
Submitted:
23 April 2025
Posted:
24 April 2025
You are already at the latest version
Abstract
In Tanzania, dengue outbreaks have occurred almost annually over the past decade, with each new outbreak becoming more severe. This study investigated the prevalence of dengue virus (DENV) serotypes in the wild Aedes aegypti and their blood sources to determine human exposure risk in Dar es Salaam, Tanzania. A two-year longitudinal survey was conducted in Ilala, Kinondoni, and Temeke districts of Dar es Salaam to sample Ae. aegypti mosquitoes using Biogents Sentinel trap (BGS), Prokopack aspiration and Gravid Aedes trap (GAT). Collected mosquitoes were pooled in groups of 10 and tested for DENV1-4 serotypes using reverse transcription polymerase chain reaction (RT-qPCR). Blood meal sources were identified using an enzyme-linked immunosorbent assay (ELISA). Of 854 tested pools, only DENV-2 serotype was detected in all districts (Temeke (3/371 pools), Ilala (1/206 pools) and Kinondoni (1/277-pools)). Blood meal analysis showed a strong preference for humans (81%) and mixed blood meals (17%). Out of 354 collected host seeking Ae. aegypypti, 78.5% were captured outdoors and 21.5% indoors. This study confirms the circulation of DENV-2 in Ae. aegypti populations, indicating a potential dengue outbreak risk in Tanzania. The mosquitoes’ strong preference for human hosts and predominance in outdoor settings pose challenges for dengue control efforts.
Keywords:
DENV
; dengue fever
; traps
; blood feeding
; longitudinal survey
; serotypes
; xenomonitoring
1. Introduction
Dengue fever is a significant global public health concern, responsible for hundreds and thousands cases of morbidity and mortality annually across tropical and sub-tropical regions [1]. Approximately 3.9 billion people, nearly half of the world’s population are estimated to be at risk of infection [2]. The global incidence of dengue has risen dramatically in recent decades, with a record high occuring in 2023 [3,4] and 2024 [5]. Nearly 100 to 400 million new dengue cases occur each year [6], of which 90 million are manifested clinically, ranging from mild symptoms to the severe, life-threatening dengue haemorrhagic fever (DHF) and dengue shock syndrome (DSS) [6,7]. However, the vast majority of the cases are asymptomatic [8], leading to a likely underestimation of the true scale of the infection.
Dengue is now endemic in over 129 countries [9], with most cases occurring in Asia, which accounts for nearly two-third of the global burden, followed by the Americas and the African region [10]. In Africa, dengue cases are likely to be underreported [6] due to misdiagnosis as malaria or urinary tract infection (UTI). Even when clinically diagnosed correctly, many health systems lack sufficient diagnostic capacity to detect dengue virus (DENV) [11]. Human activities contributing to climate change, globalization, and unplanned urbanisation fuelled by rural to urban migration, accelerate the spread of dengue [12]. Additionally, it is predicted that with rapid expansion of intra- and intercontinental trade, the disease is expected to spread further, potentially tripling in the 50 years [13].
Dengue fever is caused by four antigenically distinctive virus serotypes (DENV 1-4) [14,15] which share around 65%-70% genome similarity [16,17]. The virus is an enveloped, single-stranded ribonucleic acid (ssRNA) virus belonging to the Flaviviridae family and Flavivirus genus [18,19]. It is transmitted from a viremic individual to another individual(s) through mosquito bites. Each serotype exhibits an independent virological characteristics, where the infection by one serotype does not confer cross immunity against the others [20]. Secondary infections with another serotype or mixed infection may potentially lead to severe forms of dengue, such as (DHF) or (DSS) [21,22,23]. The severity of the secondary infection is explained by the antibody-dependent enhancement (ADE) theory [24]: antibodies from a previous infection provide a long-lasting immunity against the same serotype but only temporary cross-protection against others. Thus, during a subsequent infection with a different serotype, this short-lasting immunity fails to neutralise the new serotype and forms an immune complex that facilitates viral entry into host cells, enhancing virus replication and increasing disease severity. Although all four dengue virus serotypes (DENV 1-4) circulate in Africa [25,26], DENV-2 is the most prevalent [27,28,29], likely due to its greater transmissibility [30,31] and greater susceptibility among local vectors [32]. These factors have important epidemiological implications.
Dengue fever is primarily transmitted through mosquito bites, with Aedes aegypti and Aedes albopictus serving as the primary and secondary mosquito vectors, respectively [33]. Both species are day bitters, which poses challenges for vector control because people are active at this time and most of mosquito interventions do not protect individuals during daytime [34]. In the absence of antiviral drugs and an effective universal vaccine [35], dengue prevention and control depends on vector control. Understanding Ae. aegypti host feeding preference and location is a critical aspect towards monitoring transmission and identifying potential virus reservoirs [36]. Studies show that Ae. aegypti primarily feed on humans [36,37], but in the presence of altenative hosts, they may also feed on other hosts [38,39,40]. This suggests opportunistic feeding behaviour, dependent on host availability. Dengue vectors may bite indoors or outdoors [41,42,43,44], with the mosquito endophilic resting behaviour [45] and adaptation to artificial light influencing indoor bitting [46].
In Tanzania, the first dengue incidence was reported in 1823 [25] and subsequent studies have confirmed its circulation [27,47,48,49,50,51,52]. Like other East African countries, Tanzania has all four dengue serotypes [50,53,54,55], which have likely driven the frequent dengue outbreaks in the country. Reports have indicated the co-circulation of multiple virus serotypes [55,56] as seen in the 2018/2019 outbreak with DENV-1 and 3 serotypes [55]. This shifting and co-circulation of dengue virus serotypes likely contribute to increasing disease severity. In the past decade, Tanzania has experienced several dengue outbreaks, with each being more severe than the former one. The deadliest dengue outbreak occurred in 2019, where about 7000 cases and 13 deaths linked to DENV-1 were reported [57,58]. In 2014, over 1,000 dengue cases and four deaths were recorded [59], with DENV-2 identified as the circulating serotype.
Despite frequent outbreaks recorded in recent years, dengue surveillance in Tanzania remains limited. In humans, most studies have reported dengue seroprevalence only during the outbreaks in Tanzania [48,60,61], leaving gaps in year-round data. Additionally, little information exists on the prevalence of dengue viruses in the mosquito population, indicating the possibility that the virus may be circulating in the mosquitoes posing a silent outbreak risk. Data for targeted control efforts including Ae. aegypti host preference and the location (indoors or outdoors) where these mosquitoes are most likely to feed are lacking.
Therefore, this study aimed to determine DENV prevalence in mosquitoes to assess the risk of possible dengue outbreaks and the role of xenomonitoring for low cost and non-invasive surveillance. It also investigated the host preference and feeding location of wild Ae. aegypti to better understand the dengue transmission chain.
2. Materials and Methods
Study Area
The study was conducted in Dar es Salaam, Tanzania’s largest economic hub (Figure 1). The city is located at 6.48’S and 39.17’E along the Indian Ocean coast with a population of nearly 5.5 million [62]. Administratively, it consists of five districts: Ilala, Kigamboni, Kinondoni, Temeke, and Ubungo. Based on previous dengue outbreaks [53,63], Ilala (1’649’912 people), Kinondoni (982’328), and Temeke (1’346’674) [62] were selected for this study.
Dar es Salaam has a tropical climate with high temperatures throughout the year and the hottest period occurring between October to February. It has one dry season and two rainy seasons, with an average annual rainfall of 1100 mm. The dry season spans from June to October while the short rainy seasons occurs from November to December, and the long rainy season extends from March to May [64].
Mosquito collection
Wild adult mosquitoes were collected from June 2022 to May 2024 using BioGents Sentinel (BGS) traps (Biogents AG, Regensburg, Germany) designated for catching host-seeking Aedes mosquitoes, Ovitraps (modified Biogents Gravid Aedes traps (BG-GAT; Biogents, Germany)) [65] for sampling gravid females, and Prokopack aspirators (John W Hock Company, Florida, USA) for collecting resting adults. Four wards were selected from each district. In each ward, 20 houses were identified, and the traps (one BGS and one GAT) were deployed in one house per day for 24-hours then the trapped mosquitoes were collected, followed by the collection of resting mosquitoes around the primeses using a Prockopack aspirator. This procedure was repeated monthly per each house for 24 months. The collected mosquitoes were morphologically identified to the species level following Wilkerson et al 2021 identification key [66]. Female Ae. aegypti were pooled in groups of 10 individuals and stored in 1.5 ml Eppendoff tubes containing RNAlater locally made at Swiss tph institute. The RNAlater was prepared by mixing 60 ml of 0.5 M EDTA with 37.5 ml of 1 M sodium citrate in 1,400 ml MilliQ water. Afterwards 1,050 g ammonium sulfate were added and the solution was filtered through a 0.2 µm filter.
Additionally, an experiment was conducted in six houses per month per district for three months to determine the abundance of host seeking Ae. aegypti mosquitoes indoors and outdoors. A pair of BGS traps were deployed indoors and outdoors for 24 hours, after which female mosquitoes were collected, and recorded based on their capturing location. The collected mosquitoes in this experiment were not analysed for blood meal.
