Submitted:
28 April 2025
Posted:
29 April 2025
You are already at the latest version
Abstract
The valorization of plant biomass is one of the main strategies for sustainable development. However, its use as energy, biofuels, fertilizers, value-added products, or even food is severely affected by the complexity of the plant cell wall. Therefore, the evaluation of fungi with high production of lignocellulolytic enzymes capable of efficiently degrading these substrates constitutes a viable, clean, and eco-friendly solution, allowing, for example, an increase in the digestibility and nutritional quality of alternative animal feed sources. For these reasons, the present study evaluated the ability of the mutant strain Trichodema viride M5-2 to improve the nutritional composition of the forage legumes Lablab purpureus and Mucuna pruriens through solid-state fermentation. Endo- and exoglucanase cellulolytic activity was assessed, as well as the effect of fermentation on the fiber's physical properties and chemical composition. Molecular changes in the structure of plant fiber were analyzed using infrared spectroscopy. Increased production of the cellulolytic complex of the enzymes endoglucanase (3.29 IU/ml) and exoglucanase (0.64 IU/ml) was achieved in M. pruriens. The chemical composition showed an increase in true protein and a decrease in Neutral Fiber, Hemicellulose, and Cellulose, with a consequent improvement of nutritional quality. Fiber degradation was evident in the infrared spectrum with a significant decrease in the signals associated with cellulose and, to a lesser extent, with lignin. It can be concluded that the mutant strain T. viride M5-2 produced chemical, physical, and molecular changes in the fibrous and protein fractions of L. purpureus and M. pruriens through SSF, which improved their nutritional value as an alternative feed source for animal nutrition.
Keywords:
mucuna
; dolicho
; fungi
; degradation
; fiber
; cellulolytic enzymes
; chemical composition
; animal feed
1. Introduction
Global population growth will continue to generate greater demand for animal production. One of the solutions to the food shortage is improving the nutritional quality of agroindustrial waste and other alternative food sources. The lignocellulosic biomass produced from these wastes has low nitrogen content, high fiber content, and low nutrient density; its processing can increase its nutritional value [1]. Despite this, most of the processes used to revalue these residues are complex due to the structure of the plant cell wall.
In tropical and subtropical countries, the production of legume meals could be a very attractive alternative for feeding monogastric species due to its low cost and lack of competition with human food. Savón et al. [2] demonstrated the possibility of using forage meals from tropical legumes such as Lablab purpureus and Mucuna pruriens in poultry and pig rations. However, these feeds have limitations in terms of the biological availability of several nutrients [3], which could be ameliorated by the application of fungal solid-state fermentation (SSF) processes [4].
In recent years, solid-state fermentation has attracted the interest of scientists and industry due to its potential in the circular bioeconomy [5]. Microbial cultivation on waste-based substrates achieves the dual benefit of complete utilization of the waste and the production of value-added products, such as enzymes or secondary metabolites. They also generate biochemical and structural modifications in a large part of the fiber components, increasing their nutritional value [6], the biological value of dietary protein [7,8,9] and improving the structure of lignocellulosic compounds [10,11] of the resulting product, the production of high-value bioproducts, such as bioactive molecules for its use as ingredients in biostimulants [12].
The literature reports the cellulolytic activity of many microorganisms, including several species of fungi such as Trichoderma, Aspergillus and Penicilium, corresponding to more than 50% of the studies related to cellulases. Their use in a more productive way is conditioned by the utilizaation of highly efficient enzyme mixtures [13]. These microorganisms produce enzymes capable of improving the nutritional value and reducing the fiber content and antinutritional factors in Vigna unguiculata, L. purpureus, and M. pruriens [14]. Therefore, novel feeds obtained from processed legumes are undoubtedly useful for animal and human nutrition, due to their beneficial effects and use in disease prevention.
The mutant fungus Trichoderma viride M5-2 is capable to carry out bioconversion processes in sugarcane (Saccharum officinarum Lin) bagasse pretreated [15] and grain [16] meals in Vigna unguiculata, Canavalia ensiformis, M.pruriens, L. purpureus. This previous study promoted the analysis of modifications in feed quality, nutrient utilization characteristics, intrinsic structure, and components of the biological matrix [17] and molecular changes in the wall. Therefore, the effect of the mutant strain Trichoderma viride M5-2 on improving the nutritional composition of whole forage flour meals of legumes through solid-state fermentation was evaluated.
2. Materials and Methods
Fungal Species
The mutant strain of lignocellulolytic fungus T. viride M5-2 was used. It was isolated from sugarcane bagasse with nucleotide sequence number registered in GenBank and accession number: KY977981, which is part of the Institute of Animal Science (ICA) collection of microorganisms [18].
Whole Forage Flour Meals of Legumes Substrate Preparation
Whole forage flour meals of legumes from L. purpureus and M. pruriens forage were used for fermentation. The legumes were grown in the experimental area of the ICA. The flours were made with whole plants (leaves, stems and grains) cut at 5 cm above ground level, when 100% of the plants had pods and 50% of the pods were in a milky state. They were then spread on a plate and dried for 2-3 days until the humidity was reduced to 20-25% to allow grinding and prevent any fermentation process. The dried plants were reduced to a particle size of 1 mm ± 0.2 mm in a hammer mill.
Observation of the Cultural Characteristics of the Fungus T. viride M5-2 in Legume Flours
To determine the growth capacity of the fungus in the different flours, cultural characteristics (mycelium and sporulation) were observed at a temperature of 30±1°C and 70% humidity, without macro and micronutrients addition to the flour. This trial only considered the growth or absence of the fungus in the different legume flours, depending on the experimental conditions of the subsequent solid-state fermentation (SSF) test.
Solid State Fermentation Process
For the experiment, 500 mL Erlenmeyer flasks were used with 10 g of each of the whole forage flours meals, which were moistened with water up to 70% according to a previous analysis of the dry matter of the whole flours. The moist whole forage flours meals were sterilized at 121°C for 20 minutes (min) and inoculated with 1 cm2 of malt agar grown and completely covered with spores of the fungus, which represents 10% of the fungal inoculum. No urea or other nutrients were added during fermentation, only those from the plant material, which have a high nitrogen content. The mixture was homogenized and the Erlenmeyer flasks were placed in an incubator at 30 °C for 120 days. Sampling was carried out every 24 hours (h) to perform the corresponding chemical, physic and enzymatic analyses.
Cellulolytic Capacity of T. viride M5-2 in Solid State Fermentation Process
Enzyme Extract 5 g of the fermented substrates were taken every 24 h during 120 h, 45 mL of sterile distilled water was added, and the suspensions were stirred for 30 min. The suspensions were filtered and then centrifuged at 4000 g for 20 min. The enzyme extract obtained (E.E) was immediately used for the cellulolytic assay and pH measurement; the unused residue was stored at -20 °C. Analyses were performed in triplicate.
Enzyme Activities
The enzymatic activities (EA) of endo β1-4 glucanase (CMCase) and exo β1-4 glucanase (PFase) were determined, calculated, and expressed in international units per milliliter (IU/mL), which refer to the micromoles of glucose released per minute of reaction under the activity assay conditions [19].
CMCase Enzyme Activity
0.5 mL of (EA) was incubated in a 0.5 mL mixture of 1% carboxymethylcellulose (0.5 viscosity), 0.075 M sodium citrate buffer (pH=4.8), and 3,5-dinitrosalicylic acid (DNS) at 50°C for 30 min. The reducing sugars released were determined after incubation at 100 0C for 5 min and the reading was taken at λ=540nm.
PFase Enzyme Activity
0.5 mL of the PFase enzyme (EE) was added to a mixture of 0.075 M sodium acetate buffer (pH = 4.8) and 3,5-dinitrosalicylic acid (DNS) on Whatman No. 1 filter paper (1 x 6 cm) at 50°C for 60 min. The reducing sugars released were determined after incubation at 100°C for 5 min, and the reading was taken at λ = 540 nm.
