Submitted:
14 May 2025
Posted:
15 May 2025
You are already at the latest version
Abstract
Background: Previous syntheses on the neural effects of alcohol have been restricted to tasks assessing craving, cognitive control, and reward processing. Despite extensive research, a comprehensive synthesis of functional magnetic resonance imaging (fMRI) findings in alcohol use disorder (AUD) remains lacking. This study aimed to identify consistent brain activation alterations across all cognitive and emotional tasks administered to individuals with AUD, while distinguishing between short-term and long-term abstinence, and using Activation Likelihood Estimation meta-analysis. Sub-analyses on task types were performed. Methods: A systematic review identified 67 fMRI studies in participants with an AUD. Results: The meta-analysis revealed significant alterations in brain activity, including both hypo- and hyperactivation in the left putamen across all AUD participants. These alterations were observed more frequently during decision-making and reward tasks. Short-term abstinent individuals exhibited hypoactivation in the right middle frontal gyrus (MFG), corresponding to the dorsolateral prefrontal cortex. In contrast, long-term abstinent individuals displayed hypoactivation in the right superior frontal gyrus (SFG) and dorsal anterior cingulate cortex (dACC). This meta-analysis highlights critical neural alterations in AUD, particularly in regions associated with reward processing (putamen), executive functions (MFG and SFG), and attentional salience (dACC). Putamen changes were predominantly observed during short-term abstinence, and in decision-making as well as reward processing tasks. The dACC and SFG hypoactivation were specific to long-term abstinence, while the MFG hypoactivation was specific to short-term abstinence. Conclusions: These findings support prior research indicating a motivational imbalance and persistent executive dysfunctions in AUD. Standardizing consumption metrics and expanding task diversity in future research is essential to further refine our understanding of the neural effects of AUD.
Keywords:
alcohol use disorder
; meta-analysis
; fMRI
; cognition
1. Introduction
Alcohol is one of the most consumed psychoactive substances in the world [1]. Nearly 400 million people live with an alcohol related disorder around the world, of which 209 million suffer from dependence, resulting in approximately 2.6 million annual deaths [2]. In United States of America, the costs associated with alcohol, including accidents, lost productivity and healthcare utilization, have reached $249 billion over the last decade [3,4,5]. Heavy and chronic alcohol use raise the probabilities of experiencing anxiety, depression, bulimia nervosa, and adopting violent or suicidal behaviors [6,7]. Finally, chronic alcohol use increases the risk of neurodegenerative diseases, such as Alzheimer’s disease [8]. Due to the addiction liability of alcohol and its psychiatric and cognitive effects, several neuroimaging studies have been pursued to understand the effects of chronic alcohol use on the brain.
Structural neuroimaging research has demonstrated that chronic alcohol use is associated with brain tissue shrinkage, evidenced by reduced cortical thickness particularly in the frontal and temporal lobes as well as in the cingulate gyrus [1,9,10]. Voxel-based morphometry studies have revealed that key regions of the mesolimbic system, namely the putamen and the nucleus accumbens, are particularly impaired in chronic alcohol users [9,11,12]. Diffusion neuroimaging has shown that chronic alcohol use is associated with alterations of the integrity of white matter fibers, notably in the corpus callosum, cingulum, and fornix [13]. On the other hand, the involvement of corticospinal and thalamic tracts remains debated [14]. Chronic alcohol use impairs grey matter in (meso)-limbic and fronto-temporal circuits involved in reward and memory, and white matter fibers known to be involved in cognition, emotion and self-processing [10]. While some of these alterations may be mitigated by abstinence, the variability of results across studies complicates conclusions about the persistency of the observed changes [14,15].
As for functional neuroimaging, most task-based research using functional magnetic resonance imaging (fMRI) has focused on experimental paradigms assessing craving (with alcohol-related cues) [16,17,18,19], executive functions [17,18,20] and reward processing [17,21]. While meta-analyses on cravings have revealed that alcohol is associated with dysfunctions in the brain reward system, the default-mode network and parieto-occipital regions, meta-analyses on executive functions (e.g. cognitive control) revealed alterations in frontal and cingulate regions, and meta-analyses on reward processing revealed expected alterations of brain reward regions [17,18,19,20,21]. Echoing the results of the task-based fMRI literature, research syntheses on the functional connectivity literature has shown that alcohol is associated with impaired functional connectivity between the ventral striatum and the ventromedial prefrontal cortex (vmPFC), which are core regions of the brain reward system [22]. Despite the strengths of these meta-analytic findings, results have been heterogeneous and sometimes inconsistent across meta-analyses. Heterogeneity of findings may be explained by the heterogeneity of the included populations. While some syntheses have included only studies on alcohol use disorder (AUD) [20], others have also included studies on binge or heavy drinking [16,18,19,21]. In addition, in most previous syntheses, studies with no group of healthy controls have been included [16,17,18], making it difficult to determine if the observed patterns of activations are abnormal or not. Another issue of previous syntheses is that most of them have included studies using predefined regions of interest (ROIs) [16,17,19,20,21] although this is not recommended in the fMRI meta-analytic guidelines [23], as this approach may skew results towards a priori hypotheses. Furthermore, some meta-analyses included studies on substances other than alcohol [16,17], making it difficult to determine if results are related to alcohol per se or not. A final limitation of previous syntheses is that subs-analyses on abstinence have been performed in only a minority of cases [19], leaving the question of the potential reversibility of alcohol effects unanswered.
In addition to task assessing cravings, executive functions and reward processing, several fMRI studies have examined alternative experimental paradigms, including tasks assessing negative emotion processing, decision-making, language, memory, auditory functions and social cognitive processes [24,25,26,27,28]. This interest stems from the high co-occurrence between chronic alcohol use and anxio-depressive symptoms [29], as well as the fact that AUD is associated with cognitive deficits that are not restricted to executive functions. Indeed, two large meta-analyses have shown that AUD is associated with deficits in attention, decision-making, psychomotor speed, social cognition and verbal fluency (language) [30,31]. Unfortunately, previous meta-analyses of task-based fMRI on chronic alcohol use have not included studies examining emotion processing, decision-making, language, memory, auditory functions and social cognitive processes in the scanner. This has prevented a comprehensive synthesis of the neural alterations observed in chronic alcohol users in the whole task-based fMRI literature. In theory, the addition of the full range of tasks that have been used in the field may help unravel widespread neural alterations extending the alterations that have been observed in brain reward and executive regions in previous syntheses [17,18,19,20,21]. This assumption is coherent with the fact that alcohol produces its psychoactive effects primarily via glutamatergic and GABAergic receptors that are found in high concentrations across the brain [1].
In view of the state of knowledge, we propose to perform a meta-analysis of task-based fMRI studies on alcohol, regardless of the task used in the scanner. This across-task approach has already been successfully applied in the context of several mental disorders, including borderline personality disorder, major depressive disorder, bipolar disorder, posttraumatic stress disorder, anxiety disorders, and neurodevelopmental disorders [32,33,34,35,36,37]. Secondary analyses on task types will be performed. To reduce the heterogeneity of population, we will include studies involving participants with AUD, compared to a control group of healthy volunteers, or studies adopting dimensional analyses on AUD severity levels (from recreational use to disorder). Also, we will only include studies having used a whole-brain analysis approach, excluding predefined ROIs, to provide the most comprehensive overview of the neural effects of alcohol. By doing so, we expect to observe neural alterations that are more widespread than the alterations reported in previous fMRI syntheses on alcohol.
