Submitted:
03 May 2025
Posted:
05 May 2025
You are already at the latest version
Abstract
An ancient repertoire of ultraviolet (UV)-absorbing pigments which survive today in the phylogenetically oldest extant photosynthetic organisms, the cyanobacteria, point to a direction in evolutionary adaptation of the pigments and their associated biota; from largely UV-C absorbing pigments in the Archean to pigments covering ever more of the longer wavelength UV and visible in the Phanerozoic. Since photoprotection is not dependent on absorption, such a scenario could imply selection of photon dissipation rather than photoprotection over the evolutionary history of life, consistent with the thermodynamic dissipation theory of the origin and evolution of life which suggests that the most important hallmark of biological evolution has been the covering of Earth’s surface with organic pigment molecules and water to absorb and dissipate ever more completely the prevailing surface solar spectrum. In this article we compare a set of photophysical, photochemical, biosynthetic and other germane properties of the two dominant classes of cyanobacterial UV-absorbing pigments, the mycosporine-like amino acids (MAAs) and scytonemins. Pigment wavelengths of maximum absorption correspond with the time dependence of the prevailing Earth surface solar spectrum, and we proffer this as evidence for the selection of photon dissipation rather than photoprotection over the history of life on Earth.
Keywords:
Cyanobacteria
; UV-absorbing pigments
; mycosporine-like amino acids
; scytonemins
; photon dissipation
; thermodynamic dissipation theory
; non-equilibrium thermodynamics
; photoprotection vs. dissipation
; evolution of pigments
; Archaean Earth
; entropy production
1. Introduction
Once the subject of mystical and metaphysical interpretations, the explanation of life on Earth has slowly gained a physical-chemical grounding in biochemistry and non-equilibrium thermodynamics. Founded on Boltzmann’s 19th century insights into thermodynamics, then further elaborated by 20th century scientists, notably by Ilya Prigogine, non-equilibrium thermodynamics attempts to explain the phenomenon of life as “dissipative structuring” – an out of equilibrium organization of matter in space and time under an impressed external potential for the explicit purpose of producing entropy (Prigogine, 1967; Glansdorff and Prigogine, 1971).
Considering these non-equilibrium thermodynamic aspects of life, and using the formalism of Prigogine’s “Classical Irreversible Thermodynamics” in the non-linear regime, Michaelian (2009; 2011; 2012; 2013; 2016; 2017; 2021; 2023; 2024) has proposed a theory for life’s origin and evolution as microscopic self-organized dissipative structuring of organic pigment molecules and the dispersal of these over the entire surface of the Earth as a response to the impressed high-energy (UV-C to visible) solar photon spectrum prevailing at Earth’s surface. All physicochemical structuring associated with the pigments, such as the photosynthetic organisms primarily, and heterotrophic organisms secondarily, can be regarded as agents for the synthesis, proliferation and distribution of the pigments (Michaelian, 2023, 2024). The theory suggests that it is the thermodynamic imperative of increasing the entropy production of Earth in its solar environment that is behind the vitality of living matter in its ability to proliferate, diversify and evolve.
The theory explains satisfactorily, for example, why the three major classes of photosynthetic pigments (chlorophylls, carotenoids and phycobilins) of phototrophic organisms dissipate most of the absorbed photonic energy into heat (a process known as non-photochemical quenching, NPQ) while funneling only a minute fraction into productive photochemistry (Horton et al., 1996; Ruban et al., 2007; Staleva et al., 2015; Gupta et al., 2015; Son et al. 2020; Murchie and Ruban, 2020). Moreover, these organisms often contain a vast array of other organic pigments in addition to the photosynthetic ones, whose absorption spectra extend outside of the photosynthetically active radiation (PAR) in the visible, and into the UV-A, UV-B, and UV-C regions, hence allowing full coverage of the past and present incident surface solar spectra (Michaelian, 2012; Michaelian and Simeonov, 2015). In contradistinction to the thermodynamic dissipation theory, in the biological literature this phenomenon of full spectral coverage has been explained primarily through invoking the conventional wisdom of photoprotection (Demmig-Adams and Adams, 1992; Mulkidjanian and Junge, 1997; Wynn-Williams et al., 2002; Mulkidjanian et al., 2003; Castenholz and Garcia-Pichel, 2012; Fuentes-Tristan et al., 2019).
Photoprotective roles have especially been attributed to UV-absorbing biological pigments (e.g., mycosporine-like amino acids and scytonemins in cyanobacteria and algae; flavonoids and anthocyanins in plants, etc.) since they don’t seem to contribute to photosynthesis at all (Moisan and Mitchell, 2001; Fuentes-Tristan et al., 2019). These theories usually trace the photoprotective role of UV-pigments back to the beginnings of cellular life in the early Archean when UV radiation was far more important component of the surface solar spectrum than it is today (Sagan, 1973; Mulkidjanian and Junge, 1997; Garcia-Pichel, 1998; Cockell and Knowland, 1999; Mulkidjanian et al., 2003). UV-screening ostensibly conferred pigment-containing organisms Darwinian advantages for survival in the harsh Archean environment of intense UV radiation.
However, from a thermodynamic viewpoint, the UV-C is the region of the solar spectrum with an energy per photon sufficient for directly reconfiguring covalent bonds and it also corresponds to the greatest possible entropy production potential per unit photon dissipated. Therefore, under the high UV ambient conditions of our primitive planet, non-equilibrium thermodynamic principle of increasing the entropy production of Earth in its solar environment through dissipative structuring was probably the motive force for the dissipative synthesis of these organic pigments (Michaelian, 2017; 2021). The evidence for this inextricable link between UV light and nascent life has been reinforced with the verification of the biogenicity of ~ 3.5 Ga old euphotic stromatolite formations (Walter et al., 1980; Awramik et al., 1983; Schopf and Packer, 1987; Schopf, 1993; Schopf et al., 2002; Tice and Lowe, 2004; Van Kranendonk et al., 2008; Hickman-Lewis et al., 2023) of evidently photosynthetically active, yet UV-C bathed, microbial colonies of cyanobacteria-like organisms (Westall et al., 2006; Westall, 2009).
Here we discuss how the two major classes of cyanobacterial UV-absorbing pigments, the mycosporine-like amino acids and the scytonemins, whose occurrence in organisms today is regarded as a relic from Archean times (Garcia-Pichel, 1998; Rosic, 2021), concur with the type of microscopic dissipative structure which non-equilibrium thermodynamic principles would predict for microscopic dissipative structuring under the Archean Earth conditions.
In the following section we separately detail a set of germane properties of both pigment classes, taken from the literature. In the third section we demonstrate how these properties are consistent with several postulates concerning photon dissipation made in previous work (Michaelian and Simeonov, 2015). Also, in the third section, we compare the pigments’ optical properties with our previous construct of the most probable Earth surface solar spectrum as a function of time (see Michaelian and Simeonov, 2015).
2. Properties of Cyanobacterial UV-Absorbing Pigments
2.1. Mycosporine-Like Amino Acids (MAAs)
MAAs and a related group of organic compounds called mycosporines represent a large family of colorless, low-molecular-weight (< 400 u), water-soluble, usually intracellular secondary metabolites widespread in the biological world (Dunlap and Chalker, 1986; Carreto et al., 1990; Rosic and Dove, 2011; Sun et al., 2020). The exact number of compounds within this family is yet to be determined, since they have only relatively recently been discovered (for a historical overview, see Schick and Dunlap, 2002 and Řezanka et al., 2004), and novel molecular species are constantly being discovered. Thus far, however, their number is around 70 (Geraldes and Pinto, 2021). The name “mycosporine” has to do with them being originally isolated and chemically identified from mycelia of sporulating fungi, where it was thought they played a role in light-induced sporulation (Leach, 1965; Trione and Leach, 1969).
2.1.1. Physicochemical Properties
Chemically both MAAs and mycosporines are alicyclic compounds (see Figure 1) sharing a central 5-hydroxy-5-hydroxymethyl-2-methoxycyclohex-2-ene ring with an amino compound substituted at the third C-atom and either an oxo or an imino functionality at the first C-atom (Favre-Bonvin et al., 1976; Ito and Hirata, 1977; Arpin et al., 1979; Karentz, 2001; Geraldes and Pinto, 2021). While most authors don’t make a clear chemical distinction between the two groups, several authors (for example, Bandaranayake, 1998; Schick and Dunlap, 2002; Carreto et al., 2011; Moliné et al., 2014; Miyamoto et al., 2014) when using the term mycosporines refer only to those molecular species with a central amino-cyclohexenone chromophore (also called oxo-mycosporines), and when using the term MAAs refer only to molecules with a central amino-cyclohexenimine chromophore (also called imino-mycosporines). The N-substitution on C-3 with different amino acids or amino alcohols is what gives the diversity of molecular structures within both groups (Korbee et al., 2006; Sinha et al., 2007; Carreto and Carignan, 2011). Within the MAA group, the most common amino acid on the C-3 position is glycine, whereas they also have a second amino acid, amino alcohol, or an enaminone system attached at the C-1 position (D’Agostino et al., 2016).
This unique molecular structuring and bonding underlies their unique spectral properties.
MAAs are considered to be one of the strongest UV-A/UV-B-absorbing substances in nature (Schmid et al., 2000); with wavelength absorption maxima () in the 310-362 nm interval and molar attenuation coefficients () between 28,100 and 50,000 (Dunlap and Schick, 1998; Carreto et al., 2005; Gao and Garcia-Pichel, 2011; Singh et al., 2021).
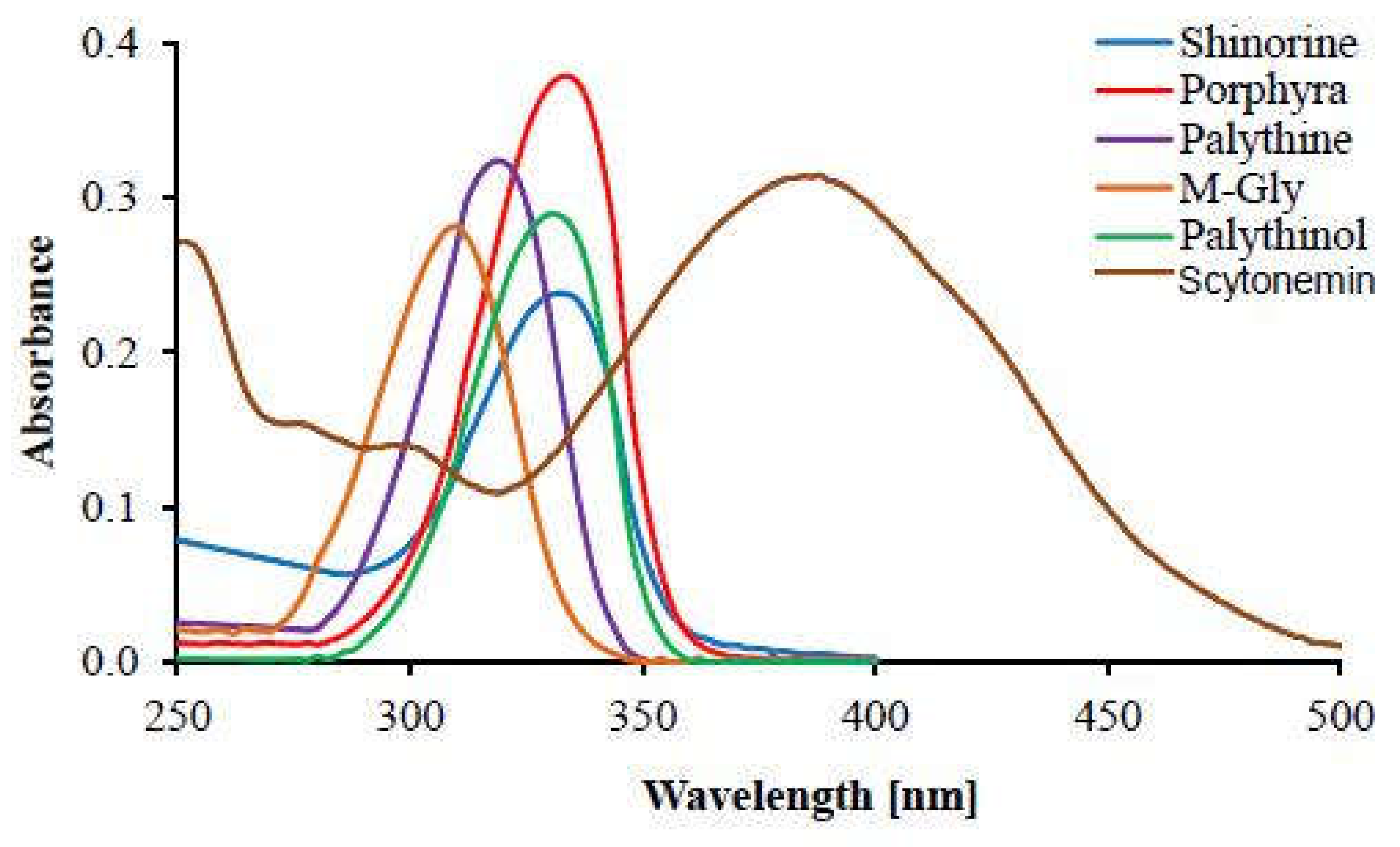
Their absorption spectra are characterized by a single sharp peak with a width of approximately 20 nm and only about 2–3 nm apart from the of structurally similar MAAs (see Figure 5), making it very difficult to distinguish these compounds based solely on their absorption spectra (Karentz, 1994; Carroll and Shick, 1996; Carreto and Carignan, 2011).
The values of and are dependent on the degree of derivatization of the central ring and the nature of the nitrogenous side groups (in particular, the presence of additional conjugated double bonds) (Singh et al., 2010; Wada et al., 2015).
Smaller, mono-substituted mycosporines (typically oxo-mycosporines) have their values shifted to shorter wavelengths in the UV-B and usually have lower values; whereas MAAs (imino-mycosporines) are normally bi-substituted, with higher values and values shifted to longer wavelengths in the UV-A (Portwich and Garcia-Pichel, 2003; Rosic, 2019).
For example, the direct metabolic precursor of all mycosporines, 4-deoxygadusol (Figure 1), which has the minimal level of derivatization, has at acidic pH, and at basic pH; mycosporine-glycine (Figure 1), the simplest oxo-mycosporine and direct precursor of all other mycosporines and MAAs has a , whereas palythine (Figure 1), a simple mono-substituted MAA (amino-MAA), has and (Carreto et al., 2005; Gao and Garcia-Pichel, 2011). Palythene (Figure 1), a bi-substituted MAA with an additional conjugated double bond, has one of the most red-shifted bands of all known MAA species, with and (Uemura et al., 1980; Wada et al., 2015).
The observed red-shift in is a consequence of the degree of resonance delocalization inside the molecules; the more efficient is the electron delocalization (i.e. the stronger the π-conjugation character) the lower is the energy requirement for an electronic transition and consequently the higher are the and values (Carreto and Carignan, 2011; Wada et al., 2015).
From a thermodynamic perspective, the fate of the electronic excitation energy is also a very relevant aspect of the absorption event since it is directly linked to the amount of entropy produced by the dissipative microscopic structure (i.e., the pigment molecule). Nonradiative, vibrational relaxation pathways of the excited state lead to more efficient energy dissipation and higher entropy production when compared to the fluorescent or phosphorescent radiative decay channels (Würfel and Ruppel, 1985; Michaelian, 2011; 2012; 2016; 2021; 2023; 2024). In this respect MAAs prove to be very efficient dissipative structures.
Aiming to fully describe their photophysical and photochemical properties and increase the evidence for the assigned UV-photoprotective role, Conde et al. (2000; 2004; 2007) made several in vitro studies on the excited-state properties and photostability of various MAAs in aqueous solution (Table 1). The results showed picosecond excited state lifetimes, very low fluorescence quantum yields (e.g.,), very low triplet formation quantum yields (e.g.,), and very low photodegradation quantum yields (e.g.,) for all the MAAs studied. These results are consistent with a very fast internal conversion (IC) process as the main deactivation pathway of the excited states, which was directly quantified by photoacoustic calorimetry experiments showing that ~ 97% of the absorbed photonic energy is promptly dissipated into the surrounding medium as heat (Conde et al., 2004; Lawrence et al., 2018; Hatakeyama and Nakamura, 2024).
