Submitted:
15 May 2025
Posted:
16 May 2025
You are already at the latest version
Abstract
Kisspeptin, the product of the KISS1 gene and a ligand of the G-protein coupled receptor, KISS1R (also known as GPR54), is a relatively well-known hypothalamic neurohormone, the secretion of which is intrinsically linked to the kisspeptin, neurokinin B, and dynorphin (KNDy) neurons and is involved in the neuromodulation of gonadotropin-releasing hormone (GnRH) secretion, playing a key role in the central mechanism controlling the hypothalamic‒pituitary‒gonadal (HPG) axis. In contrast to our knowledge of the importance of kisspeptin neuronal signaling in regulating the onset of puberty and the control of fertility through its effects on the menstrual cycle, the importance of the extraneuronal pool of this hormone after fertilization and during early human pregnancy is far less complete. The very significant increase in the serum extrahypothalamic kisspeptin concentration in the first trimester of pregnancy, which originates from peripheral reproductive tissues, is accompanied by changes in local KISS1R expression. These findings suggest an important role of kisspeptin in the regulation of embryo implantation, trophoblast invasion, placentation and early pregnancy. The aim of this review is to present the current state of knowledge regarding the involvement of kisspeptin signaling in the physiology and pathophysiology of early pregnancy in humans. The diagnostic value of kisspeptin levels for early pregnancy outcomes will also be explored to identify potential related therapeutic targets.
Keywords:
kisspeptin
; KISS-1 gene
; kisspeptin receptor (KISS1R)
; G-protein coupled receptor 54 (GPR54)
; kisspeptin signaling
; early pregnancy
; kisspeptin (KISS)/neurοkinin B (NKB)/Dynοrphin (Dyn) neurοns
; KNDy neurons
1. Introduction
Neuroendocrine control of reproduction refers to the coordinated processes in the hypothalamus and pituitary gland that govern the production of hormones [1]. Steroid sex hormones are the main products secreted by the gonads (ovaries or testicles), although they may also be produced (including conversion from other steroids) outside the gonads, namely, in the adrenal glands, liver, brain, adipose tissue, or placenta during pregnancy [2,3,4]. The neuromodulation of gonadotropin-releasing hormone (GnRH) in the hypothalamus, manifested by its pulsatile secretion, is a key phenomenon within the hypothalamic–pituitary–gonadal (HPG) axis and determines the pattern of secretion of gonadotropins, follicle-stimulating hormone (FSH) and luteinizing hormone (LH) from the anterior pituitary [5,6]. Therefore, the HPG axis is involved in the onset of puberty, gamete maturation in the gonads, menstrual cyclicity, sex drive, childbirth, milk production and lactation [7,8]. At the beginning of the 21st century, research revealed that the protein that was subsequently named after Hershey's chocolate kisses (Pennsylvania – the place of kisspeptin discovery is the home of the “Hershey’s Kisses” sweets), kisspeptin, has the ability to stimulate GnRH, which has received increased interest [9,10]. Kisspeptin is a product of the KISS1 gene and a ligand of the G-protein coupled receptor, KISS1R (also known as GPR54). Previously, kisspeptin was named metastin because, in 1996, KISS1 was identified as a human metastasis suppressor gene, the increased expression of which inhibits the metastasis of melanoma and breast cancer [11,12,13].
Confirmation of the involvement of kisspeptin in the neuroendocrine modulation of reproductive functions as a specific gatekeeper before entering the HPG axis was the starting point for numerous studies in the fields of physiology and pathophysiology [14,15,16,17,18]. Notably, research on the key role of kisspeptin in regulating the onset of puberty and modulating the activity of the HPG axis and, consequently, in the control of fertility is much more advanced than that examining the importance of kisspeptin after conception and the start of pregnancy. This disproportion is not justified in light of the results of recent studies indicating the presence of kisspeptin signaling via KISS1R outside the HPG axis [19,20]. Variable concentrations of extrahypothalamic kisspeptin in peripheral reproductive tissues, accompanied by changes in local KISS1R expression, may suggest an important role of kisspeptin in the regulation of trophoblast invasion, embryo implantation, placentation and early pregnancy [21,22,23,24,25]. Therefore, the aim of this review is to present the current state of knowledge regarding the involvement of kisspeptin signaling in the physiology and pathophysiology of early pregnancy in humans.
2. Kisspeptin
2.1. Structure
Human KISS1 is located on the long arm of chromosome 1 (1q32) and contains 4 exons [26]. The product encoded by KISS1, prepro-kisspeptin, is a 145 amino acid precursor peptide that is cleaved into four shorter peptides called, according to the number of amino acids in the molecule, kisspeptin-54 (also known as metastin), kisspeptin-14, kisspeptin-13 and kisspeptin-10. All these isoforms contain a conserved 10 amino acid core sequence at the C-terminus, containing an Arg-Phe-NH2 (RF-NH2) motif that is required for full activation of KISS1R [10,27] (see Figure 1).
The major circulating and tissue form of kisspeptin is a 54-residue fragment (Kp-54), although shorter fragments (kisspeptins 14,13 and 10) appear to be as potent in binding to KISS1R as Kp-54 is [15,29]. Polymorphisms of both KISS1 and KISS1R are associated with central precocious puberty (CPP) and idiopathic hypogonadotropic hypogonadism (IHH) [30]. With respect to KISS1, at least 14 mutations have been detected that contribute to the disruption of the amino acid sequence in the molecules of these gene products (kisspeptins) [30,31].
Owing to the existence of 4 isoforms of kisspeptin, precise assessment of its concentration using commercially available techniques is difficult [32]. There are significant differences in the detection range, sensitivity, and, above all, specificity between the individual assay methods, which may cause uncertainty as to which form of kisspeptin is detected (i.e., Kp-54, Kp-14, Kp-13, or Kp-10) [33,34]. Moreover, the ambiguous or contradictory results of the kisspeptin assay may be influenced by the varying rate of kisspeptin metabolism in the circulatory system, where the half-life of Kp-54 is approximately 30 min, whereas the half-life of Kp-10 does not exceed 4 min [35].
2.2. Sources of Kisspeptin in the Body
2.2.1. Neuronal Kisspeptin in the Central Nervous System (CNS)
The localization of kisspeptin-expressing cells in the CNS has been most fully elucidated in mice. In the mouse hypothalamus, this neuropeptide showed the strongest expression, similar to its mRNA expression, in nuclei involved in the regulation of GnRH and gonadotropin (FSH and LH) secretion, such as the anteroventral periventricular nucleus (AVPV), a small cluster of neurons along the wall of the third ventricle (AV3V, the anteroventral third ventricle) just caudal to the organum vasculosum of the lamina terminalis (OVLT), the periventricular nucleus (PeVN) and the arcuate nucleus (ARC) [29,36,37]. Furthermore, kisspeptin mRNA expression, albeit much weaker than in the above locations, was also confirmed in the anterodorsal preoptic nucleus, the medial amygdala, and the bed nucleus of the stria terminals [29,38,39]. Sexual dimorphism was found in the amount, but not the distribution, of kisspeptin in the CNS of mice, with females showing significantly greater kisspeptin expression, at least during the peripubertal period [29,40]. These differences in kisspeptin expression between the sexes may, however, vary depending on the period of life, the attainment of sexual maturity, the estrus cycle, or the species. For example, studies in rats have shown that sexual dimorphism in the expression of kisspeptin at the hypothalamic level during estrus is associated with an increase in males and a decrease in females. There was no change in the expression of GnRH or the kisspeptin receptor, regardless of sex, in this study [41].
Owing to ethical constraints, studies of the distribution of kisspeptin (+) neurons in the human brain have also yielded detailed topographic information. Studies conducted in nonhuman primates have established that in males, kisspeptin is present in the anterior part of the ARC and in the internal zone of the median eminence (ME) but is absent in the preoptic area (POA) and the AVPV [42,43]. Moreover, in females, kisspeptin (+) cells were detected in the POA in addition to the ARC, and kisspeptin expression was greater in the late follicular phase of the menstrual cycle than in the luteal phase [44]. Autopsy studies in humans have shown that kisspeptin neurons are found primarily in the infundibular nucleus and POA, which are analogous to the ARC and the rostral periventricular area of the third ventricle (RP3V) of the preoptic area in rodents, respectively [45]. In contrast to that in rodents, the sexual dimorphism of kisspeptin in humans includes not only its expression (increased in females) but also its distribution (increased expression of kisspeptin in the infundibular nucleus in females) [29,46].
2.2.2. Kisspeptin, Neurokinin B, and Dynorphin (KNDy) Neurons
Hypothalamic kisspeptin secretion is intrinsically linked to the KNDy neurons present there, the heterogeneous population of which includes a large subset of cells that, in addition to kisspeptin, coexpress tachykinin peptide neurokinin B (NKB) and the opioid peptide dynorphin (Dyn) [47,48]. The hypothalamic GnRH localization of KNDy neurons in the human brain is considered to reflect the conservation of functional anatomy across most mammalian species, and peptide colocalization was previously confirmed in sheep, rodents, cows and nonhuman primates [47,49].
Coexpression of the above protein compounds is closely related to the main function of KNDy neurons, namely, the negative feedback of GnRh within the HPG axis [50]. Increased levels of sex steroids released into the blood following GnRH stimulation cause the inhibition of kisspeptin in KNDy neurons, which is the essence of negative feedback control on the HPG axis [51]. The precise modulation of hormonal signals provided by NKB, Dyn and kisspeptin is responsible for generating the pulsatile release of GnRH, with kisspeptin secretion being dependent on NKB and Dyn [51,52,53]. The initiation of pulsatile GnRH release is mediated by the stimulating effect of NKB via NKB receptors (TACR3, tachykinin receptor 3) expressed on interconnected KNDy neurons, which leads to the release of kisspeptin in an autocrine manner. Activation of KISS1R on GnRH neurons subsequently induces pulsatile release of gonadoliberin (GnRH), whereas stimulation of KNDy neurons with KISS1R further enhances the stimulatory effect of NKB. Finally, the triggered GnRH pulse is terminated by the action of Dyn, which, via kappa opioid receptors (KORs) in KNDy neurons, inhibits both NKB and kisspeptin secretion. Furthermore, Dyn directly inhibits the activity of GnRH neurons by acting on their KORs [18,52,53]. There is still controversy about the coexpression of Dyn with kisspeptin/NKB in neurons of the infundibulum nucleus (the human homolog of the ARC) in humans, which contrasts with the results of previous immunohistochemical and in situ hybridization studies of sheep, goats, and rodents, which indicated that kisspeptin, NKB and Dyn are extensively colocalized in the hypothalamic ARC [54,55,56].
In addition to their role as GnRH pulse generators, KNDy neurons, mainly through the HPG axis, are also involved in the regulation of puberty onset [57], prolactin production [58,59], reproduction [60], the response of the reproductive system to stress [17,61], and the control of metabolic homeostasis, including thermoregulation [56,62,63,64]. Metabolic factors influencing the activity of kisspeptin (+) neurons at the subcellular, neuroendocrine and endocrine levels may therefore regulate the onset of puberty and reproductive processes associated with the HPG axis. Kisspeptin signaling pathways within KNDy neurons, particularly those involving glutamate as a cotransmitter, may also play a role in food intake, as their predominant role in energy expenditure has been demonstrated [56].
The general anatomical and functional scheme of KNDy neurons and kisspeptin neurons in relation to GnRH secretion is shown in Figure 2.
2.2.3. Extraneuronal Sources of Kisspeptin
The concentration of kisspeptin in blood in humans is not solely a consequence of the synthesis of this neurohormone by KNDy neurons in the infundibular nucleus and kisspeptin (+) neurons within the POA of the hypothalamus. The extraneuronal activation of KISS1 is responsible for the physiologically observed expression of kisspeptin in placental cell populations, including trophoblasts, within the ovaries and testes, adrenal cortex, pancreas, liver and small intestine [65,66]. The plasma concentration of kisspeptin in the circulation of healthy men and healthy nonpregnant women is low [65]. The significant increase in the plasma kisspeptin concentration during pregnancy is correlated with the development of the placenta, which is the main source of this hormone in pregnant women [19,21,67]. Moreover, in gestational trophoblastic neoplasia (GTN), which is a collective term for gestational trophoblastic diseases that invade locally or metastasize, plasma kisspeptin concentrations are elevated significantly above the norm for physiological pregnancy [68]. Therefore, kisspeptin may be a tumor biomarker in patients with GTN and is useful for monitoring the efficacy of chemotherapy, after which a gradual decrease in the plasma kisspeptin concentration is expected [68,69]. This finding may confirm the involvement of the kisspeptin-KISS1R signaling pathway in tumor biology, particularly in the previously described cancer-related disorders of cell proliferation, autophagy and apoptosis [70,71,72,73,74]. The nature of the action of kisspeptin, which is also a type of metastasis suppressor that is capable of modulating the behavior of some cancer cells by suppressing their proliferation, likely via the induction of autophagy and apoptosis, remains unclear and controversial, although many hypothetical mechanisms have been discussed [75,76,77,78,79]. It can be assumed that, in principle, any cellular stress (e.g., hypoxia) is able to increase the expression of KISS1 in different cell types, which is why, under physiological conditions associated with local angiogenesis accompanying implantation and placenta formation (placentation), an increased concentration of kisspeptin in the circulatory system of extraneuronal origin becomes understandable [80,81,82].
Notably, kisspeptin is produced in adipose tissue, where KISS1 mRNA has been detected in humans and rats [69,83]. Fasting induces an increase in KISS1 mRNA expression in rats, whereas a high-fat diet (HFD) causes it to decrease [84]. In women, however, higher KISS1 expression was demonstrated in subcutaneous adipose tissue than in visceral fat (the adipose tissue surrounding peritoneal organs), with a positive correlation with the body mass index (BMI) being found only for visceral fat [83]. The fact that, in women, including pregnant women, the kisspeptin concentration is related to BMI may indicate that this neurohormone is a link between metabolic processes and reproduction. In this context, kisspeptin may be a potential therapeutic target in the treatment of infertility due to metabolic disorders, especially obesity and insulin resistance [85,86,87].
3. Kisspeptin Receptor (KISS1R/GPR54)
All kisspeptins are endogenous ligands for the class A rhodopsin family of seven transmembrane (7TM) G protein-coupled receptors encoded by the KISS1R gene, which, previously, was named GPR54 and known as AXOR12 or hOT7T175 [88,89,90,91]. In humans, KISS1R is located on chromosome 19 (locus 19p13.3) and contains five coding exons [90,92].
Although KISS1R does not interact with galanin, significant homology with the galanin receptors GalR1, GalR2 and GalR3 within its putative transmembrane domains has been demonstrated, reaching 45% for GalR1 [93,94].
Broad expression of KISS1R in various tissues has been demonstrated in humans as well as common laboratory rodents, whereby the amino acid sequences of the human KISS1R protein (398 amino acids) share 82% identity with those of mice (396 amino acids) and display as much as 85% sequence identity (98% identity in transmembrane domains) with those of the rat KISS1R protein (396 amino acids) [49,89,95,96]. Human and rodent KISS1R homologous sequences (orthologs) are less conserved only in the carboxyl (COOH)-terminal, the amine (NH2)-terminal and within the third extracellular loop of the receptor [96,97].
Numerous studies on the etiology of CPP and IHH have been accompanied by the discovery of KISS1R point mutations resulting in the replacement of single amino acids in the KISS1R molecule chain [30,31,98,99,100]. Since all receptors belonging to the GPCR family show fundamental structural similarity, information obtained from the analysis of KISS1R mutations can be used more universally, extending it to improve the understanding of the function of different domains within the GPCR family [30,101,102].
