Submitted:
08 April 2025
Posted:
08 April 2025
You are already at the latest version
Abstract
Cadherins are key cell adhesion molecules that engage in extracellular homophilic binding. CDH15/M‐cadherin is localized to the apical surface of muscle satellite cells (SCs), which play a critical role in tissue regeneration after injury. Although CDH15 is considered a marker of SCs, there is no anti-CDH15 mAb suitable for flow cytometry. We developed anti-CDH15 monoclonal antibodies using the Cell-Based Immunization and Screening (CBIS) method containing a flow cytometry-based high-throughput screening. In flow cytometry, a clone Ca15Mab-1 (IgG1, κ) reacted with human CDH15-overexpressed Chinese hamster ovary-K1 (CHO/CDH15) cells. Furthermore, Ca15Mab-1 recognizes endogenous CDH15-expressing human osteosarcoma (Saos-2) and mouse myoblast (C2C12) cell lines. The dissociation constant values of Ca15Mab-1 for CHO/CDH15, Saos-2, and C2C12 were determined as 6.9 × 10⁻10 M, 8.4 × 10⁻10 M, and 1.6 × 10⁻⁹ M, respectively. Furthermore, Ca15Mab-1 can detect endogenous CDH15 in immunoblotting and immunohistochemistry. Ca15Mab-1, established by the CBIS method, is versatile for basic research and is expected to contribute to clinical studies such as antibody therapy.
Keywords:
CDH15
; M‐cadherin
; monoclonal antibody
; Cell‐Based Immunization and Screening
; flow cytometry
; immunohistochemistry
1. Introduction
Cadherins are a key class of cell adhesion molecules that engage in extracellular homophilic binding [1]. The epithelial cell-cell adhesion molecule cadherin 1 (CDH1/E-cadherin) forms the adherens junctions and plays a critical role in tumor development [2,3]. CDH1 is a classical cadherin with five extracellular cadherin repeats, a single transmembrane domain, and a cytoplasmic domain. Currently, the cadherin family has more than 100 members in humans, including classical cadherins, protocadherins, and cadherin-related proteins [4]. Phylogenetic analysis showed that classical cadherins include several distinct types. CDH1, CDH2/N (neuronal)-cadherin, CDH3/P (placental)-cadherin, CDH4/R (retinal)-cadherin, and CDH15/M (muscle)-cadherin are classified into the type I cadherin [5]. The intracellular domains of cadherins directly interact with β- and γ-catenin (also known as plakoglobin), which subsequently associate with α-catenin. α-Catenin connects to the F-actin cytoskeleton, contributing to cortical tension and the dynamic stability of cell-cell junctions [6,7].
CDH2 and CDH15 are expressed during skeletal muscle development [8]. CDH2 is present throughout muscle lineage development from uncommitted progenitor cells to proliferative myoblasts and nascent myofibers [9]. Although its expression decreases in adult myofibers, it is strongly re-expressed during muscle regeneration, following a pattern similar to that seen in development [9]. CDH15 was initially identified in mouse myoblast C2C12 [10], and the expression starts at the myoblast stage and is primarily restricted to the skeletal muscle lineage [11]. In adult muscle, CDH15 is localized to the apical satellite cell (SC) membrane, which is in direct contact with the myofiber plasma membrane [12,13].
Skeletal muscle SCs are responsible for the regeneration of skeletal muscle [14,15]. SCs typically remain in a long-term quiescent state and express PAX7, a transcription factor essential for maintaining this state [16]. Upon muscle injury, SCs become activated and initiate the expression of myogenic transcription factors MYOD and MYF5, which promotes proliferation as transit-amplifying myoblasts [17,18,19]. These myoblasts then differentiate and fuse with each other and with pre-existing myofibers to facilitate tissue repair [17,18,19]. A portion of SCs undergoes self-renewal, allowing muscles to regenerate through multiple cycles of SC-dependent repair [17,18,19]. However, when SC quiescence is abnormally disrupted, such as in aging, it often results in the depletion of the functional SC pool and compromised regenerative capacity [20,21,22,23].
Both CDH2 and CDH15 are believed to be involved in lineage determination, myoblast differentiation, and myoblast fusion [8]. However, CDH15-knockout (KO) mice displayed no noticeable defects in muscle development or regeneration [24]. CDH2-KO mice did not survive beyond embryonic day 10. However, cultured somites from the CDH2-KO embryos still formed elongated cells that expressed muscle markers and maintained intact adherens junctions [25]. These results suggest that neither CDH2 nor CDH15 is strictly required for myogenesis, and they may serve redundant roles in muscle development and regeneration. Therefore, the conditional depletion of CDH2 and CDH15 in SCs was performed. The CDH2 and CDH15 double KO SCs exhibited a state resembling an early stage of SC activation, suggesting that CDH2 and CDH15-mediated adhesion to neighboring cells mediates the SC quiescence [26]. Since the CDH15 expression is restricted in SC, a monoclonal antibody (mAb) against CDH15 is thought to help identify or isolate SC cells from muscle. Although several anti-CDH15 mAbs have been developed for immunoblotting [27] and immunofluorescence [12], there is no anti-CDH15 mAb suitable for flow cytometry.
We have developed various mAbs against membrane proteins such as receptor tyrosine kinases [28,29,30] and chemokine receptors [31,32,33] using the Cell-Based Immunization and Screening (CBIS) method. The CBIS method includes immunizing antigen-overexpressed cells and flow cytometry-based high-throughput screening. Therefore, mAbs obtained by the CBIS method tend to recognize conformational epitopes and are suitable for flow cytometry. Furthermore, some of these mAbs also apply to immunoblotting and immunohistochemistry (IHC). This study employed the CBIS method to generate highly versatile anti-CDH15 mAbs.
2. Materials and Methods
2.1. Cell Lines
Chinese hamster ovary (CHO)-K1, mouse myeloma P3X63Ag8.U1 (P3U1), human glioblastoma LN229, human osteosarcoma Saos-2, and mouse myoblast C2C12 were obtained from American Type Culture Collection (ATCC, Manassas, VA, USA). These cell lines were cultured as described previously [31].
2.2. Plasmid Construction and Establishment of Stable Transfectants
The genes encoding human CDH1 (NM_004360) and CDH3 (NM_001793.6) cloned into pCMV6neo-myc-DDK vector were purchased from OriGene Technologies, Inc. (Rockville, MD). The genes encoding human CDH2 (NM_001792) and CDH15 (NM_004933) were obtained from the RIKEN BioResource Research Center (Ibaraki, Japan). The gene encoding human CDH4 (NM_001794.5) was synthesized by Eurofins Genomics KK (Tokyo, Japan). The signal sequence-deleted CDH2, CDH4, and CDH15 cDNAs with an N-terminal PA16-tag [34] were subcloned into the pCAG-Ble vector (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan). The signal sequence-deleted CDH15 cDNAs with an N-terminal MAP16 tag [35] was subcloned into the pCAG-Ble vector. Using the Neon transfection system, these plasmids were transfected into LN229 or CHO-K1 cells (Thermo Fisher Scientific, Inc., Waltham, MA, USA). The transfectants were sorted using an anti-CDH1 mAb (clone DECMA-1, BioLegend, San Diego, CA, USA), an anti-CDH3 mAb (clone MM0508-9V11, Abcam, Cambridge, UK), an anti-PA16-tag mAb (clone NZ-1) [34], and an anti-MAP16 tag mAb (PMab-1) [35] using a SH800 cell sorter (Sony Corporation, Tokyo, Japan). The transfectants were maintained in a medium containing 0.5 mg/mL of Zeocin (InvivoGen, San Diego, CA, USA) or 0.5 mg/mL of G418 (Nacalai Tesque, Inc., Kyoto, Japan). Finally, we established the CDH-overexpressed stable transfectants: CHO/CDH1, CHO/PA16-CDH2 (CHO/CDH2), CHO/CDH3, CHO/PA16-CDH4 (CHO/CDH4), CHO/PA16-CDH15 (CHO/CDH15), and LN229/MAP16-CDH15.
