
Submitted:
24 April 2025
Posted:
24 April 2025
You are already at the latest version
Abstract
Phytochemical screening of extracts of Tacca leontopetaloides tubers, has afforded the isolation of two novel chalcones, tarkalynins A and B, along with taccalonolide A and its 12-propanoate. Screening of Zanthoxylum zanthoxyloides stem bark yielded taraxerol acetate, dihydrochelerythrin and fagaramide. These compounds were obtained through column and thin-layer chromatography and identified using NMR and LC-HRMS. The compounds were tested against Trypanosoma brucei brucei s427 and its multi-drug-resistant clone B48, against T. evansi, T. equiperdum, T. congolense, and against L. mexicana. Cytotoxicity was tested against the human HEK293 cell line. The highest activities were observed with dihydrochelerythrin and fagaramide against T. b. brucei s427 and B48, T. evansi, and L. mexicana with EC50 values of 0.5, 0.9, 0.4, and 1.9 µg/mL and of 4.4, 2.7, 2.7, and 3.3 µg/mL, respectively. In addition, tarkalynin A and taraxerol acetate displayed promising activity against T. equiperdum (EC50 = 7.1 and 9.98 µg/mL, respectively). None of these compounds showed significant cross-resistance with existing trypanocides (RF ≈ 1; P>0.05). The compounds displayed low toxicity to human cells, with most exhibiting no growth inhibition at concentrations of 100 or 200 µg/mL. This report provides further evidence of the potential of natural products for combating parasitic diseases.
Keywords:
Tacca leontopetaloides
; Tarkalynin A
; Tarkalynin B
; Taccalonolides
; Antitrypanosomal
; Antileishmanial activity
; Zanthoxylum zanthoxyloides
; dihydrochelerythrin
; fagaramide
1. Introduction
African trypanosomes are the etiological agents of a wide range of diseases and are usually transmitted by insect vectors feeding on humans (Human African Trypanosomiasis or sleeping sickness) and animals (African Animal Trypanosomiasis or AAT) [1]. This mode of transmission is true for all pathogenic trypanosomes, except Trypanosoma equiperdum, which is transmitted sexually by copulation in horses and other equids, causing a wasting disease known as dourine [2]. Sleeping sickness, caused by T. gambiense and T. rhodesiense, is progressing towards control and elimination [3] but animal trypanosomiasis cases continue to grow in number and geographically [4,5,6]. AAT is commonly known as nagana in sub-Saharan Africa, where it is mostly transmitted by tsetse flies, and is classified as a neglected tropical veterinary disease. It is also referred to as surra in North Africa and Asia, mal de caderas and Derrengadera in South America and as dourine for the sexually-transmitted form in equines [2,6,7]. The main species responsible for nagana are T. brucei brucei, T. congolense, and T. vivax, whereas T. evansi causes surra and together with T. vivax contributes to mal de caderas in South America [6,7,8,9]. Mutations in Trypanosoma sp. have led to the emergence of different drug-resistant strains to the current drugs used at different treatment stages [10,11]. Moreover, most of the available drugs are only effective against the hemolymphatic stage of the disease, before the infection spreads to the central nervous system [12]. There is no effective chemotherapy for several of the animal trypanosomiasis infections, most notably dourine, but drug resistance threatens the treatability of surra and nagana as well [6,13]. This implies that AAT can no longer be effectively treated, and this situation is affecting livelihoods, economies, and food security [2,6].
Leishmaniasis, caused by protozoan parasites of up to 20 different Leishmania species is also a public health and veterinary concern in many countries in the tropics and subtropics, with about one million new human cases annually [14]. It is transmitted through the bite of an infected female phlebotomine sandfly [15]. Current chemotherapies depend on treatment with drugs of various efficacies, availability, and toxicities coupled with resistance as the greatest challenge [16].
Plants produce diverse active substances used in many fields of medicine with proven templates for new drug development [17,18]. Likewise, propolis, sourced by bees from local plants or trees, has proved to be a treasure trove of anti-parasite compounds [19,20,21]. Even simple derivatives of agricultural waste products can yield highly active new antiparasitics [22,23]. Tacca leontopetaloides, commonly referred to as Polynesian arrow or bat flower, Amura or Tarayaya (in Hausa), Gbache (in Tiv), or Andru (in Idoma) [24], is a perennial herbaceous plant commonly found in the North Central and Western parts of Nigeria [25]. It is a flowering plant with a highly bitter and starchy bulb-like tuber which is used as a staple food and as a source of starch in North Central Nigeria [25,26,27]. Several compounds of pharmaceutical importance have been isolated from the plant with ethnopharmacological activities, including antimicrobial and antioxidant [28,29], anti-inflammatory and anti-pyretic [30,31] and anti-cancer properties [32]. Most of these compounds are taccalonolides, a class of highly oxygenated pentacyclic triterpenes. In cancer research, they have been shown to stabilize microtubules, similar to the anticancer agents paclitaxel and epothilone, and are active against cell lines resistant to those drugs [33]. The diversity in structure, unique mechanism of action, and low toxicity of taccalonolides have attracted interest for drug discovery. Consequently, several taccalonolides [27,33,34] have been isolated, as well as taccabulins A-E, evelynin A [35] and evelynin B [36]. The initial anti-protozoan report on taccalonolides [34] showed these compounds to possess a range of activities from promising to moderate (0.76 ≤ EC50 ≤ 12.2 µg/mL) and demonstrated their potential as drugs against Trypanosoma and Leishmania species.
We also assess the anti-parasite properties of Zanthoxylum zanthoxyloides, popularly known as Fagara, Candlewood, zanthoxylum and to some natives of Nigeria as Akenaka or Ayer (in Tiv), Ufu otachacha (in Igede) and Faschuari (in Hausa) [24]. It is a shrub, spiny and rather scandent, up to 6-8 m tall, belonging to the family Rutaceae. It has been reported to contain α-pinene, citronellol, geraniol, limonene, β-myrcene [37], tannin, saponins, flavonoids, phenol, alkaloids, terpenoids, essential oils and coumarins, and to possess several medicinal activities including antinociceptive, antimalarial, cytotoxic, antiproliferative, anthelminthic, antiviral and antifungal, antioxidant, analgesic, anti-inflammatory, antimicrobial, wound healing, larvicidal, trypanocidal, uterine contraction, antitumor and hepatoprotective properties [38,39,40,41]. Here, we report further investigation of the plant extracts to isolate compounds possessing higher activity against parasites causing HAT, AAT or leishmaniasis, with possible different modes of action and no cross-resistance to existing treatments.
2. Results and Discussion
2.1. Isolation and Structural Characterization
Purification of compounds from the ethyl acetate extracts of Tacca leontopetaloides tubers afforded a novel chalcone, tarkalynin A (1), belonging to the taccabulin class of compounds. Three previously known compounds were also identified in the T. leontopetaloides extracts. First, a methylenedioxy dihydrochalcone (2) from the combined hexane and ethyl acetate extracts of the Tacca peels in fraction TPHE 26 was identified as 1-(benzo[d][1,3]-dioxol-5-yl)-β-(2′-hydroxy-4′,6′-dimethoxyphenyl)propan-1-one (tarkalynin B), and we here report its isolation as a natural compound for the first time; it has previously only been produced synthetically, as an intermediate to the synthesis of some taccabulins [42]. Second, taccalonolide A (3) was obtained from the combined hexane and ethyl acetate fractions of the Tacca tubers (TTHE 70-76) and peels (TPHE 62-74), and from the methanol fractions of the peels (TPM 49-69). Thirdly, taccalonolide A 12-propanoate (4) was isolated from the tuber hexane and ethyl acetate fraction TTHE 73 and peel methanol extract fractions TPM 20-37. Taraxerol acetate (5) was obtained from ethyl acetate fraction of Zanthoxylum zanthoxyloides (ZSE-06), dihydrochelerythrin (6) was isolated from ethyl acetate fraction of Z. zanthoxyloides (ZSE-42) and fagaramide (7) from Z. zanthoxyloides stem bark hexane fraction ZSH-44.