The general structure of KISS1R is shown in Figure 3, which also indicates the mutation sites (amino acid exchanges) in the peptide chain leading to inactivation, possible inactivation or activation of the receptor.
3.1. Kisspeptin/KISS1R Signaling
Owing to a conserved 10 amino acid core sequence, all kisspeptin isoforms of different chain lengths (Kp-54, Kp-14, Kp-13 and Kp-10; see Figure 1.) resulting from proteolytic cleavage of prepro-kisspeptin show affinity for KISS1R [26,28]. When these ligands bind to KISS1R, a typical 7TM GPCR, they act as guanine nucleotide exchange factors (GEFs), leading to a conformational change in the three-dimensional (3-D) structure of KISS1R [90,107].
The heterotrimeric G protein is activated at the level of the Gαq/11 subunit through the exchange of a guanosine diphosphate (GDP) for a guanosine triphosphate (GTP). Activated KISS1R dissociates into Gαq/11-GTP and Gβγ to further initiate Gαq/11-dependent intracellular signaling [50]. Next, Gαq/11 activates phospholipase C (PLC)-β, an essential enzyme in the metabolism of phosphoinositide in the cytoplasm. The hydrolytic activity of PLC-β is responsible for the breakdown of the inner membrane component phosphatidylinositol-4,5-bisphosphate (PIP2) into inositol 1,4,5-trisphosphate (IP3) and diacylglycerol (DAG) [108,109,110]. Because IP3 and DAG are key signaling molecules that influence various cellular functions and are precursors of other signaling molecules, PLC can be considered a hub for numerous interconnected signaling pathways. IP3 acts as a second messenger in the release of calcium ions (Ca2+) stored in the endoplasmic reticulum (ER) [111,112], whereas DAG acts as a second messenger in the activation of protein kinase C (PKC) [113,114]. The mobilization and intracytoplasmic release of Ca2+ result from the interaction of IP3 with IP3-sensitive Ca2+ channels, which can promote a variety of different functions, depending on the cell type and its functional state. For example, an increase in the Ca2+ concentration may provide kisspeptin with the ability to release GnRH and modulate its ability to inhibit cell proliferation, including antimetastatic effects [115,116]. DAG acts as an allosteric activator of PKC, which subsequently causes a phosphorylation signaling cascade and ultimately leads to the activation of kinases belonging to the serine/threonine mitogen-activated protein kinase (MAPK) family, including extracellular signal-regulated kinase 1/2 (ERK1/2) and p38 mitogen-activated protein kinase (p38MAPK) [117,118]. Therefore, as a consequence of the action of DAG, the activation of the PKC/Ras (kinases)/Raf (kinases)/MEK/ERK1/2 pathway and the PKC/Ras/Raf/MEK/p38MAPK pathway can be distinguished [119].
Additionally, KISS1R signals independently of Gαq/11 via the adaptor proteins β-arrestins 1 and 2, leading to the inhibition or recruitment of the ERK1/2 pathway, respectively [120,121,122]. Although β-arrestins function to "arrest" G protein signaling to promote GPCR desensitization and internalization, as modulators of downstream KISS1R signaling, they have been shown to function both as passive scaffolds and as active allosteric regulators of enzymatic activity [123].
KISS1R activation may, depending on the cell type and functional state, stimulate or inhibit PI3K (phosphatidylinositol 3-kinase)/Akt (serine/threonine kinase Akt, also known as protein kinase B (PKB)) phosphorylation, although the mechanisms of this phenomenon, including intermediary compounds, have not been elucidated [124,125]. Through signaling via the PI3K/Akt/NF-κB (nuclear factor kappa-light-chain-enhancer of activated B cells) or the ERK1/2/NF-κB pathways, upon KISS1R stimulation, decreased expression of the matrix metalloproteinase-9 (MMP-9) gene also occurs [126,127]. MMP-9 directly degrades extracellular matrix (ECM) proteins and regulates tissue remodeling via cytokine/chemokine mediators in both physiological (e.g., embryonic development, reproduction, angiogenesis, and wound healing) and pathological (e.g., asthma and arthritis) processes [128,129,130,131,132,133]. Therefore, during the process of blastocyst implantation, kisspeptin may attenuate the excessive migration and invasion of trophoblasts through the inhibition of MMP-9 and, to a lesser degree, matrix metalloproteinase-2 (MMP-2) [134,135,136]. These findings confirm that the specific action of kisspeptin is dependent on the cell type, differentiation state and functional state.
The signaling pathways involved in the interaction of kisspeptin with KISS1R are shown in Figure 4.
4. Kisspeptin in Early Pregnancy
Early pregnancy, defined in humans as the first 12 weeks after fertilization, is a crucial period in the development of the fetus. This stage includes the germinal stage, which lasts 10 days and refers to the period from fertilization to the completion of blastocyst implantation in the decidual endometrium, i.e., the formation of the early embryo [137,138]. During the embryonic stage, which lasts from approximately the third week to the end of the eighth week of pregnancy, organogenesis takes place, during which a developing embryo establishes the rudimentary structures of all of its organs and tissues from the ectoderm, mesoderm, and endoderm [139]. At the beginning of the ninth week, the embryo is termed a fetus [140].
In addition to its role in the hypothalamus, nonneuronal kisspeptin action in peripheral tissues also has many regulatory functions, including during pregnancy [20,22,141]. As mentioned earlier, the concentration of circulating kisspeptin increases many times from the first weeks of pregnancy, and its main source is the placenta [19,21,33,67]. Kisspeptin is believed to regulate the manner in which this temporary organ is formed [21,25,142]. For example, the kisspeptin-54 concentration measured using an immunoenzymatic method clearly linearly increased with gestational age, reaching 1230 pmol/L in the first trimester and a maximum of 9590 pmol/L in the third trimester, after which it returned to nonpregnancy values (< 100 pmol/L) shortly after delivery [69].
The growing evidence for the involvement of kisspeptin/KISS1R signaling in embryo implantation, trophoblast invasion and placentation justifies the consideration of this neurohormone as a potential biomarker in early pregnancy; its levels correlate with the risk of miscarriage, preeclampsia/eclampsia, GTN, and fetal growth retardation [21,23,67,68,69,143,144,145,146]. Kisspeptin measurements have been shown to have potentially comparable or greater accuracy than human chorionic gonadotropin (HCG) in differentiating between miscarriage and viable intrauterine pregnancy after 6 weeks of gestation [147,148]. Kisspeptin deficiency in the serum of females with unexplained infertility was associated with a reduced chance of implantation after intracytoplasmic sperm injection (ICSI) [149].
4.1. Embryo Implantation
Implantation, the process by which a blastocyst attaches to the decidualized endometrium of the uterus to achieve pregnancy, can be divided into three distinct, increasingly advanced stages: apposition (unstable adhesion), adhesion and invasion (penetration) [150]. During this highly orchestrated process, there is an interaction between the receptive endometrium and the competent embryo within a precisely defined time frame called the "window of implantation", which includes contact of the blastocyst with the implantation site (apposition), attachment of the trophoblast cells of the blastocyst to the receptive endometrial epithelium (adhesion) and crossing the endometrial epithelial basement membrane toward the endometrial stroma by invasive trophoblast cells (invasion) [151].
The expression of the functional form of the kisspeptin/KISS1R system has been demonstrated at each of the above stages, as well as during decidual transformation of the endometrium [21,152]. In gametes, the expression of kisspeptin and KISS1R has been documented in human oocytes and in human spermatozoa [153,154,155]. In the latter, both kisspeptin and KISS1R are localized mainly in the sperm head, around the neck and in the flagellum [155]. Single-cell RNA sequencing (RNA-Seq) analyses have confirmed the expression of KISS1R in human embryos (from the zygote stage to the blastocyst stage), with some doubts regarding the production of kisspeptin by dividing germ cells [22,156].
Trophectoderm cells derived from the blastocyst give rise to cytotrophoblast cells, which, after proliferation and differentiation, finally give rise to the syncytiotrophoblast and an outwardly developing multilayer of mononuclear cells of the invasive extravillous cytotrophoblast (EVT) [157]. This invasive behavior is due to the ability of cytotrophoblasts to secrete matrix metalloproteinases (MMPs), and kisspeptin may regulate trophoblast migration and invasion by influencing the activity of MMP-9 and, to a lesser extent, MMP-2 [69,135,158]. Moreover, the contradictory results of the kisspeptin/KISS1R signaling studies may suggest that kisspeptin is a factor that bidirectionally regulates trophoblast invasiveness and migration, either by promoting or inhibiting it during embryo implantation and placentation [159]. Kisspeptin/KISS1R-dependent adhesion of EVTs to type-I collagen, a major component of the human placenta, plays a key role in this regulatory mechanism. Both the rapid and transient nature of this increased adhesion have been shown to be due to the activation of secondary effectors such as PKC and ERK1/2 in EVT cells following KISS1R activation [82].
Blastocyst implantation and the development of early pregnancy are not possible without decidual transformation of the endometrium. During decidualization, mainly under the influence of high progesterone levels, changes occur in endometrial cells that increase glandular epithelial secretion, increase glycogen storage, and enhance stromal vascularity [160]. Kisspeptin is recognized as a physiological modulator of stromal decidualization, which stimulates this process by upregulating leukemia inhibitory factor (LIF), a major cytokine regulator of decidualization in humans and mice [161,162]. Using quantitative PCR, a dynamic increase in kisspeptin and KISS1R mRNA levels was demonstrated in the mouse uterine mucosa during early pregnancy and during artificial decidualization in vivo. The expression of the kisspeptin and KISS1R proteins was then observed in a spatiotemporal manner in decidualization stromal cells in intact pregnant females as well as in pseudopregnant mice subjected to artificially induced decidualization [163]. The promotion of endometrial decidualization by the kisspeptin/KISS1R system is associated with the activation of the ERK1/2 signaling pathway, resulting in increased expression and secretion of LIF. In line with the above findings, in patients with recurrent spontaneous abortion (RSA), both serum kisspeptin levels and decidual kisspeptin expression are lower than those in normal pregnant individuals. These kisspeptin deficiencies are accompanied by reduced expression of decidualization markers, including prolactin (PRL) and insulin-like growth factor-binding protein 1 (IGFBP-1), in the decidua [152].
Recently, it has also been suggested that kisspeptin may regulate the decidualization process by modulating the neurogenic locus notch homolog protein 1 (Notch1)/Akt/forkhead box protein O1 (FOXO1) signaling cascade. In this signaling system, downregulation of kisspeptin expression may cause suboptimal decidualization and consequently contribute to the development or progression of RSA [164].
During implantation, KISS1R negatively regulates estrogen receptor alpha (ERα) signaling, in part by inhibiting ERα overexpression and preventing detrimentally high ERα activity [165]. Therefore, while the growth and development of the uterus itself are largely dependent on ovarian estradiol (E2) output via central kisspeptin signaling, peripheral kisspeptin signaling is indispensable for achieving full functional competence of the endometrium at the time of implantation [165,166].
Another important mechanism of action of kisspeptin in pregnancy is related to maternal immunological tolerance, which is necessary to avoid fetal rejection [141,167]. This immunomodulatory action of kisspeptin is revealed as early as the last stage of embryo implantation, invasion, which starts with the penetration of the syncytiotrophoblast through the uterine epithelium with subsequent infiltration of the mononuclear cytotrophoblast invading the entire endometrium [69]. In vitro studies have shown that incubation of T cells in medium containing kisspeptin at concentrations corresponding to those present during pregnancy results in increased differentiation of human naive T cells into T-regulatory cells [168]. Moreover, the transformation of CD4+ T lymphocytes into inducible regulatory T cells (iTregs) following interaction with kisspeptin was accompanied by concomitant inhibition of the differentiation of interleukin 17 (IL-17)-producing T helper (Th17) cells [169].
4.2. Trophoblast Invasion–Vascular Remodeling–Placentation
The process of invasive growth of the trophoblast into the decidualized endometrial stroma and inner third of the myometrium, initiated at the last stage of blastocyst implantation, is crucial for the formation of the placenta and the establishment of definitive maternal‒fetal circulation [150,151]. This includes endovascular trophoblast invasion, which occurs in two stages, with infiltration into the decidual segments of the spiral arteries between 8 and 10 weeks of gestation and invasion into myometrial segments of the spiral arteries between 16 and 18 weeks of gestation [151,170,171]. In the early weeks of pregnancy, migrating extravillous cytotrophoblasts plug the spiral vessels, presumably to prevent premature immunization associated with the appearance of maternal blood in the intervillous space [172,173]. This sealing of the lumen of the spiral arteries disappears around the 10th to 12th week of pregnancy, when the final vascular remodeling leads to the establishment of low-resistance circulation in the developing maternal–fetal unit [173,174]. Failure of trophoblast invasion and spiral artery remodeling results in pregnancy-induced hypertension and/or fetal growth restriction (FGR) [175,176,177].
Consistent with the original findings of the discovery of kisspeptin, which in the Kp-54 form was named metastin to emphasize its antimetastatic activity, KISS-1 gene products inhibit cellular invasion, migration and angiogenesis [178,179]. High levels of endogenous kisspeptin and marked expression of KISS1R in placental trophoblast cells have been found to be associated with direct regulation of trophoblast proliferation and invasiveness and to indirectly influence the development of the placental vascular network [21,142]. Kisspeptin limits the invasiveness of trophoblasts by inhibiting MMP-9 and MMP-2, both at the transcriptional and protein levels [158]. Reduced extracellular matrix (ECM) degradation limits the ability of the cytotrophoblast to infiltrate the uterine wall, and the effect of kisspeptin, although restricted to MMP-9 and MMP-2, mimics and supports endogenous tissue inhibitors of MMPs (TIMPS), which have been shown to be expressed in both the human decidua and the trophoblast [180,181]. Owing to this effect of kisspeptin/KISS1R signaling, trophoblastic cells, with their strictly controlled propensity for invasive growth, have limited similarity to malignant cells [182,183].
In trophoblast invasiveness studies conducted in the KISS1R knockout (KO) model, the expression of the trophoblast invasion marker MMP-2 mRNA was greater in the placental labyrinth zone of KISS1R KO mice than in that of wild-type mice [23]. Similarly, the migratory potential of cytotrophoblast-like cells was inhibited after kisspeptin treatment in a human trophoblast (BeWo) cell line cultured in vitro, with previously confirmed kisspeptin protein expression [23].
The demonstration of the role of kisspeptin/KISS1R signaling in limiting trophoblast invasiveness has become the starting point for recognizing it as a potential risk factor for the occurrence of poor trophoblast cell invasion, inadequate uterine vascular remodeling, and placental hypoperfusion. These crucial placental disorders are clearly part of the pathophysiology of preeclampsia/eclampsia, a frequently occurring metabolic/hypertensive complication that poses a threat to the health and life of women and fetuses [184,185]. It has been suggested that insufficient trophoblast invasion into spiral arteries, leading to pregnancy-induced hypertension, may be a consequence of the inhibitory effect of kisspeptin on macrophage migration inhibitory factor (MIF) [158]. MIF is a multifunctional proinflammatory cytokine highly secreted by the first-trimester placenta, mainly in villous and extravillous trophoblasts, that plays an important role in the activation of innate and adaptive immune responses. Moreover, trophoblast MIF is induced by hypoxia to promote trophoblast cell invasion and migration [186,187].