2.3. Development of Hybridomas
The female BALB/cAJcl mice were purchased from CLEA Japan (Tokyo, Japan). The mice were intraperitoneally immunized with LN229/MAP16-CDH15 cells (1 × 108 cells/injection) and Alhydrogel adjuvant 2% (InvivoGen). After three additional immunizations per week (1.5 to 2.0 × 108 cells/injection), a booster injection (2.0 × 108 cells/injection) was administered two days before harvesting the spleen cells from immunized mice. The hybridomas were generated as described previously [31]. The supernatants, which were positive for CHO/CDH15 and negative for CHO-K1, were screened by an SA3800 Cell Analyzer (Sony Corporation, Tokyo, Japan).
2.4. Flow Cytometry
Cells were washed with 0.1% bovine serum albumin (BSA) in phosphate-buffered saline (PBS, blocking buffer) and treated with mAbs for 30 minutes at 4°C. The cells were then stained with anti-mouse IgG or anti-rat IgG conjugated with Alexa Fluor 488 (2,000-fold dilution; Cell Signaling Technology, Inc., Danvers, MA, USA) for 30 minutes at 4°C. The data were collected using an SA3800 Cell Analyzer and analyzed using FlowJo software (BD Biosciences, Franklin Lakes, NJ, USA).
2.5. Determination of Dissociation Constant Values Using Flow Cytometry
CHO/CDH15, Saos-2, and C2C12 were treated with serially diluted Ca15Mab-1. Subsequently, the cells were treated with anti-mouse IgG-conjugated with Alexa Fluor 488 (200-fold dilution) for 30 minutes at 4°C. The data were collected and analyzed as described previously [31].
2.6. Immunoblotting
Whole-cell lysates (10 μg of protein) were separated into polyacrylamide gels and transferred onto polyvinylidene difluoride membranes (Merck KGaA, Darmstadt, Germany). The membranes were blocked with 4% skim milk (Nacalai Tesque, Inc.) in PBS containing 0.05% Tween 20 and incubated with 1 or 10 μg/mL of Ca15Mab-1, 1 μg/mL of NZ-1, or 1 μg/mL of an anti-β-actin mAb (clone AC-15; Sigma-Aldrich Corporation, St. Louis, MO, USA). Then, the membranes were incubated with anti-mouse IgG or anti-rat IgG conjugated with horseradish peroxidase (1,000-fold dilution; Agilent Technologies, Inc., Santa Clara, CA, USA). Chemiluminescence signals were developed and detected as described previously [29].
2.7. Immunohistochemistry (IHC) USING CELL BLOCKS
Cells were fixed with 4% paraformaldehyde, and the cell blocks were prepared using iPGell (Genostaff Co., Ltd., Tokyo, Japan) (FUJIFILM Wako Pure Chemical Corporation). The formalin-fixed paraffin-embedded (FFPE) cell sections were stained with Ca15Mab-1 (0.1 or 1 μg/mL) or NZ-9, which was produced using the VH of NZ-1, CH1, CH2 and CH3 of mouse IgG2a, and the light chain of NZ-1, (1 μg/mL) using BenchMark ULTRA PLUS with the ultraView Universal DAB Detection Kit (Roche Diagnostics, Indianapolis, IN, USA).
3. Results
3.1. Development of Anti-CDH15 mAbs
Two BALB/cAJcl mice were immunized with LN229/MAP16-CDH15 cells, and the generated hybridomas were seeded into 96-well plates. After forming colonies, supernatants were subject to flow cytometry-based high throughput screening to identify supernatants that were positive for CHO/CDH15 and negative for CHO-K1. Subsequently, anti-CDH15 mAb-producing hybridomas were cloned by limiting dilution. Finally, clones Ca15Mab-1 (IgG1, κ), Ca15Mab-2 (IgG1, κ), Ca15Mab-3 (IgG1, κ), Ca15Mab-5 (IgG1, κ), and Ca15Mab-8 (IgG1, κ) were established (Figure 1).
3.2 Flow Cytometry Using Ca15Mabs
We conducted flow cytometry using the Ca15Mabs against CHO/CDH15 and CHO-K1 cells. The Ca15Mabs recognized CHO/CDH15 in a dose-dependent manner (supplementary Figure 1) from 10 to 0.01 μg/mL but did not recognize CHO-K1 even at 10 μg/mL (supplementary Figure 2). Since Ca15Mab-1 exhibits a superior reactivity among Ca15Mabs (supplementary Figure 1 and Figure 2A), we next investigated the reactivity of Ca15Mab-1 against endogenous CDH15-expressing cell lines, human osteosarcoma Saos-2 and mouse myoblast C2C12. Ca15Mab-1 showed a dose-dependent reactivity to Saos-2 and C2C12 (Figure 2B). Other Ca15Mabs also reacted with Saos-2 (supplementary Figure 3) and C2C12 (supplementary Figure 4). These results suggest that Ca15Mab-1 recognizes human and mouse CDH15 in flow cytometry.
3.3. Specificity of Ca15Mabs to Type I Cadherin-Expressed CHO-K1
CDH15 is classified into type I cadherin, which includes CDH1/E-cadherin, CDH2/N-cadherin, CDH3/P-cadherin, CDH4/R-cadherin, and CDH15/M-cadherin [5] We established each type I cadherin-expressed CHO-K1, and the specificity of Ca15Mabs to type I cadherins was determined. As shown in Figure 3, Ca15Mab-1 recognized CHO/CDH15, and did not react with other type I cadherins-expressed CHO-K1. Other Ca15Mabs also showed the same reactivity as Ca15Mab-1 (supplementary Figure 5). These results indicate that Ca15Mab-1 is a specific mAb against CDH15 among type I cadherins.
3.4. Determination of KD Values of Ca15Mab-1 by Flow Cytometry
We performed flow cytometry to determine the binding affinity of Ca15Mab-1. The KD values of Ca15Mab-1 for CHO/CDH15, Saos-2, and C2C12 were 6.9 × 10⁻10 M, 8.4 × 10⁻10 M, and 1.6× 10⁻⁹ M, respectively (Figure 4). These results showed that Ca15Mab-1 possesses high affinity to CDH15-positive cells.
3.5. Immunoblotting Using Ca15Mab-1
We next examined whether Ca15Mab-1 is suitable for immunoblotting. Whole-cell lysates of CHO-K1, CHO/CDH15, Saos-2, and C2C12 were used. Ca15Mab-1 (1 µg/mL) showed a clear band around 110 kDa in CHO/CDH15 but not in CHO-K1 (Figure 5A). An anti-PA-tag mAb (NZ-1) also detected a similar band (Figure 5B). Furthermore, Ca15Mab-1 could detect endogenous CDH15 of C2C12 at slightly higher molecular weight compared to that of CHO/CDH15 (Figure 5A). We could also detect a band in Saos-2 using a higher concentration of Ca15Mab-1 (10 µg/mL, supplementary Figure 6). An anti-β-actin mAb (AC-15) was used as an internal control (Figure 5C). There results indicate that Ca15Mab-1 can detect exogenous and endogenous CDH15 in immunoblotting.
