
Submitted:
24 April 2025
Posted:
24 April 2025
You are already at the latest version
Abstract
A triplex-forming oligonucleotide (TFO) can form a sequence-specific triple helix via Hoogsteen hydrogen bonding to polypurine tracts within a major groove side of a DNA duplex. Triplex formation can induce a double-strand break, and this phenomenon at the amplified gene loci can selectively induce the cell death of cancer cells with specific gene amplification. However, the relationship between the binding affinity of TFO for target gene loci and the cell death response remains unclear. In this study, we aimed to develop DNA intercalator-conjugated TFOs with higher affinity for the human epidermal growth factor receptor type2 (HER2) gene, which is often amplified in breast cancer cells, than the unmodified TFO. The binding affinity of the TFOs for the target DNA duplex was analyzed using nondenaturing polyacrylamide gel electrophoresis, and one of the DNA intercalator-conjugated TFOs showed a higher binding affinity for the target duplex than the unmodified TFO. We also evaluated the cell death responses induced by these TFOs using the WST-8 assay, suggesting that the higher binding affinity of the TFO for amplified gene loci can lead to a stronger cell death response of cancer cells with specific gene amplification.
Keywords:
triplex-forming oligonucleotide
; DNA
; intercalator
; human epidermal growth factor receptor type2
; anticancer
1. Introduction
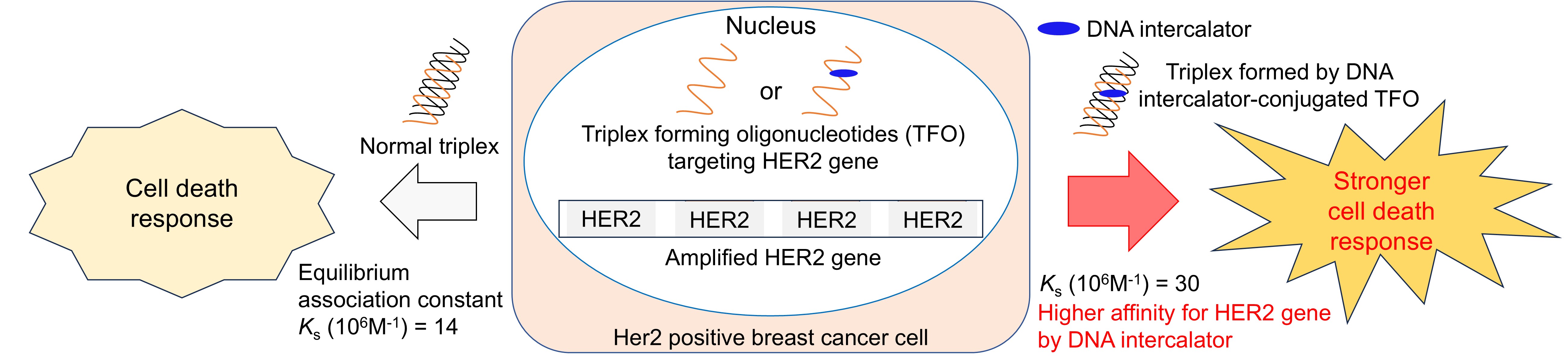
The triplex-forming oligonucleotide (TFO) can form a sequence-specific triple helix via Hoogsteen hydrogen bonding to polypurine tracts within a major groove side of the DNA duplex [1,2,3,4]. Triplex formation can be used to regulate gene functions by disturbing the interactions of DNA with protein complexes such as DNA polymerase and transcription factors [5,6,7]. Rogers et al. reported that triplex formation causes DNA perturbation, which interferes with replication fork progression, resulting in fork collapse and DNA double-strand breaks (DSBs). The efficiency of DSB induction by TFO is low because the endogenous DNA repairing system competes with this perturbation to avoid a DSB [8,9]. However, the same research group demonstrated that targeting the amplified human epidermal growth factor receptor type2 (HER2) gene loci of HER2 positive breast cancer cells by TFO was very effective in inducing cell apoptosis [10], demonstrating the potential of TFO as a new drug platform for cancer therapy. Therefore, we hypothesized that a higher binding affinity of the TFO for HER2 gene loci can induce a stronger cell death response of HER2 positive breast cancer cells (Figure 1).
To increase the binding affinity of the TFO for the target duplex, we need to address the intrinsic problems of the TFO technology, which is the requirement of a consecutive purine base sequence (polypurine sequence) in one strand of the target duplex DNA, as the Hoogsteen hydrogen bonding is formed only between the TFO and the purine bases of the duplex DNA. When we try to find the potential triplex formation site (polypurine sequence) in the target gene, the candidate sequences often contain one or two pyrimidine bases that are often mentioned as a mismatch site, which causes a reduction in the binding affinity of TFO for the target sequence. To solve this problem, several artificial nucleobase analogs have been developed that can form hydrogen bonds with pyrimidine bases within the polypurine sequence. In addition, the introduction of DNA intercalators into TFOs has been reported to effectively improve the binding affinity of TFO for target duplex [11,12]. Recently, our group developed the novel nucleoside analog P (Figure 2) that possesses a psoralen, a DNA intercalator, at its 1'-position. We successfully demonstrated that the replacement of the mismatch bases of the parallel TFO with P, named OPTO after “1´(One)-psoralen-conjugated triplex-forming oligonucleotide,” improves binding affinity of the parallel TFO for the target duplex [13].