RNA extraction and Dengue virus detection
RNA extraction
The extraction of RNA from mosquito pools was carried out using RNAzol® RT (Molecular research center, Cincinnati, Ohio USA) according to the manufacturer’s instructions. Briefly, each pool of 10 individual mosquitoes were suspended in 200 µl of RNAzol in a 1.5 ml microcentrifuge tube, and manually ground using a sterile plastic pestle designated for grinding mosquitoes. The mixture was then centrifuged at 12,000g for 15 minutes, after which the supernatant was transferred to a new 1.5 ml microcentrifuge tube. An equal volume (200 ml) of 100% isopropanol was added to precipitate the RNA followed by incubation for 15 minutes and centrifugation at 12, 000 g for 10 minutes. The supernatant was removed and discarded. The RNA pellet was washed twice with 200 µl of 75% ethanol, centrifuged at 4,000g for 3 minutes and the ethanol was carefully removed. Finally, the RNA pellets were eluted with 50 µl of RNAse-free water and stored at -800C for molecular analysis using reverse-transcription polymerase chain reaction (RT-qPCR) [67].
Dengue virus detection
A one-step multiplex RT-qPCR [68] was performed using the CFX96 Bio-rad PCR machine (Bio-Rad Laboratories Inc. Hecules, California USA). The primers and probes used in the assay were adapted with modification from Balingit et all [69] (Table 1). The reaction was performed in 25 µl reaction volumes using the Luna® Universal Probe One-Step RT-qPCR Kit (New England Biolabd, Ipswich, Massachusetts, USA) consisting of 5 µl RNA template, 10 µl of Luna Universal Probe One-Step Reaction Mix (2X), 1 µl of Luna WarmStart RT Enzyme Mix (20X), 0.8 µl each of forward and reverse primers (10 µM), and 0.4 µl of probes (10 µM). Each sample was analysed in duplicates. The RT-qPCR cycling conditions were as follows: reverse transcription at 50°C for 30min, initialization at 95°C for 2min, followed by 45 cycles of denaturation at 95°C for 15sec, and annealing/extension at 60°C for 1min. RNAse-free water was used as a template for the negative control. Samples with average cycle threshold (Ct) higher than 37 were considered negative for either DENV serotype.
Blood meal source
Blood-fed Aedes mosquito samples collected over two sampling years and preserved in 1.5 ml Eppendorf microcentrifuge tubes containing locally made RNAlater were selected and tested for polyclonal anti-IgG antibodies targeting vertebrates commonly found in the study area, including humans, dogs, chickens and bovines, using an enzyme-linked immunosorbent assay (ELISA) as described by Beier et al [70]. Briefly, the abdomen of each mosquito was separated from the rest of the body parts and triturated in 1x phosphate buffered saline (PBS) using a handheld motorised micro-pestle (DWK Life Sciences, Faust Laborbedrf AG, Schaffhausen, Switzerlan) made for grinding mosquitoes. A 96-well ELISA plate (Greiner Bio-One MicrolonTM, Monroe, North Carolina, USA) was coated with 50 µl of Mab solution at 4 µg/ml and incubated for 30 minutes. After incubation, the contents were aspirated, and the excess liquid was removed by tapping the plate on a tissue paper. The wells were then filled with 250 µl of blocking buffer (BB) and incubated for 1 hour. Following this, the buffer was drained, and 45 µl of BB was dispensed into each well. Next, 5 µl of each sample was loaded into the wells containing the 45 µl of BB and incubated for 2 hours at room temperature. Same procedures were followed for positive and negative controls. After incubation, the plate contents were aspirated, and the wells were washed three times with 250 µl of washing buffer (PBS + Tween 20). A 50 µl aliquot of the appropriate conjugate solution was then added to each well and incubated for 30 minutes at room temperature. The conjugate was removed by washing the wells four times with 250 µl of washing buffer (PBS+Tween 20). Finally, 100 µl of substrate solution (2,2’-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid, ABTS) was added to each well, followed by a 30-minute incubation at room temperature.
Data analysis
All data obtained were analysed using STATA package version 16 (Stata corp, College Station, TX).
DENV infection rate in mosquitoes
The infection rate was calculated by determining the proportion of DENV-positive mosquitoes among those tested by qRT-PCR. Usually, the minimum infection rate (MIR) and maximum infection rate (MaxIR) are computed as follows:
However, since MIR tends to underestimate and MaxIR overestimates infection rates, both are imprecise. Therefore, the Maximum Likelihood Estimate (MLE) with 95% confidance interval was used to provide a more accurate estimate.
where,
k = total number of mosquitoes tested
x = Number of positive pools
m = Number of mosquitoes per pool (assuming equal pool size)
n=total number of pools tested
Bloodmeal preference
A descriptive analysis was performed to compare the percentage of blood-fed mosquitoes across different hosts. The anthropophagy percentage was defined as the proportion of mosquitoes with human blood meals across all districts.
Host seeking preference
A descriptive analysis was performed to compare the proportion of host seeking mosquitoes collected indoors and outdoors. A negative binomial regression model was employed to determine if there was a statistically significant difference in host-seeking mosquitoes collected indoors versus those collected outdoors. The fix terms in the model were location (indoors vs outdoors), district and ward, while day and household were included as random effects. The models estimated the mean incidence rate ratios (IRR) and 95% confidence intervals around the means.
3. Results
DENV serotypes 1-4 prevalence
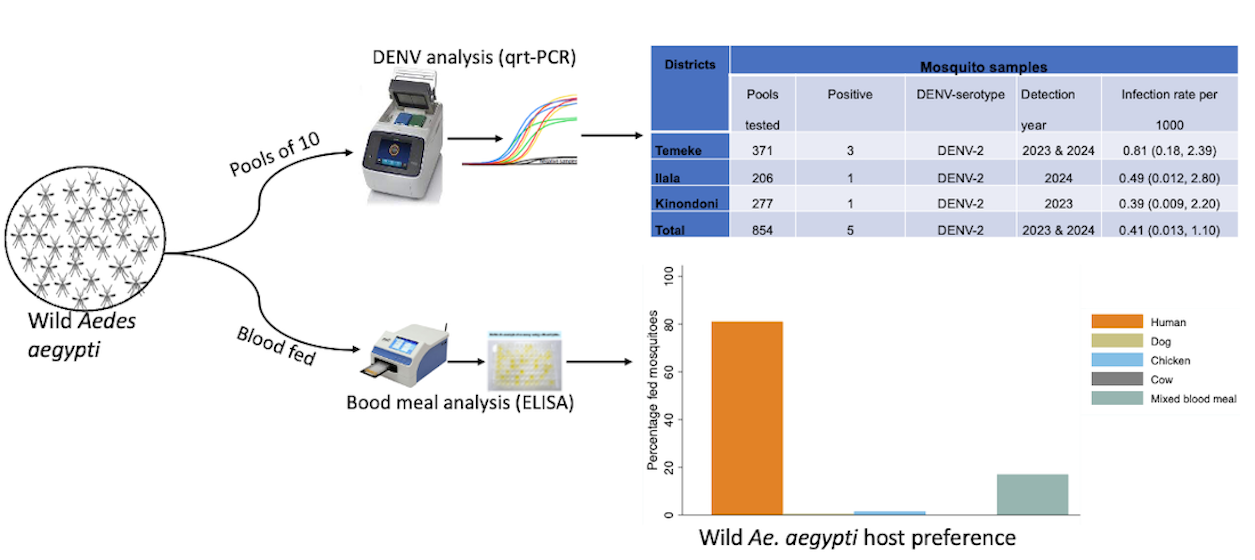
A total of 854 pools, with 10 mosquitoes per pool, were tested for DENV. Of these pools, 371 were from Temeke, 206 from Ilala and 277 from Kinondoni district (Table 2). DENV serotype 2 (DENV-2) was detected in all the three districts, with Temeke having the highest infection rate 0.81 per 1000 mosquitoes (Table 2). This likely indicates that the population in Temeke district is at a higher risk of contracting dengue fever compared to those in Ilala and Kinondoni districts. The viruses were detected in all years of mosquito sampling (Table 2). Indicating that the viruses are existing in the study area and not introduced.
Host preference
A total of 298 mosquito samples were tested for the origin of their blood meals from human, dog, chicken, and cow. Of these, 68.8% tested positive for either one or more blood meal sources (hosts) in the ELISA test, while 31.2% samples showed no reaction.
Aedes aegypti showed a strong preference for human blood, with approximately 166 mosquitoes (81.0%) feeding on humans, followed by chicken (1.5%) and dog (0.5%) (Figure 2). About 17% of the mosquitoes had taken a mixed blood meals from human and other hosts while none had fed on cow (Figure 2). The majority of blood-fed mosquitoes (69.3%) were collected using the Prockopack aspirator, followed by BGs trap 27.8% and GAT traps 2.9% (Table 3).
