
Submitted:
02 April 2025
Posted:
03 April 2025
You are already at the latest version
Abstract
Recently, many researchers have evaluated various viruses, including polyomaviruses (JCV, BKV) and EBV, as potential factors playing a role in the development of prostatę cancer (PCa), one of the most common cancers in men. Therefore, we aimed to assess the frequency of the JCPyV DNA in tissue collected from PCa patients. We detected the presence of viral DNA in 49.6% of clinical samples, including 71.9% with single EBV infection and 28.1% with EBV/JCV co-infection. We did not detect BKV or a single JCV infection. Therefore, we compared patients with EBV monoinfection with EBV/JCV coinfected patients in the context of risk group, Gleason score and TNM classification. Our results showed differences in clinicopathological features between single EBV infection and EBV/JCV co-infection. In the group of patients with single EBV infection, most patients were classified as medium/high risk, while in the group with EBV/JCV co-infection, most patients were classified as low risk. Among patients with single EBV infection, a more advanced stage of cancer was observed than in EBV/JCV co-infection. Moreover, the level of anti-EBVCA and anti-EBNA antibodies as well as EBV load was higher in the case of single infection compared to EBV/JCV co-infection. Higher antibody levels were detected in more advanced tumor stages in single EBV infection. Does JCV only “reside” in prostate cells or is it a co-factor in EBV infection? In the light of these studies, there is a need to clarify the role of the JCV virus in the development of prostate cancer.
Keywords:
prostate cancer
; JCV
; EBV/JCV co-infection
1. Introduction
High rates of morbidity and mortality due to cancer set new research directions to meet the challenges of modern and effective diagnostics, therapy and prevention of these diseases.
Prostate cancer is one of the most common cancers in men worldwide. According to IARC data, in 2022, 1,467,854 new cases of prostate cancer and 397,430 deaths from this cause were registered [1]. Also in Poland, malignant tumors constitute an increasing public health problem. In 2022, the number of new cases was 181,300, including 89,794 among men. [2]. Prostate cancer was the most frequently registered cancer in men, accounting for 23.3%.
Due to biological and clinical diversity, the main challenge of modern medicine is the ability to make an accurate prognosis [3]. Research to understand the etiology and biology of this cancer is needed to develop preventive methods and treatment strategies.
The etiology of prostate cancer is multifactorial and not fully explained. Some researchers emphasize the role of chronic inflammation, although opinions differ here [4]. Supporters of this theory believe that inflammation plays a key role in the development of prostate cancer [5,6]. Research by Gurel et al. [7] showed that chronic prostatitis increases the risk of PCa by 30%. Prostatitis was classified as acute or chronic, both bacterial and nonbacterial [4] .
Disturbances in epithelial cells, changes in their phenotype, gene mutations as well as external factors may result in dysregulation of the prostate environment, causing inflammation [8,9].
Prostate inflammation can be caused by various factors, including microorganisms such as bacteria and viruses. Among them, sexually transmitted infections (STIs) have been investigated as an etiological factor. A meta-analysis by Caini et al. [10] showed that the risk of prostate cancer was 49% higher in men who had had any sexually transmitted disease.
It is estimated that viral infections play a role in approximately 20-25% of all human cancer cases [11,12]. The group of oncogenic viruses with a proven association with carcinogenesis includes: human papillomavirus (HPV), Epstein-Barr virus (EBV), hepatitis B virus (HBV), hepatitis C virus (HCV), human herpes virus 8 (HHV8), human T-cell lymphotropic virus type 1 (HTLV-1) and Merkel cell polyomavirus (MCPyV) [13]. Oncogenic viruses constitute a taxonomically diverse group, which causes different mechanisms of development of various cancers [14].
Recently, publications have appeared in the medical literature whose authors examine the relationship between prostate cancer and viral infections. Documented, among others: the presence of viruses such as: human papilloma virus, cytomegalovirus, human herpes simplex virus type 2, human herpes virus type 8, and Epstein-Barr virus; BKV polyomavirus [15].
Polyomavirus (PyV) was discovered accidentally in 1950 during research on the infectious agent causing rodent cancer [16].
The Polyomaviridae family includes small, non-enveloped viruses with a circular dsDNA genome consisting of three functional regions: non-coding control region (NCCR) and two coding regions: early and late. The early non-structural proteins are called tumour antigens (T-Ag), the late region usually encodes three structural proteins (VP1, VP2 and VP3). The well-studied human polyomaviruses are BK PyV and JC PyV viruses [17].
JCPyV is ubiquitous and it is estimated that 70-80% of the adult population is infected [18]. Primary infection usually appears in childhood and is asymptomatic or mild. The virus can be detected in various organs such as: gastrointestinal tract, spleen, lymph nodes, lungs, bone marrow, brain, B lymphocytes, and kidneys [19].
Various viruses, including JCV, BKV and EBV, are being investigated as potential factors playing a role in the development of PCa [20,21,22,23,24]. In vitro and in vivo studies have shown that JCPyV has the ability to transform cells [25]. The main oncogenic protein of the virus is large T antigen (T-Ag), which binds retinoblastoma proteins (pRb) and p53, leading to cell cycle dysregulation. Early proteins, small tumor antigen (t-Ag) and agnoprotein are also believed to participate in the cellular transformation process.
In our previous studies, we detected EBV DNA in the tumor tissue in 49% of patients with prostate cancer, which may suggest a role of EBV in the development of this cancer [26]. Therefore, we decided to examine prostate tumor tissue for the presence of two polyomaviruses, i.e. BKV and JCV. Our goal was also to assess possible co-infection with the studied viruses. Moreover, we compared EBV single infected and EBV/JCV co-infected patients in the context of risk group, Gleason score and TNM classification. We also compared the serological status of patients. We wanted to examine the frequency and level of anti-EBV antibodies in the serum of patients with EBV mono-infection compared to patients with EBV/JCV co-infection. We then analyzed whether co-infection had an effect on anti-EBV antibody levels as well as on EBV load.
2. Materials and Methods
2.1. Characteristics of Patients
The current study comprised 115 male patients who were diagnosed with PCa and subsequently confirmed through histopathological analysis. The study participants were hospitalized at the Department of General and Oncological Urology of the 1st Military Clinical Hospital with Out-patient Clinic in Lublin from January to November of 2023. Patients who had previously undergone chemotherapy or radiotherapy were excluded from the study. It is also worthy of note that all patients underwent radical prostatectomy. The mean age of the patients was 68.9 ± 7.4 years.
