Submitted:
22 April 2025
Posted:
22 April 2025
You are already at the latest version
Abstract
Water-deficit stress is typically viewed as detrimental to agricultural yields. It has been found to enhance secondary metabolite concentrations in certain essential oil-producing plants, including Cannabis sativa L. Cannabis is a versatile plant from the Cannabaceae family used for its fibers, seeds, and bioactive compounds, including medicinal and recreational cannabinoids. Furthermore, it exhibits significant metabolic shifts under water-deficit stress conditions, which may impact the production of these resources. This review explores the physiological mechanisms underlying the metabolic responses of cannabis to water-deficit stress, focusing on how water-deficit stress could promote the accumulation of secondary metabolites. Water-deficit stress induces metabolic changes in cannabis, leading to secondary metabolite accumulation. Water shortage causes stomatal closure, significantly reducing CO2 uptake and fixation via the Calvin cycle, leading to an oversupply of NADPH+H+. This oversupply allows metabolic processes to shift toward synthesizing highly reduced compounds, such as secondary metabolites. Overall, the literature suggests that the controlled application of water-deficit stress during cannabis cultivation can enhance cannabinoid quality and yield, offering a practical strategy for optimizing plant productivity while addressing current knowledge gaps in metabolic signaling pathways.
Keywords:
Drought
; irrigation
; cannabinoids
; induced stress
; terpenes
1. Introduction
Cannabis sativa L. (Cannabaceae) is a multipurpose cultivated crop that is native to Central Asia [1]. There is consensus that C. sativa is certainly of Old-World origin, with some of the earliest indications dating back to around 12,000 years ago [2]. However, there is still some debate about its geographical origin [3,4].
C. sativa is an herbaceous and dioecious flowering plant. Depending on taxonomic classification, cannabis is frequently viewed as a single species, C. sativa, with three varieties including C. sativa var. sativa, C. sativa var. indica, and C. sativa var. ruderalis, each with distinct characteristics and uses [5]. However, the three subspecies or varieties interbreed, and their species boundaries are fluid. Therefore, it has been suggested that only a single cannabis species, C. sativa, be recognized [6]. The two most common varieties are C. sativa var. sativa and C. sativa var. indica which contain relatively high amounts of tetrahydrocannabinol (THC) and are used primarily for medicinal and recreational purposes. In contrast, the less common variety, C. sativa var. ruderalis is the smallest in height and girth and contains relatively lower amounts of THC and other cannabinoids [7].
Cannabis is usually called “hemp” when used as a source of fiber, “hemp seed” when used as a source of seed oil, and “marijuana” (more commonly spelled “marihuana” in the past) when used as a source of mood-altering substances and therapeutic compounds. Similarly, “hemp” is also used to describe cultivars grown for resin production which contains high levels of CBD (cannabidiol) and low amounts of THC. Although the terms “hemp” and “marijuana” are likely the most frequently used common names in scientific publications, it is crucial to distinguish between the two while recognizing that they belong to the same species. Specifically, the concentration of delta-9 tetrahydrocannabinol (Δ9-THC) is the primary parameter for distiguishing between them, according to legislative definitions [8].
Cannabis is often cited as a prime example of a crop that initially evolved as a “camp follower” [9]. According to this hypothesis, early humans collected plants for various uses and discarded them in disturbed areas around camps which are ideal for seed germination. Species classified as “camp followers” typically exhibit vigorous growth and colonization traits, which are key features of weedy plants, including C. sativa. As a result, C. sativa is exceptionally well suited to this habitat, with its weedy nature facilitating both natural and human-assisted dispersal. This ability to thrive in human-disturbed environments likely facilitated its early cultivation and eventual domestication. Indeed, C. sativa is one of the world’s oldest crops, cultivated for at least 6,000 years [3]. For centuries, cannabis has been utilized as a source of fiber, food, oil, and medicine, as well as for recreational and religious purposes [10]; these properties likely contributed to early domestication by prehistoric humans. It contains numerous biologically active compounds, such as cannabinoids, terpenoids, flavonoids, and alkaloids [11]. A shared characteristic of all cannabis plants is the presence of cannabinoids, which are sometimes referred to more precisely as “phytocannabinoids” to distinguish them from endocannabinods found and produced in the human body. While there are only two main endocannabinoids in humans (anadamide and 2-arachidonoylglycerol), there are over 100 different phytocannabinoids in cannabis, predominantly generated in trichome glands that develop in female inflorescences [6].
The medical potential of cannabis plants, particularly phytocannabinoids and their chemical derivatives, has drawn public attention over the past years [15]. For example, CBD has been widely recognized in treating Parkinson’s disease, schizophrenia, pain, anxiety, depression, and other neurological disorders [15], while THC has shown success in the treatment of several serious conditions, such as multiple sclerosis, cancer-related pain, nausea, and appetite loss in AIDS patients [12]. However, to be effective, THC must be administered in sufficient doses, at levels which can cause psychoactive side effects [12]. The psychoactive properties of THC are the primary reason the compound and the plants that produce it are still illegal in many places [13]. This makes it difficult for some researchers to study phytocannabinoids for their potential medicinal properties.
Along with primary metabolites essential for a plant's growth and development, plants also produce secondary metabolites that play crucial roles in plant adaptation and survival under stress conditions [14]. Environmental conditions are one of the best alternatives to breeding for the production and accumulation of cannabinoids in cannabis [16]. Like other medicinal crops, the production of cannabinoids, particularly THC and CBD, is significantly affected by a variety of environmental stresses such as light [17], temperature [16], nutrition [18], heavy metals [19], phytohormones [20], and biotic stresses including insects and microbial pathogens [1]. Hence, applying controlled water-deficit stress can potentially be an effective technique for controlled environment agriculture to boost the production of secondary metabolites in cannabis.
Water-deficit stress refers to a controlled reduction in water availability and is a more appropriate terminology for indoor controlled environments, growth chambers, and greenhouses, where water limitations are intentionally imposed to study plant responses under specific conditions. Drought, on the other hand, refers to a naturally occurring event where an area experiences below-normal precipitation, either rain or snow, due to climatic conditions [21]. This terminology is most accurately applied to field or outdoor studies, where environmental factors are uncontrolled. The accumulation of secondary metabolites due to drought and water-deficit stress has been documented in several herbaceous species, but their effects on cannabis secondary metabolism are poorly studied. The increasing medical interest in and economic value of C. sativa illustrate the importance of reviewing relevant research on how water-deficit stress affects cannabinoid production in cannabis.
2. Methodology
This review aims to assess the impact of water-deficit stress on the growth, yield, and production of secondary metabolites in cannabis, focusing on cannabinoid concentration. We searched combinations of keywords such as “cannabis cultivation”, “water-deficit stress”,” drought in Cannabis sativa”, “water-deficit stress and secondary metabolites”, and related terms. A comprehensive literature search was conducted to identify relevant journal articles, research papers, books, and published reports. The articles were sourced from ResearchGate, Google Scholar, and other academic databases.
3. Impact of Water-Deficit Stress on Plants
3.1. Overall Plant Growth
Stress may cause various effects on plants, most of which are detrimental to the health and yield of crops. It can influence primary and secondary metabolism, physiology, gene expression, the structure of organs and tissues, and overall total plant growth (Figure 1). The complexity of this response is impacted significantly by the length and severity of stress, the simultaneous presence of multiple stressors, the plant's genotype, and the developmental period at which the stress is applied [22,23]. Abiotic stressors such as inadequate water content, salt, excessive light, extreme heat/cold, and deregulated nutrient availability can impair plant growth and development [24], which results in significant crop losses globally, making abiotic stressors one of the most consequential issues that agriculture faces today [25] (Figure 2).
Water-deficit stress and drought are significant abiotic factors influencing both cultivated and wild plant species, eliciting diverse physiological responses often modulated by genetic factors. Drought is defined as a prolonged period of below-normal precipitation, which reduces soil water availability for plant growth and development. Drought typically coincides with high air temperatures, leading to increased plant evapotranspiration, stomatal closure, and reduced photosynthetic activity, ultimately resulting in lower yields. The frequency of drought stress events is rising, driven not only by global temperature increases but also by the overuse of natural resources and soil desertification [26]. While plants may adapt to water-deficit stress situations through various tolerance mechanisms, these adaptations can negatively impact yield as the complex interactions between gene expresssion and plant physiology come into play.