2. Methods
2.1. Literature Search and Selection Criteria
An exhaustive search of PubMed, Web of Science, Google Scholar and Embase was performed independently by two authors (MR, MB) with the following keywords: ‘Alcohol Use Disorder’ AND ‘AUD’AND ‘fMRI’ AND ‘Task’. In addition, cross-referencing of the most recent and large-scale meta-analyses in the field was conducted [16,17,18,19] to ensure the inclusion of all relevant studies.
The selected articles were then reviewed by all authors (SP, MB, MR) to affirm the validity of the studies in the present research. To ensure the quality of the selected articles, rigorous exclusion criteria were chosen. These criteria included: binge drinking, heavy drinking, or any other type of alcohol consumption that was not supported by a DSM or AUDIT diagnosis, acute administration, any other mental health or substance use disorder that was primary. Articles in a language other than English (or French) or which were incomplete were also excluded from our analysis. All these criteria were motives for exclusion. For studies originating from the same cohort (e.g., Boystown program), we only included studies reporting results from distinct experimental tasks.
2.2. Data Extraction
Data extraction was performed by one author (MR), and was verified by the other authors (MB, SP). Data from the selected articles were extracted manually and included: sociodemographic variables (the number of participants, the substance consumed and the age of each group, as well as the duration of abstinence and the diagnostic criterion), as well as the MRI analysis variables (power in Tesla, smoothing [mm], voxel size [mm], repetition time [TR, ms], task, contrast used, direction of activation [hyper/hypo], the coordinates [x,y,z] as well as the stereotaxic atlas used [MNI, TAL mainly]).
2.3. Activation Likelihood Estimation
The activation likelihood estimation (ALE) method was employed for the coordinate-based meta-analysis using GingerALE version 3.0.2 (https://www.brainmap.org/ale/). This technique assesses spatial convergence across studies and requires that peak foci be provided in stereotactic coordinate [38]. Consequently, coordinates in the "x, y, z" format were extracted from each article. These coordinates were then converted from Talairach to Montreal Neurological Institute (MNI) coordinates using the icbm2tal transform [39].
A main meta-analysis was conducted combining long-term abstinent (more than 4 weeks abstinent) and short-term abstinent participants with AUD (less than four weeks abstinent). As in previous studies on the neurocognitive effects of alcohol, 4 weeks (28 days) seem to be the best variable to discriminate notable differences in abstinence [40]. Sub-analyses were also conducted for the short-term and long-term abstinence studies. The ALE approach does not account for the direction of effects. Therefore, for each of group of experiments (short- and long-term abstinence, total), three main separate analyses were conducted: one for hyperactivated foci, one for hypoactivated foci, and one combining both hypoactivated and hyperactivated foci, as in several past meta-analyses using ALE [16,17,21]. For each analysis, activation maps were modeled for each foci group using a mask, the foci, and a Gaussian blur. The full-width-half-maximum (FWHM) of the Gaussian filter was determined based on the sample size of each study, incorporating spatial uncertainty associated with each foci [41]. An unthresholded ALE image was generated from the union of the modeled activation maps. ALE scores were then compared against a null distribution using a histogram, and a table of p-values was produced. This unthresholded ALE image and p-value table were used to create a 3D p-value image, which was then thresholded for statistical significance using the family-wise error (FWE) correction [41]. The uncorrected p-value threshold was set at p<0.001, with cluster-level FWE correction applied at p<0.05 using 1000 permutations. Significant results were functionally characterized using NeuroSynth meta-analytic term-based decoding, and the 5 most strongly correlated terms were identified.
Since meta-regression analyses are not supported by GingerALE, alternative steps were taken to examine the moderating effects of age, sex ratio, MRI magnetic field strength (1.5T vs 3T), MRI parameters (voxel size, time repetition, smoothing level, number of tesla), and task types [craving, executive functions (e.g. attention, cognitive control & working memory), emotion processing, decision-making, reward processing and other functions assessed in fewer than 5 studies (e.g. language, audition, etc.)] on the probabilities of activation of each significant cluster. To do this, probabilities of activation for each study were extracted and analyzed using SPSS. Binary logistic regressions were conducted for each confounding variable to assess its impact on the probabilities of activation (0 or 1) of significant clusters. These analyses were performed individually for each predictor and each significant cluster.
3. Results
3.1. Included Studies
Out of 3,090 studies identified (after duplicates removed), 2,883 were excluded based on their abstracts. Of the remaining 257 articles, 190 were further excluded for the following reasons: (1) analyses focused on predetermined regions of interest, (2) incomplete data, (3) lack of between-group comparisons or results, (4) duplicate cohorts, (5) reliance on connectivity results, (6) absence of a control group, (7) used acute administration, (8) were conference summaries, (9) were involving binge/heavy drinkers, or (10) alcohol not being the primary diagnosis (Supplementary Figure 1, Supplementary Table 1). A total of 67 studies with a sample size of 2,341 subjects met the final inclusion criteria (Table 1). Studies used tasks assessing craving, reward processing, executive functions, emotion processing, decision-making, language, memory, auditory functions and social cognitive processes (Supplementary Table 2).
3.1.1. Hypoactivation and Hyperactivation Pooled Together in the Whole Sample (Short-Term & Long-Term Abstinence)
This analysis included 730 foci from 67 experiments on 2,431 subjects with an alcohol use disorder. Aberrant activity of the left putamen (68.1%), caudate body (22.5%), and caudate head (9.3%) was found to be significant at a corrected threshold of p<0.05 (Table 2). Associations with terms like delay, gains, monetary, incentive and losses were found in Neurosynth.
3.1.2. Hypoactivated Foci in the Whole Sample
Analysis of hypoactivated foci included 345 foci from 44 experiments on 1,245 subjects with an alcohol use disorder. No clusters of significant decrease activation were found at a corrected threshold.
3.1.3. Hyperactivated Foci in the Whole Sample
For the hyperactivated foci analysis, 385 foci were included comprising 47 experiments on 1,759 subjects with an alcohol use disorder. No clusters of significant increase activation were found at a corrected threshold.
3.1.4. Hypoactivation and Hyperactivation Pooled Together in the Short-Term Abstinent Sample
This analysis included 349 foci from 37 experiments on 1,097 subjects with an alcohol use disorder. Aberrant activity of the left putamen was found to be significant at a corrected threshold of p<0.05 (Table 2; Figure 1). Since this cluster was the same as the one found in the hyper-hypoactivated foci analysis in the combined sample, the Neurosynth terms were the same, as mentioned previously.
3.1.5. Hypoactivated Foci in the Short-Term Abstinent Sample
Analysis of hypoactivated foci included 125 foci from 25 experiments on 631 subjects with an AUD. Decrease activation of a cluster located in the right middle frontal gyrus (MFG) (56.7%), the superior frontal gyrus (SFG) (40%) and the sub-gyral (3.3%) was found in participants with alcohol use disorder compared to controls (Table 2; Figure 1). This cluster was associated with Neurosynth terms like response times, task difficulty, instructions, difficulty, and signal task.
3.1.6. Hyperactivated Foci in the Short-Term Abstinent Sample
For the hyperactivated focis analysis, 224 foci were included comprising 29 experiments on 893 subjects with an AUD. No clusters of significant increase activation were found at a corrected threshold.
3.1.7. Hypoactivation and Hyperactivation Pooled Together in the Long-Term Abstinent Sample
Included are 348 foci from 26 experiments on 1,001 subjects with an AUD. This analysis yielded no clusters.
