Submitted:
21 April 2025
Posted:
22 April 2025
You are already at the latest version
Abstract
Interspecies transmission of avian influenza viruses remains a significant public health concern. H6 viruses have gained attention following the first human infection by a chicken-origin H6N1 virus (A/Taiwan/02/2013, Hu/13), which highlighted their zoonotic potential. To understand the evolutionary trajectory and mammalian adaptation of this Taiwan lineage, we compared two avian isolates (A/Chicken/Taiwan/CF19/2009, Ck/09; A/Chicken/Taiwan/2267/2012, Ck/12) and Hu/13 in vitro and in vivo. Hu/13 exhibited enhanced replication in MDCK cells, with larger plaques and higher viral titers than Ck/09 and Ck/12. In BALB/c mice, Hu/13 caused high pathogenicity and mortality, while Ck/09 induced minimal morbidity. Hu/13 replicated efficiently in respiratory tissues, eliciting robust cytokine responses and severe pulmonary lesions, with Ck/12 showing intermediate virulence. In ferrets, Hu/13 demonstrated efficient transmission, infecting all direct-contact and one airborne-contact ferret, whereas Ck/09 failed to transmit. Histopathology confirmed escalating lung pathology from Ck/09 to Ck/12 and Hu/13. Whole-genome sequencing identified adaptive mutations in Hu/13 during ferret replication, though no canonical mammalian-adaptive changes (e.g., PB2-E627K or HA-Q226L) were detected. These findings demonstrate progressive mammalian adaptation, replication efficiency, and transmissibility within the Taiwan H6N1 lineage, underscoring its zoonotic risk and emphasizing the need for enhanced surveillance to monitor mammalian-adaptive mutations, informing pandemic preparedness and public health strategies.
Keywords:
H6 subtype
; zoonotic influenza
; interspecies transmission
; spillover
; contact transmission
; airborne transmission
1. Introduction
Avian influenza viruses (AIVs) represent a persistent global threat to public health and agricultural due to their capacity for cross-species transmission and pandemic potential. Over the past decades, China has witnessed the emergence of numerous AIV subtypes, including H3N8, H5Nx, H6N1, H7N9/H7N4, H9N2, and H10Nx [1]. Several outbreaks of avian-origin influenza in humans, often referred to as “bird flu”, have underscored the critical role of domestic birds, particularly chickens, in facilitating the spillover of novel avian subtypes into human populations [1,2,3,4].
Among the diverse subtypes of influenza A viruses, H6 viruses are particularly notable for their high prevalence in both wild and domestic avian species, and their establishment in terrestrial poultry [5,6,7,8]. Despite their widespread circulation, the zoonotic risk posed by H6 viruses has historically been underestimated. The first documented human infection with an avian-origin H6N1 virus, A/Taiwan/02/2013 (Hu/13), in 2013 marked a pivotal moment in understanding the evolutionary dynamics and interspecies spillover risks of H6 viruses. This case, involving a patient with no direct poultry exposure, demonstrated the potential for H6 viruses to infect humans without prior adaptation [9].
The subsequent isolation of a closely related H6N1 strain from a dog in 2014 further highlighted the adaptive capacity of this lineage [10,11], which had been enzootically established in Taiwanese poultry since 1997 [8,12], and is now expanding its host range in mammals [9,10]. Structural and genomic analyses have identified H6 viruses harboring mammalian-adaptive mutations in key functional domains [5,11,12,13,14,15,16,17,18], emphasizing the critical need to investigate their biological properties and zoonotic potential.
Previous studies on the dog-origin H6N1 isolate revealed that mutations such as PB2-E627K significantly enhance viral polymerase activity and pathogenicity in mice [11], highlighting the role of host adaptation in cross-species transmission. However, the evolutionary trajectory of the Taiwan H6N1 lineage, spanning avian-to-mammalian hosts, remains poorly characterized. To address this gap, we investigated three temporally distinct H6N1 strains: A/Chicken/Taiwan/CF19/2009 (Ck/09), A/Chicken/Taiwan/2267/2012 (Ck/12) [12], and Hu/13 [9], using different mammalian models. Comparative analysis of these isolates enables the identification of genetic and phenotypic changes that may enhance mammalian infectivity, pathogenicity, replication efficiency, and transmissibility.
Our findings demonstrate a progressive increase in mammalian adaptation of the Taiwan H6N1 lineage, with Hu/13 exhibiting enhanced replication efficiency, heightened pathogenicity in mice, and efficient transmission in ferrets. These results provide critical insights into the evolutionary trajectory of H6 viruses and their potential to cause zoonotic infections, thereby informing surveillance and pandemic preparedness strategies. Furthermore, this study underscores the importance of continuous monitoring of H6 viruses for mammalian-adaptive mutations and highlights the need for sustained vigilance in understanding the factors that govern influenza virus evolution and cross-species transmission.
2. Materials and Methods
2.1. Ethical Compliance
All procedures involving animals were performed in accordance with institutional guidelines and internationally recognized standards for the humane treatment of laboratory animals. The experimental protocols were reviewed and approved by the Medical Animal Care and Welfare Committee of Shantou University Medical College (Approval No. SUMC2014-076) and the Committee on the Use of Live Animals in Teaching and Research (CULATR) of The University of Hong Kong (Approval No. 3265-14). Work with infectious agents was conducted in Biosafety Level 3 (BSL-3) containment facilities.
2.2. Virus Strains and Preparation
The human isolate A/Taiwan/02/2013 (Hu/13, EPI_ISL_143275) was provided by Prof. Ming-Tsan Liu (Centers for Disease Control, Taiwan) [9]. The chicken strains, A/Chicken/Taiwan/CF19/2009 (Ck/09, EPI_ISL_139240) and A/Chicken/Taiwan/2267/2012 (Ck/12, EPI_ISL_158030), were isolated as previously described and provided by Prof. Chwan-Chuen King (National Taiwan University) [12]. After plaque-purifications, the viral stocks were prepared in Madin-Darby canine kidney (MDCK) cells (ATCC CCL-34), and titrated using hemagglutination tests, plaque assays, 50% tissue culture infectious dose (TCID50), and 50% egg infectious dose (EID50) methods [19,20]. Plaque morphology was quantified by measuring the diameters of 40 randomly selected plaques using a Zeiss Axiovert 40 inverted microscope (Carl Zeiss AG, Germany) at 50× magnification. Aliquots were stored at −80°C until use.
2.3. Infection in Mice
Female BALB/c mice (Mus musculus, 8-10 weeks old, specific-pathogen-free) were obtained from Hunan SJA Laboratory Animal Co. Ltd. (China) and randomly allocated into experimental groups. Mice were intranasally inoculated with 50 μL of phosphate buffered saline (PBS) containing serial dilutions (101 to 106 PFU, plaque-forming units) of viruses (Ck/09, Ck/12, or Hu/13) under light anesthsia. Mock controls received PBS alone. Viruses with titers below 1×106 PFU/50 μL were concentrated using 100 kDa molecular weight cut-off (MWCO) Amicon® Ultra-0.5 centrifugal filter devices (Merck KGaA, Germany).
At 4 days post-inoculation (dpi), three animals per dose group were euthanized for sample collection (nasal turbinate, trachea, lungs and blood). Tissues were processed for virological titration, histopathology, and gene expression analysis. Viral titers in the supernatant from 1 ml of PBS homogenates were determined by TCID50 assays in MDCK cells. Remaining mice (n=5/group) were monitored daily for 14 days to assess survival and weight changes. Humane endpoints were defined as >25% weight loss. Mouse median lethal dose (MLD50) and 50% mouse infectious dose (MID50) were calculated using the Reed–Muench method [20].
2.4. Infection and Transmission in Ferrets
Male ferrets (Mustela putorius furo, 4-7 months old) from Wuxi Sangosho Biotechnology Co. Ltd. (China) were confirmed influenza-free and seronegative prior to use. Nine ferrets per group were intranasally inoculated (IN) with 106 PFU of virus (Ck/09, Ck/12, or Hu/13) under light anesthesia. At 1 dpi, three direct physical-contact (DC) ferrets were co-housed with inoculated animals, while three airborne-contact (AC) ferrets were placed in adjacent cages (5 cm separation; inter-cage airflow <0.2 m/s). Nasal washes and rectal swabs were collected daily for viral load quantification using TCID50. Three inoculated ferrets per group were euthanized at 4 and 7 dpi for tissue collection. Surviving animals were sacrificed at 14 dpi/dpc (days post-contact) for serum collection.
2.5. Hemagglutination Inhibition (HI) Test
Antibody titers in serum (14 dpi/dpc) were determined by HI assay using 0.5% turkey erythrocytes, as previously described [19]. HI titers of 40 or above were defined as seroconverted following influenza virus infection.
2.6. Histopathological and Immunohistochemical (IHC) Studies
2.7. Cytokine/Chemokine Quantification
Total RNA was extracted from lung homogenates using TRIzol® reagent (Thermo Fisher Scientific), treated with RNase-Free DNase (Qiagen), and reverse-transcribed using the PrimeScript™ RT reagent Kit (Takara). Transcript levels of cytokines/chemokines were quantified via SYBR Green-based real-time polymerase chain reaction (PCR) (LightCycler® 480, Roche Diagnostics) using validated primer sets (Tables S1 and S2) [23,24,25,26,27,28,29,30,31].