Productivity of Cellulolytic Enzymes in SSF
The productivity of the cellulolytic complex was calculated based on a moisture content of 70% and the initial dry matter (5 g of each legume whole forage flours meals) during the 120 h SSF process with the fungus T. viride M5-2.
Chemical Analysis of Solid State Fermentation Process
To determine the chemical composition of the fiber, 5 g of solid material were taken at 0, 24, 48 and 72 h where the indicators studied were: dry matter (DM) and crude protein (CP) according to AOAC [20], true protein (PV) by the Berstein method cited by Scull [21], neutral detergent fiber (NDF), acid detergent fiber (ADF), Hemicellulose (Hem), cellular content (CC), lignin (Lig) and cellulose (Cel), according to Van Soest [22].
Phisical Analysis of Solid State Fermentation Process
Physical properties were determined according to the techniques compiled by Savón et al. [23]. To determine packing volume and solubility, the samples were passed through a 0.8 mm sieve and placed in an oven at 105°C overnight.
Determination of Packing Volume
One gram of sample was placed in a graduated centrifuge tube. Centrifugation was then carried out at 3000 rpm for 20 min. Finally, the volume (V %) occupied by the sample after centrifugation
Determination of Solubility
60 mL of distilled water was added to two grams of the initially dried sample and left to stand for one hour. The samples were then filtered and dried in an oven at 60°C for 12 h. The difference between the dry sample weight at the beginning and end of the analysis represented the percentage of the fraction that was solubilized in water.
The Water Adsorption Capacity (WAC)
The water adsorption capacity was determined by the gravimetric method using the following formula: WAC (g/g) = (wet sample weight – dry sample weight / dry sample weight) and the acid buffering capacity (ABC) and basic buffering capacity (BBC) were determined using the method. [23]
Determination of T. viride M5-2 Structural Changes of the Whole Forage Flours Meals from the Legumes L. purpureus and M. pruriens, by Fourier Transform Infrared Spectroscopy ATR-FT-IR Analysis
The infrared (IR) spectrum was measured on an ATI Mattson Genesis Series FT-IR spectrometer with a special attachment for solid samples, set to attenuated total reflectance (ATR). The infrared spectrum was recorded in the range of 4000 to 600 cm-1 at 72h after the start of the fermentation process. Background interference from moisture and CO2 was instantly eliminated during the scanning process. Each analysis was performed in quintuplicate and the average value was taken as the final spectrum.
Statistical Analysis
A completely randomized analysis of variance (ANOVA) was performed using a factorial design (2x4) (two whole forage flours meals L. purpureus and M. pruriens and four sampling times: 0, 24, 48 and 72 h, for the chemical and physical composition of the fiber, DM and pH) with three replications. Duncan’s test was applied for p < 0.05. Main effects were reported in cases where the interaction was not significant. CMCase and PFase enzyme activities were analyzed in a factorial arrangement (two whole forage flours meals L. purpureus and M. pruriens and six sampling times: 0, 24, 48, 72, 96, 120 h) until the end of fermentation, where each Erlenmeyer flask constituted an experimental unit. The data were analyzed using the statistical package InfoStat. [24]. Differences between means were established according to the Duncan test [25].
3. Results
Growth of the Lignocellulolytic Fungus T. viride M5-2 on Whole Forage Flours Meals of L. purpureus and M. pruriens
It is known that fungi can present different adaptations depending on nutritional conditions, which directly affects their development [26]. However, the cultural characteristics of of T. viride M5-2 growth in whole forage flours meals of M. pruriens and L. purpureus showed that, after 24 hours, it developed a rapid growth (the mycelium was whitish at the beginning and, after 72 hours, a complete sporulation of deep green color was observed) in the two substrates independently of the nutritional conditions of these legumes. These results were achieved under optimal conditions of temperature and humidity. Late sporulation is an important characteristic for greater effectiveness of the degradative potential of the fungus. For this reason, the present research justifies the study with this fungus to determine the specificity of the enzymatic action in the expression of the cellulase complex through SSF over time and to obtain greater action in the cell walls of the fibrous material.
Figure 1.
Growth of the fungus T. viride M5-2 on whole forage meals of legumes (72 hours, temperature 30±1oC, 70% relative humidity). M. pruriens (left), L. purpureus (right).
Figure 1.
Growth of the fungus T. viride M5-2 on whole forage meals of legumes (72 hours, temperature 30±1oC, 70% relative humidity). M. pruriens (left), L. purpureus (right).

Cellulolytic Capacity of T. viride M5-2 in the Degradation of Legumes in the Solid State Fermentation Process (SSF)
The cellulolytic capacity of T. viride M5-2 to produce the main enzymes of the cellulase complex (endo-1,4-β-glucanase and exo-1,4-β-glucanase, respectively) during the fermentation of whole forage flours meals (L. purpureum and M. pruriens) is shown in Table 1 and Table 2. An interaction between the factors studied was detected for the activity of both enzymes (p<0.001). In the case of endo β1-4 glucanase activity (Table 1) the values are higher when the microorganism is degrading M. pruriens and a decrease in this activity was observed at 48 h. However, in L. purpureus it decreased at 24 h. In both cases they maintain cellulolytic activity up to 120 h. On the other hand, exo β1-4 glucanase activity (Table 2) showed a maximum potential for the cellulolytic production of this enzyme when the fungus is degrading L. purpureus, although in 24 hours the increase was greater in M. pruriens, with a decrease of 0.2 percentage units with respect to the other legume.These differences in fiber degradation could be due to the fact that these cellulolytic enzymes are a complex that catalyzes the progressive conversion of cellulose and guarantees the efficiency of biotransformation in a synergistic way [27], also considering the differences in the physical-chemical characteristics of the flours used in the research [21].
The productivity calculated from the expression of the main cellulolytic enzymes showed the dependence of the bioconversion of the degraded substrate on the dry matter during the fermentation dynamics, an aspect that is visualized in the range of 24 to 72 h, with the maximum degradation not coinciding with the maximum productivity. An aspect to be taken into account for the design of SSF strategies with whole forage flours meals of legumes.
As shown in the results in Table 3, during fermentation (SSF) of both whole forage flours meals, the initial pH was 6 and slightly increased over the course of fermentation (p<0.001). The lower pH values obtained in M. pruriens compared to L. purpureus are close to the optimum for cellulase activity, which could explain the higher CMCase activity in the degradation of M. pruriens forage meal. These differences show that there are changes in the levels of hydrolytic activity in the growth activity of this fungus in these legumes and that many factors may influence it, but greater stability of fermentation was observed in M. pruriens. However, the maximum values differed in time due to variations in enzyme production during carbohydrate degradation of plant fiber with fungal growth. Under these conditions it is evident that cellulolysis depends on the proportion of the enzymatic components, the nature of the fiber, the affinity for it, as well as the stereospecificity and concentration of the enzyme used [28].
Chemical Analysis of Solid State Fermentation Process by T viride M5-2
Solid-state fermentation of the whole forage flours meals of legumes L. purpureus and M. pruriens as can be observed in (Table 4), the fermentation process of the whole forage flours with T. viride M5-2 achieved crude protein values of 20% in M. pruriens and approximately 18% in L. purpureus. True protein showed a similar behavior with values of 17% for M. pruriens and 12% for L. purpureus. The most relevant aspect is that protein increased by 2 and 3 percentage units for CP and TP respectively (p<0.001), which coincided with the maximum cellulolytic activity values. Variations in dry matter during fermentation can also be observed. These variations could be the result of the volatilization of solid components, together with the production of water and CO2 as a result of the metabolism of the fungus and the beginning of the degradation of the most complex components of the plant cell wall such as cellulose and hemicellulose.