In this study, we applied our novel nucleoside analog P to antiparallel TFO to increase the binding affinity of the TFO for amplified HER2 gene loci and evaluated the apoptosis induction of HER2-amplified breast cancer cells (Figure 1). Our study shed light on the relationships between the binding affinity of the TFO for amplified gene loci under physiological conditions and the apoptosis induction levels, indicating the efficacy of DNA intercalator-conjugated TFO for anticancer therapy.
2. Materials and Methods
2.1. General Information
The phosphoramidite of P was synthesized according to our previous report [13]. The target duplexes (HER2-205-Py and HER2-205-Pu, HER2-5992-Py, and HER2-5992-Pu) and unmodified TFOs (205-TFO and 5922-TFO) were purchased from FASMAC Co., Ltd. (Kanagawa, Japan). The solid-phase syntheses of the modified TFOs with P (205-Ps1 and 205-Ps2, 5992-Ps1, and 5992-Ps2) were performed at Ajinomoto Genedesign (Osaka, Japan). The fluorescence intensity of the DNA bands in the gel was quantified using ImageJ.
2.2. Solid-Phase Synthesis of Modified TFOs with P (205-Ps1 and 205-Ps2, 5992-Ps1, and 5992-Ps2)
The syntheses of 205-Ps1 and 205-Ps2, 5992-Ps1, and 5992-Ps2 were performed at Ajinomoto Genedesign. Briefly, the phosphoramidites were introduced to oligonucleotides on the CPG support using phosphoramidite chemistry, and the purification of the oligos was performed by reverse-phased HPLC (RP-HPLC) on an X-Bridge C18 column (2.5 μm 4.6*75 mm, Waters, Co. Ltd., MA, USA) with a linear gradient of methanol in 100 mM 1,1,1,3,3,3-hexafluoro-2-propanol containing 8 mM triethylamine at a flow rate of 1.0 mL/min at 60 °C (methanol gradient: 5%–30%, 20 min). The mass spectra were obtained using MALDI-TOF-MS (Bruker DALTONICS autoflex speed).
2.3. Triplex Formation Analysis Using Nondenaturing Polyacrylamide Gel Electrophoresis (Native PAGE)
Sample solutions containing the ds-DNAs (100 nM) and different concentrations of TFOs (0, 10, 50, 60, 70, 80, 90, 100, 500, and 1,000 μM each), 20 mM Tris-HCl (pH 7.0), and 5 mM MgCl2 were denatured by heating to 95°C and subsequently cooling to 4°C at 0.5°C/min and were then incubated at 37°C for 12 h. Then, the samples were diluted with 40 wt% sucrose aq (sample: 40 wt% sucrose aq = 1:4 (v/v)) and analyzed with 20% native polyacrylamide gel (PAGE) containing 5 mM Mg2+ in TBM (37°C, 120 V, 90 min). The DNA bands were stained by SYBR® Gold, and then the gels were transferred to imaging plates. The gel images were analyzed and quantitated using ChemDoc Touch MP (BioRad, CA, USA.).
2.4. Evaluation of Anticancer Activities of TFO Using the WST-8 Assay
The BT-474 cells were planted in a 96-well plate (1 × 104 cells/well) and incubated for 24 h (5%CO2, 37ºC). Then, the cells were transfected with each TFO (25 nM) using lipofectamine 3,000 (Thermo Scientific™, MA, USA). After 24 h incubation (5%CO2, 37ºC), the cellular viability was quantified by the WST-8 assay using the Cell Counting Kit-8 (DOJINDO, Kumamoto, Japan), and the absorbance at 450 nm was measured using a multimode plate reader Cytation3 (BioTek, CA, USA).