Plants adapt to drought stress or water-deficit stress by regulating photosynthesis through physiological, biochemical, or other specialized mechanisms. Photosynthesis is highly sensitive to water scarcity and high temperatures [27]. Morales et al. (2020) [28] reviewed the impact of water deficit and high temperatures on the photosynthetic system and plant growth (Figure 1). Drought stress causes water shortages within the plant, affects its nutritional balance, alters xylem pH, changes farnesyl transferase activity, modifies xylem hydraulic conductance, and reduces the vapor pressure difference between the leaves and the air. These changes lead to stomatal closure in leaves [29]. According to Dias (2010) [30], drought stress-induced stomatal closure and decreased mesophyll conductance hinder CO2 diffusion from the atmosphere to the carboxylation sites, thereby reducing photosynthesis. Drought also inhibits plant cell growth and development, especially in stems and leaves, leading to cytoplasmic water loss, as noted by [31]. This water loss changes cell volume, vacuole size, turgor pressure sensing, membrane permeability, and intracellular osmotic balance (Figure 2) [32]. Reduced growth often occurs following drought stress because of insufficient cell expansion in plants [33]. Turgor pressure is crucial for cell expansion and growth, allowing cells to form specific differentiated regions. When drought causes turgor pressure to drop below a critical threshold, differentiation and cell expansion halt, leading to decreased plant growth (Figure 1) [34]. Additionally, drought stress reduces nutritional absorption, where water scarcity decreases potassium levels particularly in leaves, which affecs stomatal movement and the turgidity of guard cells. Consequently, this reduces photosynthesis and diminishes plant biomass production [35].
The effects of varying levels of drought stress on Plantago psyllium L. (psyllium), Achillea millefolium L. (yarrow), Salvia officinalis L. (sage), Calendula officinalis L. (marigold), and Matricaria chamomilla L. (chamomile) led to reduced shoot weight and plant height compared to non-stressed conditions [36]. Similarly, sunflower (Helianthus annuus L.) yields significantly decreased under a 40% field capacity due to reduced cellular division during seed growth and maturation phases, which resulted in lower seed weight and oil weight, but not oil concentrations [33]. Previous studies also reported that drought stress shortened the number of days to flowering and ripening in fenugreek [37] and beans [38], and reduced the growing degree days and growth period in thyme [39]. All of these observations are consistent with reduction in photosynthesis.
3.2. Photosynthesis and Physiology
Drought-stress conditions result in reduced biomass due to decreased leaf area and lower photosynthetic rates, which are caused by biochemical limitations and water scarcity, including a reduction in photosynthetic pigments [40]. Photosynthetic rates decline during drought due to mesophyll damage caused by increased stomatal resistance that prevents CO2 from entering the plant. This results in decreased net photosynthesis and reduced plant biomass [41]. Drought significantly impacts ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco), the enzyme essential for CO2 fixation, reducing its carboxylation function by approximately 47.5-50%. However, this inhibition is reversible once the drought stress is alleviated [27]. Drought affects various aspects of photosynthesis, including carbon fixation mechanisms such as stomatal aperture and Rubisco activity, as well as light-harvesting processes. During drought conditions, the photosynthetic rate decreases because the acquired light is not fully converted into chemically bound energy. This leads to photoinhibition and reduces the maximal quantum yield of PSII reaction centers (Fv/Fm) [26].
The overall process of photosynthesis is hindered because of Photosystems I and II. This decline in photosystem efficacy is suggested to be related to a general decrease in electron absorption, leading to reduced energy available for carbon dioxide fixation [42]. Excess photons are absorbed by chlorophyll, and when the photosystem's capacity to absorb photons is reduced, it produces reactive oxygen species (ROS). These ROS interfere with various cellular components, diminishing biological activities of these components [43]. Specifically, ROS can cause oxidation-induced damage to DNA, lipids, cell membranes, and cell walls [44]. During drought stress, the reduced capacity of photosystems to absorb photons prevents the efficient production of NADPH through electron transfer across thylakoid membranes, further exacerbating the stress on the photosynthetic process (Figure 3b) [43]. In alfalfa (Medicago sativa L.), under drought stress conditions, there is a reduction in Photosystem II functionality, while Photosystem I remains unaffected mainly [27]. In contrast, in spinach (Spinacia oleracea L.), Photosystem II is less severely impacted by drought stress due to a physiological response that slows electron flow to Photosystem II [42].
3.3. Plant Secondary Metabolites
In some medicinal plants, drought stress can paradoxically enhance the production of secondary metabolites, which serve as a survival mechanism against environmental stressors despite the limitations imposed by water availability (Figure 2). Drought stress significantly influences the synthesis of secondary metabolites, which is essential for these plants to thrive and produce valuable natural substances [29]. The generation of ROS under water-deficit stress conditions may have promoted the production of secondary metabolites, thereby modifying the pathways involved in plant cell signal transduction [45]. Typically, water deficits suppress biomass formation (Figure 2), causing the plants to redirect absorbed CO2 toward synthesizing carbon-based secondary metabolites. This redirection disrupts the usual sugar-mediated feedback loop that promotes photosynthesis, increasing the production of secondary metabolites (Figure 3b) [46].