3.1.8. Hypoactivated Foci in the Long-Term Abstinent Sample
Analysis of hypoactivated foci included 212 foci from 17 experiments on 532 subjects with an AUD. This analysis yielded two significant clusters: (1) a cluster located in the right SFG (63.6%) and the MFG (36.4%); (2) a bilateral cluster in the cingulate gyrus (87.1%) and the medial frontal gyrus (12.9%) (Table 2; Figure 2). The first cluster was linked to Neurosynth terms like noxious, executive, working, working memory, and abilities. The second cluster was associated to terms like monitoring, task, error, learning task, and painful.
3.1.9. Hyperactivated Foci in the Long-Term Abstinent Sample
For the hyperactivated foci analysis, 136 foci were included comprising 16 experiments on 615 subjects with an AUD. No clusters of significant increase activation were found at a corrected threshold.
3.2. Sub-Analyses on Socio-Demographic Variables, MRI Parameters and Task Types
No associations were found between socio-demographic variables and MRI parameters and probabilities of activation of each significant cluster (Table 3). Relationships with task types were identified (Table 3). In the short-term sample, significant positive associations were observed between the putamen and decision-making tasks (OR=9.33, 95% CI [1.27–68.60], p=0.028), as well as reward tasks (OR=30.00, 95% CI [2.33–386.33], p<0.009). No significant relationships were found between the MFG and any task type. In the long-term abstinent sample, a significant positive association was identified between the SFG and reward tasks (OR=9.00, 95% CI [1.03–78.57], p<0.047). However, no significant associations were observed in the case of the cingulate gyrus.
4. Discussion
The objective of this meta-analysis was to provide a comprehensive overview of the neural effects of AUD through whole-brain fMRI analyses, regardless of the task used in the scanner, while distinguishing effects based on the duration of abstinence. The activity of the left putamen emerged as being disrupted in the analysis combining both hypo- and hyper-activations. A positive association was found between the aberrant activity of the putamen and decision-making and reward tasks. In short-term abstinent AUD participants, hypoactivation was observed in the right MFG, corresponding to the dorsolateral prefrontal cortex (dlPFC). For long-term abstinent AUD participants, hypoactivation was observed in the right SFG encompassing the MFG, in the lateral part of the prefrontal cortex (PFC), as well as in the dorsal anterior cingulate cortex (dACC). In the whole sample of both short- and long-term AUD participants, the putamen was also found to be altered. Lastly, sub-analyses revealed no association between neural alterations and sociodemographic and MRI parameters.
Our results demonstrated that the activation of left putamen is bidirectionally altered in AUD, and that these alterations are mostly driven by the short abstinence studies. As such, this result is consistent with previous structural neuroimaging studies which have reliably shown that grey matter volumes/concentrations are decreased in AUD patients [9,12,13,42]. Our findings are also consistent with previous fMRI meta-analyses, which have shown a similar pattern of complex alterations in the putamen in AUD [17,21]. In healthy volunteers, fMRI studies have demonstrated that this dopamine-rich structure is involved in reward processing and associative learning [43,44]. In AUD, previous fMRI reward studies determined that the putamen shows a decreased activation during the Monetary Incentive Delay Task [21] and a hyperactivation in response to alcohol cues [17,18]. In the sub-analyses on task types, we observed associations between the putamen and reward processing and reward-based decision-making, which are coherent with the previous literature in the field. Taken together, the results from the current meta-analysis and previous meta-analyses strongly suggest that AUD is associated with a motivational imbalance whereby the brain reward system becomes hyper-activated in response to alcohol cues, while displaying a blunted response to rewarding stimuli or cues that are not alcohol-related (example: money) [45,46,47]. The fact that significant putamen alterations were only observed during short-term abstinence is coherent with the clinical literature having shown that dysphoric symptoms and alcohol cravings tend to stabilize during the first 3-4 weeks of alcohol withdrawal [48,49].
Two clusters in right MFG and SFG were found to be hypo-activated in AUD, corresponding to the right dlPFC and the right lateral PFC. These results are coherent with past structural neuroimaging studies, which have demonstrated a reduction in gray matter in the MFG and SFG in AUD, with the dlPFC appearing to be predominantly affected [11,12]. In the healthy population, large-scale fMRI evidence has shown that the right MFG and SFG play critical roles in core executive functions, including response inhibition and working memory [50,51,52]. Coherently with these findings, we performed functional decoding analyses, which revealed that the right dlPFC cluster was associated with functions like response inhibition, whereas the right lateral PFC cluster was associated with functions like working memory. Some authors have argued that executive dysfunctions are central to the development and maintenance of AUD [53,54], since these deficits weaken the ability to plan and execute effective strategies, resulting in social and occupational problems, as well as impaired impulse regulation and a loss of behavioral control [55]. Supporting the proposed interaction between executive and reward regions in AUD, the sub-analyses on task types showed and association between the SFG and reward tasks. As synthesized by Stavro and colleagues (2013) and Crowe and colleague (2019), a large body of cognitive studies have shown that working memory and response inhibition are significantly impaired in AUD, and that these deficits clearly persist after the first weeks of alcohol withdrawal, with residual effects sometimes lasting several months. The fact that the lateral PFC was found to be impaired in long-term abstinence studies is consistent with the cognitive literature on AUD.
A hypoactivation was observed in long term abstinence studies in AUD in the dACC, which is one of the core hubs of the (attentional) salience network [56]. The alteration of the dACC is supported by the structural neuroimaging literature, which has consistently shown gray matter reductions in this region in substance use disorders, including AUD [11,12,13,57]. Our result is also consistent with the results of previous fMRI meta-analyses, which has consistently shown alterations in ACC activation in AUD in separate analyses on reward processing, craving and executive functions [16,17,18,19,20,21]. In the healthy population, the dACC has been shown to be involved in top-down attention control and the detection of salient changes in the interoceptive and external environment [58,59]. In view of the well-known roles of the dACC, the reduced activity observed in this region in AUD patients may signal attentional biases towards alcohol-related stimuli and/or an inability to re-allocate attentional resources to non-alcohol stimuli [60]. The fact that the dACC hypo-activation was only observed in the long-term abstinence studies suggests that the dACC may play a key role in alcohol relapse, as recently proposed by some investigators [61].
This meta-analysis has significant strengths compared to previous fMRI meta-analyses in the field. We conducted a meta-analysis including only whole-brain fMRI studies involving participants with AUD specifically, as compared to healthy control participants. This approach allowed us to include a more homogeneous study population and to produce a more comprehensive overview of alcohol's effects on the AUD population while reducing biases associated with the selection of regions of interest. Moreover, our meta-analysis is the first to quantitatively review all tasks available, including task domains that were excluded from previous syntheses (audition, language, social cognition, etc.). This approach allowed us to include 27 fMRI studies that would have been excluded had we limited the analyses to tasks assessing executive functions, reward processing and craving (as in previous meta-analyses). Despite the variety of the tasks reviewed, our findings converge on alterations of brain regions involved in reward processing, executive functions and attentional salience, suggesting that the neural alterations observed in AUD may be restricted to these psychological domains.
Despite these strengths, a few limitations need to be acknowledged. Although the selection of tasks was comprehensive, the number of studies per task domain was rather small in several cases, namely in the case of tasks assessing language, audition, motor functions, social cognition and episodic memory. For these domains, the number of studies was lower than the minimal number studies that has been estimated to provide sufficient statistical power for fMRI meta-analyses (e.g. 17 studies) [62,63]. Therefore, statistical power was most probably insufficient for all these domains. As such, this could explain why we failed to observe neural impairments in the brain regions / networks involved in these functions. Moreover, although sub-analyses were performed on several variables that may have influenced results (e.g. age, sex, MRI parameters, etc.), we were not able to examine the influence on results of the quantities of alcohol consumed by AUD patients. Indeed, the included studies often lacked clear data on consumption in terms of standardized units, precise temporality, and some relied on subjective measures influenced by national and political standards. This is unfortunate considering that alcohol is presumed to be more neurotoxic for those who have consumed larger quantities in their lifetime, although results on this topic are more complex than previously thought [64,65,66].