Compound 1 was obtained from the ethyl acetate fractions (TTHE 48) of Tacca tubers as a brown solid. Its LC-HRMS spectrum yielded a molecular ion [M+H]+ at m/z 333.1336 (calculated 333.1338, C18H21O6), corresponding to the molecular formula C18H20O6, with seven degrees of unsaturation. This was confirmed by its 1H-NMR spectrum (Table 1) which showed four aromatic proton signals at δH (ppm) 8.09 (d, J = 8.9 Hz, H-2 and 6), 7.04 (d, J = 9.0 Hz, H-3 and H-5), 6.23 (d, J = 2.3 Hz, H-3′), 6.05 (d, J = 2.4 Hz, H-5′) and a set of methylene signals at δH (ppm) 3.46 (dd, 14.9, 2.9, H-αa), 2.86 (dd, 14.9, 7.6, H-αb). Integration of the proton spectrum showed that the signals at 7.04 and 8.09 ppm are due to two protons each, indicating symmetry in the aromatic ring. The spectrum also showed that the methylene protons are coupled to an oxymethine proton at 5.18 (dd, J = 7.6, 2.9, H-β). Three methoxy singlet signals were observed at δH (ppm) 3.95, 3.79, and 3.61. The 13C-MNR spectrum showed a total of eighteen signals including two quaternary aromatic carbons at δC (ppm) 125.9 (C-1) and 105.1 (C-1′); four aromatic tertiary carbons at δC (ppm) 164.4 (C-4), 160.4 (C-4′), 158.3 (C-6′) and 157.7 (C-2′); six aromatic CH at 131.5 (C-2 and C-6), 114.0 (C-3 and C-5), 94.8 (C-3′) and 91.3 (C-5′) indicating the presence of a disubstituted and a tetrasubstituted aromatic ring. A characteristic signal at δC 197.9 ppm was indicative of an open-chain saturated ketone. The three signals at δC (ppm) 55.6, 55.3 and 55.4 confirmed the presence of three methoxy carbons substituted on the aromatic rings. Two aliphatic signals at δC (ppm) 75.6 and 30.5 were attributed to an oxygenated carbon and a methylene carbon respectively. The structure was fully deduced through its 2D NMR (COSY, HSQC, HMBC) spectra. Its COSY spectrum gave correlations between the aromatic protons at 8.09 (H-2 and H-6) and 7.04 (H-3 and H-5) indicating they were ortho coupled. There were also correlations between the oxymethine proton at 5.18 (H-β) and the methylene protons at 3.46 (H-αa) and 2.86 (H-αb). The HMBC spectrum showed long-range correlations between the aromatic protons at 7.04 (H-3 and H-5) to the quaternary carbon at 125.9 (C-1) while the protons at 8.09 (H-2 and H-6) had correlations to the quaternary carbon at 164.4 (C-4) and the carbonyl carbon at 197.9. Long-range correlations from the protons at 8.09 (H-2 and H-6) to the carbonyl carbon at 197.9 and from the methylene proton at 2.86 (H-αb) to two aromatic quaternary carbons at 105.1 (C-1′) and 157.7 (C-2′) confirmed the propanone chain was substituted by an aromatic ring at both ends. The three methoxy groups were identified by their HSQC correlations to their respective carbons: 3.95 (δC 55.6), 3.79 (δC 55.3) and 3.61 (δC 55.4). The methoxy-bearing carbons were identified by the HMBC correlations of the methoxy protons: 3.95 (δC 164.4), 3.79 (δC 160.4), and 3.61 (δC 158.3). Thus, compound 1 was characterized as 1-(4-hydroxy)-β-(2′-hydroxy-4′,6′-dimethoxyphenyl)propan-1-one with a common name tarkalynin A (Figure 1).
Compound 2 was isolated from the combined hexane and ethyl acetate extracts of the T. leontopetaloides peels in column fraction 26 (TPHE 26) as a brown solid with the molecular formula C18H18O6 from its LC-HRMS spectrum which gave a molecular ion [M+H]+ at m/z = 331.1168 (calculated 331.1182, C18H19O6) with eight degrees of unsaturation. Its 1H-NMR spectrum gave signals (Table 1) for five aromatic protons at δH (ppm) 7.63 (dd, J = 8.3,1.8 Hz, H-2), 7.47 (d, J = 1.8 Hz, H-6), 6.84 (d, J = 8.3 Hz, H-3), 6.20 (d, J = 1.8 Hz, H-3′) and 6.07 (d, J = 2.5 Hz, H-5′) indicating the compound has two aromatic rings and was similar to compound 1. The only difference was a methylenedioxy group attached to one of the aromatic rings and the presence of neighbouring methylene protons at 3.33 (H-α) and 2.95 (H-β) as indicated by the COSY correlations between them. From correlations in its 2D spectra, it was identified as 1-(benzo[d][1,3]-dioxol-5-yl)-β-(2′-hydroxy-4′,6′-dimethoxyphenyl)propan-1-one and named tarkalynin B. The spectral data agreed with the literature reports [42].
Compound 3 was isolated from the hexane and ethyl acetate extract fractions of the T. leontopetaloides tubers (TTHE 70-76) and peels (TPHE 62-74), and the methanol extract fractions of the peels (TPM 49-69) as a brown solid. It was identified as taccalonolide A by comparison of its spectral data with literature reports [34]. Its LC-HRMS spectrum gave a molecular ion [M+H]+ at m/z 703.2973 (calculated 703.2966, C36H47O14), corresponding to the molecular formula C36H46O14.
Compound 4 was identified as Taccalonolide A 12-propanoate by comparison of its 1H-NMR data with literature reports [34]. It was also obtained as a brown solid from the hexane and ethyl acetate fractions of the T. leontopetaloides Tacca tubers (TTHE 73) and the peel methanol fractions (TPM 20-37). Its LC-HRMS spectrum yielded a molecular ion [M+H]+ at m/z 717.3156 (calculated 717.3122, C37H49O14), corresponding to the molecular formula C37H48O14.
Compound 5 was obtained from the ethyl acetate extract fraction ZSE-06 of Z. zanthoxyloides stembark as a white solid and identified as taraxerol acetate by comparison of its 1H-NMR data with literature reports [43].
Compound 6 was also obtained from the ethyl acetate extract fraction ZSE-42 of Z. zanthoxyloides stembark as a white solid and identified as dihydrochelerythrin by comparison of its spectral data with literature reports [44].
Compound 7 was obtained from the hexane extract fraction ZSH-44 of Z. zanthoxyloides stembark as a white solid and identified as fagaramide by comparison of its spectral data with earlier reports [45].
2.2. Effect of the Isolated Compounds on Trypanosomes
The in vitro activities of six of the isolated compounds were carried out against bloodstream forms of T. b. brucei (s427 wild-type), multi-drug resistant-resistant T. b. brucei (B48), T. evansi (WT) and T. equiperdum (WT), T. congolense (WT) promastigotes using a resazurin-based assay. All values are displayed in Table 2.
Dihydrochelerythrin (6) exhibited a very high activity with EC50 values between 0.48 to 1.08 µg/mL against all the trypanosomes of the Trypanozoon subgenus (i.e. T. b. brucei, T. evansi and T. equiperdum), including the multidrug-resistant clone B48 (P > 0.05 relative to s427). Against T. congolense (subgenus Nannomonas) the activity was significantly (P < 0.001) lower, although still promising with an EC50 value of 2.9 ± 0.2 µg/mL. Since toxicity against HEK 293 was low, the in vitro selectivity index (SI) was good, especially for the Trypanozoon species (30.4 < SI < 87). Fagaramide (7) also displayed good activity against all trypanosome species (2.7 < EC50 < 8.5 µg/mL), and was less selective for the Trypanozoon subgenus, as the small difference between T. b. brucei s427 and T. congolense was not statistically significant (P >0.05). With toxicity against the human cell line above 90 µg/mL, the SI values ranged from 10.6 to 33.8. This confirms a previous report by [45], who also reported an interestingly high antitrypanosomal activity of fagaramide against T. b. brucei s427, with EC50 = 7.3 µM and no toxicity to normal cell lines (macrophages RAW264.7; EC50 = 214.96 µM). This compares to the activity displayed by our compound 7 (fagaramide) with an EC50 4.4 µg/mL (~17.8 µM) against s427. Z. zanthoxyloides has been shown to possess anticancer [46,47] and antibacterial [39] properties.
Of the T. leontopetaloides-derived compounds, the novel compound Tarkalynin A (1) displayed the most promising activity, but again with a highly significant separation between the Trypanozoon and Nannomonas subgenera (P <0.001), as EC50 values against the former ranged between 7.1 µg/mL (T. equiperdum) and 15.3 µg/mL for T. evansi, whereas the compound was almost inactive against T. congolense (EC50 = 81.7 µg/mL). No EC50 could be determined against HEK 293 because it was inactive even at the highest concentration tested, 100 µg/mL.
