Submitted:
21 April 2025
Posted:
27 April 2025
Read the latest preprint version here
Abstract
Background: The epithelium of intestinal mucosa is colonized by specialized immune cells known as intraepithelial lymphocytes (IELs). Celiac disease is a gluten-sensitive enteropathy and common immune-related inflammatory disease of the small intestine. Objectives: To characterize the phenotype of IELs and the lamina propria of the mucosa in small intestine control using immuno-oncology and immune-phenotype markers, and to test the most relevant marker in celiac disease. Methods: Immunohistochemical analysis of CD3, CD4, CD8, CD103 (ITGAE), Granzyme B, TCRβ, TCRδ, CD56 (NCAM), CD16, LAIR1 (CD305), PD-L1, PD1 (CD279), BTLA (CD272), TOX2, HVEM (TNFRSF14), CD163, HLA-DP-DQ, IL4I1, and FOXP3 was performed using conventional histological analysis; and deep learning (convolutional neural network) for LAIR1. Results: IELs exhibited a cytotoxic T-cell phenotype and were positive for CD3, CD8, CD103, TCRβ, and LAIR1. The lamina propria was rich in CD163, HLA-DP-DQ, BTLA, and LAIR1-positive cells corresponding to macrophages, and T and B-lymphocytes. In celiac disease, IELs and the lamina propria were LAIR1-positive. Finally, as a proof-of-concept artificial intelligence (AI) analysis, LAIR1 image patches were successfully classified between small intestine control, celiac disease, and reactive tonsils. Conclusions: IELs are LAIR1-positive, and LAIR1 appears to be a potentially relevant marker in celiac disease.
Keywords:
small intestine
; celiac disease
; intraepithelial lymphocytes
; lamina propria
; immune-phenotype
; immuno-oncology
; LAIR1
; artificial intelligence
; deep learning
; convolutional neural network
1. Introduction
1.1. Histology of the Small Intestine
The normal intestinal mucosa has defined characteristic features, and its architecture include the villi, crypts, lamina propria, and muscularis mucosae.
The villi exhibit a digitiform shape with a 3:1 ratio between the height of the villi and the depth of the glandular crypts. The glandular crypts comprise several cell subtypes, including epithelial, Paneth, goblet, and endocrine cells. Each of these cells has different functions.
Intestinal epithelial cells (IECs) line the surface of the intestine and are responsible for the digestion of aliments, absorption of nutrients, and protection from infection by creating a physical barrier and modulating the immune response [1]. Intestinal epithelial cells are sensitive to the nutrients in the diet [1]. Paneth cells secrete alfa defensins, which are broad-spectrum microbicides that control the gut microbiota and intestinal homeostasis. In H&E staining, Paneth cells display bright red cytoplasmic granules [2]. Goblet cells produce mucus and are intimately involved in the control of the mucosal immune system [3]. Goblet cells sample luminal antigens to initiate adaptive immune response. There are several subtypes of goblet cells, with different localization and gene expression [3]. There are several types of endocrine cells in the small intestine [4]: EC cells produce serotonin (5-HT)[5,6,7]; L cells, GLP-1, GLP-2, and peptide YY [8,9,10]; K cells, GIP and 5-HT [11,12]; I cells, cholecystokinin and 5-HT [13,14]; D cells, somatostatin [15,16]; G cells, gastrin [17,18,19]; N cells, neurotensin [20,21]; M cells, motilin [22,23]; and S cells, secretin [24,25]. The main functions of endocrine cells are gut motility, appetite control, insulin release, cell proliferation control, gastric acid motility, pancreatic enzyme secretion, and intestinal absorption [4].
The lamina propria is a thin layer of connective tissue located below the epithelial basement membrane. The lamina propria is rich in fibroblasts, myofibroblasts, vascular and lymphatic vessels, elastic fibers, smooth muscle fascicles, and immune cells, including lymphocytes, plasma cells, macrophages, eosinophils, and mast cells [26].
The muscularis mucosae is composed of a very thin layer of smooth muscular cells with motor activity, which are linked to mucosal absorption and secretion functions [27].
The submucosa contains blood and lymphatic vessels, and nerves of the parasympathetic system, including the submucous plexus, also known as Meissner’s plexus [28]. The submucosal extracellular matrix is minimally immunogenic [28]. The muscularis propria [29] is composed of an inner circular and outer longitudinal layer, and within, Auerbach’s (myenteric) plexus is located.
1.2. Instraepithelial Lymphocytes
Intraepithelial lymphocytes (IELs) are found in the epithelium of the skin, genitourinary tract, respiratory tract, and intestinal tract [30]. IELs are a first lines of the immune system against pathogens that have attacked the epithelial surface. The typical phenotype is of cytotoxic T-lymphocytes, being CD3-positive and CD8-positive [31]. The T-cell receptor (TCR) is both alphabeta (αβ) and gammadelta (γδ)-positive. The fact that some IELs present with self-reactive TCR suggests an extrathymic origin [30,32,33,34,35,36].
IELs are specialized immune cells that colonize the intestinal mucosa. While B and innate cell populations may also transit inside this compartment, T-lymphocytes comprise the majority of intestinal IELs. In the intestine, IELs represent one of the largest lymphocyte populations and contribute to epithelial homeostasis and barrier integrity, including tolerance, resistance, and tissue protection [37]. There are several subsets of IELs. However, all strains share common characteristics, including restricted TCR diversity, epithelium-adapted profile, innate-like properties, and cytotoxic potential [37]. Human IELs can recognize modified self-antigens using both natural killer (NK) receptors and foreign antigens using the TCR [31]. The main characteristics of IELs are as follows:
- (1)
- (2)
- (3)
- (4)
- (5)
- (6)
- IELs are stratified into natural IELs (nIELs) and peripherally induced IELs (pIELs) [55,56,57,58]. The nIELs are generated in the thymus and migrate to the intestine. In contrast, pIELs are derived from CD4-positive or CD8-positive T cells at inductive sites, such as gut-associated lymph nodes, in response to dietary and microbial antigens [31,37,55,56,57,58,59,60,61].
- (7)
- IELs can be further subclassified according to their TCR subtype: (I) TCRγδ+ nIELs (tissue surveillance and repair), (II) TCRαβ+CD8αα+ nIELs (regulation), (III) TCRαβ+CD8αβ+ pIELs (effector memory, cytotoxicity), (IV) TCRαβ+CD4+ pIELs (regulation, cytotoxicity) [31,37]. Subtypes I and II may recognize self-antigens using their TCR, are present at birth, and are microbiota independent. On the other hand, subtypes III and IV may recognize microbial, viral, and dietary antigens using the TCRs, are absent at birth, increase with age, and are microbiota- and diet-dependent [31,37]. Of note, CD4+FOXP3+regulatory T lymphocytes (Tregs) can undergo CD4+CD8αα+ IEL differentiation in the intestinal epithelium [62,63].
- (8)
- CD8αα+ is an indication of intestinal IELs. Conventional CD8+ T cells express the CD8αβ heterodimer that is a TCR coreceptor, and enhance the TCR-MHC-I interactions during antigen presentation. Most IELs express CD8αα homodimer that decreases TCR sensitivity and prevents IEL hyperactivation via the mechanism of CD8αα homodimer interaction with thymus leukemia (TL) antigen [64], which is expressed by intestinal epithelial cells. Therefore, TL expression plays a critical role in maintaining IEL effector functions. TL deficiency is associated with colitis in a genetic model of inflammatory bowel disease [65].
- (9)
- IELs contribute to the pathogenesis of chronic intestinal inflammatory disease. Inflammatory bowel disease (IBD) includes Crohn disease and ulcerative colitis. Dysregulated intestinal immune response to microbiota is a cause of IBD [66,67]. In IBD, IELs could play a regulatory role [65,66,67,68,69,70,71,72]. A preserved villous architecture and increased IELs characterize microscopic colitis [73,74,75,76]. Celiac disease is an autoimmune disease triggered by dietary gliadin and is characterized by villous atrophy, crypt hyperplasia, and chronic inflammation of the lamina propria [77,78,79,80]. In celiac disease, there are increased CD8αβ+ pIELs and TCRγδ+ nIELs [31]. IELs can undergo neoplastic transformation into Enteropathy-associated T-cell lymphoma, a rare complication in patients with celiac disease who are unresponsive to gluten-free diet and treatment [81,82,83,84] (Figure 1).
Figure 1.
Histology of small intestine.
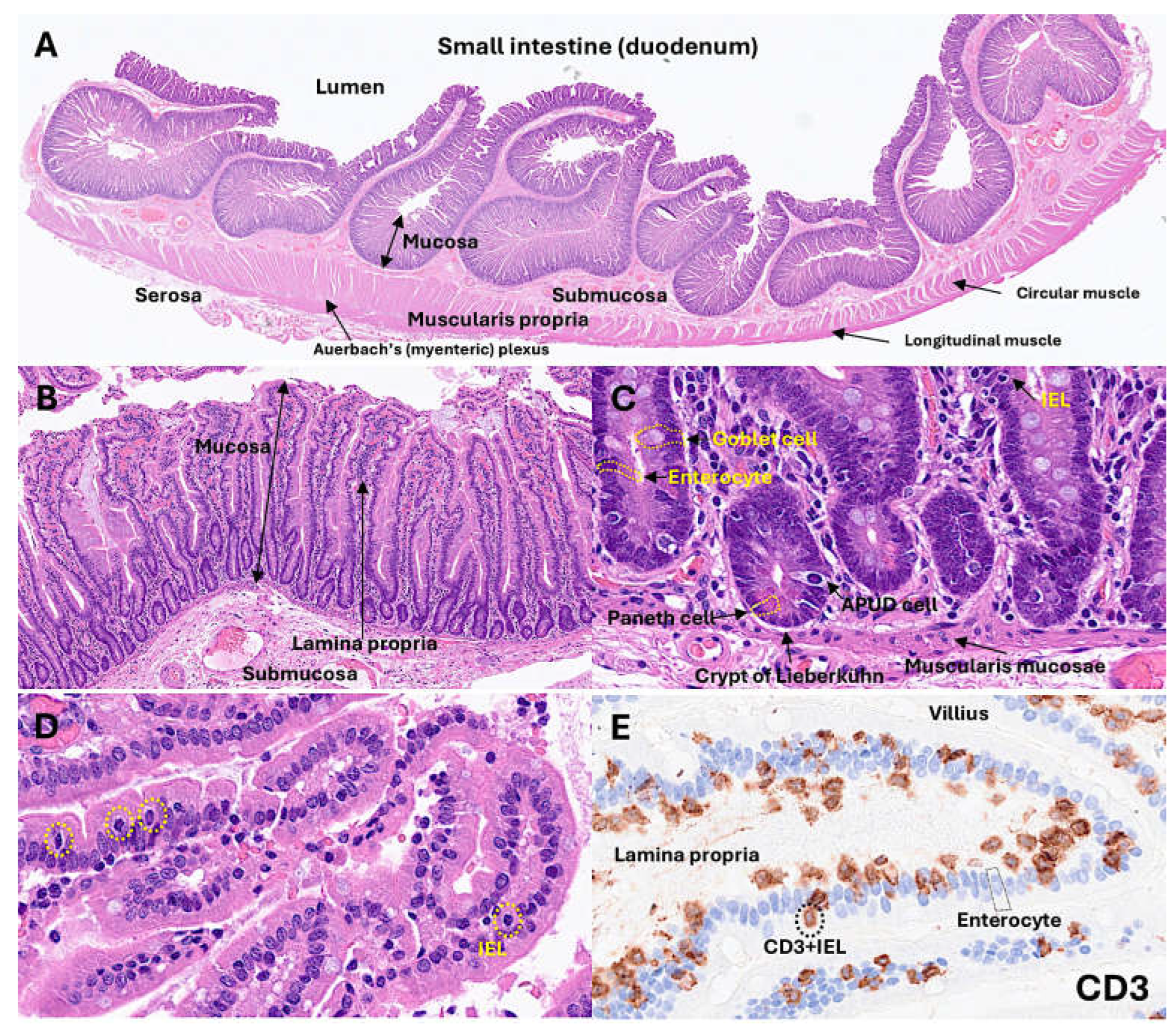
1.3. Celiac Disease
Celiac disease is a common immune-related disease, with a prevalence of approximately 1% in most populations [85]. The incidence of celiac disease has increased in recent years; the reason is unknown, but it may be related to environmental factors that are associated with the loss of tolerance to dietary gluten [85].
The pathogenesis of celiac disease is multifactorial. The pathogenesis includes a genomic background with the presence of several genetic factors [86], such as the close association with HLA-DR3-DQ2 and/or DR4-Dq8 gene locus, which is highly present in celiac disease patients [87]. Other gene loci have also been identified using genome-wide association studies (GWAS), which are related to metabolism and immune system, such as 3p21.31 (CCR3 and CCR2), 4p27 (KIAA1109, ADAD1, IL2, and IL21), 6q15 (BACH2), 6q25.3 (TAGAP), 1q24.3 (FASLG, TNFSF18, and TNFSF4), 6q22.31 (NKAIN2), 10p15.1 (PFKFB3 and PRKCQ), and 17q21.32 (HOXB9) [88]. Genome-wide gene expression studies have also highlighted similar biomarkers, including APOC3, CYP3A4, OCLN, MAD2L1, MKI67, CXCL11, and IL17A [89].