3.6. IHC Using Ca15Mab-1 in Formalin-Fixed Paraffin-Embedded Cell Blocks
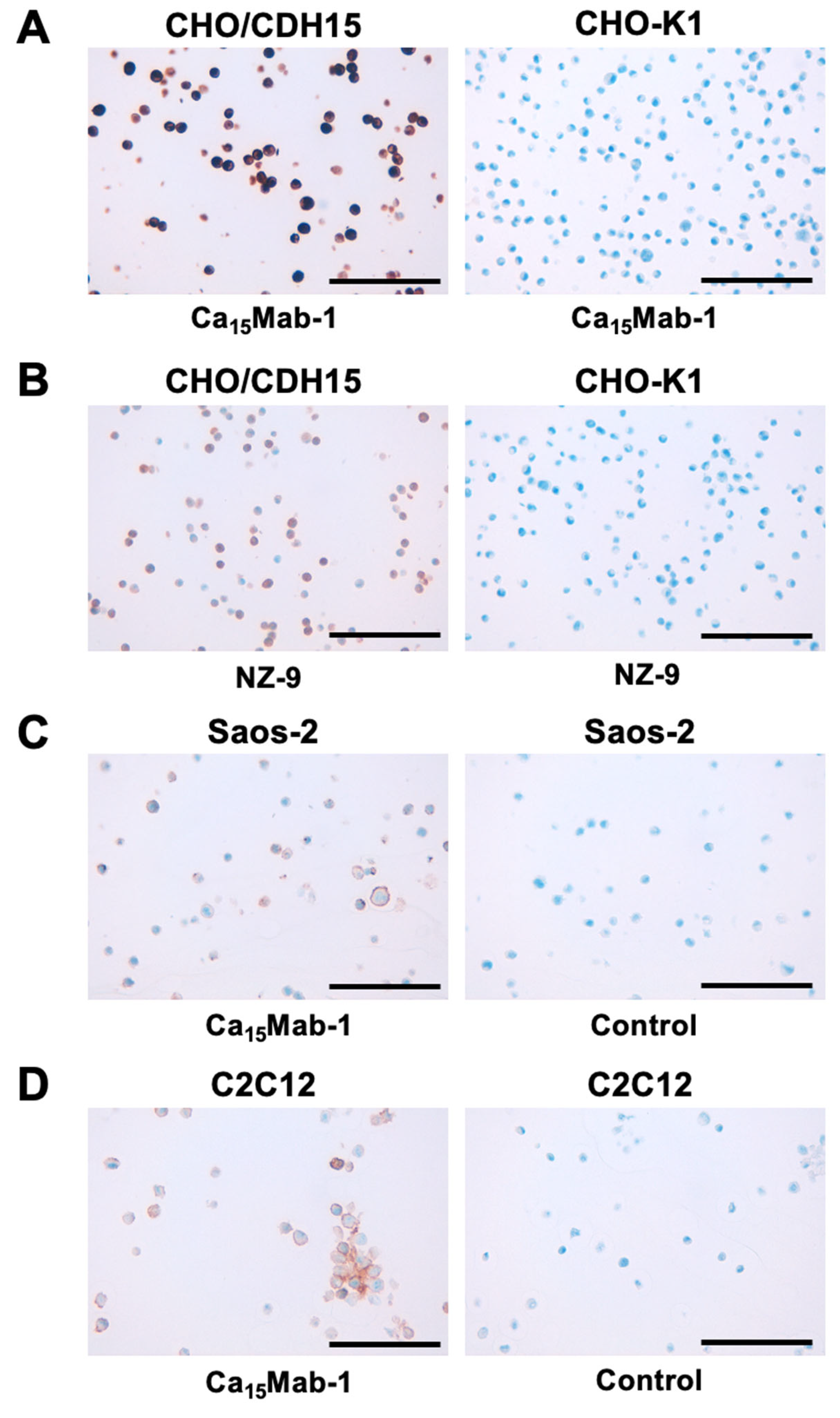
We investigated whether Ca15Mab-1 suits the IHC of FFPE sections of CHO-K1, CHO/CDH15, Saos-2, and C2C12. Both intense membranous and cytoplasmic staining by Ca15Mab-1 were detected in CHO/CDH15 but not in CHO-K1 (Figure 6A). A rat-mouse anti-PA16-tag mAb, NZ-9 also reacted with CHO/CDH15, but not CHO-K1 (Figure 6B). Furthermore, membranous staining by Ca15Mab-1 was observed in Saos-2 (Figure 6C) and C2C12 (Figure 6D). These results indicate that Ca15Mab-1 can detect exogenous and endogenous CDH15 in FFPE cell samples.
4. Discussion
The isolation of purified stem cell populations has revolutionarily changed the field of stem cell biology and become a key approach in various tissues and organisms. Skeletal muscle SCs are the most extensively studied stem cells in mammalian systems, and their targeted isolation has enabled cellular and molecular analyses. Quiescent muscle SCs are initially defined by their anatomical position to the myofiber [36]. They can be identified by the molecular markers, such as PAX7, CDH15, syndecan 3, syndecan 4, α7β1 integrin, VCAM-1, caveolin 1, and CXC chemokine receptor 4 (CXCR4) [18,37].
Studies have reported the procedures for isolating SCs from the limb muscles using fluorescence-activated cell sorting. Several mAbs against VCAM-1 [38], α7 integrin [39], and β1 integrin/CXCR4 [40] have been used for the isolation of SCs. Since these cell surface markers are expressed in other cell types, such as vascular endothelial cells and hematopoietic cells, the negative selection using anti-CD31 and/or anti-CD45 mAbs is also essential [37]. However, these protocols still have the possibility of isolating vascular endothelial cells, mesenchymal stem cells, and hematopoietic cells from muscle tissue [37]. Since the expression of CDH15 is limited in SCs compared to other markers [12,13], the isolation of CDH15-positive cells is thought to be essential for SC research. However, the isolation of CDH15-positive SCs has not been conducted, probably due to the lack of anti-CDH15 mAb for flow cytometry.
This study first reported the anti-CDH15 mAbs for flow cytometry using the CBIS method (Figure 1). An anti-CDH15 mAb, Ca15Mab-1 recognized exogenous and endogenous CDH15 in flow cytometry with high affinity (Figure 2-4). Furthermore, Ca15Mab-1 is suitable for immunoblotting (Figure 5) and IHC (Figure 6). Therefore, Ca15Mab-1 is highly versatile for basic research and would contribute to the SC biology in muscles.
Ca15Mab-1 also recognized mouse myoblast C2C12 in flow cytometry (Figure 2 and Figure 4), immunoblotting (Figure 5), and IHC (Figure 6). Cdh15 was first isolated from C2C12 cDNA [10]. C2C12 has been used in skeletal muscle research as a cell culture model [41]. C2C12 cells show potent proliferation and differentiation abilities. In low serum conditions, the cells undergo differentiation into myotubes and myofibers with cell fusion and aggregation of nuclei [42]. C2C12 has been used in various studies, such as myotube atrophy in a cancer cachexia model [43,44]. Therefore, Ca15Mab-1 will contribute to the research of myotubes and myofibers differentiation in the future.
During aging, the muscle regenerative ability is hampered due to impaired SC function [45]. A notable reduction in the number of PAX7-positive SCs has been reported in aged skeletal muscle under homeostatic conditions [46,47]. An age-dependent change in the SC niche results in the loss of ability to retain the quiescent state [23]. In several mouse models, the effects of the genetic depletion of SCs have been reported. Depletion of SCs using the Pax7CreER/+; Rosa26DTA/+ mice (DTA: diphtheria toxin A) in young adult mice impairs regeneration throughout the rest of their lives. However, the size of SC-depleted muscles was standard despite low regenerative capacity, but the muscle had increased fibrosis [48]. These results suggest that lifelong depletion of SCs did not accelerate nor exacerbate sarcopenia, the age-associated loss of skeletal muscle mass and strength. SCs were not essential to maintaining muscle size or fiber type composition during aging in mice.
A study reported that depletion of SCs suppressed the progression of muscular dystrophy. The Sgcd−/− mouse is a model of limb-girdle muscular dystrophy type 2F, devastating and life-limiting muscular dystrophy with no cure [49]. Genetic depletion of SCs in Sgcd−/− mice reduced the features of muscular dystrophy, including improved histopathology, increased sarcolemmal stability, and augmented muscle performance [50]. Mechanistically, SC-mediated induction of the fetal genes leads to sarcolemmal instability in myofibers during muscular dystrophy [50]. Since the depletion of SCs was achieved through a complicated genetic background (Sgcd−/−; Mapk3−/−; Mapk1f/f-Pax7Cre-ER), a simple depletion of SCs using mAbs is thought to be essential to obtain a proof of concept for the therapy of muscular dystrophy. Since CDH15 expression is localized to SCs [12,13], CDH15 is thought to be an essential target. We previously changed the isotype of mAbs into mouse IgG2a or human IgG1 to obtain antibody-dependent cellular cytotoxicity (ADCC), which is mainly used for the evaluation of antitumor activities in mouse xenograft models [51,52]. Since the subclass of Ca15Mab-1 is mouse IgG1, it does not exert ADCC. A class-switched Ca15Mab-1 will be useful in investigating the effect of depletion of SCs in the mouse model of muscular dystrophy.