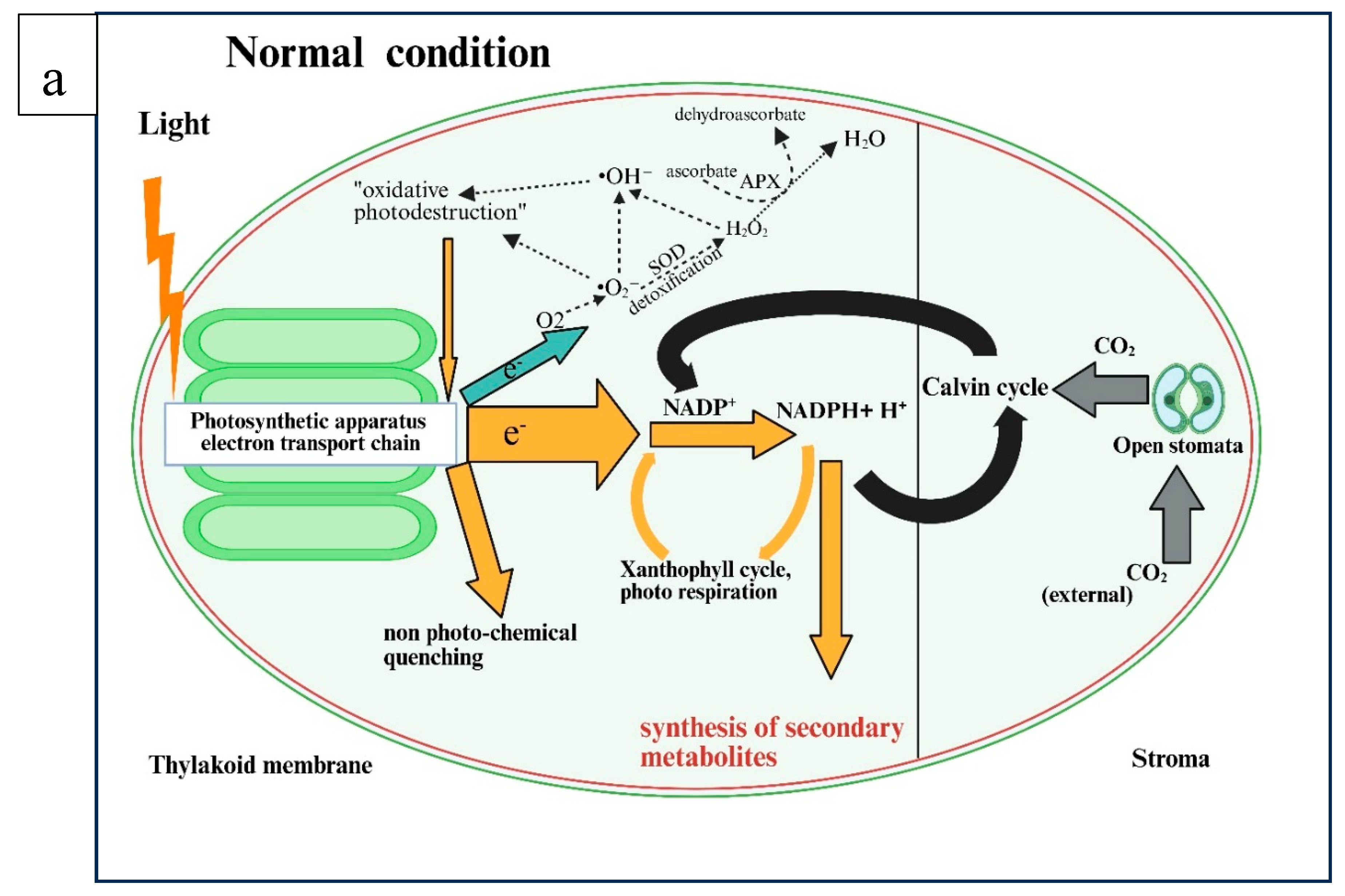
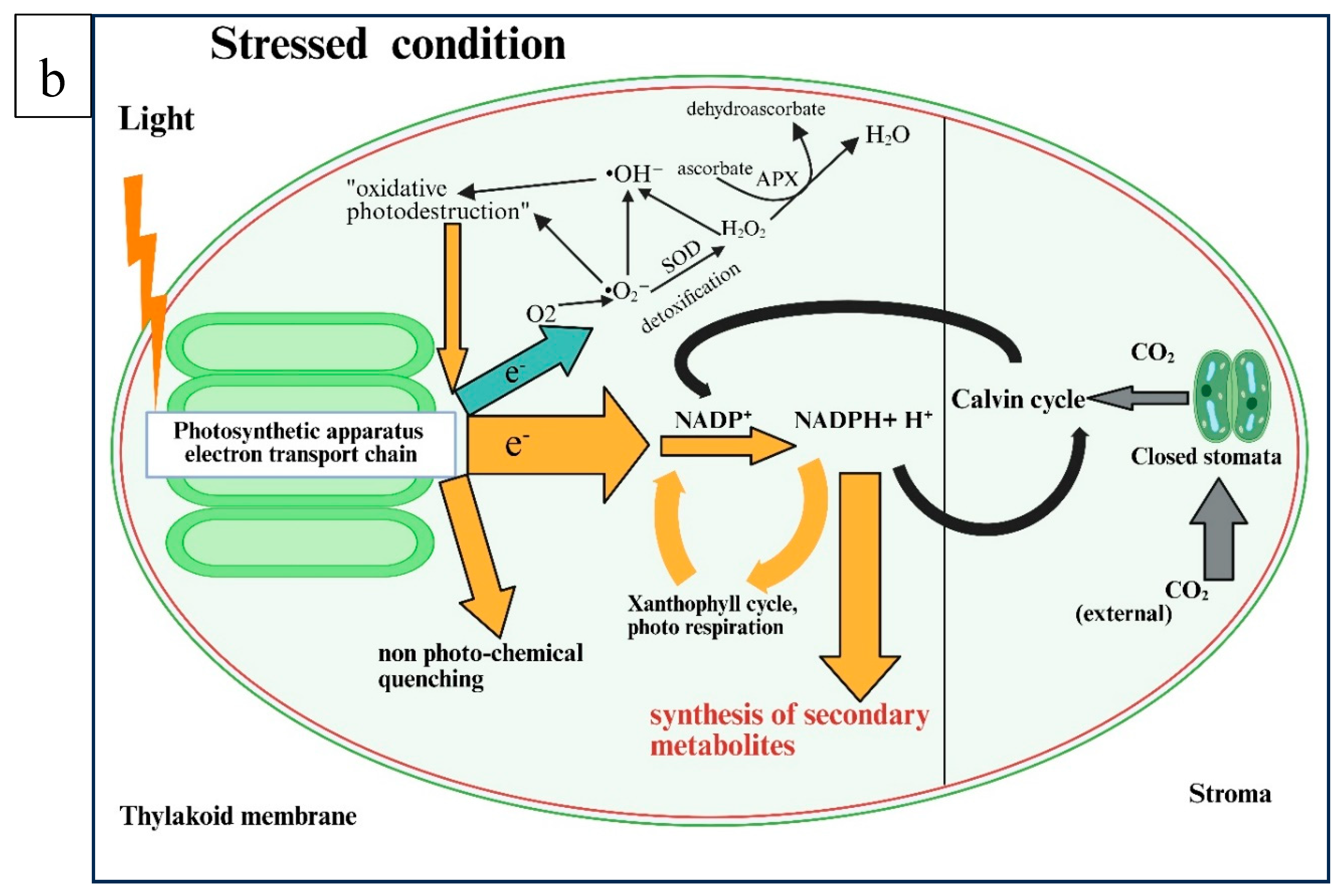
Figure 3b illustrates that the photosynthetic apparatus absorbs more light energy than is needed to fix CO2. This excess energy is dissipated through non-photochemical means such as the xanthophyll cycle and photorespiration. These processes help quench excess energy and reoxidize NADPH+H+. Under optimal water availability conditions, the excess reduction power does not produce significant amounts of radicals. However, there are noticeable changes in energy fluctuations in plants under drought-stress conditions. The higher diffusion barrier is induced by stomatal closure, which results in low endogenous CO2 levels. Consequently, in the C3 cycle, less NADPH+ H+ is used for the fixation and reduction of CO2, necessitating the release of more energy at the end [47]. The feedback system activates a protective process, including non-photochemical quenching, photorespiration, and the xanthophyll cycle to help dissipate excess energy. However, when excess energy due to limited CO2 causes a significant electron transfer to molecular oxygen. The generation of O2- ions during photosynthesis produces other ROS, like hydrogen peroxide (H2O2) and hydroxyl radicals (•OH⁻). This process, known as the Mehler reaction [48], is also called the water–water cycle [49]. The detoxification of these O2- ions is achieved through the stress-induced activation of superoxide dismutase (SOD) and ascorbate peroxidase (APX), which reduces ROS levels (Figure 3b). According to the law of mass action, plants with a higher reduction potential, indicated by a higher ratio of NADPH+ H+ to NADP+, tend to produce more secondary metabolites. However, under normal conditions, due to the co-occurrence of the various dissipation mechanisms, the balance between NADP+ and NADPH+ H+ will be achieved, and the biosynthesis of natural products is reduced (Figure 3a) [47].
Under limited water availability, Hypericum brasiliense shows a significant increase in phenolic acids [50]. Water stress also causes a notable rise in sesquiterpene (E) β-caryophyllene levels in two native Iranian subspecies of Origanum vulgare, namely gracile and virens [51]. In Labisia pumila, stress conditions increase flavonoid levels, enhancing the plant’s therapeutic properties under extreme water deficit conditions [52]. In the Lamiaceae family, essential oil content declines in Lavandula latifolia and Salvia sclarea. In contrast, the essential oil content in Mentha piperita, Salvia lavandulifolia, Thymus capitatus, and Thymus mastichina increases during drought conditions, which is attributed to a higher density of oil glands from reduced leaf area [53]. On the other hand, the content of phenolic acids simultaneously improved, while the level of flavonoids decreased in Achillea pachycephala [54]. Black cumin trials indicate drought conditions can elevate commercially valuable compounds, including essential oils [55]. Similarly, Arabidopsis plants with elevated flavonol levels due to AtMYB12 overexpression exhibit excellent resistance to salt and drought stress [56]. Additionally, different drought-tolerant plant species were found to have higher concentrations of amino acids such as glycine, asparagine, valine, isoleucine, proline, and leucine compared to drought-sensitive cultivars. Some amino acids, such as proline, glutamine, glutamate, and ornithine, function as osmolytes to enhance stress tolerance in plants like Bermuda grass (Cynodon dactylon) and Arabidopsis thaliana [57].
4. Impact of Water-deficit STRESS on Cannabis
4.1. Growth and Yield of Cannabis
Environmental stresses significantly impact the phenological stages of plants [58], and plants often respond to these stresses by accelerating flowering and shortening their growth periods [59]. Among various crops, cannabis has gained lots of attention. Several studies have highlighted its agronomic potential under such conditions, demonstrating its resistance to microbial contamination, low pesticide dependence, and ability to grow in contaminated soils, suppress pathogens, and remove heavy metals from the soil. Additionally, cannabis positively impacts crop rotation and soil quality, demonstrates resilience to water stress, thrives in diverse climatic conditions, and possesses substantial genetic diversity [60]. Cannabis, for instance, may tolerate dry conditions once established, but persistent drought conditions will induce early maturity and shorten the growth cycle [61]. According to Kumar Vijaya (2021) [62], reduced soil moisture altered cannabis phenology; specifically, the most stressed plants experienced faster flowering and seed maturation than those receiving sufficient water. Previous research has shown that plant morpho-physiological traits, such as plant height, dry weight, and lateral branches, decrease when plants experience water deficit stress [63]. One study revealed that cannabis biomass and stem yield decreased by 20%, and stem length dropped by 30% under drought stress compared to adequately hydrated plants [64]. Similarly, Arad (2016) [65] noted significant reductions in plant height, stem diameter, internode length, leaf number, and leaf area in cannabis under drought conditions. Water deficits also reduced biomass, seed yield, and stem diameter across different cannabis cultivars [66].