Taccalonolide A (3) and its 12-propanoate (4) displayed highly similar but moderate activities against the various trypanosome species. The small change from acetate to propanoate did not significantly impact its anti-trypanosomal properties. Although the compounds were not toxic to HEK 293 cells to the limited tested, their antiparasite activity was not sufficiently promising for them to be considered as lead compounds. Similarly, Taraxerol acetate (5) performed relatively poorly against most trypanosome species (EC50 > 25 µg/mL), with only the EC50 for T. equiperdum marginally below 10 µg/mL. Compounds 4 and 5 were not toxic to HEK 293 cells at the highest concentrations tested (100 and 200 µg/mL, respectively).
Cross-resistance with existing trypanocides of the diamidine class (e.g. pentamidine, diminazene, furamidine) and melaminophenyl arsenical class (melarsoprol, cymelarsan) was tested by side-by-side comparison of the standard drug-sensitive strain T. b. brucei s427 and clone B48. This clone was derived of s427 by sequentially deleting the TbAT1 drug transporter [48] and in vitro exposure to pentamidine [49]; it is highly resistant to all these important drugs against HAT and AAT [49,50]. None of the compounds here tested exhibited significant resistance in B48, nor as much as a 2-fold higher EC50, while resistance to pentamidine was approximately 50-fold (P < 0.001). The diamidine and arsenical resistance in T. brucei, T. evansi and T. equiperdum is associated with the functional loss of two drug transporters: the aminopurine transporter P2/TbAT1 and the aquaporin TbAQP2 [51,52,53,54], as has been demonstrated for B48 [55]. Thus, the trypanocidal action of the compounds used in this study is not dependent on these crucial, common drug transporters.
In general, the compounds displayed lower activity against T. congolense than against Trypanozoon group trypanosome species. This is also observed with several important trypanocides such as diminazene, pentamidine, other mitochondrion-targeting drugs, and the arsenicals [13,56]. This is a problem for the treatment of nagana since T. brucei and T. congolense incidence almost completely overlaps geographically, as they are transmitted by the same vectors, and therefore, the infecting species of a particular animal is rarely known [6]. However, dourine, in horses and other equids, is invariably caused by T. equiperdum and surra, from North Africa to South Asia and the Philippines, is caused by T. evansi only [2,57]. In this context, it is important that 6 and 7 displayed the highest activity against T. evansi and almost as promising activity against T. equiperdum, and that new treatments for dourine and surra are urgently required.
The promising antitrypanosomal ability of dihydrochelerythrin and fagaramide may be due to the presence of the methylene-dioxy moiety. A report by [47] had it that a derivative of dihydrochelerythrin: 6-Acetonyl-5, 6-dihydrochelerythrine, isolated from Zanthoxylum leprieurii showed no significant activity against s427 and the structure-activity relationships of 6 should therefore be carefully studied.
There are no anti-protozoal activity reports for compounds obtained from Tacca leontopetaloides except for our initial report on the anti-trypanosomal activity of compounds and fractions from the plant [34], where activity of 3.13 µg/mL was displayed by taccalonolide A 12 propanoate and 11.42 µg/mL by taccalonolide A against T. b. brucei s427. Although taccalonolide A showed a moderate EC50 (31.9 – 46.6 µg/mL), no cross-resistance (RF ≤ 1) was observed with the multi-drug resistant B48, T. evansi and T. equiperdum. Taccalonolide A 12 propanoate showed the least activity but exhibited moderate activity against s427 with an EC50 of 38.2 ± 6.2 µg/mL (53.4 µM) and poor activity against B48, T. evansi and T. equiperdum in the range 53.8 – 59.3 µg/mL. Generally, the tested compounds were poorly active against T. congolense (EC50 > 50 µg/mL).
2.3. Effect of the ISOLATED Compounds on Leishmania Mexicana
Dihydrochelerythrin (6) and fagaramide (7) showed significant activity, (EC50 = 1.9 ± 0.2 µg/mL (5.5 µM) and EC50 3.3 ± 0.1 µg/mL (13.3 µM) respectively against L. mexicana and thus showed genuine and broad anti-kinetoplastid activity. Tarkalynin A showed poor activity (EC50 = 193 µg/mL) against L. mexicana. Neither EC50 nor SI values for taccalonolide A and its propanoate against L. mexicana could be obtained due to low activity.
2.4. Effect of the Isolated Compounds on HEK
The compounds at a concentration of 100 µg/mL did not show toxicity to mammalian (HEK) cells as the observed EC50 values were above 90 µg/mL except for dihydrochelerythrin, which showed a moderate toxicity with EC50 = 33.0 µg/mL.
4. Materials and Methods
4.1. Plant Collection
The tubers of T. leontopetaloides and the stem bark of Z. zanthoxyloides were collected from Kusuv (Buruku LGA, Benue State) and Bunu Tai (Tai LGA, Rivers State) in Nigeria, respectively. The plants were identified by the Forestry and Wildlife Department, Joseph Sarwuan Tarka University, Makurdi, Benue State, and the Department of Forestry and Environmental Studies, Rivers State University, Port Harcourt, with voucher specimen numbers, UAM/FH/0469 and BSU/2017/ZZ-56. The tubers were rinsed with water, the bark scraped off, air-dried, ground, and sieved while the stem bark was air-dried and ground to powder.
4.2. Extraction and Isolation of Constituents
The dried and ground tubers (6.9 kg) and peel (4.0 kg) of T. leontopetaloides and 1 kg stem bark of Z. zanthoxyloides were each macerated and extracted with, successively, hexane, ethyl acetate, and methanol (2500 mL, 48 h). The filtrates were concentrated on a rotary evaporator at 40 °C and air-dried to obtain the hexane, ethyl acetate and methanol extracts. These extracts were subjected to TLC on pre-coated silica gel plates and visualized using anisaldehyde-H2SO4 reagent. The hexane and ethyl acetate extracts of T. leontopetaloides showed similar patterns on TLC and were therefore combined. Thus, four crude extracts were obtained from this plant: Tacca Tuber Hexane/Ethyl acetate (TTHE), Tacca Tuber Methanol (TTM), Tacca Peel Hexane Ethyl acetate (TPHE), and Tacca Peel Methanol (TPM). The maceration of Z. zanthoxyloides yielded three crude extracts: Zanthoxylum Stem Bark Hexane (ZSH), Zanthoxylum Stem Bark Ethyl acetate (ZSE), and Zanthoxylum Stem Bark Methanol (ZSM). The extracts were subjected to column chromatography over silica gel 60 (0.063-0.200 mm for CC, Merck, Germany). Each extract was pre-adsorbed on silica gel, loaded onto a wet-packed silica gel column, and eluted gradient-wise using hexane, ethyl acetate, and methanol. Ten fractions of ~20 mL were collected with 200 mL each of increasing ratios of ethyl acetate in hexane and then methanol in ethyl acetate to obtain a total of between 150 and 170 fractions for each extract. Fractions with similar constituents were combined based on their TLC profiles.
4.3. Spectroscopic Analysis
The spectral analysis was carried out at the Institute of Organic Chemistry, University of Glasgow, Scotland. One and two-dimensional NMR spectra of the compounds were obtained on a Bruker AVIII (400 MHz) spectrophotometer using deuterated chloroform (CDCl3) or acetone (CD3)2CO) as solvents. The spectra were processed using MestReNova (Mestrelab Research, Santiago de Compostela, Spain), and chemical shifts were referenced against residual solvent peaks. Mass spectra were acquired on a QTOF high-resolution Agilent 6545 mass spectrometer coupled to an Agilent Infinity 1290 UHPLC system.
4.4. Parasite Culture
In vitro cultures of T. b. brucei, T. evansi and T. equiperdum: Bloodstream trypomastigotes of T. b. brucei (s427 wild-type) [58], multi-drug-resistant T. b. brucei (B48) [49], T. evansi (AnTat 3/3) [59] and T. equiperdum (BoTat1) [51] were cultured in HMI-9 medium (Invitrogen, UK) supplemented with 3.0 g/L NaHCO3, 14.3 µL/L β-mercaptoethanol, adjusted to pH 7.4, 10% (v/v) heat-inactivated Foetal Bovine Serum (FBS; Gibco Life Technologies, Paisely, UK) and incubated at 37 °C in a 5% CO2 atmosphere as described. The cells were passaged every 72 h.