2.8. Whole Genomic Sequencing of the Virus
Viral RNA was extracted from inocula and ferret airway specimens using the QIAamp Viral RNA Mini Kit (Qiagen) and whole-genome amplification of the influenca virus was performed as previously described [22,32]. Libraries were prepared with the Nextera XT DNA Library Prep Kit (Illumina) and sequenced on an Illunima MiSeq benchtop sequencer (Illumina) (2×250 bp paired-end; Illumina). Raw reads were preprocessed using PRINSEQ [33] and assembled into contigs with the GS De Novo Assembler (version 2.6, Roche). Amino acid substitutions were annotated relative to reference genomes (EPI_ISL_139240, EPI_ISL_158030 and EPI_ISL_143275 for Ck/09, Ck/12 and Hu/13, respectively), with variant calling at a frequency ≥5% and minimum coverage depth of 100 reads.
2.9. Statistical Analysis
Statistical analyses were performed using GraphPad Prism 9.0.0 (GraphPad Software). Data are presented as mean ± standard deviation (SD). Two-group comparisons were analyzed using Student’s t-test, and multiple-group comparisons were analyzed using one-way analysis of variance (ANOVA). Statistical significance was defined as *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001 and ns, not significant.
3. Results
3.1. Viral Replication in MDCK Cells
Comparative analysis of Ck/09, Ck/12, and Hu/13 revealed distinct replication phenotypes. The human isolate (Hu/13) exhibited significantly larger plaque diameters (1,792 ± 227 μm) on MDCK cells compared to avian strains (Ck/09: 427 ± 102 μm; Ck/12: 1,506 ± 251 μm; P < 0.0001, unpaired two-tailed t-test; Figure 1). Viral titers quantified by EID50, TCID50, and PFU assays also demonstrated a progressive increase in replication efficiency: Ck/09 (EID50: 8.89×10⁷/ml; TCID50: 5.93×10⁶/ml; PFU: 2.25×10⁶/ml) < Ck/12 (EID50: 2.81×10⁸/ml; TCID50: 9.20×10⁷/ml; PFU: 6.00×10⁷/ml) < Hu/13 (EID50: 8.89×10⁸/ml; TCID50: 4.89×10⁸/ml; PFU: 3.00×10⁸/ml), with Hu/13 titers exceeding Ck/09 by 10-, 82.5-, and 133.3-fold, respectively (Table S3). These results suggest incremental adaptive mutations in the Taiwan H6N1 lineage enhancing mammalian cell tropism.
3.2. Viral Replication and Pathogenicity in BALB/c Mice
3.2.1. Clinical Outcomes and Survival
Mice inoculated with Hu/13 exhibited dose-dependent morbidity, with 106 PFU causing 100% mortality by 7 dpi. Intermediate virulence was observed for Ck/12 (60% survival at 10⁵ PFU; 0% at 10⁶ PFU), while Ck/09 showed minimal pathogenicity (100% survival at ≤10⁶ PFU; Figure 2G–I). MLD50 analysis confirmed Hu/13 as highly lethal (5.00 log₁₀ PFU), followed by Ck/12 (5.17 log₁₀ PFU) and Ck/09 (≥6.50 log₁₀ PFU; Table S4).
Weight loss mirrored survival trends: Hu/13 induced maximal weight reductions of over 25% (humane endpoint) at 104 to 10⁶ PFU, whereas Ck/09 caused ≤15.7% loss even at 10⁶ PFU (Figure 2D–F). Area under the curve (AUC) analysis of body weight changes revealed significant reduction for Hu/13 compared to Ck/09 at doses ≥ 103 PFU (P<0.05, Figure 2A–F).
3.2.2. Viral Loads in Respiratory Tissues
At 4 dpi, Hu/13 replicated efficiently across nasal turbinates (peak: 1.05×10⁵ TCID50/ml), trachea (5.93×10⁵ TCID50/ml), and lungs (1.05×10⁷ TCID50/ml). Ck/12 showed comparable replication in trachea (1.05×10⁵ TCID50/ml) and lung (1.87×10⁷ TCID50/ml), but higher replication in nasal turbinates at 105 PFU (1.87×106 TCID50/ml, P<0.05). Ck/09 was restricted to trachea (1.87×10⁴ TCID50/ml) and lungs (1.05×10⁴ TCID50/ml), with significantly lower replication at most doses (Figure 3A). The minimal infectious doses for productive virus replication were highest for Ck/09 and lowest for Hu/13 (Figure 3A).
3.2.3. Histopathology and Immune Responses
Immunohistochemistry revealed sparse NP-positive cells in Ck/09-infected lungs compared to dense foci in Ck/12- and Hu/13-infected tissues (Figure 3B, right panel). Histopathology confirmed escalating severity (Figure 3B, left panel) in Ck/12 and Hu/13, which showed severe pulmonary lesions, characterized by alveolar septal thickening and partial destruction of alveolar structures. Additionally, significant erythrocyte extravasation and inflammatory cell infiltration were evident within the alveoli and small bronchi, accompanied by necrosis and sloughing of epithelial cells in some small bronchioles. Contrastly, Ck/09 infection resulted only in mild bronchial epithelial sloughing (Figure 3B, left panel).
Hu/13 and Ck/12 also triggered robust cytokine upregulation compared to Ck/09, with Hu/13 consistently induced significantly higher levels of all tested cytokines/chemokines at dose ≥ 104 PFU (P<0.05; Figure 4).
3.2.4. Seroconversion and Infectivity Parameters
Serological antibody analysis revealed distinct patterns of immune response among the three H6N1 virus strains in inoculated mice (Figure S1). Ck/09 required ≥10⁴ PFU for seroconversion (HI titers ≥ 40), with complete conversion at 10⁶ PFU. Ck/12 exhibited seroconversion at ≥10² PFU, achieving full conversion at 10⁴ PFU, though mortality precluded antibody detection at higher doses. Hu/13 demonstrated the highest infectivity, with seroconversion at ≥10 PFU and complete conversion at 10³ PFU, despite mortality at elevated doses. The minimum infectious doses for seroconversion were 10⁴, 10², and 10 PFU for Ck/09, Ck/12, and Hu/13, respectively, establishing an infectivity hierarchy of Ck/09 << Ck/12 < Hu/13 (Figure S1). Likewise, the MID50 values were also found to decrease sequentially: Ck/09 (4.50 log₁₀ PFU) > Ck/12 (2.00 log₁₀ PFU) > Hu/13 (1.50 log₁₀ PFU; Table S4).
3.3. Viral Infection and Transmission Dynamics in Ferrets
3.3.1. Clinical Observations
Ferrets intranasally inoculated with 10⁶ PFU of each H6N1 virus exhibited distinct clinical responses. Hu/13 caused the most pronounced effects, including significant temperature increases (up to 2°C) and activity reduction for 1–2 days, along with marked sneezing and nasal discharge. Ck/12 induced moderate symptoms, while Ck/09 caused minimal changes, with less than 1°C temperature increase and no inactivity. No body weight loss was observed in any infected ferrets.
3.3.2. Viral Shedding and Transmission
Hu/13 demonstrated efficient replication and transmission, with peak nasal wash titers reaching 5.93 × 10⁶ TCID₅₀/mL. It infected all direct-contact ferrets (3/3) and two airborne-contact ferret (2/3), leading to marked seroconversions. Ck/12 showed moderate transmission, infecting two direct-contact ferrets (2/3) but none of the airborne-contact ferrets (0/3). Ck/09 failed to transmit to either direct- or airborne-contact ferrets (Figure 5, Table 1). Viral loads in the respiratory tracts followed the hierarchy Hu/13 > Ck/12 > Ck/09 (Figure 6A). No viable virus was detected in rectal swabs throughout the experiment.
3.3.3. Pathological Changes in Ferrets
Hu/13 induced severe pulmonary lesions, including bronchial necrosis and leukocyte infiltration, while Ck/09 caused mild pathology (Figure 6B). Ck/12 exhibited intermediate severity, with overt alveolar thickening and inflammatory cell infiltration (Figure 6B). IHC staining for viral NP antigens confirmed higher replication of Hu/13 and Ck/12 in lung tissues compared to Ck/09 (Figure 6B).
3.3.4. Seroconversion
All ferrets inoculated with Hu/13 seroconverted by 14 dpi, with HI titers ≥ 1280. Similarly, all Ck/12-inoculated ferrets seroconverted, with HI titers ranging from 640 to 1280. Ck/09-inoculated ferrets also seroconverted but with lower HI titers (320–640). Transmission groups mirrored these findings, with Hu/13 showing the highest seroconversion rates (Table 1). Notably, in the Hu/13 transmission group, one direct-contact ferret and one airborne-contact ferret showed seroconversion without detectable viral shedding (Figure 5C, Table 1).
3.3.5. Proinflammatory Responses
Proinflammatory cytokine and chemokine levels in lung tissues generally correlated with viral virulence. However, due to marked individual variations, differences among the virus groups were not statistically significant (P > 0.05, one-way ANOVA or Student’s t-tests between any groups; Figure 7).
3.3.6. Viral Mutations and Adaptation
Next-generation sequencing of Hu/13 during ferret passage identified a number of substitutions across the viral genome, with dominant mutations in PB1, HA and NP. Notably, no canonical mammalian-adaptive mutations (e.g., PB2-E627K, PB2-D701N or HA-Q226L) were observed, suggesting that Hu/13 efficiently transmits in ferrets without requiring these well-characterized adaptive changes (Table S5).
4. Discussion
The interspecies transmission of AIVs continues to pose significant threats, particularly in densely populated regions with extensive poultry production, such as China. Among the most frequently detected influenza subtypes in aquatic birds, H6 viruses have sporadically spilled over into terrestrial poultry, leading to the establishment of enzootic clades [5,6,7,8]. The Taiwan H6N1 lineage exemplifies this evolutionary trajectory, having evolved into a distinct clade in chickens since 1997 and demonstrating increasing potential for mammalian adaptation over time [5,8,9,10,11,14]. This is evidenced by the first documented human and canine infections [9,10], as well as the progressive adaptation of temporally distinct strains in mammalian models, as highlighted in this study and previous research [8].
