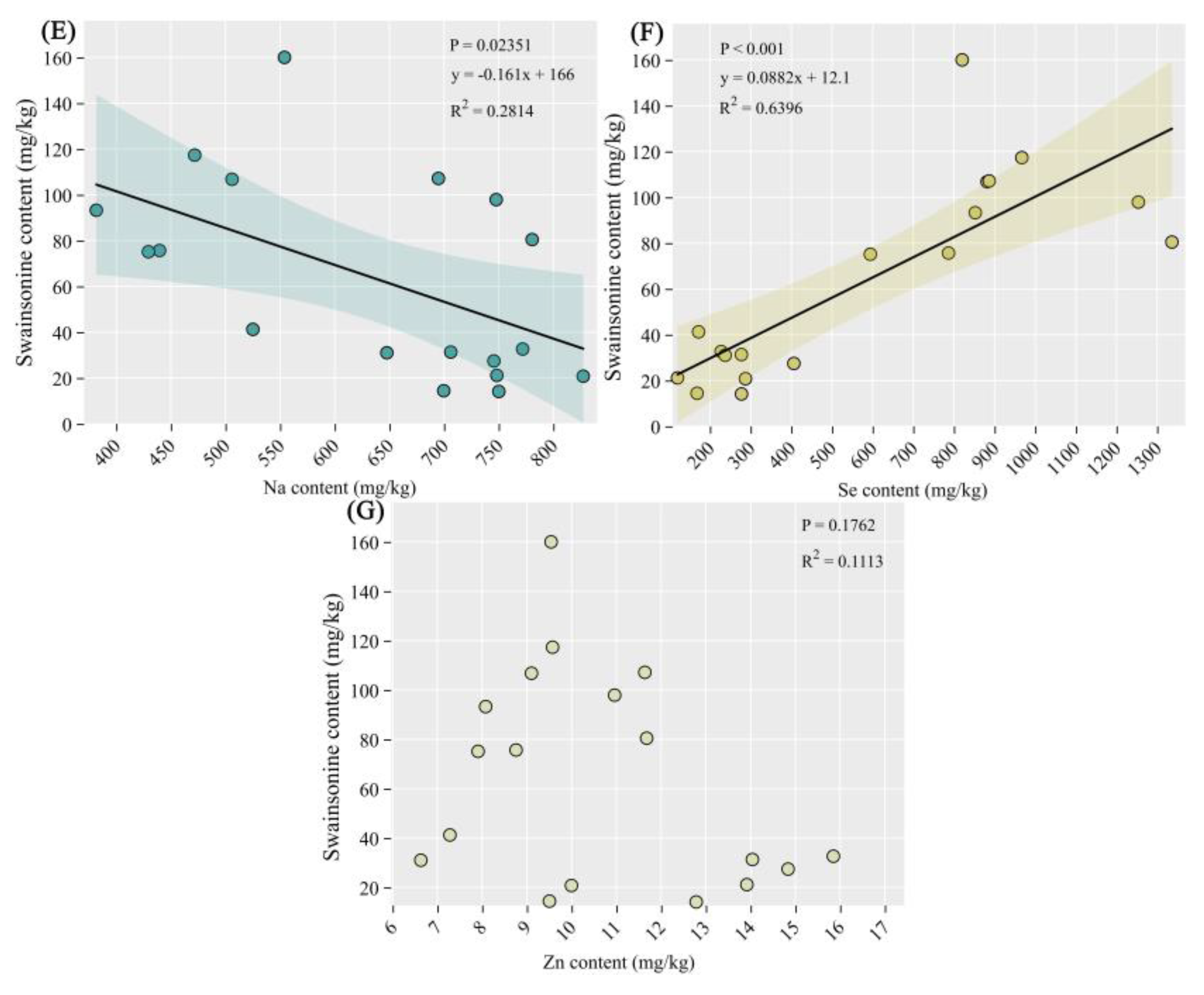
As shown in Figure 6E and F, there was a significant negative correlation between the content of swainsonine and the content of sodium element (P<0.05), while there was an extremely significant positive correlation between the content of swainsonine and the content of selenium element (P<0.001). In addition, there was no significant correlation between swainsonine and other elements.
4. Discussion
4.1. Changes in Selenium Content of Diseased Plants and Its Impact on Livestock Feeding
Selenium is an essential trace element for humans and animals, significantly influencing the activity of over thirty enzymes in organisms[34]. However, selenium exhibits a dualistic nature. In the process of selenium’s biological functions, it exhibits a remarkable dual nature. When selenium is deficient, it can lead to decreased human immunity, malnutrition, and trigger diseases such as hepatic necrosis, Keshan disease, and Kaschh-Beck msease[35,36]. In animals, selenium deficiency may cause white muscle disease, reduced immunity, and stunted growth[37]. Conversely, excessive selenium intake can also lead to poisoning in both humans and animals. The risk level for acute selenium poisoning in animals is 2−5 mg/kg body weight via a single ingestion. [38,39]. Selenium in natural plants is usually divided into two types: organic selenium and inorganic selenium. Notably, organic selenium constitutes the predominant proportion, accounting for approximately 80% of total selenium content. This organic fraction encompasses various bioactive compounds including selenoproteins, selenium-containing nucleic acids, and selenium polysaccharides, which exhibit superior bioavailability and are more readily assimilated by the animal digestive system compared to their inorganic counterparts. Organic selenium constitutes approximately 80% of total selenium content and is more readily absorbed by animal digestive systems[40]. In general plant species, the selenium concentration typically ranges from 0.05 to 1.50 ppm. However, selenium hyperaccumulator plants exhibit remarkably elevated selenium concentrations, reaching several thousand ppm. Notably, certain species within the Astragalus genus demonstrate exceptional selenium accumulation capacity, with concentrations ranging from 1000 to 10000 ppm[41,42]. These hyperaccumulators efficiently absorb selenium from soil and tolerate high selenium levels. However, excessive selenium accumulation in plants may produce foul odors, rendering forage unpalatable. Direct consumption of such plants by animals can cause acute selenium poisoning. In regions with declining soil selenium, plants may convert selenium into bioavailable forms, leading to chronic selenium toxicity in animals through the food chain[43]. As early as 1295, it was recorded in the Book of Marco Polo that in the ancient Suzhou (now Hexi Corridor) area in China, horses were found to have symptoms of hair removal and armor removal, which is generally believed in the academic community to be the earliest recorded selenium poisoning in humans[37]. Some researchers have hypothesized that the toxicity observed in certain Oxytropis species within China’s Hexi Corridor region may be attributed to excessive selenium accumulation. However, analytical data show that selenium content in Oxytropis glabra ranges from 0.052−0.174 ppm, and Oxytropis ochrocephala contains 0.058−0.078 ppm, both below the risk threshold for animal selenium poisoning[44,45]. Thus, the toxicity of Astragalus and Oxytropis species is unlikely attributable to high selenium levels.