According to the European Association of Urology classification, there are three risk groups: a low-risk group, an intermediate-risk group and a high-risk group, based on three data points: prostate-specific antigen (PSA) level, Gleason score (GS) and the TNM staging system [27,28,29]. This categorization system thus distinguishes the following risk groups: the low-risk group—PSA < 10 ng/mL, GS < 7 (ISUP grade 1), and cT1-2a; the intermediate risk group—PSA 10–20 ng/mL, GS 7 (ISUP grade 2/3), or cT2b; the high-risk group—PSA > 20 ng/mL, GS > 7 (ISUP grade 4/5), or cT2c [27,28,29].
Patients with PCa were allocated to these groups according to viral infection. Subsequent detailed comparison of the two patient groups revealed no significant disparities in their demographic and social characteristics, thus indicating that the two groups were homogeneous in this regard.

Table 1.
Epidemiological characteristics of PCa patients.
| PCa Patients | |||||
| EBV- single infection | EBV/JCV co-infection | ||||
| n | % | n | % | ||
| Total | 41 | 35.7 | 16 | 13.9 | |
| Age | < 60 | 6 | 14.6 | 1 | 6.3 |
| > 60 | 35 | 85.4 | 15 | 93.8 | |
| p | 0.6599 | ||||
| Place of residence | Urban | 28 | 68.3 | 9 | 56.3 |
| Rural | 13 | 31.7 | 7 | 43.8 | |
| p | 0.5379 | ||||
| Smoking | Never | 10 | 24.4 | 2 | 12.5 |
| Ever | 31 | 75.6 | 14 | 87.5 | |
| p | 0.4767 | ||||
| Alcoholabuse | never | 14 | 34.2 | 5 | 31.3 |
| ≤ drink per week | 23 | 56.1 | 10 | 62.5 | |
| > drink per week | 4 | 9.8 | 1 | 6.3 | |
| p | 0.9999 | ||||
Pearson’s chi-squared test.
2.2. Sample Collection
Clinical materials were collected from patients suffering from PCa. This included serum, as well as fresh frozen tumor tissues. The samples were coded using a sample identification system, in order to ensure the anonymity of the patients. The materials were collected and delivered to the laboratory within 24 hours. The tissues were collected during surgical procedures, while the blood was collected in accordance with standard hospital procedures, during routine examinations. The remaining samples were submitted to our laboratory for further analysis. Centrifugation of the blood samples (1500× g/15 min) at room temperature was performed to separate the serum. The clinical material was stored at -80°C until analysis.
2.3. Isolation and Detection of EBV DNA
As previously outlined in the study by Kiś et. al [26], the samples were subjected to a series of processing steps. Fresh tumour tissues were initially frozen and then cut and homogenised using an Omni TH/Omni International/Kennesewa homogeniser (GA, USA). Subsequently, the QIAamp DNA Mini Kit (Qiagen, Hilden, Germany) was employed for the extraction of DNA, with this process being conducted in accordance with the manufacturer's protocol. The quality of the extracted DNA was then assessed through the implementation of a β-globin assay, which was utilized to both ensure the integrity of the DNA and to identify the presence of any potential PCR inhibitors. Purified DNA was quantified using Epoch Microplate Spectrophotometer (BioTek Instruments Inc., Vinooski, VT, USA). The extracted DNA was then amplified using the GeneProof Epstein-Barr virus (EBV) PCR Kit (Brno, Czech Republic) calculating the EBV DNA copy number by the ISEX version of the EBV PCR kit. All samples, including the negative control, were analyzed in duplicate. Amplification of the specific DNA sequence for EBNA1 was performed using LightCycler 2.0 software version 4.1 (Roche Applied Science System, Penzberg, Germany) during the PCR process.
2.4. Isolation and Detection BKV and JCV
The preparation of tissues for the detection of JCV and BKV was conducted in a manner consistent with the protocol established for the detection of EBV. The extraction of genetic material was undertaken using the QIAamp DNA Minikit (Qiagen, Hilden, Germany), in accordance with the manufacturer's guidelines. Following this process, the material underwent amplification via use of the GeneProof BK/JC Virus (BK/JC) PCR Kit (Brno/Czech Republic) in conjunction with the CFX96 Dx ORM (Bio-Rad, Pleasanton, CA, USA).
2.5. Antibodies Detection—Serological Methods
The test kit has been developed for the purpose of identifying IgA, IgG, and IgM antibodies in human serum, plasma, or cerebrospinal fluid, and it utilizes specific EBV antigens, including EBNA1, EBNA2, VCA p18, VCA p23, p54 Early Antigen D (EA-D p54), EA-D p138, EA-R, Rta, ZEBRA, gp85, gp350 and latent membrane proteins (LMP1). The interpretation of the result is focused upon the presence of a reaction to a minimum of one antigen (either EBNA1 or EBVCA p18). The quantitative results are expressed in units of U/mL. Negative results are indicated by values below 185 U/mL, borderline results are observed between 185 and 210 U/mL, and positive results are indicated by values above 210 U/mL. Analysis was conducted using the Microblot-Array reader with software version 2.0.4.
2.6. Statistical Analysis
The analysis of results was conducted utilizing GraphPad Prism version 10.4.1. software, a software suite developed in San Diego, California, USA. Categorical data were presented as numbers and percentages, while the Shapiro–Wilk test was employed to evaluate the distribution of continuous variables. The baseline characteristics of patients are expressed as percentages, and the Pearson's chi-squared test and Fisher's exact test were employed to compare the frequency of antibodies in both groups. Furthermore, the Mann–Whitney and Kruskal-Wallis tests were used to assess differences in antibody levels between groups. The statistical significance of the results was determined at p ≤ 0.05
2.7. Ethics
Approval for the study was granted by the Medical University of Lublin Ethics Committee, and its implementation adhered to the principles of GCP (No. KE-0254/194/10/2022, 6 October 2022). Informed consent was obtained from each participant in written form.
3. Results
3.1. Evaluation of the Frequency EBV, JCV, BKV in the Tumor Tissue of PCa Patients
The first goal of our research was to assess the frequency of the genetic material of EB, BK and JC viruses. Among the 115 patients included in the study, viral DNA was detected in 57 people (49.6%), including 41 (71.9%) with single EBV infection and 16 (28.1%) with EBV/JCV co-infection (Figure 1). However, neither BK virus nor single JCV infections were detected in any of the samples tested.