In vitro cultures of T. congolense: Bloodstream forms of Trypanosoma congolense (IL3000) were cultured as described [60] in Tc-BSF3 medium at 34 °C in a 5% CO2 atmosphere. The basal medium (1 L) was prepared using 9.60 g MEM (Sigma M0643), 5.96 g HEPES, 2.20 g NaHCO3, 1 g D-glucose, 110 mg sodium pyruvate (Sigma P3662), 10.68 mg adenosine, 14 mg hypoxanthine, 4 mg thymidine, 14.10 mg bathocuproinesulfonic acid (Sigma B1125). The basal medium (150 mL) was then supplemented with 40 mL heat-inactivated fresh goat serum (GIBCO, UK), 10 mL heat-inactivated serum plus (Sigma-Aldrich, UK), 2.8 µL β-mercaptoethanol, 1.6 mL glutamine and 2 mL penicillin/streptomycin solution to obtain 200 mL of Tc-BSF-3 medium [61]. The cells were passaged every 72 h.
In vitro Cultures of L. mexicana: Leishmania mexicana promastigotes of strain MNY/BZ/62/M379 [62] were cultured in HOMEM medium supplemented with 10% heat-inactivated FBS and 1% penicillin/streptomycin solution (Gibco Life Technologies) at 27 °C as described [63]. The cells were passaged every 72 h.
In vitro Cultures of HEK 293 cells: The Human Embryonic Kidney cell line HEK 293 strain was grown in a standard medium containing 500 mL Dulbecco’s Modified Eagle’s Medium (DMEM) (Sigma), 50 mL of heat-inactivated FBS (GIBCO, UK), 5 mL penicillin/streptomycin solution [64]. All constituents were mixed under sterile conditions in the DMEM bottle and stored at 4 °C before use. The cells were incubated at 37 °C and 5% CO2 and passaged twice a week in a vented flask until 80-85% confluence.
4.5. Antitrypanosomal and Antileishmanial Assays
The in vitro drug sensitivity assay in bloodstream forms of trypanosomes and Leishmania and the toxicity to mammalian cells were carried out at the School of Infection and Immunity, College of Medical, Veterinary and Life Sciences, University of Glasgow, Scotland according to the Resazurin (Alamar blue) assay methods used in recent reports [52,56]. The assay is based on the reduction of the blue, non-fluorescent dye resazurin sodium salt (Sigma) by living, but not dead cells, to the red, fluorescent metabolite resorufin [65]. Stock solutions of isolated compounds were prepared at 10 mg/mL in dimethyl sulfoxide (DMSO (Merck)) from which stocks of 200 μg/mL (400 μg/mL for L. mexicana) were prepared in the appropriate respective medium for each strain according to the standard protocol [56,66]. Exactly 200 μL of each stock was added to the first well of each row on a 96-well microplate, setting up for doubling dilutions of seven different drugs per plate, each over one row of a 96-well plate. Similarly, 200 μL of current trypanocides (positive control: diminazene aceturate for T. congolense, pentamidine for all other species) prepared at appropriate concentrations for each cell was added to the first well and included in each of the triplicate experiments. Next, 100 μL medium was pipetted using a multichannel micropipette into all remaining wells, and 100 μL of the drug was taken by a multichannel pipette from the first column and mixed gently with the medium in the wells of the second column, then another 100 μL from second column wells are added to third column wells and so on to the last-but-one column, creating a doubling dilution of 11 points. The last column of the plate was the drug-free negative control. Cell counts were performed using a haemocytometer and cell density was adjusted to the desired concentration of cells/mL (2 × 105 for T. b. brucei s427, B48, T. equiperdum; 2 × 106 for L. mexicana, 4 × 105 for T. evansi, 5 × 105 for T. congolense) of which 100 μL was added to all the wells in the plate, making a final top concentration of 100 μg/mL drug (200 μg/mL for L. mexicana). This was followed by incubation of the plates at 37 oC/5% CO2 (s427, B48, T. evansi and T. equiperdum), 34 °C/5% CO2 (T. congolense) or 27 °C (L. mexicana) for 48 h (72 h for L. mexicana) before the addition of the resazurin dye (20 µL of 125 mg/L), and a further incubation under the same conditions for 24 h (48 h for L. mexicana). The extended incubation period for L. mexicana is due to the slower metabolism of the dye by Leishmania promastigotes [52]. The fluorescence of the wells was read using a FLUOstar OPTIMA Fluorimeter (BMG Labtech, Durham, NC, USA) at wavelengths 544 nm for excitation and 590 nm for emission and a gain of 1250. The fluorescence values were analysed using GraphPad Prism, GraphPad Software Inc., San Diego, CA, USA, plotting the data to an equation for a sigmoidal dose-response curve with variable slope and the EC50 (half maximal effective concentration of isolated compounds or control drug that induces a response halfway between the baseline and maximum after 72 h exposure time) values were determined. P-values were calculated using the unpaired Student’s t-test. All experiments were performed on at least three different, fully independent occasions.
4.6. Toxicity Assays
The toxicity assay of the isolated compounds to mammalian cells was carried out on human embryonic kidney (HEK 293) cells, essentially as previously described [67]. Exactly 100 μL (3 × 105 cell/mL) of a cell suspension grown in a vented flask (~80% confluence) at incubation conditions of 37 °C/5% CO2 was added to each well of a 96-well plate. The plate was incubated for 24 h cytoadherence, after which 100 µL of a serially diluted drug was added (prepared in a separate sterile plate over one row). Phenylarsine oxide (PAO (Sigma)) was used as the positive control. The cells were then incubated for a further 30 h before the addition of 10 µL sterile Alamar Blue solution (125.0 mg/mL resazurin sodium salt (Sigma) in distilled water), followed by a further 24 h incubation. Fluorescence measurements and data analysis were performed as for the anti-parasite assays. The selectivity index (SI) was calculated as EC50 (HEK)/EC50 (parasite). All experiments were performed on at least three different, fully independent occasions.
5. Conclusions
Tarkalynin A and B, taccalonolide A and its propanoate were isolated from Tacca leontopetaloides, while taraxerol acetate, dihydrochelerythrin and fagaramide were isolated from Zanthoxylum zanthoxyloides. Dihydrochelerythrin (6) showed the highest anti-kinetoplastid activity across the board (all EC50 <3 µg/mL), and the highest selectivity index values. Furthermore, fagaramide (7) also displayed broad anti-kinetoplastid activity, albeit somewhat less potently (all EC50 < 8.5 µg/mL). Tarkalynin A (1) and taraxerol acetate (5) also displayed activity with EC50 below 10 µg/mL against T. equiperdum. No loss of activity was observed towards the multi-drug-resistant T. b. brucei clone B48. The study shows that compounds from Z. zanthoxyloides, in particular, have genuine anti-kinetoplastid properties, and these should now be studied in more detail.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: title; Table S1: title; Video S1: title.
Author Contributions
Conceptualization: JOI, EOA, HDK; methodology, JVA, JOI, CTA, EOA, CA, HEA, JIA, AS, CA. Investigation: JOI, AIG, HDK. Data analysis: JOI, EOA, HDK, JVA, CTA, AIG. Writing initial draft: EOA, JOI, writing final draft: JOI, HDK. Supervision: HDK, AIG, TAT, AOI; Project administration: JOI, AIG, HDK. All authors read and approved the final manuscript.
Funding
The work was funded through a PhD studentship grant from the Tertiary Education Trust Fund (TETFund) in Nigeria.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
NMR and MS spectra accompanying this paper are provided in the supplemental materials.
Acknowledgments
E.O.A is grateful to the staff at the Specialized Equipment Centre, Joseph Sarwuan Tarka University, Makurdi-Nigeria where preliminary phytochemical studies were carried out, the School of Infection and Immunity, College of Medical, Veterinary and Life Sciences, and the Institute of Organic Chemistry, University of Glasgow, for hosting EOA during a research visit Glasgow, UK. Special thanks also go to TETFund, Nigeria for sponsoring EOA to the University of Glasgow.
Conflicts of Interest
The authors declare no conflict of interest associated with this publication.