The current study sought to identify the neural alterations associated with AUD by performing a meta-analysis of whole-brain fMRI studies including all tasks available. We found that AUD was associated with neural impairments in brain regions involved in reward processing (putamen), executive functions (MFG) and attention salience (dACC). Length of abstinence influenced results, with striatal changes being driven by short-term abstinence studies, and dACC changes being observed only in long-term abstinence studies. These findings underscore the importance of broadening the array of task domains in future fMRI research on AUD. Diversifying these domains may help move beyond the central triad of networks identified and examine more thoroughly the potential diffuse effects of AUD on brain functioning. Finally, it will be crucial to standardize, in future fMRI research, alcohol consumption units to facilitate data conversion and comparison, and thus, to refine our understanding of the neural effects of AUD.
Author Contributors
SP and MB designed the study, MR and MB did the literature search, MR extracted data, MR and MB performed the analyses, MB, MR and SP wrote the manuscript, SP and MB provided critical comments, all authors approved the final version of the manuscript.
Acknowledgments
SP is holder of the Eli Lilly Canada Chair on schizophrenia research; MB is holder of a doctoral scholarship from the Canadian Institutes of Health Research (FBD-193359).
References
- Meyer, J.; Farrar, A.M.; Bienzonski, D.; Yates, J.R. Psychopharmacology, Drugs, the Brain, and Behavior, 4th ed.; Press, O.U., Ed.; 2022.
- WHO. Over 3 million annual deaths due to alcohol and drug use, majority among men. 25 June 2024.
- Chikritzhs, T.; Livingston, M. Alcohol and the Risk of Injury. Nutrients 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Sacks, J.J.; Gonzales, K.R.; Bouchery, E.E.; Tomedi, L.E.; Brewer, R.D. 2010 National and State Costs of Excessive Alcohol Consumption. Am J Prev Med 2015, 49, e73–e79. [Google Scholar] [CrossRef] [PubMed]
- Thorrisen, M.M.; Bonsaksen, T.; Hashemi, N.; Kjeken, I.; van Mechelen, W.; Aas, R.W. Association between alcohol consumption and impaired work performance (presenteeism): a systematic review. BMJ Open 2019, 9, e029184. [Google Scholar] [CrossRef]
- Hendriks, H.F.J. Alcohol and Human Health: What Is the Evidence? Annu Rev Food Sci Technol 2020, 11, 1–21. [Google Scholar] [CrossRef]
- Palzes, V.A.; Parthasarathy, S.; Chi, F.W.; Kline-Simon, A.H.; Lu, Y.; Weisner, C.; Ross, T.B.; Elson, J.; Sterling, S.A. Associations Between Psychiatric Disorders and Alcohol Consumption Levels in an Adult Primary Care Population. Alcohol Clin Exp Res 2020, 44, 2536–2544. [Google Scholar] [CrossRef]
- Kamal, H.; Tan, G.C.; Ibrahim, S.F.; Shaikh, M.F.; Mohamed, I.N.; Mohamed, R.M.P.; Hamid, A.A.; Ugusman, A.; Kumar, J. Alcohol Use Disorder, Neurodegeneration, Alzheimer's and Parkinson's Disease: Interplay Between Oxidative Stress, Neuroimmune Response and Excitotoxicity. Front Cell Neurosci 2020, 14, 282. [Google Scholar] [CrossRef]
- Hahn, S.; Mackey, S.; Cousijn, J.; Foxe, J.J.; Heinz, A.; Hester, R.; Hutchinson, K.; Kiefer, F.; Korucuoglu, O.; Lett, T.; et al. Predicting alcohol dependence from multi-site brain structural measures. Hum Brain Mapp 2022, 43, 555–565. [Google Scholar] [CrossRef]
- Navarri, X.; Afzali, M.H.; Lavoie, J.; Sinha, R.; Stein, D.J.; Momenan, R.; Veltman, D.J.; Korucuoglu, O.; Sjoerds, Z.; van Holst, R.J.; et al. How do substance use disorders compare to other psychiatric conditions on structural brain abnormalities? A cross-disorder meta-analytic comparison using the ENIGMA consortium findings. Hum Brain Mapp 2022, 43, 399–413. [Google Scholar] [CrossRef]
- Li, L.; Yu, H.; Liu, Y.; Meng, Y.J.; Li, X.J.; Zhang, C.; Liang, S.; Li, M.L.; Guo, W.; QiangWang; et al. Lower regional grey matter in alcohol use disorders: evidence from a voxel-based meta-analysis. BMC Psychiatry 2021, 21, 247. [Google Scholar] [CrossRef]
- Yang, X.; Tian, F.; Zhang, H.; Zeng, J.; Chen, T.; Wang, S.; Jia, Z.; Gong, Q. Cortical and subcortical gray matter shrinkage in alcohol-use disorders: a voxel-based meta-analysis. Neurosci Biobehav Rev 2016, 66, 92–103. [Google Scholar] [CrossRef]
- Pando-Naude, V.; Toxto, S.; Fernandez-Lozano, S.; Parsons, C.E.; Alcauter, S.; Garza-Villarreal, E.A. Gray and white matter morphology in substance use disorders: a neuroimaging systematic review and meta-analysis. Transl Psychiatry 2021, 11, 29. [Google Scholar] [CrossRef] [PubMed]
- Spindler, C.; Mallien, L.; Trautmann, S.; Alexander, N.; Muehlhan, M. A coordinate-based meta-analysis of white matter alterations in patients with alcohol use disorder. Transl Psychiatry 2022, 12, 40. [Google Scholar] [CrossRef] [PubMed]
- Monnig, M.A.; Tonigan, J.S.; Yeo, R.A.; Thoma, R.J.; McCrady, B.S. White matter volume in alcohol use disorders: a meta-analysis. Addict Biol 2013, 18, 581–592. [Google Scholar] [CrossRef]
- Devoto, F.; Zapparoli, L.; Spinelli, G.; Scotti, G.; Paulesu, E. How the harm of drugs and their availability affect brain reactions to drug cues: a meta-analysis of 64 neuroimaging activation studies. Transl Psychiatry 2020, 10, 429. [Google Scholar] [CrossRef]
- Klugah-Brown, B.; Di, X.; Zweerings, J.; Mathiak, K.; Becker, B.; Biswal, B. Common and separable neural alterations in substance use disorders: A coordinate-based meta-analyses of functional neuroimaging studies in humans. Hum Brain Mapp 2020, 41, 4459–4477. [Google Scholar] [CrossRef]
- Oberlin, B.G.; Shen, Y.I.; Kareken, D.A. Alcohol Use Disorder Interventions Targeting Brain Sites for Both Conditioned Reward and Delayed Gratification. Neurotherapeutics 2020, 17, 70–86. [Google Scholar] [CrossRef]
- Zeng, J.; Yu, S.; Cao, H.; Su, Y.; Dong, Z.; Yang, X. Neurobiological correlates of cue-reactivity in alcohol-use disorders: A voxel-wise meta-analysis of fMRI studies. Neurosci Biobehav Rev 2021, 128, 294–310. [Google Scholar] [CrossRef]