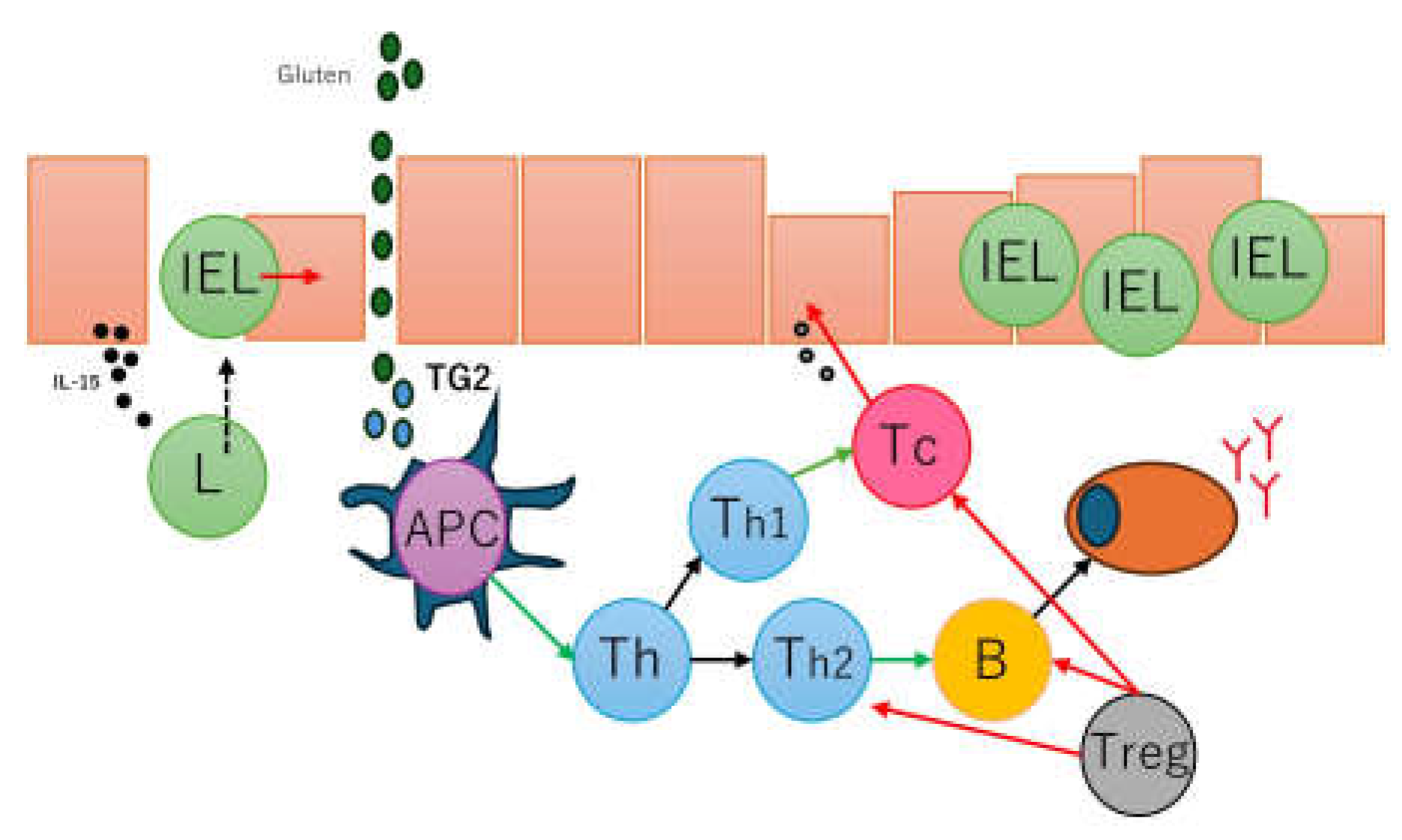
Celiac disease is characterized by an abnormal mucosal immune response to gliadin fractions, resulting in the infiltration of the lamina propria and epithelium by chronic inflammatory cells and villous atrophy [90]. Regarding adaptive immune response, key factors are peptide 56-89 (α-gliadin) that is resistant to gastrointestinal peptidases [91,92], tissue transglutaminase, and gliadin-reactive T cells. In active and gluten-sensitive celiac disease, the number of intraepithelial lymphocytes increases, express interferon gamma and IL-10 [93]. The gammadelta T-cell receptor (γδTCR) is also found to be increased in intraepithelial lymphocytes [94] in addition to the common alfa-beta T-cell receptor (αβTCR); in case of refractive celiac disease, intraepithelial lymphocytes may have an aberrant phenotype and restricted gene rearrangement [95,96]. In the serum of patients with celiac disease, several antibodies are found, including anti-gliadin antibodies (anti-AGA), anti-deamidated gliadin peptide antibodies (anti-DGP); anti-transglutaminase 2 antibodies (anti-TG2), anti-R1-type reticulin antibodies (anti-ARA), and anti-endomysial antibodies (anti-EMA) [97]. Gluten peptides also activate innate immune responses such as IL-15, intraepithelial lymphocytes, type 1 interferon (gamma), macrophages, monocytes, dendritic cells, and dysbiosis [98]. A summary of the pathogenesis of celiac disease has been presented in our previous publications [77,78] (Figure 2).
Figure 2.
Simplified pathogenesis of celiac disease. The pathogenesis of celiac disease is multifactorial and involves several factors, including dietary gluten (gliadin), genetics (HLA-DQ2/8 and other metabolic and immune-related non-HLA regions), environmental (dysbiosis, smoking), and immune factors. The immune factors include gluten-specific T-cell responses, autoantibody generation, cytokine generation, cytotoxic transformation of IELs, and innate immune activation with epithelial stress [77].
Figure 2.
Simplified pathogenesis of celiac disease. The pathogenesis of celiac disease is multifactorial and involves several factors, including dietary gluten (gliadin), genetics (HLA-DQ2/8 and other metabolic and immune-related non-HLA regions), environmental (dysbiosis, smoking), and immune factors. The immune factors include gluten-specific T-cell responses, autoantibody generation, cytokine generation, cytotoxic transformation of IELs, and innate immune activation with epithelial stress [77].
1.4. LAIR1
Leukocyte-associated immunoglobulin like receptor 1 (LAIR1), also known as CD305, is an immune-inhibitory receptor found on mature hematopoietic cells, particularly on immune cells such as mononuclear cells, natural killer cells, and T and B-lymphocytes [99]. The gene is located in the 19q13.4 region and is known as the leukocyte receptor cluster, which contains several genes that encode leukocyte receptors of the immunoglobulin superfamily. The structure of LAIR1 and the protein expression in reactive tonsils are shown in Figure 3. It is a type I glycoprotein comprising 287 ammino acids belonging to the family IR [100]. In myeloid leukemia, LAIR1 induces cell death, inhibits cytokine release and the activation of the NFKB pathway.
1.5. Aim of the Study
This study aimed to analyze the phenotype of intraepithelial lymphocytes (IELs) and the lamina propria in the small intestine, including LAIR1; and to confirm the LAIR1 expression in celiac disease.
2. Materials and Methods
2.1. Patients and Sample
This study used several biopsies of small intestine control (n = 18) and 16 cases of celiac disease (number of biopsies n = 57). The celiac cases were selected from the Department of Pathology of Hospital Clinic Barcelona, Spain, as described in our previous publications [77,78]. The cases had been diagnosed as celiac disease following the conventional diagnosis, with clinical criteria, positive celiac serology, and histological criteria, including the presence of increased intraepithelial lymphocytes with crypt hyperplasia (Marsh type 2) or villous atrophy (Marsh type 3). The detailed data are presented in Appendix A, Table A1.
The study was conducted according to the principles of the Declaration of Helsinki for human experimentation. This study was approved by the Ethical Committee of Tokai University (IRB14R-080 and IRB20-156).
2.2. Immunohistochemistry
Several immunohistochemical markers were analyzed in the tissue samples using a Leica Bond Max automated stainer according to the manufacturer’s instructions. The primary antibodies that were used were the following: CD3 (clone LN10, Leica Biosystems, Leica K.K., Tokyo, Japan), CD4 (4B12, Leica), CD8 (4B11, Leica), CD103 (EP206, Leica), granzyme B (11F1, Leica), TCRβ (TRBC1/TCRβ constant region 1 (E6Z3S) Rabbit mAb #79485, Cell Signaling Technology K.K., Tokyo, Japan), TCRδ (TRDC/TCRδ (E2E9T) XP® Rabbit mAb #55750, Cell Signaling), CD56 (CD56-504-L-CE, Leica), CD16 (CD16-L-CE, Leica), LAIR1 (CD305, JAVI82A, created by Giovanna Roncador, Spanish National Cancer Research Center (CNIO)), PD-L1 (73-10, Leica), PD1 (CD279, NAT105, CNIO), BTLA (CD272, FLO67B, CNIO), TOX2 (TOM924D, CNIO), HVEM (TNFRSF14, ab47677), CD163 (CD163-L-CE, Leica), HLA-DP-DQ (JS76, CNIO), IL4I1 (BALI265E,543H,573B, CNIO), and FOXP3 (236A, CNIO). The primary antibody details are presented in Table 1.
Confocal microscopy was performed as previously described [101] using a Fluoview FV3000 confocal laser scanning microscope (Olympus K.K, Hachioji, Japan) with Alexa Fluor 488 and 594 and DAPI dyes.
2.3. Image Classification
Image classification based on LAIR1 immunohistochemical expression was performed using transfer learning and the ResNet18 deep learning model. The images were split into images patches of 224x224x3. The image patches were pooled into 3 different folders, and the data were split into 3 sets: training set (70%) for training the network, validation set (10%) for testing its performance during training, and test set (20%) used after training to assess how well the network performed on new data. Grad-CAM analysis was used as a method of explainable AI to visualize which areas of the input image were most important for the model prediction and image classification. All the methodology was performed as previously described in our previous publications [78,102,103].
3. Results
3.1. Immunophenotype of Intraepithelial Lymphocytes (IELs) in Intestinal Mucosa Control
IELs were defined as lymphoid cells within the epithelial layer of the mucosa. CD3 staining was used as a reference. In physiological conditions, IELs are characterized by T-cell phenotype that is positive for CD3 and a cytotoxic phenotype with CD8 expression. Occasionally, cytotoxic granules were identified by granzyme B staining. Most IELs were positive for CD103/ITGAE. Most IELs expressed TCRβ chains; therefore, expressed the TCRαβ chains. Occasionally, CD56+IELs were found, as well as TCRδ chain-positive IELs (i.e., TCRγδ+IELs). All IELs were diffusely and strongly positive for LAIR1. Of note, occasional IELs were TOX2-positive. In the lamina propria, abundant CD163+macrophages/dendritic cells were found, which also expressed HLA-DP-DQ. CD4+cells were mainly found in the lamina propria; however, clusters attached below the epithelial basal membrane were found. BTLA+cells were found in the lamina propria, same result as we have previously described [77]. PD-L1 expression was limited to the lamina propria in a pattern compatible with APC (macrophages, dendritic cells). Regulatory T-lymphocytes were identified in the lamina propria using the FOXP3 marker. The characteristic images are shown in Figure 4-7. Figure 8 shows the confirmation that the IELs are CD3 and LAIR1 double-positive using confocal microscopy. Of note, LAIR1 staining revealed that many cells of the lamina propria were LAIR1-positive (Figure 4, Figure 5, Figure 6, Figure 7 and Figure 8).
3.2. Multicolor Analysis of LAIR1 and Other Immune Markers
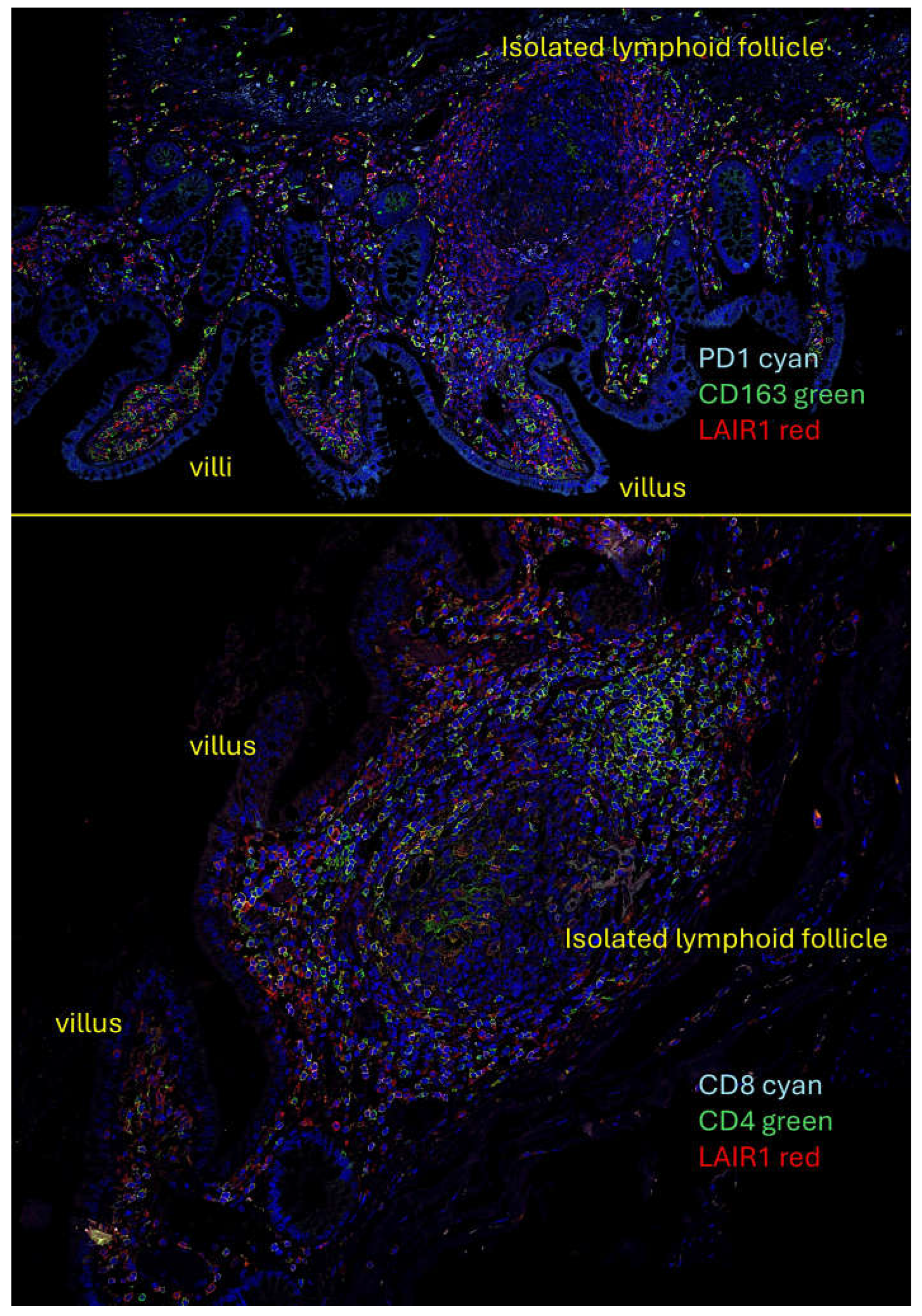
Quadruple and triple immunofluorescence analyses were performed using confocal microscopy. The combinations that also included nuclear staining were as follows: PD1 (Cyan), CD163 (green), and LAIR1 (red); and CD4 (green), CD8 (cyan), and LAIR1 (red).