A computational study by Sampedro (2011) employing palythine as a model compound, confirmed these findings. The study indicates that the fast IC processes connecting the and states are due to the presence of two energetically accessible conical intersection points that can be reached by small geometrical changes in the atomic coordinates. It is now well established that conical intersections (a.k.a. molecular funnels or diabolic points) play a very important role in fast, non-radiative de-excitation transitions from excited electronic states to ground electronic state of molecules, particularly in many fundamental biological molecules, such as DNA/RNA, amino acids and peptides (Schermann, 2008; Michaelian and Simeonov, 2015). They enable effective coupling of the electronic degrees of freedom of the molecule to its phonon degrees of freedom, thereby facilitating radiationless decay by vibrational cooling to the ground state (in the process converting the absorbed high frequency UV photon into many low frequency infrared photons), making them examples of microscopic dissipative structures, formed in response to the impressed photon potential (Michaelian, 2016; 2017; 2021).
2.1.2. Ecological Distribution
MAAs and mycosporines are cosmopolitan substances in “optical” habitats – planktonic, benthic and terrestrial; with the largest concentrations detected in environments with water exposed to high levels of solar irradiance (Castenholz and Garcia-Pichel, 2012 and references therein). They are now known to be the most common type of UV-absorbing natural substances, especially among aquatic organisms (Rastogi et al., 2010; Sun et al., 2020).
While mycosporines have been found only in the kingdom Fungi (mycosporine-glycine and mycosporine-taurine are exceptions), MAAs are more extensively distributed among taxonomically diverse organisms (Karsten, 2008; Carreto and Carignan, 2011). These include cyanobacteria; heterotrophic bacteria; dinoflagellates, diatoms and other protists; red algae; green algae; various marine animals, especially corals and their associated biota (for a database on the distribution of MAAs, see Sinha et al., 2007). They seem to be completely absent from terrestrial plants and animals but are regularly found in terrestrial cyanobacteria (Garcia-Pichel and Castenholz, 1993; Jain et al., 2017) and terrestrial algae (Karsten et al., 2007).
An interesting discovery by Ingalls et al. (2010) reveals that MAAs represent a considerable portion of the organic matter bound to diatom frustules, accounting for 3–27% of the total carbon and 2–18% of total nitrogen content of the frustules. Previously established views held that MAAs have mainly an intracellular location in these organisms.
2.1.3. Biosynthesis
The cyclohexenone core of MAAs is derived from intermediates of two fundamental anabolic pathways; the shikimate pathway (Favre-Bonvin et al., 1987; Shick et al., 1999; Portwich and Garcia-Pichel, 2003) and the pentose phosphate pathway (Balskus and Walsh, 2010), with the shikimate pathway being the predominant route for UV-induced MAA biosynthesis (Pope et al., 2015; Arsın et al., 2023).
In MAA/mycosporine biosynthesis both pathways converge at a point where their respective 6-membered cyclic intermediates with similar structures are converted to 4-deoxygadusol (Figure 1), the common precursor of all MAAs and mycosporines, a reaction catalyzed by the key enzyme O-methyltransferase (Pope et al., 2015; D’Agostino et al., 2016).
These basic biochemical pathways lie at the heart of carbon metabolism, shared by all three domains of life; the shikimate pathway links carbohydrate catabolism to the biosynthesis of the aromatic amino acids and other aromatic biomolecules; similarly, the pentose phosphate pathway uses glycolysis for the synthesis of pentose sugars, the nucleotide building blocks (Cohen, 2014). Thus, they are considered to have an ancient evolutionary origin, possibly even dating back to prebiotic times (Richards et al., 2006; Keller et al., 2014; Hirakawa et al., 2024).
As mentioned in the previous section a very interesting trait of MAAs is that they are extremely prevalent natural compounds produced by a variety of taxonomically very distant organisms from simple bacteria to algae and animals. A natural question arises: how can so evolutionary distant organisms share the same MAA encoding genes?
Several lines of evidence now suggest that the progenitor of the enzymatic machinery for MAA biosynthesis was probably a cyanobacterium or the cyanobacterial ancestor, while endosymbiotic events and prokaryote-to-eukaryote lateral gene transfer events during evolution account for their prevalence among all other biological taxa (Rozema et al., 2002; Waller et al., 2006; Starcevic et al., 2008; Singh et al., 2010; Singh et al., 2012; Sun et al., 2023).
2.1.4. Function: Traditional Protective View vs. Thermodynamic View
Since their discovery in the 1960’s, authors have struggled to confer specific physiological functions to MAAs. A UV-photoprotective role seemed most conspicuous, largely because of their unique UV-dissipating traits and the fact that their production is stimulated by UV-B light. Later, this theory faced serious challenges, for example, the failure to find a correlation between intracellular MAA accumulation and UV-resistance in certain coral zooxanthellae (Kinzie, 1993), the phytoplankton Phaeocystis antarctica (Karentz and Spero, 1995), the dinoflagellate Prorocentrum micans (Lesser, 1996), certain cyanobacterial strains (Quesada and Vincent, 1997), the red alga Gracilaria cornea (Sinha et al., 2000), Antarctic sea ice algae (Ryan et al., 2002), the red alga Palmaria palmata (Karsten et al., 2003), and others.
As a response, many researchers in the field came up with their own suggestions for MAA physiological roles, sometimes very different from the sunscreen role, such as; osmotic regulation, antioxidants, nitrogen storage, accessory pigments, protection from desiccation, protection from thermal stress, reproductive functions in fungi and marine invertebrates etc.; all of which have also been challenged or discredited (for reviews of the different theories of MAA functions and the challenges they face, see: Korbee et al., 2006; Oren and Gunde-Cimerman, 2007; Rosic and Dove, 2011; Zwerger et al., 2023).
From a traditional biological standpoint, this apparent lack of a clear defining physiological function looks extremely perplexing, especially when considering the extraordinary prevalence of these compounds in nature. Darwinian theory, in its strictest traditional formulation with evolution through natural selection operating only at the individual level, categorically dismisses this kind of phenomena. An organism would not wastefully spend free energy and resources for the synthesis of metabolically expensive nitrogen-containing compounds with no vital physiological function commensurate with their ubiquity and hence no, or little, benefit for its survival and reproduction. According to Darwinian theory, such a biosynthetic pathway, with little or no direct utility to the organism, should have been suppressed or eliminated through natural selection. However, exactly the opposite has happened during evolution; MAA biosynthetic genes have not only survived but have undergone extensive dissemination across numerous taxa through horizontal gene transfer.
We postulate that the failure of Darwinian theory to find a niche for MAAs in its classical “struggle for survival” paradigm may be related to the fact that this theory is not founded on thermodynamic principles (for a discussion on this topic, see: Michaelian, 2011; 2012; 2016; 2023). From the perspective of non-equilibrium thermodynamics MAAs do have a function, and it is a thermodynamic function of energy dissipation, or, more generally, entropy production. This thermodynamic function can be readily inferred from their physicochemical properties related to photon dissipation described above. MAAs can be regarded as typical examples of microscopic dissipative structuring of matter for the sole purpose of entropy production through highly efficient dissipation of high-frequency UV photons into heat (Michaelian, 2009, 2011; 2016; 2017; 2021). This may be the reason for their ‘coming into being’ and tendency to proliferate over the surface of the Earth, as it is the fundamental reason for the origin and evolution of life on Earth, and, in fact, the reason for the ubiquity of organic pigments in the neighborhood of stars throughout the universe (Michaelian and Simeonov, 2017; Michaelian, 2016).
This irreversible process of photon dissipation that MAAs and other bio-pigments perform, then couples to a secondary abiotic irreversible process of the water cycle through increased evaporation from surfaces through the heat of photon dissipation released into the aqueous milieu (Michaelian, 2012). Evidence exist that the profusion of life and chromophoric dissolved organic matter (CDOM) in the sea-surface microlayer (SML) causes significant heating of the ocean surface fomenting evaporation (Morel, 1988; Kahru et al., 1993; Jones et al., 2005; Patara et al., 2012; Wurl et al., 2018; Soppa et al., 2019). The heat of dissipation in the surface layer can even foment another irreversible process, hurricane formation and steering (Gnanadesikan et al., 2010).
CDOM is the fraction of dissolved organic matter in water (DOM) that interacts with solar radiation (Nelson and Siegel, 2013; Logozzo et al., 2021). Light energy absorption by CDOM at the surface of the ocean usually exceeds that of phytoplankton pigments; 5415% of the total light absorption at 440 nm and > 70% of the total light absorption at 300 nm is due to CDOM (Siegel et al., 2002; Babin et al., 2003; Bricaud et al., 2010; Organelli et al., 2014). It is a complex and extremely variable mixture of organic pigments such as pheopigments (Bricaud et al., 2010), metal-free porphyrins (Röttgers and Koch, 2012), humic and fulvic acids (Carlson and Mayer, 1980; Galgani and Engel, 2016), aromatic amino acids (Yamashita and Tanoue, 2003) and MAAs (Whitehead and Vernet, 2000; Steinberg et al., 2004; Tilstone et al., 2010; Clementson et al., 2022). While it was previously believed that CDOM in the open ocean is chiefly a byproduct of heterotrophic organisms recycling phytoplankton cell contents (Nelson et al., 1998), more recent observations (Romera-Castillo et al., 2010; Fukuzaki et al., 2014) suggest a large contribution from active plankton exudation.
Active secretion of MAAs into the surrounding water during surface blooms was demonstrated for the colonial cyanobacterium Trichodesmium spp (Subramaniam et al., 1999; Steinberg et al., 2004), for the dinoflagellate Lingulodinium polyedrum (Vernet and Whitehead, 1996; Whitehead and Vernet, 2000) and for the dinoflagellate Prorocentrum micans (Tilstone et al., 2010). Interestingly, Tilstone et al. (2010) found far greater MAA concentration in the sea-surface microlayer samples when compared to the near-surface (0–2 m), and subsurface (0–110 m) samples. Whitehead and Vernet (2000) also concluded that free-floating MAAs contributed up to 10% of the UV absorption of the total DOM pool at 330 nm during the L. polyedrum bloom. This exudation of pigments by organisms into their environment would also seem to have little Darwinian advantage.
The evidence presented above strongly suggests that MAAs are, like most of the other bio-pigments in nature, catalysts for the dissipation of photons into heat at Earth’s surface and the coupling of this heat to other abiotic entropy producing processes, such as; the water cycle, hurricanes, water and wind currents, etc. (Llewellyn et al., 2020).
2.2. Scytonemins
Scytonemin was first described in the 19th century by Swiss botanist Carl Nägeli (Nägeli, 1849; Nägeli and Schwenderer, 1877) but was not isolated until 1991 when Garcia-Pichel and Castenholz (1991) made a more in-depth study of the compound. Proteau et al. (1993) elucidated the chemical structure of scytonemin, which proved to be a completely novel indolic-phenolic dimeric structure, unique among all hitherto known natural organic substances. The carbon skeleton of this novel eight-ring homodimeric molecule was given the trivial name “the scytoneman skeleton” (Proteau et al., 1993). In 1994, another scytoneman-type molecule was isolated from the cyanobacterium Nostoc commune, and termed “nostodione A” (Kobayashi et al., 1994). Thus far, four additional substances with a scytoneman-type molecular structure, or a structure derived from it, have been isolated from cyanobacteria: dimethoxyscytonemin, tetramethoxyscytonemin, scytonine (Bultel-Poncé et al., 2004; Orellana et al., 2020) and scytonemin-imine (Grant and Louda, 2013); for which, in this review, we use the colloquial term “scytonemin” or “scytoneman” pigments.
2.2.1. Physicochemical Properties
Scytonemin (Figure 2), the representative and most common member of this yet poorly explored family of aromatic indole alkaloids, is a relatively small molecule (544 u) built from two identical condensation products of tryptophanyl- and tyrosyl-derived subunits linked through a carbon-carbon bond (Proteau et al., 1993).
Depending on the redox conditions it can exist in two inter-convertible forms: a predominant oxidized yellowish-brown form which is insoluble in water and only fairly soluble in organic solvents, such as pyridine and tetrahydrofuran, and a reduced form (Figure 2) with bright red color that is slightly more soluble in organic solvents (Garcia-Pichel and Castenholz, 1991; Proteau et al., 1993; Gao et al., 2021).
Dimethoxy- and tetramethoxyscytonemin can be considered as derivatives of reduced scytonemin, where one or both ethenyl groups in the molecule have been saturated by two or four methoxy groups, respectively (Bultel-Poncé et al., 2004; Varnali and Edwards, 2010). Another moderate degree of modification of the parent scytoneman skeleton can also be seen in scytonemin-3a-imine (a.k.a. scytonemin-imine), where the C-3a atom of scytonemin has been appended with a 2-imino-propyl radical (Grant and Louda, 2013).
Only the structure of scytonine deviates substantially from the dimeric scytoneman skeleton, due to the loss of one para-substituted phenol group and ring openings of both cyclopentenones where successive methoxylation and secondary cyclizations take place (Bultel-Poncé et al., 2004).
A full in-depth photophysical and photochemical characterization of scytonemins has yet to be attained; thus far only their elemental spectroscopic properties are known.
Scytonemin absorbs very strongly and broadly across the UVC-UVB-UVA-violet-blue spectral region (see Figure 4 and Figure 5), with in vivo at 370 nm and in vitro (tetrahydrofuran) at 386 and 252 nm, with smaller peaks at 212, 278 and 300 nm (Garcia-Pichel and Castenholz, 1991; Garcia-Pichel et al., 1992; Sinha et al., 1999; Rastogi et al., 2013). Its observed long-term persistence in cyanobacterial biocrusts or dried mats exposed to intense solar radiation might be an indication of exceptionally high photostability (Garcia-Pichel et al., 1992; Brenowitz and Castenholz, 1997; Fleming and Castenholz, 2007; Fulton et al., 2012; Lepot et al., 2014).
Reduced scytonemin has a similar spectroscopic profile, with in vitro (tetrahydrofuran) (nm) and values: 246 (30,000), 276 (14,000), 314 (15,000), 378 (22,000), 474 (14,000) and 572 (broad shoulder 7600) (Varnali and Edwards, 2014).
A comparable absorption spectrum is also exhibited by scytonemin-imine, the mahogany-colored, polar derivative of scytonemin, with slightly different values when measured in acetone (237, 366, 437 and 564 nm) and in ethanol (248, 305, 364, 440 and 553 nm) (Grant and Louda, 2013).
Contrary to these three scytoneman-type molecules, the methoxylated derivatives and scytonine do not absorb strongly in the UV-A region but have very high absorbances in the UV-C region with in vitro (methanol) (nm) and values for dimethoxyscytonemin: 215 (60,354), 316 (18,143) and 422 (23,015); for tetramethoxyscytonemin: 212 (35,928) and 562 (5944); and for scytonine: 207 (38,948), 225 (37,054) and 270 (22,484) (Bultel-Poncé et al., 2004; Orellana et al., 2020).
Concerning the monomeric scytoneman-type molecules nostodione A and prenostodione, isolated from natural cyanobacterial blooms, it remains debatable whether they are genuine cyanobacterial pigments or just intermediates in the biosynthesis of scytonemin (Ploutno and Carmeli, 2001; Soule et al., 2009a).
2.2.2. Ecological Distribution
Unlike MAAs, scytonemins are exclusively cyanobacterial sheath pigments (Pathak et al., 2016).
All phylogenetic lines of sheathed cyanobacteria contain scytonemins (Proteau et al., 1993), most notable strains are of the genera Nostoc, Calothrix, Scytonema, Rivularia, Chlorogloeopsis, Lyngbya, Hyella, etc. (Sinha and Häder, 2008); as well as cyanolichens of the genera Peltula, Collema and Gonohymenia (Büdel et al., 1997).