3. Results and Discussion
3.1. Evaluation of the Thermodynamic Stability of Triplexes with HER2 Gene Sequences
The phosphoramidite for the nucleotide analog P was synthesized according to our recent report [13]. OPTOs was synthesized at Ajinomoto Genedesign (Osaka, Japan). Briefly, the phosphoramidite for P was introduced into oligonucleotides on the CPG support using the phosphoramidite chemistry. According to the Rogers’ report [10], a sequence within the HER2 gene (HER2-205) was chosen as a target for OPTO (Figure 2a). As this target sequence contains two mismatch sites, the replacement of these two mismatch bases with P would be expected to increase the binding affinity for the target duplex. Therefore, we prepared two types of OPTOs with one or two P (205-Ps1 and 205-Ps2, respectively). We evaluated the binding affinity of the OPTOs for the HER2-205 sequence using nondenaturing polyacrylamide gel electrophoresis (Native PAGE). In this experiment, the 5´ end of the pyrimidine base-rich sequence (HER2-205Py) was labeled with tetramethylrhodamine. Further, to improve the anticancer activity, an amino linker was introduced at the 3´ end of TFOs, which has been reported to improve the cellular uptake and nuclear translocation of TFOs [14]. Sample solutions containing different concentrations of TFOs were prepared and analyzed by Native PAGE, and the fluorescence intensity of the DNA bands was quantified (Figure 2b). It is well known that the magnesium ion (Mg2+) is important in the formation of a stable triplex by mitigating the electrostatic repulsion of the phosphate backbone of oligonucleotides. For the TFO binding assay using Native PAGE, the concentration of Mg2+ in the sample solution generally varies (2.5–20 mM) [15,16,17,18,19,20,21]. However, the conformational difference of the triplex depending on the Mg2+ concentration might cause either the promotion or interference of the intercalation of the psoralen moiety in the TFO, leading either to the stabilization or destabilization of the corresponding triplex. Therefore, we conducted this experiment at two different Mg2+ concentrations (5 and 20 mM). The obtained representative gel images for each TFO (205-TFO, 205-Ps1, and 205-Ps2) at both 5 and 20 mM Mg2+ concentrations are shown in Figure 2b. The graphical charts in Figure 2b show the quantification results of the DNA bands in the obtained gels. At 20 mM Mg2+ concentration, the unmodified 205-TFO showed a higher binding affinity for the target duplex than that observed at 5 mM Mg2+. The 205-Ps1 showed a higher binding affinity for the target duplex than the unmodified 205-TFO and 205-Ps2 at both Mg2+ concentration, suggesting the triplex stabilization effect of P. In contrast, the additional introduction of P in the 205-Ps2 decreased the binding affinity of 205-Ps2 for the target duplex than the binding affinity of 205-Ps1 for the same target sequence, although it was still higher than that of the unmodified 205-TFO at 5 mM Mg2+.
We also conducted the same experiment using another sequence (HER2-5992) (Figure 3a) that was also the target site of TFO in the Rogers’ report [10]. In this case, 5992-Ps2 showed a higher binding affinity for the target duplex than did 5992-TFO and 5992-Ps1 at 20 mM Mg2+ concentration, whereas 5992-TFO showed a higher binding affinity for the target duplex than 5992-Ps1 and 5992-Ps2 at 5 mM Mg2+ concentration. This result also suggests that the replacement of mismatch bases in TFO with P does not always increase the binding affinity for the target duplex, and it depends on both the Mg2+ concentration and the target sequences.
The equilibrium association constants of the TFOs (Ks) were calculated according to the literature (Table 1) [18,19]. These Ks values also suggest that the DNA intercalator causes either stabilization or destabilization of the corresponding triplex depending on the Mg2+ concentration and target DNA sequence.
3.2. Evaluation of the Anticancer Activities of TFOs Against HER2-Positive Breast Cancer Cells
As the 205-Ps1 showed higher binding affinity for the HER2-205 sequence in a wide range of Mg2+ concentration conditions, we hypothesized that this higher binding affinity of 205-Ps1 for the target sequence would lead to stronger cell death of HER2-positive breast cancer cells (BT474). Thus, we evaluated the anticancer activities of these TFOs (205-TFO, 205-Ps1, and 205-Ps2) against BT474 cells (Figure 4). We also prepared a scramble sequence (Ps-scramble) as a negative control. The TFOs were introduced in the cancer cells by lipofection, and cell viability was analyzed using the WST-8 assay. The unmodified 205-TFO induced significant cell death of HER2-positive breast cancer cells, as demonstrated in the Rogers’ report [10]. As expected, the cell death response induced by 205-Ps1 was the strongest among the TFOs, suggesting that the binding affinity of the TFO for the target HER2 gene was related to anticancer activities. These results were correlated with the Ks values obtained at 5 mM Mg2+ concentration. When we consider the endogenous Mg2+ concentration range of a mammalian cell (0.5–10 mM) [22], it is reasonable to think that the anticancer activities of the TFOs were related to the Ks values obtained at 5 mM Mg2+ concentration. This suggests the importance of evaluating the binding affinity of the TFO for the target duplex within the Mg2+ concentration range of a mammalian cell (0.5–10 mM), especially for the DNA intercalator-conjugated TFO, as the intercalation of the DNA intercalator was affected by the conformational changes in the triplex caused by Mg2+ concentration and target DNA sequence, resulting in either the stabilization or destabilization of the corresponding triplex.
4. Conclusions
The binding affinity of our DNA intercalator-conjugated TFOs (OPTO) for the target duplex was evaluated using two target sequences in the HER2 gene (HER2-205 and HER2-5992). In the case of TFO 205-Ps1, in which a psoralen was replaced with one mismatched base of 205-TFO, the binding affinity for the target duplex was improved. However, we found that this stabilization effect of OPTO is completely dependent on the sequence and Mg2+ concentration, possibly causing the conformational differences of the triplex. As we expected, 205-Ps1, which formed a more stable triplex structure with the Her2-205 target sequence, had a stronger anticancer activity against BT-474 (one of the HER2-positive breast cancer cell) than unmodified 205-TFO. These results suggest that the enhanced binding affinity of the TFO for the target duplex structure leads to a stronger cell death and that OPTO may have potent anticancer activity against various HER2 copy number-amplified breast cancer cells other than BT-474.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Analytical data of 205-Ps1; Figure S2: Analytical data of 205-Ps2; Figure S3: Analytical data of 5992-Ps1; Figure S4: Analytical data of 5992-Ps2; Figure S5: The details of KS value calculation using datas collected for 205-Ps1; Figure S6: The whole gel images; Table S1: The row datas of WST-8 assay (absorbance at 450 nm); Figure S7: The sequence of Ps-scramble.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, A.Y. and Y.M.; methodology, Y.M.; validation, H.T., Y.U., Y.M.; investigation, H.T., Y.M.; data curation, Y.M.; writing—original draft preparation, H.T., Y.M.; writing—review and editing, H.T., A.T., T.W., A.Y., Y.M.; supervision, Y.M.; funding acquisition, A.Y., Y.M. All authors have read and agreed to the published version of the manuscript.” Please turn to the CRediT taxonomy for the term explanation. Authorship must be limited to those who have contributed substantially to the work reported.