In this study, the selenium content in plants was determined by ICP-MS. The selenium content in healthy A.adsurgens ranged from 192.42 to 366.61 μg/kg, whereas that in plants affected by yellow stunt and root rot ranged from 747.78 to 1108.95 μg/kg. Therefore, the occurrence of disease can lead to an increase in selenium content in A.adsurgens. The total selenium content in the surface soil of Lanzhou City is 0.179±0.066 mg/kg, which is at a moderate level compared to the national soil selenium content; Meanwhile, the soil selenium content in Yuzhong County is relatively high. In Heping Town, where the experimental site is located, the soil selenium content is 0.157±0.021 mg/kg[46,47]. According to the comparison between the selenium content in soil and that in A.adsurgens, plants have a certain enrichment effect on selenium, and the enrichment effect is more pronounced after infection with yellow stunt and root rot. However, the mechanism by which diseases increase the content of mineral elements in plants is still unclear.
We found the reason for the increase in selenium content in diseased plants may also be related to the decrease in dry weight of plants after disease onset. In the diseased plants, the dry weight of healthy plants per plant is approximately 67.20 g, while the average dry weight of all dwarf plants per plant is 51.24 g, a decrease of 23.75% compared to before. The single weight of plants with partially stunted branches was 46.30 g, a decrease of 31.10% compared to before[12]. The decrease in dry weight per plant leads to an increase in the proportion of selenium, but this conclusion still needs further verification.
When the selenium content in feed exceeds 5 ppm, long-term consumption can cause chronic poisoning. At 10−25 ppm, it can cause acute poisoning in most animals[43]. The selenium content in healthy plants did not reach the level of animal poisoning. After being infection with yellow stunt and root rot, the selenium content in A.adsurgens increases, with the highest exceeding 1 ppm. However, the A.adsurgens with this selenium content will not cause poisoning in animals after they consume it.
4.2. Alterations in the Concentrations of Other Mineral Elements in Diseased A.adsurgens
After infection with yellow stunt and root rot, other elements in A.adsurgens showed little change, such as manganese content. The calcium content decreased, but the difference was not significant. The contents of iron, zinc, sodium and magnesium all decreased significantly (P<0.05). After studying the nutritional substances of infected and healthy plants, it was found that the content of soluble sugars and calcium in plants significantly decreased (P<0.05) after infection12. The decrease in the content of most mineral elements may be related to the root rot of A.adsurgens. After the disease, the ability of plants roots to absorb mineral nutrients from the soil decreases, and the mineral element content in the aboveground part also decreases as a result.
4.3. The Influence of Changes in Swainsonine Content of Diseased Astragalus adsurgens on Livestock and Its Correlation with Mineral Elements
Swainsonine was detected in A.adsurgens and A.gansuense infected with yellow stunt and root rot. The content of swainsonine in healthy A.adsurgens was 0.002−0.003%, while that in diseased plants was 0.008−0.013%, and the content in A.gansuense was approximately 0.006%[48]. The swainsonine content in healthy A.adsurgens was 0.002%−0.003%, while that in diseased plants was 0.008%−0.013%, and the content in A.gansuense was approximately 0.006%. This is consistent with the results of previous studies that proved the existence of the β-ketoacyl synthase (KS) gene in A.adsurgens and detected the production of swainsonine[49].
The content of swainsonine in locoweed varies from less than the detection limit (0.001%) to more than 0.2%. The difference in content is related to a variety of factors. The content of swainsonine varies greatly between different locoweed species, such as the average content of swainsonine in A.adsurgens is less than 0.001%, while the content of A.mollissimus is higher than 0.1%. There are differences in swainsonine content between different populations of the same locoweed species due to environmental factors[50,51]; The content of the same locoweed in the same habitat is different in different plants; The content of swainsonine in different parts of the same plant is also different. The root contains less swainsonine than the stem and leaf. For example, the content of swainsonine in the aboveground part of Oxytropis sericea is nearly 10 times higher than that in the underground part. The content of swainsonine in different parts of plants at different growth stages will also change[52]. These distribution rules can provide an important basis for further study of swainsonine in A.adsurgens.
A.adsurgens containing swainsonine is similar to locoweeds and can cause animal poisoning. Locoweed feeding trials have shown that when sheep were fed with powder of O.kansuensis at a daily dose of 8−10 g per kilogram of body weight, all sheep showed poisoning symptoms within 20−25 days of the experiment and died successively after 56 days[53]. However, when the same O.kansuensis and dose were used but administered once every two weeks, the sheep did not develop poisoning symptoms[54]. The swainsonine content in these O.kansuensis plants was approximately 0.021%. The swainsonine content in O.ochrocephala is slightly lower, at 0.012%, and the symptoms of poisoning in sheep during feeding trials were relatively milder[53,55,56]. When the swainsonine content in locoweed reaches 0.001% or higher, livestock consuming 2% of their body weight in locoweed daily will be poisoned. Low swainsonine content or reduced intake may lead to weight loss but will not cause neurological poisoning[15,52]. Moreover, previous studies have proven that A.adsurgens infected by A.gansuense is toxic to mice, and the infection can increase its toxicity[57]. A.adsurgens severely affected by yellow stunt and root rot may contain 0.001% swainsonine. However, diseased A.adsurgens are stunted, with reduced biomass and poor palatability. Therefore, under natural conditions, it is unlikely that livestock will consume only diseased A.adsurgens. The likelihood of severe poisoning in livestock is relatively low. Additionally, when feeding livestock with healthy A.adsurgens, the amount should also be appropriate.
In this study, it was found that there is a significant negative correlation between the content of swainsonine and the content of sodium element (Figure 6E), while there is an extremely significant positive correlation between the content of swainsonine and the content of selenium element (Figure 6F). Therefore, we believe that the increase in the content of swainsonine may be related to the decrease of sodium element in the A.adsurgens, and it is closely associated with the increase of selenium element. Whether the causes of the disease and the increase in toxins are directly related to the amounts of these two elements still requires further exploration in the future.