3.2. Comparison of Patients with Single EBV Infection and Patients with EBV/JCV Co-Infection in the Context of Risk Group, Gleason Score and TNM Classification
Two groups of patients were included in further analysis, i.e. 41 patients with single EBV infection and 16 patients with EBV/JCV co-infection.
Due to the too small number of patients from the medium-risk group among co-infected people, it was necessary to combine groups. Thus, two groups were created, i.e. the first low-risk group and the second medium/high-risk group (Table 2).
The Gleason score was analyzed similarly, i.e. two groups were included in the analysis: one including patients with GS 6-7 and the other with GS 8-9. The allocation of patients diagnosed with PCa to these groups was made according to the above-mentioned classification system.
In the studied patient groups, there were no T3 or T4 stages, no metastases to regional lymph nodes (N0 = 100%) or distant metastases (M0 = 100%).
The objective of the present analysis was to ascertain whether EBV/JCV co-infection has an impact on the clinical features presented in Table 2. The present study revealed differences in pathological features. In the group of patients with single EBV infection, 73.2% of patients were classified as medium and high risk, while 56.2% of patients with EBV/JCV coinfection were classified as low risk. This variability was found to be statistically significant (p = 0.0365).
Moreover, 56.1% of patients with single EBV infection were classified into group 8-9 GS, while 75.0% of patients with EBV/JCV co-infection were designated as being in the 6-7 GS group. The data indicate a significant difference in the Gleason score between the group of patients with single EBV infection and co-infection. Among patients with single EBV infection, a more advanced stage of cancer predominated. (p = 0.0365).
A similar tendency was noticed regarding tumor size (T). The T2 group constituted 73.2% of patients infected only with EBV, while patients with EBV/JCV co-infection constituted 62.5% of the T1 group. The difference between these percentages was found to be statistically significant (p = 0.0121).
3.3. Frequency of Anti EBV Antibodies in PCa Patients with Ebvsingle Infection Compared with EBV/JCV Co-Infection
In the next stage of our study, we wanted to check the frequency of anti-EBV antibodies in patients with single EBV infection compared to patients with EBV/JCV coinfection in the context of risk group, Gleason Score and tumor size (T). First, we assessed the prevalence of anti-EBV antibodies
Among the numerous antigens contained in the diagnostic kit used, only antibodies against the two major EBV antigens, namely EBVCA and EBNA, were detected in the sera of the patients studied, both in the IgG and IgA classes. In contrast, the presence of anti-EBV IgM antibodies was not detected in any of the patients studied. The seroprevalence results were presented in Figure 2.
Despite the fact that anti-EBVCA and anti-EBNA antibodies were detected more often in people with single EBV infection than in co-infection, these differences were not statistically significant. There were no differences in the frequency of anti-EBNA antibodies between the single-infection and co-infection groups. However, the frequency of anti-EBNA antibodies in both groups of patients was similar.
Then, we analyzed the frequency of the studied antibodies in the context of risk groups, GS and T stages.
Table 3 shows antibody frequencies analyzed by risk group. Both in the serum of patients with single EBV infection and co-infection, anti-EBV antibodies were detected more often in higher risk groups. Anti-EBNA antibodies, both IgA (p = 0.0036) and IgG (p = 0.159), were statistically significantly more common in the intermediate/high risk group in patients with single EBV infection.
As demonstrated in Table 4, the frequency of antibodies did not differ significantly between the group with single EBV infection and EBV/JCV co-infection. Only anti-EBNA IgA antibodies were statistically more common in 8/9 GS in patients with a single EBV infection.
As illustrated in Table 5, a similar tendency was observed regarding the T trait. More often, the analyzed antibodies occurred in a more advanced stage (T2). This difference is particularly evident with respect to IgA and IgG EBNA in a single EBV infection. Anti-EBVCA IgG antibodies were detected in all cases of EBV/JCV co-infection.
3.4. Antibody Levels for EBVCA IgA and IgG and EBNA1 IgA and IgG in PCa Patients
The subsequent analysis concerned the level of anti-EBVCA and anti-EBNA antibodies in both IgA and IgG classes in the context of single EBV infection and EBV/JCV co-infection, revealing significant discrepancies in antibody levels.
The mean level of EBVCA IgA was found to be 760.08 U/mL in the EBV- single infection group; in the EBV/JCV co-infection group - 624.85 U/mL. Conversely, the mean average EBVCA IgG level in the EBV-positive group was found to be 760.79 U/mL, whereas in the EBV/JCV coinfection group this level was determined to be 611.81 U/mL. Furthermore, the mean EBNA IgA level was 675.92 U/mL (EBV-positive group) and 471.26 U/mL (EBV/JCV coinfection group). By contrast, the mean average EBNA IgG level in the EBV-single infection was 659.70 U/mL; in the EBV/JCV co-infection group - 518.96 U/mL.
Thus, statistical analysis of the antibody levels showed a statistically significant difference between these antibodies.
Figure 4 illustrates the antibody levels exhibited by PCa patients, stratified according to their respective risk groups. The intermediate and high risk group demonstrated the highest mean levels of all antibodies that were analyzed. It is notable that the observed differences in antibody levels, as measured by risk group, were found to be statistically significant exclusively in the intermediate and high-risk categories for EBVCA IgG antibodies (Figure 4b). Further details on antibody titers can be found in Table S1 in the supplementary material.
As illustrated in Figure 5, a comprehensive analysis of the levels of antibodies in the EBV single infection and EBV/JCV co-infection groups of PCa patients was conducted. The study revealed that higher antibody titers were observed in patients with single EBV infection. Statistically significant findings were observed in relation to the presence of EBNA antibodies in both IgA and IgG in the GS groups (Figure 4c, 4d). A detailed analysis of the data can be found in Supplementary Table S2.
In patients categorized by T stage, the highest antibody levels were observed in those with an EBV-positive status (with the exception of EBVCA IgA antibody levels at the T1 stage). Levels of both all antibodies were found to be significantly higher in the T2 stage compared to the T1 stage. The observed differences in antibody levels were found to be statistically significant for EBVCA IgG, EBNA IgA and EBNA IgG (Figure 6b, 6c, 6d). A detailed analysis of the data can be found in Supplementary Table S3.