References
- Büscher, P.; Cecchi, G.; Jamonneau, V.; Priotto, G. Human African trypanosomiasis. Lancet. 2017, 390, 2397–2409. [Google Scholar] [PubMed]
- Giordani, F.; Morrison, L. J.; Rowan, T. G.; De Koning, H. P.; Barrett, M. P. The animal trypanosomiasis and their chemotherapy: a review. Parasitology 2016, 143, 1862–1889. [Google Scholar] [PubMed]
- Akazue, P.I.; Ebiloma, G.U.; Ajibola, O.; Isaac, C.; Onyekwelu, K.; Ezeh, C.O.; Eze, A.A. Sustainable Elimination (Zero Cases) of Sleeping Sickness: How Far Are We from Achieving This Goal? Pathogens 2019, 8, 135. [Google Scholar] [CrossRef] [PubMed]
- Anene, B.M.; Onah, D.N.; Nawa, Y. Drug resistance in pathogenic African trypanosomes: what hopes for the future? Vet Parasitol. 2001, 96, 83–100. [Google Scholar]
- Geerts, S.; Holmes, P.H.; Eisler, M.C.; Diall, O. African bovine trypanosomiasis: the problem of drug resistance. Trends Parasitol. 2001, 17, 25–8. [Google Scholar]
- Ungogo, M.A.; De Koning, H.P. Drug resistance in animal trypanosomiases: Epidemiology, mechanisms and control strategies. Int. J. for Parasitology: Drugs and Drug Resistance 2024, 25, 1–30. [Google Scholar]
- Zheoat, A.M.; Alenezi, S.; Elmahallawy, E.K.; Ungogo, M.A.; Alghamdi, A.H.; Watson, D.G.; Igoli, J.O.; Gray, A.I.; de Koning, H.P.; Ferro, V.A. Antitrypanosomal and Antileishmanial Activity of Chalcones and Flavanones from Polygonum salicifolium. Pathogens 2021, 10, 1–9. [Google Scholar] [CrossRef]
- Desquesnes, M.; Dargantes, A.; Lai, D.H.; Lun, Z.R.; Holzmuller, P. and Jittapalapong, S. Trypanosoma evansi and surra: a review and perspectives on transmission, epidemiology and control, impact, and zoonotic aspects. BioMed Research International 2013, 321237. [Google Scholar]
- Fetene, E.; Leta, S.; Regassa, F.; Büscher, P. Global distribution, host range and prevalence of Trypanosoma vivax: a systematic review and meta-analysis. Parasit Vectors 2021, 14, 80. [Google Scholar]
- Graf, F.E.; Ludin, P.; Wenzler, T.; Kaiser, M.; Brun, R.; Pati Pyana, P.; Büscher, P.; De Koning, H.P.; Horn, D.; Mäser, P. Aquaporin 2 mutations in Trypanosoma brucei gambiense field isolates correlate with decreased susceptibility to pentamidine and melarsoprol. PLoS Negl. Trop. Dis. 2013, 7, e2475. [Google Scholar]
- Munday, J.C.; Eze, A.A.; Baker, N.; Glover, L.; Clucas, C.; Aguinaga Andrés, D.; Natto, M.J.; Teka, I.A.; McDonald, J.; Lee, R.S.; Graf, F.E.; Ludin, P.; Burchmore, R.J.S.; Turner, C.M.R.; Tait, A.; MacLeod, A.; Mäser, P.; Barrett, M.P.; Horn, D.; De Koning, H.P. Trypanosoma brucei Aquaglyceroporin 2 is a high-affinity transporter for pentamidine and melaminophenyl arsenic drugs and is the main genetic determinant of resistance to these drugs. J. Antimicrob. Chemother. 2014, 69, 651–663. [Google Scholar] [PubMed]
- De Koning, H.P. The drugs of sleeping sickness: their mechanisms of action and resistance, and a brief history. Trop. Med. Infect. Dis. 2020, 5, 14. [Google Scholar] [CrossRef] [PubMed]
- Ungogo, M.A.; Campagnaro, G.D.; Alghamdi, A.H.; Natto, M.J.; De Koning, H.P. Differences in transporters rather than drug targets are the principal determinants of the different innate sensitivities of Trypanosoma congolense and Trypanozoon subgenus trypanosomes to diamidines and melaminophenyl arsenicals. Int. J. Mol. Sci. 2022, 23, 2844. [Google Scholar] [CrossRef] [PubMed]
- Burza, S.; Crioft, S.L. Boelaert, M. Leishmaniasis. The Lancet 2018, 392, 951–970. [Google Scholar] [CrossRef]
- Maroli, M.; Feliciangeli, M.D.; Bichaud, L.; Charrel, R.N.; Gradoni, L. Phlebotomine sandflies and the spreading of leishmaniases and other diseases of public health concern. Med. Vet. Entomol. 2013, 27, 123–147. [Google Scholar]
- García-Hernández, R.; Manzano, J.I.; Castanys, S.; Gamarro, F. Leishmania donovani develops resistance to drug combinations. PLoS Negl. Trop. Dis. 2012, 6, e1974. [Google Scholar]
- Nwodo, N.J.; Ibezim, A.; Ntie-Kang, F.; Adikwu, M.U.; and Chika, J. Anti-Trypanosomal Activity of Nigerian Plants and Their Constituents. Molecules 2015, 20, 7750–7771. [Google Scholar] [CrossRef]
- Ungogo, M.A.; Ebiloma, G.U.; Ichoron, N.; Igoli, J.O.; De Koning, H.P.; Balogun, E.O. A review of the antimalarial, antitrypanosomal and antileishmanial activities of natural compounds isolated from Nigerian flora. Front. Chem. 2020, 8, 617448. [Google Scholar]
- Ebiloma, G.U.; Ichoron, N.; Watson, D.G.; Igoli, J.O.; De Koning, H.P. The strong anti-kinetoplastid properties of bee propolis: Composition, identification of the active agents and their biochemical targets. Molecules 2020, 25, 5155. [Google Scholar] [CrossRef]
- Alotaibi, A.; Ebiloma, G.U.; Williams, R.; Alenezi, S.; Donachie, A.M.; Guillaume, S.; Igoli, J.O.; Fearnley, J.; De Koning, H.P.; Watson, D.G. European propolis is highly active against trypanosomatids including Crithidia fasciculata. Sci. Rep. 2019, 9, 11364. [Google Scholar]
- Siheri, W.; Ebiloma, G.U.; Igoli, J.O.; Gray, A.I.; Biddau, M.; Akrachalanont, P.; Alenezi, S.; Edrada-Ebel, R.; Muller, S.; Lawrence, C.; Fearnley, J.; Watson, D.G.; De Koning, H.P. Isolation of a novel flavanonol and an alkylresorcinol with highly potent anti-trypanosomal activity from Libyan propolis. Molecules 2019, 24, 1041. [Google Scholar] [CrossRef] [PubMed]
- Cerone, M.; Uliassi, E.; Prati, F.; Ebiloma, G.U.; Lemgruber, L.; Bergamini, C.; Watson, D.G.; de, A.M.; Ferreira, T.; Roth Cardoso, G.S.H.; Soares Romeiro, L.A.; de Koning, H.P.; Bolognesi, M.L. Discovery of Sustainable Drugs for Neglected Tropical Diseases: Cashew Nut Shell Liquid (CNSL)-Based Hybrids Target Mitochondrial Function and ATP Production in Trypanosoma brucei. ChemMedChem. 2019, 14, 621–635. [Google Scholar] [PubMed]
- Rossi, M.; Martinengo, B.; Diamanti, E.; Salerno, A.; Rizzardi, N.; Fato, R.; Bergamini, C.; Souza de Oliveira, A.; de Araújo Marques Ferreira, T.; Andrade Holanda, C.; Roeiro, L.; Soeiro, M.N.; Nunes, K.; Ferreira de Almeida Fiuza, L.; Meuser Batista, M.; Fraga, C.; Alkhalaf, H.E.A.; Elmahallawy, E.K.; Ebiloma, G.U.; De Koning, H.; Vittorio, S.; Vistoli, G.; Blanquart, C.; Bertrand, P.; Bolognesi, M.L. Benign-by-design SAHA analogues. ACS Med Chem Lett. 2024, 15, 1506–1515. [Google Scholar] [PubMed]
- Agishi, E.C. Tiv, Idoma, Etulo, Igede, Akweya, Hausa, English and Scientific Names of Plants, 2nd ed.; Agitab Publishers ltd: Makurdi, Nigeria, 2010; p. 139. [Google Scholar]
- Ogbonna, A.I.; Adepoju, S.O.; Ogbonna, C.I.C.; Yakubu, T.; Itelima, J.U.; Dajin, V.Y. Root tuber of Tacca leontopetaloides L. (Kunze) for food and nutritional security. Microbiology: Current Research 2017, 1, 5–11. [Google Scholar]
- Ogunwusi, A.A. and Ibrahim, H.D. Prospects for Industrial Utilization of Tacca (Tacca involucrata) in Nigeria. Journal of Natural Sciences Research 2023, 14, 10–20. [Google Scholar]
- Jiang, J-H.; Yang, H-M.; Wang, Y-L. and Chen, Y.G. Phytochemical and Pharmacological Studies of the Genus Tacca: A Review. Tropical Journal of Pharmaceutical Research 2014, 13, 635–648.