In this study, we compared the infectivity, pathogenicity, and transmissibility of three Taiwan H6N1 isolates (Ck/09, Ck/12, and Hu/13) in mammalian models. The results demonstrated a progressive enhancement in replication efficiency in MDCK cells, mice, and ferrets, with Hu/13 showing the highest capability, followed by Ck/12 and then Ck/09. Pathogenicity in mice and transmissibility in ferrets showed a similar trend, with Hu/13 exhibiting efficient contact transmission among ferrets. Notably, earlier H6N1 isolates from this Taiwan lineage, such as those from 1998 and 1999, were unable to replicate efficiently in mice, while isolates from 2002 and 2005 showed moderate replication [8]. In contrast, recent isolates like Ck/12 and Hu/13 displayed significantly enhanced replication in mammalian models, with Hu/13 causing severe morbidity and 100% mortality at 10⁶ PFU in mice. These findings suggest that prolonged circulation and adaptation of H6N1 viruses in chicken populations enhance their replication efficiency and transmissibility in mammals. This pattern mirrors observations in chicken-adapted H9N2 in China and its related derivative viruses (e.g., H3N8, H7N9 and H10N8), which have also demonstrated increasing zoonotic potential over time [3,4,22,34,35]. Furthermore, the replication capabilities of Ck/12 and Hu/13 in mice exceed those of H6 viruses isolated from other regions of China, as shown in comparative studies [7,36,37,38]. Collectively, these results underscore the growing zoonotic risk posed by Taiwan H6N1 viruses and highlight the need for continuous surveillance and research to mitigate public health threats.
Chickens play a critical role in the ecology of influenza viruses, serving as an intermediate host between aquatic birds and mammals [3,4,22,39,40]. The transition of H6 viruses from waterfowl to chickens has facilitated their adaptation to mammalian systems, as evidenced by the increasing replication efficiency and pathogenicity of recent Taiwan H6N1 isolates. The shift in viral replication sites from the intestines (in waterfowl) to the trachea (in chickens) has enabled H6 viruses to acquire α2-6 receptor binding capacity, a key step in mammalian adaptation [13,14,15,16,39]. This study provides further evidence that chickens act as a “bridge” for influenza viruses, promoting their evolution and potential spillover into humans and other mammals.
The enhanced zoonotic potential of chicken-adapted H6N1 may stem from multiple factors: (1) Receptor-binding shifts toward α2,6-linked sialic acids in chicken and mammalian respiratory tracts [39]; (2) Host immune and antiviral responses driving virus evolution, such as the lack of retinoic acid-inducible gene-I (RIG-I) in chickens, which orchestrates the production of antiviral interferon-stimulated genes (ISGs) [41]; (3) High evolutionary rates in chickens, which lack the evolutionary stasis seen in waterfowl reservoirs [6,40], promoting rapid adaptation; and (4) Co-circulation of diverse subtypes (e.g., H5N2) enabling gene reassortment [12]. Chickens, as non-natural reservoir hosts, may also impose other unique selective pressures (e.g., body temperature, mucosal pH, enzymes) that inadvertently favor variants with broader host tropism.
The molecular basis for H6 virus adaptation to mammals involves key mutations in multiple viral proteins. In the HA protein, E190V and G228S substitutions, prevalent post-2000 in Taiwan H6N1, enhance human-like α2,6-sialic acid affinity and bridging avian-mammalian receptor tropism [5,9,13,14,15,16]. Compared to avian isolates, Hu/13 contains an additional P186L mutation (Table S6), which reduced its affinity for avian-type receptors [14]. Although the Taiwan H6N1 lineage retains HA-226Q, the triad of S137N, E190V and G228S in many recent isolates may alleviate the requirement for a hydrophobic residue at the 226 position, increasing preference for human receptors [13]. In the NA protein, stalk deletions at position 41–52 and 68–69 (N1-Δ14) are associated with increased adaptation to terrestrial poultry and mammals [9,18], indicating their long-term adaptation in chickens and potential spillover to mammals.
Whole-genome comparative analysis revealed that Ck/09 had 134 and 126 amino acid differences with Ck/12 and Hu/13, respectively (Table S6). While Hu/13 lacks canonical mammalian-adaptive mutations like PB2-E627K, Q591R and D701N in the internal genes [5,9], it exhibits other potential adaptive changes that may contribute to its enhanced replication and pathogenicity in mammals. These include consistent changes in Ck/12 and Hu/13, such as D249E, S265N, and I292V mutations in PB2 [43,44,45]; A85T, T97N, and L336M in PA [46,47,48,49]; and truncations in PB1-F2 (87 vs. 57 aa), a virulence modulator in mammals [50]. These findings suggest that H6 viruses can evolve to infect mammals through a combination of mutations, even in the absence of the most well-characterized adaptive markers. However, as the biological functions of these substitutions are primarily reported in H5N1, H1N1, or H7N9 strains [43,44,45,46,47,48,49,50], further investigation is needed to confirm their roles in the Taiwan H6N1 context.
While this study provides insights into the progressive evolution of H6N1 virus with increasing zoonotic potentials, several limitations must be acknowledged. First, the absence of post-2013 H6N1 isolates precludes assessment of ongoing evolution in Taiwanese poultry, and the use of limited number of strains may not fully recapitulate the genetic diversity and evolutionary dynamics of H6N1 in natural poultry populations. Second, while mouse and ferret models are widely used for influenza studies, they may not perfectly predict human infection, pathogenicity and transmission patterns. Future research should incorporate more field isolates and explore alternative models to validate these findings. Third, while we identified amino acid variations between the avian and human isolates (Table S6) and adaptive mutations in ferret-passaged Hu/13 (Table S5), functional validation of these changes is needed. Additionally, investigating the role of host immune responses in shaping viral adaptation could provide a more comprehensive understanding of the factors driving cross-species transmission.
The progressive adaptation of H6N1 viruses in mammalian systems highlights the need for enhanced surveillance and control measures. The detection of subclinical infections in ferrets and the increasing replication efficiency of recent isolates, coupled with the silent persistence of H6N1 in Taiwan, underscore the potential for H6 viruses to cause outbreaks in humans. Given the high prevalence of H6 viruses in poultry and their ability to acquire mammalian-adaptive traits, continuous monitoring of viral evolution in poultry populations is essential. This includes tracking key mutations in HA, NA, and internal genes that may enhance viral replication, transmission, and pathogenicity in mammals. A One-Health approach and proactive risk assessment, including reverse genetics studies to pinpoint transmission determinants, are critical to mitigate pandemic threats.
5. Conclusions
The Taiwan H6N1 lineage provides a compelling example of the stepwise adaptation of avian influenza viruses to mammalian hosts, demonstrating how prolonged poultry adaptation can bridge avian-mammalian barriers. The enhanced replication efficiency, pathogenicity, and transmissibility of recent isolates, such as Hu/13, highlight the zoonotic potential of H6 viruses. These findings underscore the importance of ongoing surveillance and research to identify and mitigate the risks posed by H6 viruses, particularly in regions with extensive poultry production and human-animal contact. By understanding the molecular mechanisms of viral adaptation and the role of intermediate hosts like chickens, we can better prepare for and prevent future influenza pandemics.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Table S1: Primers used for quantifying cytokine and chemokine expression in mice; Table S2: Primers used for quantifying cytokine and chemokine expression in ferrets; Table S3: Virus titers of the Taiwan H6N1 influenza viruses measured in MDCK cells and embryonated chicken eggs; Table S4: Mouse median lethal dose (MLD₅₀) and 50% mouse infectious dose (MID₅₀) of the Taiwan H6N1 influenza viruses; Table S5: Amino acid substitutions identified in the A/Taiwan/02/2013 (Hu/13) H6N1 influenza virus during infection and transmission in ferrets; Table S6. Amino acid variations identified in the three Taiwan H6N1 influenza virus isolates. Figure S1: Serological antibody titers in mice inoculated with Taiwan H6N1 influenza viruses.
Author Contributions
Conceptualization, H.Z.; methodology, Z.Z., Li.L., Y.L., H.Z. and Y.G.; validation, Z.Z. and H.Z.; formal analysis, Z.Z. and H.Z.; investigation, Z.Z., X.C., R.Z., Z.Y., Lo.L., R.C., Li.L., and Y.L.; resources, Y.G. and H.Z.; data curation, Z.Z. and H.Z.; writing—original draft preparation, Z.Z. and H.Z.; writing—review and editing, H.Z. and Z.Z.; visualization, Z.Z. and H.Z.; supervision, H.Z. and Y.G.; project administration, H.Z. and Y.G.; funding acquisition, Y.G. and H.Z.. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Health and Medical Research Fund, grant number 14131402; Hong Kong Research Grant Council, grant numbers T11-705/14-N and T11-712/19-N; National Institutes of Health, contract numbers HHSN272201400006C and 75N93021C00016, and Li Ka Shing Foundation. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the article.
Institutional Review Board Statement
The animal study protocol was approved by Committee on the Use of Live Animals in Teaching and Learning (CULATR) of The University of Hong Kong (protocol number 3265-14, 11 March 2014) and Medical Animal Care and Welfare Committee of Shantou University Medical College (protocol number SUMC2014-076, 1 March 2014).
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article and supplementary material. Further inquiries can be directed to the corresponding authors.
Acknowledgments
We gratefully acknowledge all the staff of the Guangdong-Hong Kong Joint Laboratory of Emerging Infectious Diseases/ Joint Laboratory for International Collaboration in Virology and Emerging Infectious Diseases/ Joint Institute of Virology (STU/HKU) and SKLEID for their technical support and administrative assistance. Some data were presented at the International Forum on Ecology and Evolution of Avian Influenza (EEAIV webinars) on 7 June, 2022 and at the 13th Annual CEIRS (Centres of Excellence for Influenza Research and Surveillance) Network Meeting on 12 January, 2021 (online).