3.5. Comparison of EBV Load in Tumor Tissue in PCa Patients with Single EBV Infection and in Patients with EBV/JCV Co-Infection
At the final point of our study, we assessed EBV load in the collected tumor tissue. EBV load was compared in tissue samples in which only EBV was detected and in samples with EBV/JCV co-infection. The obtained results of this analysis are presented in Figure 7. The result is shown as EBV DNA copies per 1 µg of DNA in tumor tissue. The analysis of the obtained results shows that in malignant tissues in which only EBV DNA was detected, its concentration was statistically significantly higher than in cancer tissues in which the genetic material of both viruses, i.e. EBV and JCV, was simultaneously detected. The average EBV viral load in single EBV infection was 1060 copies EBV DNA/ 1ug tumor tissue and in EBV/JCV co-infection - 656 copies EBV DNA/1 ug tissue.
4. Discussion
There are many reports in the available scientific literature on the detection of oncogenic viruses in prostate cancer, including polyomaviruses [30]. Many researchers point to the potential role of the JCV virus in causing prostatitis, which may consequently lead to the development of prostate cancer. Despite extensive research on polyomaviruses, their role in the pathogenesis of cancer has not yet been fully elucidated [31].
John Cunningham virus (JCPyV) and BKPyV have been described in 1971 [32,33]. JCV, the first human polyomavirus, was isolated from the brain tissue of a patient with Hodgkin's disease, and BK virus was isolated from the urine of a kidney transplant patient.
Already in 2015, Anzivino et al. [34] demonstrated the presence of JCV DNA in 58.3% samples obtained from PCa tissue. In conclusion, the authors defined JCV as a "common resident of the prostate", noting that this fact initiates a discussion about its possible role in the pathogenesis of PCa.
To assess the carcinogenic risk of BKV and JCV infection to humans, the IARC Monograph Working Group [35] analyzed epidemiological evidence and animal bioassays. Both viruses mentioned above have been classified as group 2B, as potentially carcinogenic to humans. The possible association of BK and JC viruses with human cancer is difficult to establish due to the common presence of these viruses in the healthy population [36,37]. Both BKV and JCV infections are usually asymptomatic, although virus particles may be excreted in the urine. In immunocompromised people, latent polyomavirus infection may be reactivated, which may result in high viral load and viruria.
The results regarding the incidence of JCV in PCa are inconsistent between studies. This variability can be attributed to a variety of factors, including differences in sample size and different detection methods. In a study conducted by Delbue et al. [19] JCV was detected in 16.4% of prostate tissue samples tested. However, the study by Zambrano et al. [20] showed a much higher incidence of JCV in prostate tissue - 50%. Gorich et al. [21] presented interesting research discovering a strong relationship between JC virus infection and the likelihood of developing PCa. Namely, these authors observed higher JCV LTag with higher viral load in the group of PCa patients compared to the control group. However, the viral load did not depend on the Gleason score. Therefore, the authors postulate that JCV infection does not influence the progression of cancer lesions. In turn, in the study conducted by Shen et al.’s [38] team 35.2% of the PC specimens extracted from paraffin-embedded tissue were positive for viral DNA. Genetic material for JCPyV was detected in 88.9% of cases, while simultaneous JCPyV and BKPyV infection occurred in 11.1% of cases. These results correlated with prostate cancer progression and prognosis.
In our own research presented here, we detected the presence of viral DNA in 49.6% of clinical samples from freshly frozen prostate cancer tissue, including 71.9% with single EBV infection and 28.1% with EBV/JCV co-infection. We did not detect BKV DNA or a single JCV infection. Therefore, having detected EBV/JCV co-infection, we compared patients with EBV monoinfection with EBV/JCV coinfected patients in the context of risk group, Gleason score and TNM classification.
Our results showed differences in clinicopathological features between single EBV infection and EBV/JCV co-infection. In the group of patients with single EBV infection, most patients were classified as intermediate/high risk, while in the group of patients with EBV/JCV co-infection, more patients were classified as low risk. Among patients with single EBV infection, a more advanced stage of cancer was observed than in patients with EBV/JCV co-infection. Moreover, as the analysis showed, the level of anti-EBVCA and anti-EBNA antibodies was higher in single infection compared to EBV/JCV co-infection. This tendency also occurred in relation to Gleason score and T stage. Higher antibody levels were detected in more advanced tumor stages in single EBV infection. In addition, higher EBV load we observed in single EBV infection.
Many viruses are known to interact with host proteins and bring about changes in genetic, immunological and inflammatory events that lead to initiation or progression of tumors [39]. Viral products, large T antigen of polyomaviruses, can transform prostate cells and interfere with the interferon (IFN) [40].
Numerous studies have shown that cell transformation is the result of JCPyV-induced genome instability [41]. Large T antigen (T-Ag) binds to β-catenin, induces its translocation into host cell nucleus, which in turn enhances the c-MYC gene, which, as a strong proto-oncogene, participates in cell cycle control, DNA metabolism and apoptosis [42,43,44,45].
Three main types of virus-virus interactions have been proposed, i.e. direct interaction of genes or their products, changes in the host environment and immunological interactions [46]. Very thoroughly, co-infection with two viruses, namely interfering, synergistic and non-interfering, has been described in a review by Du et al. [47]. These authors graphically presented the interference phenomenon in which both viral and host factors are involved (Figure 8).
Viral interference is the infection of a host cell with primary virus that inhibits infection with another virus. The phenomenon of interference between two viruses may occur at any stage of the virus replication cycle. Viral interference also often occurs in persistent infections, where viruses remain in infected cells in a state of latency, reducing the level of replication to keep the infected cell alive. Unlike interference, co-infection with some viruses can enhance the replication of other viruses. The best example is respiratory viruses [48]. Another well-known and documented phenomenon is co-infection or super-infection with HBV/HCV [49].
As stated by Moens et. al. [50], due to the fact that human polyomavirus DNA has been found in many different cell types, co-infection with other oncogenic viruses is possible. Therefore, it is assumed that polyomaviruses may act as a cofactor in the development of cancer, including cancer caused by other oncogenic viruses.