The mucilaginous extracellular sheath (matrix) consists of heteroglycans, peptides, proteins, DNA and different secondary metabolites (Pereira et al., 2009; Kehr and Dittmann, 2015), where scytonemins are usually deposited in the outer layers, giving the sheath its distinctive dark yellow to brown color (Ehling-Schulz et al., 1997; Ehling-Schulz and Scherer, 1999; Kehr and Dittmann, 2015). Up to 5% of the dry weight of cultured scytonemin-synthesizing cyanobacteria is due to the pigment, but in natural assemblages this value can be even higher (Karsten et al., 1998; Gao et al., 2021). Curiously, Abed et al. (2010) reported two to six times higher concentrations of scytonemin than chlorophyll a in cyanobacterial cryptobiotic soil crusts in the Oman Desert.
Scytonemin-producing cyanobacteria typically inhabit highly insolated terrestrial, freshwater and coastal environments such as deserts, exposed rocks, cliffs, marine intertidal flats, shallow oligotrophic fresh waters, hot springs, etc. (Castenholz and Garcia-Pichel, 2012 and references therein). In microbial mat communities, especially the extremophilic terrestrial and aquatic colonies, these cyanobacteria occupy the uppermost sunlit layers (Balskus et al., 2011). Scytonemin-imine, for example, was isolated from samples of natural Scytonema hoffmani mats growing under high to intense (300 – 2000 μmol quanta m-2 s-1) photon flux density (Grant and Louda, 2013). The methoxyscytonemins and scytonine were isolated alongside scytonemin from colonies of Scytonema sp. growing on exposed granite at the Mitaraka inselberg in French Guyana, a region subjected to intense UVR-insolation (Bultel-Poncé et al., 2004).
2.2.3. Biosynthesis
The biochemistry and genetics of cyanobacterial scytonemin biosynthesis has been extensively investigated by Soule et al. (2007; 2009a; 2009b), Balskus and Walsh (2008; 2009; 2011), Sorrels et al. (2009) and Gao et al. (2021). They confirm the assumption by Proteau et al. (1993), the discoverers of the scytonemin structure, that the scytoneman molecular scaffold is a condensation product of the aromatic amino acids tryptophan and tyrosine.
Not only do these amino acids absorb in the UV themselves (Michaelian and Simeonov, 2015 and references therein), but they also serve as biosynthetic precursors for most known aromatic UV-absorbing bio-pigments, including: anthocyanins, flavonoids and phenylpropanoids in plants, melanins in heterotrophic organisms, scytonemins in cyanobacteria, etc. (Knaggs, 2003; Tang et al., 2024).
Sorrels et al. (2009) proposed an ancient evolutionary origin for the scytonemin biosynthetic pathway based on the combination of facts that this gene cluster is highly conserved among evolutionary diverse strains of cyanobacteria (Soule et al., 2007; 2009a) and their own phylogenetic analyses implying the cluster is under strong selection pressure. Intriguingly, Soule et al. (2009a) observed scytonemin biosynthetic genes even in some cyanobacterial strains incapable of producing the pigment (e.g., Anabaena and Nodularia), and interpreted this as a case of relic genetic information.
2.2.4. Function: Traditional Protective View vs. Thermodynamic View
Similarly to MAAs, the Darwinian point of view can only describe scytonemin as an efficient protective biomolecule designed to filter out supposedly damaging high frequency UV radiation while at the same time allowing the transmittance of wavelengths necessary for photosynthesis (Ekebergh et al., 2015; Ručová et al., 2023).
Within the framework of this traditional “struggle for survival” viewpoint, the majority of authors define scytonemins as an adaptive mechanism of extremophilic cyanobacteria that colonize harsh, inhospitable habitats experiencing high doses of UVR-insolation (Ehling-Schulz et al., 1997; Wynn-Williams et al., 1999; Hunsucker et al., 2001; Sinha and Häder, 2008; Rastogi et al., 2014; Ručová et al., 2023).
Among the evidence for the accredited photoprotective role is the discovery that up to 90% of incident UV photons are prevented from entering sheathed, scytonemin-producing cyanobacterial cells, thus accomplishing significant reduction in chlorophyll a photobleaching and maintaining photosynthetic efficiency (Garcia-Pichel and Castenholz, 1991; Garcia-Pichel et al., 1992; Gao et al., 2021). Other authors, in addition to the sunscreen role, ascribe supplementary defensive roles to scytonemin such as protection from oxidative, osmotic, heat and desiccation stress (Dillon et al., 2002; Matsui et al., 2012; Gao et al., 2021).
Additionlly, scytonemin’s superior UV-C-absorbing capabilities in vivo, experimentally proven by treating cyanobacterial colonies with 0.5-1.0 Wm-2 UV-C radiation added to natural solar irradiance (Dillon and Castenholz, 1999), has led many authors to consider modern cyanobacterial production of scytonemins as a relic UV-protection mechanism from the pre-Great Oxygenation Event (Garcia-Pichel, 1998; Häder et al., 2003; Tamre and Fournier, 2022). Indeed, Raman spectral biosignatures of scytonemins, carotenoids and porphyrins were unambiguously identified in ~ 3.5 Ga old fossilized sedimentary geological specimens (Edwards et al., 2007; Pullan et al., 2008). Extracellular pigments, presumably scytonemins, also seem to occur in ~ 2.0 Ga old cyanobacterial microfossils preserved in silicified stromatolites that grew in tidal or shallow subtidal waters (Hofmann, 1976; Golubic and Hofmann, 1976; Golubic and Abed, 2010; Demoulin et al., 2019).
Although it is beyond doubt that the efficient UV absorption and dissipation properties of the scytoneman pigments provide, to some extent, a beneficiary effect for the survival of sheathed cyanobacterial cells, the opinion that this is the primary reason for the biological production of these pigments may be erroneous. Here are few examples of serious challenges and inconsistencies that the photoprotection paradigm faces:
- Inability to explain the strong visible absorption bands of scytonemin-imine, where photosynthetic pigments absorb. The question is raised by Grant and Louda (2013):
“The absorption spectrum (max 237, 366, 437, 564 nm in vitro), extending from the ultraviolet (UVB & UVA) into the blue and green of the visible, appears to indicate a photoprotective role beyond shielding only UVR. That is, going on the premise that evolution generates and retains only advantageous secondary metabolites, then what is the role of the visible bands in this case?”
- 2.
- Inability to explain the production of the strongly UV-C/UV-B-absorbing methoxyscytonemins and scytonine, in spite of the absence of UV-C wavelengths and the low intensity of UV-B in today’s surface solar spectrum. The question is raised by Varnali and Edwards (2010):
“The realization that scytonemin is the parent molecule of perhaps a whole family of related molecules is important in that an analytical challenge is generated to detect these family members in admixture and in the presence of each other naturally, and also the question is raised about the role of these molecules in the survival strategy processes involving scytonemin; what subtle changes to the radiation absorption process require molecular modification of what apparently is already a highly successful radiation protectant, especially when the molecular syntheses are accomplished in energy-poor situations?”
- 3.
- Inability to explain why many species of cyanobacteria do not synthesize scytonemins nor MAAs but, nevertheless, successfully cope with UV-induced cellular damage by employing only metabolic repair mechanisms (Quesada and Vincent, 1997; Castenholz and Garcia-Pichel, 2000; Singh et al., 2023).
- 4.
- Soule et al. (2007) developed scytoneminless mutant of the cyanobacterium Nostoc punctiforme which proved to have indistinguishable growth rate from the wild type after both were subjected to UV-A irradiation. The conclusion of the authors was that other photoprotective mechanisms can fully accommodate the absence of scytonemin in the mutant.
Finally, very efficient absorption and dissipation of high-energy photons is not a prerequisite for photoprotection, but it is for dynamical dissipative structuring of material under an external generalized chemical potential. Nature has a simpler way of creating photoprotective molecules, if this was really nature’s intention: making them either highly reflective or transparent to UV wavelengths (Michaelian, 2016).
These problems and paradoxes arising when attempting to explain scytonemins from within the Darwinian photoprotection paradigm can be resolved by viewing the situation from established non-equilibrium thermodynamic principles. In this context, we first address the questions raised by Grant and Louda (2013) and Varnali and Edwards (2010), and then, based on all the evidence presented, we assign a thermodynamic dissipative role to scytonemins.
The seemingly paradoxical absorption spectra of scytonemin-imine, the methoxyscytonemins and scytonine, which extend outside of the photoprotectively-relevant part of the spectrum, make sense only when these bio-pigments are understood as microscopic dissipative structures obeying non-equlibrium thermodynamic directives related to increasing the global solar photon dissipation rate (Michaelian, 2011, 2013; Michaelian and Simeonov, 2015; Michaelian, 2016, 2017, 2021). Under these directives, one of the several ways to increase the global solar photon dissipation rate is by evolving (inventing) novel molecular structures (pigments) that cover ever more completely the prevailing surface solar spectrum (Michaelian and Simeonov, 2015).
This is precisely what is observed in the absorption spectra of the different scytoneman pigments. The strong visible absorption peaks of scytonemin-imine at 437 nm (violet) and 564 nm (green/yellow), of tetramethoxyscytonemin at 562 nm (green/yellow), of dimethoxyscytonemin at 422 nm (violet); and the strong near-UV-C/UV-B absorption peaks of scytonine (270 nm) and dimethoxyscytonemin (316 nm) is exactly where the photosynthetic pigments do not peak in absorption (see, for example, Rowan, 1989). It is because of this rich assortment of diverse pigment molecules with complementary absorption bands that cyanobacterial biofilms, mats and soil crusts in nature tend to have high absorptivity, low albedo and appear almost black in color (Ustin et al., 2009).
These facts lead us to an important conclusion on the thermodynamic function of the scytoneman pigments. We believe that it is most reasonable to consider the photon-dissipation role of scytonemins as the terrestrial analogue of the function that MAAs perform in the open aquatic environment. This assertion may be justified on their hydrophobic character and their inextricable connection to the extracellular polymeric substances (EPSs) of the cyanobacterial sheaths. Ekebergh et al. (2015) indicate that scytonemins have the greatest photostability in vivo, where they are embedded in their natural extracellular matrix milieu. These extracellular polymeric substances may therefore be playing the role of dissipators after photon excitation of the pigment, bringing the system rapidly to the ground state and ready to absorb another photon (Mahmud et al., 2024).
In wet terrestrial regions of the planet, the thermodynamic role of photon dissipation coupled to the water cycle is performed mainly by the plant cover, but in arid and semiarid lands, where vegetation is severely restricted, this function is allotted to microscopic assemblages of cyanobacteria, heterotrophic bacteria, algae and fungi known as biological soil crusts or biocrusts (Evans and Johansen, 1999; Belnap and Lange, 2001; Lan et al., 2024). It is theorized that these types of microbial communities were pioneers on dry land and were the dominant ecosystem on the continents up until the advent of land plants and animals in the Early Devonian (Beraldi-Campesi et al., 2014; Wellman et al., 2021) brought on by the microbial retainment of water on the continents and the concomitant extension of the water cycle to regions far inland from the coasts.
Michaelian (2013) postulated that bio-pigments are generally found in association with water because they use the high frequency vibrational modes of water molecules to facilitate their de-excitation. In this context, we emphasize the fact that cyanobacteria isolated from dry regions display very high capacity to excrete large amounts of EPSs (Huang et al., 1998; Hu et al., 2003; Roeselers et al., 2007; Rossi et al., 2012; Dabravolski and Isayenkov, 2022), which are the main constituent of the biofilm matrix and together with microbial filaments play a key structural role in forming the biocrusts (Mager and Thomas, 2010; Karunakaran et al., 2011). The unique hydrophilic/hydrophobic nature of the EPSs enables highly efficient water capture and water storage within the biocrust by allowing the creation of moistened microenvironments where water dynamics is intricately regulated (Colica et al., 2014 and references therein). Hence, crust-covered soils are very hygroscopic and always exhibit higher water content compared to bare neighboring surfaces (Rossi and Phillips, 2015). This phenomenon is in accordance with our postulate that life's fundamental role is that of dispersing organic pigments and water over the entire surface of the Earth (Michaelian, 2011; 2012; 2013; 2016; 2021; 2024).
A very conspicuous analogy between these terrestrial macroscopic and microscopic photon-dissipating biological “carpets” can be drawn. In the same manner as ecological succession of plant coverage leads to old climax forests with higher pigment content and lower albedos (Pokorny et al., 2010; Maes et al., 2011; Zhu et al., 2016), ecological succession in biocrusts leads to increase in biomass of the late-stage scytonemin-producing cyanobacteria, and consequently accumulation of scytonemins in the matrix, an effect macroscopically observed as darkening of the biocrusted soil (i.e. decrease in albedo) (Couradeau et al., 2016).
During dry periods in deserts when water availability is very limited, the heat generated from scytonemin’s photon dissipation is expected to go predominantly into sensible heat of the biocrusts instead of into the latent heat of vaporization of water, and this is exactly what Couradeau et al. (2016) found when they measured ~ 10°C higher temperature of biocrust-covered, dark soils in comparison to bare soils. Dark soils absorb and dissipate photons, producing more entropy than bare soils which predominantly reflect the incident light.
3. Discussion
In a previous work (Michaelian and Simeonov, 2015) we posited five basic tendencies that organic pigment evolution on Earth would have followed: (1) increases in the photon absorption cross section with respect to the pigment physical size, (2) decreases in the electronic excited state lifetimes of the pigments, (3) quenching of the radiative de-excitation channels (e.g., fluorescence), (4) greater coverage of the surface solar spectrum, and (5) pigment proliferation and dispersion over an ever greater surface area of Earth.
To examine whether these five tendencies are satisfied with the evolutionary invention of MAAs and scytonemins we compare their properties to those of the aromatic amino acids (AAAs) (see Table 1) which are also fundamental pigments (Mejía and Michaelian, 2020).
AAAs are most likely the precursors of MAAs since; (1) they are considered to be among the earliest chromophoric organic molecules used by life with a prebiotic origin (Michaelian and Simeonov, 2015), and (2) since both MAAs and scytonemins are derived from intermediates of the shikimate pathway for AAA biosynthesis, they most likely appeared later in evolution compared to the AAAs, probably when the biosynthetic machinery for the synthesis of the AAAs was already robust; an event that most likely long predated 3.4 Ga, considering that Busch et al. (2016) demonstrated that the ancestral tryptophan synthase of the last universal common ancestor (LUCA) was already a highly sophisticated enzyme at 3.4 Ga.
Based on all the data presented in previous sections on the spectral coverage and biological and geographical ubiquity of these pigments, we can state with a high degree of certainty that the fourth and fifth requirements are satisfied with the evolutionary inventions of MAAs and scytonemins.
With respect to the first to third strategies for augmenting photon dissipation, in addition to the previously discussed material, we offer the data presented in Table 1 which has been collected from the available literature up to 2025. All compounds listed are representative members of their respective chemical groups. Gadusol is used instead of the more relevant compound 4-deoxygadusol because of lack of available data on 4-deoxygadusol and because of their chemical relatedness with similar spectroscopic properties (Losantos et al., 2015a, 2015b). The and values of gadusol in water are pH-dependent: 268 nm at pH < 7 and 296 nm at pH ≥ 7 (Losantos et al., 2015a), and in Table 1 we use the values for acidic pH since the Archean seawater was probably slightly acidic with pH ~ 6.5 (Holland, 2003; Krissansen-Totton et al., 2018).
All absorption peaks and attenuation coefficients below about 205 nm, we believe, are due to the ionization of the molecules, a process which could destroy them. Photon dissipation is not through a conical intersection at these very short wavelengths, and this is why they are omitted from Table 1 and Figure 3.
It is our hope that future experiments and studies into the nature and properties of these bio-pigments will help complete the data missing from the table. However, even with the limited data available and presented in this article, a trend compatible with our conjecture of increasing photon dissipation is evident.