The results showed that in the human mucosa, CD4- and probably CD8-positive cells were positive for LAIR1. PD1-positive cells also expressed LAIR1. In the interfollicular area and/or lamina propria, CD163-positive cells (M2-like macrophages) were partially positive for LAIR1 (Appendix A, Figure A1).
3.3. Analysis of LAIR1 Expression in Celiac Disease
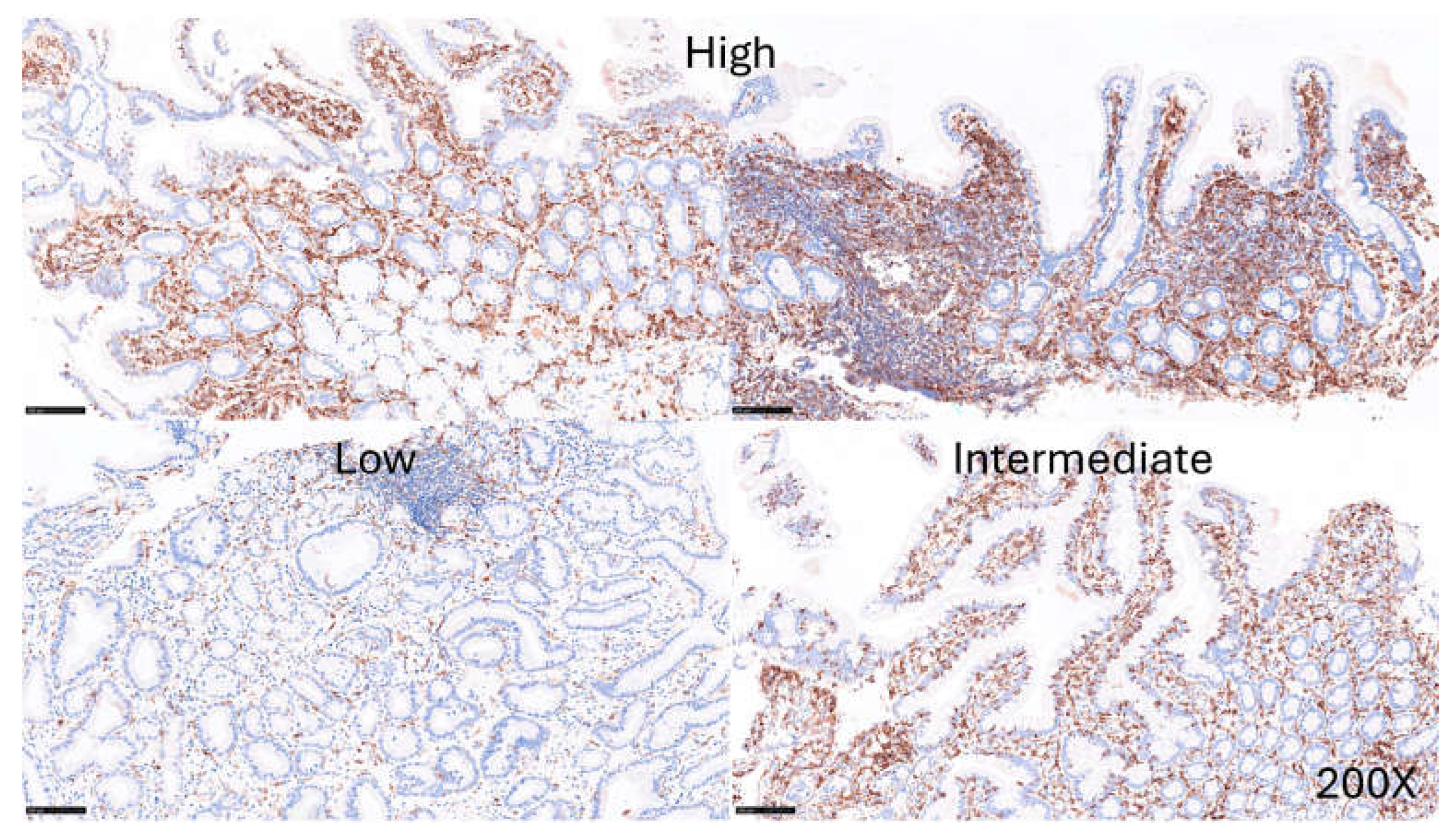
The expression of LAIR1 was evaluated in the celiac disease biopsies. LAIR1 was expressed in the IELs, and the distribution of LAIR1+IELs was heterogeneous within and between biopsies. The expression of LAIR1 in the lamina propria was also variable ranging from low (1/16, 6.25%), intermediate (6/16, 37.5%), and high (9/16, 56.25%) (Figure 9).
Figure 9.
LAIR1 expression in celiac disease. The expression of LAIR1 in the lamina propria was also variable ranging from low (1/16, 6.25%), intermediate (6/16, 37.5%), and high (9/16, 56.25%).
Figure 9.
LAIR1 expression in celiac disease. The expression of LAIR1 in the lamina propria was also variable ranging from low (1/16, 6.25%), intermediate (6/16, 37.5%), and high (9/16, 56.25%).
3.4. Image Classification of Celiac Disease, Small Intestine Control, and Reactive Tonsil Control Based on LAIR1 Immunohistochemical Expression
Images of LAIR1 protein expression analyzed by immunohistochemistry in celiac disease, small intestine control, and reactive tonsils were used as input data in a ResNet18 model.
The ResNet18 model comprises 18 layers, including convolutional layers and residual blocks. The series included 11,367 image patches of celiac disease, 11,630 patches of small intestine control, and 8147 patches of reactive tonsil control. The image patches were pooled into 3 different folders, and the data were split into 3 sets: training set (70%) for training the network, validation set (10%) for testing its performance during training, and test set (20%) used after training to assess how well the network performed on new data.
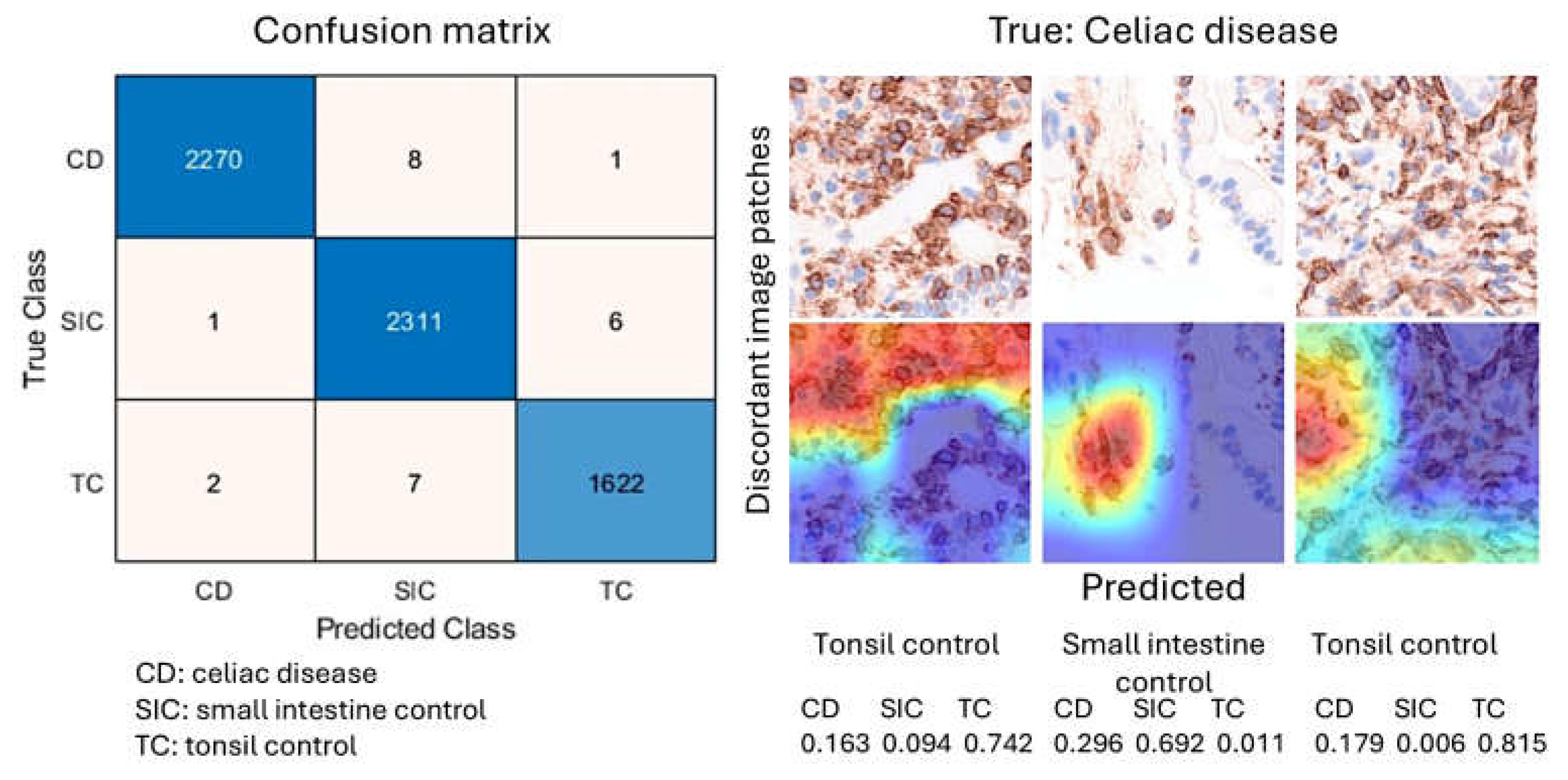
After 5 epochs in the training progress, the validation accuracy was 99.5%. After image patch classification using the test (holdout) series, the accuracy was 99.6%. The confusion matrix shows the distribution of image patches, including correctly classified and misclassified patches. The Grad-CAM technique was used to understand why the deep learning network made its classification decisions in incorrectly classified cases (Figure 10).
Figure 10.
Image classification using LAIR1 image patches. A ResNet18 model was created to classify histochemical images of celiac disease (CD), small intestine control (SIC), and reactive tonsil control (TC). The image classification results in the test set are summarized in a confusion matrix.
Figure 10.
Image classification using LAIR1 image patches. A ResNet18 model was created to classify histochemical images of celiac disease (CD), small intestine control (SIC), and reactive tonsil control (TC). The image classification results in the test set are summarized in a confusion matrix.
4. Discussion
Celiac disease is a gluten-sensitive enteropathy and common immune-mediated inflammatory condition of the small intestine caused by sensitivity to dietary gluten and related proteins in genetically predisposed individuals [104]. In western countries, celiac disease is estimated to affect approximately 1% of the population. Celiac disease clinically presents heterogeneous; therefore, it continues to be underestimated [104].
There are several phenotypes of celiac disease. Symptomatic disease includes classic and nonclassic celiac diseases. The classic celiac disease is a gluten-sensitive enteropathy characterized by diarrhea, malabsorption signs and symptoms, villous atrophy, and resolution upon withdrawal from gluten-containing foods [79]. Nonclassic celiac disease is also known as “atypical” and the patients lack the classic symptoms of malabsorption and only present with minor gastrointestinal complaints. However, duodenal biopsies show villous atrophy, the production of celiac autoantibodies such as antitissue transglutaminase, and extraintestinal manifestations are present [105,106].
The other phenotypes are subclinical or asymptomatic, potential, latent, and refractory celiac disease. Refractory celiac disease is defined the persistence of clinical symptoms and villous atrophy despite a gluten-free diet. Of note, failure to improve on a gluten-free diet is mostly due to non-compliance. However, in few cases, a pure refractory condition is found: refractory celiac disease type 1 (normal population of IELs), the semi-malignant inflammatory condition (refractory type 2; aberrant immunophenotype and T-cell receptor clonality analysis of IELs), transformation to enteropathy-associated T-cell lymphoma (EATL), collagenous sprue, or alternative diagnosis of autoimmune enteropathy [107,108,109].
The cause of refractory disease is unknown, and the treatment has focused on immunosuppression. Traditional glucocorticoids such as intravenous hydrocortisone and oral prednisolone. Alternative immunosuppressant therapy includes azathioprine, 6-mercaptopurine, and thioguanine [107,110,111,112,113]. A monoclonal antibody therapy using anti-CD52 (alemtuzumab) was reported [114].
Badran YR et al. reported eight cases of immune checkpoint inhibitor-associated celiac disease, suggesting that the drugs disrupted the mechanism of gut immune homeostasis and tolerance mechanism [115]. In that study, immunohistochemical analysis of several markers were CD3, CD8, TCRγδ, PD1, CD68, and PD-L1, and quantification of IELs [115]. In our study, we analyzed several immuno-oncology markers in small intestine control, and later LAIR1 expression in celiac disease. Our findings showed that in the small intestine, LAIR1 expression is found in IELs but also in immune cells of the lamina propria. In celiac disease, LAIR1 was diffusely expressed.
LAIR1 belongs to the family of immune-inhibitory receptors and is expressed by mature hematopoietic cells, particularly in the immune cells of natural killer (NK) and T/B-lymphocytes [114]. Beyond the physiological function of immune homeostasis and immune tolerance, LAIR1 has been involved in several autoimmune and inflammatory conditions and neoplasia [99]: allergy [116], systemic lupus erythematosus [117], rheumatoid arthritis [118,119,120], graft rejection [121], breast carcinoma [122], glioma [123], solid tumors [124], hepatocellular carcinoma [125], among others.
We recently demonstrated the usefulness of using deep learning to analyze gene expression and classify images of celiac disease [77,78] and ulcerative colitis [102,103,126]. In this study, deep learning was used to classify LAIR1 image patches between celiac disease, small intestine control, and reactive tonsils. The proposed network managed to classify images with good performance. However, the aim was to conduct a proof-of-concept analysis, not to create a trained network for use in production or commercial applications. Narrow artificial intelligence is not ready to take over the job of pathology-trained medical doctors because histological biopsies obtained from endoscopic examinations may have other diseases. Of note, other research groups such as Denholm et al., Molder et al., Scheppach et al., and Schreiber et al. (among others), have successfully used deep learning in celiac disease as well [127,128,129,130,131,132,133,134].