The CDH15 expression in cancers has not been investigated. Ca15Mab-1 recognized osteosarcoma Saos-2 in flow cytometry (Figure 2 and Figure 4), immunoblotting (supplementary Figure 6), and IHC (Figure 6. Since we have not investigated other types of sarcomas such as rhabdomyosarcoma, it is worthwhile to find more CDH15-expressing tumors. Ca15Mab-1 and derivatives would contribute to cancer cell biology and cancer therapy in the future.
Author Contributions
Rena Ubukata: Investigation, Hiroyuki Suzuki: Writing – original draft, Funding acquisition, Tomohiro Tanaka: Investigation, Funding acquisition, Mika K. Kaneko: Conceptualization, Yukinari Kato: Conceptualization, Funding acquisition, Project administration, Writing – review and editing, All authors have read and agreed to the published version of the manuscript
Funding
This research was supported in part by Japan Agency for Medical Research and Development (AMED) under Grant Numbers: JP24am0521010 (to Y.K.), JP24ama121008 (to Y.K.), JP23am0401013 (to Y.K.), JP24ama221339 (to Y.K.), JP24bm1123027 (to Y.K.), and JP24ck0106730 (to Y.K.), and by the Japan Society for the Promotion of Science (JSPS) Grants-in-Aid for Scientific Research (KAKENHI) grant nos. 22K06995 (to H.S), 24K18268 (to T.T.), and 22K07224 (to Y.K.).
Institutional Review Board Statement
The animal study protocol was approved by the Animal Care and Use Committee of Tohoku University (Permit number: 2022MdA-001) for studies involving animals.
Informed Consent Statement
Not applicable.
Data Availability Statement
All related data and methods are presented in this paper. Additional inquiries should be addressed to the corresponding authors.
Conflicts of Interest
The authors declare no conflict of interest involving this article.
References
- Niessen, C.M.; Leckband, D.; Yap, A.S. Tissue organization by cadherin adhesion molecules: dynamic molecular and cellular mechanisms of morphogenetic regulation. Physiol Rev 2011, 91, 691–731. [Google Scholar] [CrossRef] [PubMed]
- van Roy, F. Beyond E-cadherin: roles of other cadherin superfamily members in cancer. Nat Rev Cancer 2014, 14, 121–134. [Google Scholar] [CrossRef] [PubMed]
- Ratheesh, A.; Yap, A.S. A bigger picture: classical cadherins and the dynamic actin cytoskeleton. Nat Rev Mol Cell Biol 2012, 13, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Hulpiau, P.; Gul, I.S.; van Roy, F. New insights into the evolution of metazoan cadherins and catenins. Prog Mol Biol Transl Sci 2013, 116, 71–94. [Google Scholar]
- Oda, H.; Takeichi, M. Evolution: structural and functional diversity of cadherin at the adherens junction. J Cell Biol 2011, 193, 1137–1146. [Google Scholar] [CrossRef]
- Lin, W.H.; Cooper, L.M.; Anastasiadis, P.Z. Cadherins and catenins in cancer: connecting cancer pathways and tumor microenvironment. Front Cell Dev Biol 2023, 11, 1137013. [Google Scholar] [CrossRef]
- Yu, W.; Yang, L.; Li, T.; Zhang, Y. Cadherin Signaling in Cancer: Its Functions and Role as a Therapeutic Target. Front Oncol 2019, 9, 989. [Google Scholar] [CrossRef]
- Krauss, R.S.; Cole, F.; Gaio, U.; et al. Close encounters: regulation of vertebrate skeletal myogenesis by cell-cell contact. J Cell Sci 2005, 118 Pt 11, 2355–2362. [Google Scholar] [CrossRef]
- Cifuentes-Diaz, C.; Nicolet, M.; Goudou, D.; Rieger, F.; Mège, R.M. N-cadherin and N-CAM-mediated adhesion in development and regeneration of skeletal muscle. Neuromuscul Disord 1993, 3, 361–365. [Google Scholar] [CrossRef]
- Donalies, M.; Cramer, M.; Ringwald, M.; Starzinski-Powitz, A. Expression of M-cadherin, a member of the cadherin multigene family, correlates with differentiation of skeletal muscle cells. Proc Natl Acad Sci U S A 1991, 88, 8024–8028. [Google Scholar] [CrossRef]
- Moore, R.; Walsh, F.S. The cell adhesion molecule M-cadherin is specifically expressed in developing and regenerating, but not denervated skeletal muscle. Development 1993, 117, 1409–1420. [Google Scholar] [CrossRef] [PubMed]
- Hung, M.; Lo, H.F.; Beckmann, A.G.; et al. Cadherin-dependent adhesion is required for muscle stem cell niche anchorage and maintenance. Development 2024, 151. [Google Scholar] [CrossRef] [PubMed]
- Irintchev, A.; Zeschnigk, M.; Starzinski-Powitz, A.; Wernig, A. Expression pattern of M-cadherin in normal, denervated, and regenerating mouse muscles. Dev Dyn 1994, 199, 326–337. [Google Scholar] [CrossRef] [PubMed]
- Dumont, N.A.; Wang, Y.X.; Rudnicki, M.A. Intrinsic and extrinsic mechanisms regulating satellite cell function. Development 2015, 142, 1572–1581. [Google Scholar] [CrossRef]
- Brack, A.S.; Rando, T.A. Tissue-specific stem cells: lessons from the skeletal muscle satellite cell. Cell Stem Cell 2012, 10, 504–514. [Google Scholar] [CrossRef]
- von Maltzahn, J.; Jones, A.E.; Parks, R.J.; Rudnicki, M.A. Pax7 is critical for the normal function of satellite cells in adult skeletal muscle. Proc Natl Acad Sci U S A 2013, 110, 16474–16479. [Google Scholar] [CrossRef]
- Relaix, F.; Bencze, M.; Borok, M.J.; et al. Perspectives on skeletal muscle stem cells. Nat Commun 2021, 12, 692. [Google Scholar] [CrossRef]
- Fukada, S.; Ma, Y.; Ohtani, T.; et al. Isolation, characterization, and molecular regulation of muscle stem cells. Front Physiol 2013, 4, 317. [Google Scholar] [CrossRef]
- Wang, Y.X.; Rudnicki, M.A. Satellite cells, the engines of muscle repair. Nat Rev Mol Cell Biol 2011, 13, 127–133. [Google Scholar] [CrossRef]
- Rozo, M.; Li, L.; Fan, C.M. Targeting β1-integrin signaling enhances regeneration in aged and dystrophic muscle in mice. Nat Med 2016, 22, 889–896. [Google Scholar] [CrossRef]
- Mourikis, P.; Sambasivan, R.; Castel, D.; et al. A critical requirement for notch signaling in maintenance of the quiescent skeletal muscle stem cell state. Stem Cells 2012, 30, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Cheung, T.H.; Quach, N.L.; Charville, G.W.; et al. Maintenance of muscle stem-cell quiescence by microRNA-489. Nature 2012, 482, 524–528. [Google Scholar] [CrossRef] [PubMed]
- Chakkalakal, J.V.; Jones, K.M.; Basson, M.A.; Brack, A.S. The aged niche disrupts muscle stem cell quiescence. Nature 2012, 490, 355–360. [Google Scholar] [CrossRef]
- Hollnagel, A.; Grund, C.; Franke, W.W.; Arnold, H.H. The cell adhesion molecule M-cadherin is not essential for muscle development and regeneration. Mol Cell Biol 2002, 22, 4760–4770. [Google Scholar] [CrossRef]
- Radice, G.L.; Rayburn, H.; Matsunami, H.; et al. Developmental defects in mouse embryos lacking N-cadherin. Dev Biol 1997, 181, 64–78. [Google Scholar] [CrossRef] [PubMed]