Another conjecture made in Michaelian and Simeonov (2015) states that the surface solar spectrum wavelength region from approximately 285 to 310 nm has never reached the surface of the Earth during its entire geologic history; because during the Hadean and Archean eons these wavelengths were probably absorbed by atmospheric aldehydes (Sagan, 1973), and from the end of the Archean onwards gradual accumulation of oxygen and stratospheric ozone was responsible for their attenuation (Matsumi and Kawasaki, 2003; Stanley, 2008). In this earlier paper we also demonstrated how numerous fundamental molecules of life, common to all three domains of life, have strong absorbances across the UV-C and UV-A regions but little in the UV-B region (280-315 nm), consistent with the thermodynamic dissipation theory of the origin and evolution of life (Michaelian, 2009; 2011; Michaelian, 2012).
The same appears to apply to the cyanobacterial UV-absorbing pigments scytonemins and MAAs, which can be considered as evolutionary successors to the primordial pigments of life, specifically the AAAs. Indeed, the absorption spectrum of scytonemin (Figure 4) seems to adhere to this pattern. Although it is continuous from ~ 220 to ~ 700 nm, there is a dip in the ~ 275 to ~ 325 nm interval, and two large maxima at ~ 250 nm and ~ 380 nm. This is exactly the kind of shape that would be expected if the selective force for the evolution of this pigment was the Archean surface solar spectrum (Michaelian and Simeonov, 2015).
Combining these crucial facts on scytonemin, it is tempting to speculate that this pigment had a key role in photon dissipation during the Archean, being capable of dissipating almost the entire Archean surface solar spectrum. The evolutionary invention of scytonemin’s derivatives, as well as the mycosporines, the MAAs and still many other extinct and extant biological pigments, most likely resulted from the necessity to complement scytonemin’s absorption with pigments that absorbed wavelengths reaching Earth’s surface but were poorly absorbed by scytonemin itself. This kind of spectral complementary relationship between scytonemin and MAAs has been well documented by several authors (e.g., Ehling-Schulz and Scherer, 1999; Ferroni et al., 2010; Castenholz and Garcia-Pichel, 2012) and is illustrated in Figure 5.
Figure 3.
The spectrum of UV light available at Earth’s surface before the origin of life at approximately 3.9 Ga and until at least 2.9 Ga (curves black and red respectively). The spectrum in the UV-C may even have persisted throughout the entire Archean and even until 2.5 Ga. Atmospheric CO2, H2O, SO2 and probably some H2S, were responsible for absorption of wavelengths shorter than ∼205 nm, and atmospheric gas aldehydes (e.g., formaldehyde and acetaldehyde) absorbed between about 280 and 310 nm, approximately corresponding to the UV-B region. By around 2.2 Ga (green curve), UV-C light at Earth’s surface was completely extinguished by the pigments oxygen and ozone resulting from organisms performing oxygenic photosynthesis. The yellow curve corresponds to the present surface spectrum. The precursors have strong absorptions only in the UV-C (205-280 nm). We propose that the precursor molecules (also pigments) were dissipatively structured under this UV-C light. Scytonemins absorb strongly across the UV-C, UV-B, UV-A, violet and blue regions, and mycosporines and MAAs have strong absorption in the UV-B and UV-A regions. Neither scytonemins nor MAAs peak strongly in the UV-B region from ~ 280-310 nm because aldehydes produced by UV-C light on common volcanic gases such as H2S, H2O and CO2, were absorbing strongly in this gap. Energy fluxes are for the sun at the zenith. The font size is roughly proportional to the relative size of the respective molar extinction coefficient of the pigment.
Figure 3.
The spectrum of UV light available at Earth’s surface before the origin of life at approximately 3.9 Ga and until at least 2.9 Ga (curves black and red respectively). The spectrum in the UV-C may even have persisted throughout the entire Archean and even until 2.5 Ga. Atmospheric CO2, H2O, SO2 and probably some H2S, were responsible for absorption of wavelengths shorter than ∼205 nm, and atmospheric gas aldehydes (e.g., formaldehyde and acetaldehyde) absorbed between about 280 and 310 nm, approximately corresponding to the UV-B region. By around 2.2 Ga (green curve), UV-C light at Earth’s surface was completely extinguished by the pigments oxygen and ozone resulting from organisms performing oxygenic photosynthesis. The yellow curve corresponds to the present surface spectrum. The precursors have strong absorptions only in the UV-C (205-280 nm). We propose that the precursor molecules (also pigments) were dissipatively structured under this UV-C light. Scytonemins absorb strongly across the UV-C, UV-B, UV-A, violet and blue regions, and mycosporines and MAAs have strong absorption in the UV-B and UV-A regions. Neither scytonemins nor MAAs peak strongly in the UV-B region from ~ 280-310 nm because aldehydes produced by UV-C light on common volcanic gases such as H2S, H2O and CO2, were absorbing strongly in this gap. Energy fluxes are for the sun at the zenith. The font size is roughly proportional to the relative size of the respective molar extinction coefficient of the pigment.
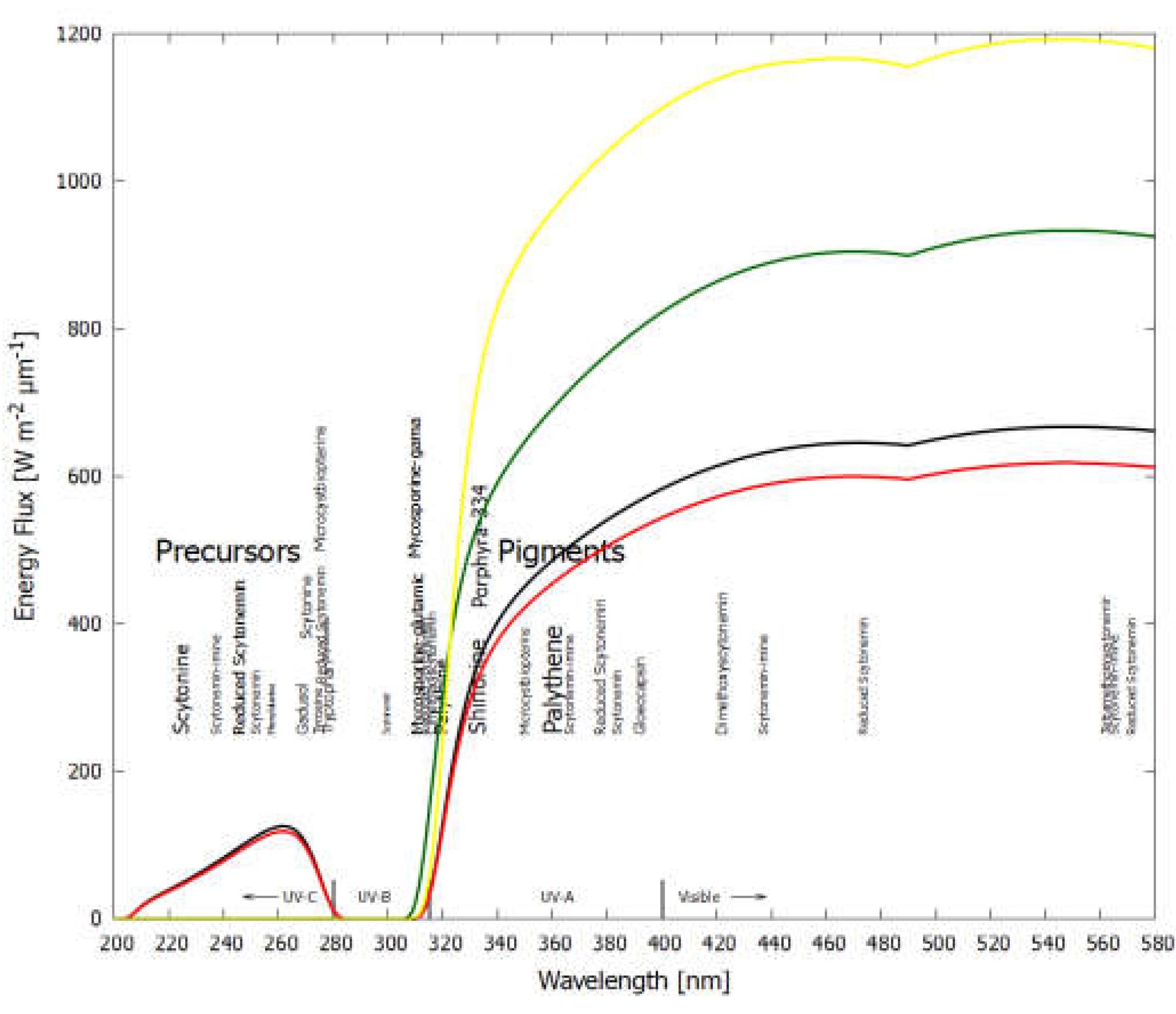
Figure 4.
In vitro absorption spectrum of scytonemin. (Adapted from Sinha et al., 1999).

Figure 5.
Complementary absorption spectra of scytonemin and MAAs. (Adapted from Rastogi and Madamwar, 2016).
Figure 5.
Complementary absorption spectra of scytonemin and MAAs. (Adapted from Rastogi and Madamwar, 2016).
4. Conclusions
The available data on the ubiquity of pigments covering the region from the UV-C to the infrared, many exuded into the environment by the organisms that produce them, make it questionable to assign to them a protective or antenna role within the Darwinian paradigm of the optimization of photosynthesis in benefit of the organism. We believe that sense can only be made of this by shifting the paradigm from one of “photoprotection” of the organism to the thermodynamic optimization of photon dissipation.
A number of contemporary pigment lines, most notably scytonemins and the mycosporine-like amino acids (MAAs), appear to harbor relics of ancient biosynthetic production routes based on the most ancient of the amino acids, the aromatics. The aromatic amino acids have known affinities to their RNA anticodons (Majerfeld and Yarus, 2005; Yarus et al., 2009) and were perhaps the first antenna pigments for photon dissipation in the UV-C at the beginnings of life (Michaelian, 2011; Mejía and Michaelian, 2020).
These pigment lines absorb and dissipate efficiently in the UV-C and UV-A, as well as somewhat in the UV-B and the visible. Some of these pigments are exuded into the environment which excludes the possibility of assigning them a role in photoprotection. Their strong absorption and dissipation in regions out of the photosynthetically active radiation (PAR) has been perplexing to perspectives within the Darwinian paradigm since these pigments appear to have little utility to the organisms themselves. In fact, they absorb exactly where the photosynthetic pigments do not (and where water does not). They appear to have completely covered the Archean surface solar spectrum and do the same today.
It should be emphasized that our current knowledge of the diversity of cyanobacterial, algal and plant pigments and the thermodynamic function they perform is incomplete. For example, there are several indications of even richer diversity of UV-absorbing pigments in cyanobacteria than those hitherto characterized and classified into the two groups, mycosporines and scytonemins. The chemical structure and other elemental properties of one of these poorly investigated pigments, named gloeocapsin, have yet to be determined, but initial results suggest that it is chemically unrelated to both MAAs and scytonemins (Storme et al., 2015; Lara et al., 2022). Still other chemically distinct UV-absorbing cyanobacterial pigments, with a unique pterin structure, have been reported elsewhere (Matsunaga et al., 1993; Lifshits et al., 2016). The wavelengths of maximum absorption of these two ill-defined groups of cyanobacterial pigments are listed in Table 1 and are plotted in Figure 3. As with the mycosporines and the scytonemins, their absorption properties are consistent with the optimization of dissipation of the prevailing photon spectrum at Earth’s surface.
Taken as a whole, these data seem to indicate that, rather than photosynthesis being optimized under a Darwinian “survival of the fittest” paradigm, that the origin and evolution of life is driven by photon dissipation with the net effect of covering Earth’s entire surface with pigments and water, reducing the albedo and the black-body temperature at which Earth radiates into space, thereby increasing the entropy production of the biosphere. It is our hope that this article will incite further investigation into the proposition that photon dissipation efficacy has been the fundamental driver of biological evolution on Earth.
References
- Abed, R.M.; Al Kharusi, S.; Schramm, A.; Robinson, M.D. Bacterial diversity, pigments and nitrogen fixation of biological desert crusts from the Sultanate of Oman. FEMS Microbiol. Ecol. 2010, 72, 418–428. [CrossRef]
- Arpin, N., Curt, R. and Favre-Bonvin, J.: Mycosporines: mise au point et donées nouvelles concernant leurs structures, leur distribution, leur localisation et leur biogenèse, Rev Mycol, 43,.
- 247–257, 1979.
- Arsın, S.; Delbaje, E.; Jokela, J.; Wahlsten, M.; Farrar, Z.M.; Permi, P.; Fewer, D. A Plastic Biosynthetic Pathway for the Production of Structurally Distinct Microbial Sunscreens. ACS Chem. Biol. 2023, 18, 1959–1967. [CrossRef]
- Awramik, S.; Schopf, J.; Walter, M. Filamentous fossil bacteria from the Archean of Western Australia. 2003, 20, 357–374. [CrossRef]
- Babin, M., Stramski, D., Ferrari, G. M., Claustre, H., Bricaud, A., Obolensky, G., Hoepffner, N.: Variations in the light absorption coefficients of phyto-plankton, nonalgal particles, and dissolved organic matter in coastal waters around Europe, J. Geophys. Res, 108(C7), 3211, 2003.
- Balskus, E.P.; Walsh, C.T. Investigating the Initial Steps in the Biosynthesis of Cyanobacterial Sunscreen Scytonemin. J. Am. Chem. Soc. 2008, 130, 15260–15261. [CrossRef]
- Balskus, E.P.; Walsh, C.T. An Enzymatic Cyclopentyl[b]indole Formation Involved in Scytonemin Biosynthesis. J. Am. Chem. Soc. 2009, 131, 14648–14649. [CrossRef]
- Balskus, E.P.; Walsh, C.T. The Genetic and Molecular Basis for Sunscreen Biosynthesis in Cyanobacteria. Science 2010, 329, 1653–1656. [CrossRef]
- Balskus, E.P.; Case, R.J.; Walsh, C.T. The biosynthesis of cyanobacterial sunscreen scytonemin in intertidal microbial mat communities. FEMS Microbiol. Ecol. 2011, 77, 322–332. [CrossRef]
- Bandaranayake, W. M.: Mycosporines: are they nature’s sunscreens?, Natural Product Reports, 1998, 159–171, 1998.
- Belnap, J. and Lange, O. L.: Biological Soil Crusts: Structure, Function, and Management,.
- Springer Verlag, Berlin, Germany, 2001.
- Beraldi-Campesi, H.; Farmer, J.D.; Garcia-Pichel, F. MODERN TERRESTRIAL SEDIMENTARY BIOSTRUCTURES AND THEIR FOSSIL ANALOGS IN MESOPROTEROZOIC SUBAERIAL DEPOSITS. PALAIOS 2014, 29, 45–54. [CrossRef]
- Berezin, M.Y.; Achilefu, S. Fluorescence Lifetime Measurements and Biological Imaging. Chem. Rev. 2010, 110, 2641–2684. [CrossRef]
- Brenowitz, S. and Castenholz, R. W.: Long-term effects of UV and visible irradiance on natural populations of a scytonemin-containing cyanobacterium (Calothrix sp.), FEMS Microbiol Ecol, 24, 343–352, 1997.
- Bricaud, A.; Babin, M.; Claustre, H.; Ras, J.; Tièche, F. Light absorption properties and absorption budget of Southeast Pacific waters. J. Geophys. Res. Oceans 2010, 115. [CrossRef]
- Büdel, B., Karsten, U. and Garcia-Pichel, F.: Ultraviolet absorbing scytonemin and mycosporine-like amino acid derivatives in exposed, rock-inhabiting cyanobacterial lichens, Oecologia, 112, 165–172, 1997.
- Bultel-Poncé, V., Felix-Theodose, F., Sarlhou, C., Ponge, J. F. and Bodo, B.: New pigments from the terrestrial cyanobacterium Scytonema sp. collected on the Mitakara Inselberg, French Guyana, J Nat Prod, 67, 678–681, 2004.