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| AIV | avian influenza virus |
| Hu/13 | A/Taiwan/02/2013 |
| Ck/09 | A/Chicken/Taiwan/CF19/2009 |
| Ck/12 | A/Chicken/Taiwan/2267/2012 |
| CULATR | Committee on the Use of Live Animals in Teaching and Learning |
| BSL-3 | Biosafety Level 3 |
| MDCK | Madin-Darby canine kidney |
| TCID50 | 50% tissue culture infectious dose |
| EID50 | 50% egg infectious dose |
| PBS | phosphate buffered saline |
| PFU | plaque-forming unit |
| MWCO | molecular weight cut-off |
| dpi | day(s) post-inoculation |
| MLD50 | Mouse median lethal dose |
| MID50 | 50% mouse infectious dose |
| IN | intranasally inoculated |
| DC | direct physical-contact |
| AC | airborne-contact |
| dpc | day(s) post-contact |
| HI | hemagglutination inhibition |
| IHC | immunohistochemical |
| HE | hematoxylin and eosin |
| NP | nucleoprotein |
| PCR | polymerase chain reaction |
| SD | standard deviation |
| ANOVA | analysis of variance |
| AUC | area under the curve |
| RIG-I | retinoic acid-inducible gene-I |
| ISGs | antiviral interferon-stimulated genes |
| CDC | Centers for Disease Control and Prevention |
| WHO | World Health Organization |
References
- CDC. Reported Human Infections with Avian Influenza A Viruses. Available online: https://www.cdc.gov/bird-flu/php/avian-flu-summary/reported-human-infections.html (accessed on 16 April, 2025).
- Alexander, D.J. An overview of the epidemiology of avian influenza. Vaccine 2007, 25, 5637–5644. [Google Scholar] [CrossRef] [PubMed]
- Lam, T.T.; Wang, J.; Shen, Y.; Zhou, B.; Duan, L.; Cheung, C.L.; Ma, C.; Lycett, S.J.; Leung, C.Y.; Chen, X.; et al. The genesis and source of the H7N9 influenza viruses causing human infections in China. Nature 2013, 502, 241–244. [Google Scholar] [CrossRef]
- Ma, C.; Lam, T.T.; Chai, Y.; Wang, J.; Fan, X.; Hong, W.; Zhang, Y.; Li, L.; Liu, Y.; Smith, D.K.; et al. Emergence and evolution of H10 subtype influenza viruses in poultry in China. J. Virol. 2015, 89, 3534–3541. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Li, Y.; Huang, S.; Wen, F. Global distribution, receptor binding, and cross-species transmission of H6 influenza viruses: risks and implications for humans. J Virol. 2023, 97, e01370–23. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Zhu, H.; Fan, X.; Wang, J.; Cheung, C.L.; Duan, L.; Hong, W.; Liu, Y.; Li, L.; Smith, D.K.; et al. Establishment and lineage replacement of H6 influenza viruses in domestic ducks in southern China. J. Virol. 2012, 86, 6075–6083. [Google Scholar] [CrossRef]
- Wang, G.; Deng, G.; Shi, J.; Luo, W.; Zhang, G.; Zhang, Q.; Liu, L.; Jiang, Y.; Li, C.; Sriwilaijaroen, N; et al. H6 influenza viruses pose a potential threat to human health. J. Virol. 2014, 88, 3953–3964. [Google Scholar] [CrossRef]
- Lee, M.S.; Chang, P.C.; Shien, J.H.; Cheng, M.C.; Chen, C.L.; Shieh, H.K. Genetic and pathogenic characterization of H6N1 avian influenza viruses isolated in Taiwan between 1972 and 2005. Avian Dis. 2006, 50, 561–571. [Google Scholar] [CrossRef]
- Wei, S.H.; Yang, J.R.; Wu, H.S.; Chang, M.C.; Lin, J.S.; Lin, C.Y.; Liu, Y.L.; Lo, Y.C.; Yang, C.H.; Chuang, J.H.; et al. Human infection with avian influenza A H6N1 virus: an epidemiological analysis. Lancet Respir Med. 2013, 1, 771–778. [Google Scholar] [CrossRef]
- Lin, H.T.; Wang, C.H.; Chueh, L.L.; Su, B.L.; Wang, L.C. Influenza A(H6N1) Virus in Dogs, Taiwan. Emerg Infect Dis. 2015, 21, 2154–2157. [Google Scholar] [CrossRef]
- Tsai, S.K.; Shih, C.H.; Chang, H.W.; Teng, K.H.; Hsu, W.E.; Lin, H.J.; Lin, H.Y.; Huang, C.H.; Chen, H.W.; Wang, L.C. Replication of a Dog-Origin H6N1 Influenza Virus in Cell Culture and Mice. Viruses 2020, 12, 704. [Google Scholar] [CrossRef]
- Lee, C.C.; Zhu, H.; Huang, P.Y.; Peng, L.; Chang, Y.C.; Yip, C.H.; Li, Y.T.; Cheung, C.L.; Compans, R.; Yang, C.; et al. Emergence and evolution of avian H5N2 influenza viruses in chickens in Taiwan. J Virol. 2014, 88, 5677–5686. [Google Scholar] [CrossRef] [PubMed]
- Ni, F.; Kondrashkina, E.; Wang, Q. Structural and Functional Studies of Influenza Virus A/H6 Hemagglutinin. PLoS One 2015, 10, e0134576. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Qi, J.; Bi, Y.; Zhang, W.; Wang, M.; Zhang, B.; Wang, M.; Liu, J.; Yan, J.; Shi, Y.; et al. Adaptation of avian influenza A (H6N1) virus from avian to human receptor-binding preference. EMBO J. 2015, 34, 1661–1673. [Google Scholar] [CrossRef]
- Yang, H.; Carney, P.J.; Chang, J.C.; Villanueva, J.M.; Stevens, J. Structure and receptor binding preferences of recombinant hemagglutinins from avian and human H6 and H10 influenza A virus subtypes. J Virol. 2015, 89, 4612–4623. [Google Scholar] [CrossRef]
- Tzarum, N.; de Vries, R.P.; Zhu, X.; Yu, W.; McBride, R.; Paulson, J.C.; Wilson, I.A. Structure and receptor binding of the hemagglutinin from a human H6N1 influenza virus. Cell Host Microbe 2015, 17, 369–376. [Google Scholar] [CrossRef]
- Yuan, J.; Zhang, L.; Kan, X.; Jiang, L.; Yang, J.; Guo, Z.; Ren, Q. Origin and molecular characteristics of a novel 2013 avian influenza A(H6N1) virus causing human infection in Taiwan. Clin Infect Dis. 2013, 57, 1367–1368. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Shi, Y.; Wu, Y.; Liu, D.; Gao, G.F. Origin and molecular characterization of the human-infecting H6N1 influenza virus in Taiwan. Protein Cell 2013, 4, 846–853. [Google Scholar] [CrossRef]
- WHO Global Influenza Surveillance Network. Manual for the laboratory diagnosis and virological surveillance of influenza. Available online: https://www.who.int/publications/i/item/manual-for-the-laboratory-diagnosis-and-virological-surveillance-of-influenza (accessed on 27 February, 2025).