- Goel, A.J.; Rieder, M.K.; Arnold, H.H.; Radice, G.L.; Krauss, R.S. Niche Cadherins Control the Quiescence-to-Activation Transition in Muscle Stem Cells. Cell Rep 2017, 21, 2236–2250. [Google Scholar] [CrossRef]
- Wang, Y.; Hao, Y.; Alway, S.E. Suppression of GSK-3β activation by M-cadherin protects myoblasts against mitochondria-associated apoptosis during myogenic differentiation. J Cell Sci 2011, 124 Pt 22, 3835–3847. [Google Scholar] [CrossRef]
- Kaneko, M.K.; Suzuki, H.; Kato, Y. Establishment of a Novel Cancer-Specific Anti-HER2 Monoclonal Antibody H(2)Mab-250/H(2)CasMab-2 for Breast Cancers. Monoclon Antib Immunodiagn Immunother 2024, 43, 35–43. [Google Scholar] [CrossRef]
- Nanamiya, R.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Development of an Anti-EphB4 Monoclonal Antibody for Multiple Applications Against Breast Cancers. Monoclon Antib Immunodiagn Immunother 2023, 42, 166–177. [Google Scholar] [CrossRef]
- Itai, S.; Yamada, S.; Kaneko, M.K.; et al. Establishment of EMab-134, a Sensitive and Specific Anti-Epidermal Growth Factor Receptor Monoclonal Antibody for Detecting Squamous Cell Carcinoma Cells of the Oral Cavity. Monoclon Antib Immunodiagn Immunother 2017, 36, 272–281. [Google Scholar] [CrossRef]
- Suzuki, H.; Tanaka, T.; Li, G.; et al. Development of a Sensitive Anti-Mouse CCR5 Monoclonal Antibody for Flow Cytometry. Monoclon Antib Immunodiagn Immunother 2024, 43, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Nanamiya, R.; Takei, J.; et al. Development of Anti-Mouse CC Chemokine Receptor 8 Monoclonal Antibodies for Flow Cytometry. Monoclon Antib Immunodiagn Immunother 2021, 40, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Nanamiya, R.; Takei, J.; Asano, T.; et al. Development of Anti-Human CC Chemokine Receptor 9 Monoclonal Antibodies for Flow Cytometry. Monoclon Antib Immunodiagn Immunother 2021, 40, 101–106. [Google Scholar] [CrossRef]
- Fujii, Y.; Kaneko, M.; Neyazaki, M.; et al. PA tag: a versatile protein tagging system using a super high affinity antibody against a dodecapeptide derived from human podoplanin. Protein Expr Purif 2014, 95, 240–247. [Google Scholar] [CrossRef]
- Fujii, Y.; Kaneko, M.K.; Kato, Y. MAP Tag: A Novel Tagging System for Protein Purification and Detection. Monoclon Antib Immunodiagn Immunother 2016, 35, 293–299. [Google Scholar] [CrossRef]
- Mauro, A. Satellite cell of skeletal muscle fibers. J Biophys Biochem Cytol 1961, 9, 493–495. [Google Scholar] [CrossRef]
- Liu, L.; Cheung, T.H.; Charville, G.W.; Rando, T.A. Isolation of skeletal muscle stem cells by fluorescence-activated cell sorting. Nat Protoc 2015, 10, 1612–1624. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Cheung, T.H.; Charville, G.W.; et al. Chromatin modifications as determinants of muscle stem cell quiescence and chronological aging. Cell Rep 2013, 4, 189–204. [Google Scholar] [CrossRef]
- Joe, A.W.; Yi, L.; Natarajan, A.; et al. Muscle injury activates resident fibro/adipogenic progenitors that facilitate myogenesis. Nat Cell Biol 2010, 12, 153–163. [Google Scholar] [CrossRef]
- Sherwood, R.I.; Christensen, J.L.; Conboy, I.M.; et al. Isolation of adult mouse myogenic progenitors: functional heterogeneity of cells within and engrafting skeletal muscle. Cell 2004, 119, 543–554. [Google Scholar] [CrossRef]
- Burattini, S.; Ferri, P.; Battistelli, M.; et al. C2C12 murine myoblasts as a model of skeletal muscle development: morpho-functional characterization. Eur J Histochem 2004, 48, 223–233. [Google Scholar] [PubMed]
- Li, L.; Huang, C.; Pang, J.; et al. Advances in research on cell models for skeletal muscle atrophy. Biomed Pharmacother 2023, 167, 115517. [Google Scholar] [CrossRef]
- Hong, Y.; Lee, J.H.; Jeong, K.W.; Choi, C.S.; Jun, H.S. Amelioration of muscle wasting by glucagon-like peptide-1 receptor agonist in muscle atrophy. J Cachexia Sarcopenia Muscle 2019, 10, 903–918. [Google Scholar] [CrossRef] [PubMed]
- Bodine, S.C.; Stitt, T.N.; Gonzalez, M.; et al. Akt/mTOR pathway is a crucial regulator of skeletal muscle hypertrophy and can prevent muscle atrophy in vivo. Nat Cell Biol 2001, 3, 1014–1019. [Google Scholar] [CrossRef] [PubMed]
- Shavlakadze, T.; McGeachie, J.; Grounds, M.D. Delayed but excellent myogenic stem cell response of regenerating geriatric skeletal muscles in mice. Biogerontology 2010, 11, 363–376. [Google Scholar] [CrossRef]
- Collins, C.A.; Zammit, P.S.; Ruiz, A.P.; Morgan, J.E.; Partridge, T.A. A population of myogenic stem cells that survives skeletal muscle aging. Stem Cells 2007, 25, 885–894. [Google Scholar] [CrossRef]
- Brack, A.S.; Bildsoe, H.; Hughes, S.M. Evidence that satellite cell decrement contributes to preferential decline in nuclear number from large fibres during murine age-related muscle atrophy. J Cell Sci 2005, 118 Pt 20, 4813–4821. [Google Scholar] [CrossRef]
- Fry, C.S.; Lee, J.D.; Mula, J.; et al. Inducible depletion of satellite cells in adult, sedentary mice impairs muscle regenerative capacity without affecting sarcopenia. Nat Med 2015, 21, 76–80. [Google Scholar] [CrossRef]
- Blain, A.M.; Straub, V.W. δ-Sarcoglycan-deficient muscular dystrophy: from discovery to therapeutic approaches. Skelet Muscle 2011, 1, 13. [Google Scholar] [CrossRef]
- Boyer, J.G.; Huo, J.; Han, S.; et al. Depletion of skeletal muscle satellite cells attenuates pathology in muscular dystrophy. Nat Commun 2022, 13, 2940. [Google Scholar] [CrossRef]
- Suzuki, H.; Ohishi, T.; Tanaka, T.; Kaneko, M.K.; Kato, Y. Anti-HER2 Cancer-Specific mAb, H(2)Mab-250-hG(1), Possesses Higher Complement-Dependent Cytotoxicity than Trastuzumab. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, M.K.; Suzuki, H.; Ohishi, T.; et al. A Cancer-Specific Monoclonal Antibody against HER2 Exerts Antitumor Activities in Human Breast Cancer Xenograft Models. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic representation of anti-CDH15 mAbs production. (A) LN229/MAP16-CDH15 was injected intraperitoneally into BALB/cAJcl mice. (B) After five times immunizations per week, spleen cells were fused with P3U1. (C) The supernatants of hybridomas were screened by flow cytometry using CHO/CDH15 and CHO-K1 cells. (D) Anti-CDH15 specific mAb-producing hybridoma clones (Ca15Mabs) were established by limiting dilution.
Figure 1.
Schematic representation of anti-CDH15 mAbs production. (A) LN229/MAP16-CDH15 was injected intraperitoneally into BALB/cAJcl mice. (B) After five times immunizations per week, spleen cells were fused with P3U1. (C) The supernatants of hybridomas were screened by flow cytometry using CHO/CDH15 and CHO-K1 cells. (D) Anti-CDH15 specific mAb-producing hybridoma clones (Ca15Mabs) were established by limiting dilution.