- Busch, F.; Rajendran, C.; Heyn, K.; Schlee, S.; Merkl, R.; Sterner, R. Ancestral Tryptophan Synthase Reveals Functional Sophistication of Primordial Enzyme Complexes. Cell Chem. Biol. 2016, 23, 709–715. [CrossRef]
- Carlson, D.J.; Mayer, L.M. Enrichment of dissolved phenolic material in the surface microlayer of coastal waters. Nature 1980, 286, 482–483. [CrossRef]
- Carreto, J. I. and Carignan, M. O.: Mycosporine-like amino acids: relevant secondary metabolites. Chemical and ecological aspects, Mar Drugs, 9, 387−446, 2011.
- Carreto, J.; Carignan, M.; Daleo, G.; Marco, S. Occurrence of mycosporine-like amino acids in the red-tide dinoflagellate Alexandrium excavatum: UV-photoprotective compounds?. J. Plankton Res. 1990, 12, 909–921. [CrossRef]
- Carreto, J.I.; Carignan, M.O.; Montoya, N.G. A high-resolution reverse-phase liquid chromatography method for the analysis of mycosporine-like amino acids (MAAs) in marine organisms. Mar. Biol. 2004, 146, 237–252. [CrossRef]
- Carreto, J. I., Roy, S., Whitehead, K., Llewellyn, C. A. and Carignan, M. O.: UV-absorbing ‘pigments’: mycosporine-like amino acids. In: Roy, S., Llewellyn, C. A., Egeland, E. S. and Johnsen, G. (eds.) Phytoplankton pigments: characterisation, chemotaxonomy and applications in oceanography, Cambridge University Press, Cambridge, 412−441, 2011.
- Carroll, A.K.; Shick, J.M. Dietary accumulation of UV-absorbing mycosporine-like amino acids (MAAs) by the green sea urchin (Strongylocentrotus droebachiensis). Mar. Biol. 1996, 124, 561–569. [CrossRef]
- Castenholz, R. W. and Garcia-Pichel, F.: Cyanobacterial responses to UV-radiation. In: Whitton, B. A. and Potts, M. (eds.) The ecology of cyanobacteria. Their diversity in time and space, Kluwer Academic Publishers, Dordrecht, pp 591–611, 669, 2000.
- Castenholz, R. W. and Garcia-Pichel, F.: Cyanobacterial responses to UV radiation, In: Whitton, B. A. (ed.) Ecology of Cyanobacteria II: Their Diversity in Space and Time, Springer Netherlands, 481-499, 2012. [CrossRef]
- Clementson, L.A.; Oubelkheir, K.; Ford, P.W.; Blondeau-Patissier, D. Distinct Peaks of UV-Absorbing Compounds in CDOM and Particulate Absorption Spectra of Near-Surface Great Barrier Reef Coastal Waters, Associated with the Presence of Trichodesmium spp. (NE Australia). Remote. Sens. 2022, 14, 3686. [CrossRef]
- Cockell, C. S. and Knowland, J.: Ultraviolet radiation screening compounds, Biol. Rev., 74, 311–345, 1999.
- Cohen, G. N.: Microbial Biochemistry, (3rd edition), Springer, Netherlands, pages: 85-90 and 415-440, 2014.
- Colica, G.; Li, H.; Rossi, F.; Li, D.; Liu, Y.; De Philippis, R. Microbial secreted exopolysaccharides affect the hydrological behavior of induced biological soil crusts in desert sandy soils. Soil Biol. Biochem. 2014, 68, 62–70. [CrossRef]
- Conde, F. R., Churio, M. S. and Previtali, C. M.: The photoprotector mechanism of mycosporine-like amino acids. Excited state properties and photostability of porphyra-334 in aqueous solution, J. Photochem. Photobiol. B, 56, 139–144, 2000.
- Conde, F.R.; Churio, M.S.; Previtali, C.M. The deactivation pathways of the excited-states of the mycosporine-like amino acids shinorine and porphyra-334 in aqueous solution. Photochem. Photobiol. Sci. 2004, 3, 960–967. [CrossRef]
- Conde, F.R.; Churio, M.S.; Previtali, C.M. Experimental study of the excited-state properties and photostability of the mycosporine-like amino acid palythine in aqueous solution. Photochem. Photobiol. Sci. 2007, 6, 669–674. [CrossRef]
- Couradeau, E.; Karaoz, U.; Lim, H.C.; da Rocha, U.N.; Northen, T.; Brodie, E.; Garcia-Pichel, F. Bacteria increase arid-land soil surface temperature through the production of sunscreens. Nat. Commun. 2016, 7, 10373–10373. [CrossRef]
- D'Agostino, P.M.; Javalkote, V.S.; Mazmouz, R.; Pickford, R.; Puranik, P.R.; Neilan, B.A. Comparative Profiling and Discovery of Novel Glycosylated Mycosporine-Like Amino Acids in Two Strains of the Cyanobacterium Scytonema cf. crispum. Appl. Environ. Microbiol. 2016, 82, 5951–5959. [CrossRef]
- Dabravolski, S.A.; Isayenkov, S.V. Metabolites Facilitating Adaptation of Desert Cyanobacteria to Extremely Arid Environments. Plants 2022, 11, 3225. [CrossRef]
- Demmig-Adams, B.; Adams, W.W., III. Photoprotection and Other Responses of Plants to High Light Stress. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1992, 43, 599–626. [CrossRef]
- Demoulin, C.F.; Lara, Y.J.; Cornet, L.; François, C.; Baurain, D.; Wilmotte, A.; Javaux, E.J. Cyanobacteria evolution: Insight from the fossil record. Free. Radic. Biol. Med. 2019, 140, 206–223. [CrossRef]
- Dillon, J.G.; Castenholz, R.W. SCYTONEMIN, A CYANOBACTERIAL SHEATH PIGMENT, PROTECTS AGAINST UVC RADIATION: IMPLICATIONS FOR EARLY PHOTOSYNTHETIC LIFE. J. Phycol. 1999, 35, 673–681. [CrossRef]
- Dillon, J. G., Tatsumi, C. M., Tandingan, P. G. and Castenholz, R. W.: Effect of environmental factors on the synthesis of scytonemin, a UV-screening pigment, in a cyanobacterium (Chroococcidiopsis sp.), Arch. Microbiol., 177, 322–331, 2002.
- Dunlap, W.C.; Chalker, B.E. Identification and quantitation of near-UV absorbing compounds (S-320) in a hermatypic scleractinian. Coral Reefs 1986, 5, 155–159. [CrossRef]
- Dunlap, W. C. and Shick, J. M.: Ultraviolet radiation absorbing mycosporine-like amino acids in coral reef organisms: a biochemical and environmental environmental perspective, J Phycol, 34, 418–430, 1998.
- Edwards, H.G.M.; Villar, S.E.J.; Pullan, D.; Hargreaves, M.D.; Hofmann, B.A.; Westall, F. Morphological biosignatures from relict fossilised sedimentary geological specimens: a Raman spectroscopic study. J. Raman Spectrosc. 2007, 38, 1352–1361. [CrossRef]
- Ehling-Schulz, M. and Scherer, S.: UV protection in cyanobacteria, Eur J Phycol, 34, 329–338, 1999.
- Ehling-Schulz, M.; Bilger, W.; Scherer, S. UV-B-induced synthesis of photoprotective pigments and extracellular polysaccharides in the terrestrial cyanobacterium Nostoc commune. J. Bacteriol. 1997, 179, 1940–1945. [CrossRef]
- Ekebergh, A.; Sandin, P.; Mårtensson, J. On the photostability of scytonemin, analogues thereof and their monomeric counterparts. Photochem. Photobiol. Sci. 2015, 14, 2179–2186. [CrossRef]
- Evans, R.D.; Johansen, J.R. Microbiotic Crusts and Ecosystem Processes. Crit. Rev. Plant Sci. 1999, 18, 183–225. [CrossRef]
- Favre-Bonvin, J.; Arpin, N.; Brevard, C. Structure de la mycosporine (P 310). Can. J. Chem. 1976, 54, 1105–1113. [CrossRef]
- Favre-Bonvin, J.; Bernillon, J.; Salin, N.; Arpin, N. Biosynthesis of mycosporines: Mycosporine glutaminol in Trichothecium roseum. Phytochemistry 1987, 26, 2509–2514. [CrossRef]
- Ferreira, D.; Garcia-Pichel, F. Mutational Studies of Putative Biosynthetic Genes for the Cyanobacterial Sunscreen Scytonemin in Nostoc punctiforme ATCC 29133. Front. Microbiol. 2016, 7, 735. [CrossRef]
- Ferroni, L.; Klisch, M.; Pancaldi, S.; Häder, D.-P. Complementary UV-Absorption of Mycosporine-like Amino Acids and Scytonemin is Responsible for the UV-Insensitivity of Photosynthesis in Nostoc flagelliforme. Mar. Drugs 2010, 8, 106–121. [CrossRef]
- Fleming, E. D. and Castenholz, R. W.: Effects of periodic desiccation on the synthesis of the UV-screening compound, scytonemin, in cyanobacteria, Environ Microbiol, 9, 1448–1455, 2007.
- Fuentes-Tristan, S.; Parra-Saldivar, R.; Iqbal, H.M.; Carrillo-Nieves, D. Bioinspired biomolecules: Mycosporine-like amino acids and scytonemin from Lyngbya sp. with UV-protection potentialities. J. Photochem. Photobiol. B: Biol. 2019, 201, 111684. [CrossRef]
- Fulton, J.M.; Arthur, M.A.; Freeman, K.H. Subboreal aridity and scytonemin in the Holocene Black Sea. Org. Geochem. 2012, 49, 47–55. [CrossRef]
- Fukuzaki, K.; Imai, I.; Fukushima, K.; Ishii, K.-I.; Sawayama, S.; Yoshioka, T. Fluorescent characteristics of dissolved organic matter produced by bloom-forming coastal phytoplankton. J. Plankton Res. 2014, 36, 685–694. [CrossRef]
- Galgani, L. and Engel, A.: Changes in optical characteristics of surface microlayers hint to photochemically and microbially mediated DOM turnover in the upwelling region off the coast of Peru, Biogeosciences, 13, 2453–2473, 2016.
- Gao, Q.; Garcia-Pichel, F. Microbial ultraviolet sunscreens. Nat. Rev. Microbiol. 2011, 9, 791–802. [CrossRef]
- Gao, X.; Jing, X.; Liu, X.; Lindblad, P. Biotechnological Production of the Sunscreen Pigment Scytonemin in Cyanobacteria: Progress and Strategy. Mar. Drugs 2021, 19, 129. [CrossRef]
- Garcia-Pichel, F. Solar Ultraviolet and the Evolutionary History of Cyanobacteria. Discov. Life 1998, 28, 321–347. [CrossRef]
- Garcia-Pichel, F. and Castenholz, R. W.: Characterization and biological implications of scytonemin, a cyanobacterial sheath pigment, J Phycol, 27, 395–409, 1991.
- Garcia-Pichel, F.; Castenholz, R.W. Occurrence of UV-Absorbing, Mycosporine-Like Compounds among Cyanobacterial Isolates and an Estimate of Their Screening Capacity. Appl. Environ. Microbiol. 1993, 59, 163–169. [CrossRef]
- Garcia-Pichel, F.; Sherry, N.D.; Castenholz, R.W. EVIDENCE FOR AN ULTRAVIOLET SUNSCREEN ROLE OF THE EXTRACELLULAR PIGMENT SCYTONEMIN IN THE TERRESTRIAL CYANOBACTERIUM Chiorogloeopsis sp.. Photochem. Photobiol. 1992, 56, 17–23. [CrossRef]
- Geraldes, V.; Pinto, E. Mycosporine-Like Amino Acids (MAAs): Biology, Chemistry and Identification Features. Pharmaceuticals 2021, 14, 63. [CrossRef]
- Glansdorff, P. and Prigogine, I.: Thermodynamic Theory of Structure, Stability, and Fluctuations, Wiley-Interscience, London, 1971.
- Gnanadesikan, A.; Emanuel, K.; Vecchi, G.A.; Anderson, W.G.; Hallberg, R. How ocean color can steer Pacific tropical cyclones. Geophys. Res. Lett. 2010, 37. [CrossRef]
- Golubic, S. and Abed, R. M. M.: Entophysalis mats as environmental regulators, In: Seckbach, J. and Oren, A. (eds.) Microbial Mats, Modern and Ancient Microorganisms in Stratified Systems, Springer, Heidelberg, pp 237–251, 2010.
- Golubic, S. and Hofmann, H. J.: Comparison of Holocene and mid-Precambrian Entophysalidaceae (Cyanophyta) in stromatolitic algal mats: cell division and degradation, J.
- Paleontol., 50, 1074–1082, 1976.
- Grant, C.S.; Louda, J. Scytonemin-imine, a mahogany-colored UV/Vis sunscreen of cyanobacteria exposed to intense solar radiation. Org. Geochem. 2013, 65, 29–36. [CrossRef]
- Gupta, S.; Guttman, M.; Leverenz, R.L.; Zhumadilova, K.; Pawlowski, E.G.; Petzold, C.J.; Lee, K.K.; Ralston, C.Y.; Kerfeld, C.A. Local and global structural drivers for the photoactivation of the orange carotenoid protein. Proc. Natl. Acad. Sci. 2015, 112, E5567–E5574. [CrossRef]
- Hatakeyama, M.; Nakamura, S. “Button-on-a-String” Mechanism in Water, the Ultrafast UV-to-Heat Conversion by Mycosporine-like Amino Acid Porphyra-334 of Natural Sunscreen Compound. ACS Phys. Chem. Au 2025. [CrossRef]
- Häder, D.-P.; Kumar, H.D.; Smith, R.C.; Worrest, R.C. Aquatic ecosystems: effects of solar ultraviolet radiation and interactions with other climatic change factors. Photochem. Photobiol. Sci. 2003, 2, 39–50. [CrossRef]
- Hickman-Lewis, K.; Cavalazzi, B.; Giannoukos, K.; D'Amico, L.; Vrbaski, S.; Saccomano, G.; Dreossi, D.; Tromba, G.; Foucher, F.; Brownscombe, W.; et al. Advanced two- and three-dimensional insights into Earth's oldest stromatolites (ca. 3.5 Ga): Prospects for the search for life on Mars. Geology 2022, 51, 33–38. [CrossRef]
- Hirakawa, Y.; Kakegawa, T.; Furukawa, Y. Hexose phosphorylation for a non-enzymatic glycolysis and pentose phosphate pathway on early Earth. Sci. Rep. 2024, 14, 1–10. [CrossRef]
- Hofmann, H. J.: Precambrian microflora, Belcher Islands, Canada: significance and systematic, J Paleontol., 50, 1040–1073, 1976.
- Holland, H. D.: The Geologic History of Seawater, In: Holland, H. D. and Turekian, K. K. (eds.), Treatise on Geochemistry, Volume 6; (ISBN: 0-08-044341-9); pp. 583–625, Elsevier Science, 2003.
- Horton, P.; Ruban, A.V.; Walters, R.G. REGULATION OF LIGHT HARVESTING IN GREEN PLANTS. Annu. Rev. Plant Biol. 1996, 47, 655–684. [CrossRef]
- Hu, C.; Zhang, D.; Huang, Z.; Liu, Y. The vertical microdistribution of cyanobacteria and green algae within desert crusts and the development of the algal crusts. Plant Soil 2003, 257, 97–111. [CrossRef]
- Huang, Z., Liu, Y., Paulsen, B. S. and Klaveness, D.: Studies on Polysaccharides from Three Edible Species of Nostoc (Cyanobacteria) with Different Colony Morphologies: Comparison of.
- 82. Monosaccharide Compositions and Viscosities of Polysaccharides from Field Colonies and.
- Suspension Cultures, J. Phycol., 34, 962–968, 1998.
- Hunsucker, S.W.; Tissue, B.M.; Potts, M.; Helm, R.F. Screening Protocol for the Ultraviolet-Photoprotective Pigment Scytonemin. Anal. Biochem. 2001, 288, 227–230. [CrossRef]
- Ingalls, A.E.; Whitehead, K.; Bridoux, M.C. Tinted windows: The presence of the UV absorbing compounds called mycosporine-like amino acids embedded in the frustules of marine diatoms. Geochim. et Cosmochim. Acta 2010, 74, 104–115. [CrossRef]
- Ito, S. and Hirata, Y.: Isolation and structure of a mycosporine from the zoanthidian Palythoa tuberculosa, Tetrahedron Lett., 28, 2429–2430, 1977.