5. Conclusions
In this study, we investigated the toxic substances (selenium and swainsonine) in the A.adsurgens infected by A.gansuense for the first time, and preliminarily concluded that the soil selenium enriched in infected A.adsurgens cannot cause animal poisoning. However, in the case of continuous feeding for multiple days, swainsonine in A.adsurgens can easily cause animal poisoning. Therefore, the main reason for the pathological changes in mice during the feeding experiment may be swainsonine. A.adsurgens with yellow stunt and root rot may contain more than 0.001% of swainsonine, which can easily lead to poisoning when animals feed on it for a long time. Therefore, caution should be exercised when using infected A.adsurgens to feed livestock in animal husbandry production. Our paper provides guidance for the subsequent feeding of livestock with A.adsurgens. In addition, the contents of the mineral elements sodium and selenium discovered in our study are closely related to the amount of swainsonine, which may also provide reference value for the subsequent prevention and control of yellow stunt and root rot.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Huaqi Liu: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing-original draft, Writing-review & editing. Yanzhong Li: Conceptualization, Data curation, Software, Formal analysis, Investigation, Methodology, Validation, Writing-original draft. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by National Forestry and Grassland Administration (20220104) and the Earmarked Fund for CARS (CARS-34).
Acknowledgments
We would like to express our gratitude to the Heping Farm of Gansu Vocational College of Agriculture for providing the experimental plots. In addition, we also appreciate the determination of swainsonine content and the measurement of mineral elements provided by the Testing Center of North Minzu University and Gansu University of Chinese Medicine.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflict interest.
References
- Sun, Q.Z.; Gui, R.; Na, R.S.; Nada, M.D.; Zhai, Z.H. Study on productivity of Astragalus adsurgens Pall. with different growth periods. Grassland of China 1999, 5, 30–35. [Google Scholar]
- Wang, J.; Lei, Z.Y.; Feng, X.Q. Gas Chromatographic Determination of Trace 3-Nitropropionic Acid in Astragalus adsurgens. Grassland of China 1992, 3, 69–72. [Google Scholar]
- Su, S.F. Revisiting the Forage Utilization Issues of Astragalus adsurgens. Pratacultural Science 1987, 2, 3–9. [Google Scholar]
- Cook, D.; Ralphs, M.; Welch, K.; Stegelmeier, B. Locoweed poisoning in livestock. Rangelands 2009, 31, 16–21. [Google Scholar] [CrossRef]
- Zhang, G.G.; Dong, S.T.; Yang, Z.B.; Yang, W.R.; Jiang, S.Z. Experiment on Feeding Small-tailed Han Sheep with Astragalus adsurgens. Pratacultural Science 2008, 3, 74–77. [Google Scholar]
- Li, Y.Z.; Nan, Z.B. Symptomology and etiology of a new disease, yellow stunt, and root rot of standing milkvetch caused by Embellisia sp. in Northern China. Mycopathologia 2007, 163, 327–334. [Google Scholar] [CrossRef]
- Li, Y.Z.; Nan, Z.B.; Hou, F.J. The roles of an Embellisia sp. causing yellow stunt and root rot of Astragalus adsurgens and other fungi in the decline of legume pastures in northern China. Australasian Plant Pathology 2007, 36, 397–402. [Google Scholar] [CrossRef]
- Liu, J.L. Study on molecular biology of Embellisia astragali. Ph.D. Thesis, Lanzhou University, Lanzhou, China, 2016. [Google Scholar]
- Li, Y.Z.; Nan, Z.B. A new species, Embellisia astragali sp. nov., causing standing milk-vetch disease in China. Mycologia 2007, 99, 406–411. [Google Scholar] [CrossRef]
- Woudenberg, J.H.C.; Groenewald, J.Z.; Binder, M.; Crous, P.W. Alternaria redefined. Studies in mycology 2013, 75, 171–212. [Google Scholar] [CrossRef]
- Liu, J.L.; Li, Y.Z.; Creamer, R. A re-examination of the taxonomic status of Embellisia astragali. Current Microbiology 2016, 72, 404–409. [Google Scholar] [CrossRef]
- Li, Y.Z. Study of yellow stunt and root rot (Embellisia astragali sp. nov. Li & Nan) of Astragalus adsurgens. Ph.D. Thesis, Lanzhou University, Lanzhou, China, 2007. [Google Scholar]
- Li, Y.Z.; Creamer, R.; Baucom, D.; Nan, Z.B. Pathogenic Embellisia astragali on Astragalus adsurgens is very closely related to locoweed endophyte. Phytopathology 2011, 101, S102–S103. [Google Scholar]
- James, L.F.; Hartley, W.J.; Kampen, K.R.V. Syndromes of astragalus poisoning in livestock. Journal of the American Veterinary Medical Association 1981, 178, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Stegelmeier, B.L.; James, L.F.; Panter, K.E.; Gardner, D.R.; Pfister, J.A.; Ralphs, M.H.; Molyneux, R.J. Dose response of sheep poisoned with locoweed (Oxytropis sericea). Journal of Veterinary Diagnostic Investigation 1999, 11, 448–456. [Google Scholar] [CrossRef]