Many other data, often requiring verification, suggest possible functional links between kisspeptin and placental trophoblast invasiveness, angiogenesis and immunology in early pregnancy. For example, the kisspeptin/KISS1R system inhibits the expression of vascular endothelial growth factor (VEGF), and increased placental KISS1R expression is positively correlated with tumor necrosis factor alpha (TNFα) synthesis, whereas interferon gamma (IFNγ), interleukin 6 (IL-6), and interleukin 10 (IL-10) synthesis are negatively correlated with placental KISS1R expression [80,184,188,189,190].
Optimal control of EVT invasiveness causes them to invade the decidua and move toward the spiral arteries in early pregnancy, where they first form trophoblast plugs and, after destroying the vascular smooth muscle layer, lead to vascular remodeling with the creation of low-resistance blood flow [172,191]. The abovementioned phenomena constitute the essence of the hemochorial placentation process, which begins in humans approximately 8–9 days after fertilization and leads to the formation of the placenta, in which maternal blood comes in direct contact with the fetal chorion. Ultimately, approximately 120–140 spiral arteries undergo remodeling, which, after developing the final form of uteroplacental circulation, are able to provide a flow at the level of 20% of the cardiac output of the pregnant woman during full-term pregnancy [171]. Kisspeptin/KISS1R signaling in trophoblast cells affects intracellular skeletal organization, and cell migration and collagenase activity may play crucial roles in this process [11,134].
Given that kisspeptin has been proposed as a biomarker of healthy placentation, it could be used to recognize both anatomical and locational abnormalities of the developing placenta in early pregnancy [67,69]. Low-lying placentas, placenta previa and abnormally invasive placentas are the most frequently occurring placental abnormalities in terms of location and anatomy, whereas ectopic pregnancy is a complication of pregnancy in which the embryo attaches outside the uterus (e.g., placentation in the fallopian tube) [192,193,194].
The evaluation of the kisspeptin concentration in the serum revealed a significant decrease in pregnant women with placenta previa compared with the control group (healthy pregnancies with normal intrauterine placenta locations). Moreover, in cases of placenta previa, biochemical, immunohistochemical and genetic analyses consistently revealed reduced expression of the KISS1 gene, accompanied by a very significant (170-fold) increase in KISS1R expression [143]. Further studies, including tissue modeling with assessments of angiogenesis, migration and the degree of cell invasiveness, are necessary to fully understand the importance of kisspeptin/KISS1R signaling disorders in the pathomechanism of placenta previa development.
In ectopic pregnancy, a significant reduction in the serum kisspeptin concentration is also observed compared with that in the first trimester of normal pregnancy. In addition, the reduced kisspeptin levels were correlated with reduced levels of KISS1 expression in human embryonic/placental tissue obtained from ectopic pregnancies, which may be due to increased expression of the microRNA miR-324-3p, a putative repressor of KISS1 in human embryonic/placental tissue at <12 weeks of gestation [195,196]. The above results provide a basis for considering circulating kisspeptin and miR-324-3p as potential biomarkers of ectopic pregnancy at early gestational stages [67,69,196].
Data on the kisspeptin concentration and KISS1 and KISS1R expression in trophoblast/placental tissue in early pregnancy (gestational age ≤ 12 weeks) with a physiological and complicated course are summarized in Table 1.
5. Concluding Remarks
In addition to its direct effects on reproductive function within the HPG axis, the neurohormone kisspeptin plays an important role in processes accompanying key stages of early human pregnancy, such as embryo implantation with trophoblast invasion and vascular remodeling, which are thought to ensure successful placentation. This is confirmed by the fact that the spectacular, approximately 900-fold increase in the serum kisspeptin concentration in the first trimester of pregnancy, compared with that in nonpregnant women, is due to its extraneuronal pool, which is functionally unrelated to KNDy neurons. The source of this extraneuronal kisspeptin is primarily populations of trophoblast cells, the dynamic proliferation of which determines the formation of the placenta, and its tissue supports the production of this hormone.
Previous findings that the kisspeptin (then called metastin)/KISS1R (then called GPR54) signaling pathway plays a role in cancer biology as a suppressor of metastatic spread by inhibiting cell proliferation, probably via the induction of autophagy and apoptosis, have been extended to the control of trophoblast invasiveness in early pregnancy. Defective trophoblast invasion has been shown to be closely associated with pregnancy complications, including ectopic pregnancy, GTN, placenta previa and miscarriage. Optimal control of KISS1 and KISS1R expression during early trophoblast development is therefore essential for initiating processes leading to eutopic placentation with adequate invasion into spiral uterine arteries to develop low-resistance vessels in the hemochorial placenta.
Considering these findings, circulating kisspeptin could be used as a potential biomarker for the detection of, among other factors, the well-being of early pregnancy and the risk of miscarriage, GTN, or its abnormal location (ectopic pregnancy). When kisspeptin is used as a marker of the above pathologies of early pregnancy, one should remember the methodological difficulties in determining kisspeptin levels, which are caused by the existence of 4 isoforms of this hormone (i.e., Kp-54, Kp-14, Kp-13, or Kp-10), which additionally differ significantly in their rate of metabolism in the body. It is necessary to refine laboratory methods for detecting kisspeptin and promote those that are characterized by adequate sensitivity and repeatability under clinical conditions.
The need for future studies arises from the demonstration that the kisspeptin concentration in women, including pregnant women, correlates with BMI and, in particular, with KISS1R mRNA expression in visceral fat. This allows for the linkage of the metabolic actions of kisspeptin with its reproductive regulatory functions at the level of the HPG axis and early pregnancy tissues. Therefore, in metabolic disorders associated with infertility, such as obesity and insulin resistance, targeting the kisspeptin signaling pathway may yield beneficial therapeutic effects.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created. Instead, the data are quoted from the available cited literature.
Conflicts of Interest
The author declares no conflict of interest.
Abbreviations
7TM GPCR – seven-transmembrane G protein-coupled receptor
ARC – arcuate nucleus
AV3V – anteroventral third ventricle
AVPV – anteroventral periventricular nucleus
BMI – body mass index
CNS – central nervous system
CPP – central precocious puberty
DAG – diacylglycerol
Dyn – dynorphin
E2 – estradiol
ECM – extracellular matrix
ER – endoplasmic reticulum
ERα – estrogen receptor alpha
ERK1/2 – extracellular signal-regulated kinase 1/2
EVT – extravillous cytotrophoblast
FGR – fetal growth restriction
FOXO1 – forkhead box protein O1
FSH – follicle stimulating hormone
GalR1, GalR2, GalR3 – galanin receptors type 1, 2 and 3 respectively
GEFs – guanine nucleotide exchange factors
GnRH – gonadotropin-releasing hormone
GTN – gestational trophoblastic neoplasia
HCG – human chorionic gonadotropin
HFD – high fat diet
HPG axis – hypothalamic-pituitary-gonadal axis
iTreg – inducible regulatory T cells
ICSI – intracytoplasmic sperm injection
IFNγ – interferon gamma
IGFBP-1 – insulin-like growth factor-binding protein 1
IHH – idiopathic hypogonadotropic hypogonadism
IL-6, IL-10, IL-17 – interleukins 6, 10 and 17, respectively
IP3 – inositol 1,4,5-trisphosphate
KISS1 – gene encoding kisspeptin
KISS1R – KISS1-derived peptide receptor is a G protein-coupled receptor (also known as GPR54)
KNDy neurons – kisspeptin, neurokinin B, dynorphin neurons
KOR – kappa opioid receptor
Kp-54, Kp-14, Kp-13, Kp-10 – the four kisspeptin isoforms
LH – luteinizing hormone
LIF – leukemia inhibitory factor
MAPKs – mitogen-activated protein kinases
MIF – macrophage migration inhibitory factor
MMP-2, MMP-9 – matrix metalloproteinase 2 and 9, respectively
MMPs – matric metalloproteinases
NF-κB – nuclear factor kappa-light-chain-enhancer of activated B cells
NKB – neurokinin B
Notch1 – neurogenic locus notch homolog protein 1
OVLT – organum vasculosum of the lamina terminalis
p38MAPK – p38 mitogen-activated protein kinase
PeVN – periventricular nucleus
PIP2 – phosphatidylinositol-4,5-bisphosphate
PKB – protein kinase B (also known as serine/threonine kinase Akt)
PKC – protein kinase C
PLC-β – phospholipase C beta
POA – preoptic area
PRL – prolactin
RNA-Seq – RNA sequencing
RSA – recurrent spontaneous abortion
TACR3 – tachykinin receptor 3 (NKB receptor)
Th17 – T helper 17 cells
TIMPS – tissue inhibitors of matrix metalloproteinases (MMPs)
TNFα – tumor necrosis factor alpha
VEGF – vascular endothelial growth factor
References
- Dufour S, Quérat B, Tostivint H, Pasqualini C, Vaudry H, Rousseau K. Origin and Evolution of the Neuroendocrine Control of Reproduction in Vertebrates, With Special Focus on Genome and Gene Duplications. Physiol Rev. 2020;100(2):869-943. [CrossRef]
- Pillerová M, Borbélyová V, Hodosy J, Riljak V, Renczés E, Frick KM, Tóthová Ľ. On the role of sex steroids in biological functions by classical and non-classical pathways. An update. Front Neuroendocrinol. 2021;62:100926. [CrossRef]
- Colldén H, Nilsson ME, Norlén AK, Landin A, Windahl SH, Wu J, Gustafsson KL, Poutanen M, Ryberg H, Vandenput L, Ohlsson C. Comprehensive Sex Steroid Profiling in Multiple Tissues Reveals Novel Insights in Sex Steroid Distribution in Male Mice. Endocrinology. 2022;163(3):bqac001. [CrossRef]
- Chatuphonprasert W, Jarukamjorn K, Ellinger I. Physiology and Pathophysiology of Steroid Biosynthesis, Transport and Metabolism in the Human Placenta. Front Pharmacol. 2018;9:1027. [CrossRef]
- Goodman RL, Herbison AE, Lehman MN, Navarro VM. Neuroendocrine control of gonadotropin-releasing hormone: Pulsatile and surge modes of secretion. J Neuroendocrinol. 2022;34(5):e13094. [CrossRef]
- Filicori, M. Pulsatile gonadotropin-releasing hormone: clinical applications of a physiologic paradigm. F S Rep. 2023;4(2 Suppl):20-26. [CrossRef]
- Garg D, Berga SL. Neuroendocrine mechanisms of reproduction. Handb Clin Neurol. 2020;171:3-23. [CrossRef]
- Koysombat K, Dhillo WS, Abbara A. Assessing hypothalamic pituitary gonadal function in reproductive disorders. Clin Sci (Lond). 2023;137(11):863-879. [CrossRef]
- Tsutsui K, Bentley GE, Kriegsfeld LJ, Osugi T, Seong JY, Vaudry H. Discovery and evolutionary history of gonadotrophin-inhibitory hormone and kisspeptin: new key neuropeptides controlling reproduction. J Neuroendocrinol. 2010;22(7):716-27. [CrossRef]
- Dhillo, W. Timeline: kisspeptins. Lancet Diabetes Endocrinol. 2013;1(1):12-3. [CrossRef]
- Ohtaki T, Shintani Y, Honda S, Matsumoto H, Hori A, Kanehashi K, Terao Y, Kumano S, Takatsu Y, Masuda Y, Ishibashi Y, Watanabe T, Asada M, Yamada T, Suenaga M, Kitada C, Usuki S, Kurokawa T, Onda H, Nishimura O, Fujino M. Metastasis suppressor gene KiSS-1 encodes peptide ligand of a G-protein-coupled receptor. Nature. 2001;411(6837):613-7. [CrossRef]
- Stafford LJ, Xia C, Ma W, Cai Y, Liu M. Identification and characterization of mouse metastasis-suppressor KiSS1 and its G-protein-coupled receptor. Cancer Res. 2002;62(19):5399-404.