- Jain, S.; Prajapat, G.; Abrar, M.; Ledwani, L.; Singh, A.; Agrawal, A. Cyanobacteria as efficient producers of mycosporine-like amino acids. J. Basic Microbiol. 2017, 57, 715–727. [CrossRef]
- Jones, I.; George, G.; Reynolds, C. Quantifying effects of phytoplankton on the heat budgets of two large limnetic enclosures. Freshw. Biol. 2005, 50, 1239–1247. [CrossRef]
- Kahru, M.; Leppanen, J.-M.; Rud, O. Cyanobacterial blooms cause heating of the sea surface. Mar. Ecol. Prog. Ser. 1993, 101, 1–7. [CrossRef]
- Karentz, D.: Ultraviolet tolerance mechanisms in Antarctic marine organisms, Antarctic Research Series, 62, 93-110, 1994.
- Karentz, D.: Chemical defenses of marine organisms against solar radiation exposure: UV absorbing mycosporine-like amino acids and scytonemin. In: McClintock, J. B. and Baker, B. J. (eds.) Marine chemical ecology, CRC Press, Boca Raton, pp 481-519, 2001.
- Karentz, D.; Spero, H.J. Response of a natural Phaeocysris population to ambient fluctuations of UVB radiation caused by Antarctic ozone depletion. J. Plankton Res. 1995, 17, 1771–1789. [CrossRef]
- Karsten, U.: Defense strategies of algae and cyanobacteria against solar ultraviolet radiation. In: Amsler, C. D. (ed.), Algal Chemical Ecology, Springer-Verlag, Berlin Heidelberg, 2008.
- Karsten, U.; Dummermuth, A.; Hoyer, K.; Wiencke, C. Interactive effects of ultraviolet radiation and salinity on the ecophysiology of two Arctic red algae from shallow waters. Polar Biol. 2003, 26, 249–258. [CrossRef]
- Karsten, U., Lembcke, S. and Schumann, R: The effect of ultraviolet radiation on photosynthetic performance, growth and sunscreen compound in aeroterrestrial biofilm algae isolated from building facades, Planta, 225, 991–1000, 2007.
- Karsten, U.; Maier, J.; Garcia-Pichel, F. Seasonality in UV-absorbing compounds of cyanobacterial mat communities from an intertidal mangrove flat. Aquat. Microb. Ecol. 1998, 16, 37–44. [CrossRef]
- Karunakaran, E., Mukherjee, J., Ramalingam, B. and Biggs, C. A.: “Biofilmology”: a multidisciplinary review of the study of microbial biofilms, Appl. Microbiol. Biotechnol., 90, 1869-1881, 2011.
- Kehr, J.-C.; Dittmann, E. Biosynthesis and Function of Extracellular Glycans in Cyanobacteria. Life 2015, 5, 164–180. [CrossRef]
- A Keller, M.; Turchyn, A.V.; Ralser, M. Non-enzymatic glycolysis and pentose phosphate pathway-like reactions in a plausible Archean ocean. Mol. Syst. Biol. 2014, 10, 725. [CrossRef]
- Kinzie, R.A. Effects of ambient levels of solar ultraviolet radiation on zooxanthellae and photosynthesis of the reef coral Montipora verrucosa. Mar. Biol. 1993, 116, 319–327. [CrossRef]
- Knaggs, A.R. The biosynthesis of shikimate metabolites. Nat. Prod. Rep. 2002, 20, 119–136. [CrossRef]
- Kobayashi, A.; Kajiyama, S.-I.; Inawaka, K.; Kanzaki, H.; Kawazu, K. Nostodione A, a Novel Mitotic Spindle Poison from a Blue-Green Alga Nostoc commune. Z. Fur Naturforschung Sect. C-A J. Biosci. 1994, 49, 464–470. [CrossRef]
- Korbee, N., Figueroa, F. L. and Aguilera, J.: Accumulation of mycosporine-like amino acids (MAAs): biosynthesis, photocontrol and ecophysiological functions, Revista Chilena de Historia Natural, 79, 119–132, 2006.
- Krissansen-Totton, J.; Arney, G.N.; Catling, D.C. Constraining the climate and ocean pH of the early Earth with a geological carbon cycle model. Proc. Natl. Acad. Sci. 2018, 115, 4105–4110. [CrossRef]
- Lan, S.; Elliott, D.R.; Chamizo, S.; Felde, V.J.M.N.L.; Thomas, A.D.; Lan, \. Editorial: Biological soil crusts: spatio-temporal development and ecological functions of soil surface microbial communities across different scales. Front. Microbiol. 2024, 15, 1447058. [CrossRef]
- Lara, Y.J.; McCann, A.; Malherbe, C.; François, C.; Demoulin, C.F.; Sforna, M.C.; Eppe, G.; De Pauw, E.; Wilmotte, A.; Jacques, P.; et al. Characterization of the Halochromic Gloeocapsin Pigment, a Cyanobacterial Biosignature for Paleobiology and Astrobiology. Astrobiology 2022, 22, 735–754. [CrossRef]
- Lawrence, K.P.; Long, P.F.; Young, A.R. Mycosporine-Like Amino Acids for Skin Photoprotection. Curr. Med. Chem. 2018, 25, 5512–5527. [CrossRef]
- Leach, C.M. ULTRAVIOLET-ABSORBING SUBSTANCES ASSOCIATED WITH LIGHT-INDUCED SPORULATION IN FUNGI. Can. J. Bot. 1965, 43, 185–200. [CrossRef]
- Lepot, K., Deremiens, L., Namsaraev, Z., Compère, P., Gérard, E., Verleyen, E., Tavernier, I., Hodgson, D. A., Wilmotte, A., and Javaux, E. J.: Organo-mineral imprints in fossil cyanobacterial mats of an Antarctic lake, Geobiology, 12, 424–450, 2014.
- Lesser, M. Acclimation of phytoplankton to UV-B radiation:oxidative stress and photoinhibition of photosynthesis are not prevented by UV-absorbing compounds in the dinoflagellate Prorocentrum micans. Mar. Ecol. Prog. Ser. 1996, 132, 287–297. [CrossRef]
- Lifshits, M.; Kovalerchik, D.; Carmeli, S. Microcystbiopterins A–E, five O-methylated biopterin glycosides from two Microcystis spp. bloom biomasses. Phytochemistry 2016, 123, 69–74. [CrossRef]
- Llewellyn, C.A.; Greig, C.; Silkina, A.; Kultschar, B.; Hitchings, M.D.; Farnham, G. Mycosporine-like amino acid and aromatic amino acid transcriptome response to UV and far-red light in the cyanobacterium Chlorogloeopsis fritschii PCC 6912. Sci. Rep. 2020, 10, 1–13. [CrossRef]
- Logozzo, L.; Tzortziou, M.; Neale, P.; Clark, J.B. Photochemical and Microbial Degradation of Chromophoric Dissolved Organic Matter Exported From Tidal Marshes. J. Geophys. Res. Biogeosciences 2021, 126. [CrossRef]
- Losantos, R.; Churio, M.S.; Sampedro, D. Computational Exploration of the Photoprotective Potential of Gadusol. ChemistryOpen 2015, 4, 155–160. [CrossRef]
- Losantos, R., Sampedro, D. and Churio, M. S.: Photochemistry and photophysics of mycosporine-like amino acids and gadusols, nature’s ultraviolet screens, Pure Appl. Chem., 87(9-10), 979–996, 2015b.
- Maes, W.; Pashuysen, T.; Trabucco, A.; Veroustraete, F.; Muys, B. Does energy dissipation increase with ecosystem succession? Testing the ecosystem exergy theory combining theoretical simulations and thermal remote sensing observations. Ecol. Model. 2011, 222, 3917–3941. [CrossRef]
- Mager, D. M. and Thomas, A. D.: Carbohydrates in cyanobacterial soil crusts as a source of carbon in the southwest Kalahari, Botswana, Soil Biol. Biochem., 42, 313–318, 2010.
- Mahmud, T.; Moore, D.C.; Dergachev, I.D.; Sun, H.; Varganov, S.A.; Tucker, M.J.; Jeffrey, C.S. Cortical polysaccharides play a crucial role in lichen UV protection. Cell Rep. Phys. Sci. 2024, 5. [CrossRef]
- Majerfeld, I. and Yarus, M.: A diminutive and specific RNA binding site for L-tryptophan, Nucleic Acids Res., 33, 5482–5493, 2005.
- Matsui, K.; Nazifi, E.; Hirai, Y.; Wada, N.; Matsugo, S.; Sakamoto, T. The cyanobacterial UV-absorbing pigment scytonemin displays radical-scavenging activity. J. Gen. Appl. Microbiol. 2012, 58, 137–144. [CrossRef]
- Matsumi, Y.; Kawasaki, M. Photolysis of Atmospheric Ozone in the Ultraviolet Region. Chem. Rev. 2003, 103, 4767–4782. [CrossRef]
- Matsunaga, T.; Burgess, J.G.; Yamada, N.; Komatsu, K.; Yoshida, S.; Wachi, Y. An ultraviolet (UV-A) absorbing biopterin glucoside from the marine planktonic cyanobacterium Oscillatoria sp.. Appl. Microbiol. Biotechnol. 1993, 39, 250–253. [CrossRef]
- Michaelian, K.: Thermodynamic origin of life. https://arxiv.org/abs/0907.0042 (last access: 25 July 2015), 2009.
- Michaelian, K. Thermodynamic dissipation theory for the origin of life. Earth Syst. Dyn. 2011, 2, 37–51. [CrossRef]
- Michaelian, K. HESS Opinions "Biological catalysis of the hydrological cycle: life's thermodynamic function". Hydrol. Earth Syst. Sci. 2012, 16, 2629–2645. [CrossRef]
- Michaelian, K. A non-linear irreversible thermodynamic perspective on organic pigment proliferation and biological evolution.CONFERENCE NAME, LOCATION OF CONFERENCE, COUNTRYDATE OF CONFERENCE; p. 012010.
- Michaelian, K.: Thermodynamic Dissipation Theory of the Origin and Evolution of Life: Salient characteristics of RNA and DNA and other fundamental molecules suggest an origin of life driven by UV-C light, Self-published. Printed by CreateSpace, Mexico City, ISBN: 978-1541317482, 2016.
- Michaelian, K. Microscopic dissipative structuring and proliferation at the origin of life. Heliyon 2017, 3, e00424. [CrossRef]
- Michaelian, K.; Simeonov, A. Fundamental molecules of life are pigments which arose and co-evolved as a response to the thermodynamic imperative of dissipating the prevailing solar spectrum. Biogeosciences 2015, 12, 4913–4937. [CrossRef]
- Michaelian, K., Simeonov, A.: Thermodynamic Explanation for the Cosmic Ubiquity of.
- Organic Pigments. Astrobiol Outreach (2017) 5:156. [CrossRef]
- Michaelian, K. The Dissipative Photochemical Origin of Life: UVC Abiogenesis of Adenine. Entropy 2021, 23, 217. [CrossRef]
- 2021; 133. Entropy 2021, 23(2), 217, ISSN 1099-4300. [CrossRef]
- Michaelian, K.: The Non-Equilibrium Thermodynamics of Natural Selection: From Molecules.
- Michaelian, K. The Non-Equilibrium Thermodynamics of Natural Selection: From Molecules to the Biosphere. Entropy 2023, 25, 1059. [CrossRef]
- Michaelian, K. The Pigment World: Life’s Origins as Photon-Dissipating Pigments. Life 2024, 14, 912. [CrossRef]
- Miyamoto, K.T.; Komatsu, M.; Ikeda, H. Discovery of Gene Cluster for Mycosporine-Like Amino Acid Biosynthesis from Actinomycetales Microorganisms and Production of a Novel Mycosporine-Like Amino Acid by Heterologous Expression. Appl. Environ. Microbiol. 2014, 80, 5028–5036. [CrossRef]
- Moisan, T.A.; Mitchell, B.G. UV absorption by mycosporine-like amino acids in Phaeocystis antarctica Karsten induced by photosynthetically available radiation. Mar. Biol. 2001, 138, 217–227. [CrossRef]
- Moliné, M., Libkind, D., de Garcia, V. and Giraudo, M. R.: Production of Pigments and Photo-Protective Compounds by Cold-Adapted Yeasts, In: Buzzini, P. and Margesin, R. (eds.) Cold-adapted Yeasts: Biodiversity, Adaptation Strategies and Biotechnological Significance, Springer Berlin Heidelberg, pp 193-224, 2014.
- Morel, A. Optical modeling of the upper ocean in relation to its biogenous matter content (case I waters). J. Geophys. Res. Oceans 1988, 93, 10749–10768. [CrossRef]
- Mulkidjanian, A.Y.; Junge, W. On the origin of photosynthesis as inferred from sequence analysis. Photosynth. Res. 1997, 51, 27–42. [CrossRef]
- Mulkidjanian, A.Y.; A Cherepanov, D.; Galperin, M.Y. Survival of the fittest before the beginning of life: selection of the first oligonucleotide-like polymers by UV light. BMC Evol. Biol. 2003, 3, 12–12. [CrossRef]
- Murchie, E. H., & Ruban, A. V. (2020). Non-photochemical quenching in plants: From molecular mechanisms to crop improvement. Annual Review of Plant Biology, 71, 739–766. [CrossRef]
- Nägeli, C. Gattungen einzelliger Algen physiologisch und systematisch bearbeitet, von Carl Nägeli.; Smithsonian Institution: Washington, DC, United States, 1849; ISBN: .
- Nägeli, C. and Schwenderer, S.: Das Mikroskop, 2nd edn. Willhelm Engelmann Verlag, Leipzig, 505 p., 1877.
- Nelson, N.B.; Siegel, D.A. The Global Distribution and Dynamics of Chromophoric Dissolved Organic Matter. Annu. Rev. Mar. Sci. 2013, 5, 447–476. [CrossRef]
- Nelson, N.; Siegel, D.; Michaels, A. Seasonal dynamics of colored dissolved material in the Sargasso Sea. Deep. Sea Res. Part I: Oceanogr. Res. Pap. 1998, 45, 931–957. [CrossRef]
- Olivucci, M. (Ed.): Computational Photochemistry, Elsevier, Amsterdam, 2005.
- Orellana, G.; Gómez-Silva, B.; Urrutia, M.; Galetović, A. UV-A Irradiation Increases Scytonemin Biosynthesis in Cyanobacteria Inhabiting Halites at Salar Grande, Atacama Desert. Microorganisms 2020, 8, 1690. [CrossRef]
- Oren, A. and Gunde-Cimerman, N.: Mycosporines and mycosporinelike amino acids: UV protectants or multipurpose secondary metabolites? FEMS Microbiol Lett, 269, 1–10, 2007.
- Organelli, E.; Bricaud, A.; Antoine, D.; Matsuoka, A. Seasonal dynamics of light absorption by chromophoric dissolved organic matter (CDOM) in the NW Mediterranean Sea (BOUSSOLE site). Deep. Sea Res. Part I: Oceanogr. Res. Pap. 2014, 91, 72–85. [CrossRef]
- Patara, L.; Vichi, M.; Masina, S.; Fogli, P.G.; Manzini, E. Global response to solar radiation absorbed by phytoplankton in a coupled climate model. Clim. Dyn. 2012, 39, 1951–1968. [CrossRef]
- Pathak, J., Rajneesh, Richa, Sonker, A.S., Kannaujiya, V. K. and Sinha, R. P.: Cyanobacterial extracellular polysaccharide sheath pigment, scytonemin: A novel multipurpose pharmacophore, In: Se-Kwon, K. (ed.) Marine Glycobiology: Principles and Applications, 1st edition, Taylor & Francis Group, 6000 Broken Sound Parkway NW, Suite 300, Boca Raton, FL 33487-2742 CRC Press, Pages 323–337, 2016.