- Williams, M.C.; James, L.F. Toxicity of nitro-containing Astragalus to sheep and chicks. Journal of Range Management 1975, 28, 260–263. [Google Scholar] [CrossRef]
- Wu, W.H. Plant Biology, 2th ed.Science: Beijing, China, 2008. [Google Scholar]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.M.; Bakker, P.A.H.M. Induced Systemic Resistance by Beneficial Microbes. Annual Review of Phytopathology 2014, 52, 347–375. [Google Scholar] [CrossRef] [PubMed]
- Lazar, T.; Taiz, L. and Zeiger, E. Plant physiology. 3rd edn. Annals of Botany 2003, 91, 750–751. [Google Scholar] [CrossRef]
- Guo, Y.Y.; Xu, K.; Wang, X.F.; Zhu, Y.H. Research progress on mineral nutrition and plant disease mechanism. Journal of Gansu Agricultural University 2003, (4), 385–393. [Google Scholar]
- Marschner, H. Marschner’s mineral nutrition of higher plants. Science Press: Beijing, China, 2013. [Google Scholar]
- Tian, Y.Q.; Huang, L.P.; Zhang, Z. Research Progress of the Relationship Between Mineral Nutrients Deficiency orImbalance and Plant Disease. Chinese Agricultural Science Bulletin 2016, 32, 174–176. [Google Scholar]
- Fidanza, M.A.; Dernoeden, P.H. Interaction of nitrogen source, application timing, and fungicide on Rhizoctonia blight in ryegrass. Hortscience A Publication of the American Society for Horticultural Science 1996, 31, 389–392. [Google Scholar] [CrossRef]
- Bhaskar, C.V.; Rao, G.R.; Reddy, K.B. Effect of nitrogen and potassium nutrition on sheath rot incidence and phenol content in rice(Oryz asativa L.). Indian Journal of Plant Physiology 2001, 16, 254–257. [Google Scholar]
- Sharma, S.R.; Kolte, S.J. Effect of soil-applied NPK fertilizers on severity of black spot disease (Alternaria brassicae) and yield of oilseed rape. plant and soil 1994, 167, 313–320. [Google Scholar] [CrossRef]
- Liu, X.Y.; He, P.; Jin, J.Y. Advances in effect of potassium nutrition on plant disease resistance and its mechanism. Plant Nutrition and Fertilizer Science 2006, 12, 445–450. [Google Scholar] [CrossRef]
- Mu, K.G.; Zhao, X.Q.; Li, J.Q.; Liu, X.L. Progressing on the Relation between Mineral Nutrients and Plant Disease. Journal of China Agricultural University 2000, 5, 84–90. [Google Scholar]
- Conway, W.S.; Sams, C.E.; Kelman, A. Enhancing the natural resistance of plant tissues to postharvest diseases through calcium applications. HortScience 1994, 29, 66–69. [Google Scholar] [CrossRef]
- Huber, D.M.; Jones, J.B. The role of magnesium in plant disease. Plant and soil 2013, 368, 73–85. [Google Scholar] [CrossRef]
- Bhargava, A.K.; Singh, R.D. Effect of Nitrogenous Fertilizers and Trace Elements on the Severity of Alternaria Blight of Bottle Gound. Annals of Arid Zone 1992, 31, 63–64. [Google Scholar]
- Gadi, B.; Jeffrey, G. Copper as a biocidal tool. Current Medicinal Chemistry 2005, 12, 2163–2175. [Google Scholar] [CrossRef]
- Sharma, A.K.; Sawant, I.S.; Sawant, S.D.; Saha, S.; Kadam, P.; Somkuwar, R.G. Aqueous chlorine dioxide for the management of powdery mildew vis-a-vis maintaining quality of grapes and raisins. Journal of Eco-friendly Agriculture 2017, 12, 59–64. [Google Scholar]
- Ehret, D.L.; Utkhede, R.S.; Frey, B.; Menzies, J.G.; Bogdanoff, C. Foliar applications of fertilizer salts inhibit powdery mildew on tomato. Canadian Journal of Plant Pathology 2002, 24, 437–444. [Google Scholar] [CrossRef]
- Panigati, M.; Falciola, L.; Mussini, P.; Beretta, G.; Facino, R.M. Determination of selenium in Italian rices by differential pulse cathodic stripping voltammetry. Food Chemistry 2007, 105, 1091–1098. [Google Scholar] [CrossRef]
- Schwarz, K.; Bieri, J.G.; Briggs, G.M.; Scott, M.L. Prevention of Exudative Diathesis in Chicks by Factor 3 and Selenium. Proceedings of the Society for Experimental Biology & Medicine 1957, 95, 621. [Google Scholar] [CrossRef]
- Zhao, G.Q.; Yin, Z.F.; Wei, L.H.; Wang, Y.J.; Yang, X.Y. The duality of selenium and its related health issues. Journal of chengde medical college 1999, 1, 92–93. [Google Scholar] [CrossRef]
- Wu, Y.Y.; Peng, Z.K.; Luo, Z.M. The multiple biological functions of selenium and its impact on human and animal health. Journal of Hunan Agricultural University 1997, 23, 294–300. [Google Scholar] [CrossRef]
- Rayman, M.P.; Margaret, P. Selenium and human health. The Lancet 2012, 379, 1256–1268. [Google Scholar] [CrossRef]