- Reed, L. J.; Muench, H. A simple method of estimating fifty percent endpoints. Am J Hyg. 1938, 27, 493–497. [Google Scholar]
- Li, L.; Chen, R.; Yan, Z.; Cai, Q.; Guan, Y.; Zhu, H. Experimental infection of rats with influenza A viruses: Implications for murine rodents in influenza A virus ecology. Viruses 2025, 17, 495. [Google Scholar] [CrossRef]
- Chen, P.; Jin, Z.; Peng, L.; Zheng, Z.; Cheung, Y.M.; Guan, J.; Chen, L.; Huang, Y.; Fan, X.; Zhang, Z.; et al. Characterization of an Emergent Chicken H3N8 Influenza Virus in Southern China: a Potential Threat to Public Health. J Virol. 2023, 97, e0043423. [Google Scholar] [CrossRef]
- Giulietti, A.; Overbergh, L.; Valckx, D.; Decallonne, B.; Bouillon, R.; Mathieu, C. An overview of real-time quantitative PCR: applications to quantify cytokine gene expression. Methods 2001, 25, 386–401. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Friggeri, A.; Yang, Y.; Park, Y.J.; Tsuruta, Y.; Abraham, E. miR-147, a microRNA that is induced upon Toll-like receptor stimulation, regulates murine macrophage inflammatory responses. Proc Natl Acad Sci U S A. 2009, 106, 15819–15824. [Google Scholar] [CrossRef]
- Martinez-Gil, L.; Goff, P.H.; Hai, R.; Garcia-Sastre, A.; Shaw, M.L.; Palese, P. A Sendai virus-derived RNA agonist of RIG-I as a virus vaccine adjuvant. J Virol. 2013, 87, 1290–1300. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ma, X. Interferon regulatory factor 8 regulates RANTES gene transcription in cooperation with interferon regulatory factor-1, NF-kappaB, and PU.1. J Biol Chem. 2006, 281, 19188–19195. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Kemper, A.; McCarthy, K.D.; Pytel, P.; Wang, J.P.; Campbell, I.L.; Utset, M.F.; Popko, B. Interferon-gamma induced medulloblastoma in the developing cerebellum. J Neurosci. 2004, 24, 10074–10083. [Google Scholar] [CrossRef]
- Arima, Y.; Harada, M.; Kamimura, D.; Park, J.H.; Kawano, F.; Yull, F.E.; Kawamoto, T.; Iwakura, Y.; Betz, U.A.; Marquez, G.; et al. Regional neural activation defines a gateway for autoreactive T cells to cross the blood-brain barrier. Cell 2012, 148, 447–457. [Google Scholar] [CrossRef]
- Yoshida, S.; Yoshida, A.; Ishibashi, T.; Elner, S.G.; Elner, V.M. Role of MCP-1 and MIP-1alpha in retinal neovascularization during postischemic inflammation in a mouse model of retinal neovascularization. J Leukoc Biol. 2003, 73, 137–144. [Google Scholar] [CrossRef]
- Svitek, N.; von Messling, V. Early cytokine mRNA expression profiles predict Morbillivirus disease outcome in ferrets. Virology 2007, 362, 404–410. [Google Scholar] [CrossRef]
- Maines, T.R.; Belser, J.A.; Gustin, K.M.; van Hoeven, N.; Zeng, H.; Svitek, N.; von Messling, V.; Katz, J.M.; Tumpey, T.M. Local innate immune responses and influenza virus transmission and virulence in ferrets. J Infect Dis. 2012, 205, 474–485. [Google Scholar] [CrossRef]
- Zhou, B.; Donnelly, M.E.; Scholes, D.T.; St George, K.; Hatta, M.; Kawaoka, Y.; Wentworth, D.E. Single-reaction genomic amplification accelerates sequencing and vaccine production for classical and Swine origin human influenza a viruses. J Virol. 2009, 83, 10309–10313. [Google Scholar] [CrossRef]
- Schmieder, R.; Edwards, R. Quality control and preprocessing of metagenomic datasets. Bioinformatics 2011, 27, 863–864. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Belser, J.A.; Maines, T.R. Adaptation of H9N2 Influenza Viruses to Mammalian Hosts: A Review of Molecular Markers. Viruses. 2020, 12, 541. [Google Scholar] [CrossRef]
- Li, B.; Su, G.; Xiao, C.; Zhang, J.; Li, H.; Sun, N.; Lao, G.; Yu, Y.; Ren, X.; Qi, W.; et al. The PB2 co-adaptation of H10N8 avian influenza virus increases the pathogenicity to chickens and mice. Transbound Emerg Dis. 2022, 69, 1794–1803. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Cui, H.; Teng, Q.; Li, L.; Shi, Y.; Li, X.; Yang, J.; Liu, Q.; Deng, J.; Li, Z. Evolution and pathogenicity of H6 avian influenza viruses isolated from Southern China during 2011 to 2017 in mice and chickens. Sci Rep. 2020, 10, 20583. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, M.; Tian, J.; Bai, X.; Li, Y. Genetic characteristics and pathogenicity of novel reassortant H6 viruses isolated from wild birds in China. Vet Microbiol. 2021, 254, 108978. [Google Scholar] [CrossRef]
- Xu, X.; Chen, Q.; Tan, M.; Liu, J.; Li, X.; Yang, L.; Shu, Y.; Wang, D.; Zhu, W. Epidemiology, evolution, and biological characteristics of H6 avian influenza viruses in China. Emerg Microbes Infect. 2023, 12, 2151380. [Google Scholar] [CrossRef]
- Kuchipudi, S.V.; Nelli, R.; White, G.A.; Bain, M.; Chang, K.C.; Dunham, S. Differences in influenza virus receptors in chickens and ducks: Implications for interspecies transmission. J Mol Genet Med. 2009, 3, 143–151. [Google Scholar] [CrossRef]
- Webster, R.G.; Bean, W.J.; Gorman, O.T.; Chambers, T.M.; Kawaoka, Y. Evolution and ecology of influenza A viruses. Microbiol. Rev. 1992, 56, 152–179. [Google Scholar] [CrossRef]
- Barber, M.R.W.; Aldridge, J.R., Jr.; Webster, R.G.; Magor, K.E. Association of RIG-I with innate immunity of ducks to influenza. Proc Natl Acad Sci U S A. 2010, 107, 5913–5918. [Google Scholar] [CrossRef]
- Li, J.; Zu Dohna, H.; Cardona, C.J.; Miller, J.; Carpenter, T.E. Emergence and Genetic Variation of Neuraminidase Stalk Deletions in Avian Influenza Viruses. PLoS One. 2011, 6, e14722. [Google Scholar] [CrossRef]
- Yamaji, R.; Yamada, S.; Le, M.Q; Li, C.; Chen, H.; Qurnianingsi, E.; Nidom, C.A.; Ito, M.; Sakai-Tagawa, Y.; Kawaoka, Y. Identification of PB2 mutations responsible for the efficient replication of H5N1 influenza viruses in human lung epithelial cells. J Virol. 2015, 89, 3947–3956. [Google Scholar] [CrossRef] [PubMed]
- Cox, A.; Schmierer, J.; D’Angelo, J.; Smith, A.; Levenson, D.; Treanor, J.; Kim, B.; Dewhurst, S. A Mutated PB1 Residue 319 Synergizes with the PB2 N265S Mutation of the Live Attenuated Influenza Vaccine to Convey Temperature Sensitivity. Viruses 2020, 12, 1246. [Google Scholar] [CrossRef] [PubMed]
- Kong, H.; Ma, S.; Wang, J.; Gu, C.; Wang, Z.; Shi, J.; Deng, G.; Guan, Y.; Chen, H. Identification of Key Amino Acids in the PB2 and M1 Proteins of H7N9 Influenza Virus That Affect Its Transmission in Guinea Pigs. J Virol. 2019, 94, e01180–19. [Google Scholar] [CrossRef]
- Bussey, K.A.; Desmet, E.A.; Mattiacio, J.L.; Hamilton, A.; Bradel-Tretheway, B.; Bussey, H.E.; Kim, B.; Dewhurst, S.; Takimoto, T. PA residues in the 2009 H1N1 pandemic influenza virus enhance avian influenza virus polymerase activity in mammalian cells. J Virol. 2011, 85, 7020–8. [Google Scholar] [CrossRef] [PubMed]
- Song, M.S.; Pascua, P.N.Q.; Lee, J.H.; Baek, Y.H.; Lee, O.J.; Kim, C.J.; Kim, H.; Webby, R.J.; Webster, R.G.; Choi, Y.K. The polymerase acidic protein gene of influenza a virus contributes to pathogenicity in a mouse model. J Virol. 2009, 83, 12325–12335. [Google Scholar] [CrossRef]
- Taft, A.S.; Ozawa, M.; Fitch, A.; Depasse, J.V.; Halfmann, P.J.; Hill-Batorski, L.; Hatta, M.; Friedrich, T.C.; Lopes, T.J.S.; Maher, E.A.; et al. Identification of mammalian-adapting mutations in the polymerase complex of an avian H5N1 influenza virus. Nat Commun. 2015, 6, 7491. [Google Scholar] [CrossRef]
- Yu, Z.; Cheng, K.; Xin, Y.; Sun, W.; Li, X.; Huang, J.; Zhang, K.; Yang, S.; Wang, T.; Zheng, X.; et al. Multiple amino acid substitutions involved in the adaptation of H6N1 avian influenza virus in mice. Vet Microbiol. 2014, 174, 316–321. [Google Scholar] [CrossRef]
- Kamal, R.P. , Alymova, I.V., York, I.A. Evolution and Virulence of Influenza A Virus Protein PB1-F2. Int J Mol Sci. 2017, 19, 96. [Google Scholar] [CrossRef]
Figure 1.
Plaques formed by the Taiwan H6N1 influenza viruses on MDCK cells. Mean plaque diameter for each virus (n=40) is plotted above, error bars denote standard deviation (SD), **** indicates P<0.0001. Plaque phenotypes are shown below; scale bars indicate 5 mm. Ck/09, A/Chicken/Taiwan/CF19/2009 (green); Ck/12, A/Chicken/Taiwan/2267/2012 (blue); Hu/13, A/Taiwan/02/2013 (red).
Figure 1.
Plaques formed by the Taiwan H6N1 influenza viruses on MDCK cells. Mean plaque diameter for each virus (n=40) is plotted above, error bars denote standard deviation (SD), **** indicates P<0.0001. Plaque phenotypes are shown below; scale bars indicate 5 mm. Ck/09, A/Chicken/Taiwan/CF19/2009 (green); Ck/12, A/Chicken/Taiwan/2267/2012 (blue); Hu/13, A/Taiwan/02/2013 (red).