5. Conclusions
This study used several immuno-oncology and immune-phenotype markers to characterize the intraepithelial lymphocytes and the lamina propria of small intestine control, and to confirm the expression of LAIR1 in celiac disease. This study also successfully performed a proof-of-concept deep learning histological analysis of LAIR1 between small intestine control, celiac disease, and reactive tonsils. Therefore, LAIR1 appears to be a potentially relevant marker in celiac disease.
Supplementary Materials
N/A
Author Contributions
N/A
Funding
This research was funded to J.C. by the Ministry of Education, Culture, Sports, Science and Technology (MEXT) and Japan Society for the Promotion of Science (JSPS), grant numbers KAKEN 15K19061, 18K15100, and 23K06454.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Institutional Review Board of TOKAI UNIVERSITY, SCHOOL OF MEDICINE (protocol code IRB14R-080 and IRB20-156).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Data are uploaded into Zenodo CERN OpenAIRE repository.
Acknowledgments
I want to thank Josep A. Bombi from the Department of Pathology, Hospital Clinic of Barcelona (Spain) for the celiac disease cases.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| IELs | Intraepithelial lymphocytes |
| EATL | Enteropathy-associated T-cell lymphoma |
| LAIR1 | Leukocyte-associated immunoglobulin like receptor 1 |
Appendix A
Appendix A.1.

Table A1.
Clinicopatholgical characteristics of celiac disease cases.
| Age | Sex | Biopsy Location | Diagnosis | Marsh-Oberhuber Classification |
|---|---|---|---|---|
| 70 | Male | Duodenum | Celiac Disease | 3a |
| 62 | Male | Pylorus/duodenum | Celiac Disease/Chronic gastritis | 2 |
| 62 | Male | Duodenum | Celiac Disease | 2 |
| 78 | Female | Duodenum | Celiac Disease | 3b |
| 59 | Male | Duodenum | Celiac Disease | 3a |
| 44 | Female | Duodenum | Celiac Disease | 2 |
| 17 | Female | Duodenum | Celiac Disease | 3b |
| 56 | Female | Duodenum | Celiac Disease | 3a |
| 54 | Female | Duodenum | Celiac Disease | 2 |
| 58 | Female | Duodenum | Celiac Disease | 3b |
| 61 | Female | Duodenum | Celiac Disease | 3c |
| 45 | Male | Duodenum | Celiac Disease | 3a |
| 70 | Female | Duodenum | Celiac Disease | 2 |
| 40 | Female | Duodenum | Celiac Disease | 3a |
| 61 | Female | Duodenum | Celiac Disease | 3c |
| 44 | Female | Duodenum | Celiac Disease | 3a |
Figure A1.
Multicolor immunofluorescence. Multicolor immunofluorescence of LAIR1 in relationship with other immune markers in human intestinal control.
Figure A1.
Multicolor immunofluorescence. Multicolor immunofluorescence of LAIR1 in relationship with other immune markers in human intestinal control.
References
- Kong, S.; Zhang, Y.H.; Zhang, W. Regulation of Intestinal Epithelial Cells Properties and Functions by Amino Acids. Biomed Res Int 2018, 2018, 2819154. [Google Scholar] [CrossRef]
- Yang, E.; Shen, J. The roles and functions of Paneth cells in Crohn’s disease: A critical review. Cell Prolif 2021, 54, e12958. [Google Scholar] [CrossRef]
- Gustafsson, J.K.; Johansson, M.E.V. The role of goblet cells and mucus in intestinal homeostasis. Nat Rev Gastroenterol Hepatol 2022, 19, 785–803. [Google Scholar] [CrossRef]
- Atanga, R.; Singh, V.; In, J.G. Intestinal Enteroendocrine Cells: Present and Future Druggable Targets. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Kanova, M.; Kohout, P. Serotonin-Its Synthesis and Roles in the Healthy and the Critically Ill. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Pithadia, A.B.; Jain, S.M. 5-Hydroxytryptamine Receptor Subtypes and their Modulators with Therapeutic Potentials. J Clin Med Res 2009, 1, 72–80. [Google Scholar] [CrossRef]
- Shajib, M.S.; Khan, W.I. The role of serotonin and its receptors in activation of immune responses and inflammation. Acta Physiol (Oxf) 2015, 213, 561–574. [Google Scholar] [CrossRef]
- Drucker, D.J. Mechanisms of Action and Therapeutic Application of Glucagon-like Peptide-1. Cell Metab 2018, 27, 740–756. [Google Scholar] [CrossRef]
- Cinci, L.; Faussone-Pellegrini, M.S.; Rotondo, A.; Mule, F.; Vannucchi, M.G. GLP-2 receptor expression in excitatory and inhibitory enteric neurons and its role in mouse duodenum contractility. Neurogastroenterol Motil 2011, 23, e383–e392. [Google Scholar] [CrossRef]
- Guan, X.; Shi, X.; Li, X.; Chang, B.; Wang, Y.; Li, D.; Chan, L. GLP-2 receptor in POMC neurons suppresses feeding behavior and gastric motility. Am J Physiol Endocrinol Metab 2012, 303, E853–E864. [Google Scholar] [CrossRef]
- Overton, H.A.; Fyfe, M.C.; Reynet, C. GPR119, a novel G protein-coupled receptor target for the treatment of type 2 diabetes and obesity. Br J Pharmacol 2008, 153 Suppl 1, S76–81. [Google Scholar] [CrossRef]
- Martin, B.; Lopez de Maturana, R.; Brenneman, R.; Walent, T.; Mattson, M.P.; Maudsley, S. Class II G protein-coupled receptors and their ligands in neuronal function and protection. Neuromolecular Med 2005, 7, 3–36. [Google Scholar] [CrossRef]
- Szewczyk, J.R.; Laudeman, C. CCK1R agonists: a promising target for the pharmacological treatment of obesity. Curr Top Med Chem 2003, 3, 837–854. [Google Scholar] [CrossRef]
- Tanaka, T.; Katsuma, S.; Adachi, T.; Koshimizu, T.A.; Hirasawa, A.; Tsujimoto, G. Free fatty acids induce cholecystokinin secretion through GPR120. Naunyn Schmiedebergs Arch Pharmacol 2008, 377, 523–527. [Google Scholar] [CrossRef]
- Theodoropoulou, M.; Stalla, G.K. Somatostatin receptors: from signaling to clinical practice. Front Neuroendocrinol 2013, 34, 228–252. [Google Scholar] [CrossRef]
- Harda, K.; Szabo, Z.; Juhasz, E.; Dezso, B.; Kiss, C.; Schally, A.V.; Halmos, G. Expression of Somatostatin Receptor Subtypes (SSTR-1-SSTR-5) in Pediatric Hematological and Oncological Disorders. Molecules 2020, 25. [Google Scholar] [CrossRef]
- Schmassmann, A.; Reubi, J.C. Cholecystokinin-B/gastrin receptors enhance wound healing in the rat gastric mucosa. J Clin Invest 2000, 106, 1021–1029. [Google Scholar] [CrossRef]
- Larsson, L.I. Developmental biology of gastrin and somatostatin cells in the antropyloric mucosa of the stomach. Microsc Res Tech 2000, 48, 272–281. [Google Scholar] [CrossRef]
- Jiang, X.; Liu, Y.; Zhang, X.Y.; Liu, X.; Liu, X.; Wu, X.; Jose, P.A.; Duan, S.; Xu, F.J.; Yang, Z. Intestinal Gastrin/CCKBR (Cholecystokinin B Receptor) Ameliorates Salt-Sensitive Hypertension by Inhibiting Intestinal Na(+)/H(+) Exchanger 3 Activity Through a PKC (Protein Kinase C)-Mediated NHERF1 and NHERF2 Pathway. Hypertension 2022, 79, 1668–1679. [Google Scholar] [CrossRef]
- Xiao, Y.; Yan, W.; Lu, Y.; Zhou, K.; Cai, W. Neurotensin contributes to pediatric intestinal failure-associated liver disease via regulating intestinal bile acids uptake. EBioMedicine 2018, 35, 133–141. [Google Scholar] [CrossRef]
- Li, J.; Song, J.; Yan, B.; Weiss, H.L.; Weiss, L.T.; Gao, T.; Evers, B.M. Neurotensin differentially regulates bile acid metabolism and intestinal FXR-bile acid transporter axis in response to nutrient abundance. FASEB J 2021, 35, e21371. [Google Scholar] [CrossRef]
- Takeshita, E.; Matsuura, B.; Dong, M.; Miller, L.J.; Matsui, H.; Onji, M. Molecular characterization and distribution of motilin family receptors in the human gastrointestinal tract. J Gastroenterol 2006, 41, 223–230. [Google Scholar] [CrossRef]
- Miedzybrodzka, E.L.; Foreman, R.E.; Lu, V.B.; George, A.L.; Smith, C.A.; Larraufie, P.; Kay, R.G.; Goldspink, D.A.; Reimann, F.; Gribble, F.M. Stimulation of motilin secretion by bile, free fatty acids, and acidification in human duodenal organoids. Mol Metab 2021, 54, 101356. [Google Scholar] [CrossRef]
- Modvig, I.M.; Andersen, D.B.; Grunddal, K.V.; Kuhre, R.E.; Martinussen, C.; Christiansen, C.B.; Orskov, C.; Larraufie, P.; Kay, R.G.; Reimann, F.; et al. Secretin release after Roux-en-Y gastric bypass reveals a population of glucose-sensitive S cells in distal small intestine. Int J Obes (Lond) 2020, 44, 1859–1871. [Google Scholar] [CrossRef]
- Fukuhara, S.; Kobayashi, K.; Kusakizako, T.; Iida, W.; Kato, M.; Shihoya, W.; Nureki, O. Structure of the human secretin receptor coupled to an engineered heterotrimeric G protein. Biochem Biophys Res Commun 2020, 533, 861–866. [Google Scholar] [CrossRef]
- Roulis, M.; Flavell, R.A. Fibroblasts and myofibroblasts of the intestinal lamina propria in physiology and disease. Differentiation 2016, 92, 116–131. [Google Scholar] [CrossRef]
- Uchida, K.; Kamikawa, Y. Muscularis mucosae - the forgotten sibling. J Smooth Muscle Res 2007, 43, 157–177. [Google Scholar] [CrossRef]
- Zhao, Y.; Peng, H.; Sun, L.; Tong, J.; Cui, C.; Bai, Z.; Yan, J.; Qin, D.; Liu, Y.; Wang, J.; et al. The application of small intestinal submucosa in tissue regeneration. Mater Today Bio 2024, 26, 101032. [Google Scholar] [CrossRef]
- Lai, S.; Yu, W.; Wallace, L.; Sigalet, D. Intestinal muscularis propria increases in thickness with corrected gestational age and is focally attenuated in patients with isolated intestinal perforations. J Pediatr Surg 2014, 49, 114–119. [Google Scholar] [CrossRef]
- Beagley, K.W.; Husband, A.J. Intraepithelial lymphocytes: origins, distribution, and function. Crit Rev Immunol 1998, 18, 237–254. [Google Scholar] [CrossRef]
- Mayassi, T.; Jabri, B. Human intraepithelial lymphocytes. Mucosal Immunol 2018, 11, 1281–1289. [Google Scholar] [CrossRef]
- Lin, T.; Matsuzaki, G.; Kenai, H.; Nakamura, T.; Nomoto, K. Thymus influences the development of extrathymically derived intestinal intraepithelial lymphocytes. Eur J Immunol 1993, 23, 1968–1974. [Google Scholar] [CrossRef]
- Matsuzaki, G.; Lin, T.; Nomoto, K. Differentiation and function of intestinal intraepithelial lymphocytes. Int Rev Immunol 1994, 11, 47–60. [Google Scholar] [CrossRef]
- Lin, T.; Matsuzaki, G.; Kenai, H.; Kishihara, K.; Nabeshima, S.; Fung-Leung, W.P.; Mak, T.W.; Nomoto, K. Characteristics of fetal thymus-derived T cell receptor gamma delta intestinal intraepithelial lymphocytes. Eur J Immunol 1994, 24, 1792–1798. [Google Scholar] [CrossRef]
- Trejdosiewicz, L.K. Intestinal intraepithelial lymphocytes and lymphoepithelial interactions in the human gastrointestinal mucosa. Immunol Lett 1992, 32, 13–19. [Google Scholar] [CrossRef]
- Hamerman, J.A.; Page, S.T.; Pullen, A.M. Distinct methylation states of the CD8 beta gene in peripheral T cells and intraepithelial lymphocytes. J Immunol 1997, 159, 1240–1246. [Google Scholar]
- Lockhart, A.; Mucida, D.; Bilate, A.M. Intraepithelial Lymphocytes of the Intestine. Annu Rev Immunol 2024, 42, 289–316. [Google Scholar] [CrossRef]
- Morikawa, R.; Nemoto, Y.; Yonemoto, Y.; Tanaka, S.; Takei, Y.; Oshima, S.; Nagaishi, T.; Tsuchiya, K.; Nozaki, K.; Mizutani, T.; et al. Intraepithelial Lymphocytes Suppress Intestinal Tumor Growth by Cell-to-Cell Contact via CD103/E-Cadherin Signal. Cell Mol Gastroenterol Hepatol 2021, 11, 1483–1503. [Google Scholar] [CrossRef]
- Hartl, C.; Finke, J.; Hasselblatt, P.; Kreisel, W.; Schmitt-Graeff, A. Diagnostic and therapeutic challenge of unclassifiable enteropathies with increased intraepithelial CD103(+) CD8(+) T lymphocytes: a single center case series. Scand J Gastroenterol 2021, 56, 889–898. [Google Scholar] [CrossRef]
- Dietz, S.B.; Whitaker-Menezes, D.; Lessin, S.R. The role of alpha E beta 7 integrin (CD103) and E-cadherin in epidermotropism in cutaneous T-cell lymphoma. J Cutan Pathol 1996, 23, 312–318. [Google Scholar] [CrossRef]
- Xu, W.; Bergsbaken, T.; Edelblum, K.L. The multifunctional nature of CD103 (alphaEbeta7 integrin) signaling in tissue-resident lymphocytes. Am J Physiol Cell Physiol 2022, 323, C1161–C1167. [Google Scholar] [CrossRef]