- Makri A, Pissimissis N, Lembessis P, Polychronakos C, Koutsilieris M. The kisspeptin (KiSS-1)/GPR54 system in cancer biology. Cancer Treat Rev. 2008;34(8):682-92. [CrossRef]
- Rhie, YJ. Kisspeptin/G protein-coupled receptor-54 system as an essential gatekeeper of pubertal development. Ann Pediatr Endocrinol Metab. 2013;18(2):55-9. [CrossRef]
- Joy KP, Chaube R. Kisspeptin control of hypothalamus-pituitary-ovarian functions. Vitam Horm. 2025;127:153-206. [CrossRef]
- Sobrino V, Avendaño MS, Perdices-López C, Jimenez-Puyer M, Tena-Sempere M. Kisspeptins and the neuroendocrine control of reproduction: Recent progress and new frontiers in kisspeptin research. Front Neuroendocrinol. 2022;65:100977. [CrossRef]
- Acevedo-Rodriguez A, Kauffman AS, Cherrington BD, Borges CS, Roepke TA, Laconi M. Emerging insights into hypothalamic-pituitary-gonadal axis regulation and interaction with stress signalling. J Neuroendocrinol. 2018;30(10):e12590. [CrossRef]
- Koysombat K, Tsoutsouki J, Patel AH, Comninos AN, Dhillo WS, Abbara A. Kisspeptin and neurokinin B: roles in reproductive health. Physiol Rev. 2025;105(2):707-764. [CrossRef]
- Radovick S, Babwah AV. Regulation of Pregnancy: Evidence for Major Roles by the Uterine and Placental Kisspeptin/KISS1R Signaling Systems. Semin Reprod Med. 2019;37(4):182-190. [CrossRef]
- Cao Y, Li Z, Jiang W, Ling Y, Kuang H. Reproductive functions of Kisspeptin/KISS1R Systems in the Periphery. Reprod Biol Endocrinol. 2019;17(1):65. [CrossRef]
- Hu KL, Chang HM, Zhao HC, Yu Y, Li R, Qiao J. Potential roles for the kisspeptin/kisspeptin receptor system in implantation and placentation. Hum Reprod Update. 2019;25(3):326-343. [CrossRef]
- D'Occhio MJ, Campanile G, Baruselli PS. Peripheral action of kisspeptin at reproductive tissues-role in ovarian function and embryo implantation and relevance to assisted reproductive technology in livestock: a review. Biol Reprod. 2020;103(6):1157-1170. [CrossRef]
- Panting EN, Weight JH, Sartori JA, Coall DA, Smith JT. The role of placental kisspeptin in trophoblast invasion and migration: an assessment in Kiss1r knockout mice, BeWo cell lines and human term placenta. Reprod Fertil Dev. 2024; 36:RD23230. [CrossRef]
- Szydełko-Gorzkowicz M, Poniedziałek-Czajkowska E, Mierzyński R, Sotowski M, Leszczyńska-Gorzelak B. The Role of Kisspeptin in the Pathogenesis of Pregnancy Complications: A Narrative Review. Int J Mol Sci. 2022;23(12):6611. [CrossRef]
- Musa E, Salazar-Petres E, Vatish M, Levitt N, Sferruzzi-Perri AN, Matjila MJ. Kisspeptin signalling and its correlation with placental ultrastructure and clinical outcomes in pregnant South African women with obesity and gestational diabetes. Placenta. 2024;154:49-59. [CrossRef]
- López-Ojeda W, Hurley RA. Kisspeptin in the Limbic System: New Insights Into Its Neuromodulatory Roles. J Neuropsychiatry Clin Neurosci. 2022;34(3):190-195. [CrossRef]
- Pinilla L, Aguilar E, Dieguez C, Millar RP, Tena-Sempere M. Kisspeptins and reproduction: physiological roles and regulatory mechanisms. Physiol Rev. 2012; 92(3):1235-316. [CrossRef]
- Tena-Sempere, M. GPR54 and kisspeptin in reproduction. Hum Reprod Update. 2006; 12(5):631-9. [CrossRef]
- Hu KL, Chen Z, Li X, Cai E, Yang H, Chen Y, Wang C, Ju L, Deng W, Mu L. Advances in clinical applications of kisspeptin-GnRH pathway in female reproduction. Reprod Biol Endocrinol. 2022;20(1):81. [CrossRef]
- Ke R, Ma X, Lee LTO. Understanding the functions of kisspeptin and kisspeptin receptor (Kiss1R) from clinical case studies. Peptides. 2019;120:170019. [CrossRef]
- Silveira LG, Noel SD, Silveira-Neto AP, Abreu AP, Brito VN, Santos MG, Bianco SD, Kuohung W, Xu S, Gryngarten M, Escobar ME, Arnhold IJ, Mendonca BB, Kaiser UB, Latronico AC. Mutations of the KISS1 gene in disorders of puberty. J Clin Endocrinol Metab. 2010;95(5):2276-80. [CrossRef]
- Gorkem U, Kan O, Bostanci MO, Taskiran D, Inal HA. Kisspeptin and Hematologic Parameters as Predictive Biomarkers for First-Trimester Abortions. Medeni Med J. 2021;36(2):98-105. [CrossRef]
- Horikoshi Y, Matsumoto H, Takatsu Y, Ohtaki T, Kitada C, Usuki S, Fujino M. Dramatic elevation of plasma metastin concentrations in human pregnancy: metastin as a novel placenta-derived hormone in humans. J Clin Endocrinol Metab. 2003;88(2):914-9. [CrossRef]
- Cetković A, Miljic D, Ljubić A, Patterson M, Ghatei M, Stamenković J, Nikolic-Djurovic M, Pekic S, Doknic M, Glišić A, Bloom S, Popovic V. Plasma kisspeptin levels in pregnancies with diabetes and hypertensive disease as a potential marker of placental dysfunction and adverse perinatal outcome. Endocr Res. 2012;37(2):78-88. [CrossRef]
- d'Anglemont de Tassigny X, Jayasena CN, Murphy KG, Dhillo WS, Colledge WH. Mechanistic insights into the more potent effect of KP-54 compared to KP-10 in vivo. PLoS One. 2017;12(5):e0176821. Erratum in: PLoS One. 2018; 13(1):e0192014. doi: 10.1371/journal.pone.0192014. [CrossRef]
- Wang L, Moenter SM. Differential Roles of Hypothalamic AVPV and Arcuate Kisspeptin Neurons in Estradiol Feedback Regulation of Female Reproduction. Neuroendocrinology. 2020;110(3-4):172-184. [CrossRef]
- Mills EG, Dhillo WS. Invited review: Translating kisspeptin and neurokinin B biology into new therapies for reproductive health. J Neuroendocrinol. 2022;34(10):e13201. [CrossRef]
- Gottsch ML, Cunningham MJ, Smith JT, Popa SM, Acohido BV, Crowley WF, Seminara S, Clifton DK, Steiner RA. A role for kisspeptins in the regulation of gonadotropin secretion in the mouse. Endocrinology. 2004;145(9):4073-7. [CrossRef]
- Clarkson J, d'Anglemont de Tassigny X, Colledge WH, Caraty A, Herbison AE. Distribution of kisspeptin neurones in the adult female mouse brain. J Neuroendocrinol. 2009;21(8):673-82. [CrossRef]
- Semaan SJ, Kauffman AS. Developmental sex differences in the peri-pubertal pattern of hypothalamic reproductive gene expression, including Kiss1 and Tac2, may contribute to sex differences in puberty onset. Mol Cell Endocrinol. 2022;551:111654. [CrossRef]
- Muñoz de la Torre LP, Trujillo Hernández A, Eguibar JR, Cortés C, Morales-Ledesma L. Sex-specific hypothalamic expression of kisspeptin, gonadotropin releasing hormone, and kisspeptin receptor in progressive demyelination model. J Chem Neuroanat. 2022;123:102120. [CrossRef]
- Shibata M, Friedman RL, Ramaswamy S, Plant TM. Evidence that down regulation of hypothalamic KiSS-1 expression is involved in the negative feedback action of testosterone to regulate luteinising hormone secretion in the adult male rhesus monkey (Macaca mulatta). J Neuroendocrinol. 2007;19(6):432-8. [CrossRef]
- Ramaswamy S, Guerriero KA, Gibbs RB, Plant TM. Structural interactions between kisspeptin and GnRH neurons in the mediobasal hypothalamus of the male rhesus monkey (Macaca mulatta) as revealed by double immunofluorescence and confocal microscopy. Endocrinology. 2008;149(9):4387-95. [CrossRef]
- Smith JT, Shahab M, Pereira A, Pau KY, Clarke IJ. Hypothalamic expression of KISS1 and gonadotropin inhibitory hormone genes during the menstrual cycle of a non-human primate. Biol Reprod. 2010;83(4):568-77. [CrossRef]
- Rometo AM, Krajewski SJ, Voytko ML, Rance NE. Hypertrophy and increased kisspeptin gene expression in the hypothalamic infundibular nucleus of postmenopausal women and ovariectomized monkeys. J Clin Endocrinol Metab. 2007; 92(7):2744-50. [CrossRef]
- Hrabovszky E, Ciofi P, Vida B, Horvath MC, Keller E, Caraty A, Bloom SR, Ghatei MA, Dhillo WS, Liposits Z, Kallo I. The kisspeptin system of the human hypothalamus: sexual dimorphism and relationship with gonadotropin-releasing hormone and neurokinin B neurons. Eur J Neurosci. 2010;31(11):1984-98. [CrossRef]
- Moore AM, Coolen LM, Porter DT, Goodman RL, Lehman MN. KNDy Cells Revisited. Endocrinology. 2018;159(9):3219-3234. [CrossRef]
- Velasco I, Franssen D, Daza-Dueñas S, Skrapits K, Takács S, Torres E, Rodríguez-Vazquez E, Ruiz-Cruz M, León S, Kukoricza K, Zhang FP, Ruohonen S, Luque-Cordoba D, Priego-Capote F, Gaytan F, Ruiz-Pino F, Hrabovszky E, Poutanen M, Vázquez MJ, Tena-Sempere M. Dissecting the KNDy hypothesis: KNDy neuron-derived kisspeptins are dispensable for puberty but essential for preserved female fertility and gonadotropin pulsatility. Metabolism. 2023;144:155556. [CrossRef]
- Beltramo M, Robert V, Decourt C. The kisspeptin system in domestic animals: what we know and what we still need to understand of its role in reproduction. Domest Anim Endocrinol. 2020;73:106466. [CrossRef]
- Xie Q, Kang Y, Zhang C, Xie Y, Wang C, Liu J, Yu C, Zhao H, Huang D. The Role of Kisspeptin in the Control of the Hypothalamic-Pituitary-Gonadal Axis and Reproduction. Front Endocrinol (Lausanne). 2022;13:925206. [CrossRef]
- Uenoyama Y, Nagae M, Tsuchida H, Inoue N, Tsukamura H. Role of KNDy Neurons Expressing Kisspeptin, Neurokinin B, and Dynorphin A as a GnRH Pulse Generator Controlling Mammalian Reproduction. Front Endocrinol (Lausanne). 2021;12: 724632. [CrossRef]
- Oride A, Kanasaki H. The role of KNDy neurons in human reproductive health. Endocr J. 2024;71(8):733-743. [CrossRef]
- Moore AM, Novak AG, Lehman MN. KNDy Neurons of the Hypothalamus and Their Role in GnRH Pulse Generation: an Update. Endocrinology. 2023 Dec;165(2): bqad194. [CrossRef]
- Hrabovszky E, Sipos MT, Molnár CS, Ciofi P, Borsay BÁ, Gergely P, Herczeg L, Bloom SR, Ghatei MA, Dhillo WS, Liposits Z. Low degree of overlap between kisspeptin, neurokinin B, and dynorphin immunoreactivities in the infundibular nucleus of young male human subjects challenges the KNDy neuron concept. Endocrinology. 2012;153(10):4978-89. [CrossRef]
- Lehman MN, He W, Coolen LM, Levine JE, Goodman RL. Does the KNDy Model for the Control of Gonadotropin-Releasing Hormone Pulses Apply to Monkeys and Humans? Semin Reprod Med. 2019;37(2):71-83. [CrossRef]
- Navarro, VM. Metabolic regulation of kisspeptin - the link between energy balance and reproduction. Nat Rev Endocrinol. 2020;16(8):407-420. [CrossRef]
- Nandankar N, Negrón AL, Wolfe A, Levine JE, Radovick S. Deficiency of arcuate nucleus kisspeptin results in postpubertal central hypogonadism. Am J Physiol Endocrinol Metab. 2021;321(2):E264-E280. [CrossRef]
- Campideli-Santana AC, Gusmao DO, Almeida FRCL, Araujo-Lopes R, Szawka RE. Partial loss of arcuate kisspeptin neurons in female rats stimulates luteinizing hormone and decreases prolactin secretion induced by estradiol. J Neuroendocrinol. 2022; 34(11):e13204. [CrossRef]
- Szukiewicz, D. Current Insights in Prolactin Signaling and Ovulatory Function. Int J Mol Sci. 2024;25(4):1976. [CrossRef]
- Torres E, Pellegrino G, Granados-Rodríguez M, Fuentes-Fayos AC, Velasco I, Coutteau-Robles A, Legrand A, Shanabrough M, Perdices-Lopez C, Leon S, Yeo SH, Manchishi SM, Sánchez-Tapia MJ, Navarro VM, Pineda R, Roa J, Naftolin F, Argente J, Luque RM, Chowen JA, Horvath TL, Prevot V, Sharif A, Colledge WH, Tena-Sempere M, Romero-Ruiz A. Kisspeptin signaling in astrocytes modulates the reproductive axis. J Clin Invest. 2024;134(15):e172908. [CrossRef]
- Huang Y, Liu Q, Huang G, Wen J, Chen G. Hypothalamic Kisspeptin Neurons Regulates Energy Metabolism and Reproduction Under Chronic Stress. Front Endocrinol (Lausanne). 2022;13:844397. [CrossRef]
- Harter CJL, Kavanagh GS, Smith JT. The role of kisspeptin neurons in reproduction and metabolism. J Endocrinol. 2018;238(3):R173-R183. [CrossRef]
- Patel R, Smith JT. Novel actions of kisspeptin signaling outside of GnRH-mediated fertility: a potential role in energy balance. Domest Anim Endocrinol. 2020;73: 106467. [CrossRef]
- Oyedokun PA, Akangbe MA, Akhigbe TM, Akhigbe RE. Regulatory Involvement of Kisspeptin in Energy Balance and Reproduction. Cell Biochem Biophys. 2025;83(1): 247-261. [CrossRef]
- Dhillo WS, Murphy KG, Bloom SR. The neuroendocrine physiology of kisspeptin in the human. Rev Endocr Metab Disord. 2007;8(1):41-6. [CrossRef]
- Bhattacharya M, Babwah AV. Kisspeptin: beyond the brain. Endocrinology. 2015; 156(4):1218-27. [CrossRef]
- Hu KL, Zhao H, Yu Y, Li R. Kisspeptin as a potential biomarker throughout pregnancy. Eur J Obstet Gynecol Reprod Biol. 2019;240:261-266. [CrossRef]
- Dhillo WS, Savage P, Murphy KG, Chaudhri OB, Patterson M, Nijher GM, Foggo VM, Dancey GS, Mitchell H, Seckl MJ, Ghatei MA, Bloom SR. Plasma kisspeptin is raised in patients with gestational trophoblastic neoplasia and falls during treatment. Am J Physiol Endocrinol Metab. 2006;291(5):E878-84. [CrossRef]
- Tsoutsouki J, Patel B, Comninos AN, Dhillo WS, Abbara A. Kisspeptin in the Prediction of Pregnancy Complications. Front Endocrinol (Lausanne). 2022;13: 942664. [CrossRef]
- Li C, Yuan L, Han S, Xuan M, Liu D, Tian B, Yu W. Reduced Kiss-1 expression is associated with clinical aggressive feature of gastric cancer patients and promotes migration and invasion in gastric cancer cells. Oncol Rep. 2020;44(3):1149-1157. [CrossRef]
- Wu HM, Chen LH, Chiu WJ, Tsai CL. Kisspeptin Regulates Cell Invasion and Migration in Endometrial Cancer. J Endocr Soc. 2024;8(3):bvae001. [CrossRef]
- Loosen SH, Luedde M, Lurje G, Spehlmann M, Paffenholz P, Ulmer TF, Tacke F, Vucur M, Trautwein C, Neumann UP, Luedde T, Roderburg C. Serum Levels of Kisspeptin Are Elevated in Patients with Pancreatic Cancer. Dis Markers. 2019; 2019:5603474. [CrossRef]
- Kim CW, Lee HK, Nam MW, Choi Y, Choi KC. Overexpression of KiSS1 Induces the Proliferation of Hepatocarcinoma and Increases Metastatic Potential by Increasing Migratory Ability and Angiogenic Capacity. Mol Cells. 2022;45(12):935-949. [CrossRef]
- Zhu N, Zhao M, Song Y, Ding L, Ni Y. The KiSS-1/GPR54 system: Essential roles in physiological homeostasis and cancer biology. Genes Dis. 2020;9(1):28-40. [CrossRef]