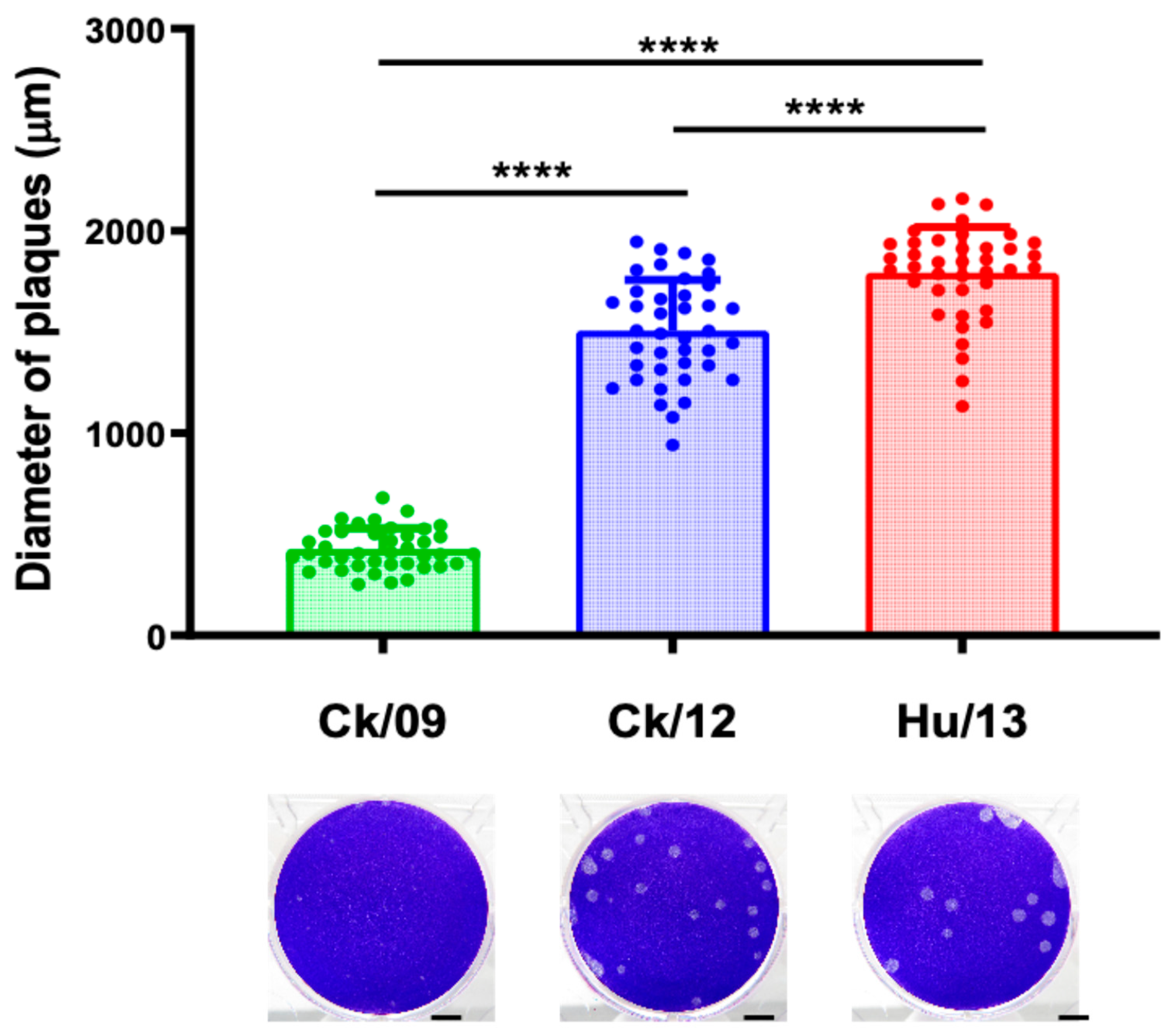
Figure 2.
Clinical outcomes of Taiwan H6N1 influenza virus infection in mice. Groups of five BALB/c mice were inoculated with Ck/09 (green), Ck/12 (blue) or Hu/13 (red) at the indicated dose. Mock controls received phosphate-buffered saline (PBS, grey) alone. Body weight changes (A-F) and survival rates (G-I) were monitored for 14 days post-inoculation (dpi). At the inoculation dose of 10-1,000 PFU, no deaths were observed. Results are presented as mean ± SD.
Figure 2.
Clinical outcomes of Taiwan H6N1 influenza virus infection in mice. Groups of five BALB/c mice were inoculated with Ck/09 (green), Ck/12 (blue) or Hu/13 (red) at the indicated dose. Mock controls received phosphate-buffered saline (PBS, grey) alone. Body weight changes (A-F) and survival rates (G-I) were monitored for 14 days post-inoculation (dpi). At the inoculation dose of 10-1,000 PFU, no deaths were observed. Results are presented as mean ± SD.
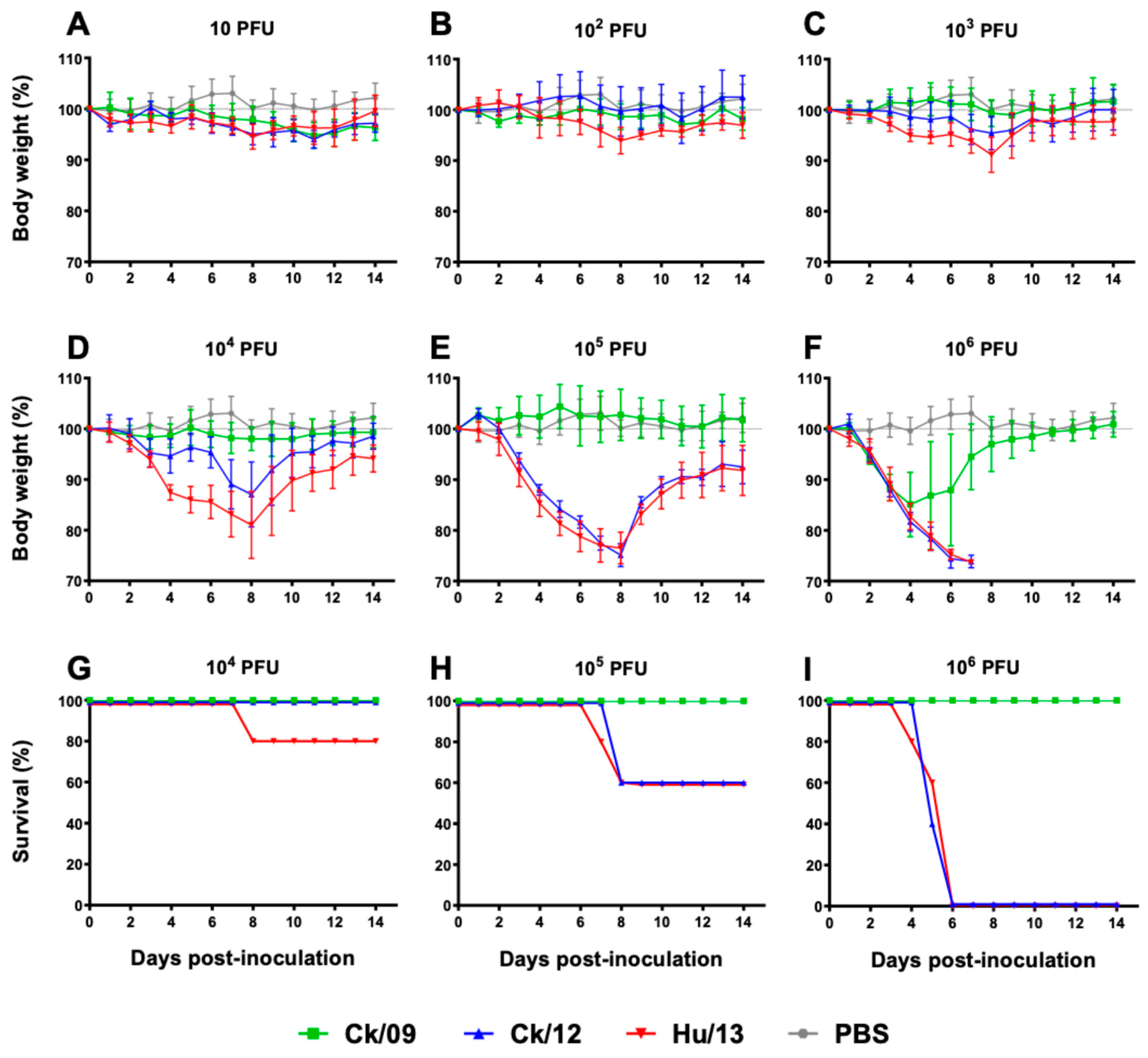
Figure 3.
Replication of Taiwan H6N1 influenza viruses in the respiratory tissues of mice. Nasal turbinate, trachea and lungs were harvested at 4 dpi (n=3 per dose). (A) Viral titers were determined by TCID50 assays. Results are presented as mean ± SD. Statistical significance is denoted as *P<0.05; **P<0.01; ***P<0.001; ****P<0.0001. Comparative results are presented as follows: comparisons between Ck/09 (green) and Hu/13 (red) are indicated above the Ck/09 groups; those between Ck/09 and Ck/12 (blue) are shown above the Ck/12 groups; and comparisons between Ck/12 and Hu/13 are labeled above the Hu/13 groups. No virus was detected in the mock-infected group inoculated with PBS (grey). The lower limit of detection is indicated by the horizontal dashed line. (B) Histopathological and immunohistochemical (IHC) analyses were performed on lung tissues from mice inoculated with 105 PFU of each virus. The left panel shows hematoxylin and eosin (HE) staining, while the right panel demonstrates viral nucleoprotein (NP) detection, with brown color indicating positively stained cells. Scale bars indicate 100 μm.
Figure 3.
Replication of Taiwan H6N1 influenza viruses in the respiratory tissues of mice. Nasal turbinate, trachea and lungs were harvested at 4 dpi (n=3 per dose). (A) Viral titers were determined by TCID50 assays. Results are presented as mean ± SD. Statistical significance is denoted as *P<0.05; **P<0.01; ***P<0.001; ****P<0.0001. Comparative results are presented as follows: comparisons between Ck/09 (green) and Hu/13 (red) are indicated above the Ck/09 groups; those between Ck/09 and Ck/12 (blue) are shown above the Ck/12 groups; and comparisons between Ck/12 and Hu/13 are labeled above the Hu/13 groups. No virus was detected in the mock-infected group inoculated with PBS (grey). The lower limit of detection is indicated by the horizontal dashed line. (B) Histopathological and immunohistochemical (IHC) analyses were performed on lung tissues from mice inoculated with 105 PFU of each virus. The left panel shows hematoxylin and eosin (HE) staining, while the right panel demonstrates viral nucleoprotein (NP) detection, with brown color indicating positively stained cells. Scale bars indicate 100 μm.