- Cao, Y.; Tian, F.; Zeng, J.; Gong, Q.; Yang, X.; Jia, Z. The brain activity pattern in alcohol-use disorders under inhibition response Task. J Psychiatr Res 2023, 163, 127–134. [Google Scholar] [CrossRef]
- Zeng, J.; You, L.; Yang, F.; Luo, Y.; Yu, S.; Yan, J.; Liu, M.; Yang, X. A meta-analysis of the neural substrates of monetary reward anticipation and outcome in alcohol use disorder. Hum Brain Mapp 2023, 44, 2841–2861. [Google Scholar] [CrossRef]
- Dugré, J.R.; Orban, P.; Potvin, S. Disrupted functional connectivity of the brain reward system in substance use problems: A meta-analysis of functional neuroimaging studies. Addict Biol 2023, 28, e13257. [Google Scholar] [CrossRef]
- Müller, V.I.; Cieslik, E.C.; Laird, A.R.; Fox, P.T.; Radua, J.; Mataix-Cols, D.; Tench, C.R.; Yarkoni, T.; Nichols, T.E.; Turkeltaub, P.E.; et al. Ten simple rules for neuroimaging meta-analysis. Neurosci Biobehav Rev 2018, 84, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Bagga, D.; Singh, N.; Modi, S.; Kumar, P.; Bhattacharya, D.; Garg, M.L.; Khushu, S. Assessment of lexical semantic judgment abilities in alcohol-dependent subjects: an fMRI study. J Biosci 2013, 38, 905–915. [Google Scholar] [CrossRef] [PubMed]
- Hermann, D.; Smolka, M.N.; Klein, S.; Heinz, A.; Mann, K.; Braus, D.F. Reduced fMRI activation of an occipital area in recently detoxified alcohol-dependent patients in a visual and acoustic stimulation paradigm. Addict Biol 2007, 12, 117–121. [Google Scholar] [CrossRef]
- Li, C.S.; Luo, X.; Yan, P.; Bergquist, K.; Sinha, R. Altered impulse control in alcohol dependence: neural measures of stop signal performance. Alcohol Clin Exp Res 2009, 33, 740–750. [Google Scholar] [CrossRef]
- Maurage, P.; Joassin, F.; Philippot, P.; Heeren, A.; Vermeulen, N.; Mahau, P.; Delperdange, C.; Corneille, O.; Luminet, O.; de Timary, P. Disrupted regulation of social exclusion in alcohol-dependence: an fMRI study. Neuropsychopharmacology 2012, 37, 2067–2075. [Google Scholar] [CrossRef]
- van Holst, R.J.; Clark, L.; Veltman, D.J.; van den Brink, W.; Goudriaan, A.E. Enhanced striatal responses during expectancy coding in alcohol dependence. Drug Alcohol Depend 2014, 142, 204–208. [Google Scholar] [CrossRef]
- McHugh, R.K.; Weiss, R.D. Alcohol Use Disorder and Depressive Disorders. ALCOHOL RESEARCH Current Reviews, 8. [CrossRef]
- Crowe, S.F.; Cammisuli, D.M.; Stranks, E.K. Widespread cognitive deficits in alcoholism persistent following prolonged abstinence: An updated meta-analysis of studies that used standardized neuropsychological assessment tools. Archives of Clinical Neuropsychology 2019, 35(1), 31–45. [Google Scholar] [CrossRef]
- Stavro, K.; Pelletier, J.; Potvin, S. Widespread and sustained cognitive deficits in alcoholism: a meta-analysis. Addict Biol 2013, 18, 203–213. [Google Scholar] [CrossRef]
- Degasperi, G.; Cristea, I.A.; Di Rosa, E.; Costa, C.; Gentili, C. Parsing variability in borderline personality disorder: a meta-analysis of neuroimaging studies. Transl Psychiatry 2021, 11, 314. [Google Scholar] [CrossRef]
- Janiri, D.; Moser, D.A.; Doucet, G.E.; Luber, M.J.; Rasgon, A.; Lee, W.H.; Murrough, J.W.; Sani, G.; Eickhoff, S.B.; Frangou, S. Shared Neural Phenotypes for Mood and Anxiety Disorders A Meta-Analysis of 226 Task-Related Functional Imaging Studies. Focus (Am Psychiatr Publ) 2021, 19, 256–263. [Google Scholar] [CrossRef]
- Pico-Perez, M.; Vieira, R.; Fernandez-Rodriguez, M.; De Barros, M.A.P.; Radua, J.; Morgado, P. Multimodal meta-analysis of structural gray matter, neurocognitive and social cognitive fMRI findings in schizophrenia patients. Psychol Med 2022, 52, 614–624. [Google Scholar] [CrossRef] [PubMed]
- Solomonov, N.; Victoria, L.W.; Lyons, K.; Phan, D.K.; Alexopoulos, G.S.; Gunning, F.M.; Fluckiger, C. Social reward processing in depressed and healthy individuals across the lifespan: A systematic review and a preliminary coordinate-based meta-analysis of fMRI studies. Behav Brain Res 2023, 454, 114632. [Google Scholar] [CrossRef] [PubMed]
- Tamon, H.; Fujino, J.; Itahashi, T.; Frahm, L.; Parlatini, V.; Aoki, Y.Y.; Castellanos, F.X.; Eickhoff, S.B.; Cortese, S. Shared and Specific Neural Correlates of Attention Deficit Hyperactivity Disorder and Autism Spectrum Disorder: A Meta-Analysis of 243 Task-Based Functional MRI Studies. Am J Psychiatry 2024, 181, 541–552. [Google Scholar] [CrossRef]
- Wang, Z.; He, D.; Yang, L.; Wang, P.; Xiao, J.; Zou, Z.; Min, W.; He, Y.; Yuan, C.; Zhu, H.; et al. Similarities and differences between post-traumatic stress disorder and major depressive disorder: Evidence from task-evoked functional magnetic resonance imaging meta-analysis. J Affect Disord 2024, 361, 712–719. [Google Scholar] [CrossRef]
- Eickhoff, S.B.; Bzdok, D.; Laird, A.R.; Kurth, F.; Fox, P.T. Activation likelihood estimation meta-analysis revisited. Neuroimage 2012, 59, 2349–2361. [Google Scholar] [CrossRef]
- Lancaster, J.L.; Tordesillas-Gutierrez, D.; Martinez, M.; Salinas, F.; Evans, A.; Zilles, K.; Mazziotta, J.C.; Fox, P.T. Bias between MNI and Talairach coordinates analyzed using the ICBM-152 brain template. Hum Brain Mapp 2007, 28, 1194–1205. [Google Scholar] [CrossRef]
- Wallach, J.D.; Gueorguieva, R.; Phan, H.; Witkiewitz, K.; Wu, R.; O'Malley, S.S. Predictors of abstinence, no heavy drinking days, and a 2-level reduction in World Health Organization drinking levels during treatment for alcohol use disorder in the COMBINE study. Alcohol Clin Exp Res 2022, 46, 1331–1339. [Google Scholar] [CrossRef]
- Eickhoff, S.B.; Laird, A.R.; Grefkes, C.; Wang, L.E.; Zilles, K.; Fox, P.T. Coordinate-based activation likelihood estimation meta-analysis of neuroimaging data: a random-effects approach based on empirical estimates of spatial uncertainty. Hum Brain Mapp 2009, 30, 2907–2926. [Google Scholar] [CrossRef]
- Fein, G.; Greenstein, D.; Cardenas, V.A.; Cuzen, N.L.; Fouche, J.P.; Ferrett, H.; Thomas, K.; Stein, D.J. Cortical and subcortical volumes in adolescents with alcohol dependence but without substance or psychiatric comorbidities. Psychiatry Res 2013, 214, 1–8. [Google Scholar] [CrossRef]
- Ghandili, M.; Munakomi, S. Neuroanatomy, Putamen. 2023.