- Ulasov IV, Borovjagin AV, Timashev P, Cristofanili M, Welch DR. KISS1 in breast cancer progression and autophagy. Cancer Metastasis Rev. 2019;38(3):493-506. [CrossRef]
- Guzman S, Brackstone M, Wondisford F, Babwah AV, Bhattacharya M. KISS1/KISS1R and Breast Cancer: Metastasis Promoter. Semin Reprod Med. 2019;37(4):197-206. [CrossRef]
- Ly T, Harihar S, Welch DR. KISS1 in metastatic cancer research and treatment: potential and paradoxes. Cancer Metastasis Rev. 2020;39(3):739-754. [CrossRef]
- Fratangelo F, Carriero MV, Motti ML. Controversial Role of Kisspeptins/KiSS-1R Signaling System in Tumor Development. Front Endocrinol (Lausanne). 2018;9:192. [CrossRef]
- Stathaki M, Stamatiou ME, Magioris G, Simantiris S, Syrigos N, Dourakis S, Koutsilieris M, Armakolas A. The role of kisspeptin system in cancer biology. Crit Rev Oncol Hematol. 2019;142:130-140. [CrossRef]
- Hiden U, Bilban M, Knöfler M, Desoye G. Kisspeptins and the placenta: regulation of trophoblast invasion. Rev Endocr Metab Disord. 2007;8(1):31-9. [CrossRef]
- Santos BR, Dos Anjos Cordeiro JM, Santos LC, Barbosa EM, Mendonça LD, Santos EO, de Macedo IO, de Lavor MSL, Szawka RE, Serakides R, Silva JF. Kisspeptin treatment improves fetal-placental development and blocks placental oxidative damage caused by maternal hypothyroidism in an experimental rat model. Front Endocrinol (Lausanne). 2022; 13: 908240. [CrossRef]
- Taylor J, Pampillo M, Bhattacharya M, Babwah AV. Kisspeptin/KISS1R signaling potentiates extravillous trophoblast adhesion to type-I collagen in a PKC- and ERK1/2-dependent manner. Mol Reprod Dev. 2014;81(1):42-54. [CrossRef]
- Cockwell H, Wilkinson DA, Bouzayen R, Imran SA, Brown R, Wilkinson M. KISS1 expression in human female adipose tissue. Arch Gynecol Obstet. 2013;287(1):143-7. [CrossRef]
- Dudek M, Kołodziejski PA, Pruszyńska-Oszmałek E, Sassek M, Ziarniak K, Nowak KW, Sliwowska JH. Effects of high-fat diet-induced obesity and diabetes on Kiss1 and GPR54 expression in the hypothalamic-pituitary-gonadal (HPG) axis and peripheral organs (fat, pancreas and liver) in male rats. Neuropeptides. 2016;56:41-9. [CrossRef]
- Pruszyńska-Oszmałek E, Wojciechowska M, Krauss H, Sassek M, Leciejewska N, Szczepankiewicz D, Nowak KW, Piątek J, Nogowski L, Sliwowska JH, Kołodziejski PA. Obesity is associated with increased level of kisspeptin in mothers' blood and umbilical cord blood - a pilot study. Eur Rev Med Pharmacol Sci. 2021;25(19):5993-6002. [CrossRef]
- Wolfe A, Hussain MA. The Emerging Role(s) for Kisspeptin in Metabolism in Mammals. Front Endocrinol (Lausanne). 2018;9:184. [CrossRef]
- Sliwowska JH, Woods NE, Alzahrani AR, Paspali E, Tate RJ, Ferro VA. Kisspeptin a potential therapeutic target in treatment of both metabolic and reproductive dysfunction. J Diabetes. 2024;16(4):e13541. [CrossRef]
- Gottsch ML, Clifton DK, Steiner RA. From KISS1 to kisspeptins: An historical perspective and suggested nomenclature. Peptides. 2009;30(1):4-9. [CrossRef]
- Kirby HR, Maguire JJ, Colledge WH, Davenport AP. International Union of Basic and Clinical Pharmacology. LXXVII. Kisspeptin receptor nomenclature, distribution, and function. Pharmacol Rev. 2010;62(4):565-78. [CrossRef]
- Wu Z, Chen G, Qiu C, Yan X, Xu L, Jiang S, Xu J, Han R, Shi T, Liu Y, Gao W, Wang Q, Li J, Ye F, Pan X, Zhang Z, Ning P, Zhang B, Chen J, Du Y. Structural basis for the ligand recognition and G protein subtype selectivity of kisspeptin receptor. Sci Adv. 2024;10(33):eadn7771. [CrossRef]
- Muir AI, Chamberlain L, Elshourbagy NA, Michalovich D, Moore DJ, Calamari A, Szekeres PG, Sarau HM, Chambers JK, Murdock P, Steplewski K, Shabon U, Miller JE, Middleton SE, Darker JG, Larminie CG, Wilson S, Bergsma DJ, Emson P, Faull R, Philpott KL, Harrison DC. AXOR12, a novel human G protein-coupled receptor, activated by the peptide KiSS-1. J Biol Chem. 2001;276(31):28969-75. [CrossRef]
- Acierno JS Jr, Shagoury JK, Bo-Abbas Y, Crowley WF Jr, Seminara SB. A locus for autosomal recessive idiopathic hypogonadotropic hypogonadism on chromosome 19p13.3. J Clin Endocrinol Metab. 2003;88(6):2947-50. [CrossRef]
- Cardoso JC, Félix RC, Bjärnmark N, Power DM. Allatostatin-type A, kisspeptin and galanin GPCRs and putative ligands as candidate regulatory factors of mantle function. Mar Genomics. 2016;27:25-35. [CrossRef]
- Quillet R, Ayachi S, Bihel F, Elhabazi K, Ilien B, Simonin F. RF-amide neuropeptides and their receptors in Mammals: Pharmacological properties, drug development and main physiological functions. Pharmacol Ther. 2016;160:84-132. [CrossRef]
- Hasegawa K, Maedomari R, Sato Y, Gotoh K, Kudoh S, Kojima A, Okada S, Ito T. Kiss1R Identification and Biodistribution Analysis Employing a Western Ligand Blot and Ligand-Derivative Stain with a FITC-Kisspeptin Derivative. ChemMedChem. 2020;15(18):1699-1705. Erratum in: ChemMedChem. 2021;16(4):725. doi: 10.1002/cmdc.202000990. [CrossRef]
- Pasquier J, Kamech N, Lafont AG, Vaudry H, Rousseau K, Dufour S. Molecular evolution of GPCRs: Kisspeptin/kisspeptin receptors. J Mol Endocrinol. 2014;52(3): T101-17. [CrossRef]
- Kotani M, Detheux M, Vandenbogaerde A, Communi D, Vanderwinden JM, Le Poul E, Brézillon S, Tyldesley R, Suarez-Huerta N, Vandeput F, Blanpain C, Schiffmann SN, Vassart G, Parmentier M. The metastasis suppressor gene KiSS-1 encodes kisspeptins, the natural ligands of the orphan G protein-coupled receptor GPR54. J Biol Chem. 2001;276(37):34631-6. [CrossRef]
- Wang Y, Qin M, Fan L, Gong C. Correlation Analysis of Genotypes and Phenotypes in Chinese Male Pediatric Patients With Congenital Hypogonadotropic Hypogonadism. Front Endocrinol (Lausanne). 2022;13:846801. [CrossRef]
- Chen X, Hu M, Du T, Yang L, Li Y, Feng L, Luo J, Yao H, Chen X. Homozygous mutation of KISS1 receptor (KISS1R) gene identified in a Chinese patient with congenital hypogonadotropic hypogonadism (CHH): case report and literature review. J Pediatr Endocrinol Metab. 2024;37(11):999-1008. [CrossRef]
- Geng D, Zhang H, Liu X, Fei J, Jiang Y, Liu R, Wang R, Zhang G. Identification of KISS1R gene mutations in disorders of non-obstructive azoospermia in the northeast population of China. J Clin Lab Anal. 2020;34(4):e23139. [CrossRef]
- Schöneberg T, Liebscher I. Mutations in G Protein-Coupled Receptors: Mechanisms, Pathophysiology and Potential Therapeutic Approaches. Pharmacol Rev. 2021;73(1):89-119. [CrossRef]
- Heydenreich FM, Marti-Solano M, Sandhu M, Kobilka BK, Bouvier M, Babu MM. Molecular determinants of ligand efficacy and potency in GPCR signaling. Science. 2023;382(6677):eadh1859. [CrossRef]
- Roa J, Aguilar E, Dieguez C, Pinilla L, Tena-Sempere M. New frontiers in kisspeptin/GPR54 physiology as fundamental gatekeepers of reproductive function. Front Neuroendocrinol. 2008;29(1):48-69. [CrossRef]
- Abbara A, Clarke SA, Dhillo WS. Clinical Potential of Kisspeptin in Reproductive Health. Trends Mol Med. 2021;27(8):807-823. [CrossRef]
- Gianetti E, Seminara S. Kisspeptin and KISS1R: a critical pathway in the reproductive system. Reproduction. 2008;136(3):295-301. [CrossRef]
- Franssen D, Tena-Sempere M. The kisspeptin receptor: A key G-protein-coupled receptor in the control of the reproductive axis. Best Pract Res Clin Endocrinol Metab. 2018;32(2):107-123. [CrossRef]
- Shen S, Wang D, Liu H, He X, Cao Y, Chen J, Li S, Cheng X, Xu HE, Duan J. Structural basis for hormone recognition and distinctive Gq protein coupling by the kisspeptin receptor. Cell Rep. 2024;43(7):114389. [CrossRef]
- Lyon AM, Tesmer JJ. Structural insights into phospholipase C-β function. Mol Pharmacol. 2013; 84(4):488-500. [CrossRef]
- Kanemaru K, Nakamura Y. Activation Mechanisms and Diverse Functions of Mammalian Phospholipase C. Biomolecules. 2023;13(6):915. [CrossRef]
- Ubeysinghe S, Wijayaratna D, Kankanamge D, Karunarathne A. Molecular regulation of PLCβ signaling. Methods Enzymol. 2023;682:17-52. [CrossRef]
- Paknejad N, Hite RK. Structural basis for the regulation of inositol trisphosphate receptors by Ca2+ and IP3. Nat Struct Mol Biol. 2018;25(8):660-668. Erratum in: Nat Struct Mol Biol. 2018;25(9):902. doi: 10.1038/s41594-018-0119-4. [CrossRef]
- Schmitz EA, Takahashi H, Karakas E. Structural basis for activation and gating of IP3 receptors. Nat Commun. 2022;13(1):1408. [CrossRef]
- Lučić I, Truebestein L, Leonard TA. Novel Features of DAG-Activated PKC Isozymes Reveal a Conserved 3-D Architecture. J Mol Biol. 2016;428(1):121-141. [CrossRef]
- Kolczynska K, Loza-Valdes A, Hawro I, Sumara G. Diacylglycerol-evoked activation of PKC and PKD isoforms in regulation of glucose and lipid metabolism: a review. Lipids Health Dis. 2020; 19(1):113. [CrossRef]
- Krebs J, Agellon LB, Michalak M. Ca(2+) homeostasis and endoplasmic reticulum (ER) stress: An integrated view of calcium signaling. Biochem Biophys Res Commun. 2015;460(1):114-21. [CrossRef]
- Bagur R, Hajnóczky G. Intracellular Ca2+ Sensing: Its Role in Calcium Homeostasis and Signaling. Mol Cell. 2017;66(6):780-788. [CrossRef]
- Peng J, Tang M, Zhang BP, Zhang P, Zhong T, Zong T, Yang B, Kuang HB. Kisspeptin stimulates progesterone secretion via the Erk1/2 mitogen-activated protein kinase signaling pathway in rat luteal cells. Fertil Steril. 2013;99(5):1436-1443.e1. [CrossRef]
- Kim GL, Dhillon SS, Belsham DD. Kisspeptin directly regulates neuropeptide Y synthesis and secretion via the ERK1/2 and p38 mitogen-activated protein kinase signaling pathways in NPY-secreting hypothalamic neurons. Endocrinology. 2010;151(10):5038-47. [CrossRef]
- Yang C, Kazanietz MG. Divergence and complexities in DAG signaling: looking beyond PKC. Trends Pharmacol Sci. 2003;24(11):602-8. [CrossRef]
- Szereszewski JM, Pampillo M, Ahow MR, Offermanns S, Bhattacharya M, Babwah AV. GPR54 regulates ERK1/2 activity and hypothalamic gene expression in a Gα(q/11) and β-arrestin-dependent manner. PLoS One. 2010;5(9):e12964. [CrossRef]
- Ahow M, Min L, Pampillo M, Nash C, Wen J, Soltis K, Carroll RS, Glidewell-Kenney CA, Mellon PL, Bhattacharya M, Tobet SA, Kaiser UB, Babwah AV. KISS1R signals independently of Gαq/11 and triggers LH secretion via the β-arrestin pathway in the male mouse. Endocrinology. 2014; 155(11):4433-46. [CrossRef]
- Pampillo M, Camuso N, Taylor JE, Szereszewski JM, Ahow MR, Zajac M, Millar RP, Bhattacharya M, Babwah AV. Regulation of GPR54 signaling by GRK2 and {beta}-arrestin. Mol Endocrinol. 2009;23(12):2060-74. [CrossRef]
- Kahsai AW, Shah KS, Shim PJ, Lee MA, Shreiber BN, Schwalb AM, Zhang X, Kwon HY, Huang LY, Soderblom EJ, Ahn S, Lefkowitz RJ. Signal transduction at GPCRs: Allosteric activation of the ERK MAPK by β-arrestin. Proc Natl Acad Sci U S A. 2023;120(43):e2303794120. [CrossRef]
- Hu KL, Zhao H, Chang HM, Yu Y, Qiao J. Kisspeptin/Kisspeptin Receptor System in the Ovary. Front Endocrinol (Lausanne). 2018;8:365. [CrossRef]
- Gan DM, Zhang PP, Zhang JP, Ding SX, Fang J, Liu Y. KISS1/KISS1R mediates Sertoli cell apoptosis via the PI3K/AKT signalling pathway in a high-glucose environment. Mol Med Rep. 2021; 23(6):477. [CrossRef]
- Yabluchanskiy A, Ma Y, Iyer RP, Hall ME, Lindsey ML. Matrix metalloproteinase-9: Many shades of function in cardiovascular disease. Physiology (Bethesda). 2013;28(6):391-403. [CrossRef]
- Wahab F, Atika B, Shahab M, Behr R. Kisspeptin signalling in the physiology and pathophysiology of the urogenital system. Nat Rev Urol. 2016;13(1):21-32. [CrossRef]
- Quintero-Fabián S, Arreola R, Becerril-Villanueva E, Torres-Romero JC, Arana-Argáez V, Lara-Riegos J, Ramírez-Camacho MA, Alvarez-Sánchez ME. Role of Matrix Metalloproteinases in Angiogenesis and Cancer. Front Oncol. 2019;9:1370. [CrossRef]
- Li X, Wu C, Shen Y, Wang K, Tang L, Zhou M, Yang M, Pan T, Liu X, Xu W. Ten-eleven translocation 2 demethylates the MMP9 promoter, and its down-regulation in preeclampsia impairs trophoblast migration and invasion. J Biol Chem. 2018;293(26):10059-10070. [CrossRef]
- Plaks V, Rinkenberger J, Dai J, Flannery M, Sund M, Kanasaki K, Ni W, Kalluri R, Werb Z. Matrix metalloproteinase-9 deficiency phenocopies features of preeclampsia and intrauterine growth restriction. Proc Natl Acad Sci U S A. 2013;110(27):11109-14. [CrossRef]
- Yan Y, Fang L, Li Y, Yu Y, Li Y, Cheng JC, Sun YP. Association of MMP2 and MMP9 gene polymorphisms with the recurrent spontaneous abortion: A meta-analysis. Gene. 2021;767:145173. [CrossRef]
- Chiu KL, He JL, Chen GL, Shen TC, Chen LH, Chen JC, Tsai CW, Chang WS, Hsia TC, Bau DT. Impacts of Matrix Metalloproteinase-9 Promoter Genotypes on Asthma Risk. In Vivo. 2024;38(5): 2144-2151. [CrossRef]
- Pulito-Cueto V, Atienza-Mateo B, Batista-Liz JC, Sebastián Mora-Gil M, Mora-Cuesta VM, Iturbe-Fernández D, Izquierdo Cuervo S, Aguirre Portilla C, Blanco R, López-Mejías R. Matrix metalloproteinases and their tissue inhibitors as upcoming biomarker signatures of connective tissue diseases-related interstitial lung disease: towards an earlier and accurate diagnosis. Mol Med. 2025; 31(1):70. [CrossRef]
- Bilban M, Ghaffari-Tabrizi N, Hintermann E, Bauer S, Molzer S, Zoratti C, Malli R, Sharabi A, Hiden U, Graier W, Knöfler M, Andreae F, Wagner O, Quaranta V, Desoye G. Kisspeptin-10, a KiSS-1/metastin-derived decapeptide, is a physiological invasion inhibitor of primary human trophoblasts. J Cell Sci. 2004;117(Pt 8):1319-28. [CrossRef]
- Roseweir AK, Katz AA, Millar RP. Kisspeptin-10 inhibits cell migration in vitro via a receptor-GSK3 beta-FAK feedback loop in HTR8SVneo cells. Placenta. 2012;33(5):408-15. [CrossRef]
- Francis VA, Abera AB, Matjila M, Millar RP, Katz AA. Kisspeptin regulation of genes involved in cell invasion and angiogenesis in first trimester human trophoblast cells. PLoS One. 2014;9(6):e99680. [CrossRef]
- Moore KL, Persaud TVN, The developing human: clinically oriented embryology, 7th edition, Saunders, 2003:520.