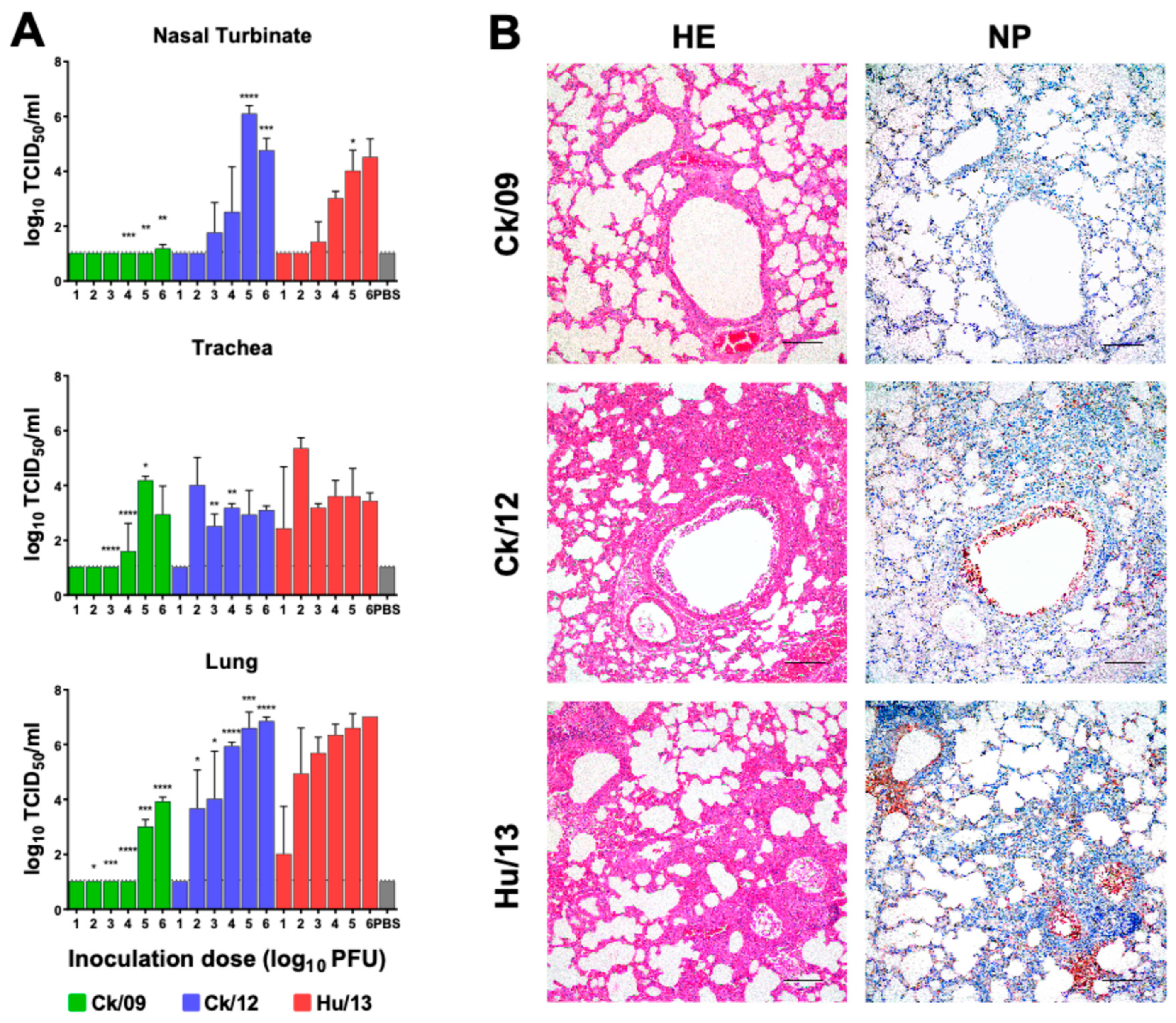
Figure 4.
Transcriptional expression of cytokines/chemokines induced by the Taiwan H6N1 influenza viruses in the lungs of mice. Lung lobes of mice inoculated with PBS (grey), Ck/09 (green), Ck/12 (blue) or Hu/13 (red) were harvested at 4 dpi (n=3 per inoculation dose). RNA expression levels of INFβ (A), IL-6 (B), IL-12P40 (C), CXCL9 (D), CXCL10 (E), CXCL11 (F), MCP-1 (G), MIP-1α (H) and TNFα (I) were measure using real-time PCR. Fold changes over the PBS inoculated group (mock controls) are shown, with the x axis indicated the inoculation doses (log10 PFU) of each virus. Results are presented as mean ± SD. Statistical significance is denoted as *P<0.05; **P<0.01; ***P<0.001; ****P<0.0001. Comparisons between Ck/09 and Hu/13 are shown above the Ck/09 groups; those between Ck/09 and Ck/12 are indicated above the Ck/12 groups; and comparisons between Ck/12 and Hu/13 are labeled above the Hu/13 groups.
Figure 4.
Transcriptional expression of cytokines/chemokines induced by the Taiwan H6N1 influenza viruses in the lungs of mice. Lung lobes of mice inoculated with PBS (grey), Ck/09 (green), Ck/12 (blue) or Hu/13 (red) were harvested at 4 dpi (n=3 per inoculation dose). RNA expression levels of INFβ (A), IL-6 (B), IL-12P40 (C), CXCL9 (D), CXCL10 (E), CXCL11 (F), MCP-1 (G), MIP-1α (H) and TNFα (I) were measure using real-time PCR. Fold changes over the PBS inoculated group (mock controls) are shown, with the x axis indicated the inoculation doses (log10 PFU) of each virus. Results are presented as mean ± SD. Statistical significance is denoted as *P<0.05; **P<0.01; ***P<0.001; ****P<0.0001. Comparisons between Ck/09 and Hu/13 are shown above the Ck/09 groups; those between Ck/09 and Ck/12 are indicated above the Ck/12 groups; and comparisons between Ck/12 and Hu/13 are labeled above the Hu/13 groups.
Figure 5.
Infection and transmission of Taiwan H6N1 influenza viruses in ferrets. Groups of three ferrets were intranasally inoculated (IN, green) with 106 PFU of Ck/09 (A), Ck/12 (B) or Hu/13 (C). For each group, three direct physical-contact (DC, blue) and three airborne-contact (AC, red) ferrets were introduced at 1 dpi. Virus shedding titers in nasal washes were monitored daily for 14 days post-inoculation or post-contact (dpi/dpc). The area under the curve (AUC) of viral shedding titers was calculated for each IN ferret and plotted (D). Results are presented as mean ± SD. The lower limit of detection is indicated by the horizontal dashed line.
Figure 5.
Infection and transmission of Taiwan H6N1 influenza viruses in ferrets. Groups of three ferrets were intranasally inoculated (IN, green) with 106 PFU of Ck/09 (A), Ck/12 (B) or Hu/13 (C). For each group, three direct physical-contact (DC, blue) and three airborne-contact (AC, red) ferrets were introduced at 1 dpi. Virus shedding titers in nasal washes were monitored daily for 14 days post-inoculation or post-contact (dpi/dpc). The area under the curve (AUC) of viral shedding titers was calculated for each IN ferret and plotted (D). Results are presented as mean ± SD. The lower limit of detection is indicated by the horizontal dashed line.
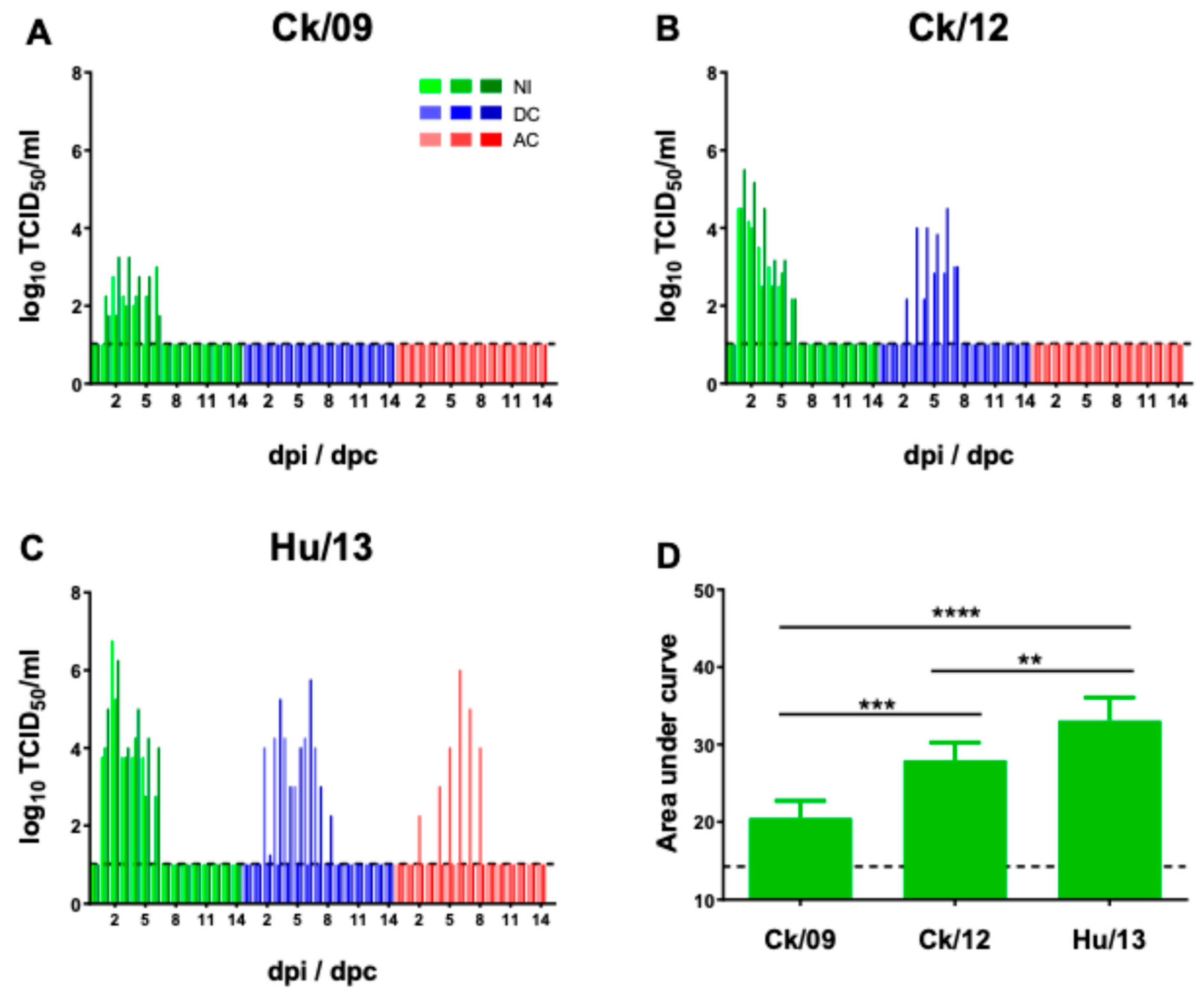
Figure 6.