- Hu, W.; Zhao, X.; Liu, Y.; Ren, Y.; Wei, Z.; Tang, Z.; Tian, Y.; Sun, Y.; Yang, J. Reward sensitivity modulates the brain reward pathway in stress resilience via the inherent neuroendocrine system. Neurobiol Stress 2022, 20, 100485. [Google Scholar] [CrossRef]
- Kim, S.I. Neuroscientific model of motivational process. Front Psychol 2013, 4, 98. [Google Scholar] [CrossRef] [PubMed]
- Poulton, A.; Hester, R. Transition to substance use disorders: impulsivity for reward and learning from reward. Soc Cogn Affect Neurosci 2020, 15, 1182–1191. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, A.M. Reward, motivation and brain imaging in human healthy participants - A narrative review. Front Behav Neurosci 2023, 17, 1123733. [Google Scholar] [CrossRef]
- Petit, G.; Deschietere, G.; Loas, G.; Luminet, O.; de Timary, P. Link Between Anhedonia and Depression During Early Alcohol Abstinence: Gender Matters. Alcohol Alcohol 2020, 55, 71–77. [Google Scholar] [CrossRef]
- Seo, D.; Sinha, R. Neuroplasticity and predictors of alcohol recovery. Alcohol Research: Current Reviews.
- Depue, B.E.; Banich, M.T. Increased inhibition and enhancement of memory retrieval are associated with reduced hippocampal volume. Hippocampus 2012, 22, 651–655. [Google Scholar] [CrossRef]
- Miller, J.A.; Tambini, A.; Kiyonaga, A.; D'Esposito, M. Long-term learning transforms prefrontal cortex representations during working memory. Neuron 2022, 110, 3805–3819. [Google Scholar] [CrossRef]
- Oldrati, V.; Patricelli, J.; Colombo, B.; Antonietti, A. The role of dorsolateral prefrontal cortex in inhibition mechanism: A study on cognitive reflection test and similar tasks through neuromodulation. Neuropsychologia 2016, 91, 499–508. [Google Scholar] [CrossRef]
- Lopez-Caneda, E.; Rodriguez Holguin, S.; Cadaveira, F.; Corral, M.; Doallo, S. Impact of alcohol use on inhibitory control (and vice versa) during adolescence and young adulthood: a review. Alcohol Alcohol 2014, 49, 173–181. [Google Scholar] [CrossRef]
- Si, Y.; Wang, L.; Zhao, M. Anti-saccade as a Tool to Evaluate Neurocognitive Impairment in Alcohol Use Disorder. Front Psychiatry 2022, 13, 823848. [Google Scholar] [CrossRef]
- Rure, D.; Shakya, M.; Singhal, A.; Varma, A.; Mishra, N.; Pathak, U. A Study of the association of neurocognition with relapse and quality of life in patients of alcohol dependence. Ind Psychiatry J 2024, 33, 133–140. [Google Scholar] [CrossRef]
- Ham, T.; Leff, A.; de Boissezon, X.; Joffe, A.; Sharp, D.J. Cognitive control and the salience network: an investigation of error processing and effective connectivity. J Neurosci 2013, 33, 7091–7098. [Google Scholar] [CrossRef] [PubMed]
- Xiao, P.; Dai, Z.; Zhong, J.; Zhu, Y.; Shi, H.; Pan, P. Regional gray matter deficits in alcohol dependence: A meta-analysis of voxel-based morphometry studies. Drug Alcohol Depend 2015, 153, 22–28. [Google Scholar] [CrossRef]
- Hardee, J.E.; Weigard, A.S.; Heitzeg, M.M.; Martz, M.E.; Cope, L.M. Sex differences in distributed error-related neural activation in problem-drinking young adults. Drug Alcohol Depend 2024, 263, 112421. [Google Scholar] [CrossRef]
- Luo, Q.; Mitchell, D.; Jones, M.; Mondillo, K.; Vythilingam, M.; Blair, R.J. Common regions of dorsal anterior cingulate and prefrontal–parietal cortices provide attentional control of distracters varying in emotionality and visibility. NeuroImage 2007, 38(3), 631–639. [Google Scholar] [CrossRef] [PubMed]
- Murray, L.; Welsh, J.C.; Johnson, C.G.; Kaiser, R.H.; Farchione, T.J.; Janes, A.C. Alcohol- and non-alcohol-related interference: An fMRI study of treatment-seeking adults with alcohol use disorder. Drug Alcohol Depend 2022, 235, 109462. [Google Scholar] [CrossRef]
- Padula, C.B.; Tenekedjieva, L.T.; McCalley, D.M.; Al-Dasouqi, H.; Hanlon, C.A.; Williams, L.M.; Kozel, F.A.; Knutson, B.; Durazzo, T.C.; Yesavage, J.A.; et al. Targeting the Salience Network: A Mini-Review on a Novel Neuromodulation Approach for Treating Alcohol Use Disorder. Front Psychiatry 2022, 13, 893833. [Google Scholar] [CrossRef]
- Eickhoff, S.B.; Nichols, T.E.; Laird, A.R.; Hoffstaedter, F.; Amunts, K.; Fox, P.T.; Bzdok, D.; Eickhoff, C.R. Behavior, sensitivity, and power of activation likelihood estimation characterized by massive empirical simulation. Neuroimage 2016, 137, 70–85. [Google Scholar] [CrossRef]
- Garrison, J.R.; Done, J.; Simons, J.S. Interpretation of published meta-analytical studies affected by implementation errors in the GingerALE software. Neurosci Biobehav Rev 2019, 102, 424–426. [Google Scholar] [CrossRef]
- Jeon, K.H.; Han, K.; Jeong, S.M.; Park, J.; Yoo, J.E.; Yoo, J.; Lee, J.; Kim, S.; Shin, D.W. Changes in Alcohol Consumption and Risk of Dementia in a Nationwide Cohort in South Korea. JAMA Netw Open 2023, 6, e2254771. [Google Scholar] [CrossRef]
- Xu, W.; Wang, H.; Wan, Y.; Tan, C.; Li, J.; Tan, L.; Yu, J.T. Alcohol consumption and dementia risk: a dose-response meta-analysis of prospective studies. Eur J Epidemiol 2017, 32, 31–42. [Google Scholar] [CrossRef]
- Zarezadeh, M.; Mahmoudinezhad, M.; Faghfouri, A.H.; Mohammadzadeh Honarvar, N.; Regestein, Q.R.; Papatheodorou, S.I.; Mekary, R.A.; Willett, W.C. Alcohol consumption in relation to cognitive dysfunction and dementia: A systematic review and dose-response meta-analysis of comparative longitudinal studies. Ageing Res Rev 2024, 100, 102419. [Google Scholar] [CrossRef]
Figure 1.
Short-term abstinence fMRI Results. Note. A = Putamen (Hyper and hypoactivation); B = Middle frontal gyrus (hypoactivation).
Figure 1.
Short-term abstinence fMRI Results. Note. A = Putamen (Hyper and hypoactivation); B = Middle frontal gyrus (hypoactivation).