Funding
Please add: This research was funded by the Grant-in-Aid for Transformative Research Areas (A) “Material Symbiosis” (Grant Number:20H05874 awarded to A.Y.) from MEXT, Japan. This study was also supported by JSPS KAKENHI (Grant Num-bers 22H00593 and 22K14839 to A.Y. and Y.M., respectively), Japan, and the Cooperative Research Program of “Network Joint Research Center for Materials and Devices” (Grant Numbers 20224030 and 20241141 to A.Y. and Y.M., respectively). This work was also supported by AMED under Grant Number 25ak0101227h0002 and 24ym0126811j0003.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article and supplementary materials.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Felsenfeld, G.; Davies, D.R.; Rich, A. FORMATION OF A THREE-STRANDED POLYNUCLEOTIDE MOLUCULE. J. Am. Chem. Soc. 1957, 79, 2023–2024. [Google Scholar] [CrossRef]
- Frank-Kamenetskii, M.D.; Mirkin, S.M. TRIPLEX DNA STRUCTURES. Annu. Rev. Biochem. 1995, 64, 65–95. [Google Scholar] [CrossRef] [PubMed]
- Fossella, J.A.; Kim, Y.J.; Shin, H.; Richards, E.G.; Fresco, J.R. Relative specificities in binding of Watson - Crick base pairs by third strand residues in a DNA pyrimidine triplex motif. Nucleic Acids Res. 1993, 21, 4511–4515. [Google Scholar] [CrossRef] [PubMed]
- Thuong, N.T.; Hélène, C. Sequence-Specific Recognition and Modification of Double-Helical DNA by Oligonucleotides. Angew. Chem. Intl. Ed. Engl. 1993, 32, 666–690. [Google Scholar] [CrossRef]
- Karympalis, V.; Kalopita, K.; Zarros, A.; Carageorgiou, H. Regulation of Gene Expression via Triple Helical Formations. Biochemistry, 2004, 69, 855–860. [Google Scholar] [CrossRef]
- Duca, M.; Vekhoff, P.; Oussedik, K.; Halby, L.; Arimondo, P.B. The triple helix: 50 years later, the outcome. Nucleic Acids Res. 2008, 36, 5123–5138. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhou, Z.; Ren, C.; Deng, Y.; Peng, F.; Wang, Q.; Zhang, H.; Jiang, Y. Triplex-forming oligonucleotides as an anti-gene technique for cancer therapy. Front. Pharmacol. 2022, 13, 1007723. [Google Scholar] [CrossRef] [PubMed]
- Kaushik Tiwari, M.; Rogers, F.A. XPD-dependent activation of apoptosis in response to triplex-induced DNA damage. Nucleic Acids Res. 2013, 41, 8979–8994. [Google Scholar] [CrossRef] [PubMed]
- Kaushik Tiwari, M.; Adaku, N.; Peart, N.; Rogers, F.A. Triplex structures induce DNA double strand breaks via replication fork collapse in NER deficient cells. Nucleic Acids Res. 2016, 44, 7742–7754. [Google Scholar] [CrossRef] [PubMed]
- Kaushik Tiwari, M.; Colon-Rios, D.A.; Rao Tumu, H.C.; Liu, Y.; Quijano, E.; Krysztofiak, A.; Chan, C.; Sang, E.; Braddock, D.T.; Suh, H.-W.; Saltzman, W.M.; Rogers, F.A. Direct targeting of amplified gene loci for proapoptotic anticancer therapy. Nat. Biotechnol. 2022, 40, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Mikame, Y.; Yamayoshi, A. Recent Advancements in Development and Therapeutic Applications of Genome-Targeting Triplex-Forming Oligonucleotides and Peptide Nucleic Acids. Pharmaceutics. 2023, 15, 2515. [Google Scholar] [CrossRef] [PubMed]