Host seeking Aedes aegypti mosquitoes
Using BGS traps, 354 female Ae. aegypti mosquitoes were collected from both indoor and outdoor locations. More than three quarters 78.5% (n=278) were caught outdoors, while 21.5% were collected indoors. The outdoors mosquito count was 4.33 higher than the indoor count (95%CI: [2.38-7.89], p-value<0.001) (Table 4). Among the three districts, Temeke had the higher count than Ilala and were statistically significant (Table 4).
4. Discussion
Understanding pathogens circulation in vectors is crucial for disease control. This study reports the presence of DENV-2 circulating in mosquitoes from Dar es Salaam city throughout the survey period, suggesting ongoing virus circulation rather than a new introduction. The virus serotype reported in this study is the same as the one detected in 2014 outbreak [53], highlighting the possibility that the virus has been persistent in the ecosystem since then. However, whole genome sequencing would be required to elucidate whether this is the case or if a new introduction of a different DENV-2 genotype occured.
All four DENV serotypes (DENV 1-4) have circulated in Tanzania [50], with different serotypes predominating in each outbreak. This shifting pattern may explain the increasing number of dengue cases and deaths during subsequent outbreaks [58], a trend also observed in West Africa [71] and other endemic regions [4]. In 2019, WHO reported a dramatic increase in dengue cases across several countries in Africa, particularly in the sub-Saharan region [72]. This rise reflects a broader global increase in dengue in all global WHO regions [6]. Despite the growing evidence, African countries including Tanzania, lack comprehensive data on the exact magnitude of dengue virus distribution due to limited epidemiological, entomological and virological surveillance, since dengue is a neglected tropical disease and African vector control efforts focus mainly on malaria.
Genotyping studies have shown a relatedness of dengue virus genotypes detected in East Africa, particularly Tanzania [53,59] and Kenya [73,74], to virus genotypes from Asian countries such as India and Singapore [55] as well as China [59]. This indicates that the viruses are being imported from the East via international travel and trade [59]. Africa’s rapid population growth and urbanisation will likely further accelerate virus spread. By 2050, nearly 60% of the continent’s population is expected to live in cities [75]. Increased human mobility and urbanisation will be inevitable, therefore, deliberate dengue monitoring efforts are needed. In this context, routine screening at national and international entry points could be implemented to reduce introductions of new virus genotypes, although this may be cost prohibitive.
This study identified Temeke, Ilala and Kinondoni districts as areas with dengue infected-mosquitoes, suggesting that these are priority areas for dengue xenomonitoring. The infected mosquitoes indicate a potential risk of dengue outbreaks in these areas, necessitating proactive Aedes mosquito surveillance. Additionally, it emphasizes the need for the government authorities to implement dengue control measures such as larval source reduction, targeted insecticide spraying, and educating citizens on larval control and the importance of seeking health care in case of potential dengue like symptoms. Furthermore, it highlights the need for the implementation of the International Health Regulations 2005 (IHR) to reduce further risk of transmiting viruses to other areas [76]. The IHR is a WHO legal framework for managing public health events and emergencies that have the potential to cross borders.
In Tanzania, dengue prevalence often exceeds 10% in human samples [48,59,60,77]. This study reports a minimal 0.08% dengue mosquito infection rate, lower than what was reported by Mboera et al [53] in 2014 during an outbreak. However, in that study, they collected larvae instead of adults, which might has overestimated the infection rates, given they do not represent the host-seeking population, and the samples could have been biased if siblings from the same transovarilly-infected egg batch were sampled. The findings from the present study align with those of Chilongola et al. [78] and Joseph et al. [79] in East Africa, Mojica et al. [80] in Nicaragua and Ecuador in Latin America, and Maneerattanasak et al. [81] in Southeast and South Asia. Similarly, this is consistent with reports on other female arthropod-transmitted diseases such as malaria [82,83], where Plasmodium infection rates in Anopheles mosquitoes are typically very low even in high endemic areas.
This study has demonstrated that, Ae. aegypti from Dar es Salaam are highly anthropophagic, with nearly more than 80% feeding on humans. This behaviour significantly increases dengue transmission risk, as human-mosquito contact is a key driver of virus spread [84]. The findings of this study are consistent with research from West Africa (Senegal and Burkina Faso) [41,85,86] as well as India [87], South East Asia (Thailand) [36], Australia [88], Latin America (Brazil, Equador and Peru) [89] and North America (USA) [90].
Aedes aegypti aegypti (Aaa) and Aedes aegypti formosus (Aaf) are the Ae. aegypti subspecies commonly found in Africa [91]. Aaa is considered an urban mosquito primarily responsible for urban dengue and yellow fever transmission [92]. Aaf inhabits peri-urban environments and serves as an agent for sylvatic dengue as well as yellow fever transmission [92,93], is less competent for dengue [94] and less anthropophilic [95]. In this study, we were unable to distinguish between the two subspecies. Given the lack of comprehensive research on their coexistence in the country, our findings emphasize the need for genomic studies to accurately characterize them.
As reported in other studies [40,41,85], human-animal mixed blood meals were frequently observed in this study, highlighting the feeding flexibility of Ae. aegypti. This mosquito species is known for transmitting dengue virus and other viruses, including yellow fever virus (YFV), Chikungunya virus (CHIKV) and Zika virus (ZIKV), all of which pose a substantial public health burden. While the ability of Ae. aegypti mosquitoes to feed on multiple animal species has implications for virus transmission, the role of animals as reservoirs for DENV, CHIKV and ZIKV in urban settings remains unlikely. While YFV does have non-human primates that serve as reservoir hosts [96], saving as a potential virus spillover source. Studies have described systems demonstrating the potential co-infection of DENV serotypes in an individual [97,98]. With all four DENV serotypes circulating in the country [50], it is likely that the mosquitoes’ ability to feed on multiple human hosts could essentially lead to individuals being infected with more than one virus serotype. This co-infection may result in complications, including dengue haemorrhagic fever (DHF) and dengue shock syndrome (DSS).
Moreover, the location where mosquitoes seek blood meals from hosts and rest after feeding has significant implications for the application of interventions to control mosquito vectors. This study demonstrated that more than three quarters of Ae. aegypti preferentially seek blood meals outdoors, highlighting a challenge in controlling this vector because the majority of mosquito control interventions, particularly those developed for malaria control, are applied indoors, targeting indoor host seeking and resting mosquitoes. This finding is similar to studies of Ae. aegypti in Burkina Faso [99,100] and Ghana [101]. This indicates that larval source reduction [102] might be the most likely intervention to be successful at the community level. In addition, the release of Wolbachia has shown excellent efficacy in preventing dengue when deployed at a city scale [103]. Further work is ongoing to measure the resting behaviour and susceptibility of the Dar es Salaam population of Ae. aegypti to insecticides used for mosquito control.
Study Limitations
To fully understand the indoor and outdoor ecology of Ae. aegypti, traps targeting host-seeking, resting and oviposition behaviours need to be deployed concurrently both indoors and outdoors. However, we were unable to collect indoor mosquitoes using GATs because of the smell of the infusion, and with Prokopack aspirators due to house entry restrictions, as most of the time the residents were absent. As a result, we collected mosquitoes with only BGS, a trap type designated for collecting host-seeking mosquitoes. Therefore, we recommend that for future studies, the indoor and outdoor mosquito collection should also include resting collections because the use of traps with a lure may bias the collections towards human-fed mosquitoes [89]. Additionally, blood meal analysis for host preference was performed on only four hosts (humans, dog, chicken and cow). However, we found that 93 samples did not react suggesting that the mosquitoes may have contained blood meals from hosts not included in the analysis. In Kenya, Ae aegypti has been found to feed on goats, rats and cats [104], which were also present in the present study site, but not tested for. Therefore, future studies should include a broader range of potential hosts.
5. Conclusions
This study confirms the circulation of DENV-2 in the mosquito population in Dar es Salaam, highlighting the risk of a potential dengue outbreak in Tanzania. Dar es Salaam is one of Africa’s major metropolitan cities, with a population of nearly six million. It serves as the economic hub of Tanzania, so it experiences a significant influx of local and international travellers. The presence of DENV-2 in mosquitoes as well as the strong human feeding preference of Ae. aegypti indicates the potential risk of DENV transmission to humans. These findings emphasise the need for enhanced surveillance and targeted proactive vector control measures including removal of breeding sites to mitigate dengue outbreaks.
6. Patents
Author Contributions
Conceptualisation: FSCT and SJM; Data curation: FSCT, OD, MH, and HM; Formal analysis: FSCT; Investigation: FSCT, OD, MH, HM, SH, LDB, JM, JJM, NSL and TGM; Methodology: FSCT, PM and SJM; Supervision; SJM; Writing original draft: FSCT; Visualization: SJM, PM, SH and LMH; Writing review and editing SJM, PM, SH and LMH; Funding acquisition: SJM. All authors read and approved the final manuscript draft.
Funding
This study was funded by Rudolf Geigy–Stiftung (RGS) and Armasuisse through Swiss TPH. The findings and conclusions contained within are those of the authors, herein stated is Ifakara Health Institute and Swiss TPH, and do not necessarily reflect positions or policies of the Rudolf Geigy Foundation and and Armasuisse.