On the other hand, the results of the fiber fractionation analysis of the whole forage flours meals are shown. An interaction was found between fermentation time (h) and of the whole forage flours meals (L. purpureus and M. pruriens) for all indicators, which were expressed as a percentage of dry matter. As can be seen in the table, the main component of the meals is insoluble fiber. The concentrations of NDF, ADF, and cellulose in the forages showed differences between species, with M. pruriens presenting the highest NDF value. The lowest NDF content was observed at 48 h in the L. purpureus forage meal and at 24 h for the M. pruriens forage meal. This indicator decreased by 4.18 percentage units in M. pruriens and presented a cellular content of 31.52%. However, in L. purpureus it is only 1.09 percentage units lower than the initial value and the cellular content of this fraction is 33.26%. With respect to hemicellulose, this fraction decreases due to the cellulolytic activity of the fungus on the plant fiber, but a higher percentage of enzymatic hydrolysis is observed in M. pruriens compared to the control.
The ADF did not vary in the first 48 h for the forage meal of L. purpureus and M. pruriens respectively, then showed a relative increase (p <0.001) in both complete forage meals. In the most notable decreases in the NDF and ADF fractions, it could be determined that the hemicellulose polysaccharide present in the cell wall of the plant fibers under study was on the order of 15.94% and 13.77% in 24 h of degradation for M. pruriens and 48 h for L. purpureus respectively. Using the ADF, the cellulose and lignin contents were determined during the fermentation of whole-wheat flour from the legumes studied. The results showed a decrease in cellulose of 2.19 percentage units for L. purpureus after 24 h of degradation, while in M. pruriens it reached a decrease of only 1.29 percentage units over a longer period (48 h). Differences were observed with the NDF, which did not coincide with the moments of maximum fungal activity. Regarding the values obtained for lignin, the results showed that it was concentrated during the fermentation of L. purpureus; however, for M. pruriens, it decreased 24 h after the start of the process by 1.7 percentage units.
The analysis of the physical properties showed differences between the two whole forage flours meals with respect to fermentation time in four of the five indicators analyzed (Table 5). Overall, they presented low physical composition values despite the high fiber and protein content of these legumes. However, M. pruriens showed a greater water retention capacity in relation to its fiber volume. The results suggest greater acid buffering capacity than base buffering in both legumes, which favors the quality of the fiber for the introduction of unconventional feeds into animal diets. Of note is the low solubility of both legumes (Table 6), which did not differ between treatments. However, they did differ with respect to the enzymatic biodegradation time of the fungus, with lower solubility at 72 h. This coincides with the lignin concentration found. This aspect is also related to the differences between the polymers that make up the cell wall of the legume fiber studied, which allow water retention.
Determination of T. viride M5-2 Molecular Changes of Whole Forage Flours Meals from the Legumes L. purpureus and M. pruriens, by Fourier Transform Infrared Spectroscopy (ATR-FT-IR) Analysis
The ATR-FT-IR spectroscopic analysis (Figure 2 and Figure 3) shows the first spectra corresponding to whole forage flours meals from the legumes L. purpureus and M. pruriens. It can be observed that there is degradation of structures belonging to the carbon skeleton of the cellulose fibers and lesser signals corresponding to the lignin in the fermented samples. The spectra of the forage meals generally present well-defined bands with good resolution. In the range of 1600-1400 cm⁻¹, the bands are more expanded in the biodegraded meals. However, between 1445–1390 cm⁻¹ the bands of bond formation in the OH plane of carbohydrates are of low intensity. The observed structural modification shows a characteristic band of the carbonyl groups present in amides 1636–1449 cm⁻¹. This intense valence vibration band of the carbonyl group C=O γ C=O appears slightly split in two, suggesting the presence of amino acids or proteins in the samples. Furthermore, this band is so intense that it is unlikely to be associated with another bond, and its presence can be explained by the possible degradation of polysaccharides from the legume wall.
When comparing spectra of the whole forage flours meals respect to zero hour of fermentation of each one, a lower intensity of the vibration bands of benzene (C=CH) is observed in the range 670±20 cm-1 and a new band also appears with intensity at 774.30±20 cm-1 where the aromatic rings mono, di and tri-substituted by OH groups or methyl groups derived from benzene are found, which implies hydrolytic action of the fungus on the fiber of the legumes under study. In addition, an intense valence vibration band of the O-H γ OH bond can be observed in the 3291-3283 cm-1 range. This band is a product of the polymeric association of the OH groups of carbohydrates. In the spectral area from 2852 to 2920 cm-1, bands corresponding to CH2 and CH3 vibrations of carbonyl chains of organic compounds can be seen. The band is of medium intensity, according to the valence vibration of the C-H γ CH bond of alkyl groups in the same carbohydrate chain. Around 1730 cm-1, bands characteristic of ester-type compounds are observed. Around 1028-1051 cm-1, an intense band characteristic of C-O type vibrations is found. The bands corresponding to carbohydrates and aliphatic ethers are observed in all spectra; however, in the fermented samples, they are wider and more intense. This could be related to the hydrolytic action of the fungus on the carbohydrates present in the legumes, as cellulose decreases with fermentation time. These data obtained from the ATR-FT-IR spectra correspond to those obtained in the chemical composition analysis when observing the degradation of NDF in the range of 24-72 h compared to the control based on dry matter, suggesting a degradation of the fiber caused by the cellulolytic action in these whole forage flours meals legume.
4. Discussion
Cellulolytic enzymes are a complex that catalyzes the progressive conversion of cellulose. The catalytic mechanisms of this enzymatic complex develop synergistically, ensuring the efficiency of bioconversion. Cellulolytic fungi such as T. viride M5-2 produce these enzymes under conditions of deficit of alternative carbon sources with greater degradability and absorption and of other nitrogen sources also rich in metabolizable energy and suppliers of carbon chains. This characteristic is due to the strict control by catabolic repression to which the cellulase genes are subject [29]. Under such conditions, these microorganisms use enzymes to release simple sugars from the solid substrate and use them as a carbon source [30].
As shown in the results, the T. viride M 5-2 strain has the ability to grow on whole forage meals from the legumes L. purpureus and M. pruriens, without the addition of nutrients.
The quantitative evaluation of the activity of the main enzymes of the cellulase complex (endo-1,4-β-glucanase and exo-1,4-β-glucanase), together with the productivity of the enzymatic system during fermentation, the degradation of plant fiber and its structural changes observed by infrared spectroscopy, as well as the increase in protein, demonstrated that this fungus, through a simple biotechnological process with SSF, was able to modify the nutritional composition of legume meals. According to this research, there are several factors that could have affected to some degree the degradation of whole legume flours, such as: nitrogen concentration, enzyme inhibition by reaction products, adsorption of the enzyme to the substrate and pH [31,32], the latter reaching values higher than 7 after 48 h of fermentation, even so, the cellulolytic activity was not affected in either of the two flours.
This relationship between pH and activity of the cellulase complex depends on the acid-base behavior of the medium and the nature and concentration of the substrate [33,34,35]. It is known that the initial optimum pH for the hydrolytic action of cellulases is between 5 and 6 [36] depending on the substrate to be fermented and the growth temperature of each microorganism. However, Valiño et al. [15] demonstrated that the T. viride M5-2 strain is resistant to catabolic repression and exhibits hydrolytic activity at pH=7. Therefore, despite the basic pH levels during the fermentation of both whole legume flours, this allowed for enzymatic activity, thereby increasing the nutritional value of the resulting product. It is also known that plant proteins are sensitive to pH, and that their modification through this pathway can improve their functionalities, as well as through hydrolysis induced by laccase and protease enzymes [37] in research on oxidative protein polymerization.
On the other hand, knowledge of the chemical composition of unconventional raw materials allows for estimating their nutritional quality and performing specific evaluations for their use in animal feed [38]. That is why fungal fermentation results in the degradation of plant food carbohydrates into simpler sugars and ensures the biosynthesis of various metabolites with nutritional changes in fiber [39]. Therefore, the comprehensive characterization of the nutritional value of whole forage flours meals meals from these legumes fermented with T. viride M5-2 offers a novel aspect for research in digestive physiology of monogastric animals.