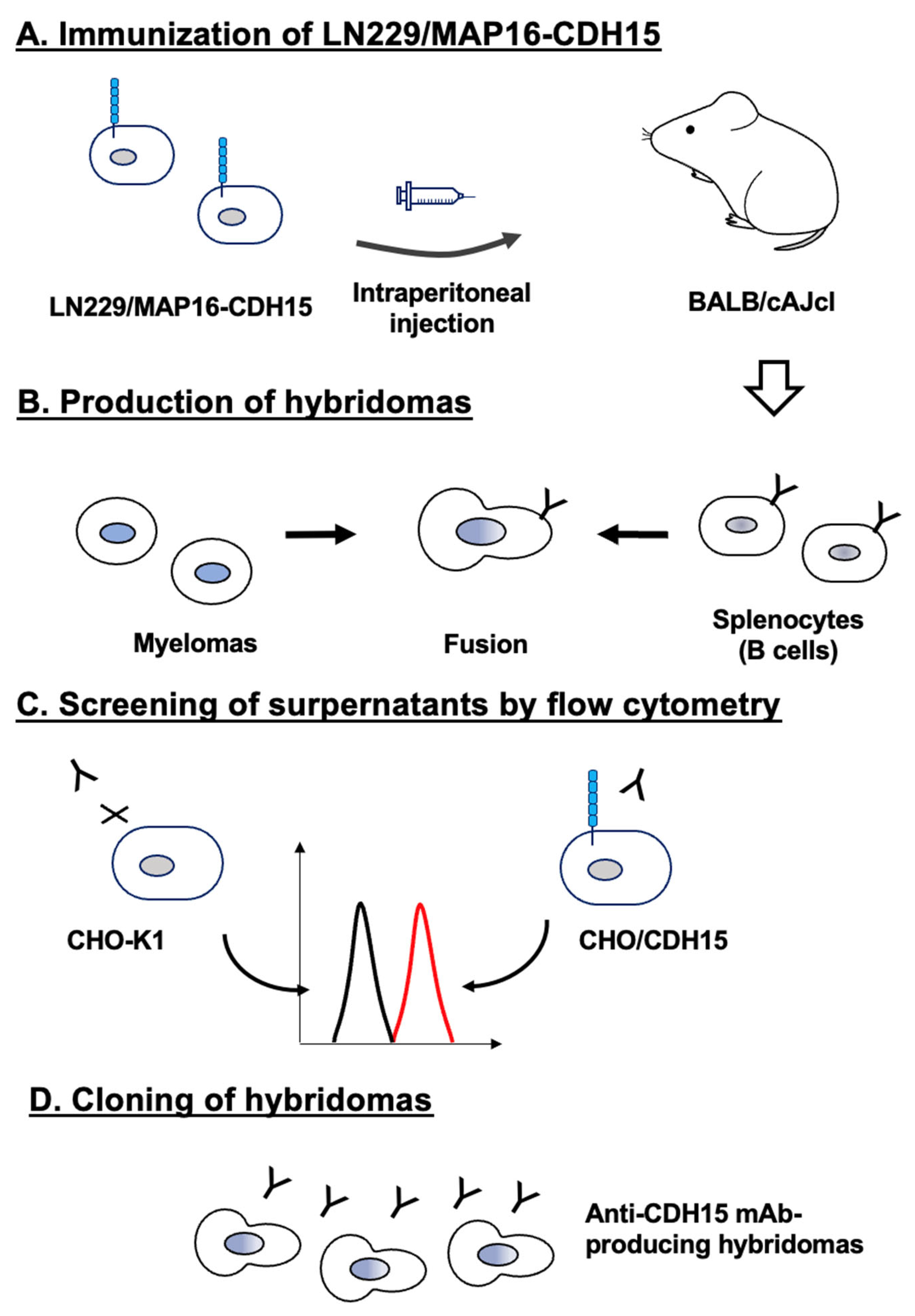
Figure 2.
Flow cytometry analysis of Ca15Mab-1 against CHO/CDH15, CHO-K1, Saos-2, and C2C12. (A) CHO/CDH15 and CHO-K1 were treated with Ca15Mab-1 at the indicated concentrations (red) or blocking buffer (black). (B) Saos-2 and C2C12 were treated with Ca15Mab-1 at the indicated concentrations (red) or blocking buffer (black). The mAbs-treated cells were incubated with anti-mouse IgG conjugated with Alexa Fluor 488. The fluorescence data were collected using the SA3800 Cell Analyzer.
Figure 2.
Flow cytometry analysis of Ca15Mab-1 against CHO/CDH15, CHO-K1, Saos-2, and C2C12. (A) CHO/CDH15 and CHO-K1 were treated with Ca15Mab-1 at the indicated concentrations (red) or blocking buffer (black). (B) Saos-2 and C2C12 were treated with Ca15Mab-1 at the indicated concentrations (red) or blocking buffer (black). The mAbs-treated cells were incubated with anti-mouse IgG conjugated with Alexa Fluor 488. The fluorescence data were collected using the SA3800 Cell Analyzer.
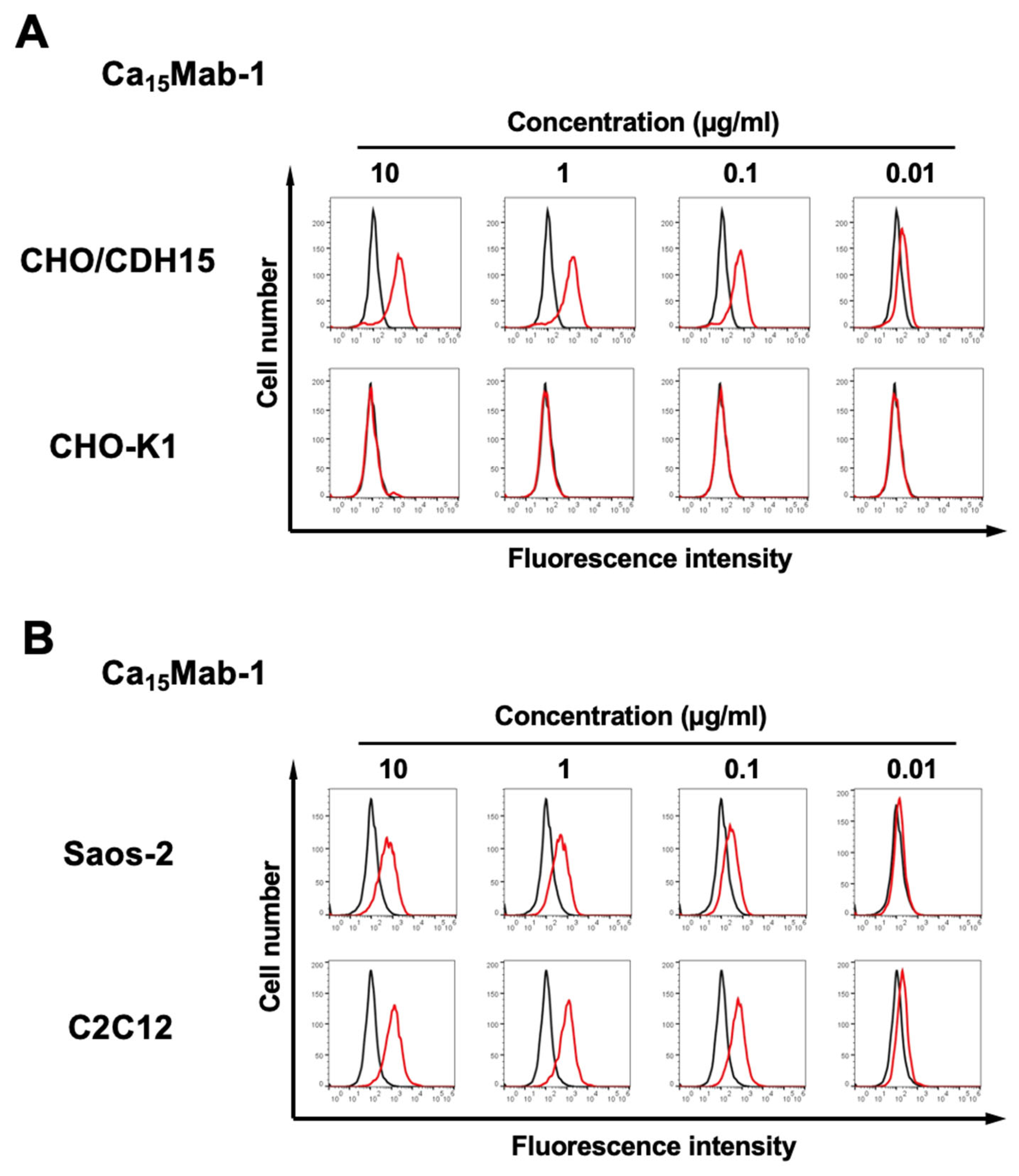
Figure 3.