Institutional Review Board Statement
This study received an ethical approval from Ifakara Health Institute Review Board (IHI-IRB) IHI/IRB/No: 7-2021 and National Institute for Medical Research Review Board (NIMR-RB) No. NIMR/HQ/R.8a/Vol. IX/3641.
Data Availability Statement
The datasets generated during the study are available from Ifakara Health Institute and the corresponding author on reasonable request.
Acknowledgments
The authors would like to thank Ifakara Health Institute staff for their support. Also, we would like to extend our heartfelt thanks to the RGS for funding this study and the excellent support from Dr Lukas Meier at RGS.
Conflicts of Interest
The authors declare that they have no competing interests.
Consent for publication
All authors contributed to the manuscript and approved the final manuscript. The National Institute for Medical Research Review Board (NIMR-RB) gave permission for the publication of results Ref No. BD.242/437/01C/127.
Abbreviations
| BGS | Biogent sentinel trap |
| GAT | Gravid Aedes trap |
| DENV | Dengue virus |
| DHF | Dengue haemorrhagic fever |
| DSS | Dengue shock syndrome |
| ITN | Insecticide treated net |
| IRS | Indoor residual spray |
| DF | Dengue fever |
| YF | Yellow fever |
| CHIK | Chikungunya |
| ZIK | Zika |
| IHR | International health regulations |
| WHO | World health organisation |
| IRR | Incidence rate ratio |
| OR | Odds ratio |
| ELISA | Enzyme-linked immunosorbent assay. |
| qRT-PCR | Quantitative reverse transcriptase polymerase chain reaction |
| MIR | Mosquito infection rate |
References
- Guzman, A.; Istúriz, R.E. Update on the global spread of dengue. Int. J. Antimicrob. Agents 2010, 36, S40–S42. [Google Scholar] [CrossRef] [PubMed]
- Brady OJ, Gething PW, Bhatt S, Messina JP, Brownstein JS, Hoen AG, et al. Refining the global spatial limits of dengue virus transmission by evidence-based consensus. PlosNegl Trop Dis. 2012. [CrossRef]
- Control ECfDPa. Dengue cases January-December 2023. 2023.
- World Health Organization. Dengue - Global situation. Geneva, Switzerland. 2024.
- Control ECfDPa. Dengue worldwide overview. 2024.
- Who. Dengue and severe dengue. World Health Organisation, Geneva, Switzerland. 2024.
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; et al. The global distribution and burden of dengue. Nature. 2013, 496, 504–7. [Google Scholar] [CrossRef] [PubMed]
- Waggoner, J.J.; Gresh, L.; Vargas, M.J.; Ballesteros, G.; Tellez, Y.; Soda, K.J.; et al. Viremia and clinical presentation in Nicaraguan patients infected with Zika virus, chikungunya virus, and dengue virus. Clin Infect Dis. 2016, 589. [Google Scholar] [CrossRef]
- World Health Organization. Dengue - Global situation. Geneva, Switzerland. 2023.
- World Health Organization. Dengue and severe dengue. Geneva, Switzerland. 2022.
- Boillat-Blanco, N.; Klaassen, B.; Mbarack, Z.; Samaka, J.; Mlaganile, T.; Masimba, J.; Franco Narvaez, L.; Mamin, A.; Genton, B.; Kaiser, L. Dengue fever in Dar es Salaam, Tanzania: clinical features and outcome in populations of black and non-black racial category. BMC Infect Dis. 2018, 18, 644. [Google Scholar] [CrossRef]
- Guzman, M.G.; Harris, E. Dengue. Lancet 2015, 385, 453–465. [Google Scholar] [CrossRef]
- Messina, J.P.; Brady, O.J.; Golding, N.; Kraemer, M.U.; Wint, G.; Ray, S.E.; Pigott, D.M.; Shearer, F.M.; Johnson, K.; Earl, L. The current and future global distribution and population at risk of dengue. Nat. Microbiol. 2019, 4, 1508–1515. [Google Scholar] [CrossRef]
- Bennett, S.N. 17 Taxonomy and Evolutionary Relationships of Flaviviruses. Dengue and dengue hemorrhagic fever 2014, 322. [Google Scholar]
- Guzman, M.G.; Halstead, S.B.; Artsob, H.; Buchy, P.; Farrar, J.; Gubler, D.J.; Hunsperger, E.; Kroeger, A.; Margolis, H.S.; Martínez, E. Dengue: a continuing global threat. Nat. Rev. Microbiol. 2010, 8, S7–S16. [Google Scholar] [CrossRef]
- Harapan, H.; Michie, A.; Sasmono, R.T.; Imrie, A. Dengue: a minireview. Viruses 2020, 12, 829. [Google Scholar] [CrossRef]
- Najri, N.; Mazlan, Z.; Jaimin, J.; Mohammad, R.; Yusuf, N.M.; Kumar, V.S.; Hoque, M. Genotypes of the dengue virus in patients with dengue infection from Sabah, Malaysia. Journal of Phys. Conference Series; 2019.
- Holmes, E.C.; Twiddy, S.S. The origin, emergence and evolutionary genetics of dengue virus. Infect. Genet. Evol. 2003, 3, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Sasmono, R.T.; Wahid, I.; Trimarsanto, H.; Yohan, B.; Wahyuni, S.; Hertanto, M.; Yusuf, I.; Mubin, H.; Ganda, I.J.; Latief, R. Genomic analysis and growth characteristic of dengue viruses from Makassar, Indonesia. Infect. Genet. Evol. 2015, 32, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Rodenhuis-Zybert, I.A.; Wilschut, J.; Smit, J.M. Dengue virus life cycle: viral and host factors modulating infectivity. Cell. Mol. Life Sci. 2010, 67, 2773–2786. [Google Scholar] [CrossRef] [PubMed]
- Guzman, M.G.; Alvarez, M.; Halstead, S.B. Secondary infection as a risk factor for dengue hemorrhagic fever/dengue shock syndrome: an historical perspective and role of antibody-dependent enhancement of infection. Arch. Virol. 2013, 158, 1445–1459. [Google Scholar] [CrossRef]
- Guzman, M.G.; Gubler, D.J.; Izquierdo, A.; Martinez, E.; Halstead, S.B. Dengue infection. Nat. Rev. Dis. Primers 2016, 2, 1–25. [Google Scholar] [CrossRef]
- Vaughn, D.W.; Green, S.; Kalayanarooj, S.; Innis, B.L.; Nimmannitya, S.; Suntayakorn, S.; Endy, T.P.; Raengsakulrach, B.; Rothman, A.L.; Ennis, F.A. Dengue viremia titer, antibody response pattern, and virus serotype correlate with disease severity. J. Infect. Dis. 2000, 181, 2–9. [Google Scholar] [CrossRef]
- Moi, M.L.; Takasaki, T.; Omatsu, T.; Nakamura, S.; Katakai, Y.; Ami, Y.; Suzaki, Y.; Saijo, M.; Akari, H.; Kurane, I. Demonstration of marmosets (Callithrix jacchus) as a non-human primate model for secondary dengue virus infection: high levels of viraemia and serotype cross-reactive antibody responses consistent with secondary infection of humans. J. Gen. Virol. 2014, 95, 591–600. [Google Scholar] [CrossRef]
- Amarasinghe, A.; Kuritsky, J.N.; Letson, G.W.; Margolis, H.S. Dengue virus infection in Africa. Emerg. Infect. Dis. 2011, 17, 1349. [Google Scholar] [CrossRef]
- Sang RC, editor Dengue in Africa. Report of the scientific working group meeting on dengue Geneva: WHO Special Programme for Research and Training in Tropical Diseases. 2007.