In recent years, especially in fiber hemp, the optimal conditions and their dependence on environmental factors such as irradiation, photoperiod, temperature, relative humidity, production techniques like plant density, mineral nutrition, irrigation regime, and genetic factors have been intensively investigated [67]. Studies on hemp have shown variable effects on yield under drought stress, with some cultivar strains remaining unaffected while others experience significant yield losses [68]. In a fully irrigated condition where hemp plants were watered at 100% evapotranspiration, significantly higher yields were observed relative to trials with reduced irrigation at 75% of the calculated evapotranspiration [69]. The final yields of well-watered controls were 43-64% higher than those under drought treatments [69].
Previous research on hemp has examined the effects of water-deficit stress starting in the vegetative stage. Garcia-Tejero et al. (2014) [69] highlighted how consistent water-deficit stress, initiated early in the vegetative stage and continuing through harvest, adversely affected yield and physiological parameters such as photosynthesis, stomatal conductance, and carbon isotope discrimination. García-Caparrós et al. (2019) [53] reported a significant reduction in plant height and the length of the first internode in cannabis under water-deficit stress. Consistent with these findings, Bahador & Tadayon (2020) [70] observed that cannabis plants subjected to drought stress were shorter in height over a two-year study period. The study attributed the decreased stem diameter to reduced lateral meristem activity, cell proliferation/expansion, and a reduction in epidermal and vascular tissue thickness [71]. Recent research on various cannabis genotypes also confirmed that water deficits lead to decreases in plant height, stem diameter, and internode length [66]. Additional studies have shown that the cannabis cultivars Futura 75 and Black Label experienced reduced growth and yield when the available water in the root zone was limited [64,72]. Conversely, Caplan et al. (2019) [73] found that applying water-deficit stress to medicinal cannabis in the final two weeks of the growing cycle did not adversely affect flower yield. These findings highlight the complex responses of cannabis to water-deficit stress, which can vary depending on the stress's timing, duration, and severity, as well as the specific cultivar, production purpose, and growing conditions.
4.2. Production of Secondary Metabolites of Cannabis
The biological functions of glandular trichomes and their resinous secretion are primarily linked to defense mechanisms against various biotic and abiotic stresses imposed by the environment and herbivores [74]. Therefore, there have been numerous attempts to manage resin accumulation in medical cannabis using stress induction [75]. The production of secondary metabolites in cannabis is influenced by external factors, including light duration, oxygen levels, and harvest timing, particularly during floral maturation [76]. Research has established a connection between water-deficit stress and cannabinoid synthesis in cannabis, as discussed below.
According to Selmar & Kleinwachter (2013) [46], plants experience stomatal closure under drought conditions to conserve water loss, significantly reducing CO2 uptake. In return, it limits the CO2 activity of the Calvin cycle, which means less NADPH+H+ and ATP formed during light-dependent photosynthesis reactions in the thylakoid membranes used for photosynthetic carbon fixation. Ultimately, large amounts of energy must be dissipated through non-photochemical quenching and effective re-oxidation of NADP+ H+ through the xanthophyll cycle and photorespiration (Figure 3b). However, the excess NADPH+H+ generated during light reactions causes excess electrons not utilized by NADP+ to be transferred to molecular oxygen, forming highly damaging ROS (Figure 3b). Kleinwächter & Selmar (2014) [47] suggested that plants activate protective mechanisms to dissipate surplus energy to avoid ROS damage and harmful effects. The excess NADPH+H+ generated during light reactions has been linked to increased levels of THC, THCa (tetrahydrocannabinolic acid), CBDa (cannabidiolic acid), and CBD [73], suggesting that unused NADPH+H+ might contribute to altered secondary metabolite profiles in cannabis.
It has been demonstrated that the production of cannabinoids, such as THC, CBD, and cannabigerol (CBG), is influenced by simple genetic traits. However, the variability in cannabinoid content is significantly influenced by environmental conditions, including drought stress [77]. Ecological studies have revealed that cannabis plants develop a higher trichome density in arid climates [78]. Additionally, decreased humidity has also been associated with increased THC content in cannabis [79]. Similar research findings have been reported in non-THC-producing hemp plants, which demonstrated significant cannabinoid production when cultivated in dry regions [80,81]. However, these findings are not entirely conclusive, and further investigation is needed to fully understand how water stress comprehensively affects cannabinoid production.
Minimizing dry weight loss during drought stress is critical for maximizing the production of essential oils and secondary metabolites in Cannabis sativa [82]. This species exhibit notable physiological adaptability, adjusting their production of secondary metabolites in response to varying levels of dry conditions. Previous studies have suggested that cannabis cultivated in hotter, drier environments tends to display higher densities of trichomes [78]. Recent research has shown a positive correlation between hemp's accumulation of CBD and its ability to tolerate drought stress in both field and controlled environment experiments [83]. Studies on the effect of drought stress have been documented to increase secondary metabolite accumulation in other crops like Satureja hortensis (summer savory) [84]. This indicates a general trend in increased secondary metabolite concentrations under drought stress, which may also apply to Cannabis sativa.
Caplan et al. (2019) [73] observed that when cannabis plants were subjected to mild drought stress, it significantly increased the levels of tetrahydrocannabinol acid (THCa), cannabidiolic acid (CBDa), THC and CBD without reducing inflorescence dry weight. Similarly, recent research on four medical cannabis varieties has shown that exposure to a wide range of biotic stresses (such as Golovinomyces spadiceus and Manduca sexta) and abiotic stresses (herbicide treatment, flooding, wound injury, heat, and drought) significantly influences cannabinoid production. The floral parts produced at least 2.5 times greater cannabinoids than the leaf tissues under stress conditions [85,86].
However, according to Park et al. (2022) [85], cannabinoid production at the onset of flowering was found to be negatively correlated with herbivory, extreme heat, and drought, whereas mechanical wounding showed no impact. Applying induced water-deficit stress during the early stages of flowering has been observed to significantly alter cannabinoid profiles, with significant increases in CBG alongside reductions of up to 70-80% in THC and CBD accumulation [85]. This suggests that applications of water-deficit stress at the early stages of flowering are likely to disrupt the conversion of CBGa (cannabigerolic acid) into its downstream products, CBDa and THCa; instead, it raises CBG concentration. The catalytic enzymes, CBDa and THCa synthases responsible for the conversion may have reduced activities, or their gene expression may have been downregulated, leading to decreased downstream THC and CBD levels, while CBG levels accumulate [85].
Morgan (2023) [87] has found that moderate water-deficit stress and drought intensities did not affect the levels of THC or CBD. However, severe drought treatments reduced the concentrations of these cannabinoids due to impaired photosynthesis and carbon fixation. In contrast, the “Nebula” cultivar has shown enhanced production of primary cannabinoids THCa/THC and CBDa/CBD following water-deficit stress initiation, particularly about 5-7 weeks after flowering began [73]. The observed variation in THC and CBD wasnot only related to water-deficit stress, but also related to the specific cultivar, water-deficit stress timing, and the original cannabinoid content. However, the limited research on cannabis cultivation practices has led to gaps in knowledge related to water usage, and yield potential [88,89].
5. Conclusions
Water-deficit stress is usually recognized as a negative factor that reduces plant height, stem diameter, and internode length in crops, ultimately leading to significant yield losses in agriculture. Similar detrimental effects have been seen in different aromatic and medicinal plants, including Cannabis sativa. However, recent research reveals a direct relationship between water-deficit stress and plant metabolism. Water-deficit stress, induced directly or indirectly by water shortage, has been shown to enhance the accumulation of secondary metabolites in cannabis, especially the cannabinoids. However, the impact of water-deficit stress on cannabis plants can vary based on the cultivars, intensity of the stress, and timing of the stress events, with some studies indicating substantial losses. In contrast, others suggest that late-stage drought stress might not significantly affect flower yield. This variability highlights the need to understand the specific requirements and stress responses of different cannabis cultivar strains to optimize growth and productivity under different growing conditions. Further research is essential to unravel how drought stress can be strategically managed to maximize biomass yield and secondary metabolite production in cannabis under various environmental conditions.