Many researchers have shown a high frequency of EBV in prostate cancer [51,52,53]. Our results regarding EBV are consistent with the findings of the above-mentioned authors. Among the examined patients, EBV DNA was detected in 49.5% of cases. However, a co-infection of EBV/JCV was observed in 13.9% of all PCa cases, but no single JCV infection was detected. This fact may suggest that although JCV co-occurs with other viral infections, it does not necessarily appear to be the sole or main viral factor in the development of PCa. It can be assumed that JCV inhibits EBV replication. Does JCV only “reside” in prostate cells or is it a co-factor in EBV infection? Was EBV infection primary and JCV secondary or vice versa? In the light of these studies, there is a need to clarify the role of the JCV virus in the development of prostate cancer.
According to Pietropaolo et al. [54] immunosuppression resulting from the underlying disease or treatment plays a key role in the reactivation of the latent phase of JCV infection. Unfortunately, how JCV latency is reactivated is not well understood.
There is no direct cause-and-effect relationship between infection with any virus and the development of cancer. It is rather persistent viral infections that promote cancer. Cancer transformation is a complex and multi-stage process in which the virus may be one of the factors. To properly determine whether the JCV virus is indeed a risk factor for prostate cancer or is involved in co-infection, the following issues should be taken into account. It is necessary to assess whether PCa patients were exposed to contact with JCV, which will be indirectly confirmed by the presence of specific antibodies. Strategies to prove or exclude „hit-and-run” oncogenesis seem to be an interesting challenge [55]. The „hit and run” theory suggests that viruses encourage the accumulation of mutations, which causes genome instability until the virus is no longer needed for the cell to survive.
Lawson et al. [56] reviewing the available literature on the role of various pathogens, including viruses, in prostate cancer, confirmed that Epstein Barr virus may play a role in chronic prostatitis and subsequent prostate cancer. However, according to these authors, a role for polyomaviruses is unlikely, but not impossible.
Limitation of Our Study
These studies were conducted only in one clinical center, which does not allow generalization of the conclusions. The study group was relatively small, especially the one in which EBV/JCV co-infection was detected. JCV load was not analyzed quantitatively, only qualitatively. In tissue from PCa patients, JCV viral load was classified as low or high based on the cycle threshold (Ct) value of the viral gene. There were only 16 cases of co-infection and low JCV viremia was found in all samples.
Given both the results obtained and the limitations of this study, this will require further research on a much larger group of patients. Further research is needed to evaluate the complex interplay between host genetics, host immunity, and viral infection that may lead to the initiation or promotion of prostate cancer. Only further detailed studies can elucidate the clinical significance of JCV and its possible role in co-infection with other viruses. However, it seems that the presented research may be an inspiration for other researchers.
Finally, it is worth quoting the words of Nobel Prize winner prof. Harald zur Hausen [57], who stated: “Although we know that currently slightly more than 20 % of the global cancer incidence is linked to infectious events, some epidemiological observations suggest that this percentage will increase in the future. The recognition that no cancer linked to infections develops without additional modifications within the host-cell genome permits the speculation that even cancers with well-established chromosomal modifications deserve careful analysis for an additional involvement of infectious agents…. Although still hypothetical, this proposal is accessible to experimental verification. Even if only one of these speculations turns out to be correct, this would have profound implications for the prevention, diagnosis, and hopefully also for the therapy of the respective malignancy”.
5. Conclusions
The role of JCV in the development of PCa remains unclear due to the paucity of research in this area and divergent results among individual researchers. Some studies have shown a potential correlation between the JCV virus and the development and progression of prostate cancer. The results of our study indicate that EBV/JCV co-infection does not have a significant impact on the development of PCa. On the other hand, our studies indirectly confirmed the involvement of the EBV virus in the development and progression of prostate cancer. Lower levels of anti-EBV antibodies and lower EBV viral load in co-infection may suggest that JCV inhibits EBV replication. However, these observations require confirmation in a larger group of cases. Further studies can elucidate the clinical significance of JCV and its possible role in PCa as well as in co-infection with other viruses.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Analysis of EBVCA and EBNA1 antibodies levels in PCa patients with EBV single infection and EBV/JCV co-infection, according to risk groups; Table S2: Analysis of EBVCA and EBNA1 antibodies levels in PCa patients with EBV single infection and EBV/JCV co-infection, according to Gleason score; Table S3: Analysis of EBVCA and EBNA1 antibodies levels in PCa patients with EBV single infection and EBV/JCV co-infection, according to the T stage.
Author Contributions
Conceptualization, J.K., D.S., M.J.J., and M.P-D.; methodology, J.K., M.J.J. and M.P-D.; software, D.S. and M.J.J.; validation, D.S.; formal analysis, J.K., D.S. and M.J.J.; investigation, D.S.; resources, J.K. and D.S.; data curation, J.K., M.J.J. and D.S.; writing—original draft preparation, J.K., and D.S.; writing—review and editing, M.P-D.; visualization, D.S. and M.J.J.; supervision, M.P-D.; project administration, J.K. and M.P-D.; funding acquisition, M.P-D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by Research Grant No. DS 233 of the Medical University of Lublin.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Medical University of Lublin Ethics Committee (no. KE-0254/194/10/2022, 6 October 2022).
Informed Consent Statement
Written informed consent has been obtained from the patients to publish this paper.
Data Availability Statement
The data presented in this study are available in the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- World Health Organization, International Agency for Research of Cancer Global Cancer Observatory 2022 available online:. Available online: https://gco.iarc.who.int/media/globocan/factsheets/cancers/27-prostate-fact-sheet.pdf (accessed on 6 March 2025).
- Wojciechowska, U.; Didkowska, J.A.; Barańska, K.; Miklewska, M.; Michałek, I.; Olasek, P.; Jawołowska, A. Cancer in Poland in 2022. Maria Skłodowska-Curie National Institute of Oncology. Warsaw, Poland; 2024. Available online: https://onkologia.org.