- Hari, Y.; Obika, S.; Imanishi, T. Towards the Sequence-Selective Recognition of Double-Stranded DNA Containing Pyrimidine-Purine Interruptions by Triplex-Forming Oligonucleotides. Eur. J. Org. Chem. 2012, 2875–2887. [Google Scholar] [CrossRef]
- Mikame, Y.; Toyama, H.; Dohno, C.; Wada, T.; Yamayoshi, A. Development and functional evaluation of a psoralen-conjugated nucleoside mimic for triplex-forming oligonucleotides. Commun. Chem. 2025, 8, 18. [Google Scholar] [CrossRef] [PubMed]
- Catapano, C.V.; McGuffie, E.M.; Pacheco, D.; Carbone, G.M.R. Inhibition of gene expression and cell proliferation by triple helix-forming oligonucleotides directed to the c-myc gene. Biochemistry 2000, 39, 5126–5138. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-. Y, Bi, G.; Miller, P.S. Triplex Formation by Oligonucleotides Containing Novel Deoxycytidine Derivatives. Nucleic Acids Res. 1996, 24, 2606–2613. [Google Scholar] [CrossRef] [PubMed]
- Amosova, O.A.; Fresco, J.R. A search for base analogs to enhance third-strand binding to ‘inverted’ target base pairs of triplexes in the pyrimidine/parallel motif. Nucleic Acids Res. 1999, 27, 4632–4635. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.L.; McLaughlin, L.W. Use of pKa Differences to Enhance the Formation of Base Triplets Involving C−G and G−C Base Pairs. J. Org. Chem. 2000, 65, 7468–7474. [Google Scholar] [CrossRef] [PubMed]
- Okamura, H.; Taniguchi, Y.; Sasaki, S. Aminopyridinyl–Pseudodeoxycytidine Derivatives Selectively Stabilize Antiparallel Triplex DNA with Multiple CG Inversion Sites. Angew. Chem. Int. Ed. 2016, 55, 12445–12449. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Taniguchi, Y.; Okamura, H.; Sasaki, S. Modification of the aminopyridine unit of 2′-deoxyaminopyridinyl-pseudocytidine allowing triplex formation at CG interruptions in homopurine sequences. Nucleic Acids Res. 2018, 46, 8679–8688. [Google Scholar] [CrossRef] [PubMed]
- Notomi, R.; Sasaki, S.; Taniguchi, Y. Recognition of 5-methyl-CG and CG base pairs in duplex DNA with high stability using antiparallel-type triplex-forming oligonucleotides with 2-guanidinoethyl-2′-deoxynebularine. Nucleic Acids Res. 2022, 50, 12071–12081. [Google Scholar] [CrossRef] [PubMed]
- Milo, R.; Phillips, R. CELL BIOLOGY by the numbers, Garland Science, Oxford, 2016; pp. 92–93.
Figure 1.
Working hypothesis of this work. The higher binding affinity of OPTOs for amplified gene loci will result in the formation of a larger number of triplexes in the nucleus, which will induce a stronger cell death response of cancer cells.
Figure 1.
Working hypothesis of this work. The higher binding affinity of OPTOs for amplified gene loci will result in the formation of a larger number of triplexes in the nucleus, which will induce a stronger cell death response of cancer cells.
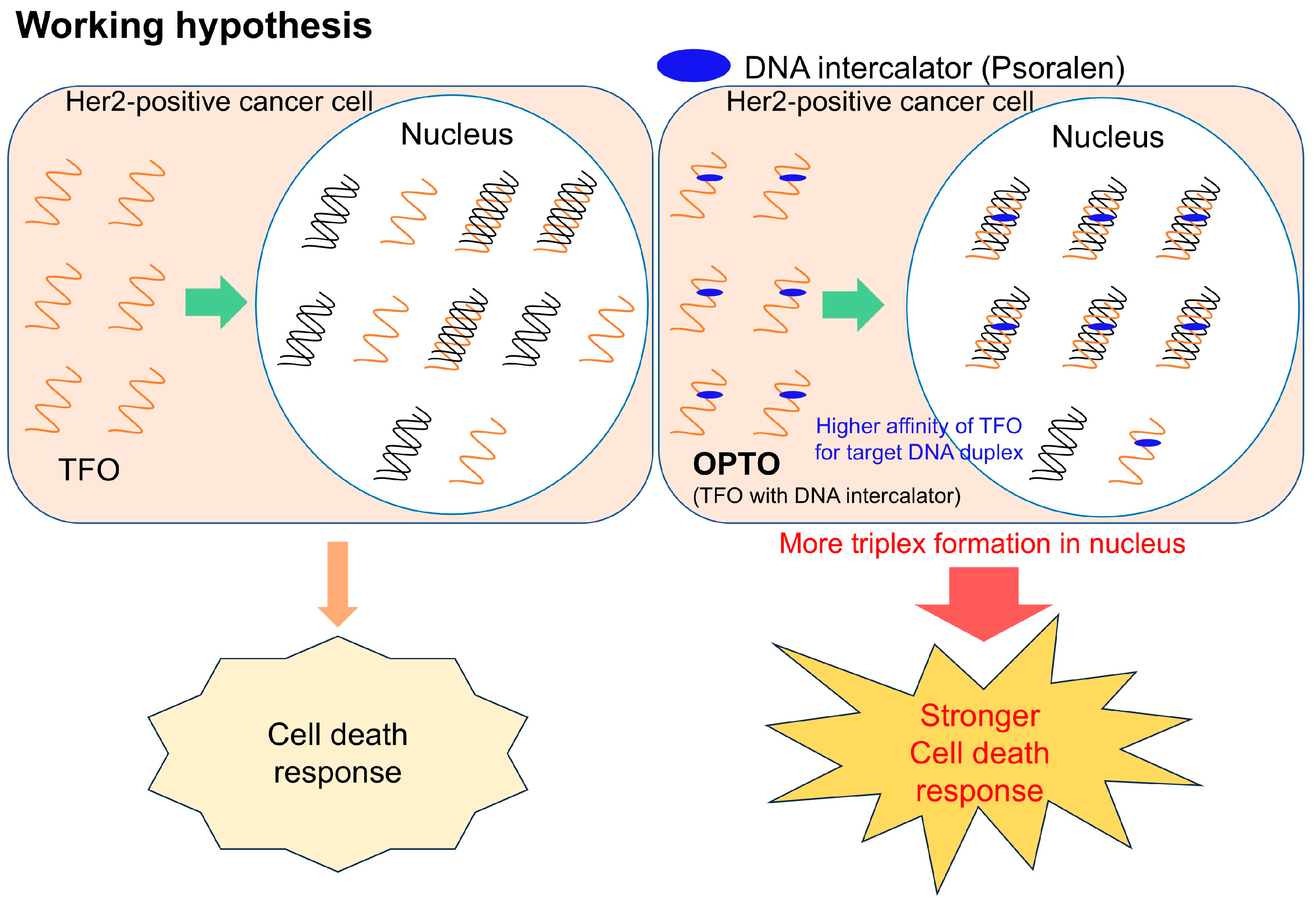
Figure 2.