According to Shubha et al. [40] and Sowdhanyaa et al. [41] they stated that the protein nitrogen content of legumes accounts for 75-85% of the total nitrogen in the plant. With this feature, it was possible to perform SSF without the need to add urea or other nitrogen source, although the nitrogen concentration of whole forage flours meals from the legumes could influence the enzyme activity variably. Furthermore, fermentation processes of these legumes have been shown to produce a number of positive changes in nutrient content, such as increased essential amino acids, soluble proteins, and in vitro protein digestibility [2,42,43].
On the other hand, the fiber content of whole legume forage meals affects their nutritional quality by negatively influencing the digestibility of the nutrients that make up the feed.44 Savón et al. [45] evaluated a group of legumes and other tree forage plants and suggested that the high fiber content of these legumes could be due to the complexes formed between carbohydrates and phenolic compounds.
In this experiment, the differences in the percentages of NDF and cellulose degradation by the fungus in both forages are related to the cellulolytic activity observed during SSF with T. viride M5-2. This fungus, in addition to utilizing easily degradable wall components, such as hemicellulose, was also able to utilize some of the nitrogen associated with the fiber as a nutrient thanks to its rapid growth and short adaptation period before beginning to degrade lignin. This demonstrated its valuable potential for the bioconversion of whole forage meals from the legumes used. However, Vázquez et al. [46] report that greater modifications in plant fiber can be obtained by combining cellulolytic and ligninolytic enzymes.
The observed lignin concentration was due to two possible causes: one, the physical barrier of this cell wall component and the other, to the chemical analysis of the fiber that could not be standardized in the SSF, mainly for whole forage flours meals from fungal delignified forage. The nature and chemical composition of the plant material to be fermented and the results that may arise should also be taken into account [47]. From the results obtained, an apparent concentration of ADF is observed, which could be due to the reduction of dry matter in both whole grain flours. However, comparing them with other results makes their interpretation difficult since the dry matter losses during this process are not reported. This is aggravated in the determination of lignin, since its value could be overestimated mainly in M. pruriens, due to the presence of antinutritional factors such as tannins, of a chemical nature similar to lignin [48,49] and phytates and trypsin inhibitors that can have harmful effects on the animals that consume those [50].
For this reason, legume forage meals [51] and their seeds are consumed not only for their excellent nutritional properties, [52] but their quality must also be taken into account, which are modified by their physical properties, such as fiber volume, solubility, water adsorption capacity and acid-base buffering capacity, which also influence biological processes such as digestion and nutrient consumption [53].
The potential for greater water absorption favors fiber, due to the moisture content reached, which facilitated hydrolysis by cellulase enzymes in the matrix of polysaccharides, due to the greater hygroscopic power of hemicellulose compared to cellulose and despite the hydrophobic nature of lignin. This explains the behavior of this indicator in whole forage flours meals. This process can occur not only during SSF with the fungus T. viride M5-2, but also due to the acidic buffering capacity of the legume flours studied for pH regulation in the gastrointestinal tract (GIT) of monogastric species. This is of great importance, since during digestion (GIT) in monogastric animals, a pH change occurs, from very acidic (pH 1) to almost neutral (pH 6.8-7.2) [54].
The formation of alcoholic, carboxylic, and carbonyl groups derived from the biodegradation of carbohydrates, as well as some groups formed by the oxidation of aromatic structures, confirm the molecular changes in the cell wall of legume flours by the fungus T. viride M5-2, capable of expressing specific enzymes responsible for degrading fiber components. Studies can be mentioned where the intensity of lignin and cellulose signals decreased with another ascomycete, Curvularia kusanoi L7 [55] corroborating previous approaches suggesting the expression of enzymes that modify lignin and cellulose and allow for more thorough degradation of the plant cell wall. This result provides useful information for fine-tuning solid-state fermentation processes of whole legume forage meals, enabling their efficient use in animal feed.
5. Conclusions
It can be concluded that the mutant strain T. viride M5-2 produced chemical, physical, and molecular changes in the fibrous and protein fractions of L. purpureus and M. pruriens through SSF, which improved their nutritional value as an alternative feed source for animal nutrition. The valorization of this type of source significantly contributes to improving bioabsorption in animal nutrition, reducing competition with human food, and preserving the environment. Therefore, the conversion of fibrous biomass by cellulolytic fungi develops more efficient, sustainable, and environmentally friendly agricultural production.
Author Contributions
Conceptualization and design of the study; E.V.C, L.R.S. Investigation, L.R.S., E.V.C, L. S. V., M. A.V., J. C. D. M., Y.G.H. Methodology, E.V.C and L. S. V; Data curation, L.R.S, L. S. V, E.V.C, M. A.V., Formal analysis L. S. V., E.V.C, M. A.V., J. C. D. M., Y.G.H. L.R.S, Supervision, E.V.C, M. A.V., Writing—original draft preparation, E.V.C, M. A.V., L.R.S.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The microorganism sequences were deposited to GenBank under accession number KY977981.
Acknowledgments
Special thanks are extended to Nereyda Albelo Dorta of the Institute of Animal Science for the maintaining of the strains and the execution of the experiments
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ikusika, O.O.; Akinmoladun,O.F.; Mpendulo, C.T. Enhancement of the Nutritional Composition and Antioxidant Activities of FruitPomaces and Agro-Industrial Byproducts through Solid-State Fermentation for Livestock Nutrition: A Review. Fermentation. 2024, 10, 227. [CrossRef]
- Savón, L. ; Scull, I. ; Dihigo, L. E. ; Martínez, M. ; Albert, A. ; Leiva, L. Tropical forage meals: an alternative for sustainable monogastric species production. Multifunctional grasslands in a changing world. 2008, 2, 487-497.
- Ezegbe, C.C.; Nwosu, J. N.; Owuamanam, C.I.; Victor-Aduloju, T.A.; Nkhata, S.G. Proximate composition and anti-nutritional factors in Mucuna pruriens (velvet bean) seed flour as affected by several processing methods. 2023. Heliyon, 9 No. 8. [CrossRef]
- Yang, C.; Yoo, J.; Zhang, T.; Yang, K.; Guo, J.; Pan, S. Mixed fermentationof navel orange peel by Trichoderma viride and Aspergillus niger: Effects on the structural and functional properties of soluble dietary fiber. Food Bioscience. 2024, 57, No. 103545. [CrossRef]
- McConnell, L. L. ; Osorio, C. ; Hofmann, T. The future of agriculture and food: sustainable approaches to achieve zero hunger. Journal of Agricultural and Food Chemistry. 2023, 71, No. 36, 13165-13167. [CrossRef]
- Pandey, D. K.; Singh, S.; Kumar, S. D.; Sing, T. M.; Dixit, S.; Sawargaonkar, G. Nutrient profiling of lablab bean (Lablab purpureus) from northeastern India: A potential legume for plant-based meat alternatives. Journal of Food Composition and Analysis. 2023, 119, No. 105252. [CrossRef]
- Yoon, L.W.; Ang, T.N.; Ngoh, G.C.; Seak, A.M.C. Fungal solid-state fermentationand various methods of enhancement in cellulase production. Biomass and Bioenergy. 2014, 67, 319-338. [CrossRef]
- Malgas, S.; Thorensen, M.; van Dyk, J.S.; Pletschke, B.I. Time dependence of enzyme synergism during the degradation of model and natural lignocellulosic substrates. Enzyme and Microbial Technology. 2017, 103, 1-11. [CrossRef]
- Cebrián, M.; Ibarruri, J. Filamentous Fungi Biorefinery. Filamentous fungi processing by solid-state fermentation. Current Developments in Biotechnology and Bioengineering. 2023, 251-292.
- Hernández, D.J.M; Ferrera, C.R.; Alarcón, A. Trichoderma: importancia agrícola, biotecnológica, y sistemas de fermentación para producir biomasa y enzimas de interés industrial. Chilean Journal of Agricultural ; Animal Sciences, ex Agro-Ciencia. 2019, 35, No. 1, 98-112.