- Yomogida, K.; Trsan, T.; Sudan, R.; Rodrigues, P.F.; Ulezko Antonova, A.; Ingle, H.; Luccia, B.D.; Collins, P.L.; Cella, M.; Gilfillan, S.; et al. The transcription factor Aiolos restrains the activation of intestinal intraepithelial lymphocytes. Nat Immunol 2024, 25, 77–87. [Google Scholar] [CrossRef]
- Jabri, B.; de Serre, N.P.; Cellier, C.; Evans, K.; Gache, C.; Carvalho, C.; Mougenot, J.F.; Allez, M.; Jian, R.; Desreumaux, P.; et al. Selective expansion of intraepithelial lymphocytes expressing the HLA-E-specific natural killer receptor CD94 in celiac disease. Gastroenterology 2000, 118, 867–879. [Google Scholar] [CrossRef]
- Melandri, D.; Zlatareva, I.; Chaleil, R.A.G.; Dart, R.J.; Chancellor, A.; Nussbaumer, O.; Polyakova, O.; Roberts, N.A.; Wesch, D.; Kabelitz, D.; et al. The gammadeltaTCR combines innate immunity with adaptive immunity by utilizing spatially distinct regions for agonist selection and antigen responsiveness. Nat Immunol 2018, 19, 1352–1365. [Google Scholar] [CrossRef]
- Girardi, M.; Lewis, J.M.; Filler, R.B.; Hayday, A.C.; Tigelaar, R.E. Environmentally responsive and reversible regulation of epidermal barrier function by gammadelta T cells. J Invest Dermatol 2006, 126, 808–814. [Google Scholar] [CrossRef]
- Nakandakari-Higa, S.; Canesso, M.C.C.; Walker, S.; Chudnovskiy, A.; Jacobsen, J.T.; Bilanovic, J.; Parigi, S.M.; Fiedorczuk, K.; Fuchs, E.; Bilate, A.M.; et al. Universal recording of cell-cell contacts in vivo for interaction-based transcriptomics. bioRxiv 2023. [Google Scholar] [CrossRef]
- Hariss, F.; Delbeke, M.; Guyot, K.; Zarnitzky, P.; Ezzedine, M.; Certad, G.; Meresse, B. Cytotoxic innate intraepithelial lymphocytes control early stages of Cryptosporidium infection. Front Immunol 2023, 14, 1229406. [Google Scholar] [CrossRef]
- Zhou, C.; Qiu, Y.; Yang, H. CD4CD8alphaalpha IELs: They Have Something to Say. Front Immunol 2019, 10, 2269. [Google Scholar] [CrossRef]
- Yakou, M.H.; Ghilas, S.; Tran, K.; Liao, Y.; Afshar-Sterle, S.; Kumari, A.; Schmid, K.; Dijkstra, C.; Inguanti, C.; Ostrouska, S.; et al. TCF-1 limits intraepithelial lymphocyte antitumor immunity in colorectal carcinoma. Sci Immunol 2023, 8, eadf2163. [Google Scholar] [CrossRef]
- Kornberg, A.; Botella, T.; Moon, C.S.; Rao, S.; Gelbs, J.; Cheng, L.; Miller, J.; Bacarella, A.M.; Garcia-Vilas, J.A.; Vargas, J.; et al. Gluten induces rapid reprogramming of natural memory alphabeta and gammadelta intraepithelial T cells to induce cytotoxicity in celiac disease. Sci Immunol 2023, 8, eadf4312. [Google Scholar] [CrossRef]
- Russell, G.J.; Nagler-Anderson, C.; Anderson, P.; Bhan, A.K. Cytotoxic potential of intraepithelial lymphocytes (IELs). Presence of TIA-1, the cytolytic granule-associated protein, in human IELs in normal and diseased intestine. Am J Pathol 1993, 143, 350–354. [Google Scholar]
- Abadie, V.; Discepolo, V.; Jabri, B. Intraepithelial lymphocytes in celiac disease immunopathology. Semin Immunopathol 2012, 34, 551–566. [Google Scholar] [CrossRef]
- Iijima, H.; Takahashi, I.; Kiyono, H. Mucosal immune network in the gut for the control of infectious diseases. Rev Med Virol 2001, 11, 117–133. [Google Scholar] [CrossRef]
- Santiago, L.; Castro, M.; Pardo, J.; Arias, M. Mouse Model of Colitis-Associated Colorectal Cancer (CAC): Isolation and Characterization of Mucosal-Associated Lymphoid Cells. Methods Mol Biol 2019, 1884, 189–202. [Google Scholar] [CrossRef]
- Gui, Y.; Cheng, H.; Zhou, J.; Xu, H.; Han, J.; Zhang, D. Development and function of natural TCR(+) CD8alphaalpha(+) intraepithelial lymphocytes. Front Immunol 2022, 13, 1059042. [Google Scholar] [CrossRef]
- Klose, C.S.N.; Hummel, J.F.; Faller, L.; d’Hargues, Y.; Ebert, K.; Tanriver, Y. A committed postselection precursor to natural TCRalphabeta(+) intraepithelial lymphocytes. Mucosal Immunol 2018, 11, 333–344. [Google Scholar] [CrossRef]
- Harada, Y.; Sujino, T.; Miyamoto, K.; Nomura, E.; Yoshimatsu, Y.; Tanemoto, S.; Umeda, S.; Ono, K.; Mikami, Y.; Nakamoto, N.; et al. Intracellular metabolic adaptation of intraepithelial CD4(+)CD8alphaalpha(+) T lymphocytes. iScience 2022, 25, 104021. [Google Scholar] [CrossRef]
- Morrow, N.M.; Morissette, A.; Mulvihill, E.E. Immunomodulation and inflammation: Role of GLP-1R and GIPR expressing cells within the gut. Peptides 2024, 176, 171200. [Google Scholar] [CrossRef]
- Canesso, M.C.C.; Lemos, L.; Neves, T.C.; Marim, F.M.; Castro, T.B.R.; Veloso, E.S.; Queiroz, C.P.; Ahn, J.; Santiago, H.C.; Martins, F.S.; et al. The cytosolic sensor STING is required for intestinal homeostasis and control of inflammation. Mucosal Immunol 2018, 11, 820–834. [Google Scholar] [CrossRef]
- Gao, J.; Xu, C.; Zhang, M.; Liu, J.; Wu, X.; Cui, C.; Wei, H.; Peng, J.; Zheng, R. Functional fiber enhances the effect of every-other-day fasting on insulin sensitivity by regulating the gut microecosystem. J Nutr Biochem 2022, 110, 109122. [Google Scholar] [CrossRef]
- Kadowaki, A.; Miyake, S.; Saga, R.; Chiba, A.; Mochizuki, H.; Yamamura, T. Gut environment-induced intraepithelial autoreactive CD4(+) T cells suppress central nervous system autoimmunity via LAG-3. Nat Commun 2016, 7, 11639. [Google Scholar] [CrossRef]
- Sujino, T.; London, M.; Hoytema van Konijnenburg, D.P.; Rendon, T.; Buch, T.; Silva, H.M.; Lafaille, J.J.; Reis, B.S.; Mucida, D. Tissue adaptation of regulatory and intraepithelial CD4(+) T cells controls gut inflammation. Science 2016, 352, 1581–1586. [Google Scholar] [CrossRef]
- London, M.; Bilate, A.M.; Castro, T.B.R.; Sujino, T.; Mucida, D. Stepwise chromatin and transcriptional acquisition of an intraepithelial lymphocyte program. Nat Immunol 2021, 22, 449–459. [Google Scholar] [CrossRef]
- Olivares-Villagomez, D.; Van Kaer, L. TL and CD8alphaalpha: Enigmatic partners in mucosal immunity. Immunol Lett 2010, 134, 1–6. [Google Scholar] [CrossRef]
- Olivares-Villagomez, D.; Mendez-Fernandez, Y.V.; Parekh, V.V.; Lalani, S.; Vincent, T.L.; Cheroutre, H.; Van Kaer, L. Thymus leukemia antigen controls intraepithelial lymphocyte function and inflammatory bowel disease. Proc Natl Acad Sci U S A 2008, 105, 17931–17936. [Google Scholar] [CrossRef]
- Xavier, R.J.; Podolsky, D.K. Unravelling the pathogenesis of inflammatory bowel disease. Nature 2007, 448, 427–434. [Google Scholar] [CrossRef]
- Ramos, G.P.; Papadakis, K.A. Mechanisms of Disease: Inflammatory Bowel Diseases. Mayo Clin Proc 2019, 94, 155–165. [Google Scholar] [CrossRef]
- Hung, C.T.; Ma, C.; Panda, S.K.; Trsan, T.; Hodel, M.; Frein, J.; Foster, A.; Sun, S.; Wu, H.T.; Kern, J.; et al. Western diet reduces small intestinal intraepithelial lymphocytes via FXR-Interferon pathway. Mucosal Immunol 2024, 17, 1019–1028. [Google Scholar] [CrossRef]
- Tougaard, P.; Skov, S.; Pedersen, A.E.; Krych, L.; Nielsen, D.S.; Bahl, M.I.; Christensen, E.G.; Licht, T.R.; Poulsen, S.S.; Metzdorff, S.B.; et al. TL1A regulates TCRgammadelta+ intraepithelial lymphocytes and gut microbial composition. Eur J Immunol 2015, 45, 865–875. [Google Scholar] [CrossRef]
- Abuquteish, D.; Putra, J. Upper gastrointestinal tract involvement of pediatric inflammatory bowel disease: A pathological review. World J Gastroenterol 2019, 25, 1928–1935. [Google Scholar] [CrossRef]
- Hu, M.D.; Edelblum, K.L. Sentinels at the frontline: the role of intraepithelial lymphocytes in inflammatory bowel disease. Curr Pharmacol Rep 2017, 3, 321–334. [Google Scholar] [CrossRef]
- Patterson, E.R.; Shmidt, E.; Oxentenko, A.S.; Enders, F.T.; Smyrk, T.C. Normal villous architecture with increased intraepithelial lymphocytes: a duodenal manifestation of Crohn disease. Am J Clin Pathol 2015, 143, 445–450. [Google Scholar] [CrossRef]
- van Hemert, S.; Skonieczna-Zydecka, K.; Loniewski, I.; Szredzki, P.; Marlicz, W. Microscopic colitis-microbiome, barrier function and associated diseases. Ann Transl Med 2018, 6, 39. [Google Scholar] [CrossRef]
- Miehlke, S.; Verhaegh, B.; Tontini, G.E.; Madisch, A.; Langner, C.; Munch, A. Microscopic colitis: pathophysiology and clinical management. Lancet Gastroenterol Hepatol 2019, 4, 305–314. [Google Scholar] [CrossRef]
- Yuan, L.; Wu, T.T.; Zhang, L. Microscopic colitis: lymphocytic colitis, collagenous colitis, and beyond. Hum Pathol 2023, 132, 89–101. [Google Scholar] [CrossRef]
- Burke, K.E.; D’Amato, M.; Ng, S.C.; Pardi, D.S.; Ludvigsson, J.F.; Khalili, H. Microscopic colitis. Nat Rev Dis Primers 2021, 7, 39. [Google Scholar] [CrossRef]
- Carreras, J. Artificial Intelligence Analysis of Celiac Disease Using an Autoimmune Discovery Transcriptomic Panel Highlighted Pathogenic Genes including BTLA. Healthcare (Basel) 2022, 10. [Google Scholar] [CrossRef]
- Carreras, J. Celiac Disease Deep Learning Image Classification Using Convolutional Neural Networks. J Imaging 2024, 10. [Google Scholar] [CrossRef]
- Catassi, C.; Verdu, E.F.; Bai, J.C.; Lionetti, E. Coeliac disease. Lancet 2022, 399, 2413–2426. [Google Scholar] [CrossRef]
- Villanacci, V.; Vanoli, A.; Leoncini, G.; Arpa, G.; Salviato, T.; Bonetti, L.R.; Baronchelli, C.; Saragoni, L.; Parente, P. Celiac disease: histology-differential diagnosis-complications. A practical approach. Pathologica 2020, 112, 186–196. [Google Scholar] [CrossRef]
- Al Somali, Z.; Hamadani, M.; Kharfan-Dabaja, M.; Sureda, A.; El Fakih, R.; Aljurf, M. Enteropathy-Associated T cell Lymphoma. Curr Hematol Malig Rep 2021, 16, 140–147. [Google Scholar] [CrossRef]
- Marchi, E.; Craig, J.W.; Kalac, M. Current and upcoming treatment approaches to uncommon subtypes of PTCL (EATL, MEITL, SPTCL, and HSTCL). Blood 2024, 144, 1898–1909. [Google Scholar] [CrossRef]
- Abdullah, S.A.A.; Goa, P.; Vandenberghe, E.; Flavin, R. Update on the Pathogenesis of Enteropathy-Associated T-Cell Lymphoma. Diagnostics (Basel) 2023, 13. [Google Scholar] [CrossRef]
- Liu, H.; Brais, R.; Lavergne-Slove, A.; Jeng, Q.; Payne, K.; Ye, H.; Liu, Z.; Carreras, J.; Huang, Y.; Bacon, C.M.; et al. Continual monitoring of intraepithelial lymphocyte immunophenotype and clonality is more important than snapshot analysis in the surveillance of refractory coeliac disease. Gut 2010, 59, 452–460. [Google Scholar] [CrossRef]
- Lebwohl, B.; Rubio-Tapia, A. Epidemiology, Presentation, and Diagnosis of Celiac Disease. Gastroenterology 2021, 160, 63–75. [Google Scholar] [CrossRef]
- Cerqueira, J.X.; Saavalainen, P.; Kurppa, K.; Laurikka, P.; Huhtala, H.; Nykter, M.; LEKoskinen, L.; Yohannes, D.A.; Kilpeläinen, E.; Shcherban, A.; Palotie, A. Independent and cumulative coeliac disease-susceptibility loci are associated with distinct disease phenotypes. J Hum Genet 2021, 66, 613–623. [Google Scholar] [CrossRef]
- Liu, E.; Lee, H.S.; Aronsson, C.A.; Hagopian, W.A.; Koletzko, S.; Rewers, M.J.; Eisenbarth, G.S.; Bingley, P.J.; Bonifacio, E.; Simell, V.; et al. Risk of pediatric celiac disease according to HLA haplotype and country. N Engl J Med 2014, 371, 42–49. [Google Scholar] [CrossRef]
- Garner, C.; Ahn, R.; Ding, Y.C.; Steele, L.; Stoven, S.; Green, P.H.; Fasano, A.; Murray, J.A.; Neuhausen, S.L. Genome-wide association study of celiac disease in North America confirms FRMD4B as new celiac locus. PLoS One 2014, 9, e101428. [Google Scholar] [CrossRef]
- Bragde, H.; Jansson, U.; Jarlsfelt, I.; Soderman, J. Gene expression profiling of duodenal biopsies discriminates celiac disease mucosa from normal mucosa. Pediatr Res 2011, 69, 530–537. [Google Scholar] [CrossRef]
- Ludvigsson, J.F.; Yao, J.; Lebwohl, B.; Green, P.H.R.; Yuan, S.; Leffler, D.A. Coeliac disease: complications and comorbidities. Nat Rev Gastroenterol Hepatol 2025. [Google Scholar] [CrossRef]