- O’Toole, D.; Raisbeck, M.F. Pathology of experimentally induced chronic selenosis (alkali disease) in yearling cattle. Journal of Veterinary diagnostic investigation 1995, 7, 364–373. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.X.; Zhou, W.S.; Guo, S.H. Advances of Studies on Microelement Selenium in Plants. Journal of Anhui Agricultural Sciences 2009, 37, 5844–5845. [Google Scholar] [CrossRef]
- Brown, T.A.; Shrift, A. Selenium: toxicity and tolerance in higher plants. Biological Reviews 1982, 57, 59–84. [Google Scholar] [CrossRef]
- Wu, J.; Liu, X.F.; Xu, H.S. Functions of Selenium in Plants. Plant Physiology Journal 1999, 35, 417–423. [Google Scholar] [CrossRef]
- Wang, J.H. Selenosis. Chinese Journal of Veterinary Science 1993, 13, 401–408. [Google Scholar] [CrossRef]
- Shao, S.X.; Zheng, B.S.; Zhao, C.Z.; Yin, C.Q.; Lang, Y.B.; Zhang, A.L. Study on Selenium in Poisonous Oxytropis Plants (Locoweed) from the Hexi Corridor and Its Association with Livestock Poisoning. Advances in Earth Science 2004, 19, 502–505. [Google Scholar] [CrossRef]
- Shao, S.X.; Zheng, B.S.; Wang, M.S.; Li, X.Y.; Liu, X.J.; Ling, H.W.; Luo, C. Environmental Geochemistry of Selenium in the Hexi Corridor Region and Investigation into the Causes of Livestock Poisoning by Toxic Plants. Acta Mineralogica Sinica 2006, 26, 448–452. [Google Scholar] [CrossRef]
- Li, W.; Li, F.; Bi, D.; Wang, H.K.; Li, H.Y.; Zhou, S.B.; Zhao, Q.G.; Yin, X.B. Distribution and Speciation of Selenium in Alkaline Soils and Agricultural Products of Lanzhou. Soils 2012, 44, 632–638. [Google Scholar] [CrossRef]
- Xie, C.J.; Li, H.Y.; Wang, G.Y.; Wang, P. Analysis of Selenium Speciation in Soils and Key Agricultural Products from Lanzhou City. Gansu Science and Technology Information 2016, 45, 89–91. [Google Scholar]
- Shi, M.; Li, Y.Z. Alternaria gansuense, a Plant Systematic Fungal Pathogen Producing Swainsonine in Vivo and in Vitro. Current Microbiology 2023, 80, 232. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.H.; Zhai, X.H.; Li, Q.F.; Wang, J.H.; Geng, G.X. Determination of Swainsonine in Astragalus locoweed by internal standard gas chromatography. Journal of Northwest Sci-Tech University of Agriculture and Forestry 2008, 36, 17–21. [Google Scholar] [CrossRef]
- Cook, D.; Gardner, D.R.; Pfister, J.A. Swainsonine-containing plants and their relationship to endophytic fungi. Journal of Agricultural and Food Chemistry 2014, 62, 7326–7334. [Google Scholar] [CrossRef]
- Cook, D.; Gardner, D.R.; Lee, S.T.; Pfister, J.A.; Stonecipher, C.A.; Welsh, S.L. A swainsonine survey of North American Astragalus and Oxytropis taxa implicated as locoweeds. Toxicon 2016, 118, 104–111. [Google Scholar] [CrossRef]
- Cook, D.; Gardner, D.R.; Ralphs, M.H.; Pfister, J.A.; Welch, K.D.; Green, B.T. Swainsoninine concentrations and endophyte amounts of Undifilum oxytropis in different plant parts of Oxytropis sericea. Journal of chemical ecology 2009, 35, 1272–1278. [Google Scholar] [CrossRef]
- Wang, K.; He, B.X. A study on the toxicity of Oxytropis kansuonisis in sheep. Chinese Journal of Veterinary Medicine 1995, 21, 5–6. [Google Scholar]
- Wang, K.; Mo, C.H.; Zhao, B.Y.; Cao, G.R. Study on the Toxicity of Intermittent Feeding of Oxytropis kansuensis to Sheep. Animal Husbandry & Veterinary Medicine 1998, 30, 29–30. [Google Scholar]
- Gu, B.Q.; Xue, D.M.; Cao, G.R.; Duan, D.X. Pathological Observations of Oxytropis kansuensis Poisoning in Goats. Journal of Animal Science and Veterinary Medicine 1991, 3, 9–11. [Google Scholar]
- Li, Q.F.; Wang, J.H.; Yuan, Y.; Qi, X.R.; Zhao, Y. Pathology of Oxytropis glacialis Poisoning in Goats. Chinese Journal of Veterinary Science 2001, 21, 75–78. [Google Scholar] [CrossRef]
- Yang, B.; Nan, Z.B.; Li, Y.Z. Toxicity of standing milkvetch infected with Alternaria gansuense in white mice. Frontiers in Veterinary Science 2025, 11, 1477970. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Location of the sampling sites. (A) The experimental field of A.adsurgens is located at the Heping Campus of Gansu Agricultural Vocational and Technical College in Yuzhong County, Lanzhou City, Gansu Province, China. (B) Plant material samples of A.adsurgens (Left: Healthy branches; Right: Diseased branches). (C) The experimental field of 4-year-old A.adsurgens.
Figure 1.
Location of the sampling sites. (A) The experimental field of A.adsurgens is located at the Heping Campus of Gansu Agricultural Vocational and Technical College in Yuzhong County, Lanzhou City, Gansu Province, China. (B) Plant material samples of A.adsurgens (Left: Healthy branches; Right: Diseased branches). (C) The experimental field of 4-year-old A.adsurgens.
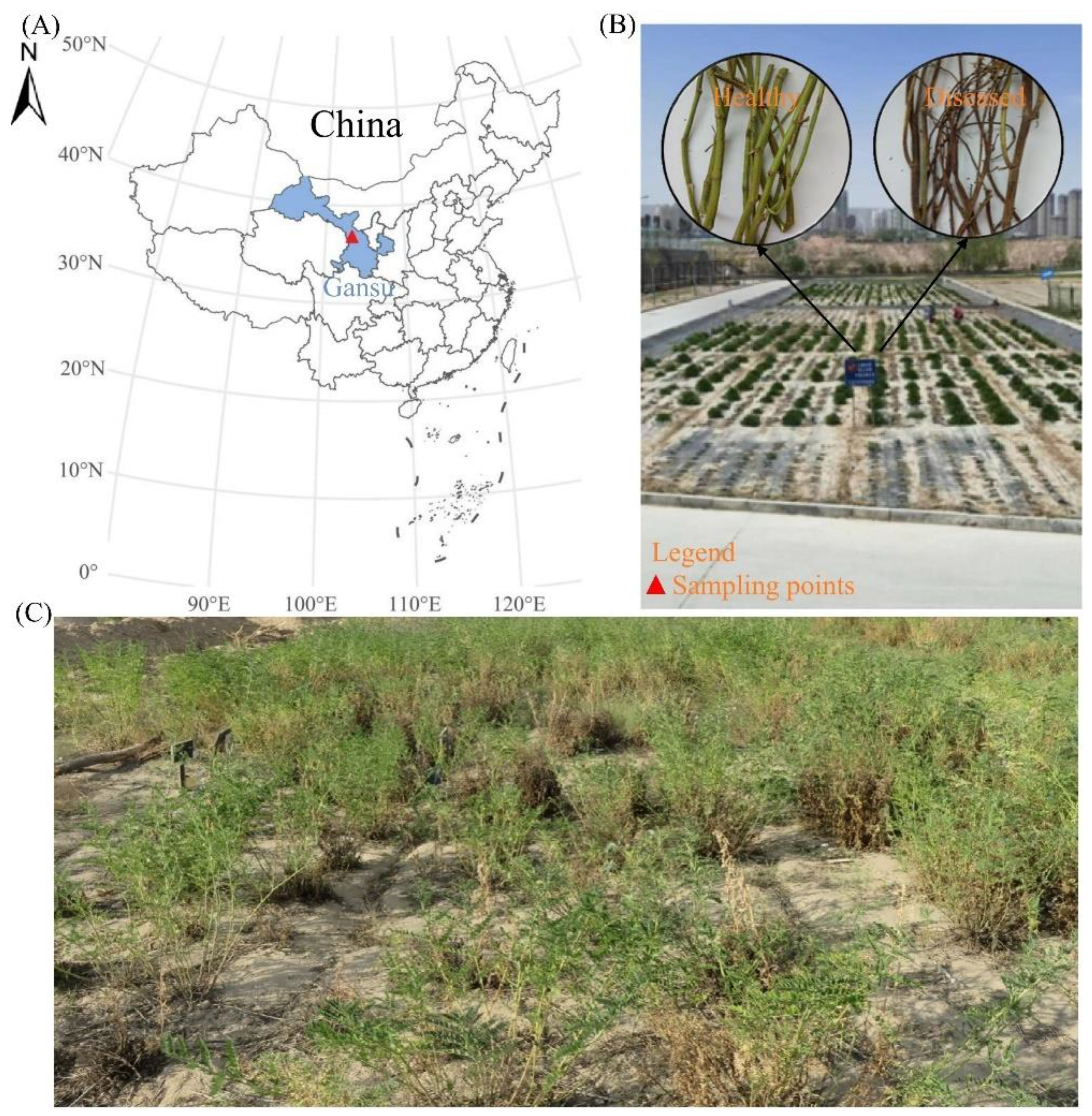
Figure 2.