- Plouhinec, L.; Neugnot, V.; Lafond, M.; Berrin, J.G. Carbohydrate-active enzymes in animal feed. Biotechnology Advances. 2023, 65, No. 10814. [CrossRef]
- Bulgari, D.; Alias, C.; Peron, G.; Ribaudo, G.; Gianoncelli, A.; Savino, S.; Gobbi, E. Solid-state fermentation of Trichoderma spp.: A new way to valorize the agricultural digestate and produce value-added bioproducts. Journal of Agricultural and Food Chemistry. 2023, 71, No. 9, 3994-4004. [CrossRef]
- De França, P. D.; Pereira, Jr.N.; de Castro, A.M. A. Comparative review of recent advances in cellulases production by Aspergillus, Penicillium and Trichoderma strains and their use for lignocellulose deconstruction. Curr. Opin. Green Sustain. Chem. 2018, 14, 60–66.
- Valiño, E.C.; Dustet, J.C.; Pérez, H.; Brandão, L.R.; Rosa, A.C.; Scull, I. Transformation of Mucuna pruriens with cellulolytics fungi strains as functional food”. Academia Journal of Microbiology Research. 2016, 4, No. 4, 62–71.
- Valiño, E.C., Elías, A., Rodríguez, M.; Albelo, N. Evaluation using Fourier Transformed-infrared spectroscopy (FT-IR) of fermentationby the strain T. viride M5-2 from the cell walls of sugarcane (Saccharum officinarum Lin) bagasse pretreated International Journal of Chemical and Biomolecular Science. 2015b,1, No. 3, 134-140.
- Valiño, E.C.; Savón, L.; Elías, A.; Rodríguez, M.; Albelo, N. Nutritive value improvement of seasonal legumes Vigna unguiculata, Canavalia ensiformis, M.pruriens, Lablab purpureus, through processing their grains with Trichoderma viride M5-2. Cuban Journal of Agricultural Science. 2015a, 49, No. 1, 81-89.
- Wang, L.Y.; Cheng, G.N. ; May, A,S. Fungal solid-state fermentationand various methods of enhancement in cellulases production. Biomass Bioenergy. 2014, 67, 319-338.
- Sosa, A.; González, N.; García, Y.; Marrero, Y.; Valiño, E.; Galindo, J.; Sosa, D. ;Alberto, M.; Roque, D.; Albelo, N.; Colomina, L.; Moreira, O. Collection of microorganisms with potential as additives for animal nutrition at the Institute of Animal Science. Cuban Journal of Agricultural Science. 2018, 51, No. 3, 311-319.
- Mandels, M.; Medeiros, J. E.; Andreotti, R.E.; Bissett, F.H. Enzymatic hydrolysis of cellulose: evaluation of cellulase culture filtrates under use conditions. Biotechnology and bioengineering. 1981, 23, No. 9, 2009-2026. [CrossRef]
- AOAC. Official Methods of Analysis of AOAC International. Chapter 4. Animal Feed. Dr. George Latimer Jr. Editor, 21st ed; AOAC International, Gaithersburg, USA. 2019.
- Scull, I.R.; Savón, L.V.; Spengler, I.S.; Herrera, M.V.; González, V.L. Potentiality of the forage meal of Stizolobium niveum and Stizolobium aterrimum as a nutraceutical for animal feeding. Cuban Journal of Agricultural Science. 2018, 52, No. 2, 223-234.
- Van Soest, P.; Robertson, J. B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and non-starch polysaccharides in relation to animal nutrition. Journal of Dairy Science. 1991, 74, No.10, 3583–3597.
- Savón, L.; Scull, I.; Orta, M.; Martínez, M. Whole-grain foliage flours from three tropical legumes for poultry feed. Chemical composition, physical properties, and phytochemical screening. Cuban Journal of Agricultural Science. 2007, 41 No. 4, 359-361.
- Di Rienzo, J.; Casanoves, F.; Balzarini, M.; González, L.; Tablada, M.; Robledo, Y.C. InfoStat versión 2017. Grupo InfoStat, FCA, Universidad Nacional de Córdoba: Argentina. 2017.
- Duncan, B. Multiple ranges and multiple F test. Biometrics. 1955, 11, No. 3, 1-42.
- Matas Baca, M. Á.; Flores-Córdova, M. A.; Pérez Álvarez, S.; Rodríguez Roque, M.J.; Salas, Salazar; N.A.; Soto, Caballero, M.C.; Sánchez, Chávez, E. Trichoderma fungi as an agricultural biological control in México. Revista Chapingo. Serie horticultura. 2023, 29, No. 3, 79-114.
- Bézier, S.; Stiegler, M.; Hitzenhammer, E.; Schmoll, M. Screening for genes involved in cellulase regulation by expression under the control of a novel constitutive promoter in Trichoderma reesei. Current Research in Biotechnology. 2022, 4, 238–246. [CrossRef]
- Bamidele, M. O.; Bamikale, M. B.; Cárdenas-Hernández, E.; Bamidele, M. A.; Castillo-Olvera, G.; Sandoval-Cortes, J.; Aguilar, C. N. Bioengineering in Solid-State Fermentation for next sustainable food bioprocessing. Next Sustainability. 2025, 6, No. 100105. [CrossRef]
- Beier, S.; Stiegler, M.; Hitzenhammer, E.; Schmoll, M. Screening for genes involved in cellulase regulation by expression under the control of a novel constitutive promoter in Trichoderma reesei. Current Research in Biotechnology. 2022, 4, 238–246. [CrossRef]
- Singh, A.; Bajarb, S.; Devia, A.; Pantc, D. An overview on the recent developments in fungal cellulase production and their industrial applications Bioresource Technology Reports journal Bioresource Technology. 2021, 14, No. 100652.
- Bhandari, S.; Pandey, K.R.; Joshi, Y.R.; Lamichhane, S. K. An overview of multifaceted role of Trichoderma spp. for sustainable agriculture. Archives of Agriculture and Environmental Science. 2021, 6, No. 1, 72-79. [CrossRef]
- Zhao, C.; Deng, L.; Fang, H. Mixed culture of recombinant Trichoderma reesei and Aspergillus niger for cellulase production to increase the cellulose degrading capability. Biomass and Bioenergy. 2018, 112, 93–98. [CrossRef]
- Vázquez, M. A.; Cabrera, E.C.V.; Aceves, M.A.; Mallol, J.L.F. Cellulolytic and ligninolytic potential of new strains of fungi for the conversion of fibrous substrates. Biotechnology Research and Innovation. 2019, 3, No.1,177-186. [CrossRef]
- Liu, H.; Sun, J.; Leu, S.Y.; Chen, S. Toward a fundamental understanding of cellulase-lignin interactions in the whole slurry enzymatic saccharification process. Biofuels Bioproduct and Biorefinering. 2016, 10, No. 5, 648-663. [CrossRef]
- Dustet, J.C.; Izquierdo, E. Aplicación de balances de masa y energía al proceso de fermentación en estado sólido de bagazo de caña de azúcar con Aspergillus niger. Biotecnología Aplicada. 2004, 21, 85-91.
- Ghose, T. K. Measurement of Cellulase Activities. Pure Appl. Chem. 1987, 59, 257−268.
- Yi, J.; Chen, X.; Wen, Z. ; Fan, Y. Improving the functionality of pea protein with laccase-catalyzed crosslinking mediated by chlorogenic acid. Food chemistry. 2024, 433, No. 137344. [CrossRef]
- Martínez, M.; Sarmiento, L.; Santos, R. H.; Villafranca, M.; Londres, S. Digestive and carcass indicators of Rhode Island Red chickens, which intake rocessed Mucuna pruriens, in two rearing systems. Technical note. Cuban Journal of Agricultural Science. 2022, 56, No. 2, 121-126.
- Alberto, Vazquez, M.; Saa, L.R., Valiño, E.; Torta, L.; Laudicina, V.A. Microbiological Aspects and Enzymatic Characterization of Curvularia kusanoi L7: Ascomycete with Great Biomass Degradation Potentialities. Journal of Fungi. 2024, 10, No. 12, 807.