- Shan, L.; Molberg, O.; Parrot, I.; Hausch, F.; Filiz, F.; Gray, G.M.; Sollid, L.M.; Khosla, C. Structural basis for gluten intolerance in celiac sprue. Science 2002, 297, 2275–2279. [Google Scholar] [CrossRef]
- Sakly, W.; Thomas, V.; Quash, G.; El Alaoui, S. A role for tissue transglutaminase in alpha-gliadin peptide cytotoxicity. Clin Exp Immunol 2006, 146, 550–558. [Google Scholar] [CrossRef]
- Forsberg, G.; Hernell, O.; Melgar, S.; Israelsson, A.; Hammarstrom, S.; Hammarstrom, M.L. Paradoxical coexpression of proinflammatory and down-regulatory cytokines in intestinal T cells in childhood celiac disease. Gastroenterology 2002, 123, 667–678. [Google Scholar] [CrossRef]
- Eggesbo, L.M.; Risnes, L.F.; Neumann, R.S.; Lundin, K.E.A.; Christophersen, A.; Sollid, L.M. Single-cell TCR sequencing of gut intraepithelial gammadelta T cells reveals a vast and diverse repertoire in celiac disease. Mucosal Immunol 2020, 13, 313–321. [Google Scholar] [CrossRef]
- de Mascarel, A.; Belleannee, G.; Stanislas, S.; Merlio, C.; Parrens, M.; Laharie, D.; Dubus, P.; Merlio, J.P. Mucosal intraepithelial T-lymphocytes in refractory celiac disease: a neoplastic population with a variable CD8 phenotype. Am J Surg Pathol 2008, 32, 744–751. [Google Scholar] [CrossRef]
- Soderquist, C.R.; Lewis, S.K.; Gru, A.A.; Vlad, G.; Williams, E.S.; Hsiao, S.; Mansukhani, M.M.; Park, D.C.; Bacchi, C.E.; Alobeid, B.; et al. Immunophenotypic Spectrum and Genomic Landscape of Refractory Celiac Disease Type II. Am J Surg Pathol 2021, 45, 905–916. [Google Scholar] [CrossRef]
- Caja, S.; Maki, M.; Kaukinen, K.; Lindfors, K. Antibodies in celiac disease: implications beyond diagnostics. Cell Mol Immunol 2011, 8, 103–109. [Google Scholar] [CrossRef]
- Kim, S.M.; Mayassi, T.; Jabri, B. Innate immunity: actuating the gears of celiac disease pathogenesis. Best Pract Res Clin Gastroenterol 2015, 29, 425–435. [Google Scholar] [CrossRef]
- Van Laethem, F.; Donaty, L.; Tchernonog, E.; Lacheretz-Szablewski, V.; Russello, J.; Buthiau, D.; Almeras, M.; Moreaux, J.; Bret, C. LAIR1, an ITIM-Containing Receptor Involved in Immune Disorders and in Hematological Neoplasms. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Meyaard, L.; Adema, G.J.; Chang, C.; Woollatt, E.; Sutherland, G.R.; Lanier, L.L.; Phillips, J.H. LAIR-1, a novel inhibitory receptor expressed on human mononuclear leukocytes. Immunity 1997, 7, 283–290. [Google Scholar] [CrossRef]
- Carreras, J.; Lopez-Guillermo, A.; Kikuti, Y.Y.; Itoh, J.; Masashi, M.; Ikoma, H.; Tomita, S.; Hiraiwa, S.; Hamoudi, R.; Rosenwald, A.; et al. High TNFRSF14 and low BTLA are associated with poor prognosis in Follicular Lymphoma and in Diffuse Large B-cell Lymphoma transformation. J Clin Exp Hematop 2019, 59, 1–16. [Google Scholar] [CrossRef]
- Carreras, J.; Roncador, G.; Hamoudi, R. Ulcerative Colitis, LAIR1 and TOX2 Expression, and Colorectal Cancer Deep Learning Image Classification Using Convolutional Neural Networks. Cancers (Basel) 2024, 16. [Google Scholar] [CrossRef]
- Carreras, J.; Roncador, G.; Hamoudi, R. Dataset and AI Workflow for Deep Learning Image Classification of Ulcerative Colitis and Colorectal Cancer. Preprints 2024, 2024121201. [Google Scholar] [CrossRef]
- Zingone, F.; Bai, J.C.; Cellier, C.; Ludvigsson, J.F. Celiac Disease-Related Conditions: Who to Test? Gastroenterology 2024, 167, 64–78. [Google Scholar] [CrossRef]
- Sahin, Y. Celiac disease in children: A review of the literature. World J Clin Pediatr 2021, 10, 53–71. [Google Scholar] [CrossRef]
- Roshanzamir, N.; Zakeri, Z.; Rostami-Nejad, M.; Sadeghi, A.; Pourhoseingholi, M.A.; Shahbakhsh, Y.; Asadzadeh-Aghdaei, H.; Elli, L.; Zali, M.R.; Rezaei-Tavirani, M. Prevalence of celiac disease in patients with atypical presentations. Arab J Gastroenterol 2021, 22, 220–223. [Google Scholar] [CrossRef]
- Malamut, G.; Soderquist, C.R.; Bhagat, G.; Cerf-Bensussan, N. Advances in Nonresponsive and Refractory Celiac Disease. Gastroenterology 2024, 167, 132–147. [Google Scholar] [CrossRef]
- Green, P.H.R.; Paski, S.; Ko, C.W.; Rubio-Tapia, A. AGA Clinical Practice Update on Management of Refractory Celiac Disease: Expert Review. Gastroenterology 2022, 163, 1461–1469. [Google Scholar] [CrossRef]
- Scarmozzino, F.; Pizzi, M.; Pelizzaro, F.; Angerilli, V.; Dei Tos, A.P.; Piazza, F.; Savarino, E.V.; Zingone, F.; Fassan, M. Refractory celiac disease and its mimickers: a review on pathogenesis, clinical-pathological features and therapeutic challenges. Front Oncol 2023, 13, 1273305. [Google Scholar] [CrossRef]
- Rolny, P.; Sigurjonsdottir, H.A.; Remotti, H.; Nilsson, L.A.; Ascher, H.; Tlaskalova-Hogenova, H.; Tuckova, L. Role of immunosuppressive therapy in refractory sprue-like disease. Am J Gastroenterol 1999, 94, 219–225. [Google Scholar] [CrossRef]
- Vaidya, A.; Bolanos, J.; Berkelhammer, C. Azathioprine in refractory sprue. Am J Gastroenterol 1999, 94, 1967–1969. [Google Scholar] [CrossRef]
- Tack, G.J.; van Asseldonk, D.P.; van Wanrooij, R.L.; van Bodegraven, A.A.; Mulder, C.J. Tioguanine in the treatment of refractory coeliac disease--a single centre experience. Aliment Pharmacol Ther 2012, 36, 274–281. [Google Scholar] [CrossRef]
- Maurino, E.; Niveloni, S.; Chernavsky, A.; Pedreira, S.; Mazure, R.; Vazquez, H.; Reyes, H.; Fiorini, A.; Smecuol, E.; Cabanne, A.; et al. Azathioprine in refractory sprue: results from a prospective, open-label study. Am J Gastroenterol 2002, 97, 2595–2602. [Google Scholar] [CrossRef]
- Vivas, S.; Ruiz de Morales, J.M.; Ramos, F.; Suarez-Vilela, D. Alemtuzumab for refractory celiac disease in a patient at risk for enteropathy-associated T-cell lymphoma. N Engl J Med 2006, 354, 2514–2515. [Google Scholar] [CrossRef]
- Badran, Y.R.; Shih, A.; Leet, D.; Mooradian, M.J.; Coromilas, A.; Chen, J.; Kem, M.; Zheng, H.; Borowsky, J.; Misdraji, J.; et al. Immune checkpoint inhibitor-associated celiac disease. J Immunother Cancer 2020, 8. [Google Scholar] [CrossRef]
- Omiya, R.; Tsushima, F.; Narazaki, H.; Sakoda, Y.; Kuramasu, A.; Kim, Y.; Xu, H.; Tamura, H.; Zhu, G.; Chen, L.; et al. Leucocyte-associated immunoglobulin-like receptor-1 is an inhibitory regulator of contact hypersensitivity. Immunology 2009, 128, 543–555. [Google Scholar] [CrossRef]
- Colombo, B.M.; Canevali, P.; Magnani, O.; Rossi, E.; Puppo, F.; Zocchi, M.R.; Poggi, A. Defective expression and function of the leukocyte associated Ig-like receptor 1 in B lymphocytes from systemic lupus erythematosus patients. PLoS One 2012, 7, e31903. [Google Scholar] [CrossRef]
- Myers, L.K.; Winstead, M.; Kee, J.D.; Park, J.J.; Zhang, S.; Li, W.; Yi, A.K.; Stuart, J.M.; Rosloniec, E.F.; Brand, D.D.; et al. 1,25-Dihydroxyvitamin D3 and 20-Hydroxyvitamin D3 Upregulate LAIR-1 and Attenuate Collagen Induced Arthritis. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Spiliopoulou, A.; Iakovliev, A.; Plant, D.; Sutcliffe, M.; Sharma, S.; Cubuk, C.; Lewis, M.; Pitzalis, C.; Barton, A.; McKeigue, P.M. Genome-Wide Aggregated Trans Effects Analysis Identifies Genes Encoding Immune Checkpoints as Core Genes for Rheumatoid Arthritis. Arthritis Rheumatol 2025. [Google Scholar] [CrossRef]
- Zhang, X.; He, X.; Zhang, M.; Wu, T.; Liu, X.; Zhang, Y.; Xie, Z.; Liu, S.; Xia, T.; Wang, Y.; et al. Efficient delivery of the lncRNA LEF1-AS1 through the antibody LAIR-1 (CD305)-modified Zn-Adenine targets articular inflammation to enhance the treatment of rheumatoid arthritis. Arthritis Res Ther 2023, 25, 238. [Google Scholar] [CrossRef]
- Agashe, V.V.; Jankowska-Gan, E.; Keller, M.; Sullivan, J.A.; Haynes, L.D.; Kernien, J.F.; Torrealba, J.R.; Roenneburg, D.; Dart, M.; Colonna, M.; et al. Leukocyte-Associated Ig-like Receptor 1 Inhibits T(h)1 Responses but Is Required for Natural and Induced Monocyte-Dependent T(h)17 Responses. J Immunol 2018, 201, 772–781. [Google Scholar] [CrossRef]
- Joseph, C.; Alsaleem, M.A.; Toss, M.S.; Kariri, Y.A.; Althobiti, M.; Alsaeed, S.; Aljohani, A.I.; Narasimha, P.L.; Mongan, N.P.; Green, A.R.; et al. The ITIM-Containing Receptor: Leukocyte-Associated Immunoglobulin-Like Receptor-1 (LAIR-1) Modulates Immune Response and Confers Poor Prognosis in Invasive Breast Carcinoma. Cancers (Basel) 2020, 13. [Google Scholar] [CrossRef]
- Tripathi, S.; Najem, H.; Dussold, C.; Pacheco, S.; Miska, J.; McCortney, K.; Steffens, A.; Walshon, J.; Winkowski, D.; Cloney, M.; et al. Cancer-associated fibroblast-secreted collagen is associated with immune inhibitor receptor LAIR1 in gliomas. J Clin Invest 2024, 134. [Google Scholar] [CrossRef]
- Rodriguez, B.L.; Huang, J.; Gibson, L.; Fradette, J.J.; Chen, H.H.; Koyano, K.; Cortez, C.; Li, B.; Ho, C.; Ashique, A.M.; et al. Antitumor Activity of a Novel LAIR1 Antagonist in Combination with Anti-PD1 to Treat Collagen-Rich Solid Tumors. Mol Cancer Ther 2024, 23, 1144–1158. [Google Scholar] [CrossRef]
- Pan, B.; Ke, X.; Qiu, J.; Ye, D.; Zhang, Z.; Zhang, X.; Luo, Y.; Yao, Y.; Wu, X.; Wang, X.; et al. LAIR1-mediated resistance of hepatocellular carcinoma cells to T cells through a GSK-3beta/beta-catenin/MYC/PD-L1 pathway. Cell Signal 2024, 115, 111039. [Google Scholar] [CrossRef]
- Carreras, J. Artificial Intelligence Analysis of Ulcerative Colitis Using an Autoimmune Discovery Transcriptomic Panel. Healthcare (Basel) 2022, 10. [Google Scholar] [CrossRef]
- Denholm, J.; Schreiber, B.A.; Evans, S.C.; Crook, O.M.; Sharma, A.; Watson, J.L.; Bancroft, H.; Langman, G.; Gilbey, J.D.; Schonlieb, C.B.; et al. Multiple-instance-learning-based detection of coeliac disease in histological whole-slide images. J Pathol Inform 2022, 13, 100151. [Google Scholar] [CrossRef]
- Molder, A.; Balaban, D.V.; Molder, C.C.; Jinga, M.; Robin, A. Computer-Based Diagnosis of Celiac Disease by Quantitative Processing of Duodenal Endoscopy Images. Diagnostics (Basel) 2023, 13. [Google Scholar] [CrossRef]
- Scheppach, M.W.; Rauber, D.; Stallhofer, J.; Muzalyova, A.; Otten, V.; Manzeneder, C.; Schwamberger, T.; Wanzl, J.; Schlottmann, J.; Tadic, V.; et al. Detection of duodenal villous atrophy on endoscopic images using a deep learning algorithm. Gastrointest Endosc 2023, 97, 911–916. [Google Scholar] [CrossRef]
- Schreiber, B.A.; Denholm, J.; Gilbey, J.D.; Schonlieb, C.B.; Soilleux, E.J. Stain normalization gives greater generalizability than stain jittering in neural network training for the classification of coeliac disease in duodenal biopsy whole slide images. J Pathol Inform 2023, 14, 100324. [Google Scholar] [CrossRef]
- Stoleru, C.A.; Dulf, E.H.; Ciobanu, L. Automated detection of celiac disease using Machine Learning Algorithms. Sci Rep 2022, 12, 4071. [Google Scholar] [CrossRef] [PubMed]
- DiPalma, J.; Suriawinata, A.A.; Tafe, L.J.; Torresani, L.; Hassanpour, S. Resolution-based distillation for efficient histology image classification. Artif Intell Med 2021, 119, 102136. [Google Scholar] [CrossRef] [PubMed]
- Gruver, A.M.; Lu, H.; Zhao, X.; Fulford, A.D.; Soper, M.D.; Ballard, D.; Hanson, J.C.; Schade, A.E.; Hsi, E.D.; Gottlieb, K.; et al. Pathologist-trained machine learning classifiers developed to quantitate celiac disease features differentiate endoscopic biopsies according to modified marsh score and dietary intervention response. Diagn Pathol 2023, 18, 122. [Google Scholar] [CrossRef]
- Alharbi, E.; Rajaram, A.; Cote, K.; Farag, M.; Maleki, F.; Gao, Z.H.; Maedler-Kron, C.; Marcus, V.; Fiset, P.O. A Deep Learning-Based Approach to Estimate Paneth Cell Granule Area in Celiac Disease. Arch Pathol Lab Med 2024, 148, 828–835. [Google Scholar] [CrossRef] [PubMed]
Figure 3.