Author Contributions
Writing-original draft preparation, S.S.; writing, data curation, T.A.; data curation, figure preparation, R.S.R.; methodology, review, A.M.A.; methodology, review, J.W; conceptualization, review, A.A.D; conceptualization, methodology, resources, writing, review, editing, J.F.D.C.L.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The original contributions presented in the study are included in the article; further inquiries can be directed to the corresponding authors.
Conflicts of Interest
The authors declare no conflict of interest.
References
- McPartland, J. M.; Hegman, W.; Long, T. Cannabis in Asia: its center of origin and early cultivation, based on a synthesis of subfossil pollen and archaeobotanical studies. Vegetation History and Archaeobotany 2019, 28(6), 691–702. [Google Scholar] [CrossRef]
- Li, H.-L. An archaeological and historical account of cannabis in China. Economic botany 1974, 28(4), 437–448. [Google Scholar] [CrossRef]
- Small, E. Evolution and Classification of Cannabis sativa (Marijuana, Hemp) in Relation to Human Utilization. The Botanical Review 2015, 81(3), 189–294. [Google Scholar] [CrossRef]
- Clarke, R.; Merlin, M. Cannabis: evolution and ethnobotany; Univ of California Press, 2016.
- Small, E.; Cronquist, A. A practical and natural taxonomy for Cannabis. Taxon 1976, 405–435. [Google Scholar] [CrossRef]
- Schilling, S.; Melzer, R.; McCabe, P. F. Cannabis sativa . Curr Biol 2020, 30(1), R8–R9. [Google Scholar] [CrossRef]
- Hudak, J. Marijuana: A short history; Brookings Institution Press, 2020.
- Skorbiansky, S. R.; Thornsbury, S.; Camp, K. M. Legal Risk Exposure Heightens Uncertainty in Developing US Hemp Markets. Choices 2021, 36(1), 1–10. [Google Scholar]
- Anderson, E. Plants, life, and man. Andrew Melrose, London 1954, 120-133.
- Bonini, S. A.; Premoli, M.; Tambaro, S.; Kumar, A.; Maccarinelli, G.; Memo, M.; Mastinu, A. Cannabis sativa: A comprehensive ethnopharmacological review of a medicinal plant with a long history. J Ethnopharmacol 2018, 227, 300–315. [Google Scholar] [CrossRef]
- Andre, C. M.; Hausman, J. F.; Guerriero, G. Cannabis sativa: The Plant of the Thousand and One Molecules. Front Plant Sci 2016, 7, 19. [Google Scholar] [CrossRef]
- Pacher, P.; Kogan, N. M.; Mechoulam, R. Beyond THC and Endocannabinoids. Annu Rev Pharmacol Toxicol 2020, 60, 637–659. [Google Scholar] [CrossRef]
- Schlosser, E. Reefer madness: Sex, drugs, and cheap labor in the American black market; HMH, 2004.
- Ramakrishna, A.; Ravishankar, G. A. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal Behav 2011, 6(11), 1720–1731, From NLM Medline. [Google Scholar] [CrossRef]
- Montoya, Z.; Conroy, M.; Vanden Heuvel, B. D.; Pauli, C. S.; Park, S. H. Cannabis Contaminants Limit Pharmacological Use of Cannabidiol. Front Pharmacol 2020, 11, 571832. [Google Scholar] [CrossRef]
- Gorelick, J.; Bernstein, N. Chemical and Physical Elicitation for Enhanced Cannabinoid Production in Cannabis. In Cannabis sativa L. - Botany and Biotechnology, 2017; pp 439-456.
- Eichhorn Bilodeau, S.; Wu, B. S.; Rufyikiri, A. S.; MacPherson, S.; Lefsrud, M. An Update on Plant Photobiology and Implications for Cannabis Production. Front Plant Sci 2019, 10, 296. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, N.; Gorelick, J.; Koch, S. Interplay between chemistry and morphology in medical cannabis (Cannabis sativa L.). Industrial Crops and Products 2019, 129, 185–194. [Google Scholar] [CrossRef]
- Husain, R.; Weeden, H.; Bogush, D.; Deguchi, M.; Soliman, M.; Potlakayala, S.; Katam, R.; Goldman, S.; Rudrabhatla, S. Enhanced tolerance of industrial hemp (Cannabis sativa L.) plants on abandoned mine land soil leads to overexpression of cannabinoids. PLoS One 2019, 14(8), e0221570. [Google Scholar] [CrossRef] [PubMed]
- Burgel, L.; Hartung, J.; Schibano, D.; Graeff-Honninger, S. Impact of Different Phytohormones on Morphology, Yield and Cannabinoid Content of Cannabis sativa L. Plants (Basel) 2020, 9. [Google Scholar] [CrossRef]
- Verma, K.; Pratibha; Priya Soni, R.; Kumar, P.; Chauhan, D.; Augustine, A. A. Drought; Influence of Drought in Agriculture and Management Strategies for Drought-a Review. Asian Journal of Microbiology, Biotechnology & Environmental Sciences 2023, 25(04), 638–642. [CrossRef]
- He, M.; He, C. Q.; Ding, N. Z. Abiotic Stresses: General Defenses of Land Plants and Chances for Engineering Multistress Tolerance. Front Plant Sci 2018, 9, 1771. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Rivero, R. M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and biotic stress combinations. New Phytol 2014, 203(1), 32–43. [Google Scholar] [CrossRef]
- Kapoor, D.; Bhardwaj, S.; Landi, M.; Sharma, A.; Ramakrishnan, M.; Sharma, A. The Impact of Drought in Plant Metabolism: How to Exploit Tolerance Mechanisms to Increase Crop Production. Applied Sciences 2020, 10(16). [Google Scholar] [CrossRef]
- Godoy, F.; Olivos-Hernandez, K.; Stange, C.; Handford, M. Abiotic Stress in Crop Species: Improving Tolerance by Applying Plant Metabolites. Plants (Basel) 2021, 10(2). [Google Scholar] [CrossRef]
- Giordano, M.; Petropoulos, S. A.; Rouphael, Y. Response and Defence Mechanisms of Vegetable Crops against Drought, Heat and Salinity Stress. Agriculture 2021, 11(5). [Google Scholar] [CrossRef]
- Xu, C.; He, C. G.; Wang, Y. J.; Bi, Y. F.; Jiang, H. Effect of drought and heat stresses on photosynthesis, pigments, and xanthophyll cycle in alfalfa (Medicago sativa L.). Photosynthetica 2020, 58(5), 1226–1236. [Google Scholar] [CrossRef]
- Morales, F.; Ancin, M.; Fakhet, D.; Gonzalez-Torralba, J.; Gamez, A. L.; Seminario, A.; Soba, D.; Ben Mariem, S.; Garriga, M.; Aranjuelo, I. Photosynthetic Metabolism under Stressful Growth Conditions as a Bases for Crop Breeding and Yield Improvement. Plants (Basel) 2020, 9(1). [Google Scholar] [CrossRef] [PubMed]
- Emami Bistgani, Z.; Barker, A. V.; Hashemi, M. Physiology of medicinal and aromatic plants under drought stress. The Crop Journal 2024. [Google Scholar] [CrossRef]
- M. C. DIAS, W. B. Limitations of photosynthesis in Phaseolus vulgaris under drought stress: gas exchange, chlorophyll fluorescence and Calvin cycle enzymes. PHOTOSYNTHETICA 2010, 48(1), 96–102. [Google Scholar] [CrossRef]
- Huang, L.; Li, M.; Zhou, K.; Sun, T.; Hu, L.; Li, C.; Ma, F. Uptake and metabolism of ammonium and nitrate in response to drought stress in Malus prunifolia. Plant Physiol Biochem 2018, 127, 185–193. [Google Scholar] [CrossRef]
- Filippou, P.; Antoniou, C.; Fotopoulos, V. Effect of drought and rewatering on the cellular status and antioxidant response of Medicago truncatula plants. Plant Signal Behav 2011, 6(2), 270–277. [Google Scholar] [CrossRef] [PubMed]
- Keipp, K.; Hütsch, B. W.; Ehlers, K.; Schubert, S. Drought stress in sunflower causes inhibition of seed filling due to reduced cell-extension growth. Journal of Agronomy and Crop Science 2020, 206(5), 517–528. [Google Scholar] [CrossRef]
- Coussement, J. R.; Villers, S. L. Y.; Nelissen, H.; Inze, D.; Steppe, K. Turgor-time controls grass leaf elongation rate and duration under drought stress. Plant Cell Environ 2021, 44(5), 1361–1378. [Google Scholar] [CrossRef]