- Jung, N.; Meyer, A.S. Solid-State Fungal Fermentation for Better Plant Foods. Food Science and Nutrition Cases. 2025, fsncases, No. 20250001. [CrossRef]
- Shubha, K.; Choudhary, A. K.; Mukherjee, A.; Kumar, S.; Saurabh, K.; Kumar, R.; Das, A.A. Chemometric study comparing nutritional profiles and functional attributes of two botanical forms of Lablab Bean (Lablab purpureus (L.) Sweet). South African Journal of Botany. 2024, 173, 320-329. [CrossRef]
- Sowdhanyaa, D.; Singha, J.; Rasanea, P.; Kaura, S.; Kaura, J.; Ercislib, S.; Vermac, H. Nutritional significance of velvet bean (Mucuna pruriens) and opportunities for its processing into value-added products. Journal of Agriculture and Food Research. 2024, 15, No. 100921.
- Díaz, M.; Martín-Cabrejas, M.Á.; González, A.; Torres, V.; Noda, A. Biotransformation of Vigna unguiculata during the germination process. Cuban Journal of Agricultural Science. 2007, 41, No. 2,161.
- Williams, B.A.; Mikkelsen, D.; Flanagan, B.M. “Dietary fibre”: moving beyond the “soluble/insoluble” classification for monogastric nutrition, with an emphasis on humans and pigs. J Animal Sci Biotechno. 2019, 10, No. 45, 2-12.
- Savón, L.; Scull, I., Orta, M.; Torres, V. Physicochemical characterization of the fibrous fraction of five tropical foliage meals for monogastric species. Cuban Journal of Agricultural Science. 2004, 38, No.3, 281-286.
- Vázquez, M.A.; Valiño, E.C.; Torta, L.; Laudicina, A.; Sardina, M.T.; Mirabile, G. Potencialidades del consorcio microbiano Curvularia kusanoi -Trichoderma pleuroticola como pretratamiento biológico para la degradación de fuentes fibrosas. Rev MVZ Córdoba. 2022, 27, No. 2, 2559.
- Valiño, E.C.; Alberto, M.; Dustet, J. C ; Albelo, N. Production of lignocellulases enzymes from Trichoderma viride M5-2 in wheat bran (Triticum aestivum) and purification of their laccases. Cuban Journal of Agricultural Science. 2020, 54, No. 1, 55-59.
- García, Y.; Ibarra, A.; Valiño, E. C.; Dustet, J.; Oramas, A.; Albelo, N. Study of a solid fermentationsystem with agitation in the Biotransformation of sugarcane bagasse by the Trichoderma viride strain M5-2. Cuban Journal of Agricultural Science. 2002, 36, No. 3, 265-270.
- Sadh, P. K.; Saharan, P.; Duhan, J. S. Bio-augmentation of antioxidants and phenolic content of Lablab purpureus by solid state fermentation with GRAS filamentous fungi. Resource-Efficient Technologies. 2017, 3, No. 3, 285-292.
- Murphy, A. M.; Colucci, P.E. A tropical forage solution to poor quality diets: A review of Lablab purpureus. Livestock Research for Rural Development. 1999, 11, No. 2, 1999.
- Guo, Q.; Luo, J.; Zhang, X.; Zhi, J.; Yin, Z.; Zhang, J.; Zhang, B.; Chen, L. A Comprehensive Review of the Chemical Constituents and Functional Properties of Adzuki Beans (Vigna angulariz). Journal of Agricultural and Food Chemistry. 2025, 73, No. 11, 6361- 6384. [CrossRef]
- Hettiarachchi, H.A.C.O.; Gunathilake, K.D.P.P. Physicochemical and functional properties of seed flours obtained from germinated and non-germinated Canavalia gladiata and Mucuna pruriens. Heliyon. 2023, 9, No.19653. [CrossRef]
- Alcívar, J.L.; Martínez, M.P.; Lezcano, P. ; Scull, I.; Valverde, A. Technical note on physical-chemical composition of Sacha inchi (Plukenetia volubilis) cake Cuban Journal of Agricultural Science. 2020, 54, No. 1, 19-23.
- Martínez, M.P.; Vives, Y.H.; Rodríguez, B.; Pérez, G.O.A.; Herrera M.V. Nutritional value of palm kernel meal, fruit of the royal palm tree (Roystonea regia), for feeding broilers. Cuban Journal of Agricultural Science, 55th Anniversary. 2021, 55, No. 3, 303-313.
- Alberto, Vazquez, M.; Saa, L. R.; Valiño, E.; Torta, L.; Laudicina, V.A. Microbiological Aspects and Enzymatic Characterization of Curvularia kusanoi L7: Ascomycete with Great Biomass Degradation Potentialities. Journal of Fungi. 2024,10, No. 12, 807-820.
Figure 2.
ATR−FT−IR spectrum of whole forage flours meals from the legume L. purpureus (blue line) after 72 hours of fermentation with the T. viride M5-2 strain (red line). Working range: 4000 to 600 cm⁻¹. Main groups affected by fermentative degradation, unfolded band 3291-3283 cm-1: intense valence vibration of the O-H bond of the phenolic and aliphatic groups, bands 2852 - 2920 cm-1: CH2, CH3 vibrations, 1730 cm-1 ester type bands, band 1636-1449 cm⁻¹ of the carbonyl groups present in the amides, bands 1445-1390 cm⁻¹ of the bond in the OH plane, unfolded band 1028-1051 cm-1 of intense C-O type vibration.
Figure 2.
ATR−FT−IR spectrum of whole forage flours meals from the legume L. purpureus (blue line) after 72 hours of fermentation with the T. viride M5-2 strain (red line). Working range: 4000 to 600 cm⁻¹. Main groups affected by fermentative degradation, unfolded band 3291-3283 cm-1: intense valence vibration of the O-H bond of the phenolic and aliphatic groups, bands 2852 - 2920 cm-1: CH2, CH3 vibrations, 1730 cm-1 ester type bands, band 1636-1449 cm⁻¹ of the carbonyl groups present in the amides, bands 1445-1390 cm⁻¹ of the bond in the OH plane, unfolded band 1028-1051 cm-1 of intense C-O type vibration.