LAIR1 expression in reactive tonsils and crystal structure of human LAIR1. LAIR1 is expressed in both follicles and interfollicular areas of reactive tonsils. In the secondary follicles, the pattern is characteristic of macrophage/dendritic cells in the germinal center, and of naïve B-lymphocytes in the mantle zone. In the interfollicular area, the pattern is compatible with dendritic cells, including in the lymphoepithelial area. Occasional intraepithelial lymphocytes are also positive. The crystal structure of human LAIR1 in the C2 space group is also shown; experimental data using the X-ray diffraction method (website: https://www.rcsb.org/structure/3RP1; accessed on March 25, 2025).
Figure 3.
LAIR1 expression in reactive tonsils and crystal structure of human LAIR1. LAIR1 is expressed in both follicles and interfollicular areas of reactive tonsils. In the secondary follicles, the pattern is characteristic of macrophage/dendritic cells in the germinal center, and of naïve B-lymphocytes in the mantle zone. In the interfollicular area, the pattern is compatible with dendritic cells, including in the lymphoepithelial area. Occasional intraepithelial lymphocytes are also positive. The crystal structure of human LAIR1 in the C2 space group is also shown; experimental data using the X-ray diffraction method (website: https://www.rcsb.org/structure/3RP1; accessed on March 25, 2025).
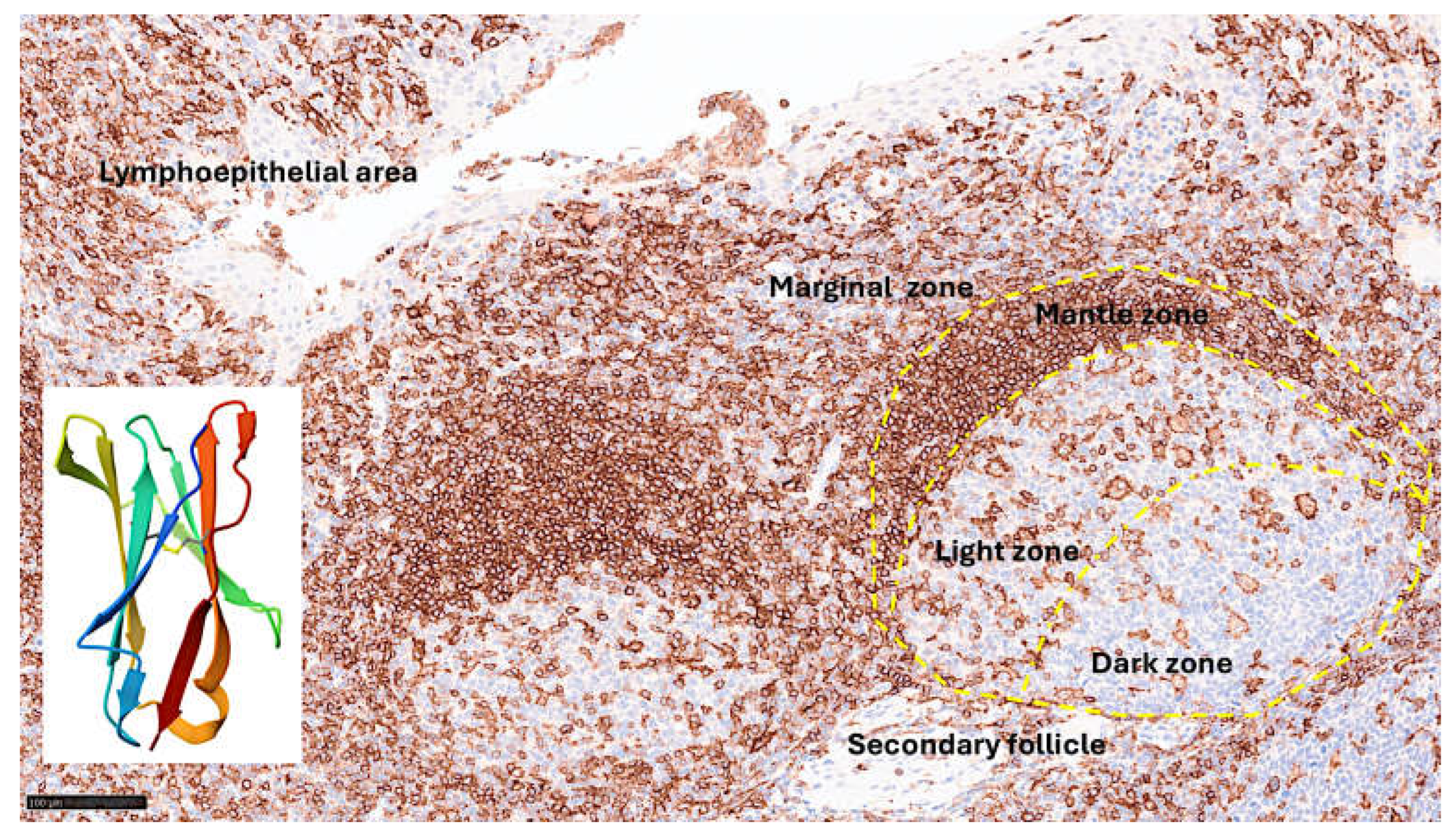
Figure 4.
Immunophenotype characterization of IELs in intestinal mucosa control. Most IELs were CD3, CD103, and CD8-positive. CD4-positive cells were mainly found in the lamina propria.
Figure 4.
Immunophenotype characterization of IELs in intestinal mucosa control. Most IELs were CD3, CD103, and CD8-positive. CD4-positive cells were mainly found in the lamina propria.
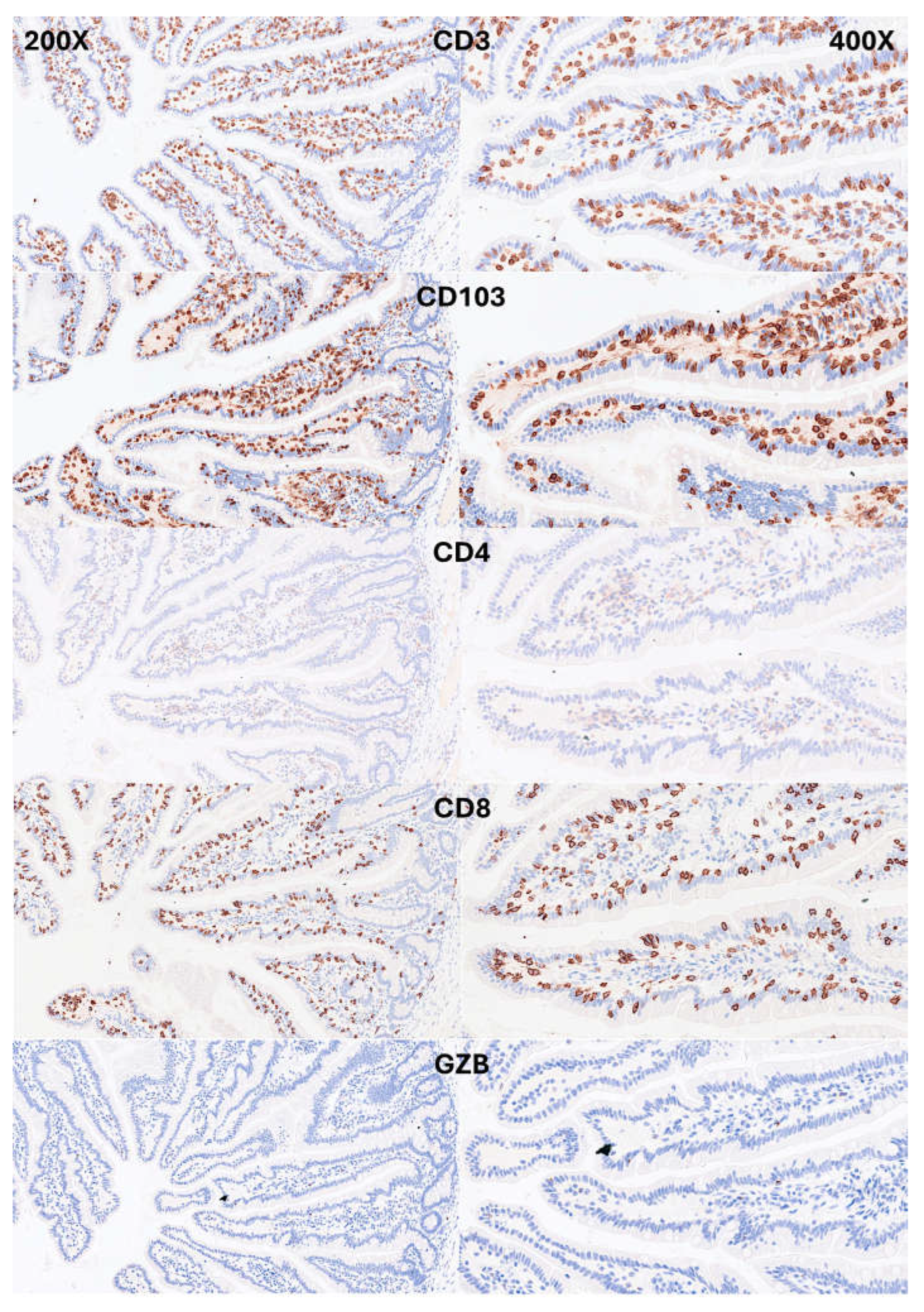
Figure 5.
Immunophenotype characterization of IELs in intestinal mucosa control. Most IELs expressed TCRβ chains; therefore, expressed the TCRαβ chains. Occasionally, CD56+IELs were found, as well as TCRδ chain-positive IELs (i.e., TCRγδ+IELs).
Figure 5.
Immunophenotype characterization of IELs in intestinal mucosa control. Most IELs expressed TCRβ chains; therefore, expressed the TCRαβ chains. Occasionally, CD56+IELs were found, as well as TCRδ chain-positive IELs (i.e., TCRγδ+IELs).
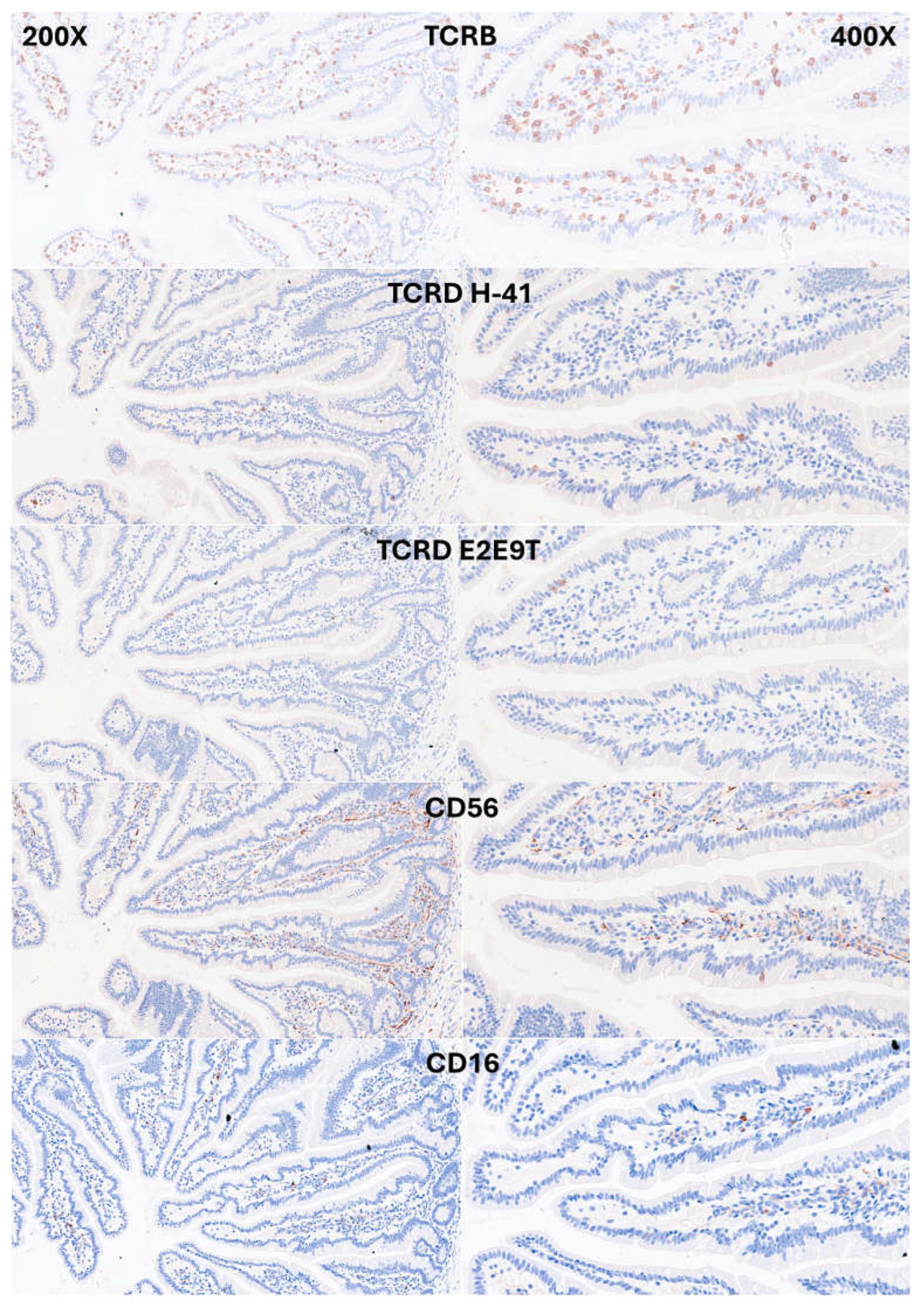
Figure 6.