- Habila, J.D.; Bello, I.A.; Dzikwe, A.A.; Ladan, Z. and Sabiu, I.M. Comparative Evaluation of Phytochemicals, Antioxidant and Antimicrobial Activity of Four Medicinal Plants Native to Northern Nigeria. Australian Journal of Basic and Applied Sciences 2011, 5, 537–543. [Google Scholar]
- Vu, Q.T.H.; Le, P.T.K.; Vo, H.P.H.; Nguyen, T.T.; Nguyen, T.K.M. Characteristics of Tacca leontopetaloides L. Kuntze Collected from a Giang in Vietnam. International Conference on Chemical Engineering, Food and Biotechnology. AIP Conf. Proc. 2017, 1878, 020022-1–020022-6. [Google Scholar] [CrossRef]
- Ahmed, S.; Rakib, A.; Islam, Md. A.; Khanam, B.H.; Faiz, F.B.; Arkajyoti, P. ; Md. Nazim Uddin Chy, Md. N.U.; Bhuiya, N.M.M.A.; Uddin, M.M.N.; Ullah, S.M.A.; Rahman, Md. A. and Emran, T.B. In vivo and in vitro pharmacological activities of Tacca integrifolia rhizome and investigation of possible lead compounds against breast cancer through in silico approaches. Clinical Phytoscience 2019, 5, 1–13. [Google Scholar]
- Yen, P.H.; Chi, V.T.Q.; Kiem, P.V.; Tai, B.H.; Quang, T.H.; Nhiem, N.X.; Anh, H.L.T.; Ban, N.K.; Thanh, B.V.; Minh, C.V.; Park, S.J.; Kim, S.H. Spirostanol saponins from Tacca veitnamensis and their inflammatory activity. Bioorganic and Medicinal Chemistry Letters 2016, 26, 3780–3784. [Google Scholar]
- Li, L.; Ni, W.; Li, X.R.; Hua, Y.; Fang, P.L.; Kong, L.M.; Pan, L.L.; Li, Y.; Chen, C.X.; Liu, H.Y. Taccasubosides A-D, four new steroidal glycosides from Tacca subflabellata. Steroids 2011, 76, 1037–1042. [Google Scholar] [PubMed]
- Li. Y.; Du, Y-F.; Gao, F.; Xu, J-B.; Zheng, L-L.; Liu, G. and Lei, Y. Taccalonolides: Structure, semi-synthesis, and biological activity. Frontiers in Pharmacology 2022, 13, 1–15. [Google Scholar] [CrossRef]
- Dike, V.T.; Burbwa, V.; Bosha, J.A.; Yin, T.M.; Ebiloma, G.U.; de Koning, H.P.; Igoli, J.O. and Gray, A.I. Antitrypanosomal Activity of a Novel Taccalonolide from the Tubers of Tacca leontopetaloides. Phytochemical Analysis 2016, 27, 217–221. [Google Scholar] [PubMed]
- Peng, J.; Jackson, E. M.; Babinski, D. J.; Risinger, A. L.; Helms, G.; Frantz, D. E.; Mooberry, S. L. Evelynin, a Cytotoxic Benzoquinone-type retro-Dihydrochalcone from Tacca chantrieri. J. Nat. Prod. 2010, 73, 1590–1592. [Google Scholar]
- Peng, J.; Risinger, A.L.; Da, C.; Fest, G.A.; Kellogg, G.E. and Mooberry, S.L. Structure−Activity Relationships of Retro-dihydrochalcones Isolated from Tacca sp. Journal of Natural Products 2013, 76, 2189–2194. [Google Scholar]
- Nna., P.J.; Tor-Anyiin, T.A. Nna. P.J.; Tor-Anyiin, T.A. and Igoli, J.O. Fagaramide and Pellitorine from the Stem Bark of Zanthoxylum zanthoxyloides and Their Antimicrobial Activities. South Asian Research Journal of Natural Products 2019, 2, 1–8. [Google Scholar]
- Balekar, N.; Nakpheng, T. and Chiang, T.S. Wedelia trilobata L.: A Phytochemical and Pharmacological Review. Mai J. Sci. 2014, 41, 590–605, http://epg.science.cmu.ac.th/ejournal/Contributed Paper. [Google Scholar]
- Ikome, H.N.; Tamfu, A.N.; Abdou, J.P.; Fouotsa, H.; Nangmo, P.K.; Lah, F.C.W.; Tchinda, A.T.; Ceylan, O.; Frederich, M.; Nkengfack, A.E. Disruption of Biofilm Formation and Quorum Sensing in Pathogenic Bacteria by Compounds from Zanthoxylum Gilletti (De Wild) P.G. Waterman. Applied Biochemistry and Biotechnology 2023, 195, 6113–6131. [Google Scholar]
- Omosa, L. K.; and Okemza, E. K. Antiplasmodial activities of the stem bark extract and compounds of Zanthoxylum gilletii (De wild) PG Waterman. Pharmacognosy Communications 2017, 7, 41–46. [Google Scholar]
- Claudio, R. N.; and Lopes, L. M. X. Antiplasmodial natural products (review). Molecules 2011, 16, 2146–2190. [Google Scholar]
- Rao, M.L.N. and Ramakrishna, B.S. Rh-Catalyzed Decarbonylative Addition of Salicylaldehydes with Vinyl Ketones: Synthesis of Taccabulins A–E. European Journal of Organic Chemistry 2019, 7545–7554. [Google Scholar]
- Abouelela, M.E.; Orabi, M.A.A.; Abdelhamid, R.A.; Abdelkader, M.S.A.; Darwish, F.M.M. Chemical and Cytotoxic Investigation of Non-Polar Extract from Ceiba Pentandra (L.) Gaertn.: A Study Supported by Computer Based Screening. Journal of Applied Pharmaceutical Science 2018, 8, 057–064. [Google Scholar]
- Joshi, B.S.; Moore, K.M.; Pelletier, S.W.; Puar, M.S. Alkaloids of Zanthoxylum budrunga Wall.: NMR Assignments of Dihydrochelerythrine, (±)-Evodiamine and Zanthobungeanine. Phytochemical analysis 1991, 2, 20–25. [Google Scholar]
- Dofuor, A.K.; Kwain, S.; Osei, E.; Tetevi, G.M.; Okine, L.K.; Ohashi, M.; Gwira, T.M.; and Kyeremeh, K. N-(Isobutyl)-3,4-methylenedioxy Cinnamoyl Amide. Molbank 2019, M1070, 1–11. [Google Scholar]
- Ali, I.; Li, J.; Cui, L.; Zhao, H.; He, Q.; Wang, D. Efficient extraction and purification of benzo[c]phenanthridine alkaloids from Macleaya cordata (Willd) R. Br. by combination of ultrahigh pressure extraction and pH-zone-refining counter-current chromatography with anti-breast cancer activity in vitro. Phyto. Anal 2021, 32, 423–432. [Google Scholar]
- Eze, F.I.; Siwe-Noundou, X.; Isaac, M.; Patnala, S.; Osadebe, P.O.; Krause, R.W.M. Anti-cancer and anti-trypanosomal properties of alkaloids from the root bark of Zanthoxylum leprieurii Guill and Perr. Tropical Journal of Pharmaceutical Research 2020, 19, 2377–2383. [Google Scholar]
- Matovu, E.; Stewart, M.; Geiser, F.; Brun, R.; Maser, P.; Wallace, L.J.M.; Burchmore, R.J.; Enyaru, J.C.K.; Barrett, M.P.; Kaminsky, R.; Seebeck, T.; and de Koning, H.P. Mechanisms of arsenical and diamidine uptake and resistance in Trypanosoma brucei. Eukaryotic Cell 2003, 2, 1003–1008. [Google Scholar] [CrossRef]
- Bridges, D.; Gould, M.K.; Nerima, B.; Mäser, P.; Burchmore, R.J.S. and De Koning, H.P. Loss of the High Affinity Pentamidine Transporter is responsible for high levels of cross-resistance between arsenical and diamidine drugs in African trypanosomes. Mol Pharmacol 2007, 71, 1098–1108. [Google Scholar]
- Ward, C.P.; Wong, P.E.; Burchmore, R.J.; De Koning, H.P.; and Barrett, M.P. Trypanocidal furamidine analogues: influence of pyridine nitrogens on trypanocidal activity, transport kinetics and resistance patterns. Antimicrob Agents Chemother. 2011, 55, 2352–2361. [Google Scholar]
- Stewart, M.L.; Burchmore, R.J.S.; Clucas, C.; Hertz-Fowler, C.; Brook, K.; Tait, A.; McLeod, A.; Turner, C.M.R.; De Koning, H.P.; Wong, P. E. and Barrett, M.P. Multiple genetic mechanisms lead to the loss of functional TbAT1 expression in drug resistant trypanosomes. Eukaryot Cell 2010, 9, 336–343. [Google Scholar]
- Alghamdi, A.H.S.; 2020. Drug sensitivity and drug resistance in Trypanosoma brucei and Leishmania: the aquaporins. PhD thesis. http://theses.gla.ac.uk/82107/.