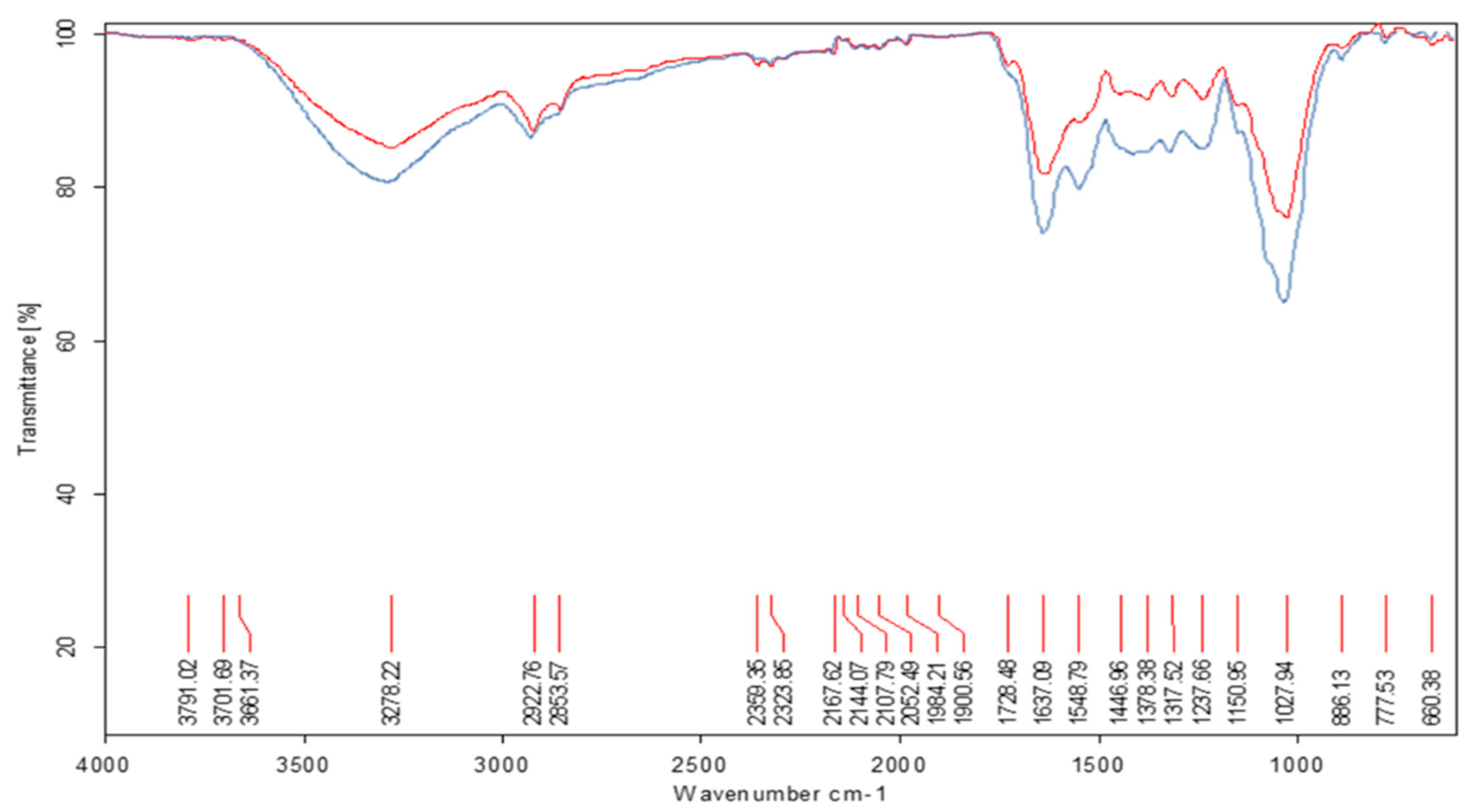
Figure 3.
ATR−FT−IR spectrum of whole forage flours meals from the legume M. pruriens (red line) after 72 hours of fermentation with the T. viride M5-2 strain (blue line). Working range: 4000 to 600 cm⁻¹. Main groups affected by the degradation, band 3291-3283 cm-1: intense valence vibration of the O-H bond of the phenolic and aliphatic groups, bands 2852 - 2920 cm-1: CH2, CH3 vibrations, 1730 cm-1 ester type bands, band 1636-1449 cm⁻¹ of the carbonyl groups present in the amides, bands 1445-1390 cm⁻¹ of the planar OH bond, band 1028-1051 cm-1 of intense C-O type vibration, bands 620-778 cm-1 vibration of the benzene (C=CH).
Figure 3.
ATR−FT−IR spectrum of whole forage flours meals from the legume M. pruriens (red line) after 72 hours of fermentation with the T. viride M5-2 strain (blue line). Working range: 4000 to 600 cm⁻¹. Main groups affected by the degradation, band 3291-3283 cm-1: intense valence vibration of the O-H bond of the phenolic and aliphatic groups, bands 2852 - 2920 cm-1: CH2, CH3 vibrations, 1730 cm-1 ester type bands, band 1636-1449 cm⁻¹ of the carbonyl groups present in the amides, bands 1445-1390 cm⁻¹ of the planar OH bond, band 1028-1051 cm-1 of intense C-O type vibration, bands 620-778 cm-1 vibration of the benzene (C=CH).
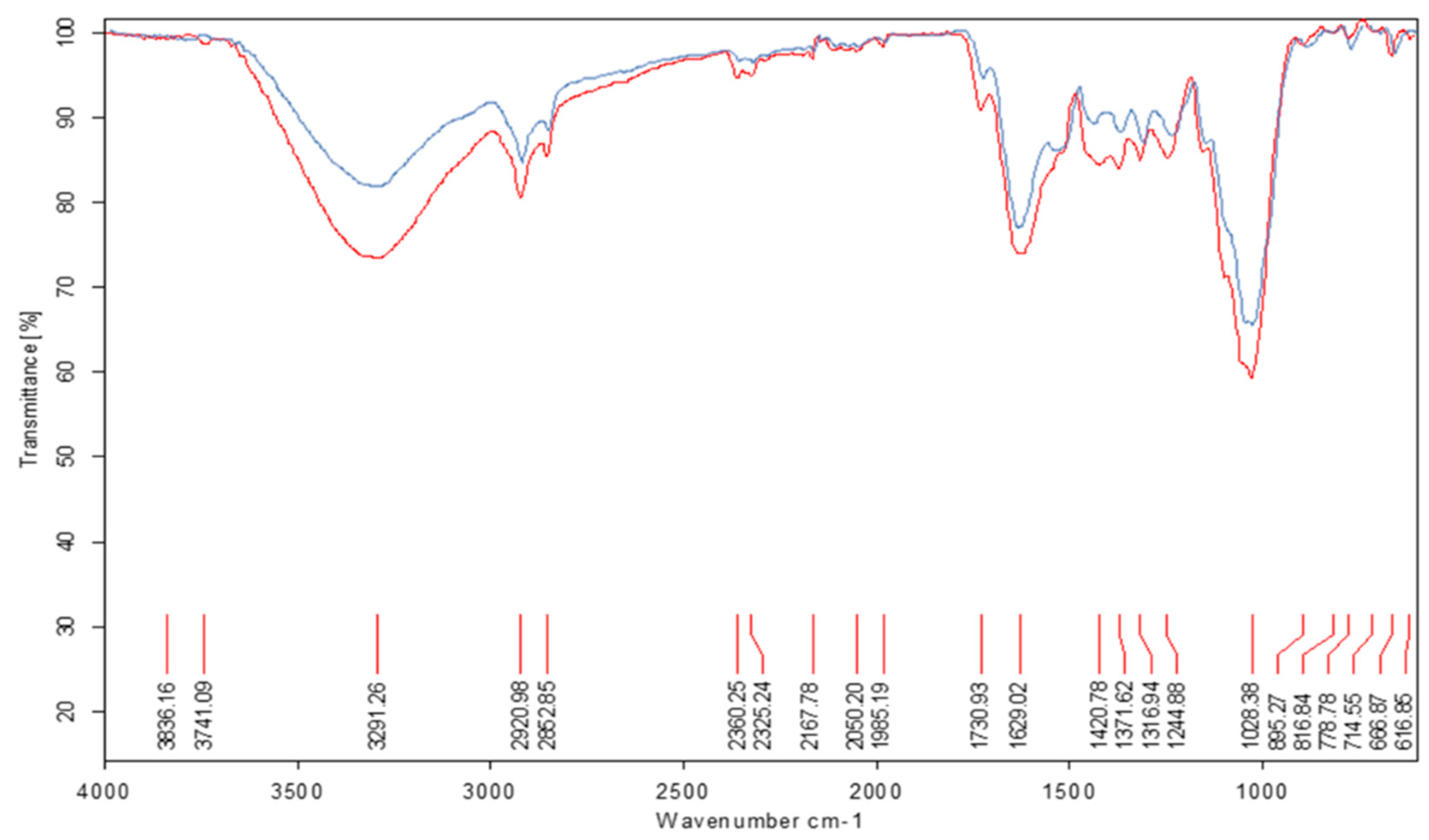

Table 1.
Cellulolytic activity endo β1-4 glucanase (CMCase) of the T. viride M 5-2 strain in the whole forage flours meals of legumes.
Table 1.