- Thowfeequ S, Srinivas S. Embryonic and extraembryonic tissues during mammalian development: shifting boundaries in time and space. Philos Trans R Soc Lond B Biol Sci. 2022;377(1865):20210255. [CrossRef]
- Donovan MF, Cascella M. Embryology, Weeks 6-8. [Updated 2022 Oct 10]. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan- [(accessed on 10 May 2025)]. Available from: https://www.ncbi.nlm.nih.gov/books/NBK563181/.
- Findlay JK, Gear ML, Illingworth PJ, Junk SM, Kay G, Mackerras AH, Pope A, Rothenfluh HS, Wilton L. Human embryo: a biological definition. Hum Reprod. 2007; 22(4):905-11. [CrossRef]
- Gorbunova O, Shirshev S. The effect of kisspeptin on the functional activity of peripheral blood monocytes and neutrophils in the context of physiological pregnancy. J Reprod Immunol. 2022;151:103621. [CrossRef]
- Katirci Y, Kocaman A, Ozdemir AZ. Kisspeptin expression levels in patients with placenta previa: A randomized trial. Medicine (Baltimore). 2024;103(28):e38866. [CrossRef]
- Kapustin RV, Drobintseva AO, Alekseenkova EN, Onopriychuk AR, Arzhanova ON, Polyakova VO, Kvetnoy IM. Placental protein expression of kisspeptin-1 (KISS1) and the kisspeptin-1 receptor (KISS1R) in pregnancy complicated by diabetes mellitus or preeclampsia. Arch Gynecol Obstet. 2020;301(2):437-445. [CrossRef]
- Hu KL, Zhang Y, Yang Z, Zhao H, Xu H, Yu Y, Li R. Predictive value of serum kisspeptin concentration at 14 and 21 days after frozen-thawed embryo transfer. Reprod Biomed Online. 2019;39(1):161-167. [CrossRef]
- Ye S, Zhou L. Role of serum kisspeptin as a biomarker to detect miscarriage: a systematic review and meta-analysis. Hum Fertil (Camb). 2024;27(1):2417934. [CrossRef]
- Silva PHAD, Romão LGM, Freitas NPA, Carvalho TR, Porto MEMP, Araujo Júnior E, Cavalcante MB. Kisspeptin as a predictor of miscarriage: a systematic review. J Matern Fetal Neonatal Med. 2023;36(1):2197097. [CrossRef]
- Guo J, Feng Q, Chaemsaithong P, Appiah K, Sahota DS, Leung BW, Chung JP, Li TC, Poon LC. Biomarkers at 6 weeks' gestation in the prediction of early miscarriage in pregnancy following assisted reproductive technology. Acta Obstet Gynecol Scand. 2023;102(8):1073-1083. [CrossRef]
- Abbara A, Al-Memar M, Phylactou M, Kyriacou C, Eng PC, Nadir R, et al. Performance of Plasma Kisspeptin as a Biomarker for Miscarriage Improves With Gestational Age During the First Trimester. Fertil Steril 2021;116(3):809–19. [CrossRef]
- Mumtaz A, Khalid A, Jamil Z, Fatima SS, Arif S, Rehman R. Kisspeptin: A Potential Factor for Unexplained Infertility and Impaired Embryo Implantation. Int J Fertil Steril. 2017;11(2):99-104. [CrossRef]
- Kim SM, Kim JS. A Review of Mechanisms of Implantation. Dev Reprod. 2017;21(4): 351-359. [CrossRef]
- Cohen M, Bischof P. Factors regulating trophoblast invasion. Gynecol Obstet Invest. 2007;64(3):126-30. [CrossRef]
- Zhang S, Xiao Y, Wang Y, Qian C, Zhang R, Liu J, Wang Q, Zhang H. Role of kisspeptin in decidualization and unexplained recurrent spontaneous abortion via the ERK1/2 signalling pathway. Placenta. 2023;133:1-9. [CrossRef]
- Saadeldin IM, Koo OJ, Kang JT, Kwon DK, Park SJ, Kim SJ, Moon JH, Oh HJ, Jang G, Lee BC. Paradoxical effects of kisspeptin: it enhances oocyte in vitro maturation but has an adverse impact on hatched blastocysts during in vitro culture. Reprod Fertil Dev. 2012;24(5):656-68. [CrossRef]
- Dorfman MD, Garcia-Rudaz C, Alderman Z, Kerr B, Lomniczi A, Dissen GA, Castellano JM, Garcia-Galiano D, Gaytan F, Xu B, Tena-Sempere M, Ojeda SR. Loss of Ntrk2/Kiss1r signaling in oocytes causes premature ovarian failure. Endocrinology. 2014;155(8):3098-111. [CrossRef]
- Pinto FM, Cejudo-Román A, Ravina CG, Fernández-Sánchez M, Martín-Lozano D, Illanes M, Tena-Sempere M, Candenas ML. Characterization of the kisspeptin system in human spermatozoa. Int J Androl. 2012;35(1):63-73. [CrossRef]
- Yan L, Yang M, Guo H, Yang L, Wu J, Li R, Liu P, Lian Y, Zheng X, Yan J, Huang J, Li M, Wu X, Wen L, Lao K, Li R, Qiao J, Tang F. Single-cell RNA-Seq profiling of human preimplantation embryos and embryonic stem cells. Nat Struct Mol Biol. 2013;20(9):1131-9. [CrossRef]
- Bischof P, Campana A. Molecular mediators of implantation. Baillieres Best Pract Res Clin Obstet Gynaecol. 2000;14(5):801-14. [CrossRef]
- Kleimenova T, Polyakova V, Linkova N, Drobintseva A, Medvedev D, Krasichkov A. The Expression of Kisspeptins and Matrix Metalloproteinases in Extragenital Endometriosis. Biomedicines. 2024;12(1):94. [CrossRef]
- Babwah, AV. Uterine and placental KISS1 regulate pregnancy: what we know and the challenges that lie ahead. Reproduction. 2015;150(4):R121-8. [CrossRef]
- Okada H, Tsuzuki T, Murata H. Decidualization of the human endometrium. Reprod Med Biol. 2018;17(3):220-227. [CrossRef]
- Calder M, Chan YM, Raj R, Pampillo M, Elbert A, Noonan M, Gillio-Meina C, Caligioni C, Bérubé NG, Bhattacharya M, Watson AJ, Seminara SB, Babwah AV. Implantation failure in female Kiss1-/- mice is independent of their hypogonadic state and can be partially rescued by leukemia inhibitory factor. Endocrinology. 2014;155(8):3065-78. [CrossRef]
- Shuya LL, Menkhorst EM, Yap J, Li P, Lane N, Dimitriadis E. Leukemia inhibitory factor enhances endometrial stromal cell decidualization in humans and mice. PLoS One. 2011;6(9):e25288. [CrossRef]
- Zhang P, Tang M, Zhong T, Lin Y, Zong T, Zhong C, Zhang B, Ren M, Kuang H. Expression and function of kisspeptin during mouse decidualization. PLoS One. 2014;9(5):e97647. [CrossRef]
- Yang Y, Zhang J, Li D, Liu C, Guo R, Xiao Y, Zhou L, Tong L, Zhang H. kisspeptin调控Notch1/Akt/Foxo1通路参与复发性流产患者子宫内膜蜕膜化 [Notch1/Akt/Foxo1 Pathway Regulated by Kisspeptin Is Involved in Endometrial Decidualization in Patients With Recurrent Spontaneous Abortion]. Sichuan Da Xue Xue Bao Yi Xue Ban. 2024;55(3):542-551. Chinese. [CrossRef]
- Schaefer J, Vilos AG, Vilos GA, Bhattacharya M, Babwah AV. Uterine kisspeptin receptor critically regulates epithelial estrogen receptor α transcriptional activity at the time of embryo implantation in a mouse model. Mol Hum Reprod. 2021;27(10): gaab060. [CrossRef]
- León S, Fernandois D, Sull A, Sull J, Calder M, Hayashi K, Bhattacharya M, Power S, Vilos GA, Vilos AG, Tena-Sempere M, Babwah AV. Beyond the brain-Peripheral kisspeptin signaling is essential for promoting endometrial gland development and function. Sci Rep. 2016;6:29073. Erratum in: Sci Rep. 2016;6:30954. doi: 10.1038/srep30954. [CrossRef]
- Yang Y, Song S, Gu S, Gu Y, Zhao P, Li D, Cheng W, Liu C, Zhang H. Kisspeptin prevents pregnancy loss by modulating the immune microenvironment at the maternal-fetal interface in recurrent spontaneous abortion. Am J Reprod Immunol. 2024;91(2):e13818. [CrossRef]
- Williams, Z. Inducing tolerance to pregnancy. N Engl J Med. 2012;367(12):1159-61. [CrossRef]
- Gorbunova OL, Shirshev SV. Molecular mechanisms of the regulation by kisspeptin of the formation and functional activity of Treg and Th17. Biochem Moscow 2016;Suppl. Ser. A 10, 180–187. [CrossRef]
- Lyall F, Bulmer JN, Duffie E, Cousins F, Theriault A, Robson SC. Human trophoblast invasion and spiral artery transformation: the role of PECAM-1 in normal pregnancy, preeclampsia, and fetal growth restriction. Am J Pathol. 2001;158(5):1713-21. [CrossRef]
- Moser G, Windsperger K, Pollheimer J, de Sousa Lopes SC, Huppertz B. Human trophoblast invasion: new and unexpected routes and functions. Histochem Cell Biol. 2018; 150(4):361-370. [CrossRef]
- James JL, Saghian R, Perwick R, Clark AR. Trophoblast plugs: impact on utero-placental haemodynamics and spiral artery remodelling. Hum Reprod. 2018;33(8): 1430-1441. [CrossRef]
- Saghian R, Bogle G, James JL, Clark AR. Establishment of maternal blood supply to the placenta: insights into plugging, unplugging and trophoblast behaviour from an agent-based model. Interface Focus. 2019;9(5):20190019. [CrossRef]
- Velicky P, Knöfler M, Pollheimer J. Function and control of human invasive trophoblast subtypes: Intrinsic vs. maternal control. Cell Adh Migr. 2016;10(1-2):154-62. [CrossRef]
- Barrientos G, Pussetto M, Rose M, Staff AC, Blois SM, Toblli JE. Defective trophoblast invasion underlies fetal growth restriction and preeclampsia-like symptoms in the stroke-prone spontaneously hypertensive rat. Mol Hum Reprod. 2017;23(7):509-519. [CrossRef]
- Huppertz, B. The Critical Role of Abnormal Trophoblast Development in the Etiology of Preeclampsia. Curr Pharm Biotechnol. 2018;19(10):771-780. [CrossRef]
- Staff AC, Fjeldstad HE, Fosheim IK, Moe K, Turowski G, Johnsen GM, Alnaes-Katjavivi P, Sugulle M. Failure of physiological transformation and spiral artery atherosis: their roles in preeclampsia. Am J Obstet Gynecol. 2022;226(2S):S895-S906. [CrossRef]
- Li Z, Liu J, Inuzuka H, Wei W. Functional analysis of the emerging roles for the KISS1/KISS1R signaling pathway in cancer metastasis. J Genet Genomics. 2022;49(3):181-184. [CrossRef]
- Harihar S, Welch DR. KISS1 metastasis suppressor in tumor dormancy: a potential therapeutic target for metastatic cancers? Cancer Metastasis Rev. 2023;42(1):183-196. [CrossRef]
- Anacker J, Segerer SE, Hagemann C, Feix S, Kapp M, Bausch R, Kämmerer U. Human decidua and invasive trophoblasts are rich sources of nearly all human matrix metalloproteinases. Mol Hum Reprod. 2011;17(10):637-52. [CrossRef]
- Hisamatsu Y, Murata H, Tsubokura H, Hashimoto Y, Kitada M, Tanaka S, Okada H. Matrix Metalloproteinases in Human Decidualized Endometrial Stromal Cells. Curr Issues Mol Biol. 2021;43(3):2111-2123. [CrossRef]
- Janneau JL, Maldonado-Estrada J, Tachdjian G, Miran I, Motté N, Saulnier P, Sabourin JC, Coté JF, Simon B, Frydman R, Chaouat G, Bellet D. Transcriptional expression of genes involved in cell invasion and migration by normal and tumoral trophoblast cells. J Clin Endocrinol Metab. 2002;87(11):5336-9. [CrossRef]
- Ferretti C, Bruni L, Dangles-Marie V, Pecking AP, Bellet D. Molecular circuits shared by placental and cancer cells, and their implications in the proliferative, invasive and migratory capacities of trophoblasts. Hum Reprod Update. 2007;13(2):121-41. [CrossRef]
- Gomes VCL, Sones JL. From inhibition of trophoblast cell invasion to proapoptosis: what are the potential roles of kisspeptins in preeclampsia? Am J Physiol Regul Integr Comp Physiol. 2021;321(1):R41-R48. [CrossRef]
- Gomes VCL, Woods AK, Crissman KR, Landry CA, Beckers KF, Gilbert BM, Ferro LR, Liu CC, Oberhaus EL, Sones JL. Kisspeptin Is Upregulated at the Maternal-Fetal Interface of the Preeclamptic-like BPH/5 Mouse and Normalized after Synchronization of Sex Steroid Hormones. Reprod Med (Basel). 2022;3(4):263-279. [CrossRef]
- Todros T, Paulesu L, Cardaropoli S, Rolfo A, Masturzo B, Ermini L, Romagnoli R, Ietta F. Role of the Macrophage Migration Inhibitory Factor in the Pathophysiology of Pre-Eclampsia. Int J Mol Sci. 2021;22(4):1823. [CrossRef]
- Ietta F, Ferro EAV, Bevilacqua E, Benincasa L, Maioli E, Paulesu L. Role of the Macrophage Migration Inhibitory Factor (MIF) in the survival of first trimester human placenta under induced stress conditions. Sci Rep. 2018;8(1):12150. [CrossRef]
- Martino NA, Rizzo A, Pizzi F, Dell'Aquila ME, Sciorsci RL. Effects of kisspeptin-10 on in vitro proliferation and kisspeptin receptor expression in primary epithelial cell cultures isolated from bovine placental cotyledons of fetuses at the first trimester of pregnancy. Theriogenology. 2015;83(6):978-987.e1. [CrossRef]
- Zhai J, Liu J, Zhao S, Zhao H, Chen ZJ, Du Y, Li W. Kisspeptin-10 inhibits OHSS by suppressing VEGF secretion. Reproduction. 2017;154(4):355-362. [CrossRef]
- Santos LC, Dos Anjos Cordeiro JM, da Silva Santana L, Santos BR, Barbosa EM, da Silva TQM, Corrêa JMX, Niella RV, Lavor MSL, da Silva EB, de Melo Ocarino N, Serakides R, Silva JF. Kisspeptin/Kiss1r system and angiogenic and immunological mediators at the maternal-fetal interface of domestic cats. Biol Reprod. 2021;105(1):217-231. [CrossRef]
- Cai J, Han X, Peng S, Chen J, Zhang JV, Huang C. Chemerin facilitates placental trophoblast invasion and spiral artery remodeling through the pentose phosphate pathway. Life Sci. 2025 :123645. [CrossRef]
- Jansen CHJR, Kastelein AW, Kleinrouweler CE, Van Leeuwen E, De Jong KH, Pajkrt E, Van Noorden CJF. Development of placental abnormalities in location and anatomy. Acta Obstet Gynecol Scand. 2020;99(8):983-993. [CrossRef]
- Fox, H. (1992). Ectopic Sites of Placentation. In: Barnea, E.R., Hustin, J., Jauniaux, E. (eds) The First Twelve Weeks of Gestation. Springer, Berlin, Heidelberg. [CrossRef]
- Rathbun KM, Hildebrand JP. Placenta Abnormalities. StatPearls [Internet]. [Updated 2022 Oct 17]. StatPearls Publishing; Treasure Island, FL, USA: 2022. [(accessed on 5 May 2025)]. Available online: https://www.ncbi.nlm.nih.gov/books/NBK459355/.