- Sarani Mahnaz, H. S. M. , Mahdi M.R. The effect of drought stress on chlorophyll content, root growth, glucosinolate and proline in crop plants. International Journal of Farming and Allied Sciences 2014, 3(9), 994–997. [Google Scholar]
- Lebaschy, M. H.; Sharifi ashoor abadi, E. Growth indices of some medicinal plants under different water stresses. Iranian Journal of Medicinal and Aromatic Plants Research 2004, 20(3), 249–261. [Google Scholar]
- Bazzazi, N.; Khodambashi, M.; Mohammadi, S. The effect of drought stress on morphological characteristics and yield components of medicinal plant fenugreek. Isfahan University of Technology-Journal of Crop Production and Processing 2013, 3(8), 11–23. [Google Scholar]
- Kumar, A.; Omae, H.; Egawa, Y.; Kashiwaba, K.; Shono, M. Adaptation to heat and drought stresses in snap bean (Phaseolus vulgaris) during the reproductive stage of development. Japan Agricultural Research Quarterly: JARQ 2006, 40(3), 213–216. [Google Scholar] [CrossRef]
- Asadi, S.; Moghaddam, H.; Naghdi Badi, H.; Naghavi, M. R.; Salami, S. A.; Solaiman, Z. Agronomic, phytochemical and drought tolerance evaluation of Iranian cannabis (Cannabis sativa L.) ecotypes under different soil moisture levels: a step towards identifying pharmaceutical and industrial populations. Crop & Pasture Science 2023, 74(12), 1238–1257. [Google Scholar] [CrossRef]
- Zhang, S.-h.; Xu, X.-f.; Sun, Y.-m.; Zhang, J.-l.; Li, C.-z. Influence of drought hardening on the resistance physiology of potato seedlings under drought stress. Journal of Integrative Agriculture 2018, 17(2), 336–347. [Google Scholar] [CrossRef]
- Wang, L.; Liu, L.; Ma, Y.; Li, S.; Dong, S.; Zu, W. Transcriptome profilling analysis characterized the gene expression patterns responded to combined drought and heat stresses in soybean. Computational Biology and Chemistry 2018, 77, 413–429. [Google Scholar] [CrossRef]
- Leverne, L.; Krieger-Liszkay, A. Moderate drought stress stabilizes the primary quinone acceptor Q(A) and the secondary quinone acceptor Q(B) in photosystem II. Physiol Plant 2021, 171(2), 260–267. [Google Scholar] [CrossRef]
- Pospisil, P. Production of reactive oxygen species by photosystem II. Biochim Biophys Acta 2009, 1787(10), 1151–1160. [Google Scholar] [CrossRef]
- Berni, R.; Luyckx, M.; Xu, X.; Legay, S.; Sergeant, K.; Hausman, J.-F.; Lutts, S.; Cai, G.; Guerriero, G. Reactive oxygen species and heavy metal stress in plants: Impact on the cell wall and secondary metabolism. Environmental and Experimental Botany 2019, 161, 98–106. [Google Scholar] [CrossRef]
- Khodabin, G.; Lightburn, K.; Hashemi, S. M.; Moghada, M. S. K.; Jalilian, A. Evaluation of nitrate leaching, fatty acids, physiological traits and yield of rapeseed (Brassica napus) in response to tillage, irrigation and fertilizer management. Plant and Soil 2022, 473((1-2)), 423–440. [Google Scholar] [CrossRef]
- Selmar, D.; Kleinwachter, M. Stress enhances the synthesis of secondary plant products: the impact of stress-related over-reduction on the accumulation of natural products. Plant Cell Physiol 2013, 54(6), 817–826. [Google Scholar] [CrossRef] [PubMed]
- Kleinwächter, M.; Selmar, D. New insights explain that drought stress enhances the quality of spice and medicinal plants: potential applications. Agronomy for Sustainable Development 2014, 35(1), 121–131. [Google Scholar] [CrossRef]
- Hideg, É.; Spetea, C.; Vass, I. Superoxide radicals are not the main promoters of acceptor-side-induced photoinhibitory damage in spinach thylakoids. Photosynthesis Research 1995, 46, 399–407. [Google Scholar] [CrossRef]
- Hormann, H.; Neubauer, C.; Asada, K.; Schreiber, U. Intact chloroplasts display pH 5 optimum of O 2-reduction in the absence of methyl viologen: Indirect evidence for a regulatory role of superoxide protonation. Photosynthesis research 1993, 37, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Nacif de Abreu, I.; Mazzafera, P. Effect of water and temperature stress on the content of active constituents of Hypericum brasiliense Choisy. Plant Physiol Biochem 2005, 43(3), 241–248. [Google Scholar] [CrossRef]
- Morshedloo, M. R.; Craker, L. E.; Salami, A.; Nazeri, V.; Sang, H.; Maggi, F. Effect of prolonged water stress on essential oil content, compositions and gene expression patterns of mono- and sesquiterpene synthesis in two oregano (Origanum vulgare L.) subspecies. Plant Physiol Biochem 2017, 111, 119–128. [Google Scholar] [CrossRef]
- Jaafar, H. Z.; Ibrahim, M. H.; Mohamad Fakri, N. F. Impact of soil field water capacity on secondary metabolites, phenylalanine ammonia-lyase (PAL), maliondialdehyde (MDA) and photosynthetic responses of Malaysian kacip fatimah (Labisia pumila Benth). Molecules 2012, 17(6), 7305–7322. [Google Scholar] [CrossRef]
- García-Caparrós, P.; Romero, M.; Llanderal, A.; Cermeño, P.; Lao, M.; Segura, M. Effects of Drought Stress on Biomass, Essential Oil Content, Nutritional Parameters, and Costs of Production in Six Lamiaceae Species. Water 2019, 11(3). [Google Scholar] [CrossRef]
- Gharibi, S.; Sayed Tabatabaei, B. E.; Saeidi, G.; Talebi, M.; Matkowski, A. The effect of drought stress on polyphenolic compounds and expression of flavonoid biosynthesis related genes in Achillea pachycephala Rech.f. Phytochemistry 2019, 162, 90–98. [Google Scholar] [CrossRef]
- Bayati, P.; Karimmojeni, H.; Razmjoo, J. Changes in essential oil yield and fatty acid contents in black cumin (Nigella sativa L.) genotypes in response to drought stress. Industrial Crops and Products 2020, 155. [Google Scholar] [CrossRef]
- Wang, F.; Kong, W.; Wong, G.; Fu, L.; Peng, R.; Li, Z.; Yao, Q. AtMYB12 regulates flavonoids accumulation and abiotic stress tolerance in transgenic Arabidopsis thaliana. Mol Genet Genomics 2016, 291(4), 1545–1559. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Chan, Z. Improvement of plant abiotic stress tolerance through modulation of the polyamine pathway. J Integr Plant Biol 2014, 56(2), 114–121. [Google Scholar] [CrossRef]
- Da Cunha Leme Filho, J. F.; Chim, B. K.; Bermand, C.; Diatta, A. A.; Thomason, W. E. Effect of organic biostimulants on cannabis productivity and soil microbial activity under outdoor conditions. J Cannabis Res 2024, 6(1), 16. [Google Scholar] [CrossRef]
- Chaves Manuela M., M. J. P. P. J. Understanding plant responses to drought — from genes to the whole plant. Functional Plant Biology 2003, 30, 239–264. [Google Scholar] [CrossRef] [PubMed]
- Amaducci, S.; Zatta, A.; Pelatti, F.; Venturi, G. Influence of agronomic factors on yield and quality of hemp (Cannabis sativa L.) fibre and implication for an innovative production system. Field Crops Research 2008, 107(2), 161–169. [Google Scholar] [CrossRef]
- Adesina, I.; Bhowmik, A.; Sharma, H.; Shahbazi, A. A Review on the Current State of Knowledge of Growing Conditions, Agronomic Soil Health Practices and Utilities of Hemp in the United States. Agriculture 2020, 10(4). [Google Scholar] [CrossRef]
- Kumar Vijaya, I. M. K. Production and Quality of Industrial Hemp (Cannabis sativa L.) in Response to Water Regimes. Doctoral dissertation, University Of Tasmania 2021.