Cellulolytic activity endo β1-4 glucanase (CMCase) of the T. viride M 5-2 strain in the whole forage flours meals of legumes.
| Cellulolytic Activity | Legumes | Fermentation Time (h) | SE and p | ||||
| 24 | 48 | 72 | 96 | 120 | |||
| CMCase (IU/mL) |
L. purpureus |
2.09f | 1.18c | 1.22c | 1.01e | 0.99d | ±0.03 p<0,0001 |
|
M. pruriens |
3.15h | 3.29i | 2.57g | 2.01b | 1.90b | ||
| Productivity enzymatic (UI/gMSh) |
L. purpureus |
3.32 | 1.91 | 1.96 | 1.61 | 1.59 | |
|
M. pruriens |
5.36 | 5.46 | 4.26 | 3.32 | 3.15 | ||
a,b,c,d,e,f,g,h,i. Means with different letters in each row differ at p < 0.05 (Duncan, 1955). S.E ± 0.03; p<0.001, ( SE: standard error, p: level of significance).
Table 2.
Cellulolytic activity exo β1-4 glucanase (PFase) of the T. viride M 5-2 strain in the of whole forage flours meals of legumes.
Table 2.
Cellulolytic activity exo β1-4 glucanase (PFase) of the T. viride M 5-2 strain in the of whole forage flours meals of legumes.
| Cellulolytic Activity | Legumes | Fermentation Time (h) | SE and p | ||||
| 24 | 48 | 72 | 96 | 120 | |||
| PFase (IU/mL) |
L. purpureus |
0.74d | 0.46e | 0.39e | 0.38ef | 0.20d | ±0.01 p<0,0001 |
|
M. pruriens |
0.49g | 0.60d | 0.64c | 0,63c | 0.50b | ||
| Productivity enzymatic (UI/gMSh) |
L. purpureus |
1.17 | 0.74 | 0.62 | 0.61 | 0.32 | |
|
M. pruriens |
0.84 | 0.99 | 1.06 | 1.04 | 0.83 | ||
a,b,c,d,e,f,g. Means with different letters in each row differ at (p < 0.005) (Duncan, 1955). S.E ± 0.01; p<0.0001 (SE: standard error, p: level of significance).
Table 3.
Determination of pH during fermentation of the whole forage flours meals of legumes with the T. viride M5-2 strain.
Table 3.
Determination of pH during fermentation of the whole forage flours meals of legumes with the T. viride M5-2 strain.
| Indicator | Legumes | Fermentation Time (h) | SE and p | |||||
| pH | 0 | 24 | 48 | 72 | 96 | 120 | ||
| L. purpureus |
6.13d | 6.83f | 7.46i | 7.37h | 7.28g | 7.20b | ±0.03 p<0.001 |
|
| M. pruriens |
6.03c | 6.71e | 6.72e | 7.12g | 7.11a | 7.10g | ||
a,b,c,d,e,f,g,h,i. Means with different letters in each row differ at (p<0.05) (Duncan, 1955). (S.E ± 0.03; p<0.001). SE: standard error, p: level of significance.
Table 4.
Chemical composition of the fiber of the whole forage flours meals of the legumes L. purpureus and M. pruriens, and protein composition during fermentation with T. viride M5-2.
Table 4.
Chemical composition of the fiber of the whole forage flours meals of the legumes L. purpureus and M. pruriens, and protein composition during fermentation with T. viride M5-2.
| Indicators (% de DM) |
Legumes | Fermentation Time (h) | SE and p | |||
| 0 | 24 | 48 | 72 | |||
| DM* | L. purpureus | 28.1d | 28.3d | 27.8c | 27.9b | ±0.11 p<0.05 |
| M. pruriens | 28.8c | 26.4a | 27.1b | 27.1b | ||
| CP | L. purpureus | 13.63a | 17.15c | 15.36a | 17.03c | ±0.13 p<0.001 |
| M. pruriens | 19.04b | 21.34d | 22.05e | 21.27d | ||
| TP | L. purpureus | 11.30a | 13.24a | 13.21a | 14.40b | ±0.14 p<0.001 |
| M. pruriens | 17.58c | 19.53e | 21.74g | 20.61f | ||
| NDF | L. purpureus | 67.83 b | 67.83b | 66.74a | 67.90b | ±0.29 p<0.001 |
| M. pruriens | 73.06d | 68.48b | 69.56c | 69.83c | ||
| ADF | L. purpureus | 51.44a | 51.18a | 52.97c | 54.31d | ±0.18 p<0.001 |
| M. pruriens | 52.27b | 52.54bc | 52.64bc | 54.78d | ||
| Cel | L. purpureus | 42.07f | 39.28b | 41.67e | 43.88g | ±0.13 p<0.001 |
| M. pruriens | 39.66c | 39.97cd | 38.41a | 40.14d | ||
| Lig | L. purpureus | 9.17a | 10.87 c | 10.93c | 10.20b | ±0.13 p<0.001 |
| M. pruriens | 12.33e | 11.38d | 13.35f | 13.98g | ||
| Hem | L. purpureus | 16.39d | 16.65d | 13.27a | 13.59a | ±0.08 p<0.0001 |
| M. pruriens | 20.79f | 15.74c | 16.92e | 15.05b | ||
a,b,c,d *Means with different letters in each row differ at (p < 0.05) (Duncan, 1955). p: level of significance. a,b,c,d,e,f,g,h,i. Means with different letters in each row differ at p < 0.05 (Duncan, 1955). SE: standard error, p: level of significance. The values highlighted in bold correspond to the greatest increase in the quemical indicators carried out according to analysis (drye matter [DM], crude protein [CP], true protein [CT], neutral detergent fiber [NDF], acid detergent fiber [ADF], cellulose [Cel], lignin [lig], hemicellulose [Hem].
Table 5.
Physical composition of the fiber of the whole forage flours meals of the legumes L. purpureus and M. pruriens, during of fermentation with T. viride M5-2.
Table 5.
Physical composition of the fiber of the whole forage flours meals of the legumes L. purpureus and M. pruriens, during of fermentation with T. viride M5-2.
| Physical indicators | Legumes | Fermentation Time (h) | ||||
| 0 | 24 | 48 | 72 | SE and p | ||
| V (g/ml) | L. purpureus | 4.05a | 3.60b | 3.35d | 4.05a | ±0,04 p<0,0001 |
| M. pruriens | 3.48c | 3.21e | 3.41de | 3.25de | ||
| WAC (g/g) | L. purpureus | 3.71d | 3.76d | 2.85e | 4.36c | ±0,13 p=0,0005 |
| M. pruriens | 4.66bc | 5.14a | 4.68bc | 4.78ab | ||
| ABC (meq) | L. purpureus | 0.51f | 0.42h | 0.59b | 0.47g | ±0,002 p<0,0001 |
| M. pruriens | 0.63a | 0.58c | 0.53d | 0.56e | ||
| BBC (meq) | L. purpureus | 0.36c | 0.31g | 0.43a | 0.36c | ±0,001 p<0,0001 |
| M. pruriens | 0.32f | 0.41b | 0.35d | 0.33e | ||
a,b,c,d,e,f,g,h,i. Means with different letters in each row differ at p < 0.05 (Duncan, 1955). SE: standard error, p: level of significance. The values highlighted in bold correspond to the greatest increase in the indicators. Physical composition of the fiber carried out according to analysis: Volume (V), Water adsorption capacity (WAC), acid and base buffering capacity (ABC, BBC).
Table 6.
Main effects on the solubility of the whole forage flours meals of the legumes L. purpureus and M. pruriens, during of fermentation with T. viride M5-2.
Table 6.
Main effects on the solubility of the whole forage flours meals of the legumes L. purpureus and M. pruriens, during of fermentation with T. viride M5-2.
| Physical indicator | Legumes | SE and p | |||
|
Solubility (%) |
L. purpureus | M. pruriens | ±0,07 p=0,0613 |
||
| 7.22 | 7.22 | ||||
| Fermentation time (h) |
±0,11 p=0,0024 |
||||
| 0 | 24 | 48 | 72 | ||
| 7,05bc | 7,16ab | 7,47a | 6,77c | ||
a,b,c Means with different letters in each row differ at p < 0.05 (Duncan, 1955). SE: standard error, p: level of significance. Physical composition of the fiber carried out according to analysis: solubility (S).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