Analysis of the triplex formation with the HER2-205 sequence using Native PAGE. (a) Sequences of target DNA duplexes and TFOs. (b) Representative gel images of each TFO with the target duplex and the quantification results of Native PAGE, which are displayed as the mean ± standard deviation (s.d.) for n = 3 replicates.
Figure 2.
Analysis of the triplex formation with the HER2-205 sequence using Native PAGE. (a) Sequences of target DNA duplexes and TFOs. (b) Representative gel images of each TFO with the target duplex and the quantification results of Native PAGE, which are displayed as the mean ± standard deviation (s.d.) for n = 3 replicates.
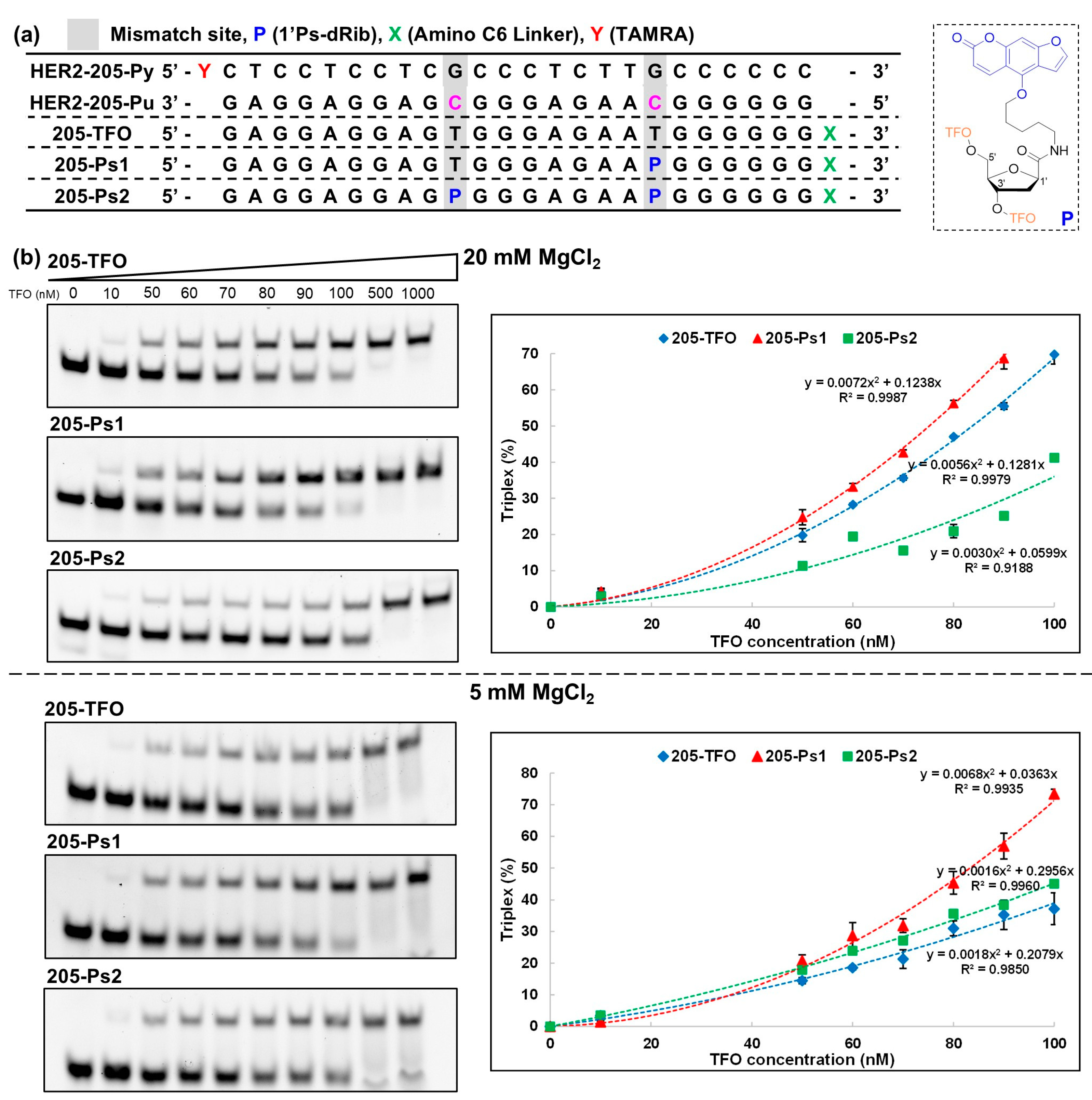
Figure 3.