- Chipwaza, B.; Mugasa, J.P.; Selemani, M.; Amuri, M.; Mosha, F.; Ngatunga, S.D.; Gwakisa, P.S. Dengue and Chikungunya fever among viral diseases in outpatient febrile children in Kilosa district hospital, Tanzania. PLoS Neglected Trop. Dis. 2014, 8, e3335. [Google Scholar] [CrossRef]
- Parreira, R.; Centeno-Lima, S.; Lopes, A.; Portugal-Calisto, D.; Constantino, A.; Nina, J. Dengue virus serotype 4 and chikungunya virus coinfection in a traveller returning from Luanda, Angola, January 2014. Eurosurveillance 2014, 19, 20730. [Google Scholar] [CrossRef]
- Gautret, P.; Simon, F.; Askling, H.H.; Bouchaud, O.; Leparc-Goffart, I.; Ninove, L.; Parola, P. Dengue type 3 virus infections in European travellers returning from the Comoros and Zanzibar, February-April 2010. Eurosurveillance 2010, 15, 19541. [Google Scholar] [CrossRef] [PubMed]
- Gubler, D.J.; Nalim, S.; Tan, R.; Saipan, H. Variation in susceptibility to oral infection with dengue viruses among geographic strains of Aedes aegypti. Am. J. Trop. Med. Hyg. 1979, 28, 1045–1052. [Google Scholar] [CrossRef] [PubMed]
- Rosen, L.; Roseboom, L.E.; Gubler, D.J.; Lien, J.C.; Chaniotis, B.N. Comparative susceptibility of mosquito species and strains to oral and parenteral infection with dengue and Japanese encephalitis viruses. Am. J. Trop. Med. Hyg. 1985, 34, 603–615. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, P.M.; Rico-Hesse, R. Efficiency of dengue serotype 2 virus strains to infect and disseminate in Aedes aegypti. Am. J. Trop. Med. Hyg. 2003, 68, 539. [Google Scholar] [CrossRef]
- Leta, S.; Beyene, T.J.; De Clercq, E.M.; Amenu, K.; Kraemer, M.U.; Revie, C.W. Global risk mapping for major diseases transmitted by Aedes aegypti and Aedes albopictus. Int. J. Infect. Dis. 2018, 67, 25–35. [Google Scholar] [CrossRef]
- Brady, O.J.; Hay, S.I. The Global Expansion of Dengue: How Aedes aegypti Mosquitoes Enabled the First Pandemic Arbovirus. Annu Rev Entomol 2020, 65, 191–208. [Google Scholar] [CrossRef]
- Sridhar, S.; Luedtke, A.; Langevin, E.; Zhu, M.; Bonaparte, M.; Machabert, T.; Savarino, S.; Zambrano, B.; Moureau, A.; Khromava, A. Effect of dengue serostatus on dengue vaccine safety and efficacy. New Engl. J. Med. 2018, 379, 327–340. [Google Scholar] [CrossRef]
- Ponlawat, A.; Harrington, L.C. Blood feeding patterns of Aedes aegypti and Aedes albopictus in Thailand. J. Med. Entomol. 2005, 42, 844–849. [Google Scholar] [CrossRef]
- Mann, J.G.; Washington, M.; Guynup, T.; Tarrand, C.; Dewey, E.M.; Fredregill, C.; Duguma, D.; Pitts, R.J. Feeding habits of vector mosquitoes in Harris County, TX, 2018. J. Med. Entomol. 2020, 57, 1920–1929. [Google Scholar] [CrossRef]
- Diallo, D.; Chen, R.; Diagne, C.T.; Ba, Y.; Dia, I.; Sall, A.A.; Weaver, S.C.; Diallo, M. Bloodfeeding patterns of sylvatic arbovirus vectors in southeastern Senegal. Trans. R. Soc. Trop. Med. Hyg. 2013, 107, 200–203. [Google Scholar] [CrossRef]
- Sivan, A.; Shriram, A.; Sunish, I.; Vidhya, P. Host-feeding pattern of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) in heterogeneous landscapes of South Andaman, Andaman and Nicobar Islands, India. Parasitol. Res. 2015, 114, 3539–3546. [Google Scholar] [CrossRef] [PubMed]
- Olson, M.F.; Ndeffo-Mbah, M.L.; Juarez, J.G.; Garcia-Luna, S.; Martin, E.; Borucki, M.K.; Frank, M.; Estrada-Franco, J.G.; Rodríguez-Pérez, M.A.; Fernández-Santos, N.A. High rate of non-human feeding by Aedes aegypti reduces Zika virus transmission in South Texas. Viruses 2020, 12, 453. [Google Scholar] [CrossRef] [PubMed]
- Sene, N.M.; Diouf, B.; Gaye, A.; Gueye, A.; Seck, F.; Diagne, C.T.; Dia, I.; Diallo, D.; Diallo, M. Blood feeding patterns of Aedes aegypti populations in Senegal. Am. J. Trop. Med. Hyg. 2022, 106, 1402. [Google Scholar] [CrossRef] [PubMed]
- Pruszynski, C.A.; Stenn, T.; Acevedo, C.; Leal, A.L.; Burkett-Cadena, N.D. Human blood feeding by Aedes aegypti (Diptera: Culicidae) in the Florida Keys and a review of the literature. J. Med. Entomol. 2020, 57, 1640–1647. [Google Scholar] [CrossRef]
- Zahid, M.H.; Van Wyk, H.; Morrison, A.C.; Coloma, J.; Lee, G.O.; Cevallos, V.; Ponce, P.; Eisenberg, J.N. The biting rate of Aedes aegypti and its variability: A systematic review (1970–2022). PLoS Neglected Trop. Dis. 2023, 17, e0010831. [Google Scholar] [CrossRef]
- Martin, E.; Medeiros, M.C.; Carbajal, E.; Valdez, E.; Juarez, J.G.; Garcia-Luna, S.; Salazar, A.; Qualls, W.A.; Hinojosa, S.; Borucki, M.K. Surveillance of Aedes aegypti indoors and outdoors using Autocidal Gravid Ovitraps in South Texas during local transmission of Zika virus, 2016 to 2018. Acta Trop. 2019, 192, 129–137. [Google Scholar] [CrossRef]
- Dalpadado, R.; Amarasinghe, D.; Gunathilaka, N.; Ariyarathna, N. Bionomic aspects of dengue vectors Aedes aegypti and Aedes albopictus at domestic settings in urban, suburban and rural areas in Gampaha District, Western Province of Sri Lanka. Parasites Vectors 2022, 15, 148. [Google Scholar] [CrossRef]
- Chadee, D.D.; Martinez, R. Landing periodicity of Aedes aegypti with implications for dengue transmission in Trinidad, West Indies. J. Soc. Vector Ecol. 2000, 25, 158–163. [Google Scholar]
- Vairo, F.; Nicastri, E.; Meschi, S.; Schepisi, M.S.; Paglia, M.G.; Bevilacqua, N.; Mangi, S.; Sciarrone, M.R.; Chiappini, R.; Mohamed, J. Seroprevalence of dengue infection: a cross-sectional survey in mainland Tanzania and on Pemba Island, Zanzibar. Int. J. Infect. Dis. 2012, 16, e44–e46. [Google Scholar] [CrossRef]
- Vairo, F.; Nicastri, E.; Yussuf, S.M.; Cannas, A.; Meschi, S.; Mahmoud, M.A.; Mohamed, A.H.; Maiko, P.M.; De Nardo, P.; Bevilacqua, N. IgG against dengue virus in healthy blood donors, Zanzibar, Tanzania. Emerg. Infect. Dis. 2014, 20, 465. [Google Scholar] [CrossRef]
- Moi, M.L.; Takasaki, T.; Kotaki, A.; Tajima, S.; Lim, C.-K.; Sakamoto, M.; Iwagoe, H.; Kobayashi, K.; Kurane, I. Importation of dengue virus type 3 to Japan from Tanzania and Côte d’Ivoire. Emerg. Infect. Dis. 2010, 16, 1770. [Google Scholar] [CrossRef] [PubMed]
- Chipwaza B, Sumaye RD, Weisser M, Gingo W, Yeo NK-W, Amrun SN, et al., editors. Occurrence of 4 dengue virus serotypes and chikungunya virus in Kilombero Valley, Tanzania, during the dengue outbreak in 2018. Open forum infectious diseases; 2021: Oxford Univ Press US.
- Hertz, J.T.; Munishi, O.M.; Ooi, E.E.; Howe, S.; Lim, W.Y.; Chow, A.; Morrissey, A.B.; Bartlett, J.A.; Onyango, J.J.; Maro, V.P. Chikungunya and dengue fever among hospitalized febrile patients in northern Tanzania. Am. J. Trop. Med. Hyg. 2012, 86, 171. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Dengue outbreak in the United Republic of Tanzania. 2014.
- Mboera, L.E.; Mweya, C.N.; Rumisha, S.F.; Tungu, P.K.; Stanley, G.; Makange, M.R.; Misinzo, G.; De Nardo, P.; Vairo, F.; Oriyo, N.M. The risk of dengue virus transmission in Dar es Salaam, Tanzania during an epidemic period of 2014. PLoS Neglected Trop. Dis. 2016, 10, e0004313. [Google Scholar] [CrossRef]
- Kelly, M.E.; Msafiri, F.; Affara, M.; Gehre, F.; Moremi, N.; Mghamba, J.; Misinzo, G.; Thye, T.; Gatei, W.; Whistler, T. Molecular Characterization and Phylogenetic Analysis of Dengue Fever Viruses in Three Outbreaks in Tanzania Between 2017 and 2019. PLoS Neglected Trop. Dis. 2023, 17, e0011289. [Google Scholar] [CrossRef]
- Okada, K.; Morita, R.; Egawa, K.; Hirai, Y.; Kaida, A.; Shirano, M.; Kubo, H.; Goto, T.; Yamamoto, S.P. Dengue virus type 1 infection in traveler returning from Tanzania to Japan, 2019. Emerg. Infect. Dis. 2019, 25, 1782. [Google Scholar] [CrossRef]
- Mwanyika, G.O.; et al. Co-circulation of Dengue Virus Serotypes 1 and 3 during the 2019 epidemic in Dar es Salaam, Tanzania. PloS Negl Trop Dis. 2019.