The differences in selenium content between healthy plants and diseased plants among different varieties of A.adsurgens. Those marked with *** in the figure indicate that there are extremely significant differences in the selenium content between the diseased and healthy plants of the same variety (P < 0.001).
Figure 2.
The differences in selenium content between healthy plants and diseased plants among different varieties of A.adsurgens. Those marked with *** in the figure indicate that there are extremely significant differences in the selenium content between the diseased and healthy plants of the same variety (P < 0.001).
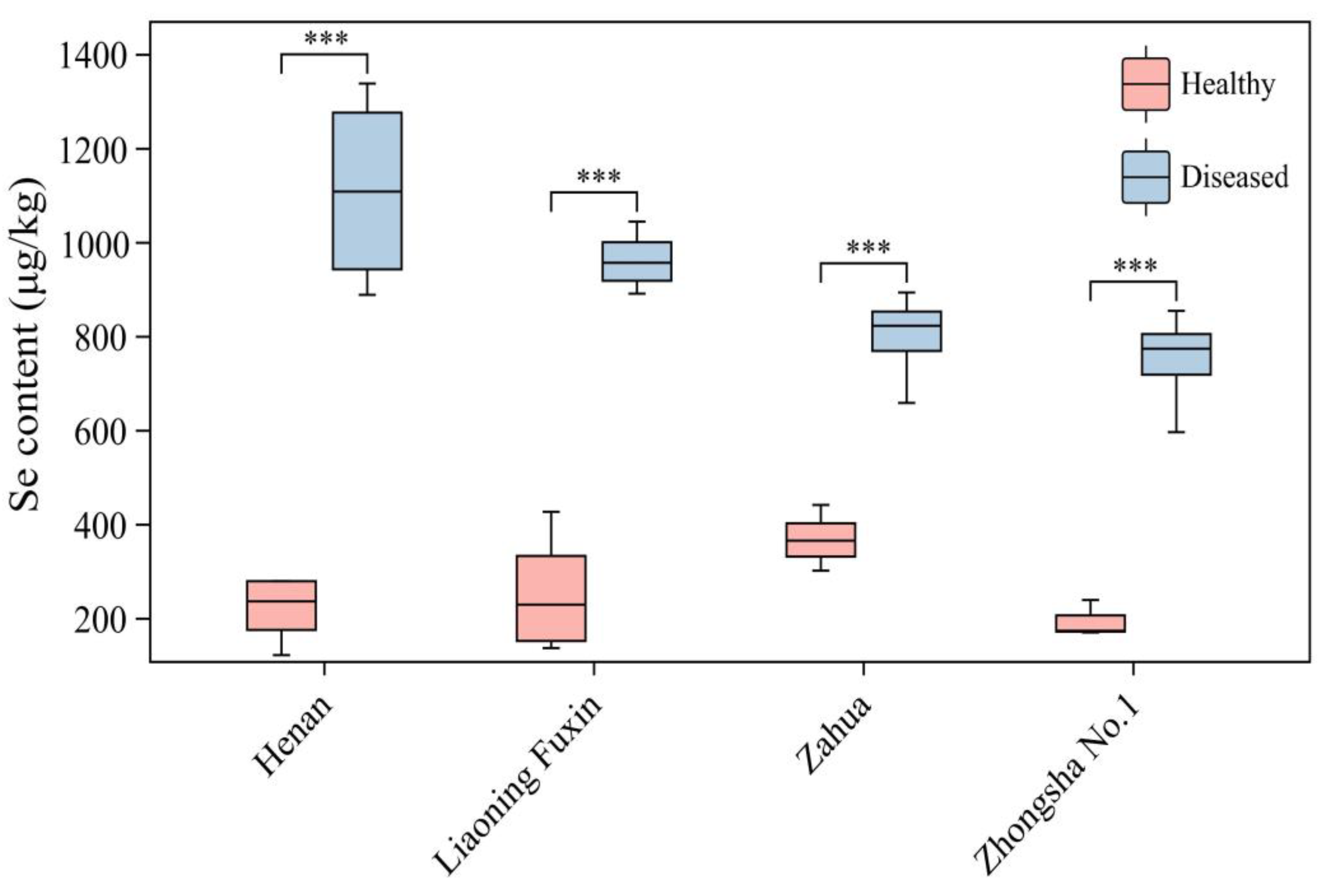
Figure 3.
Comparison of the contents of mineral elements between healthy plants and diseased plants of different varieties of A.adsurgens. (A) The differences in Ca, (B) Fe, (C) Mg, (D) Mn, (E) Na and (F) Zn content between healthy plants and diseased plants of same variety. Different lowercase letters represent significant differences in the elemental content between healthy plants and diseased plants.
Figure 3.
Comparison of the contents of mineral elements between healthy plants and diseased plants of different varieties of A.adsurgens. (A) The differences in Ca, (B) Fe, (C) Mg, (D) Mn, (E) Na and (F) Zn content between healthy plants and diseased plants of same variety. Different lowercase letters represent significant differences in the elemental content between healthy plants and diseased plants.