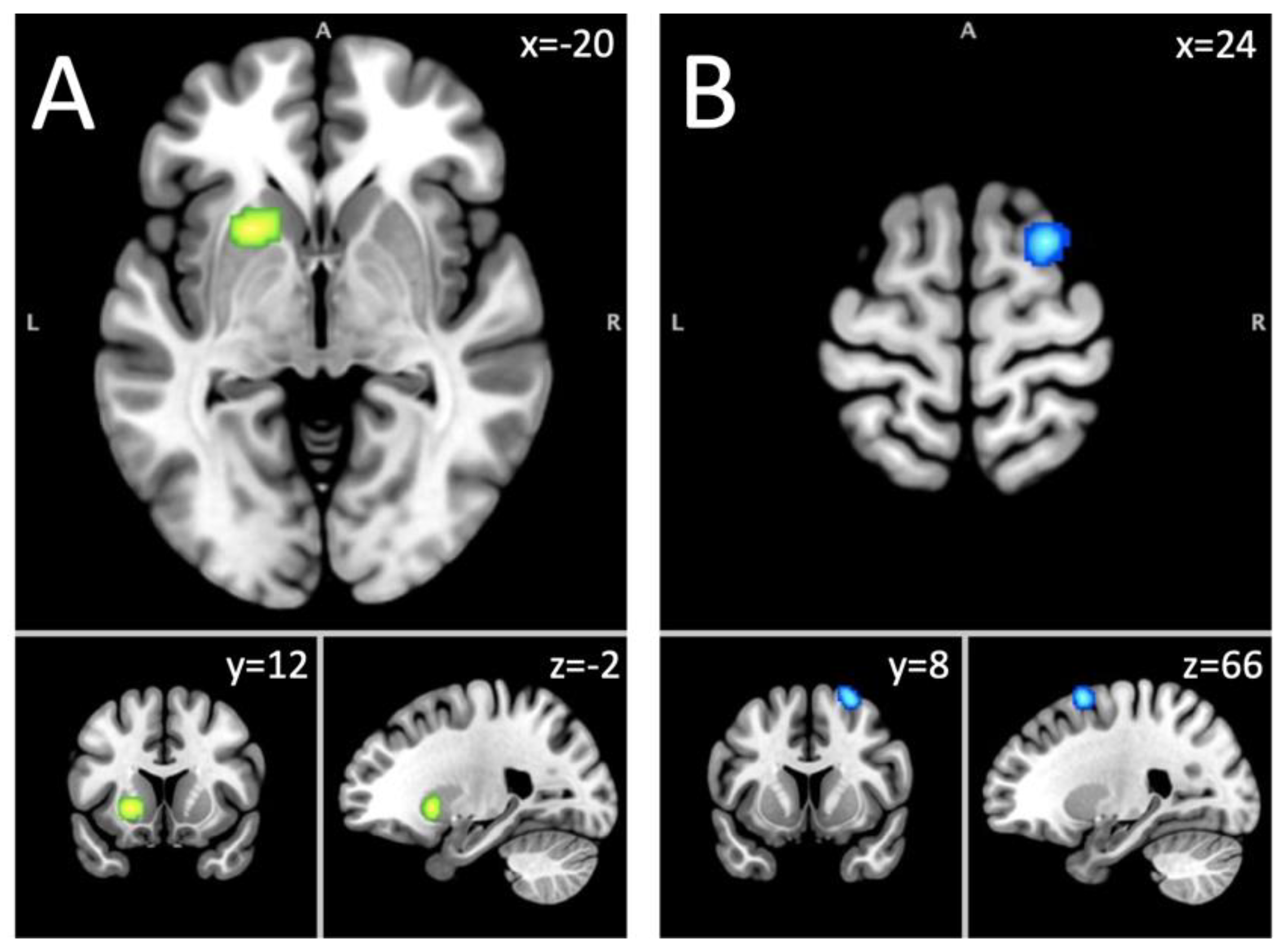
Figure 2.
Long-term abstinence fMRI Results. Note. A = Superior frontal gyrus (hypoactivation); B = Dorsal anterior cingulate cortex (hypoactivation).
Figure 2.
Long-term abstinence fMRI Results. Note. A = Superior frontal gyrus (hypoactivation); B = Dorsal anterior cingulate cortex (hypoactivation).
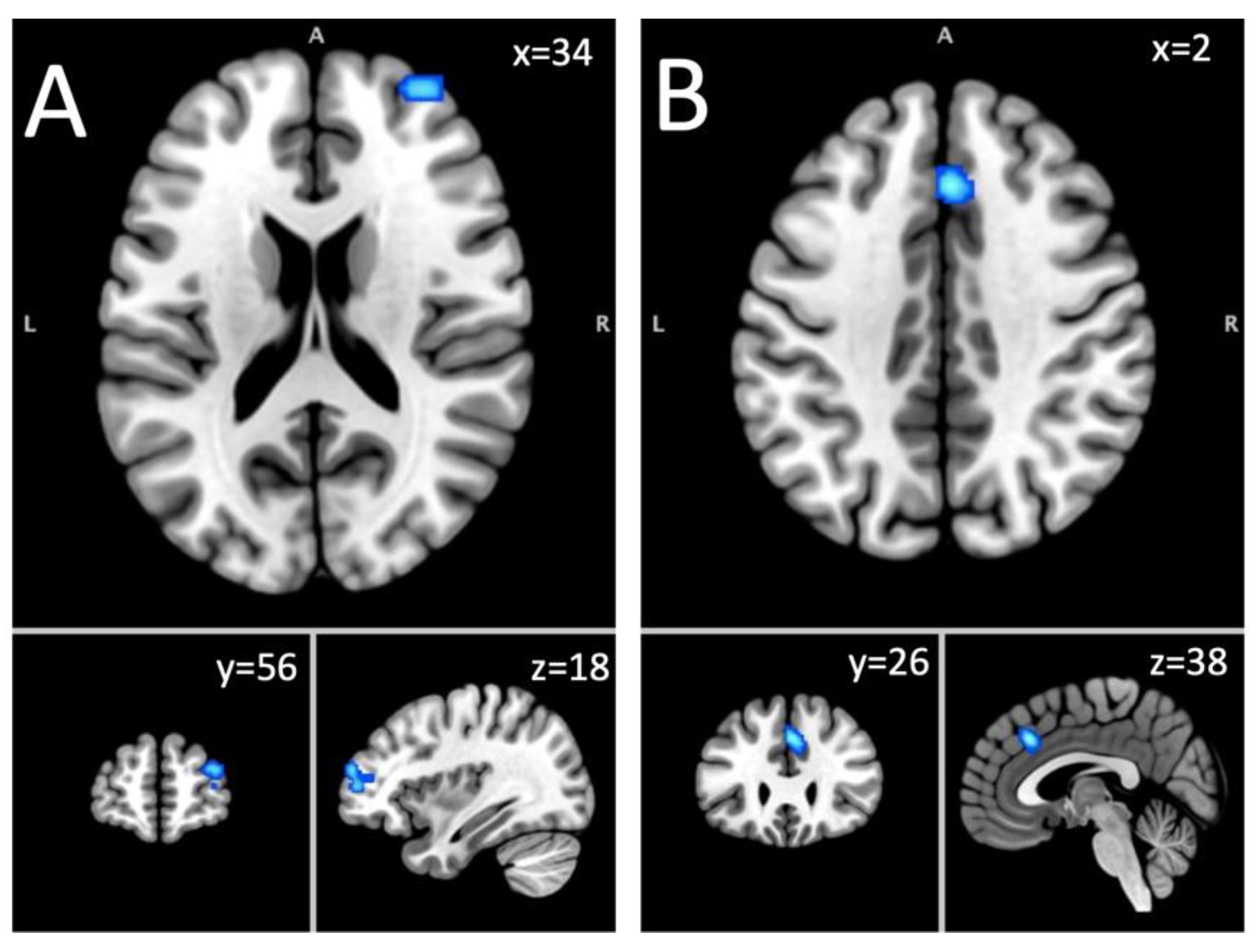

Table 1.