- Mwanyika, G.O.; Sindato, C.; Rugarabamu, S.; Rumisha, S.F.; Karimuribo, E.D.; Misinzo, G.; Rweyemamu, M.M.; Hamid, M.M.A.; Haider, N.; Vairo, F. Seroprevalence and associated risk factors of chikungunya, dengue, and Zika in eight districts in Tanzania. Int. J. Infect. Dis. 2021, 111, 271–280. [Google Scholar] [CrossRef]
- SACIDS. Dengue Outbreaks in Tanzania: Recent Trends and Importance of Research Data in Disease Surveillance. Morogoro, Tanzania: Southern African Centre for Infectious Disease Surveillance; 2019.
- Vairo, F.; Mboera, L.E.; De Nardo, P.; Oriyo, N.M.; Meschi, S.; Rumisha, S.F.; Colavita, F.; Mhina, A.; Carletti, F.; Mwakapeje, E. Clinical, virologic, and epidemiologic characteristics of dengue outbreak, Dar es Salaam, Tanzania, 2014. Emerg. Infect. Dis. 2016, 22, 895. [Google Scholar] [CrossRef]
- Shauri, H.S.; Ngadaya, E.; Senkoro, M.; Buza, J.J.; Mfinanga, S. Seroprevalence of Dengue and Chikungunya antibodies among blood donors in Dar es Salaam and Zanzibar, Tanzania: a cross-sectional study. BMC Infect. Dis. 2021, 21, 1–6. [Google Scholar] [CrossRef]
- Kajeguka, D.C.; Kaaya, R.D.; Mwakalinga, S.; Ndossi, R.; Ndaro, A.; Chilongola, J.O.; Mosha, F.W.; Schiøler, K.L.; Kavishe, R.A.; Alifrangis, M. Prevalence of dengue and chikungunya virus infections in north-eastern Tanzania: a cross sectional study among participants presenting with malaria-like symptoms. BMC Infect. Dis. 2016, 16, 1–9. [Google Scholar]
- National Bureau of Statistics Ministry of Finance and Office of Chief Government Statistician President’s Office, F., Economy and Development Planning. Population and Housing Census. Administrative units Population Distribution and Age and Sex Distribution Report Tanzania- volume1a. 2022.
- Msellemu, D.; Gavana, T.; Ngonyani, H.; Mlacha, Y.P.; Chaki, P.; Moore, S.J. Knowledge, attitudes and bite prevention practices and estimation of productivity of vector breeding sites using a Habitat Suitability Score (HSS) among households with confirmed dengue in the 2014 outbreak in Dar es Salaam, Tanzania. PLoS Neglected Trop. Dis. 2020, 14, e0007278. [Google Scholar] [CrossRef] [PubMed]
- Machange, J.J.; Maasayi, M.S.; Mundi, J.; Moore, J.; Muganga, J.B.; Odufuwa, O.G.; Moore, S.J.; Tenywa, F.C. Comparison of the Trapping Efficacy of Locally Modified Gravid Aedes Trap and Autocidal Gravid Ovitrap for the Monitoring and Surveillance of Aedes aegypti Mosquitoes in Tanzania. Insects 2024, 15, 401. [Google Scholar] [CrossRef] [PubMed]
- Wilkerson, R.C.; Linton, Y.-M.; Strickman, D. Mosquitoes of the World; Johns Hopkins University Press: 2021.
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L. The MIQE Guidelines: M inimum I nformation for Publication of Q uantitative Real-Time PCR E xperiments. Oxford University Press: 2009.
- Pérez-Castro, R.; Castellanos, J.E.; Olano, V.A.; Matiz, M.I.; Jaramillo, J.F.; Vargas, S.L.; Sarmiento, D.M.; Stenström, T.A.; Overgaard, H.J. Detection of all four dengue serotypes in Aedes aegypti female mosquitoes collected in a rural area in Colombia. Mem. Do Inst. Oswaldo Cruz 2016, 111, 233–240. [Google Scholar] [CrossRef]
- Balingit, J.C.; Carvajal, T.M.; Saito-Obata, M.; Gamboa, M.; Nicolasora, A.D.; Sy, A.K.; Oshitani, H.; Watanabe, K. Surveillance of dengue virus in individual Aedes aegypti mosquitoes collected concurrently with suspected human cases in Tarlac City, Philippines. Parasites Vectors 2020, 13, 1–13. [Google Scholar] [CrossRef]
- Beier, J.C.; Perkins, P.V.; Wirtz, R.A.; Koros, J.; Diggs, D.; Gargan, T.P.; Koech, D.K. Bloodmeal identification by direct enzyme-linked immunosorbent assay (ELISA), tested on Anopheles (Diptera: Culicidae) in Kenya. J. Med. Entomol. 1988, 25, 9–16. [Google Scholar] [CrossRef]
- Gyasi, P.; Bright Yakass, M.; Quaye, O. Analysis of dengue fever disease in West Africa. Exp. Biol. Med. 2023, 248, 1850–1863. [Google Scholar] [CrossRef]
- World Health Organization. Dengue and Severe Dengue Fact Sheet. Geneva, Switzerland. 2019.
- Masika, M.M.; Korhonen, E.M.; Smura, T.; Uusitalo, R.; Vapalahti, K.; Mwaengo, D.; Jääskeläinen, A.J.; Anzala, O.; Vapalahti, O.; Huhtamo, E. Detection of dengue virus type 2 of Indian origin in acute febrile patients in rural Kenya. PLoS Neglected Trop. Dis. 2020, 14, e0008099. [Google Scholar] [CrossRef]
- Langat, S.K.; Eyase, F.L.; Berry, I.M.; Nyunja, A.; Bulimo, W.; Owaka, S.; Ofula, V.; Limbaso, S.; Lutomiah, J.; Jarman, R. Origin and evolution of dengue virus type 2 causing outbreaks in Kenya: Evidence of circulation of two cosmopolitan genotype lineages. Virus Evol. 2020, 6, veaa026. [Google Scholar] [CrossRef]
- Bank, A.D. Urbanization in Africa. 2012.
- World Health Organization. Urban yellow fever risk management: preparedness and response: Handbook for national operational planning. Geneva, Switzerland. 2023.
- Mwanyika, G.; Mboera, L.E.; Rugarabamu, S.; Lutwama, J.; Sindato, C.; Paweska, J.T.; Misinzo, G. Co-circulation of Dengue Virus Serotypes 1 and 3 during the 2019 epidemic in Dar es Salaam, Tanzania. bioRxiv 2019, 763003. [Google Scholar]
- Chilongola, J.O.; Mwakapuja, R.S.; Horumpende, P.G.; Vianney, J.-M.; Shabhay, A.; Mkumbaye, S.I.; Semvua, H.S.; Mmbaga, B.T. Concurrent Infection With Dengue and Chikungunya Viruses in Humans and Mosquitoes: A Field Survey in Lower Moshi, Tanzania. East Afr. Sci. 2022, 4, 78–86. [Google Scholar] [CrossRef]
- Joseph, N.K.; Mumo, E.; Morlighem, C.; Macharia, P.M.; Snow, R.W.; Linard, C. Mosquito-borne diseases in urban East African Community region: a scoping review of urban typology research and mosquito genera overlap, 2000-2024. Front. Trop. Dis. 2024, 5, 1499520. [Google Scholar] [CrossRef]
- Mojica, J.; Arévalo, V.; Juarez, J.G.; Galarza, X.; Gonzalez, K.; Carrazco, A.; Suazo, H.; Harris, E.; Coloma, J.; Ponce, P. A numbers game: mosquito-based arbovirus surveillance in two distinct geographic regions of Latin America. J. Med. Entomol. 2025, 62, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Maneerattanasak, S.; Ngamprasertchai, T.; Tun, Y.M.; Ruenroengbun, N.; Auewarakul, P.; Boonnak, K. Prevalence of dengue, Zika, and chikungunya virus infections among mosquitoes in Asia: A systematic review and meta-analysis. Int. J. Infect. Dis. 2024, 107226. [Google Scholar] [CrossRef]
- Mapua, S.A.; Hape, E.E.; Kihonda, J.; Bwanary, H.; Kifungo, K.; Kilalangongono, M.; Kaindoa, E.W.; Ngowo, H.S.; Okumu, F.O. Persistently high proportions of plasmodium-infected Anopheles funestus mosquitoes in two villages in the Kilombero valley, South-Eastern Tanzania. Parasite Epidemiol. Control 2022, 18, e00264. [Google Scholar] [CrossRef]
- Lwetoijera, D.W.; Harris, C.; Kiware, S.S.; Dongus, S.; Devine, G.J.; McCall, P.J.; Majambere, S. Increasing role of Anopheles funestus and Anopheles arabiensis in malaria transmission in the Kilombero Valley, Tanzania. Malar. J. 2014, 13, 1–10. [Google Scholar] [CrossRef]
- Macdonald, G. Epidemiologic models in studies of vetor-borne diseases: The re dyer lecture. Public Health Rep. 1961, 76, 753. [Google Scholar] [CrossRef]
- Diouf, B.; Sene, N.M.; Ndiaye, E.H.; Gaye, A.; Ngom, E.H.M.; Gueye, A.; Seck, F.; Diagne, C.T.; Dia, I.; Diallo, M. Resting behavior of blood-fed females and host feeding preferences of Aedes aegypti (Diptera: Culicidae) morphological forms in Senegal. J. Med. Entomol. 2021, 58, 2467–2473. [Google Scholar] [CrossRef]
- Badolo, A.; Sombié, A.; Yaméogo, F.; Wangrawa, D.W.; Sanon, A.; Pignatelli, P.M.; Sanon, A.; Viana, M.; Kanuka, H.; Weetman, D. First comprehensive analysis of Aedes aegypti bionomics during an arbovirus outbreak in west Africa: Dengue in Ouagadougou, Burkina Faso, 2016–2017. PLOS Neglected Trop. Dis. 2022, 16, e0010059. [Google Scholar] [CrossRef]
- Sivan, A.; Shriram, A.N.; Sunish, I.P.; Vidhya, P.T. Host-feeding pattern of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) in heterogeneous landscapes of South Andaman, Andaman and Nicobar Islands, India. Parasitol Res 2015, 114, 3539–3546. [Google Scholar] [CrossRef]
- Jansen, C.C.; Webb, C.E.; Graham, G.C.; Craig, S.B.; Zborowski, P.; Ritchie, S.A.; Russell, R.C.; van den Hurk, A.F. Blood sources of mosquitoes collected from urban and peri-urban environments in eastern Australia with species-specific molecular analysis of avian blood meals. Am J Trop Med Hyg 2009, 81, 849–857. [Google Scholar] [CrossRef] [PubMed]
- Melgarejo-Colmenares, K.; Cardo, M.V.; Vezzani, D. Blood feeding habits of mosquitoes: hardly a bite in South America. Parasitol Res 2022, 121, 1829–1852. [Google Scholar] [CrossRef] [PubMed]
- Pruszynski, C.A.; Stenn, T.; Acevedo, C.; Leal, A.L.; Burkett-Cadena, N.D. Human Blood Feeding by Aedes aegypti (Diptera: Culicidae) in the Florida Keys and a Review of the Literature. J Med Entomol 2020, 57, 1640–1647. [Google Scholar] [CrossRef] [PubMed]
- Powell JR, Tabachnick WJ. History of domestication and spread of Aedes aegypti-a review. Memórias do Instituto Oswaldo Cruz. 2013;108(suppl 1):11-7.