Flow cytometry analysis of Ca15Mab-1 in type I cadherins-expressed CHO-K1 cells. The type I cadherins (CDH1, CDH2, CDH3, CDH4, and CDH15)-expressed CHO-K1 cells were treated with 10 µg/mL of Ca15Mab-1 (red) or control blocking buffer (black line), followed by the treatment with anti-mouse IgG conjugated with Alexa Fluor 488. Each cadherin expression was confirmed by 10 µg/mL of an anti-CDH1 mAb (clone DECMA-1), 1 µg/mL of an anti-CDH3 mAb (clone MM0508-9V11), and 1 µg/mL of an anti-PA16-tag mAb (clone NZ-1), followed by the treatment with anti-mouse IgG or anti-rat IgG conjugated with Alexa Fluor 488.
Figure 3.
Flow cytometry analysis of Ca15Mab-1 in type I cadherins-expressed CHO-K1 cells. The type I cadherins (CDH1, CDH2, CDH3, CDH4, and CDH15)-expressed CHO-K1 cells were treated with 10 µg/mL of Ca15Mab-1 (red) or control blocking buffer (black line), followed by the treatment with anti-mouse IgG conjugated with Alexa Fluor 488. Each cadherin expression was confirmed by 10 µg/mL of an anti-CDH1 mAb (clone DECMA-1), 1 µg/mL of an anti-CDH3 mAb (clone MM0508-9V11), and 1 µg/mL of an anti-PA16-tag mAb (clone NZ-1), followed by the treatment with anti-mouse IgG or anti-rat IgG conjugated with Alexa Fluor 488.
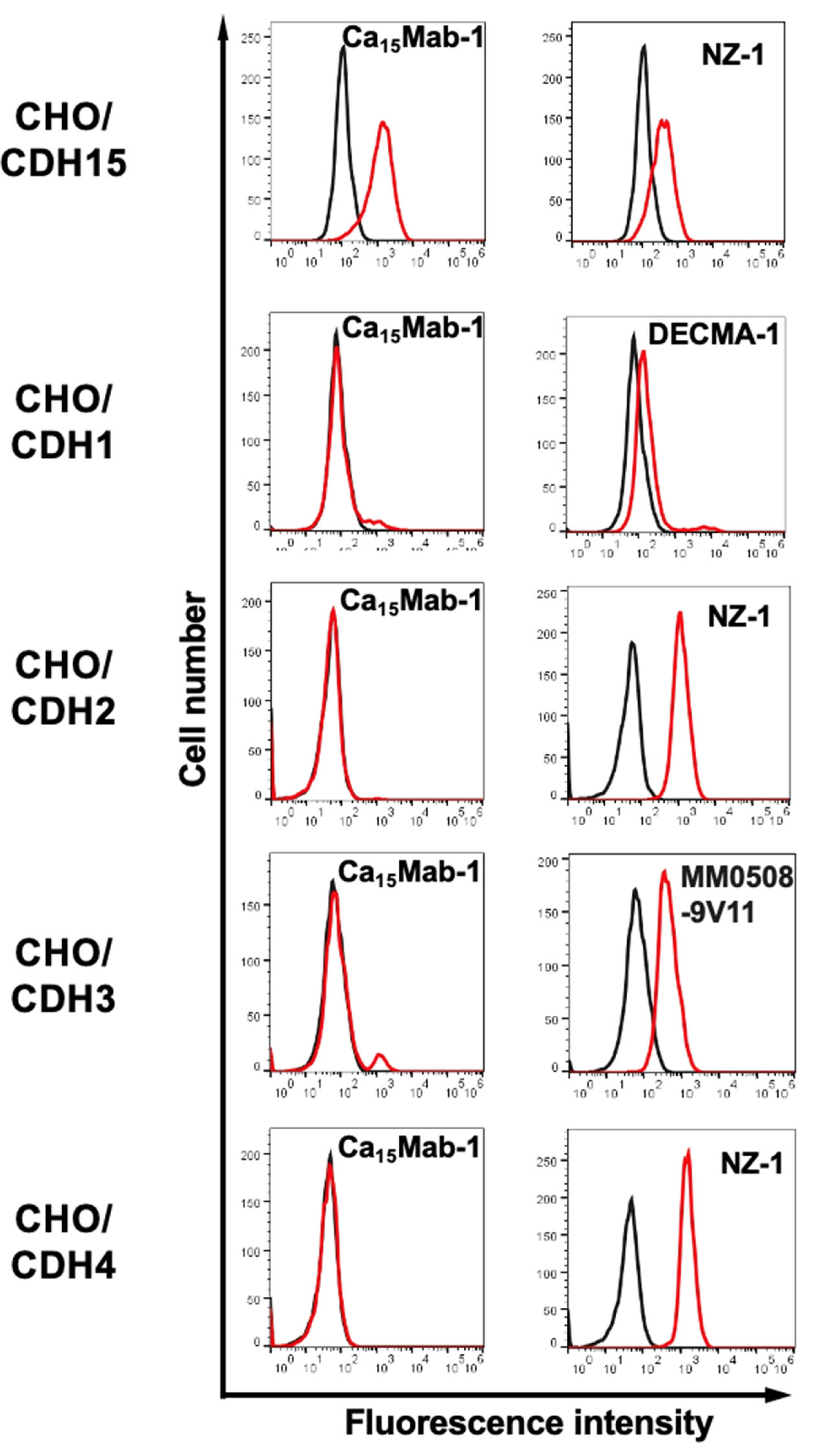
Figure 4.
Measurement of binding affinity of Ca15Mab-1. CHO/CDH15 (A), Saos-2 (B), and C2C12 (C) were treated with serially diluted Ca15Mab-1, followed by anti-mouse IgG conjugated with Alexa Fluor 488. The KD values were determined using GraphPad PRISM 6.
Figure 4.
Measurement of binding affinity of Ca15Mab-1. CHO/CDH15 (A), Saos-2 (B), and C2C12 (C) were treated with serially diluted Ca15Mab-1, followed by anti-mouse IgG conjugated with Alexa Fluor 488. The KD values were determined using GraphPad PRISM 6.