- Munday, J.C.; Settimo, L. and De Koning, H.P. Transport proteins determine drug sensitivity and resistance in a protozoan parasite, Trypanosoma brucei. Frontiers Pharmacol. 2015, 6, 32. [Google Scholar]
- Suswam, E.A.; Ross, C.A.; Martin, R.J. Changes in adenosine transport associated with melaminophenyl arsenical (Mel CY) resistance in Trypanosoma evansi: down-regulation and affinity changes of the P2 transporter. Parasitology 2003, 127, 543–9. [Google Scholar] [PubMed]
- Munday, J.C.; Eze, A.A.; Baker, N.; Glover, L.; Clucas, C.; Aguinaga Andrés, D.; Natto, M.J.; Teka, I.A.; McDonald, J.; Lee, R.S.; Graf, F.E.; Ludin, P.; Burchmore, R.J.S.; Turner, C.M.R.; Tait, A.; MacLeod, A.; Mäser, P.; Barrett, M.P.; Horn, D.; De Koning, H.P. Trypanosoma brucei Aquaglyceroporin 2 is a high-affinity transporter for pentamidine and melaminophenyl arsenic drugs and is the main genetic determinant of resistance to these drugs. J. Antimicrob. Chemother. 2014, 69, 651–663. [Google Scholar] [PubMed]
- Ebiloma, G.U.; Ayuga, T.D.; Balogun, E.O.; Gil, L.A.; Donachie, A.; Kaiser, M.; Herraiz, T.; Inaoka, D.K.; Shiba, T.; Harada, S.; Kita, K.; De Koning, H.P.; Dardonville, C. Inhibition of trypanosome alternative oxidase without its N-terminal mitochondrial targeting signal (ΔMTS-TAO) by cationic and non-cationic 4-hydroxybenzoate and 4-alkoxybenzaldehyde derivatives active against T. brucei and T. congolense. Eur. J. Med. Chem. 2018, 150, 385–402. [Google Scholar]
- Elata, A.; Galon, E.M.; Moumouni, P.F.A.; Ybanez, R.H.D.; Mossaad, E.; Salces, C.B.; Bajenting, G.P.; Ybanez, A.P.; Xuan, X.; Inoue, N.; Suganuma, K. First molecular detection and identification of Trypanosoma evansi in goats from Cebu, Philippines using a PCR-based assay. Vet Parasitol Reg Stud Reports 2020, 21, 100414. [Google Scholar]
- De Koning, H.P.; MacLeod, A.; Barrett, M.P.; Cover, B.; Jarvis, S.M. . Further evidence for a link between melarsoprol resistance and P2 transporter function in African trypanosomes. Mol Biochem Parasitol. 2000, 106, 181–185. [Google Scholar]
- Dean, S.; Gould, M.K.; Dewar, C.E.; Schnaufer, A.C. Single point mutations in ATP synthase compensate for mitochondrial genome loss in trypanosomes. Proc. Natl. Acad. Sci. USA. 2013, 110, 14741–14746. [Google Scholar]
- Giordani, F.; Khalaf, A.I.; Gillingwater, K.; Munday, J.C.; De Koning, H.P.; Suckling, C.J.; Barrett, M.P.; Scott, F.J. Novel minor groove binders cure animal African trypanosomiasis in an in vivo mouse model. J. Med. Chem. 2019, 62, 3021–3035. [Google Scholar]
- Coustou, V.; Guegan, F.; Plazolles, N.; Baltz, T. Complete in vitro life cycle of Trypanosoma congolense: Development of genetic tools. PLoS Negl. Trop. Dis. 2010, 4, e618. [Google Scholar]
- Al-Salabi, M.I.; De Koning, H.P. Purine nucleobase transport in amastigotes of Leishmania mexicana: involvement in allopurinol uptake. Antimicrob Agents Chemother. 2005, 49, 3682–3689. [Google Scholar]
- Al-Salabi, M.I.; Wallace, L.J.M.; and De Koning, H.P. A Leishmania major nucleobase transporter responsible for allopurinol uptake is a functional homologue of the Trypanosoma brucei H2 transporter. Mol Pharmacol. 2003, 63, 814–820. [Google Scholar] [PubMed]
- Ebiloma, G.U.; Igoli, J.O.; Katsoulis, E.; Donachie, A.M.; Eze, A.; Gray, A.I.; De Koning, H.P. Bioassay-guided isolation of active principles from Nigerian medicinal plants identifies new trypanocides with low toxicity and no cross-resistance to diamidines and arsenicals. J. Ethnopharmacol. 2017, 20, 256–264. [Google Scholar]
- Gould, M.K.; Vu, X.L.; Seebeck, T.; and De Koning, H.P. Propidium iodide-based methods for monitoring drug action in the kinetoplastidae: comparison with the Alamar Blue assay. Anal Biochem. 2008, 382, 87–93. [Google Scholar] [PubMed]
- Ene, A.C.; Atawodi, S.E.; Apeh, Y.E.O. In vitro and In vivo antitrypanosomal effects of petroleum ether, chloroform and methanol extracts of Artemisia maritime Linn. Brit. J. Pharm. Res. 2014, 4, 751–58. [Google Scholar]
- Ebiloma, G.U.; Katsoulis, E.; Igoli, J.O.; Gray, A.I.; De Koning, H.P. Multi-target mode of action of a Clerodane-type diterpenoid from Polyalthia longifolia targeting African trypanosomes. Sci. Rep. 2018, 8, 4613. [Google Scholar]
Figure 1.
Structures of isolated compounds: 1, Tarkalynin A; 2, Tarkalynin B; 3, Taccalonolide A; 4, Taccalonolide A 12-propanoate; 5, Taraxerol acetate; 6, Dihydrochelerythrin; 7, Fagaramide.
Figure 1.
Structures of isolated compounds: 1, Tarkalynin A; 2, Tarkalynin B; 3, Taccalonolide A; 4, Taccalonolide A 12-propanoate; 5, Taraxerol acetate; 6, Dihydrochelerythrin; 7, Fagaramide.
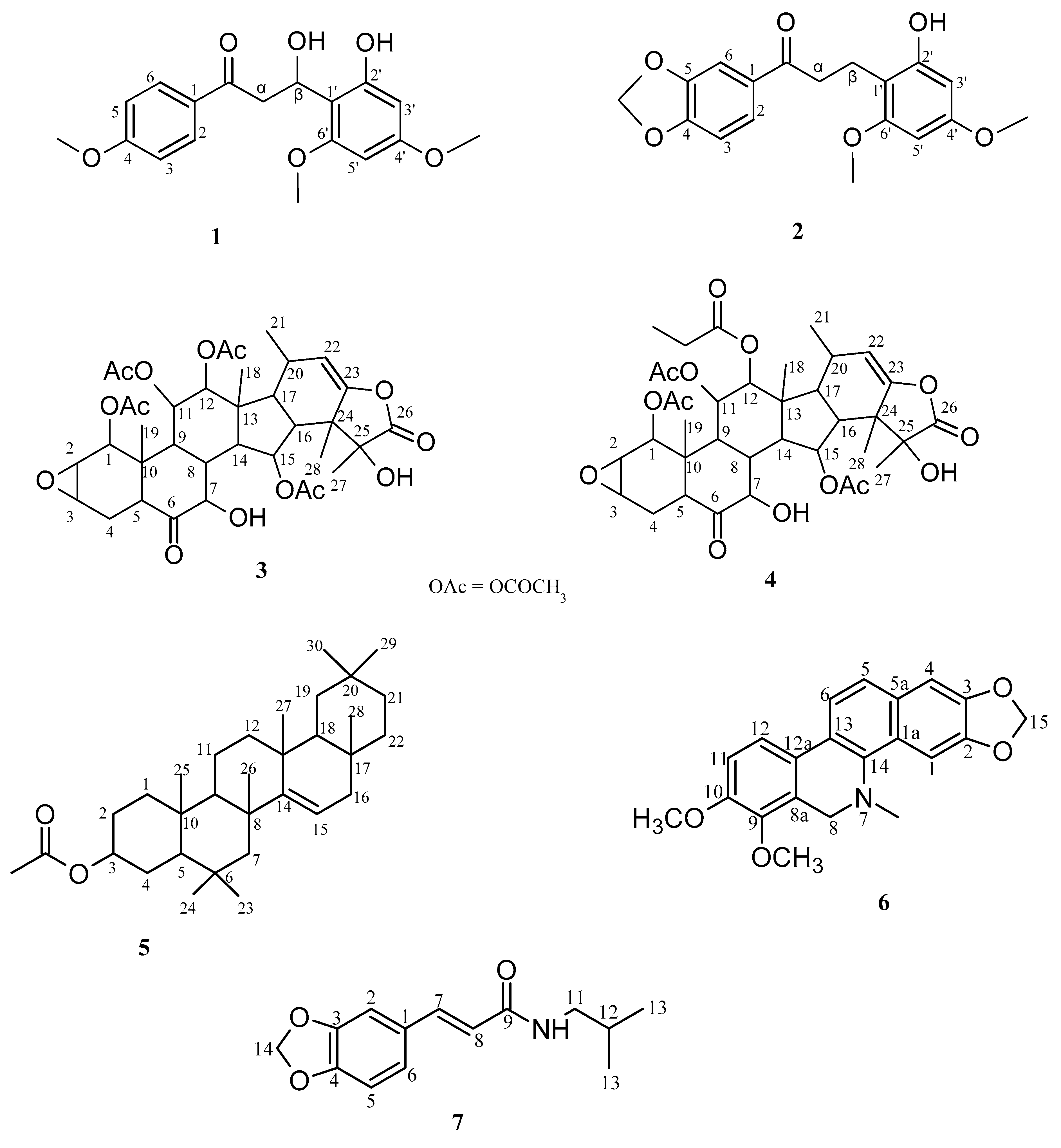

Table 1.