- Huang, L.; La Bonte, M.J.; Craig, S.G.; Finn, S.P.; Allott, E.H. Inflammation and Prostate Cancer: A Multidisciplinary Approach to Identifying Opportunities for Treatment and Prevention. Cancers 2022, 14, 1367. [Google Scholar] [CrossRef]
- Tafuri, A.; Ditonno, F.; Panunzio, A.; Gozzo, A.; Porcaro, A.B.; Verratti, V.; Cerruto, M.A.; Antonelli, A. Prostatic Inflammation in Prostate Cancer: Protective Effect or Risk Factor? Uro 2021, 1, 54–59. [Google Scholar] [CrossRef]
- Maitland, N.J.; Collins, A.T. Inflammation as the Primary Aetiological Agent of Human Prostate Cancer: A Stem Cell Connection? J. Cell Biochem. 2008, 105, 931–939. [Google Scholar] [CrossRef]
- Fernandes, R.; Costa, C.; Fernandes, R.; Barros, A.N. Inflammation in Prostate Cancer: Exploring the Promising Role of Phenolic Compounds as an Innovative Therapeutic Approach. Biomedicines 2023, 11, 3140. [Google Scholar] [CrossRef] [PubMed]
- Gurel, B.; Lucia, M.S.; Thompson, I.M.; Goodman, P.J.; Tangen, C.M.; Kristal, A.R.; Parnes, H.L.; Hoque, A.; Lippman, S.M.; Sutcliffe, S. Chronic inflammation in benign prostate tissue is associated with high-grade prostate cancer in the placebo arm of the prostate cancer prevention trial. Cancer Epidemiol. Prev. Biomark. 2014, 23, 847–856. [Google Scholar] [CrossRef] [PubMed]
- Grisanzio, C.; Signoretti, S. P63 in Prostate Biology and Pathology. J. Cell Biochem. 2008, 103, 1354–1368. [Google Scholar] [CrossRef]
- Vogelstein, B.; Papadopoulos, N.; Velculescu, V.E.; Zhou, S.; Diaz, L.A.; Kinzler, K.W. Cancer Genome Landscapes. Science 2013, 339, 1546–1558. [Google Scholar] [CrossRef]
- Caini, S.; Gandini, S.; Dudas, M.; Bremer, V.; Severi, E.; Gherasim, A. Sexually transmitted infections and prostate cancer risk: A systematic review and meta-analysis. Cancer Epidemiol. 2014, 38, 329–338. [Google Scholar] [CrossRef]
- Akram, N.; Imran, M.; Noreen, M.; Ahmed, F.; Atif, M.; Fatima, Z.; Bilal Waqar, A. Oncogenic role of tumor viruses in humans. Viral Immunol. 2017, 30, 20–27. [Google Scholar] [CrossRef]
- Moore, P.S.; Chang, Y. Why do viruses cause cancer? Highlights of the first century of human tumour virology. Nat. Rev. Cancer 2010, 10, 878–889. [Google Scholar] [CrossRef]
- Chang, Y.; Moore, P.S.; Weiss, R.A. Human oncogenic viruses: nature and discovery. Phil. Trans. R. Soc. B. 2017, 372, 20160264. [Google Scholar] [CrossRef] [PubMed]
- MacLennan, S.A.; Marra, M.A. Oncogenic Viruses and the Epigenome: How Viruses Hijack Epigenetic Mechanisms to Drive Cancer. Int. J. Mol. Sci. 2023, 24, 9543. [Google Scholar] [CrossRef]
- Abumisimir, B.; Mahasneh, I.; Kasmi, Y.; Saif, I.; Hammou, R.A.; Mustafa, M. Prostate cancer and viral infections: epidemiological and clinical indications. Editor(s): Moulay Mustapha Ennaji, Oncogenic Viruses, Academic Press, 2023, 263-272.
- Dalianis, T.; Hirsch, H.H. Human polyomaviruses in disease and cancer. Virology 2013, 437, 63–72. [Google Scholar] [CrossRef]
- Frisque, R.J.; Bream, G.L.; Cannella, M.T. Human polyomavirus JC virus genome. J. Virol. 1984, 51, 458–469. [Google Scholar] [CrossRef]
- Kean, J.M.; Rao, S.; Wang, M.; Garcea, R.L. Seroepidemiology of human polyomaviruses. PLoS Pathog. 2009, 5, e1000363. [Google Scholar] [CrossRef] [PubMed]
- Delbue, S.; Comar, M.; Ferrante, P. Review on the role of the human Polyomavirus JC in the development of tumors. Infect Agent Cancer. 2017, 12, 10. [Google Scholar] [CrossRef]
- Zambrano, A.; Kalantari, M.; Simoneau, A.; Jensen, J.L.; Villarreal, L.P. Detection of human polyomaviruses and papillomaviruses in prostatic tissue reveals the prostate as a habitat for multiple viral infections. Prostate. 2002, 53, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Gorish, B.M.T. JC Polyoma Virus as a Possible Risk Factor for Prostate Cancer Development - Immunofluorescence and Molecular Based Case Control Study. Cancer Control. 2022, 29, 10732748221140785. [Google Scholar] [CrossRef]
- Villani, S.; Gagliano, N.; Procacci, P.; Sartori, P.; Comar, M.; Provenzano, M.; Favi, E.; Ferraresso, M.; Ferrante, P.; Delbue, S. Characterization of an in vitro model to study the possible role of polyomavirus BK in prostate cancer. J Cell Physiol. 2019, 234, 11912–11922. [Google Scholar] [CrossRef]
- Delbue, S.; Matei, D.V.; Carloni, C.; Pecchenini, V.; Carluccio, S.; Villani, S.; Tringali, V.; Brescia, A.; Ferrante, P. Evidence supporting the association of polyomavirus BK genome with prostate cancer. Med Microbiol Immunol. 2013, 202, 425–430. [Google Scholar] [CrossRef]
- Ahmed, K.; Sheikh, A.; Fatima, S.; Haider, G.; Ghias, K.; Abbas, F.; Mughal, N.; Abidi, S.H. Detection and characterization of latency stage of EBV and histopathological analysis of prostatic adenocarcinoma tissues. Sci. Rep. 2022, 12, 10399. [Google Scholar] [CrossRef] [PubMed]
- Moens, U.; Prezioso, C.; Pietropaolo, V. Functional Domains of the Early Proteins and Experimental and Epidemiological Studies Suggest a Role for the Novel Human Polyomaviruses in Cancer. Front Microbiol. 2022, 13, 834368. [Google Scholar] [CrossRef]
- Kiś, J.; Góralczyk, M.; Sikora, D.; Stępień, E.; Drop, B.; Polz-Dacewicz, M. Can the Epstein–Barr Virus Play a Role in the Development of Prostate Cancer? Cancers 2024, 16, 328. [Google Scholar] [CrossRef]
- Epstein, J.I.; Egevad, L.; Amin, M.B.; Delahunt, B.; Srigley, J.R.; Humphrey, P.A.; Grading Committee. The 2014 International Society of Urological Pathology (ISUP) Consensus Conference on Gleason Grading of Prostatic Carcinoma: Definition of Grading Patterns and Proposal for a New Grading System. Am. J. Surg. Pathol. 2016, 40, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Epstein, J.I.; Zelefsky, M.J.; Sjoberg, D.D.; Nelson, J.B.; Egevad, L.; Magi-Galluzzi, C.; Vickers, A.J.; Parwani, A.V.; Reuter, V.E.; Fine, S.W.; et al. A Contemporary Prostate Cancer Grading System: A Validated Alternative to the Gleason Score. Eur. Urol. 2016, 69, 428–435. [Google Scholar] [CrossRef]
- EAU Guidelines for Prostate Cancer. Available online: https://uroweb.org/guidelines/prostate-cancer/chapter/classification-and-staging-systems (accessed on 8 February 2025).