Analysis of the triplex formation with the HER2-5992 sequence using Native PAGE. (a) Sequences of target DNA duplexes and TFOs. (b) Representative gel images of each TFO with the target duplex and the quantification results of Native PAGE, which are displayed as the mean ± standard deviation (s.d.) for n = 3 replicates.
Figure 3.
Analysis of the triplex formation with the HER2-5992 sequence using Native PAGE. (a) Sequences of target DNA duplexes and TFOs. (b) Representative gel images of each TFO with the target duplex and the quantification results of Native PAGE, which are displayed as the mean ± standard deviation (s.d.) for n = 3 replicates.
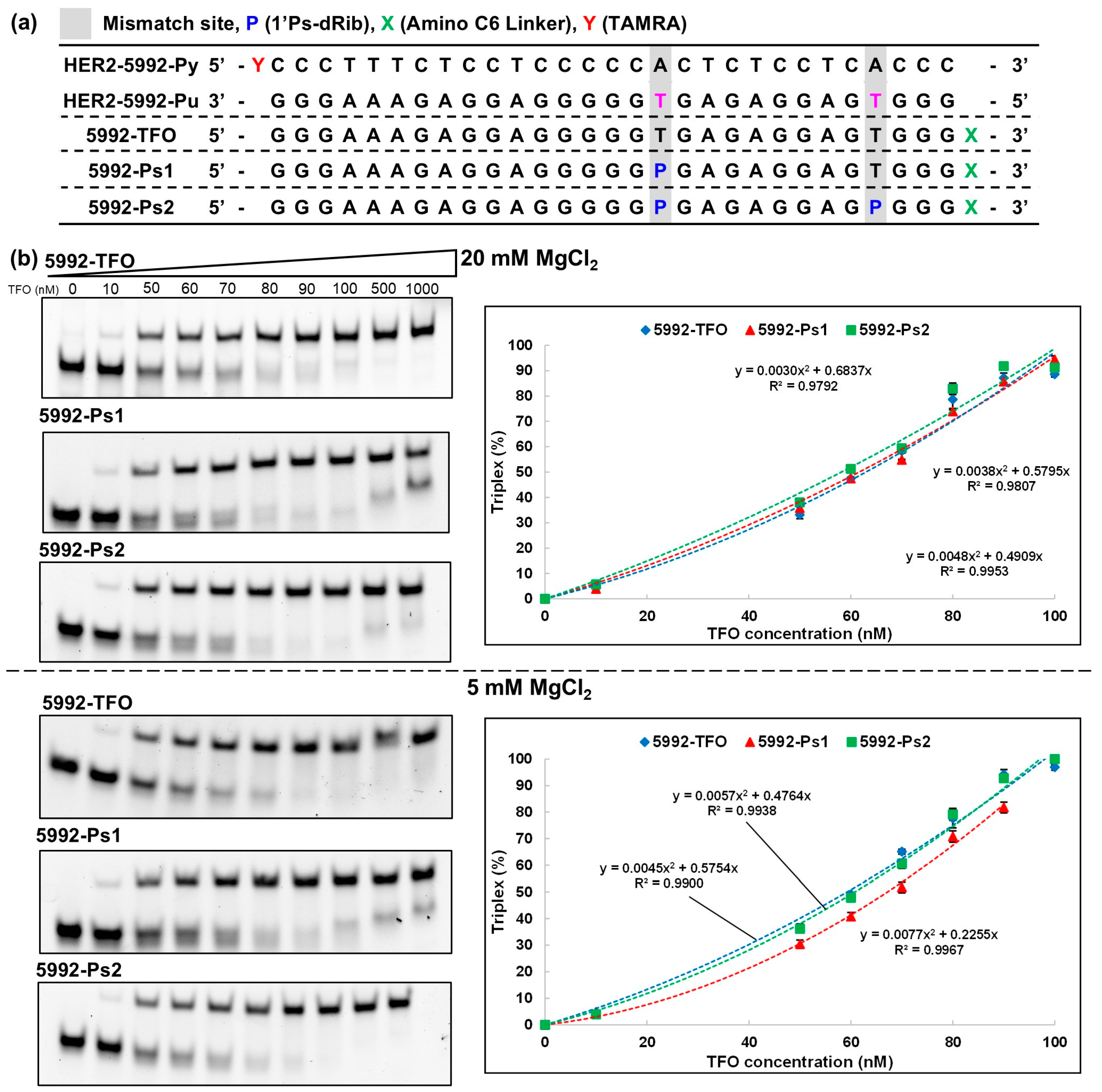
Figure 4.