Immunophenotype characterization of IELs in the intestinal mucosa control. IELs were diffusely and strongly positive for LAIR1. LAIR1 also marked the inflammatory infiltrate of the lamina propria. PD-L1 and BTLA expression was limited in the lamina propria. Occasional PD1-positive cells were identified. TOX2+IELs were occasionally found.
Figure 6.
Immunophenotype characterization of IELs in the intestinal mucosa control. IELs were diffusely and strongly positive for LAIR1. LAIR1 also marked the inflammatory infiltrate of the lamina propria. PD-L1 and BTLA expression was limited in the lamina propria. Occasional PD1-positive cells were identified. TOX2+IELs were occasionally found.
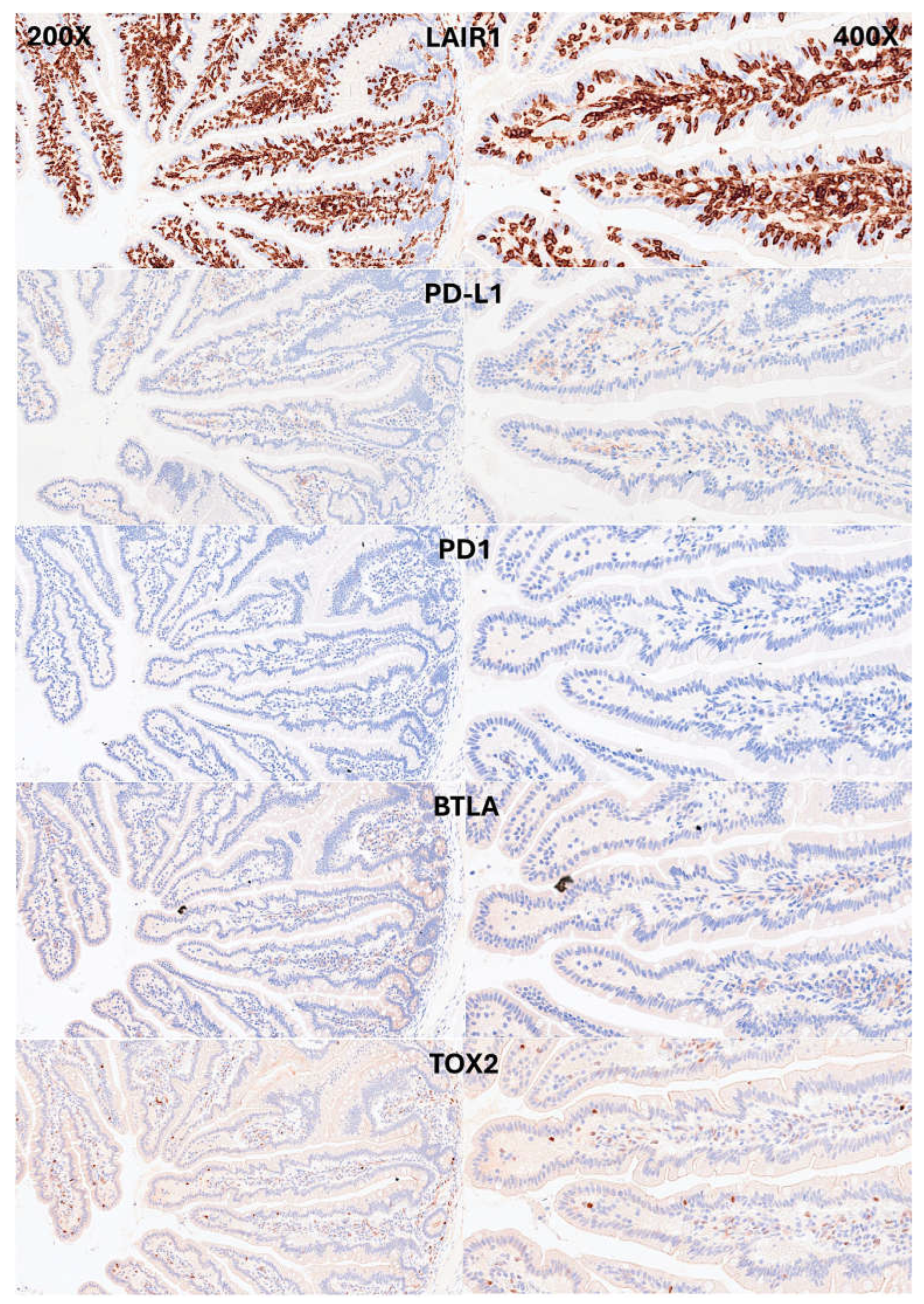
Figure 7.
Immunophenotype characterization of IELs in intestinal mucosa control. CD163 and HLA-DPDR identified antigen-presenting cells (APCs), mainly macrophages and dendritic cells in the lamina propria. Few FOXP3+Tregs were identified in the lamina propria.
Figure 7.
Immunophenotype characterization of IELs in intestinal mucosa control. CD163 and HLA-DPDR identified antigen-presenting cells (APCs), mainly macrophages and dendritic cells in the lamina propria. Few FOXP3+Tregs were identified in the lamina propria.
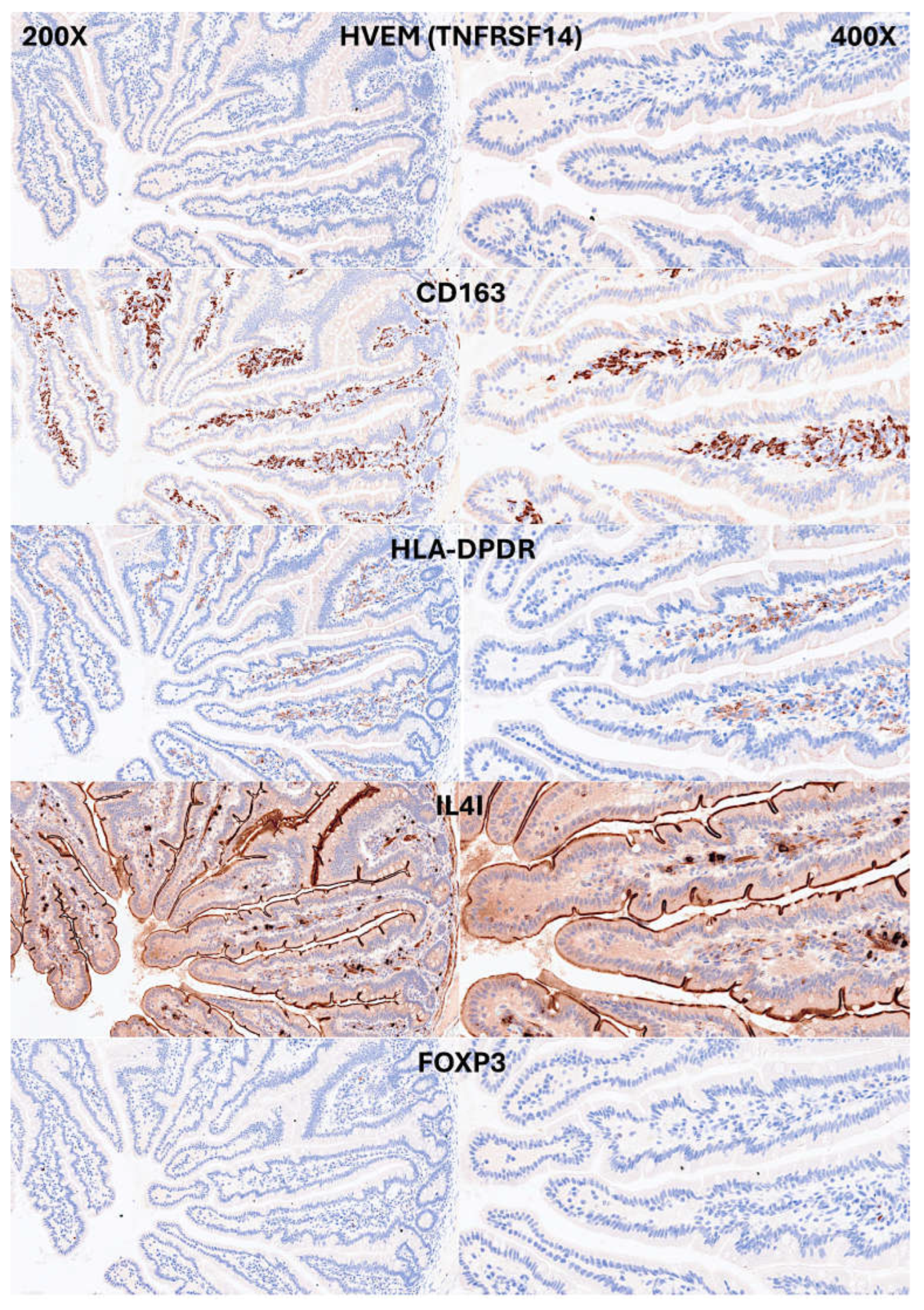
Figure 8.
Confocal microscopy showing double immunofluorescence between CD3 (red) and LAIR1(green) in the small intestine control. The IELs were double CD3 and LAIR1-positive.
Figure 8.
Confocal microscopy showing double immunofluorescence between CD3 (red) and LAIR1(green) in the small intestine control. The IELs were double CD3 and LAIR1-positive.

Table 1.
Details of primary antibodies.
| Antibody | Target/pathway | Source | Details |
| CD3 | T-lymphocytes | Leica | Mouse monoclonal, clone LN10, IgG1, C-terminal region |
| CD4 | Helper T-lymphocytes (+antigen presenting cells) | Leica | Mouse monoclonal, clone 4B12, IgG1, external domain |
| CD8 | Cytotoxic T-lymphocytes | Leica | Mouse monoclonal, clone 4B11, IgG2b, alpha chain cytoplasmic portion |
| CD103 | Alpha E integrin & human mucosal lymphocyte antigen 1 (ITGAE), intraepithelial T lymphocytes, FOXP3+Tregs, CD4+ and CD8+Tcells, dendritic cells, and mast cells in mucosal tissues. Interacts with E-cadherin (epithelial cells) | Leica | Rabbit monoclonal, clone EP206, IgG, residues of human CD103/ITGAE protein |
| Granzyme B | Lytic granules of cytotoxic-T lymphocytes (CTL) and in natural killer (NK) cells | Leica | Mouse monoclonal, clone 11F1, IgG2a, N-terminus of the mature granzyme B molecule |
| TCRβ | T-cell receptor | CST | Rabbit IgG, residues near the amino terminus of human TRBC1/TCRβ constant region 1 protein |
| TCRδ | T-cell receptor | CST | Rabbit IgG, total TRDC/TCRδ protein |
| CD56 (NCAM) | Neurons, astrocytes, Schwann cells, NK cells and a subset of activated T-lymphocytes | Leica | Mouse monoclonal, clone CD564, IgG2b, extracellular domain |
| CD16 | NK cells, granulocytes, activated macrophages and subset T cells (TCRαβ and TCRγδ) | Leica | Mouse monoclonal, clone 2H7, IgG2a, external domain (both transmembrane and GPI-linked forms) |
| LAIR1 (CD305) | Co-inhibitory receptor | CNIO | Rat monoclonal, clone JAVI82A, IgG2a, k |
| PD-L1 | Immune suppression and inhibition of T-cell activity | Leica | Rabbit IgG, clone 73-10, C-terminal domain |
| PD1 (CD279) | Co-inhibitory receptor | CNIO | Mouse monoclonal, clone NAT105, IgG1 |
| BTLA (CD272) | Co-inhibitory receptor | CNIO | Mouse monoclonal, clone FLO67B, IgG1 |
| TOX2 | Transcription factor, maturation of NK cells and differentiation of T follicular helper (TFH) cells | CNIO | Rat monoclonal, clone TOM924D, IgG2a |
| HVEM (TNFRSF14) | Ligand of BTLA | Abcam | Rabbit polyclonal, IgG, exact immunogen is proprietary information |
| CD163 | M2-like macrophages | Leica | Mouse monoclonal, clone 10D6, IgG1, N-terminal region |
| HLA-DP-DQ | Antigen presentation by APC | CNIO | Mouse monoclonal, clone JS76, IgG2a |
| IL4I1 | APC, T-cell inhibition | CNIO | Rat monoclonal, clone BALI265E,543H,573B, IgG2a |
| FOXP3 | Regulatory T-lymphocytes (Tregs) | CNIO | Mouse monoclonal, clone 236A, IgG1 |
APC, antigen-presenting cells (B-lymphocytes, dendritic cells, and macrophages); CST, Cell Signaling Technology; Leica, Leica Biosystems K.K.; NK, natural killer cells; Spanish National Cancer Research Center (CNIO);.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