- Diallo, M.; Sall, A.A.; Moncayo, A.C.; Ba, Y.; Fernandez, Z.; Ortiz, D.; Coffey, L.L.; Mathiot, C.; Tesh, R.B.; Weaver, S.C. Potential role of sylvatic and domestic African mosquito species in dengue emergence. Am. J. Trop. Med. Hyg. 2005, 73, 445–449. [Google Scholar] [CrossRef]
- Valentine, M.J.; Murdock, C.C.; Kelly, P.J. Sylvatic cycles of arboviruses in non-human primates. Parasit Vectors 2019, 12, 1–18. [Google Scholar] [CrossRef]
- Chepkorir, E.; Lutomiah, J.; Mutisya, J.; Mulwa, F.; Limbaso, K.; Orindi, B.; Ng'ang'a, Z.; Sang, R. Vector competence of Aedes aegypti populations from Kilifi and Nairobi for dengue 2 virus and the influence of temperature. Parasit Vectors 2014, 7, 435. [Google Scholar] [CrossRef]
- Gouck, H.K. Host preferences of various strains of Aedes aegypti and A. simpsoni as determined by an olfactometer. Bull World Health Organ 1972, 47, 680–683. [Google Scholar]
- Monath, T.P.; Vasconcelos, P.F. Yellow fever. J. Clin. Virol. 2015, 64, 160–173. [Google Scholar] [CrossRef]
- Esteva, L.; Vargas, C. Coexistence of different serotypes of dengue virus. J. Math. Biol. 2003, 46, 31–47. [Google Scholar] [CrossRef]
- Feng, Z.; Velasco-Hernández, J.X. Competitive exclusion in a vector-host model for the dengue fever. J. Math. Biol. 1997, 35, 523–544. [Google Scholar] [CrossRef]
- Ouédraogo, W.M.; Zanré, N.; Sombié, A.; Yameogo, F.; Gnémé, A.; Sanon, A.; Costantini, C.; Kanuka, H.; Viana, M.; Weetman, D.; et al. Blood-Feeding Patterns and Resting Behavior of Aedes aegypti from Three Health Districts of Ouagadougou City, Burkina Faso. Am J Trop Med Hyg 2024, 111, 1295–1301. [Google Scholar] [CrossRef] [PubMed]
- Badolo, A.; Sombié, A.; Yaméogo, F.; Wangrawa, D.W.; Sanon, A.; Pignatelli, P.M.; Sanon, A.; Viana, M.; Kanuka, H.; Weetman, D.; McCall, P.J. First comprehensive analysis of Aedes aegypti bionomics during an arbovirus outbreak in west Africa: Dengue in Ouagadougou, Burkina Faso, 2016-2017. PLoS Negl Trop Dis 2022, 16, e0010059. [Google Scholar] [CrossRef] [PubMed]
- Captain-Esoah, M.; Kweku Baidoo, P.; Frempong, K.K.; Adabie-Gomez, D.; Chabi, J.; Obuobi, D.; Kwame Amlalo, G.; Balungnaa Veriegh, F.; Donkor, M.; Asoala, V.; et al. Biting Behavior and Molecular Identification of Aedes aegypti (Diptera: Culicidae) Subspecies in Some Selected Recent Yellow Fever Outbreak Communities in Northern Ghana. J Med Entomol 2020, 57, 1239–1245. [Google Scholar] [CrossRef] [PubMed]
- Montenegro-Quiñonez, C.A.; Louis, V.R.; Horstick, O.; Velayudhan, R.; Dambach, P.; Runge-Ranzinger, S. Interventions against Aedes/dengue at the household level: a systematic review and meta-analysis. eBioMedicine 2023, 93. [Google Scholar] [CrossRef]
- Utarini, A.; Indriani, C.; Ahmad, R.A.; Tantowijoyo, W.; Arguni, E.; Ansari, M.R.; Supriyati, E.; Wardana, D.S.; Meitika, Y.; Ernesia, I.; et al. Efficacy of Wolbachia-Infected Mosquito Deployments for the Control of Dengue. N Engl J Med 2021, 384, 2177–2186. [Google Scholar] [CrossRef]
- Agha, S.B.; Tchouassi, D.P.; Turell, M.J.; Bastos, A.D.S.; Sang, R. Entomological assessment of dengue virus transmission risk in three urban areas of Kenya. PLoS Negl Trop Dis 2019, 13, e0007686. [Google Scholar] [CrossRef]
Figure 1.
Study sites of the mosquito collection. The collection of the mosquitoes was collected in three districts and four wards from each district.
Figure 1.
Study sites of the mosquito collection. The collection of the mosquitoes was collected in three districts and four wards from each district.
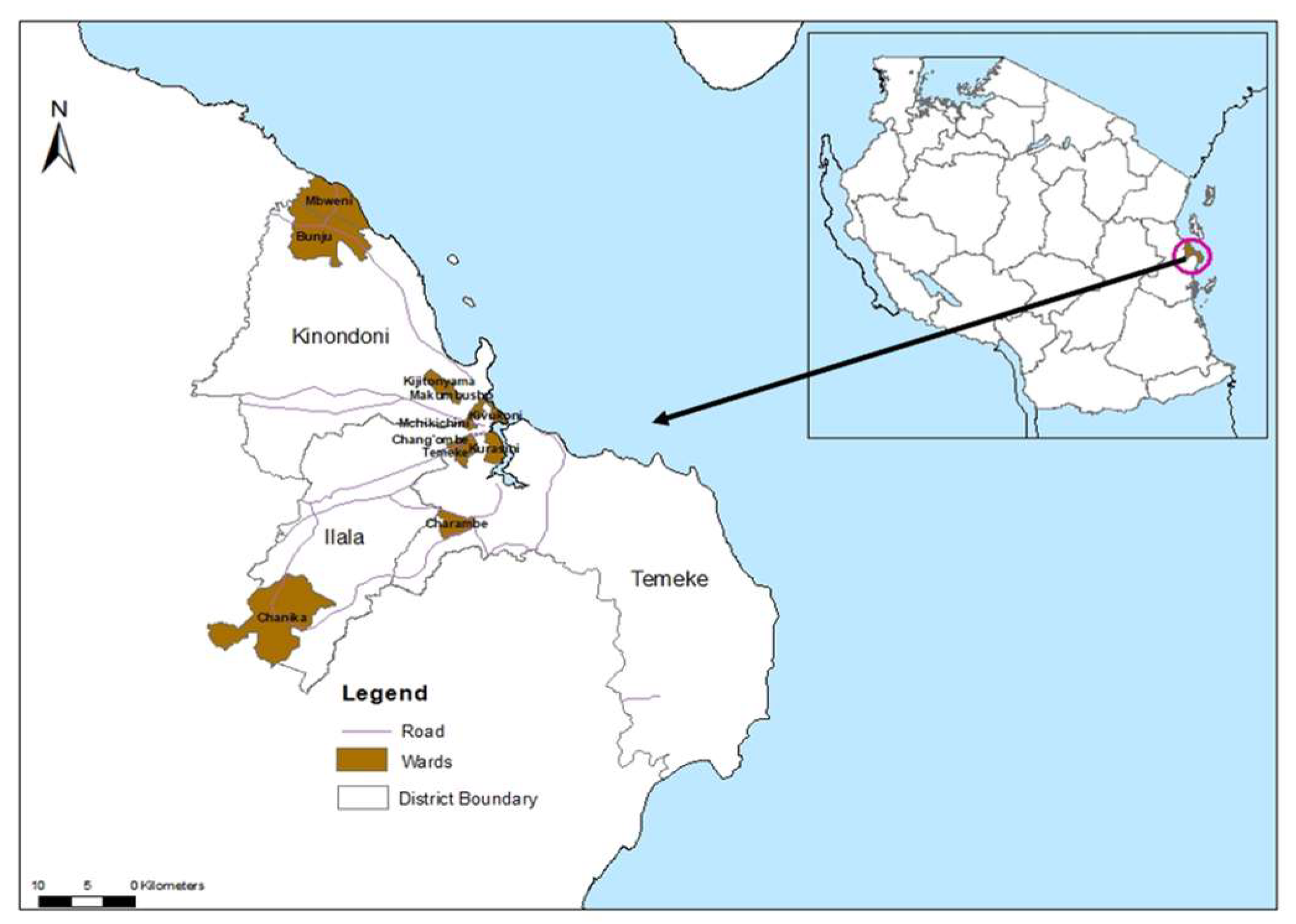
Figure 2.