The anticancer activities of TFOs against HER2-positive breast cancer cells (BT474) are displayed as the mean ± standard deviation (s.d.) for n = 4 replicates. The final concentration of TFOs was 25 nM. Statistical significance determined by two-tailed t-test. ***P < 0.001, ****P < 0.0001.
Figure 4.
The anticancer activities of TFOs against HER2-positive breast cancer cells (BT474) are displayed as the mean ± standard deviation (s.d.) for n = 4 replicates. The final concentration of TFOs was 25 nM. Statistical significance determined by two-tailed t-test. ***P < 0.001, ****P < 0.0001.
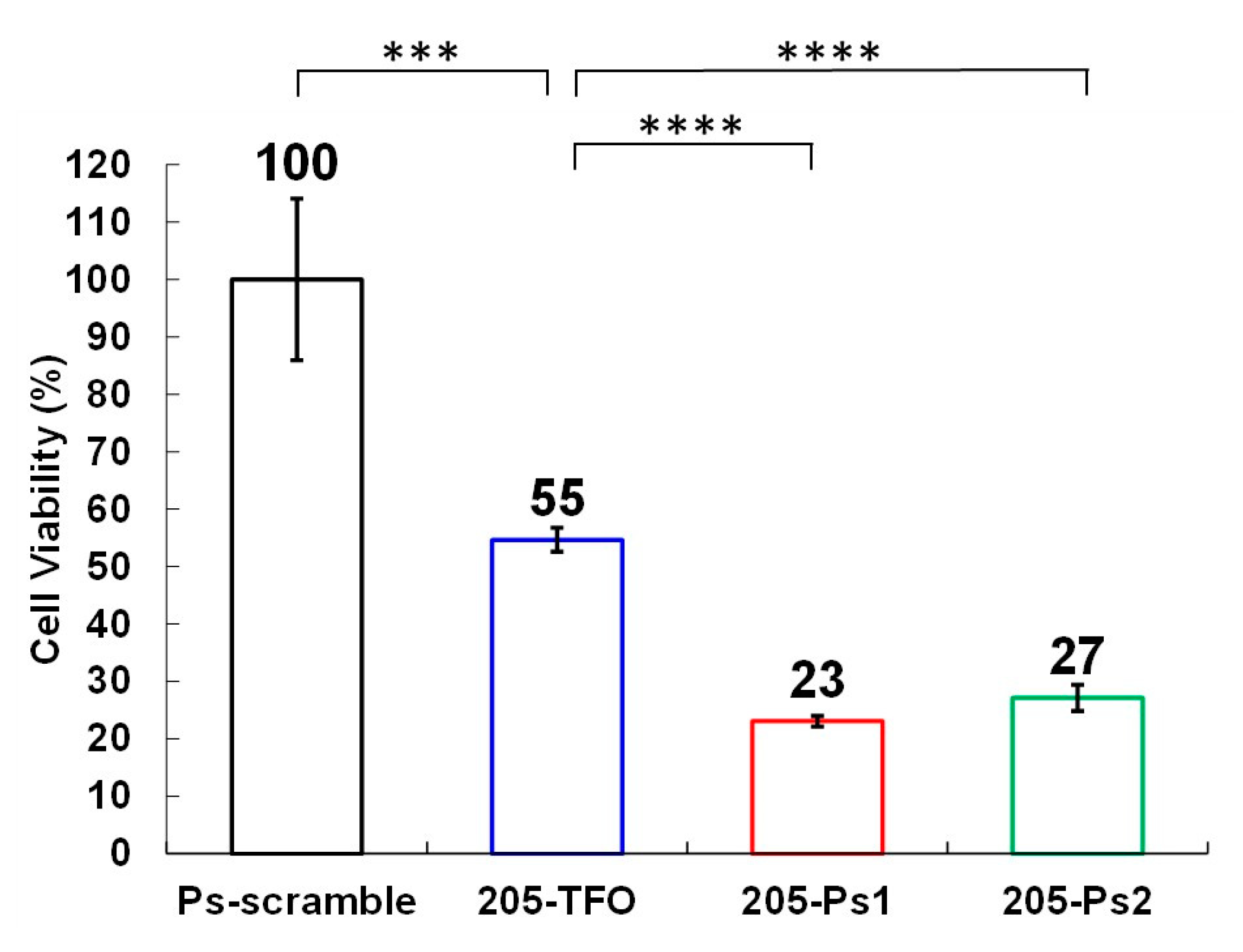

Table 1.
Ks values of the TFOs.
|
KS value (106 M−1) at 20 mM Mg2+ |
KS value (106 M−1) at 5 mM Mg2+ |
|
|---|---|---|
| 205-TFO | 30 | 14 |
| 205-Ps1 | 40 | 30 |
| 205-Ps2 | 8 | 18 |
| 5992-TFO | 77 | 107 |
| 5992-Ps1 | 87 | 58 |
| 5992-Ps2 | 121 | 93 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