Demographics.
| n cases | n controls | Mean age cases | Mean age controls | % of male for cases | % of male for controls | Mean days of abstinence (range) |
|---|---|---|---|---|---|---|
| Whole sample | ||||||
| 2,421 | 1,458 | 36.94 | 37.07 | 76.73 | 73.26 | 189.54 (5-2994)a |
| Short-term Abstinent sample | ||||||
| 1,097 | 653 | 35.71 | 34.46 | 76.32 | 70.71 | 16.10 (5-25.30)b |
| Long-term Abstinent sample | ||||||
| 991 | 488 | 37.42 | 40.15 | 77.56 | 78.35 | 416.47 (34-2994)c |
Note. a = information available only for 44/67 studies; b = information available only for 23/37 studies; c = information available only for 19/26 studies.
Table 2.
Results from the ALE meta-analysis.
| Regions | L/R | Cluster size (mm3) | ALE value | Z-score | Coordinates (MNI) |
|---|---|---|---|---|---|
| Hyper-Hypoactivation combined: whole sample | |||||
| Putamen, caudate body, caudate head | L | 1840 | 0.0367 | 5.66 | -20,12,-2 |
| Hyperactivation: whole sample | |||||
| No clusters found | |||||
| Hypoactivation: whole sample | |||||
| No clusters found | |||||
| Hyper-Hypoactivation combined: acute sample | |||||
| Putamen | L | 1096 | 0.0236 | 4.88 | -20,12,-2 |
| Hyperactivation: acute sample | |||||
| No clusters found | |||||
| Hypoactivation: acute sample | |||||
| Middle frontal gyrus, superior frontal gyrus, sub-gyral | R | 856 | 0.016 | 4.48 | 24,8,66 |
| Hyper-Hypoactivation combined: abstinent sample | |||||
| No clusters found | |||||
| Hyperactivation: abstinent sample | |||||
| No clusters found | |||||
| Hypoactivation: abstinent sample | |||||
| Superior frontal gyrus, middle frontal gyrus | R | 856 | 0.0147 | 3.85 | 34,56,18 |
| Cingulate gyrus, medial frontal gyrus | R & L | 800 | 0.017 | 4.26 | 2,26,38 |
Note. L = Left; R = Right; MNI = Montreal neurologic institute.
Table 3.
Binary logistic regressions for confounding variables and probabilities of activation of each cluster.
Table 3.
Binary logistic regressions for confounding variables and probabilities of activation of each cluster.
| 95 C.I. for odds ratio | ||||
|---|---|---|---|---|
| Predictors | Odds ratio | Lower | Higher | p-value |
| Hyper-hypoactivation in the short-term abstinent sample: Putamen | ||||
| Age | 0.985 | 0.900 | 1.077 | 0.735 |
| Sex ratio (% male) | 0.981 | 0.951 | 1.013 | 0.245 |
| Days of abstinence | 0.939 | 0.746 | 1.182 | 0.590 |
| MRI field strength | 0.737 | 0.110 | 4.955 | 0.753 |
| Smoothing level | 0.701 | 0.422 | 1.164 | 0.170 |
| Voxel size | 1.047 | 0.993 | 1.103 | 0.088 |
| Time repetition | 0.998 | 0.996 | 1.000 | 0.115 |
| Craving studies | 0.489 | 0.050 | 4.793 | 0.539 |
| Decision-making studies | 9.333 | 1.270 | 68.597 | 0.028* |
| Emotion studies | 1.350 | 0.124 | 14.734 | 0.806 |
| Executive functions studies | 0.364 | 0.038 | 3.518 | 0.382 |
| Reward processing studies | 30.000 | 2.330 | 386.325 | 0.009* |
| Other task studies | 3.375 | 0.459 | 24.837 | 0.232 |
| Hypoactivation in the short-term abstinent sample: Middle frontal gyrus | ||||
| Age | 0.942 | 0.854 | 1.040 | 0.234 |
| Sex ratio (% male) | 0.985 | 0.951 | 1.021 | 0.417 |
| Days of abstinence | 0.859 | 0.664 | 1.113 | 0.251 |
| MRI field strength | 0.762 | 0.060 | 9.611 | 0.833 |
| Smoothing level | 1.068 | 0.648 | 1.760 | 0.797 |
| Voxel size | 1.054 | 0.989 | 1.123 | 0.108 |
| Time repetition | 1.000 | 0.999 | 1.001 | 0.924 |
| Craving studies | 0.000 | 0.000 | 0.000 | 0.999 |
| Decision-making studies | 7.250 | 0.786 | 66.842 | 0.080 |
| Emotion studies | 0.000 | 0.000 | 0.000 | 0.999 |
| Executive functions studies | 2.300 | 0.283 | 18.705 | 0.436 |
| Reward processing studies | 3.333 | 0.259 | 42.925 | 0.356 |
| Other task studies | 1.867 | 0.160 | 21.742 | 0.618 |
| Hypoactivation in the long-term abstinent sample: Superior frontal gyrus | ||||
| Age | 0.963 | 0.894 | 1.038 | 0.323 |
| Sex ratio (% male) | 0.993 | 0.950 | 1.037 | 0.746 |
| Days of abstinence | 1.002 | 0.999 | 1.004 | 0.153 |
| MRI field strength | 0.500 | 0.066 | 3.770 | 0.501 |
| Smoothing level | 0.586 | 0.278 | 1.233 | 0.159 |
| Voxel size | 0.980 | 0.939 | 1.022 | 0.337 |
| Time repetition | 1.001 | 0.999 | 1.004 | 0.267 |
| Craving studies | 0.000 | 0.000 | 0.000 | 0.999 |
| Decision-making studies | 0.000 | 0.000 | 0.000 | 1.000 |
| Emotion studies | 3.800 | 0.201 | 72.000 | 0.374 |
| Executive functions studies | 1.167 | 0.166 | 8.186 | 0.877 |
| Reward processing studies | 9.000 | 1.031 | 78.574 | 0.047* |
| Other task studies | 1.500 | 0.208 | 10.823 | 0.688 |
| Hypoactivation in the long-term abstinent sample: Cingulate gyrus | ||||
| Age | 1.025 | 0.931 | 1.128 | 0.617 |
| Sex ratio (% male) | 1.022 | 0.964 | 1.084 | 0.456 |
| Days of abstinence | 0.997 | 0.990 | 1.004 | 0.387 |
| MRI field strength | 1.125 | 0.097 | 13.036 | 0.925 |
| Smoothing level | 0.956 | 0.450 | 2.031 | 0.906 |
| Voxel size | 0.999 | 0.948 | 1.052 | 0.955 |
| Time repetition | 1.000 | 0.997 | 1.002 | 0.708 |
| Craving studies | 1.500 | 0.122 | 18.441 | 0.751 |
| Decision-making studies | 0.000 | 0.000 | 0.000 | 1.000 |
| Emotion studies | 0.000 | 0.000 | 0.000 | 0.999 |
| Executive functions studies | 0.000 | 0.000 | 0.000 | 0.999 |
| Reward processing studies | 6.333 | 0.630 | 63.639 | 0.117 |
| Other task studies | 3.400 | 0.377 | 30.655 | 0.275 |
Note. MRI = magnetic resonance imaging.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