- Pereira, S.; Zille, A.; Micheletti, E.; Moradas-Ferreira, P.; De Philippis, R.; Tamagnini, P. Complexity of cyanobacterial exopolysaccharides: composition, structures, inducing factors and putative genes involved in their biosynthesis and assembly. FEMS Microbiol. Rev. 2009, 33, 917–941. [CrossRef]
- putative genes involved in their biosynthesis and assembly. FEMS Microbiol. Rev. 33, 917–941, 2009.
- Ploutno, A. and Carmeli, S.: Prenostodione, a novel UV-absorbing metabolite from a natural bloom of the cyanobacterium Nostoc species, J Nat Prod, 64, 544-545, 2001.
- Pokorny, J.; Brom, J.; Cermak, J.; Hesslerova, P.; Huryna, H.; Nadezhdina, N.; Rejskova, A. Solar energy dissipation and temperature control by water and plants. Int. J. Water 2010, 5, 311. [CrossRef]
- Pope, M.A.; Spence, E.; Seralvo, V.; Gacesa, R.; Heidelberger, S.; Weston, A.J.; Dunlap, W.C.; Shick, J.M.; Long, P.F. O-Methyltransferase Is Shared between the Pentose Phosphate and Shikimate Pathways and Is Essential for Mycosporine-Like Amino Acid Biosynthesis in Anabaena variabilis ATCC 29413. ChemBioChem 2014, 16, 320–327. [CrossRef]
- Portwich, A. and Garcia-Pichel, F.: A novel prokaryotic UVB photoreceptor in the cyanobacterium Chlorogloeopsis PCC 6912, Photochem Photobiol, 71, 493–498, 2000.
- Portwich, A.; Garcia-Pichel, F. Biosynthetic pathway of mycosporines (mycosporine-like amino acids) in the cyanobacterium Chlorogloeopsis sp. strain PCC 6912. Phycologia 2003, 42, 384–392. [CrossRef]
- Prigogine, I.: Thermodynamics of Irreversible Processes, Wiley, New York, 1967.
- Proteau, P.J.; Gerwick, W.H.; Garcia-Pichel, F.; Castenholz, R. The structure of scytonemin, an ultraviolet sunscreen pigment from the sheaths of cyanobacteria. Cell. Mol. Life Sci. 1993, 49, 825–829. [CrossRef]
- Pullan, D.; Westall, F.; Hofmann, B.A.; Parnell, J.; Cockell, C.S.; Edwards, H.G.; Villar, S.E.J.; Schröder, C.; Cressey, G.; Marinangeli, L.; et al. Identification of Morphological Biosignatures in Martian Analogue Field Specimens Using In Situ Planetary Instrumentation. Astrobiology 2008, 8, 119–156. [CrossRef]
- Quesada, A. and Vincent, W. F.: Strategies of adaptation by Antarctic cyanobacteria to ultraviolet radiation, Eur J Phycol, 32, 335–342, 1997.
- Rastogi, R. P. and Incharoensakdi, A.: Characterization of UV-screening compounds, mycosporine-like amino acids, and scytonemin in the cyanobacterium Lyngbya sp. CU2555, FEMS Microbiol Ecol., 87(1), 244-256, 2014.
- Rastogi, R. P. and Madamwar, D.: Cyanobacteria Synthesize their own UV-Sunscreens for Photoprotection, Bioenergetics, 5, 138, 2016. [CrossRef]
- Rastogi, R. P., Richa, S. R. P., Singh, S. P. and Häder, D. P.: Photoprotective compounds from marine organisms, J Ind Microbiol Biotechnol, 37, 537–558, 2010.
- Rastogi, R. P., Sinha, R. P., Moh, S. H., Lee, T. K., Kottuparambil, S., Kim, Y. J., Rhee, J. S., Choi, E. M., Brown, M. T., Häder, D. P. and Han, T.: Ultraviolet radiation and cyanobacteria, Journal of Photochemistry and Photobiology B: Biology, 141, 154–169, 2014.
- Rastogi, R.P.; Sinha, R.P.; Incharoensakdi, A. Partial characterization, UV-induction and photoprotective function of sunscreen pigment, scytonemin from Rivularia sp. HKAR-4. Chemosphere 2013, 93, 1874–1878. [CrossRef]
- Řezanka, T.; Temina, M.; Tolstikov, A.G.; Dembitsky, V.M. Natural microbial UV radiation filters — Mycosporine-like amino acids. Folia Microbiol. 2004, 49, 339–352. [CrossRef]
- Richards, T. A., Dacks, J. B., Campbell, S. A., Blanchard, J. L., Foster, P. G., McLeod, R. and Roberts, C. W.: Evolutionary origins of the eukaryotic shikimate pathway: gene fusions, horizontal gene transfer, and endosymbiotic replacements, Eukaryot Cell, 5(9), 1517-1531, 2006.
- Roeselers, G.; Norris, T.B.; Castenholz, R.W.; Rysgaard, S.; Glud, R.N.; Kühl, M.; Muyzer, G. Diversity of phototrophic bacteria in microbial mats from Arctic hot springs (Greenland). Environ. Microbiol. 2006, 9, 26–38. [CrossRef]
- Romera-Castillo, C., Sarmento, H., Álvarez-Salgado, X. A., Gasol, J. M. and Marrasé, C.: Production of chromophoric dissolved organic matter by marine phyto-plankton, Limnol.Oceanogr., 55(1), 446–454, 2010.
- Rosic, N.N. Mycosporine-Like Amino Acids: Making the Foundation for Organic Personalised Sunscreens. Mar. Drugs 2019, 17, 638. [CrossRef]
- Rosic, N.N. Recent advances in the discovery of novel marine natural products and mycosporine-like amino acid UV-absorbing compounds. Appl. Microbiol. Biotechnol. 2021, 105, 7053–7067. [CrossRef]
- Rosic, N.N.; Dove, S. Mycosporine-Like Amino Acids from Coral Dinoflagellates. Appl. Environ. Microbiol. 2011, 77, 8478–8486. [CrossRef]
- Rossi, F. and De Philippis, R.: Role of Cyanobacterial Exopolysaccharides in Phototrophic Biofilms and in Complex Microbial Mats, Life (Basel), 5(2), 1218–1238, 2015.
- Rossi, F.; Potrafka, R.M.; Pichel, F.G.; De Philippis, R. The role of the exopolysaccharides in enhancing hydraulic conductivity of biological soil crusts. Soil Biol. Biochem. 2012, 46, 33–40. [CrossRef]
- Röttgers, R.; Koch, B.P. Spectroscopic detection of a ubiquitous dissolved pigment degradation product in subsurface waters of the global ocean. Biogeosciences 2012, 9, 2585–2596. [CrossRef]
- Rowan, K. S.: Photosynthetic Pigments of Algae, Cambridge University Press, Cambridge, 112–210, 1989.
- Rozema, J.; Björn, L.; Bornman, J.; Gaberščik, A.; Häder, D.-P.; Trošt, T.; Germ, M.; Klisch, M.; Gröniger, A.; Sinha, R.; et al. The role of UV-B radiation in aquatic and terrestrial ecosystems—an experimental and functional analysis of the evolution of UV-absorbing compounds. J. Photochem. Photobiol. B: Biol. 2002, 66, 2–12. [CrossRef]
- Ruban, A.V.; Berera, R.; Ilioaia, C.; Van Stokkum, I.H.M.; Kennis, J.T.M.; Pascal, A.; van Amerongen, H.; Robert, B.; Horton, P.; Van Grondelle, R. Identification of a mechanism of photoprotective energy dissipation in higher plants. Nature 2007, 450, 575–578. [CrossRef]
- Ručová, D.; Vilková, M.; Sovová, S.; Vargová, Z.; Kostecká, Z.; Frenák, R.; Routray, D.; Bačkor, M. Photoprotective and antioxidant properties of scytonemin isolated from Antarctic cyanobacterium Nostoc commune Vaucher ex Bornet & Flahault and its potential as sunscreen ingredient. J. Appl. Phycol. 2023, 35, 2839–2850. [CrossRef]
- Ryan, K.G.; McMinn, A.; Mitchell, K.A.; Trenerry, L. Mycosporine-Like Amino Acids in Antarctic Sea Ice Algae, and Their Response to UVB Radiation. Z. Fur Naturforschung Sect. C-A J. Biosci. 2002, 57, 471–477. [CrossRef]
- Sagan, C. Ultraviolet selection pressure on the earliest organisms. J. Theor. Biol. 1973, 39, 195–200. [CrossRef]
- Sampedro, D. Computational exploration of natural sunscreens. Phys. Chem. Chem. Phys. 2011, 13, 5584–5586. [CrossRef]
- Schermann, J.-P. Spectroscopy and Modeling of Biomolecular Building Blocks; Elsevier: Amsterdam, NX, Netherlands, 2008; ISBN: .
- Schmid, D., Schürch, C. and Zülli, F.: UV-A sunscreen from red algae for protection against premature skin aging, Cosmet. Toilet. Manuf. Worldw., 115, 139–143, 2000.
- Schopf, J.W. Microfossils of the Early Archean Apex Chert: New Evidence of the Antiquity of Life. Science 1993, 260, 640–646. [CrossRef]
- Schopf, J.W.; Packer, B.M. Early Archean (3.3-Billion to 3.5-Billion-Year-Old) Microfossils from Warrawoona Group, Australia. Science 1987, 237, 70–73. [CrossRef]
- Schopf, J.W.; Kudryavtsev, A.B.; Agresti, D.G.; Wdowiak, T.J.; Czaja, A.D. Laser–Raman imagery of Earth's earliest fossils. Nature 2002, 416, 73–76. [CrossRef]
- Shick, J. M. and Dunlap, W. C.: Mycosporine-like amino acids and related gadusols: biosynthesis, accumulation, and UV protective functions in aquatic organisms, Annu Rev Plant.
- Physiol., 64, 223–262, 2002.
- Shick, J.M.; Romaine-Lioud, S.; Ferrier-Pagès, C.; Gattuso, J.-P. Ultraviolet-B radiation stimulates shikimate pathway-dependent accumulation of mycosporine-like amino acids in the coral Stylophora pistillata despite decreases in its population of symbiotic dinoflagellates. Limnol. Oceanogr. 1999, 44, 1667–1682. [CrossRef]
- Siegel, D.A.; Maritorena, S.; Nelson, N.B.; Hansell, D.A.; Lorenzi-Kayser, M. Global distribution and dynamics of colored dissolved and detrital organic materials. J. Geophys. Res. Oceans 2002, 107, 21-1–21-14. [CrossRef]
- Singh, S.P.; Häder, D.-P.; Sinha, R.P. Bioinformatics evidence for the transfer of mycosporine-like amino acid core (4-deoxygadusol) synthesizing gene from cyanobacteria to dinoflagellates and an attempt to mutate the same gene (YP_324358) in Anabaena variabilis PCC 7937. Gene 2012, 500, 155–163. [CrossRef]
- Singh, S.P.; Klisch, M.; Sinha, R.P.; Häder, D.-P. Genome mining of mycosporine-like amino acid (MAA) synthesizing and non-synthesizing cyanobacteria: A bioinformatics study. Genomics 2009, 95, 120–128. [CrossRef]
- Singh, A.; Čížková, M.; Bišová, K.; Vítová, M. Exploring Mycosporine-Like Amino Acids (MAAs) as Safe and Natural Protective Agents against UV-Induced Skin Damage. Antioxidants 2021, 10, 683. [CrossRef]
- Singh, V.K.; Jha, S.; Rana, P.; Mishra, S.; Kumari, N.; Singh, S.C.; Anand, S.; Upadhye, V.; Sinha, R.P. Resilience and Mitigation Strategies of Cyanobacteria under Ultraviolet Radiation Stress. Int. J. Mol. Sci. 2023, 24, 12381. [CrossRef]
- Sinha, R. P. and Häder, D. P.: UV-protectants in cyanobacteria, Plant Sci., 174, 278–289, 2008.
- Sinha, R.P.; Klisch, M.; Helbling, E.W.; Häder, D.-P. Induction of mycosporine-like amino acids (MAAs) in cyanobacteria by solar ultraviolet-B radiation. J. Photochem. Photobiol. B: Biol. 2001, 60, 129–135. [CrossRef]
- Sinha, R. P., Klisch, M., Vaishampayan, A. and Häder, D. P.: Biochemical and spectroscopic characterization of the cyanobacterium Lyngbya sp. Inhabiting Mango (Mangifera indica) trees: presence of an ultraviolet-absorbing pigment, scytonemin, Acta Protozool, 38, 291–298, 1999.
- Sinha, R. P., Singh, S. P. and Häder, D. P.: Database on mycosporines and mycosporine-like amino acids (MAAs) in fungi, cyanobacteria, macroalgae, phytoplankton and animals, J. Photochem. Photobiol. B, 89, 29–35, 2007.
- Sinha, R.P.; Klisch, M.; Gröniger, A.; Häder, D.-P. Mycosporine-like amino acids in the marine red alga Gracilaria cornea — effects of UV and heat. Environ. Exp. Bot. 2000, 43, 33–43. [CrossRef]
- Son, M.; Pinnola, A.; Gordon, S.C.; Bassi, R.; Schlau-Cohen, G.S. Observation of dissipative chlorophyll-to-carotenoid energy transfer in light-harvesting complex II in membrane nanodiscs. Nat. Commun. 2020, 11, 1–8. [CrossRef]
- Soppa, M.A.; Pefanis, V.; Hellmann, S.; Losa, S.N.; Hölemann, J.; Martynov, F.; Heim, B.; Janout, M.A.; Dinter, T.; Rozanov, V.; et al. Assessing the Influence of Water Constituents on the Radiative Heating of Laptev Sea Shelf Waters. Front. Mar. Sci. 2019, 6. [CrossRef]
- Sorrels, C.M.; Proteau, P.J.; Gerwick, W.H. Organization, Evolution, and Expression Analysis of the Biosynthetic Gene Cluster for Scytonemin, a Cyanobacterial UV-Absorbing Pigment. Appl. Environ. Microbiol. 2009, 75, 4861–4869. [CrossRef]
- Soule, T.; Garcia-Pichel, F.; Stout, V. Gene Expression Patterns Associated with the Biosynthesis of the Sunscreen Scytonemin in Nostoc punctiforme ATCC 29133 in Response to UVA Radiation. J. Bacteriol. 2009, 191, 4639–4646. [CrossRef]
- Soule, T.; Palmer, K.; Gao, Q.; Potrafka, R.M.; Stout, V.; Garcia-Pichel, F. A comparative genomics approach to understanding the biosynthesis of the sunscreen scytonemin in cyanobacteria. BMC Genom. 2009, 10, 336–336. [CrossRef]
- Soule, T.; Stout, V.; Swingley, W.D.; Meeks, J.C.; Garcia-Pichel, F. Molecular Genetics and Genomic Analysis of Scytonemin Biosynthesis in Nostoc punctiforme ATCC 29133. J. Bacteriol. 2007, 189, 4465–4472. [CrossRef]
- Staleva, H.; Komenda, J.; Shukla, M.K.; Šlouf, V.; Kaňa, R.; Polívka, T.; Sobotka, R. Mechanism of photoprotection in the cyanobacterial ancestor of plant antenna proteins. Nat. Chem. Biol. 2015, 11, 287–291. [CrossRef]
- Stanley, S. M.: Earth System History, 3rd edn., W. H. Freeman and Company, New York, NY, 263–287, 2008.
- Starcevic, A., Akthar, S., Dunlap, W. C., Shick, J. M., Hranueli, D., Cullum, J. and Long, P. F.: Enzymes of the shikimic acid pathway encoded in the genome of a basal metazoan, Nematostella vectensis, have microbial origins, Proc. Natl. Acad. Sci. USA, 105, 2533–2537, 2008.