- Ghafouri-Fard S, Shoorei H, Taheri M. The role of microRNAs in ectopic pregnancy: A concise review. Noncoding RNA Res. 2020;5(2):67-70. [CrossRef]
- Romero-Ruiz A, Avendaño MS, Dominguez F, Lozoya T, Molina-Abril H, Sangiao-Alvarellos S, Gurrea M, Lara-Chica M, Fernandez-Sanchez M, Torres-Jimenez E, Perdices-Lopez C, Abbara A, Steffani L, Calzado MA, Dhillo WS, Pellicer A, Tena-Sempere M. Deregulation of miR-324/KISS1/kisspeptin in early ectopic pregnancy: mechanistic findings with clinical and diagnostic implications. Am J Obstet Gynecol. 2019;220(5):480.e1-480.e17. [CrossRef]
- Meena P, Mathur R, Meena ML. Kisspeptin: new biomarker of pregnancy. Int J Reprod Contracept Obstet Gynecol. 2019;8(8):3021–3024. [CrossRef]
- Sullivan-Pyke C, Haisenleder DJ, Senapati S, Nicolais O, Eisenberg E, Sammel MD, Barnhart KT. Kisspeptin as a new serum biomarker to discriminate miscarriage from viable intrauterine pregnancy. Fertil Steril. 2018;109(1):137-141.e2. [CrossRef]
- Jayasena CN, Abbara A, Izzi-Engbeaya C, Comninos AN, Harvey RA, Gonzalez Maffe J, Sarang Z, Ganiyu-Dada Z, Padilha AI, Dhanjal M, Williamson C, Regan L, Ghatei MA, Bloom SR, Dhillo WS. Reduced levels of plasma kisspeptin during the antenatal booking visit are associated with increased risk of miscarriage. J Clin Endocrinol Metab. 2014;99(12):E2652-60. [CrossRef]
- Yuksel S, Ketenci Gencer F. Serum kisspeptin, to discriminate between ectopic pregnancy, miscarriage and first trimester pregnancy. J Obstet Gynaecol. 2022; 42(6):2095-2099. [CrossRef]
- Hu KL, Chen Z, Deng W, Li X, Ju L, Yang H, Zhang H, Mu L. Diagnostic Value of Kisspeptin Levels on Early Pregnancy Outcome: a Systematic Review and Meta-analysis. Reprod Sci. 2022;29(12):3365-3372. [CrossRef]
- Kavvasoglu S, Ozkan ZS, Kumbak B, Sımsek M, Ilhan N. Association of kisspeptin-10 levels with abortus imminens: a preliminary study. Arch Gynecol Obstet. 2012; 285(3):649-53. [CrossRef]
- Park DW, Lee SK, Hong SR, Han AR, Kwak-Kim J, Yang KM. Expression of Kisspeptin and its receptor GPR54 in the first trimester trophoblast of women with recurrent pregnancy loss. Am J Reprod Immunol. 2012;67(2):132-9. [CrossRef]
- Li L, Tian J, Zhou L, Wu S, Zhang S, Qi L, Zhang H. Role of kisspeptin/GPR54 in the first trimester trophoblast of women with a history of recurrent spontaneous abortion. Int J Clin Exp Pathol. 2017;10(8):8161-8173.
Figure 1.
Formation and structure of kisspeptin molecules (inspired by Tena-Sempere [28], modified from López-Ojeda and Hurley [26]). In humans, KISS1 gene located on the long arm of chromosome 1 (1q32), produces a 145-amino acid precursor protein, prepro-kisspeptin, via two coding exons (III. and IV.) of its four exons. Different kisspeptins are generated by proteolytic cleavage of prepro-kisspeptin, with the 19-amino acid signal peptide cut off from the N-terminus by matrix metalloproteinase 16 (MMP-16), and excision of the central 54-amino acid region (the cleavage sites at 68 and 121), corresponding to kisspeptin-54 (Kp-54, previously known as metastin) by aminopeptidase Q (AMPQ) and matrix metalloproteinases 5 and 24 (MMP-5 and MMP-24). Kp-54 can be cleaved into shorter forms of lower molecular weight (MW), giving rise to Kp-14, Kp-13, and Kp-10. All kisspeptins share a common C-terminus with Kp-54 and contain the RF-amide motif that is able to bind and activate kisspeptin receptor. COOH – carboxy terminal; kDa – protein product size in kilodaltons; NH2 – amino terminal. .
Figure 1.
Formation and structure of kisspeptin molecules (inspired by Tena-Sempere [28], modified from López-Ojeda and Hurley [26]). In humans, KISS1 gene located on the long arm of chromosome 1 (1q32), produces a 145-amino acid precursor protein, prepro-kisspeptin, via two coding exons (III. and IV.) of its four exons. Different kisspeptins are generated by proteolytic cleavage of prepro-kisspeptin, with the 19-amino acid signal peptide cut off from the N-terminus by matrix metalloproteinase 16 (MMP-16), and excision of the central 54-amino acid region (the cleavage sites at 68 and 121), corresponding to kisspeptin-54 (Kp-54, previously known as metastin) by aminopeptidase Q (AMPQ) and matrix metalloproteinases 5 and 24 (MMP-5 and MMP-24). Kp-54 can be cleaved into shorter forms of lower molecular weight (MW), giving rise to Kp-14, Kp-13, and Kp-10. All kisspeptins share a common C-terminus with Kp-54 and contain the RF-amide motif that is able to bind and activate kisspeptin receptor. COOH – carboxy terminal; kDa – protein product size in kilodaltons; NH2 – amino terminal. .
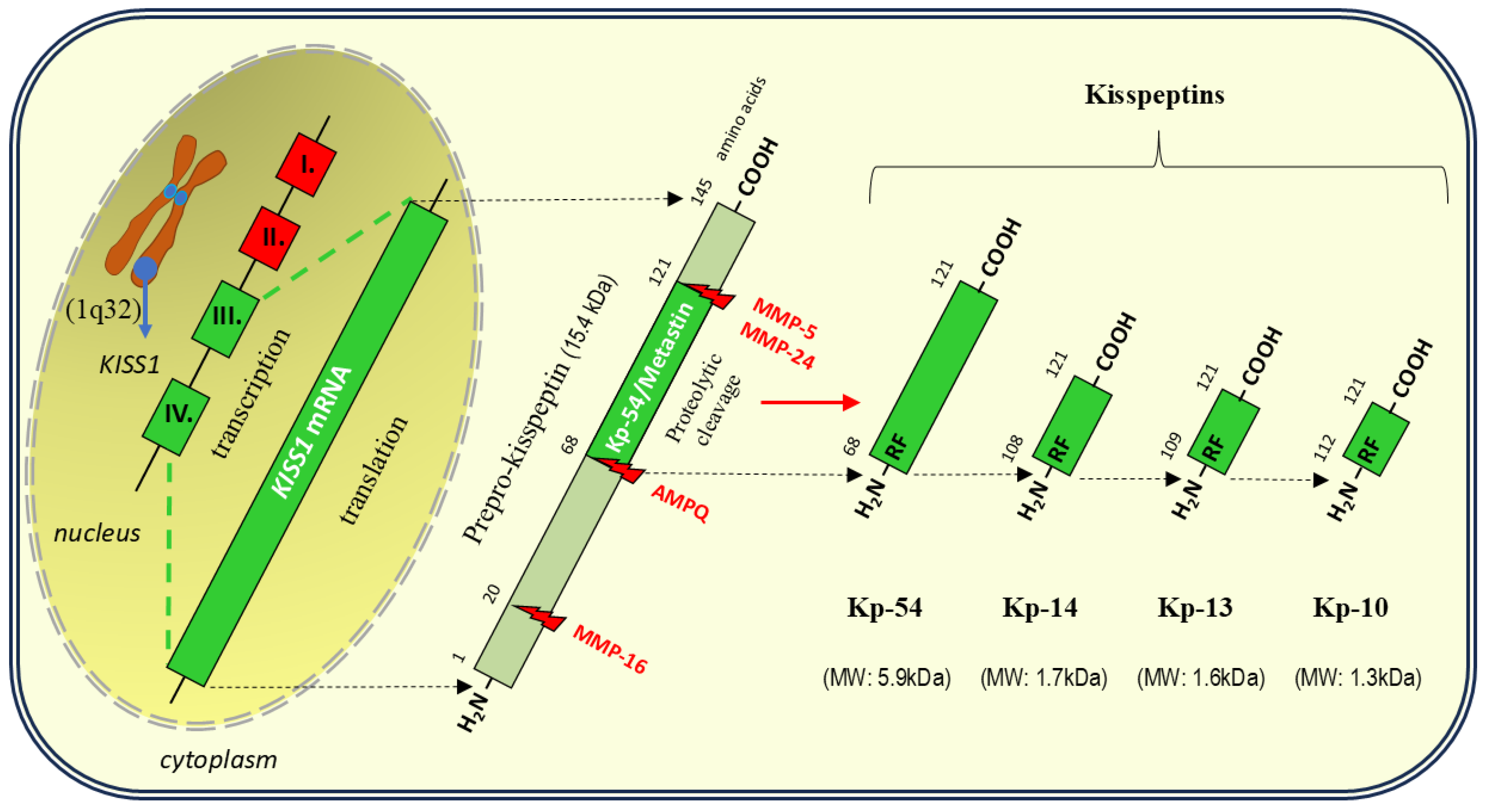
Figure 2.
Hypothalamic control of pulsatile gonadotropin-releasing hormone (GnRH) secretion by kisspeptin/neurokinin B/dynorphin (KNDy) neurons and kisspeptin (KP) neurons [18,51,52,53]. Neurokinin B (NKB) stimulating tachykinin receptor 3 (TACR3) on interconnected KNDy neurons causes the release of KP, which, using an autocrine mechanism, acts on KP receptors (KISS1R) in GnRH neurons and triggers pulsatile GnRH secretion. At the same time, autocrine stimulation of KISS1R by KP on KNDy neurons further enhances the stimulatory effect of NKB. GnRH pulsatility is inhibited by the action of dynorphin (Dyn) on kappa opioid receptors (KOR) in KNDy neurons, leading to inhibition of both NKB and KP secretion. In addition to KNDy neurons located in the infundibular nucleus, GnRH release via KISS1R is also stimulated by KP neurons of the preoptic area (POA). As a result of the action of GnRH, gonadotropins released from the gonadotrophs of the anterior pituitary gland in the form of follicle-stimulating hormone (FSH) and luteinizing hormone (LH) stimulate the production of sex steroids in the gonads (e.g., ovaries). Then, sex steroids inhibit the secretion of KP from KNDy neurons via a negative feedback mechanism, whereas they stimulate KP neurons within the POA (positive feedback). Metabolic stimuli associated with the action of insulin, ghrelin, and also leptin, produced in white adipose tissue (WAT), exert a stimulating effect on KNDy neurons. the "(+)" sign and arrow indicate stimulation; the "(-)" sign and a line ending with a crossbar indicate inhibition.
Figure 2.
Hypothalamic control of pulsatile gonadotropin-releasing hormone (GnRH) secretion by kisspeptin/neurokinin B/dynorphin (KNDy) neurons and kisspeptin (KP) neurons [18,51,52,53]. Neurokinin B (NKB) stimulating tachykinin receptor 3 (TACR3) on interconnected KNDy neurons causes the release of KP, which, using an autocrine mechanism, acts on KP receptors (KISS1R) in GnRH neurons and triggers pulsatile GnRH secretion. At the same time, autocrine stimulation of KISS1R by KP on KNDy neurons further enhances the stimulatory effect of NKB. GnRH pulsatility is inhibited by the action of dynorphin (Dyn) on kappa opioid receptors (KOR) in KNDy neurons, leading to inhibition of both NKB and KP secretion. In addition to KNDy neurons located in the infundibular nucleus, GnRH release via KISS1R is also stimulated by KP neurons of the preoptic area (POA). As a result of the action of GnRH, gonadotropins released from the gonadotrophs of the anterior pituitary gland in the form of follicle-stimulating hormone (FSH) and luteinizing hormone (LH) stimulate the production of sex steroids in the gonads (e.g., ovaries). Then, sex steroids inhibit the secretion of KP from KNDy neurons via a negative feedback mechanism, whereas they stimulate KP neurons within the POA (positive feedback). Metabolic stimuli associated with the action of insulin, ghrelin, and also leptin, produced in white adipose tissue (WAT), exert a stimulating effect on KNDy neurons. the "(+)" sign and arrow indicate stimulation; the "(-)" sign and a line ending with a crossbar indicate inhibition.
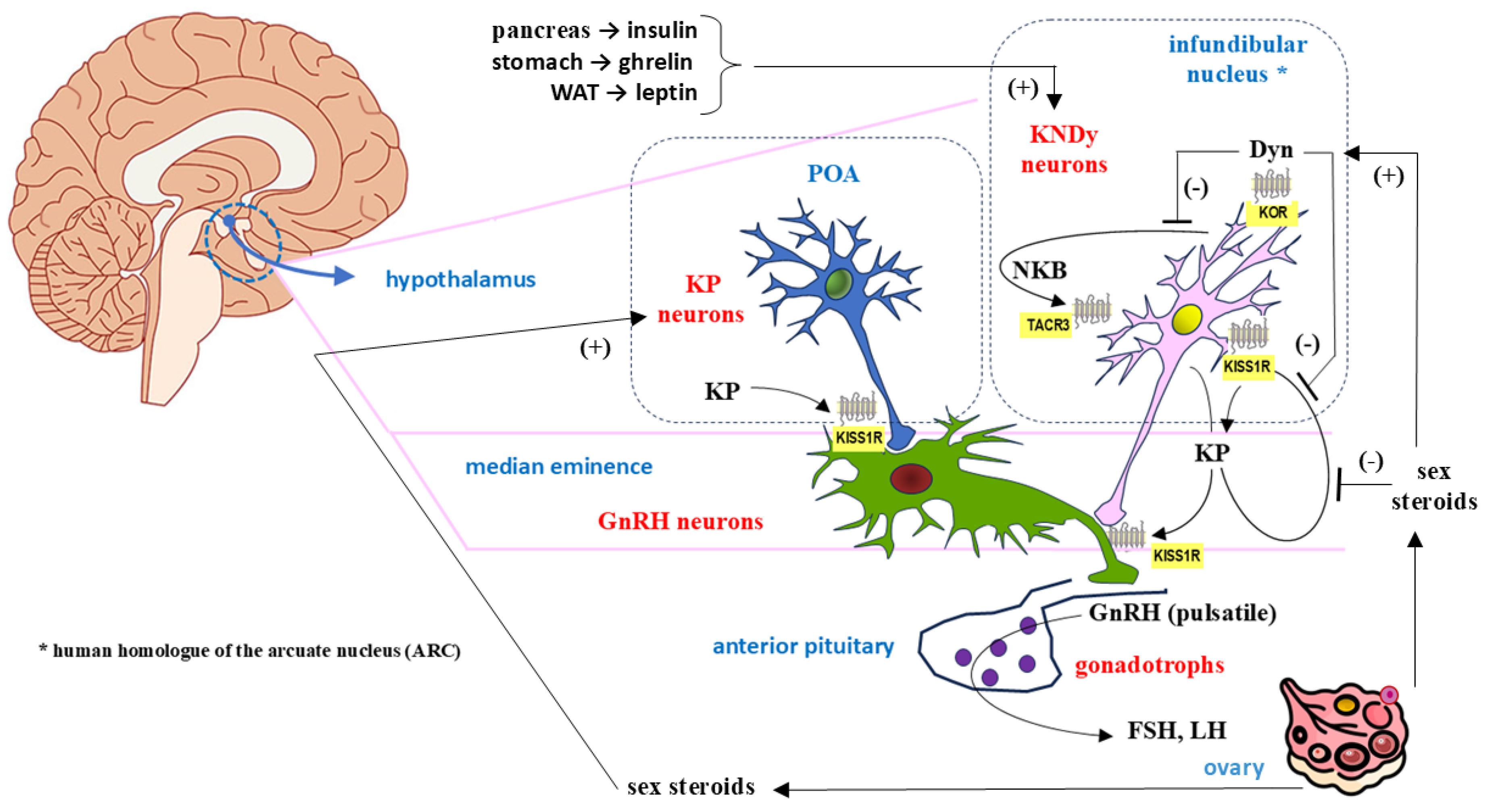
Figure 3.