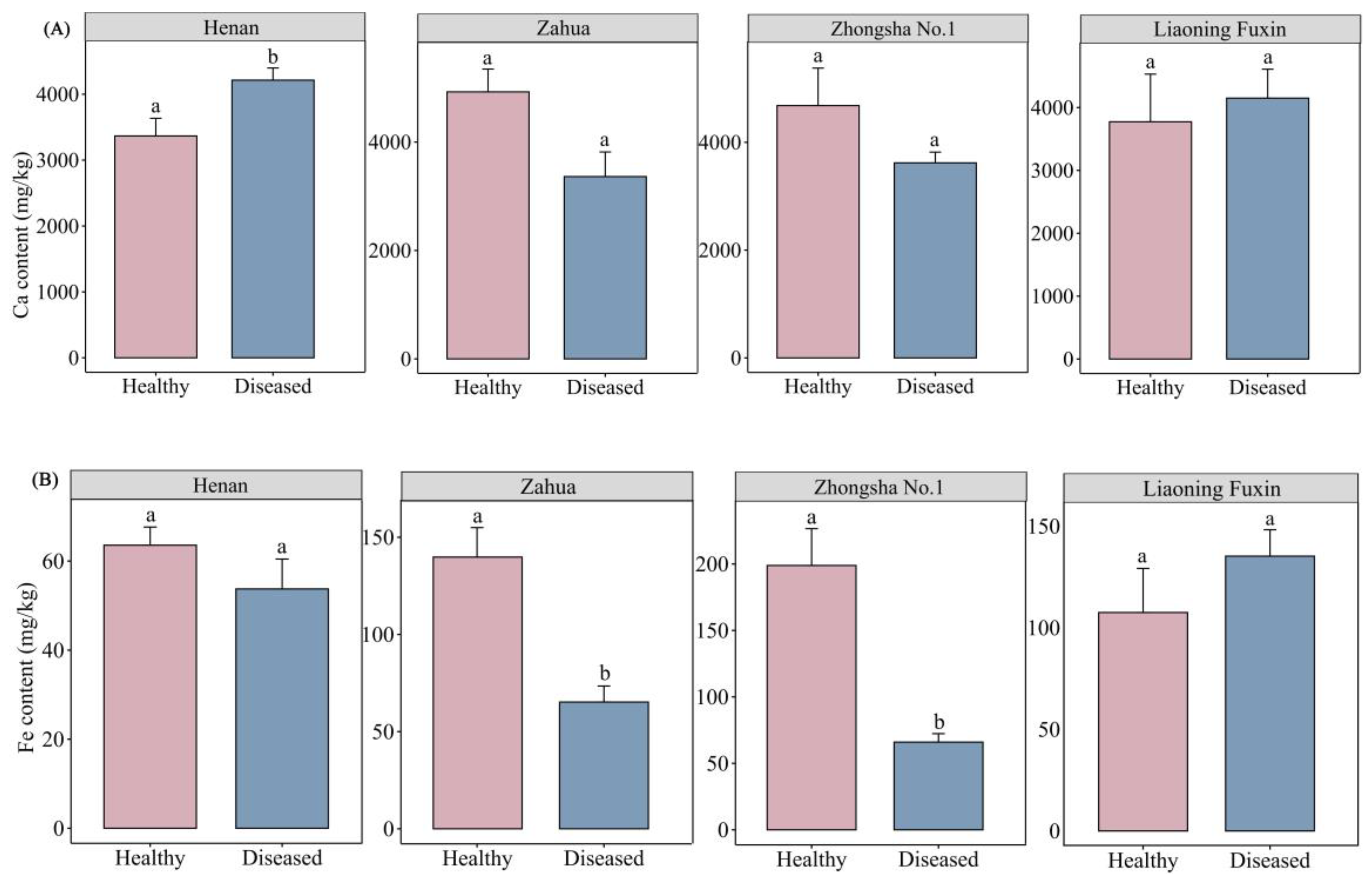
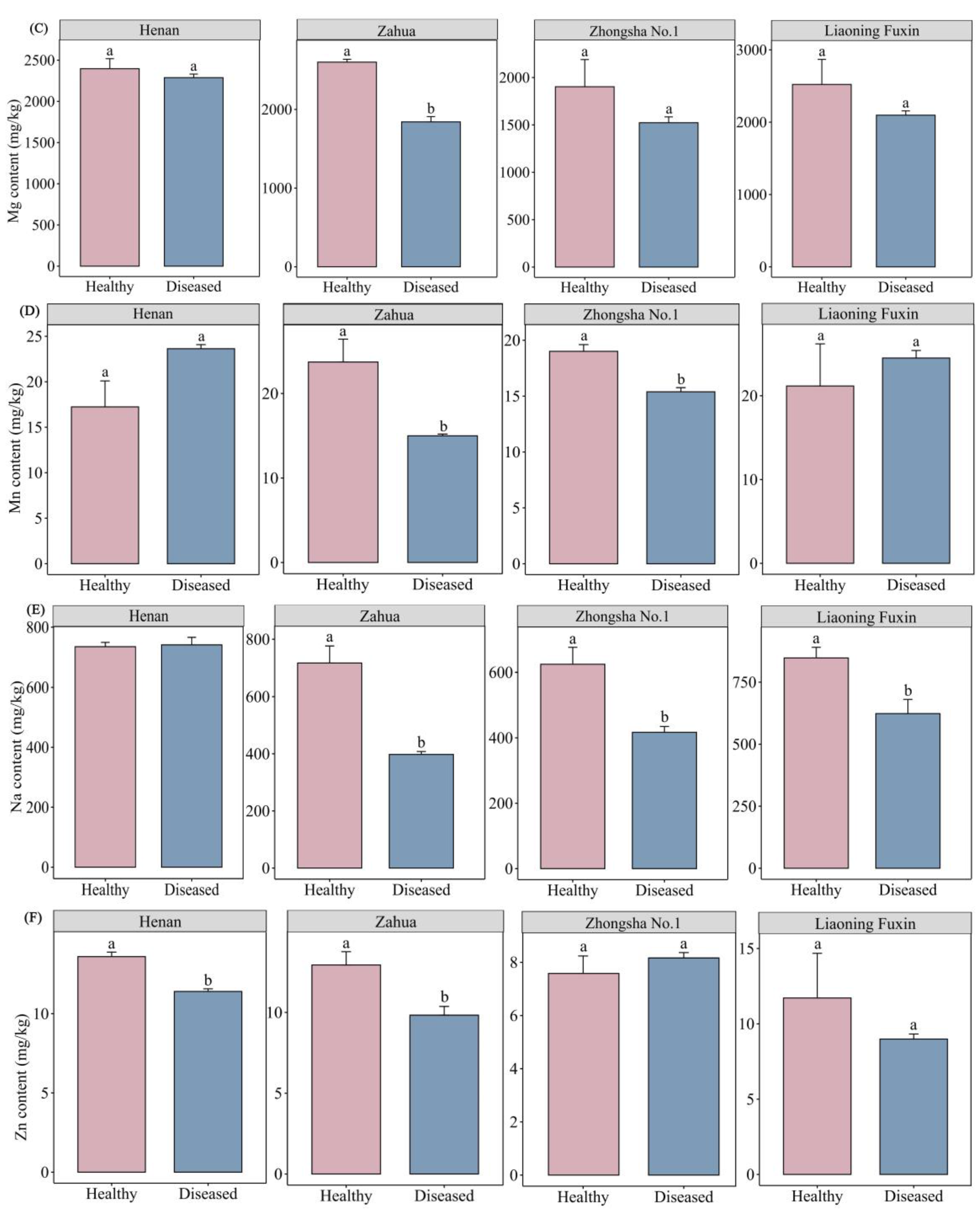
Figure 4.