Host-feeding preference of Aedes aegypti. *Mixed blood meal refers to blood meals from human and other hosts.
Figure 2.
Host-feeding preference of Aedes aegypti. *Mixed blood meal refers to blood meals from human and other hosts.
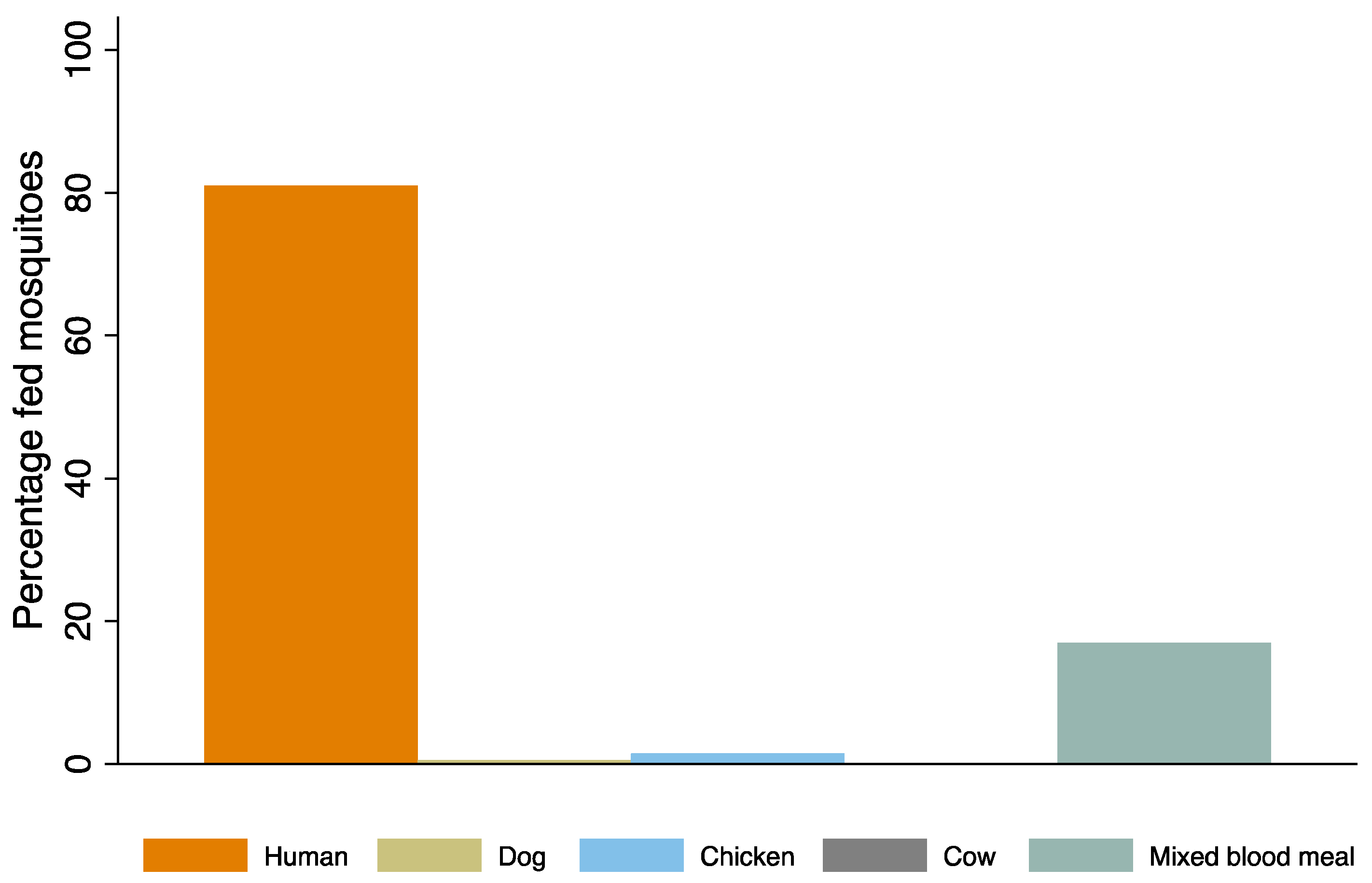

Table 1.
Primers and Probes used for DENV serotyping from Aedes aegypti mosquito samples.
| DENV serotype detected | Primer and probes | Nucleotide sequence (5’ → 3’) | Fluorophore and 3’ Quencher |
|---|---|---|---|
| DENV-1 | DEN-1 forward | CAAAAGGAAGTCGTGCAATA | FAM |
| DEN-1 reverse | CTGAGTGAATTCTCTCTACTGAACC | ||
| DEN-1 probe | CATGTGGTTGGGAGCACGC | ||
| DENV-2 | DEN-2 forward | CAGGCTATGGCACTGTCAC | HEX |
| DEN-2 reverse | CCATTTGCAGCAACACCATC | ||
| DEN-2 probe | CTCTCCGAGAACGGGCCTCGACTTCAA | ||
| DENV-3 | DEN-3 forward | GGACTGGACACACGCACTCA | CY5 |
| DEN-3 reverse | CATGTCTCTACCTTCTCGACTTGTCT | ||
| DEN-3 probe | ACCTGGATGTCGGCTGAAGGAGCTTG | ||
| DENV-4 | DEN-4 forward | TTGTCCTAATGATGCTGGTCG | CY5.5 CY5/BHQ3 |
| DEN-4 reverse | TCCACCTGAGACTCCTTCCA | ||
| DEN-4 probe | TTCCTACTCCTACGCATCGCATTCCG |
Table 2.
DENV serotype detected from pooled Aedes aegypti mosquitoes in Dar es Salaam.
| Districts | Mosquito samples | ||||
|---|---|---|---|---|---|
| Pools tested | Positive | DENV-serotype | Detection year | Infection rate per 1000 | |
| Temeke | 371 | 3 | DENV-2 | 2023 & 2024 | 0.81 (0.18, 2.39) |
| Ilala | 206 | 1 | DENV-2 | 2024 | 0.49 (0.012, 2.80) |
| Kinondoni | 277 | 1 | DENV-2 | 2023 | 0.39 (0.009, 2.20) |
| Total | 854 | 5 | DENV-2 | 2023 & 2024 | 0.41 (0.013, 1.10) |
Table 3.
Blood-fed Aedes aegypti mosquitoes collected by trap type.
| Traps | BGS trap | Prokopack aspirator | GAT |
|---|---|---|---|
| Blood fed Aedes aegypti | 57 | 142 | 6 |
| Percentage blood fed | 27.8 | 69.3 | 2.9 |
Table 4.
Percentage and incidence rate ratio (IRR) of host-seeking Aedes aegypti collected indoors and outdoors.
Table 4.
Percentage and incidence rate ratio (IRR) of host-seeking Aedes aegypti collected indoors and outdoors.
| N | n(%) | IRR (95%CI) | P-value | |
|---|---|---|---|---|
| Collection location | ||||
| Indoors | 54 | 76 (21.5) | 1 | - |
| Outdoors | 54 | 278 (78.5) | 4.33 (2.38-7.89) | <0.001 |
| Districts | ||||
| Ilala | 18 | 42 (11.9) | 1 | - |
| Kinondoni | 18 | 135 (38.1) | 4.24 (1.98-9.06) | <0.001 |
| Temeke | 18 | 177 (50.0) | 5.03 (2.39-10.58) | <0.001 |
Legend: N= collection days, n=number of mosquitoes, IRR=Incidance rate ratio, 95%CI=95% confidence interval.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