Replication of Taiwan H6N1 influenza viruses in the respiratory tissues of ferrets. Groups of three ferrets intranasally inoculated with 106 PFU of Ck/09 (green), Ck/12 (blue) or Hu/13 (red) were sacrificed at 4 dpi. Viral titers in their nasal turbinate, trachea and lungs were determined by TCID50 assays (A). Results are presented as mean ± SD. Statistical significance is denoted as *P<0.05; **P<0.01; ***P<0.001; ****P<0.0001. The lower limit of detection is indicated by the horizontal dashed line. Histopathological examination of lung tissues using HE staining and IHC analysis for viral NP detection (brown staining) are shown (B). Scale bars indicate 100 μm.
Figure 6.
Replication of Taiwan H6N1 influenza viruses in the respiratory tissues of ferrets. Groups of three ferrets intranasally inoculated with 106 PFU of Ck/09 (green), Ck/12 (blue) or Hu/13 (red) were sacrificed at 4 dpi. Viral titers in their nasal turbinate, trachea and lungs were determined by TCID50 assays (A). Results are presented as mean ± SD. Statistical significance is denoted as *P<0.05; **P<0.01; ***P<0.001; ****P<0.0001. The lower limit of detection is indicated by the horizontal dashed line. Histopathological examination of lung tissues using HE staining and IHC analysis for viral NP detection (brown staining) are shown (B). Scale bars indicate 100 μm.
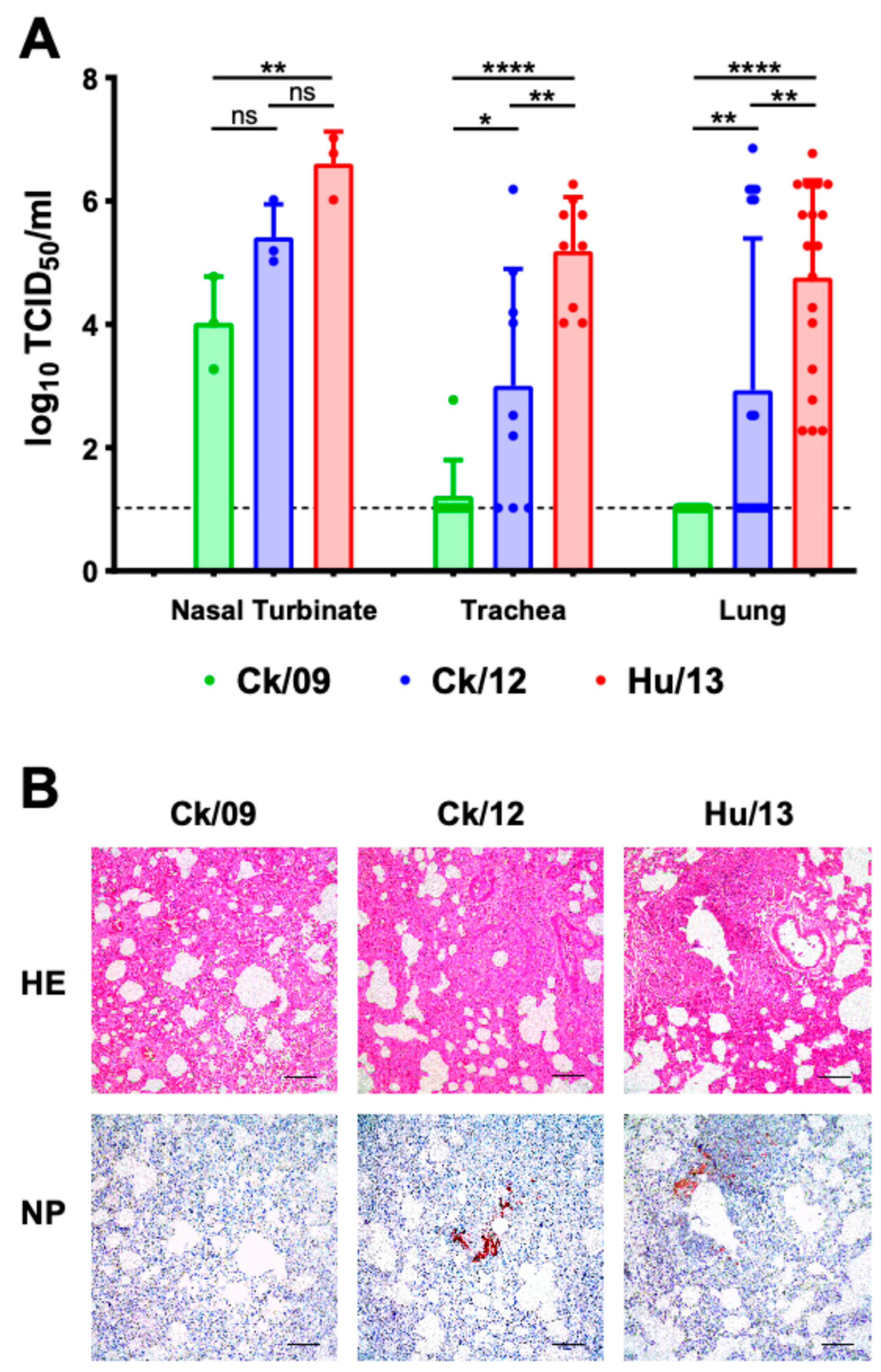
Figure 7.
Transcriptional expression of cytokines/chemokines induced by the Taiwan H6N1 influenza viruses in the lungs of ferrets. Lung lobes from ferrets inoculated with PBS (grey), Ck/09 (green), Ck/12 (blue) and Hu/13 (red) were harvested at 4 dpi (n=3 per group). RNA expression levels of IL-6 (A), IL-8 (B), IL-12P40 (C), CXCL9 (D), CXCL10 (E), CXCL11 (F), IFNα (G), INFβ (H) and IFNγ (I) were measure using real-time PCR. Fold changes over the PBS-inoculated group (mock controls) are shown. Results are presented as mean ± SD. No statistically significant differences (P > 0.05) were observed using one-way analysis of variance (ANOVA), or Student’s t-tests between any groups.
Figure 7.
Transcriptional expression of cytokines/chemokines induced by the Taiwan H6N1 influenza viruses in the lungs of ferrets. Lung lobes from ferrets inoculated with PBS (grey), Ck/09 (green), Ck/12 (blue) and Hu/13 (red) were harvested at 4 dpi (n=3 per group). RNA expression levels of IL-6 (A), IL-8 (B), IL-12P40 (C), CXCL9 (D), CXCL10 (E), CXCL11 (F), IFNα (G), INFβ (H) and IFNγ (I) were measure using real-time PCR. Fold changes over the PBS-inoculated group (mock controls) are shown. Results are presented as mean ± SD. No statistically significant differences (P > 0.05) were observed using one-way analysis of variance (ANOVA), or Student’s t-tests between any groups.

Table 1.
Infection and transmission of the Taiwan H6N1 viruses in ferrets.
| Ferret Groups | Virus | Shedding of virus | Seroconversion (HI titers at 14 dpi/dpc) |
||
|---|---|---|---|---|---|
| Animal numbers | Peak titers (log10TCID50/mL) |
Duration (days) | |||
|
Intranasally inoculated (IN) |
Ck/09 | 3/3 | 2.8, 3.0, 3.3 | 3, 6, 6 | 320 ,640, 640 |
| Ck/12 | 3/3 | 4.5, 4.5, 5.5 | 5, 6, 6 | 640, 640, 1280 | |
| Hu/13 | 3/3 | 5.3, 6.3, 6.8 | 5, 6, 6 | 1280, 1280, 1280 | |
|
Direct physical contact (DC) |
Ck/09 | 0/3 | -, -, - | -, -, - | -, -, - |
| Ck/12 | 2/3 | 3.0, 4.5, - | 4, 6, - | 640, 640, - | |
| Hu/13 | 2/3 | 4.3, 5.8, - | 6, 7, - | 640, 1280, 1280 | |
|
Airborne contact (AC) |
Ck/09 | 0/3 | -, -, - | -, -, - | -, -, - |
| Ck/12 | 0/3 | -, -, - | -, -, - | -, -, - | |
| Hu/13 | 1/3 | 6.0, -, - | 7, -, - | 1280, 1280, - | |
-, Undetectable (virus titer < 10 TCID50/ml, or HI titer < 10). .
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