- Abidi, S.H.; Bilwani, F. , Ghias, K. , Abbas F. Viral etiology of prostate cancer: Genetic alterations and immune response. A literature review, International Journal of Surgery 2018, 52, 136–140. [Google Scholar] [PubMed]
- Balis, V.; Sourvinos, G.; Soulitzis, N.; Giannikaki, E.; Sofras, F.; Spandidos, D.A. Prevalence of BK virus and human papillomavirus in human prostate cancer. Int J Biol Markers. 2007, 22(4):245-51.
- Padgett, B.L.; Zu Rhein, G.M.; Walker, D.L.; Echroade, R.; Dessel, B. Cultivation of papova-like virus from human brain with progressive multifocal leukoencephalopathy. Lancet 1971, 1, 1257–60. [Google Scholar] [CrossRef]
- Gardner, S.D.; Field, A.M.; Coleman, D.V.; Hulme, B. New human papovavirus (B.K.) isolated from urine after renal transplantation. Lancet 1971, 1, 1:1253–7. [Google Scholar] [CrossRef]
- Anzivino, E.; Rodio, D.M.; Mischitelli, M.; Bellizzi, A.; Sciarra, A.; Salciccia, S.; Gentile, V.; Pietropaolo, V. High Frequency of JCV DNA Detection in Prostate Cancer Tissues. Cancer Genomics Proteomics, 2015,12(4):189-200.
- IARC Monographs on the Evaluation of Carcinogenic Risks to Humans, Malaria and Some Polyomaviruses (SV40, BK, JC, and Merkel Cell Viruses), Lyon 2013, 104.
- Jiang, M.; Abend, J.R.; Johnson, S.F.; Imperiale, M.J. The role of polyomaviruses in human disease. Virology, 2009, 384(2):266-73.
- Prado, J.C.M.; Monezi, T.A.; Amorim, A.T.; Lino, V.; Paladino, A.; Boccardo, E. Human polyomaviruses and cancer: an overview. Clinics (Sao Paulo), 2018, 73(suppl 1):e558s.
- Shen, C. , Tung, C., Chao, C. Jou Y. Huang S.; Meng M.; Chang D.; Chen P. The differential presence of human polyomaviruses, JCPyV and BKPyV, in prostate cancer and benign prostate hypertrophy tissues. BMC Cancer 2021, 21: 1141. [Google Scholar] [CrossRef]
- Martinez-Fierro, M.L.; Leach, R.J.; Gomez-Guerra, L.S.; Gomez-Guerra, L.; Garza-Guajardo, R.; et al. Identification of viral infections in the prostate and evaluation of their association with cancer, BMC Cancer 2010, 10: 326. 10.
- Zheng, Z.M. Viral oncogenes, noncoding RNAs, and RNA splicing in human tumor viruses. Int. J. Biol. Sci, 2010, 6(7): 730–755. 41.
- Barbier, M.T.; Del Valle, L. Co-Detection of EBV and Human Polyomavirus JCPyV in a Case of AIDS-Related Multifocal Primary Central Nervous System Diffuse Large B-Cell Lymphoma. Viruses 2023, 15, 755. [Google Scholar] [CrossRef]
- Darbinyan, A.; White, M.K.; Akan, S.; Radhakrishnan, S.; Del Valle, L.; Amini, S.; Khalili, K. Alterations of DNA damage repair pathways resulting from JCV infection. Virology 2007, 364, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Del Valle, L.; Gordon, J.; Assimakopoulou, M.; Enam, S.; Geddes, J.F.; Varakis, J.N.; Katsetos, C.D.; Croul, S.; Khalili, K. Detection of JC Virus DNA sequences and expression of the viral regulatory protein T-Antigen in tumors of the Central Nervous System. Cancer Res. 2001, 61, 4287–4293. [Google Scholar]
- Ripple, M.J.; Parker Struckhoff, A.; Trillo-Tinoco, J.; Li, L.; Margolin, D.A.; McGoey, R.; Del Valle, L. Activation of c-Myc and Cyclin D1 by JCV T-Antigen and beta-catenin in colon cancer. PLoS ONE 2014, 9, e106257. [Google Scholar] [CrossRef]
- Nguyen, L.; Papenhausen, P.; Shao, H. The Role of c-MYC in B-Cell Lymphomas: Diagnostic and Molecular Aspects. Genes 2017, 8, 116. [Google Scholar] [CrossRef] [PubMed]
- Escobedo-Bonilla, C.M. Mini Review: Virus Interference: History, Types and Occurrence in Crustaceans. Front. Immunol, 2021, 12:674216.
- Du, Y.; Wang, C.; Zhang, Y. Viral Coinfections. Viruses, 2022;14(12):2645.
- Goto, H. , Ihira H., Morishita K., Tsuchiya M., Ohta K., Yumine N., Tsurudome M., Nishio M. Enhanced growth of influenza A virus by coinfection with human parainfluenza virus type 2. Med. Microbiol. Immunol. 2016, 205: 209–218.
- Chu, C.-J. , Lee S.-D. Hepatitis B virus/hepatitis C virus coinfection: Epidemiology, clinical features, viral interactions and treatment. J. Gastroenterol. Hepatol. 2008, 23, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Moens, U. ; Van Ghule, M; Ehlers, B. Are human polyomaviruses co-factors for cancers induced by other oncoviruses? Rev Medical Virology, 2014, 24(5): 343-360.