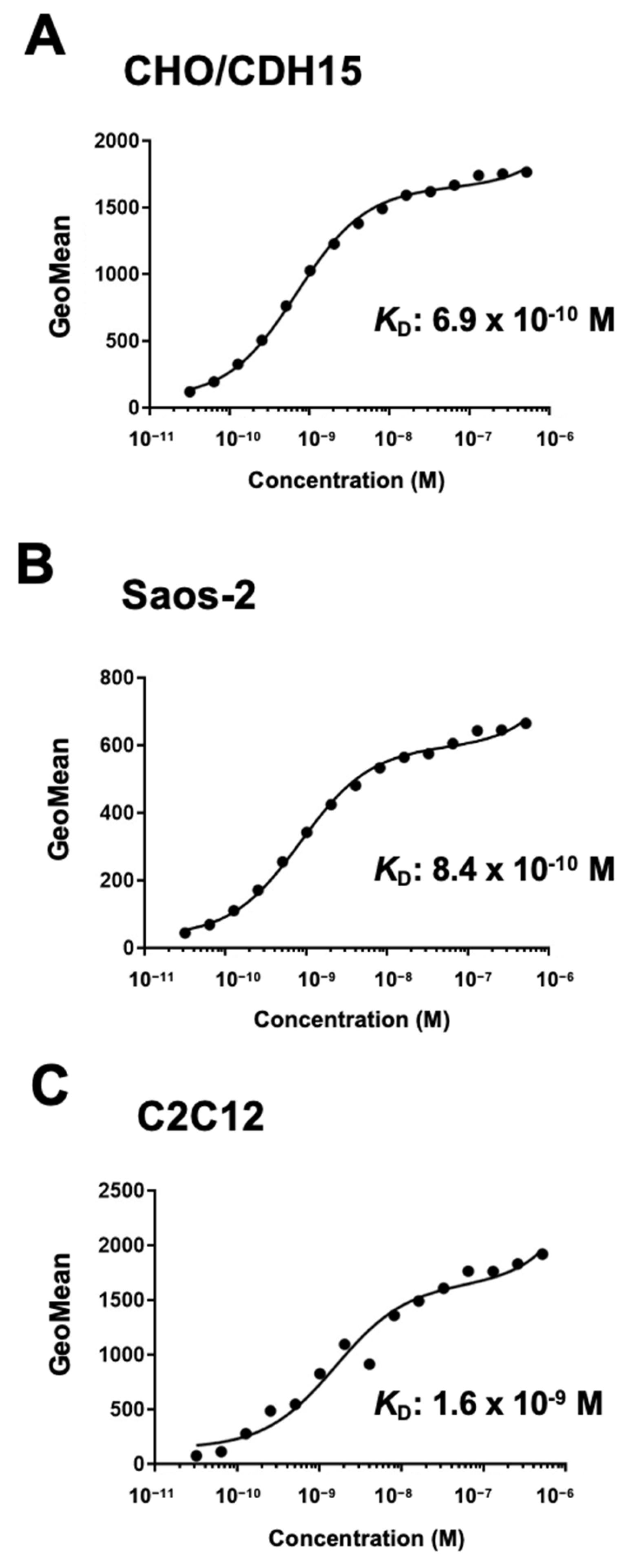
Figure 5.
Immunoblotting using Ca15Mab-1. The cell lysate (10 μg/lane) of CHO-K1, CHO/CDH15, Saos-2, and C2C12 were electrophoresed and transferred onto polyvinylidene difluoride membranes. The membranes were incubated with 1 μg/mL of Ca15Mab-1 (A), 1 μg/mL of NZ-1 (B), and 1 μg/mL of AC-15 (an anti-β-actin mAb) (C), followed by the treatment with anti-mouse or anti-rat IgG conjugated with horseradish peroxidase.
Figure 5.
Immunoblotting using Ca15Mab-1. The cell lysate (10 μg/lane) of CHO-K1, CHO/CDH15, Saos-2, and C2C12 were electrophoresed and transferred onto polyvinylidene difluoride membranes. The membranes were incubated with 1 μg/mL of Ca15Mab-1 (A), 1 μg/mL of NZ-1 (B), and 1 μg/mL of AC-15 (an anti-β-actin mAb) (C), followed by the treatment with anti-mouse or anti-rat IgG conjugated with horseradish peroxidase.
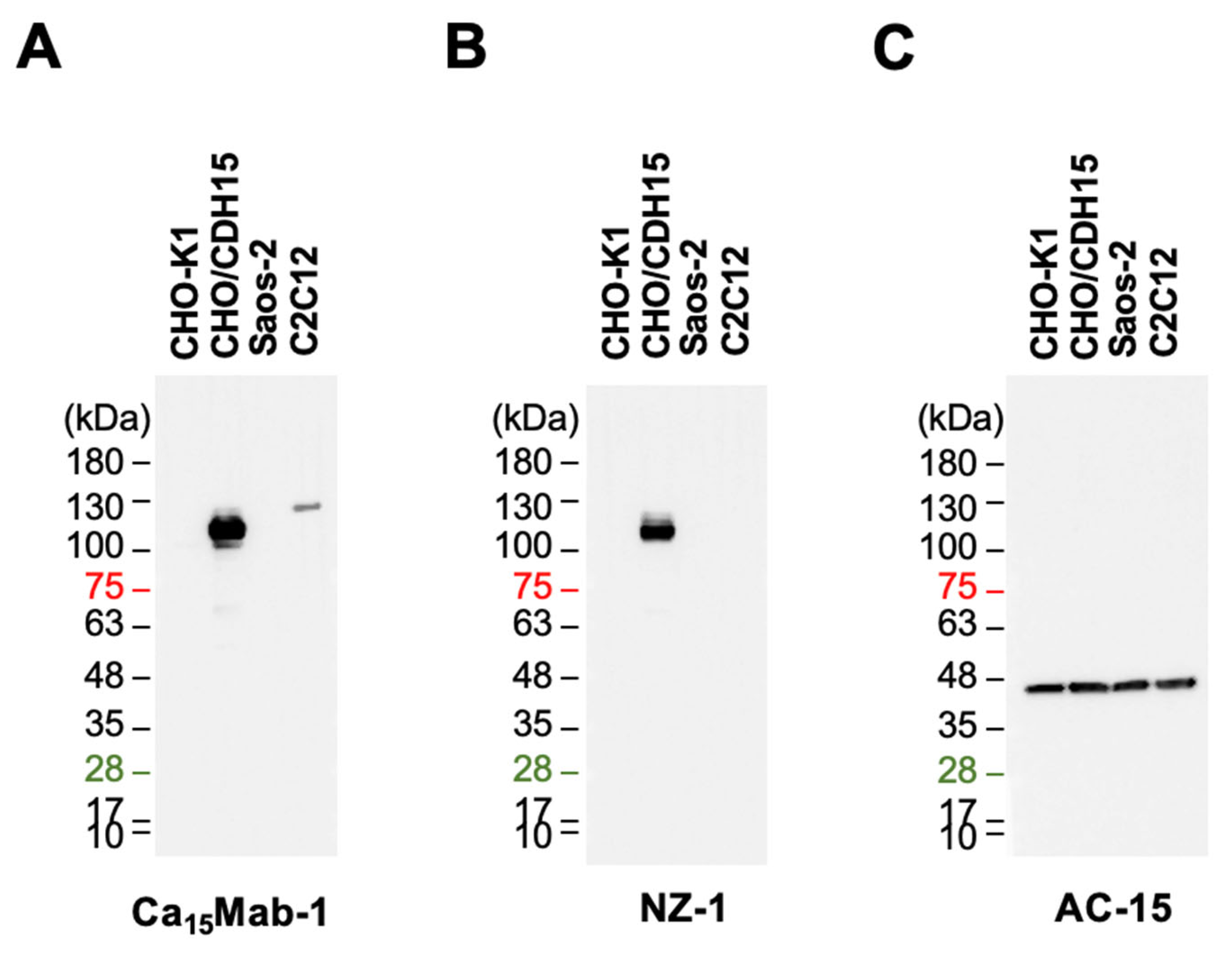
Figure 6.
Immunohistochemistry using Ca15Mab-1 in formalin-fixed paraffin-embedded cell blocks. (A) CHO/CDH15 and CHO-K1 sections were treated with 0.1 μg/mL of Ca15Mab-1. (B) CHO/CDH15 and CHO-K1 sections were treated with 1 μg/mL of NZ-9. (C) The sections of Saos-2 were treated with 1 μg/mL of Ca15Mab-1 or control (without primary Ab). (D) The sections of C2C12 were treated with 1 μg/mL of Ca15Mab-1 or control (without primary Ab). The staining was performed using BenchMark ULTRA PLUS with the ultraView Universal DAB Detection Kit. Scale bar = 100 μm.
Figure 6.
Immunohistochemistry using Ca15Mab-1 in formalin-fixed paraffin-embedded cell blocks. (A) CHO/CDH15 and CHO-K1 sections were treated with 0.1 μg/mL of Ca15Mab-1. (B) CHO/CDH15 and CHO-K1 sections were treated with 1 μg/mL of NZ-9. (C) The sections of Saos-2 were treated with 1 μg/mL of Ca15Mab-1 or control (without primary Ab). (D) The sections of C2C12 were treated with 1 μg/mL of Ca15Mab-1 or control (without primary Ab). The staining was performed using BenchMark ULTRA PLUS with the ultraView Universal DAB Detection Kit. Scale bar = 100 μm.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