NMR data (400Hz, in CDCl3) (δ, ppm) for compounds 1 and 2.
| Position | Compound 1 | Compound 2 | ||
|---|---|---|---|---|
| 1H (ẟ ppm, m, J (Hz)) | 13C (m) | 1H (ẟ ppm, m, J (Hz)) | 13C (m) | |
| 1 | - | 125.9 (C) | - | 131.4 (C) |
| 2 | 8.09 (d, 8.9) | 131.5 (CH) | 7.63 (dd, 8.3, 1.8) | 125.0 (CH) |
| 3 | 7.04 (d, 9.0) | 114.0 (CH) | 6.84 (d, 8.3) | 107.9 (CH) |
| 4 | - | 164.4 (C) | - | 152.3 (C) |
| 5 | 7.04 (d, 9.0) | 114.0 (CH) | - | 147.7 (C) |
| 6 | 8.09 (d, 8.9) | 131.5 (CH) | 7.47 (d, 1.8) | 108.1 (CH) |
| 1′ | - | 105.1 (C) | - | 108.7 (C) |
| 2′ | - | 157.7 (C) | - | 156.3 (C) |
| 3′ | 6.23 (d, 2.3) | 94.8 (CH) | 6.20 (d, 2.4) | 94.7 (CH) |
| 4′ | - | 160.4 (C) | - | 159.9 (C) |
| 5′ | 6.05 (d, 2.4) | 91.3 (CH) | 6.07 (d, 2.5) | 91.3 (CH) |
| 6′ | - | 158.3 (C) | - | 159.3 (C) |
| α | 3.46 (dd, 14.9, 2.9)2.86 (dd, 14.9, 7.6) | 30.5 (CH2) | 3.33 (m) | 38.8 (CH2) |
| Β | 5.18 (dd, 7.6, 2.9) | 75.3 (CH) | 2.95 (dd, 6.6,4.4) | 16.6 (CH2) |
| -C=O | - | 197.9 (C) | - | 201.4 (C) |
| O-CH2-O | - | - | 6.02 (s) | 101.9 (CH2) |
| 4-OCH3 | 3.95 (s) | 55.6 (CH3) | - | - |
| 4′-OCH3 | 3.79 (s) | 55.3 (CH3) | 3.77 (s) | 55.3 (CH3) |
| 6′ -OCH3 | 3.61 (s) | 55.4 (CH3) | 3.81 (s) | 55.5 (CH3) |
| 2′-OH | 6.13 (s, br) | - | 8.80 (s, br) | - |
Table 2.
Effect of isolated compounds on trypanosomes and leishmania. EC50 values are average ± SEM of at least three independent determinations. RF = Resistance factor, being EC50 (parasite)/ EC50(s427WT). SI = Selectivity index, being EC50 (HEK)/ EC50(parasite).
Table 2.
Effect of isolated compounds on trypanosomes and leishmania. EC50 values are average ± SEM of at least three independent determinations. RF = Resistance factor, being EC50 (parasite)/ EC50(s427WT). SI = Selectivity index, being EC50 (HEK)/ EC50(parasite).
| Compound | T. b. brucei s427 | T. b. brucei B48 | T. evansi | T. equiperdum | T. congolense | L. mexicana | HEK 293 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EC50 (µg/mL) | SI | EC50 (µg/mL) | RF | SI | EC50 (µg/mL) | RF | SI | EC50 (µg/mL) | RF | SI | EC50 (µg/mL) | RF | SI | EC50 (µg/mL) | SI | EC50 (µg/mL) | |
| 1 | 11.8 ± 1.2 |
>8.5 | 16.7 ± 5.0 |
1.4 | >6.0 | 15.3 ± 0.8 | 1.3 | >6.5 | 7.1 ± 1.3 | 0.60 | >14 | 81.7 ± 1.7*** | 6.9 | >1.2 | 193 ± 1*** | 0.52 | >100 |
| 3 | 45.5 ± 14.9 |
>2.2 | 31.9 ± 2.9 |
0.7 | >3.1 | 46.6 ± 6.7 | 1.0 | >2.2 | 39.2 ± 0.0 | 0.86 | >2.6 | 73.9 ± 28.9 | 1.6 | >1.4 | >200 | ND | >100 |
| 4 | 38.2 ± 6.2 |
>2.6 | 53.8 ± 8.8 | 1.4 | >1.9 | 59.3 ± 5.3 | 1.6 | >1.7 | 55.7 ± 7.5 | 1.5 | >1.8 | >100 | >2.8 | ND | >200 | ND | >100 |
| 5 | 64.5 ± 28.5 | >3.1 | 59.8 ± 2.1 | 0.75 | >3.4 | 29.5 ± 0.7 | 0.4 | >6.8 | 9.98 ± 0.12 | 0.13 | >20 | 83.6 ± 6.7 | 1.1 | 2.4 | >200 | ND | >200 |
| 6 | 0.48 ± 0.15 | 68.0 | 0.89 ± 0.10 | 1.8 | 37.0 | 0.38 ± 0.02 | 0.8 | 87.0 | 1.08 ± 0.18 | 2.2 | 30.4 | 2.9 ± 0.2*** | 6.0 | 11.4 | 1.9 ± 0.2** | 18 | 33.0 ± 8.1 |
| 7 | 4.4 ± 1.8 | 20.4 | 2.7 ± 0.1 | 0.6 | 33.8 | 2.7 ± 0.1 | 0.6 | 33.0 | 6.70 ± 0.03 | 1.5 | 13.4 | 8.5 ± 1.4 | 1.9 | 10.6 | 3.3 ± 0.1 | 27 | 90.1 ± 10.8 |
| PMD (µM) |
a0.0066 ± 0.0001 b0.0052 ± 0.0006 |
- |
a0.312 ± 0.0349*** b0.288 ± 0.05*** |
a47.3 b55.4 |
a0.016 ± 0.004** b0.0025 ± 0.0003** |
a0.0081 ± 0.001 b0.0033 ± 0.001 |
ND |
a1.10 ± 0.03*** b0.76 ± 0.05*** |
ND | ||||||||
| DA (µM) |
ND | ND | ND | ND |
a0.51 ± 0.01 b0.46 ± 0.15 |
ND | ND | ||||||||||
| PAO (µM) | ND | ND | ND | ND | ND | ND |
a0.17 ± 0.01 b0.048 ± 0.011 |
||||||||||
Control: PMD = Pentamidine isethionate, DA = Diminazene aceturate, PAO = Phenyl arsine oxide. ND = not determined. **, P <0.01; ***, P <0.001. a = EC50 of control for 1, Tarkalynin A; 3, Taccalonolide A; 4, Taccalonolide A 12-propanoate; b = EC50 of control for 5, Taraxerol acetate; 6, Chelerythrine; 7, Fagaramide.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