Comparison of the contents of mineral elements between healthy plants and diseased plants of A.adsurgens of all varieties. (A) The total contents of Zn, Fe and Mn (B) Na, Ca and Mg (C) Se in all varieties of A.adsurgens, as well as the content differences between healthy plants and diseased plants. Significance level: ***P<0.001; **P< 0.01; *P< 0.05.
Figure 4.
Comparison of the contents of mineral elements between healthy plants and diseased plants of A.adsurgens of all varieties. (A) The total contents of Zn, Fe and Mn (B) Na, Ca and Mg (C) Se in all varieties of A.adsurgens, as well as the content differences between healthy plants and diseased plants. Significance level: ***P<0.001; **P< 0.01; *P< 0.05.
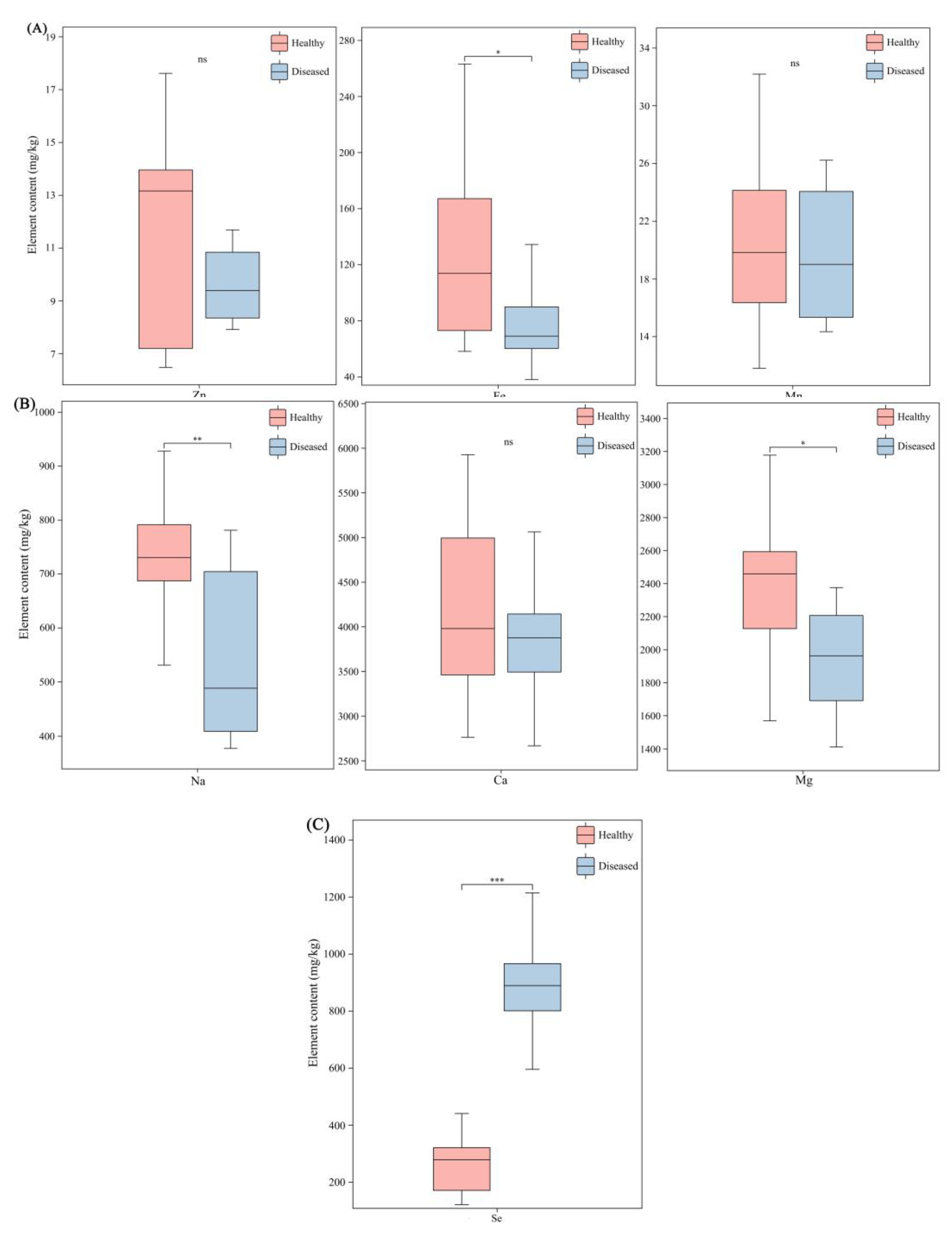
Figure 5.
Comparison of the swainsonine content between healthy plants and diseased plants of different varieties of A.adsurgens. (A) The content of swainsonine in healthy plants and diseased plants of the mixed varieties, (B) Zhongsha No.1 and (C) Henan. In the figure, plants with the same letters show no significant difference, while those with different letters have significant differences (P<0.05).
Figure 5.
Comparison of the swainsonine content between healthy plants and diseased plants of different varieties of A.adsurgens. (A) The content of swainsonine in healthy plants and diseased plants of the mixed varieties, (B) Zhongsha No.1 and (C) Henan. In the figure, plants with the same letters show no significant difference, while those with different letters have significant differences (P<0.05).
Figure 6.
The correlation between swainsonine and elements. (A) The correlation between Ca (B) Fe, (C) Mg, (D) Mn, (E) Na, (F) Se and (G) Zn element content and swainsonine. Significance level: P<0.001 indicates an extremely significant difference; P< 0.05 indicates significance.
Figure 6.
The correlation between swainsonine and elements. (A) The correlation between Ca (B) Fe, (C) Mg, (D) Mn, (E) Na, (F) Se and (G) Zn element content and swainsonine. Significance level: P<0.001 indicates an extremely significant difference; P< 0.05 indicates significance.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