- Petropoulos, S. A.; Daferera, D.; Polissiou, M. G.; Passam, H. C. The effect of water deficit stress on the growth, yield and composition of essential oils of parsley. Scientia Horticulturae 2008, 115(4), 393–397. [Google Scholar] [CrossRef]
- Cosentino, S. L.; Riggi, E.; Testa, G.; Scordia, D.; Copani, V. Evaluation of European developed fibre hemp genotypes (Cannabis sativa L.) in semi-arid Mediterranean environment. Industrial Crops and Products 2013, 50, 312–324. [Google Scholar] [CrossRef]
- Arad, N. Effect of drought stress on relative expression of some key genes involved in cannabisis in medicinal cannabis. Master’s thesis, University of Tehran, Karaj, Iran.[In Persian], 2016.
- Campbell, B. J.; Berrada, A. F.; Hudalla, C.; Amaducci, S.; McKay, J. K. Genotype × Environment Interactions of Industrial Hemp Cultivars Highlight Diverse Responses to Environmental Factors. Agrosystems, Geosciences & Environment 2019, 2(1), 1–11. [Google Scholar] [CrossRef]
- Herppich, W. B.; Gusovius, H.-J.; Flemming, I.; Drastig, K. Effects of Drought and Heat on Photosynthetic Performance, Water Use and Yield of Two Selected Fiber Hemp Cultivars at a Poor-Soil Site in Brandenburg (Germany). Agronomy 2020, 10(9). [Google Scholar] [CrossRef]
- Babaei, M.; Ajdanian, L. Screening of different Iranian ecotypes of cannabis under water deficit stress. Scientia Horticulturae 2020, 260. [Google Scholar] [CrossRef]
- Garcia Tejero, I.; Duran Zuazo, V.; Pérez-Álvarez, R.; Hernández, A.; Casano, S.; Morón, M.; Muriel-Fernández, J. Impact of plant density and irrigation on yield of hemp (Cannabis sativa L.) in a Mediterranean semi-arid environment. Journal of Agricultural Science and Technology 2014, 16(4), 887–895. [Google Scholar]
- Bahador, M.; Tadayon, M. R. Investigating of zeolite role in modifying the effect of drought stress in hemp: Antioxidant enzymes and oil content. Industrial crops and products 2020, 144, 112042. [Google Scholar] [CrossRef]
- Selim, A.-F. H.; El-Nady, M. F. Physio-anatomical responses of drought stressed tomato plants to magnetic field. Acta Astronautica 2011, 69(7-8), 387–396. [Google Scholar] [CrossRef]
- Gill, A. R.; Loveys, B. R.; Cowley, J. M.; Hall, T.; Cavagnaro, T. R.; Burton, R. A. Physiological and morphological responses of industrial hemp (Cannabis sativa L.) to water deficit. Industrial Crops and Products 2022, 187, 115331. [Google Scholar] [CrossRef]
- Caplan, D.; Dixon, M.; Zheng, Y. Increasing Inflorescence Dry Weight and Cannabinoid Content in Medical Cannabis Using Controlled Drought Stress. HortScience 2019, 54(5), 964–969. [Google Scholar] [CrossRef]
- Tanney, C. A. S.; Backer, R.; Geitmann, A.; Smith, D. L. Cannabis Glandular Trichomes: A Cellular Metabolite Factory. Front Plant Sci 2021, 12, 721986. [Google Scholar] [CrossRef] [PubMed]
- Chandra, S.; Lata, H.; ElSohly, M. A.; Walker, L. A.; Potter, D. Cannabis cultivation: Methodological issues for obtaining medical-grade product. Epilepsy Behav 2017, 70 (Pt B), 302–312. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Fernando, D.; Daniel, G.; Madsen, B.; Meyer, A. S.; Ale, M. T.; Thygesen, A. Effect of harvest time and field retting duration on the chemical composition, morphology and mechanical properties of hemp fibers. Industrial Crops and Products 2015, 69, 29–39. [Google Scholar] [CrossRef]
- De Meijer, E. P.; Bagatta, M.; Carboni, A.; Crucitti, P.; Moliterni, V. C.; Ranalli, P.; Mandolino, G. The inheritance of chemical phenotype in Cannabis sativa L. Genetics 2003, 163(1), 335–346. [Google Scholar] [CrossRef]
- G. Sharma, "Altidunal variation in leaf epidermal patterns of Cannabis sativa,". Bulletin of the Torrey Botanical Club 1975, 102(4), 199–200. [Google Scholar] [CrossRef]
- Paris M, B. F. , Cosson L. The Constituents of Cannabis sativa Pollen. Economic Botany 1975, 29(3), 245–253. [Google Scholar] [CrossRef]
- H. Hakim, Y. E. Kheir, and M. Mohamed, "Effect of the climate on the content of a CBD-rich variant of cannabis,". Fitoterapia 1987, 57 (4), 239-241.
- Murari, G.; Lombardi, S.; Puccini, A.; Sanctis, R. d. Influence of environmental conditions on tetrahydrocannabinol (Δ9-TCH) in different cultivars of Cannabis sativa L. Fitoterapia 1984, 5, 195–201. [Google Scholar]
- Nakawuka, P.; Peters, T. R.; Gallardo, K. R.; Toro-Gonzalez, D.; Okwany, R. O.; Walsh, D. B. Effect of Deficit Irrigation on Yield, Quality, and Costs of the Production of Native Spearmint. Journal of Irrigation and Drainage Engineering 2014, 140. [Google Scholar] [CrossRef]
- Sheldon, K.; Shekoofa, A.; Walker, E.; Kelly, H. Physiological screening for drought-tolerance traits among hemp (Cannabis sativa L.) cultivars in controlled environments and in field. Journal of Crop Improvement 2021, 35(6), 816–831. [Google Scholar] [CrossRef]
- Baher, Z. F.; Mirza, M.; Ghorbanli, M.; Bagher Rezaii, M. The influence of water stress on plant height, herbal and essential oil yield and composition in Satureja hortensis L. Flavour and Fragrance Journal 2002, 17(4), 275–277. [Google Scholar] [CrossRef]
- Park, S. H.; Pauli, C. S.; Gostin, E. L.; Staples, S. K.; Seifried, D.; Kinney, C.; Vanden Heuvel, B. D. Effects of short-term environmental stresses on the onset of cannabinoid production in young immature flowers of industrial hemp (Cannabis sativa L.). J Cannabis Res 2022, 4(1), 1. [Google Scholar] [CrossRef]
- Toth, J. A.; Smart, L. B.; Smart, C. D.; Stack, G. M.; Carlson, C. H.; Philippe, G.; Rose, J. K. C. Limited effect of environmental stress on cannabinoid profiles in high-cannabidiol hemp (Cannabis sativa L.). GCB Bioenergy 2021, 13(10), 1666–1674. [Google Scholar] [CrossRef]
- Morgan, G. W. Effects of drought stress on floral hemp (Cannabis sativa L.) agricultural systems. Auburn University, 2023.
- V. Butsic and J. C. Brenner, "Cannabis ( Cannabis sativa or C. indica) agriculture and the environment: a systematic, spatially-explicit survey and potential impacts," Environmental Research Letters 2016, vol. 11, no. 4. [CrossRef]
- J. F. Da Cunha Leme Filho et al., "Biochemical and physiological responses of Cannabis sativa to an integrated plant nutrition system," Agronomy Journal, 2020, vol. 112, no. 6, pp. 5237-5248. [CrossRef]
Figure 1.
An overview of the physiological process that causes growth reduction in Cannabis sativa under water-deficit stress. The figure was created using Biorender (https://BioRender.com, accessed 6 April 2025).
Figure 1.
An overview of the physiological process that causes growth reduction in Cannabis sativa under water-deficit stress. The figure was created using Biorender (https://BioRender.com, accessed 6 April 2025).