- Steinberg, D.K.; Nelson, N.B.; Carlson, C.A.; Prusak, A. Production of chromophoric dissolved organic matter (CDOM) in the open ocean by zooplankton and the colonial cyanobacterium Trichodesmium spp.. Mar. Ecol. Prog. Ser. 2004, 267, 45–56. [CrossRef]
- Storme, J.-Y.; Golubic, S.; Wilmotte, A.; Kleinteich, J.; Velázquez, D.; Javaux, E.J. Raman Characterization of the UV-Protective Pigment Gloeocapsin and Its Role in the Survival of Cyanobacteria. Astrobiology 2015, 15, 843–857. [CrossRef]
- Subramaniam, A., Carpenter, E. J., Karentz, D. and Falkowski, P. G.: Bio-optical properties of the marine diazotrophic cyanobacteria Trichodesmium spp. I. Absorption and photosynthetic action spectra, Limnol Oceanogr, 44, 608–617, 1999.
- Sun, Y.; Zhang, N.; Zhou, J.; Dong, S.; Zhang, X.; Guo, L.; Guo, G. Distribution, Contents, and Types of Mycosporine-Like Amino Acids (MAAs) in Marine Macroalgae and a Database for MAAs Based on These Characteristics. Mar. Drugs 2020, 18, 43. [CrossRef]
- Zhao, Y.-C.; Wang, C.-C.; Li, X.-Y.; Wang, D.-D.; Wang, Y.-M.; Xue, C.-H.; Wen, M.; Zhang, T.-T. Supplementation of n-3 PUFAs in Adulthood Attenuated Susceptibility to Pentylenetetrazol Induced Epilepsy in Mice Fed with n-3 PUFAs Deficient Diet in Early Life. Mar. Drugs 2023, 21, 354. [CrossRef]
- Tamre, E.; Fournier, G.P. Inferred ancestry of scytonemin biosynthesis proteins in cyanobacteria indicates a response to Paleoproterozoic oxygenation. Geobiology 2022, 20, 764–775. [CrossRef]
- Tang, Q.; Li, Z.; Chen, N.; Luo, X.; Zhao, Q. Natural pigments derived from plants and microorganisms: classification, biosynthesis, and applications. Plant Biotechnol. J. 2024, 23, 592–614. [CrossRef]
- Tice, M. M and Lowe, D. R.: Photosynthetic microbial mats in the 3,416-Myr-old ocean, Nature, 431(7008), 549–552, 2004.
- Tilstone, G.H.; Airs, R.L.; Vicente, V.M.; Widdicombe, C.; Llewellyn, C. High concentrations of mycosporine-like amino acids and colored dissolved organic matter in the sea surface microlayer off the Iberian Peninsula. Limnol. Oceanogr. 2010, 55, 1835–1850. [CrossRef]
- Trione, E.J.; Leach, C.M. Light-induced sporulation and sporogenic substances in fungi.. 1969, 59, 1077–83.
- Uemura, D.; Katayama, C.; Wada, A.; Hirata, Y. CRYSTAL AND MOLECULAR STRUCTURE OF PALYTHENE POSSESSING A NOVEL 360 NM CHROMOPHORE. Chem. Lett. 1980, 9, 755–756. [CrossRef]
- Ustin, S.L.; Valko, P.G.; Kefauver, S.C.; Santos, M.J.; Zimpfer, J.F.; Smith, S.D. Remote sensing of biological soil crust under simulated climate change manipulations in the Mojave Desert. Remote. Sens. Environ. 2008, 113, 317–328. [CrossRef]
- Van Kranendonk, M. J., Philippot, P., Lepot, K., Bodorkos, S. and Pirajno, F.: Geological setting of Earth’s oldest fossils in the c. 3.5 Ga Dresser Formation, Pilbara craton, Western Australia, Precambr. Res., 167, 93–124, 2008.
- Varnali, T.; Edwards, H.G.M. Ab initio calculations of scytonemin derivatives of relevance to extremophile characterization by Raman spectroscopy. Philos. Trans. R. Soc. A: Math. Phys. Eng. Sci. 2010, 368, 3193–3203. [CrossRef]
- Varnali, T.; Edwards, H.G.M. Raman spectroscopic identification of scytonemin and its derivatives as key biomarkers in stressed environments. Philos. Trans. R. Soc. A: Math. Phys. Eng. Sci. 2014, 372, 20140197. [CrossRef]
- Vernet, M.; Whitehead, K. Release of ultraviolet-absorbing compounds by the red-tide dinoflagellateLingulodinium polyedra. Mar. Biol. 1996, 127, 35–44. [CrossRef]
- Wada, N.; Sakamoto, T.; Matsugo, S. Mycosporine-Like Amino Acids and Their Derivatives as Natural Antioxidants. Antioxidants 2015, 4, 603–646. [CrossRef]
- Waller, R.F.; Slamovits, C.H.; Keeling, P.J. Lateral Gene Transfer of a Multigene Region from Cyanobacteria to Dinoflagellates Resulting in a Novel Plastid-Targeted Fusion Protein. Mol. Biol. Evol. 2006, 23, 1437–1443. [CrossRef]
- Walter, M.R.; Buick, R.; Dunlop, J.S.R. Stromatolites 3,400–3,500 Myr old from the North Pole area, Western Australia. Nature 1980, 284, 443–445. [CrossRef]
- Wellman, C. H., Strother, P. K., & Servais, T. (2021). The terrestrialization process: A palaeobotanical and palynological perspective. *Annual Review of Earth and Planetary Sciences*, 49, 423–451. [CrossRef]
- Westall, F. Life on an Anaerobic Planet. Science 2009, 323, 471–472. [CrossRef]
- Westall, F., de Ronde, C. E. J., Southam, G., Grassineau, N., Colas, M., Cockell, C. and Lammer, H.: Implications of a 3.472-3.333 Gyr-old subaerial microbial mat from the Barberton greenstone belt, South Africa for the UV environmental conditions on the early Earth, Philosophical Transactions of The Royal Society B, 361, 1857-1875, 2006.
- Whitehead, K.; Vernet, M. Influence of mycosporine-like amino acids (MAAs) on UV absorption by particulate and dissolved organic matter in La Jolla Bay. Limnol. Oceanogr. 2000, 45, 1788–1796. [CrossRef]
- Wurl, O.; Bird, K.; Cunliffe, M.; Landing, W.M.; Miller, U.; Mustaffa, N.I.H.; Ribas-Ribas, M.; Witte, C.; Zappa, C.J. Warming and Inhibition of Salinization at the Ocean's Surface by Cyanobacteria. Geophys. Res. Lett. 2018, 45, 4230–4237. [CrossRef]
- Wurfel, P.; Ruppel, W. The flow equilibrium of a body in a radiation field. J. Phys. C: Solid State Phys. 1985, 18, 2987–3000. [CrossRef]
- Wynn-Williams, D. D., Edwards, H. G. M. and Garcia-Pichel, F.: Functional biomolecules of Antarctic stromatolitic and endolithic cyanobacterial communities, Eur. J. Phycol., 34, 381–391, 1999.
- Wynn-Williams, D.; Edwards, H.; Newton, E.; Holder, J. Pigmentation as a survival strategy for ancient and modern photosynthetic microbes under high ultraviolet stress on planetary surfaces. Int. J. Astrobiol. 2002, 1, 39–49. [CrossRef]
- Yamashita, Y.; Tanoue, E. Chemical characterization of protein-like fluorophores in DOM in relation to aromatic amino acids. Mar. Chem. 2003, 82, 255–271. [CrossRef]
- Yarus, M.; Widmann, J.J.; Knight, R. RNA–Amino Acid Binding: A Stereochemical Era for the Genetic Code. J. Mol. Evol. 2009, 69, 406–429. [CrossRef]
- Zhu, H.; Zhang, T.; Zhang, P.; Peng, C. Pigment patterns and photoprotection of anthocyanins in the young leaves of four dominant subtropical forest tree species in two successional stages under contrasting light conditions. Tree Physiol. 2016, 36, 1092–1104. [CrossRef]
- Zwerger, M.J.; Hammerle, F.; Siewert, B.; Ganzera, M. Application of feature-based molecular networking in the field of algal research with special focus on mycosporine-like amino acids. J. Appl. Phycol. 2023, 35, 1377–1392. [CrossRef]
Figure 1.
Chemical structures of some common mycosporines and MAAs.
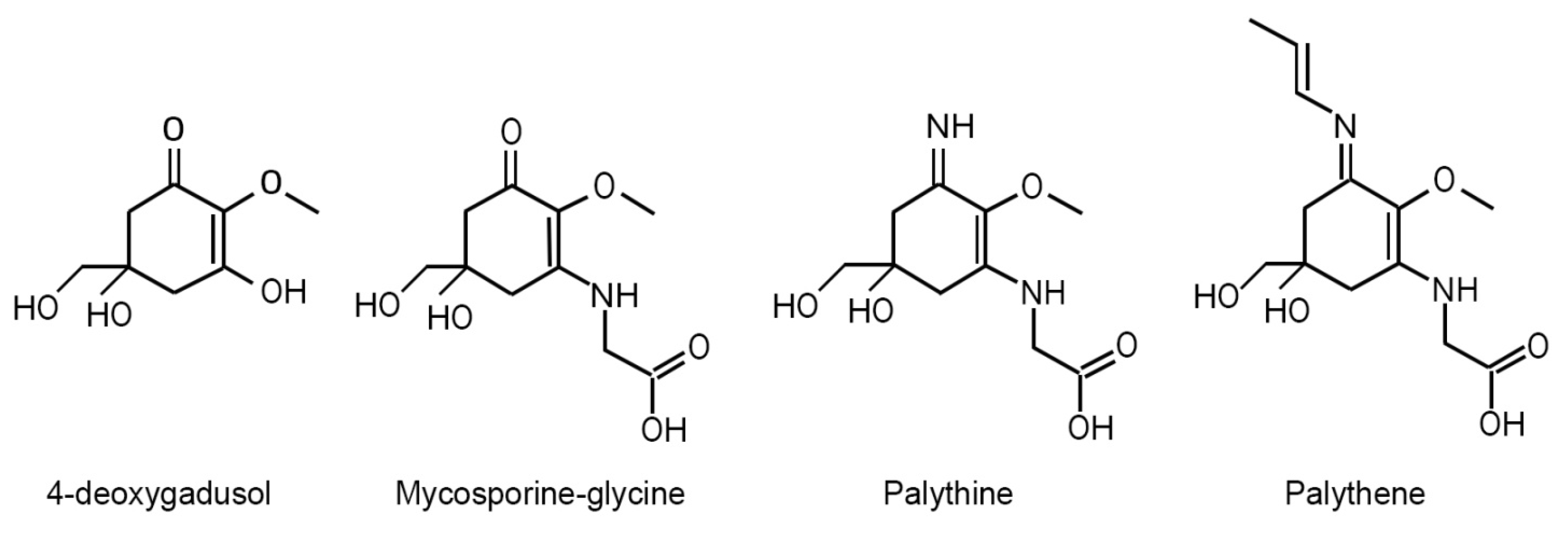
Figure 2.
Chemical structures of scytonemin and reduced scytonemin.
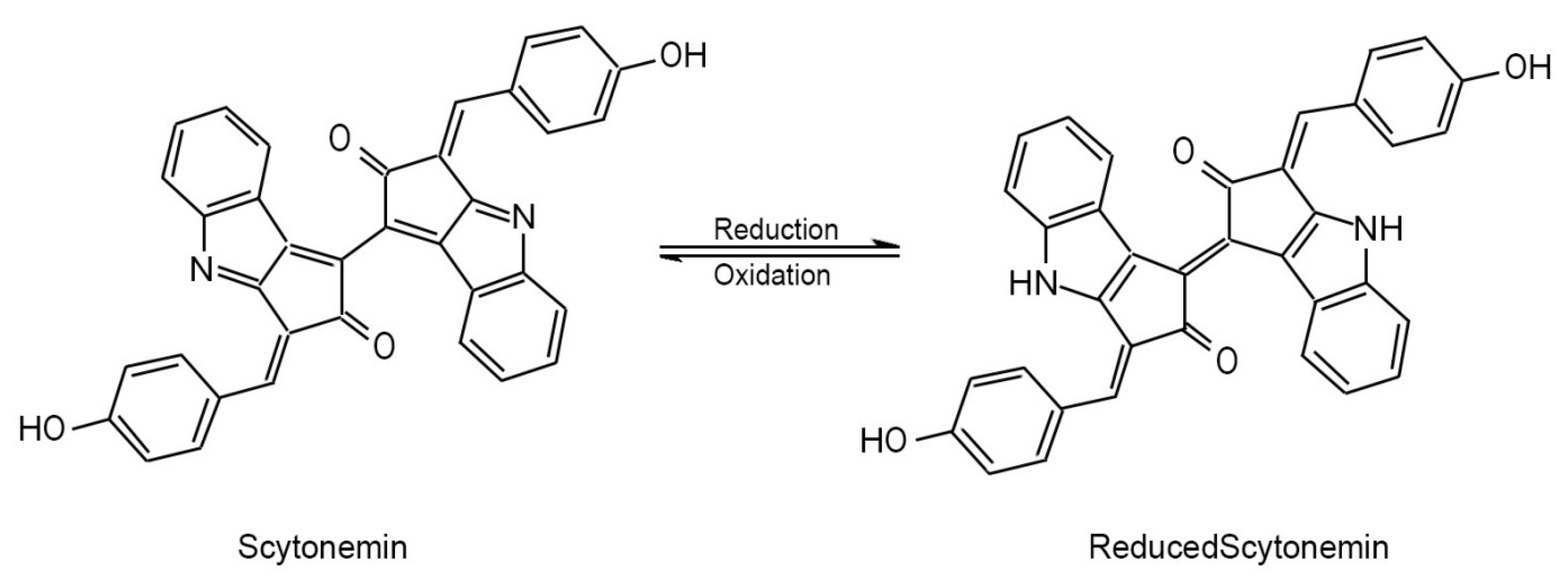

Table 1.
Comparison between the photophysical properties of the aromatic amino acids and representative compounds of the major classes of cyanobacterial UV-absorbing pigments.
Table 1.
Comparison between the photophysical properties of the aromatic amino acids and representative compounds of the major classes of cyanobacterial UV-absorbing pigments.
| UV bio-pigments | (nm) | (M-1cm-1) | Electronic excited state lifetime (ns) | Fluorescence quantum yield () |
| Aromatic amino acids | ||||
| Phenylalaninea | 257 | 195 | 7.5 | 0.024 |
| Tyrosinea | 274 | 1405 | 2.5 | 0.14 |
| Tryptophana | 278 | 5579 | 3.03 | 0.13 |
| Mycosporines and MAAs | ||||
| Gadusolb | 269 | 12400 | / | non-fluorescent |
| Mycosporine-γ-aminobutyric acidc | 310 | 28900 | / | / |
| Mycosporine-glutamic acidc | 311 | 20900 | / | / |
| Palythineb, c | 320 | 36200 | / | non-fluorescent |
| Shinorineb | 333 | 44700 | 0.35 | 0.002 |
| Porphyra-334b | 334 | 42300 | 0.4 | 0.0016 |
| Palythenec | 360 | 50000 | / | / |
| Scytonemins | ||||
|
Scytonemind, e |
212, 252, 278, 300, 384 |
/ / / / 16200 |
/ |
non-fluorescent |
|
Reduced Scytonemind |
246, 276, 314, 378, 474, 572 |
30000, 14000, 15000, 22000, 14000, 7600 |
/ | / |
|
Scytonemin-iminef |
237, 366, 437, 564 |
/ / / / |
/ | / |
| Dimethoxyscytonemind | 215, 316, 422 |
60354 18143, 23015 |
/ | / |
| Tetramethoxyscytonemind | 212, 562 |
35928 5944 |
/ | / |
| Scytonined | 207, 225, 270 |
38948 37054, 22484 |
/ | / |
| Other poorly characterized cyanobacterial UV-absorbing pigments | ||||
| Gloeocapsing | 392 | / | / | / |
| Microcystbiopterinsh | ~ 275, ~ 350 |
10000, 3500 |
/ | / |
aBerezin and Achilefu (2010); bLosantos et al. (2015a); cWada et al. (2015); dVarnali and Edwards (2014); eBalskus et al. (2011); fGrant and Louda (2013); gStorme et al. (2015); hLifshits et al. (2016).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