Human kisspeptin receptor (KISS1R/GPR54): general structural diagram showing mutations in the peptide chain that result in amino acid exchange leading to inactivation, possible inactivation or activation of the receptor (marked in pink, yellow and green, respectively). Modified and updated from Pinilla et al. [27], Roa et al. [103], Abbara et al. [104], Gianetti and Seminara [105] and Franssen and Tena-Sempere [106].
Figure 3.
Human kisspeptin receptor (KISS1R/GPR54): general structural diagram showing mutations in the peptide chain that result in amino acid exchange leading to inactivation, possible inactivation or activation of the receptor (marked in pink, yellow and green, respectively). Modified and updated from Pinilla et al. [27], Roa et al. [103], Abbara et al. [104], Gianetti and Seminara [105] and Franssen and Tena-Sempere [106].
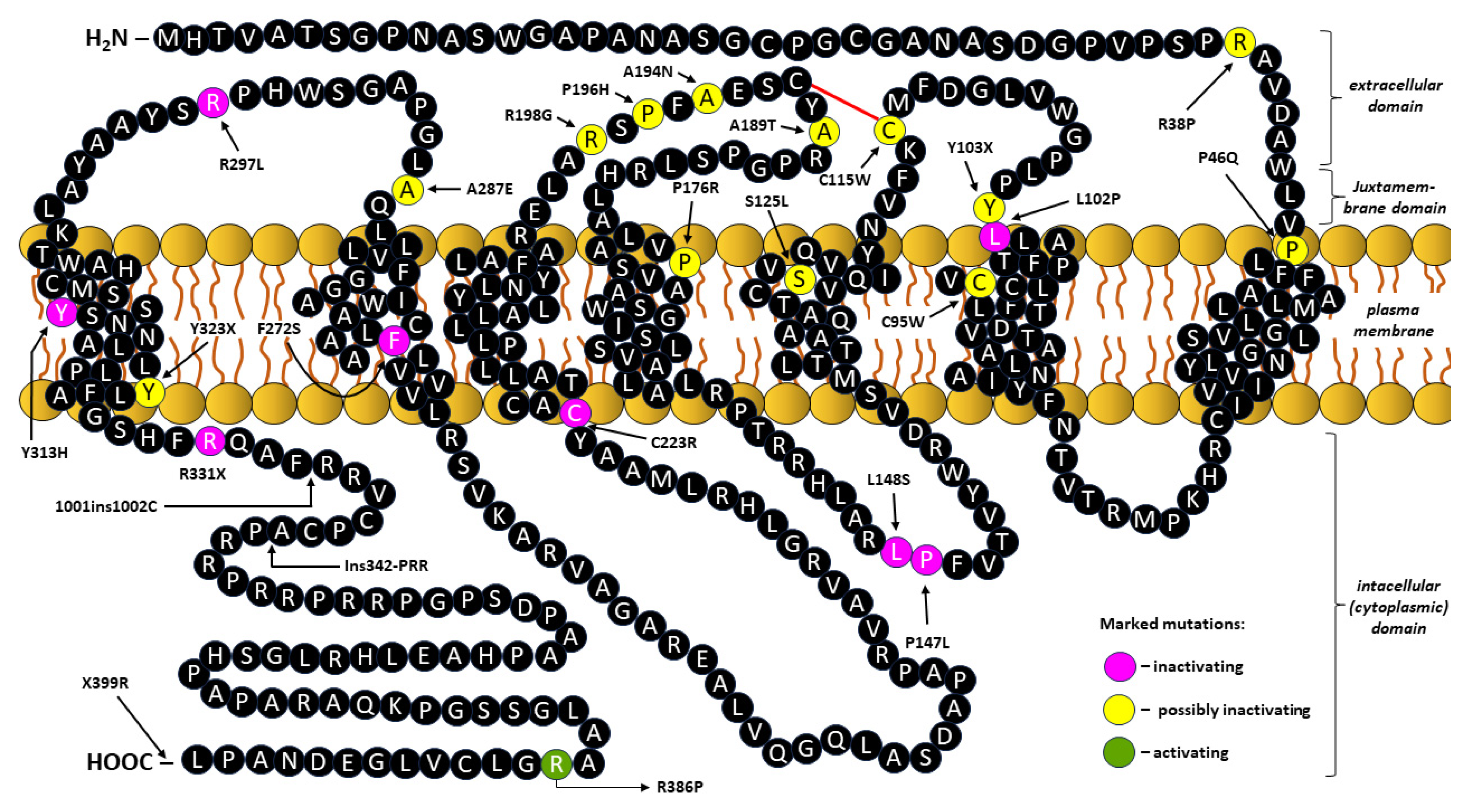
Figure 4.
Kisspeptin signaling via its receptor KISS1R [also known as G-protein-coupled receptor-54 (GPR54)]. After binding of the ligand (kisspeptin) to KISS1R, changes in the structure of the receptor protein occur within the extracellular domain. The activation of the intracellular signal cascade is initiated by the exchange of the guanosine diphosphate (GDP) for a guanosine triphosphate (GTP) in the Gαq/11 subunit of the heterotrimeric G protein, with subsequent dissociation into Gαq/11-GTP and Gβγ. Next, the hydrolytic action of Gαq/11-GTP-activated phospholipase C (PLC)-β leads to the disintegration of the inner membrane component, phosphatidylinositol-4,5-bisphosphate (PIP2), into inositol 1,4,5-trisphosphate (IP3) and diacylglycerol (DAG). IP3 acts as a second messenger in the release of calcium ions (Ca2+) stored in the endoplasmic reticulum (ER), which plays a key role in KISS1R-dependent gonadotropin-releasing hormone (GnRH) release, as well as in antiproliferative and antimetastatic effects. As an allosteric activator of protein kinase C (PKC), DAG leads to the phosphorylation signaling cascade via activation of the serine/threonine mitogen-activated protein kinases (MAPK), including extracellular signal-regulated kinase 1/2 (ERK1/2) and p38 mitogen-activated protein kinase (p38MAPK). Independently of Gαq/11, KISS1R can transduce signals through adaptor proteins, β-arrestins 1 and 2, which leads to inhibition or recruitment of the ERK1/2 pathway, respectively. Signaling through PI3K/Akt/NF-κB (nuclear factor kappa-light-chain-enhancer of activated B cells) or the ERK1/2/NF-κB pathways is accompanied by decreased cellular synthesis of matrix metalloproteinase-9 (MMP-9), which is involved in tissue remodeling via cytokine/chemokine mediators. * Depending on the cell type and its functional state, KISS1R activation can both stimulate and inhibit PI3K (phosphatidylinositol 3-kinase)/Akt [serine/threonine kinase Akt, also known as protein kinase B (PKB)] phosphorylation through a mechanism that has not been sufficiently explained. ATP – adenosine triphosphate; P – phosphorylation; Raf – Raf kinases; Ras – Ras kinases.
Figure 4.
Kisspeptin signaling via its receptor KISS1R [also known as G-protein-coupled receptor-54 (GPR54)]. After binding of the ligand (kisspeptin) to KISS1R, changes in the structure of the receptor protein occur within the extracellular domain. The activation of the intracellular signal cascade is initiated by the exchange of the guanosine diphosphate (GDP) for a guanosine triphosphate (GTP) in the Gαq/11 subunit of the heterotrimeric G protein, with subsequent dissociation into Gαq/11-GTP and Gβγ. Next, the hydrolytic action of Gαq/11-GTP-activated phospholipase C (PLC)-β leads to the disintegration of the inner membrane component, phosphatidylinositol-4,5-bisphosphate (PIP2), into inositol 1,4,5-trisphosphate (IP3) and diacylglycerol (DAG). IP3 acts as a second messenger in the release of calcium ions (Ca2+) stored in the endoplasmic reticulum (ER), which plays a key role in KISS1R-dependent gonadotropin-releasing hormone (GnRH) release, as well as in antiproliferative and antimetastatic effects. As an allosteric activator of protein kinase C (PKC), DAG leads to the phosphorylation signaling cascade via activation of the serine/threonine mitogen-activated protein kinases (MAPK), including extracellular signal-regulated kinase 1/2 (ERK1/2) and p38 mitogen-activated protein kinase (p38MAPK). Independently of Gαq/11, KISS1R can transduce signals through adaptor proteins, β-arrestins 1 and 2, which leads to inhibition or recruitment of the ERK1/2 pathway, respectively. Signaling through PI3K/Akt/NF-κB (nuclear factor kappa-light-chain-enhancer of activated B cells) or the ERK1/2/NF-κB pathways is accompanied by decreased cellular synthesis of matrix metalloproteinase-9 (MMP-9), which is involved in tissue remodeling via cytokine/chemokine mediators. * Depending on the cell type and its functional state, KISS1R activation can both stimulate and inhibit PI3K (phosphatidylinositol 3-kinase)/Akt [serine/threonine kinase Akt, also known as protein kinase B (PKB)] phosphorylation through a mechanism that has not been sufficiently explained. ATP – adenosine triphosphate; P – phosphorylation; Raf – Raf kinases; Ras – Ras kinases.
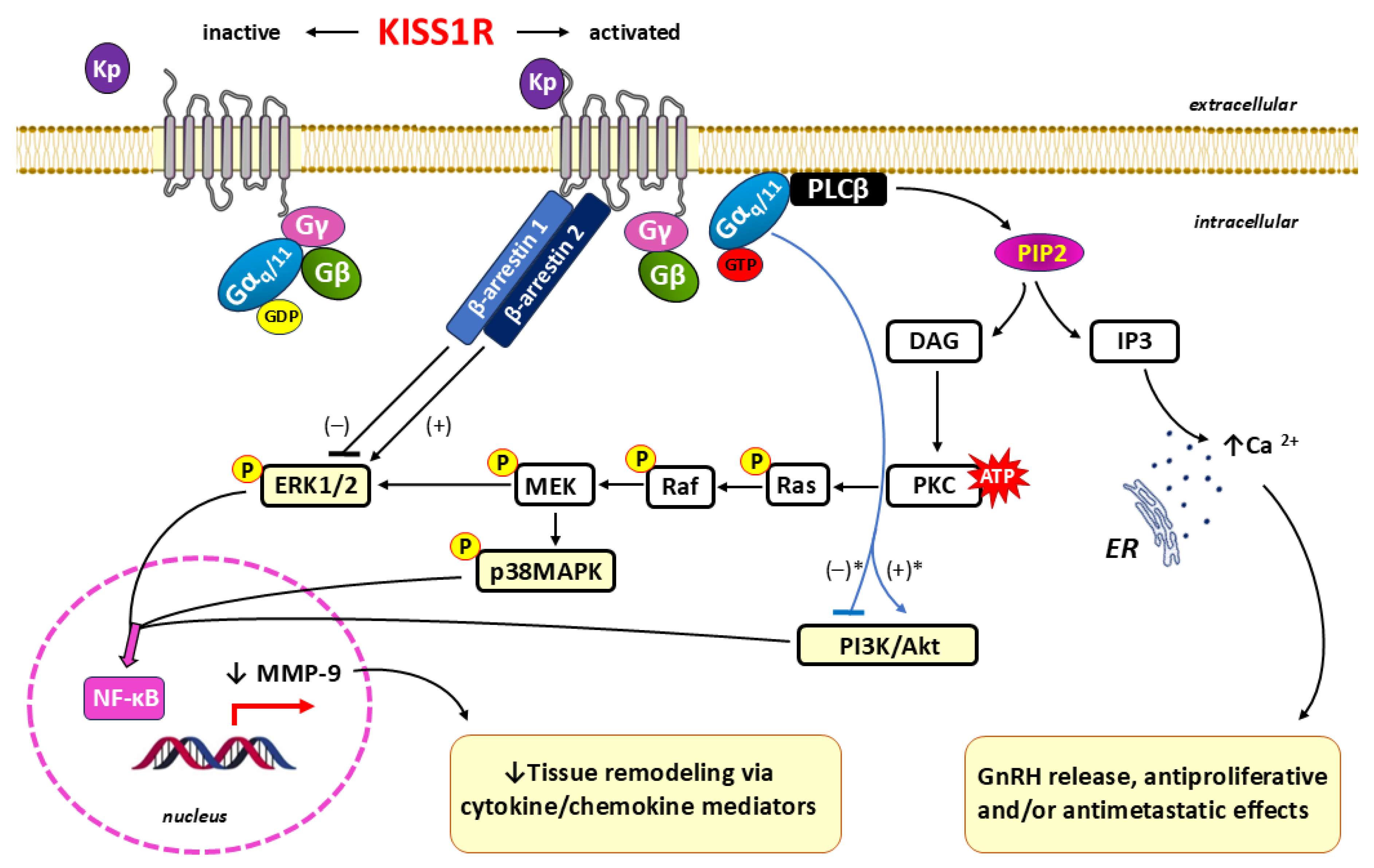

Table 1.
Kisspeptin concentration and expression of kisspeptin and kisspeptin receptor genes (KISS1 and KISS1R, respectively) in early human pregnancy (gestational age ≤ 12 weeks): normal pregnancy versus selected pregnancy complications.
Table 1.
Kisspeptin concentration and expression of kisspeptin and kisspeptin receptor genes (KISS1 and KISS1R, respectively) in early human pregnancy (gestational age ≤ 12 weeks): normal pregnancy versus selected pregnancy complications.
| Parameter | Normal pregnancy | Abnormal pregnancy # | |||
|---|---|---|---|---|---|
| Ectopic pregnancy | GTN | Placenta previa | Miscarriage | ||
| Kisspeptin concentration in plasma or serum |
↑* [33,197,198] |
↓ [67,69,195,196,200] |
↑ [68] |
↓ [143] |
↓ [145,146,148,198,199,200,201,202,203] or INC [32] |
| KISS1 expression in trophoblast/ placental tissue | N/A |
↓ [196] |
↑ [182] |
↓ [143] |
↓ [32,203,204] |
| KISS1R expression in trophoblast/ placental tissue | N/A | N/D |
↑ [182] |
↑ [143] |
↓ [32,204] or WNR [203] |
* compared to non-pregnant women; #compared to normal pregnancy; GTN – gestational trophoblastic neoplasia; INC – inconclusive; N/A – not applicable; N/D – no data; WNR – within normal range.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