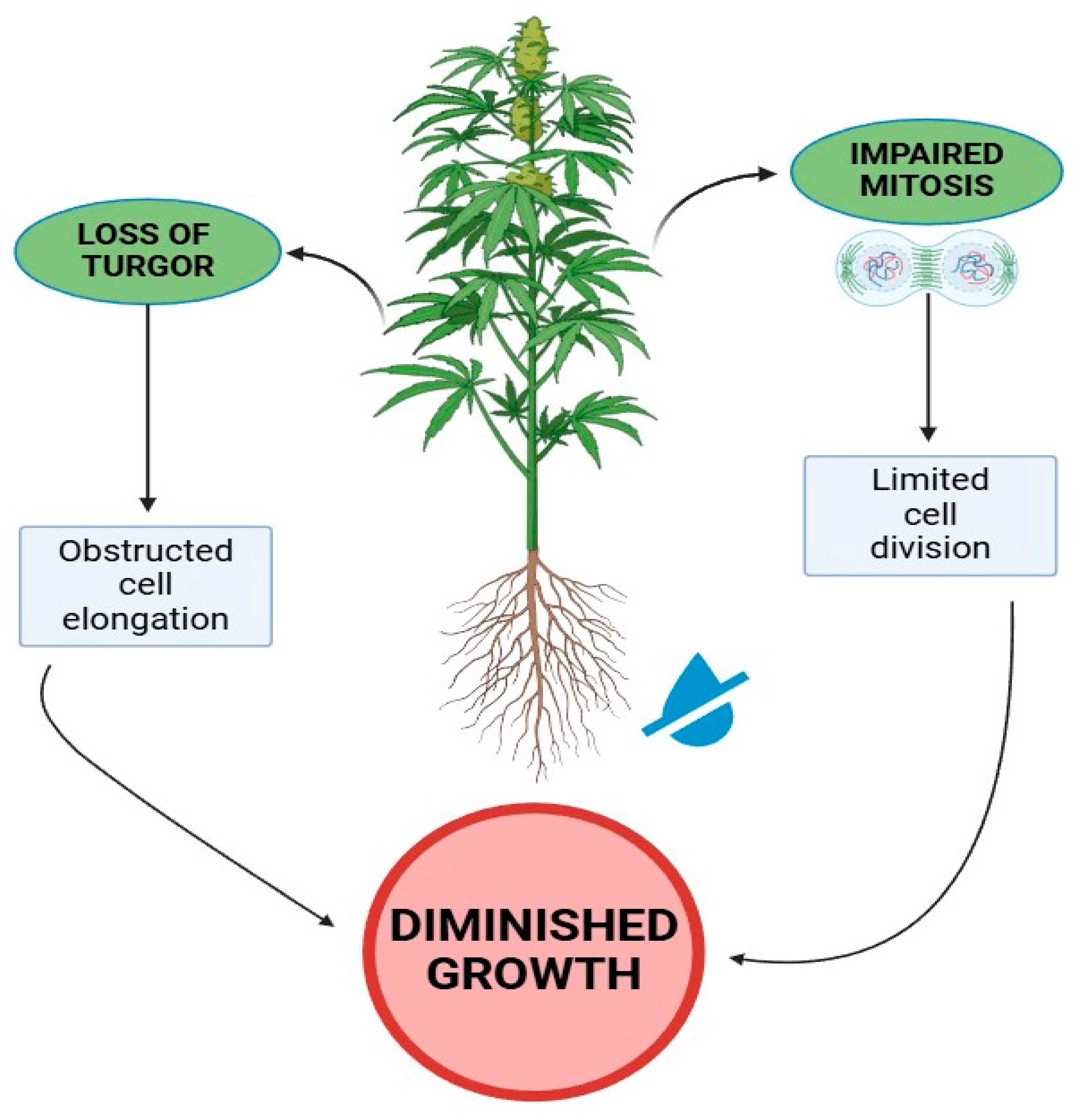
Figure 2.
Schematics illustrate the role of secondary metabolites (SMs) in response to abiotic stress, including water-deficit stress. Abiotic stress induces SM production, activating metabolic pathways that adjust plant physiology to mitigate the effects of stress. The figure was created using Biorender (https://BioRender.com/, accessed 6 April 2025).
Figure 2.
Schematics illustrate the role of secondary metabolites (SMs) in response to abiotic stress, including water-deficit stress. Abiotic stress induces SM production, activating metabolic pathways that adjust plant physiology to mitigate the effects of stress. The figure was created using Biorender (https://BioRender.com/, accessed 6 April 2025).
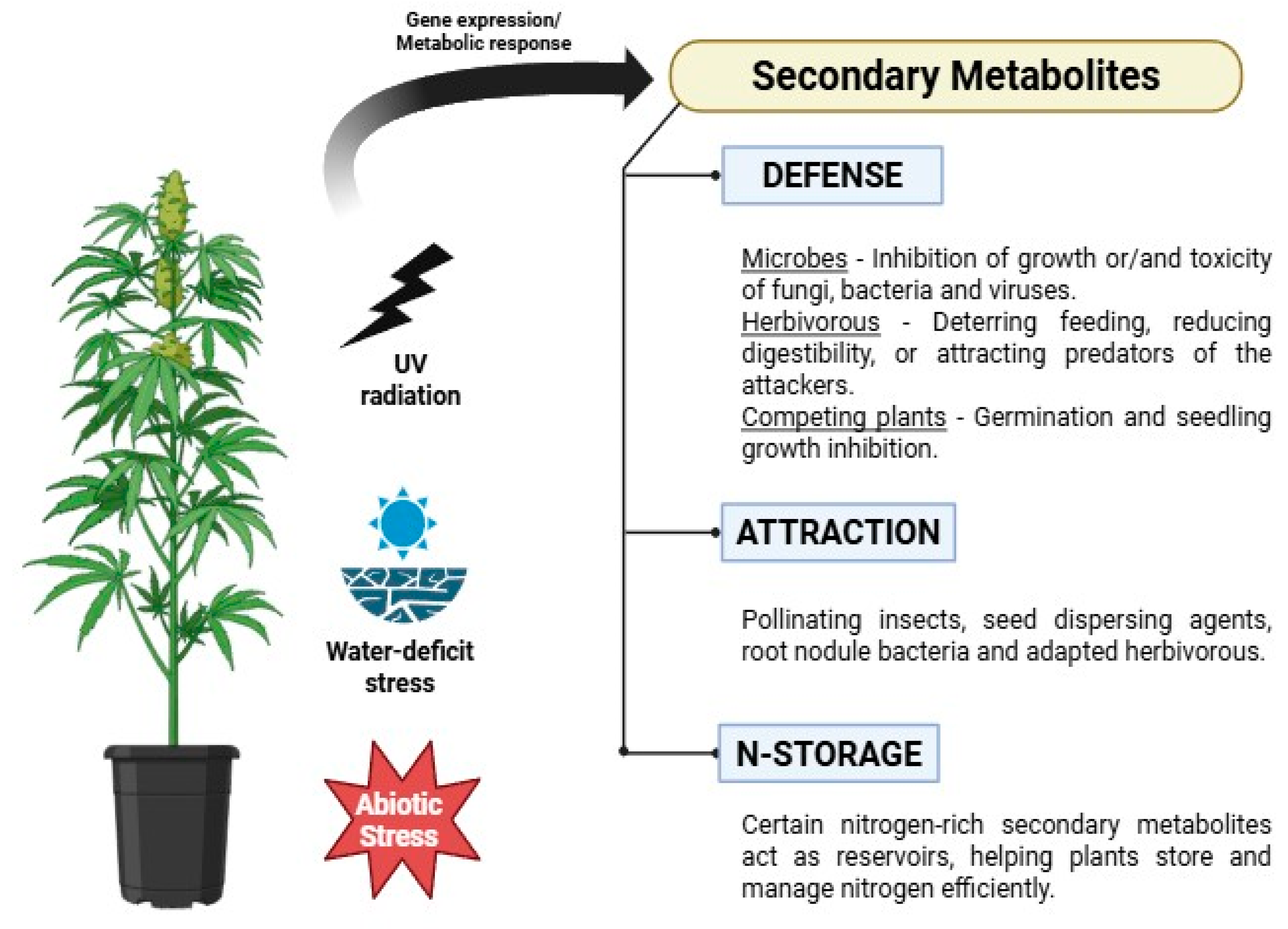
Figure 3.
An illustration of the pathways involved in the biosynthesis of secondary metabolites under water-deficit stress (a) under normal conditions and (b) under stressed conditions. The figure was created using Biorender (https://BioRender.com, accessed 10 April 2025).
Figure 3.
An illustration of the pathways involved in the biosynthesis of secondary metabolites under water-deficit stress (a) under normal conditions and (b) under stressed conditions. The figure was created using Biorender (https://BioRender.com, accessed 10 April 2025).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