- Ahmed, K. , Sheikh A., Fatima S., Haider G., Ghias K., Abbas F., Mughal N., Abidi S.H. Detection and characterization of latency stage of EBV and histopathological analysis of prostatic adenocarcinoma tissues. Sci. Rep. 2022, 12:10399.. [CrossRef]
- Whitaker, N.J. , Glenn W.K., Sahrudin A., Orde M.M., Delprado W., Lawson J.S. Human papillomavirus and Epstein Barr virus in prostate cancer: Koilocytes indicate potential oncogenic influences of human papillomavirus in prostate cancer. Prostate 2013, 73, 236–241. [Google Scholar] [CrossRef]
- Nahand, J.S. , Khanaliha K., Mirzaei H., Moghoofei M., Baghi H.B., Esghaei M., Khatami A.R., Fatemipour M., Bokharaei-Salim F. Possible role of HPV/EBV coinfection in anoikis resistance and development in prostate cancer. BMC Cancer 2021, 21:926. [Google Scholar] [CrossRef]
- Pietropaolo V, Prezioso C, Bagnato F, Antonelli G. John Cunningham virus: an overview on biology and disease of the etiological agent of the progressive multifocal leukoencephalopathy. New Microbiol, 2018,41(3):179-186.
- Ferreira, A.D.; Tayyar, Y.; Idris, A.; McMillan, N.A.J. A “hit-and-run” affair – A possible link for cancer progression in virally driven cancers. Biochimica et Biophysica Acta (BBA) - Reviews on Cancer, 2021, 1875(1):188476.
- Lawson JS, Glenn WK. Multiple pathogens and prostate cancer. Infect Agent Cancer, 2022, 17(1):23.
- zur Hausen The Search for Infectious Causes of Human Cancers: Where and Why (Nobel Lecture), Angewandte Chemie International Edition 2009, 48(32), 2009, 48(32): 5769-5969.
Figure 1.
Frequency of EBV- single infection and EBV/ JCV co-infection in tumor tissue isolated from patients with prostate cancer. Green color – infected patients; Gray color – no infected patients; Red color – EBV-single infection; Blue color – EBV/JCV co-infection.
Figure 1.
Frequency of EBV- single infection and EBV/ JCV co-infection in tumor tissue isolated from patients with prostate cancer. Green color – infected patients; Gray color – no infected patients; Red color – EBV-single infection; Blue color – EBV/JCV co-infection.
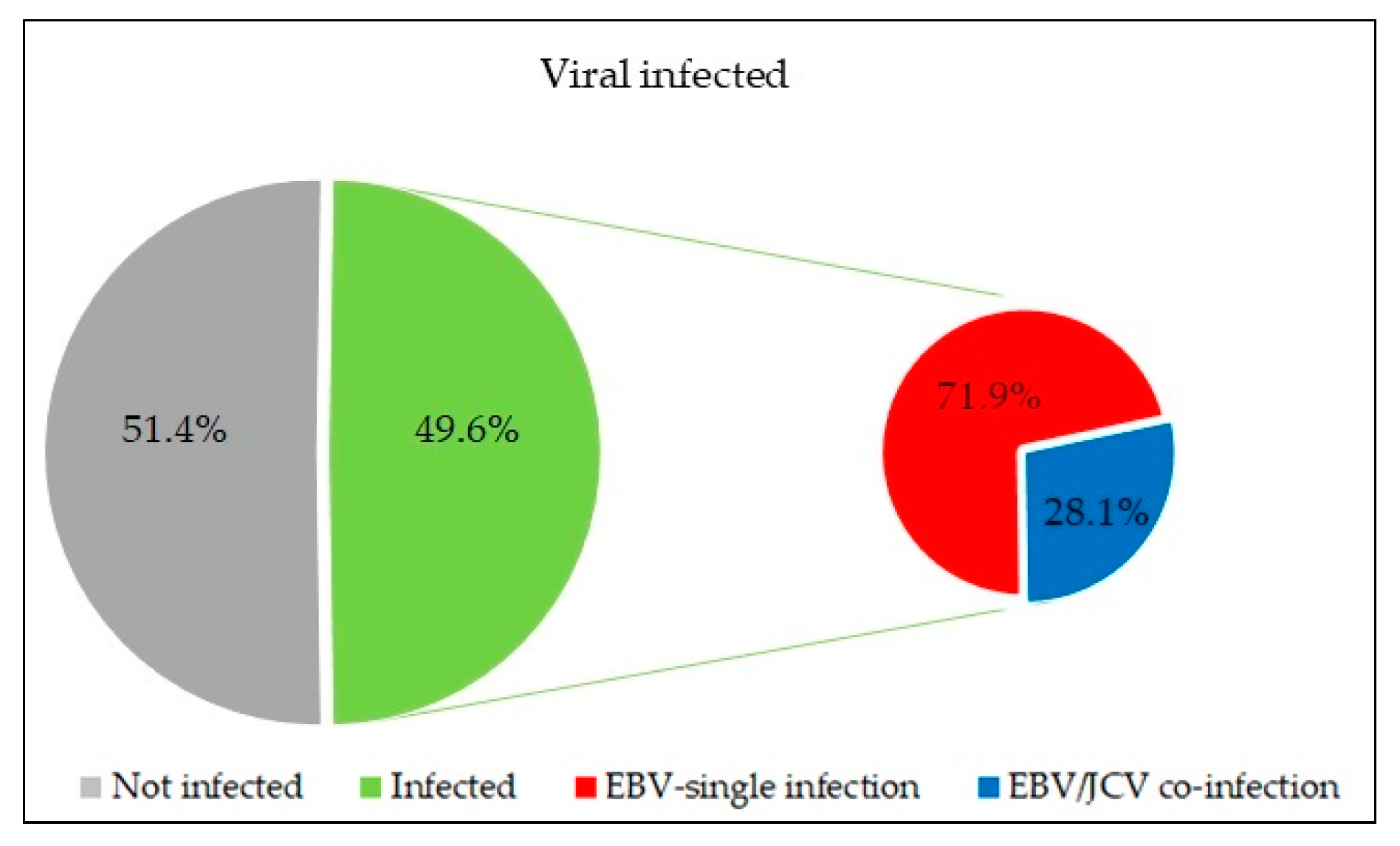
Figure 2.
Prevalence of EBVCA and EBNA1 antibodies in PCa patients. Red color – EBV-single infection; Blue color – EBV/JCV co-infection.
Figure 2.
Prevalence of EBVCA and EBNA1 antibodies in PCa patients. Red color – EBV-single infection; Blue color – EBV/JCV co-infection.
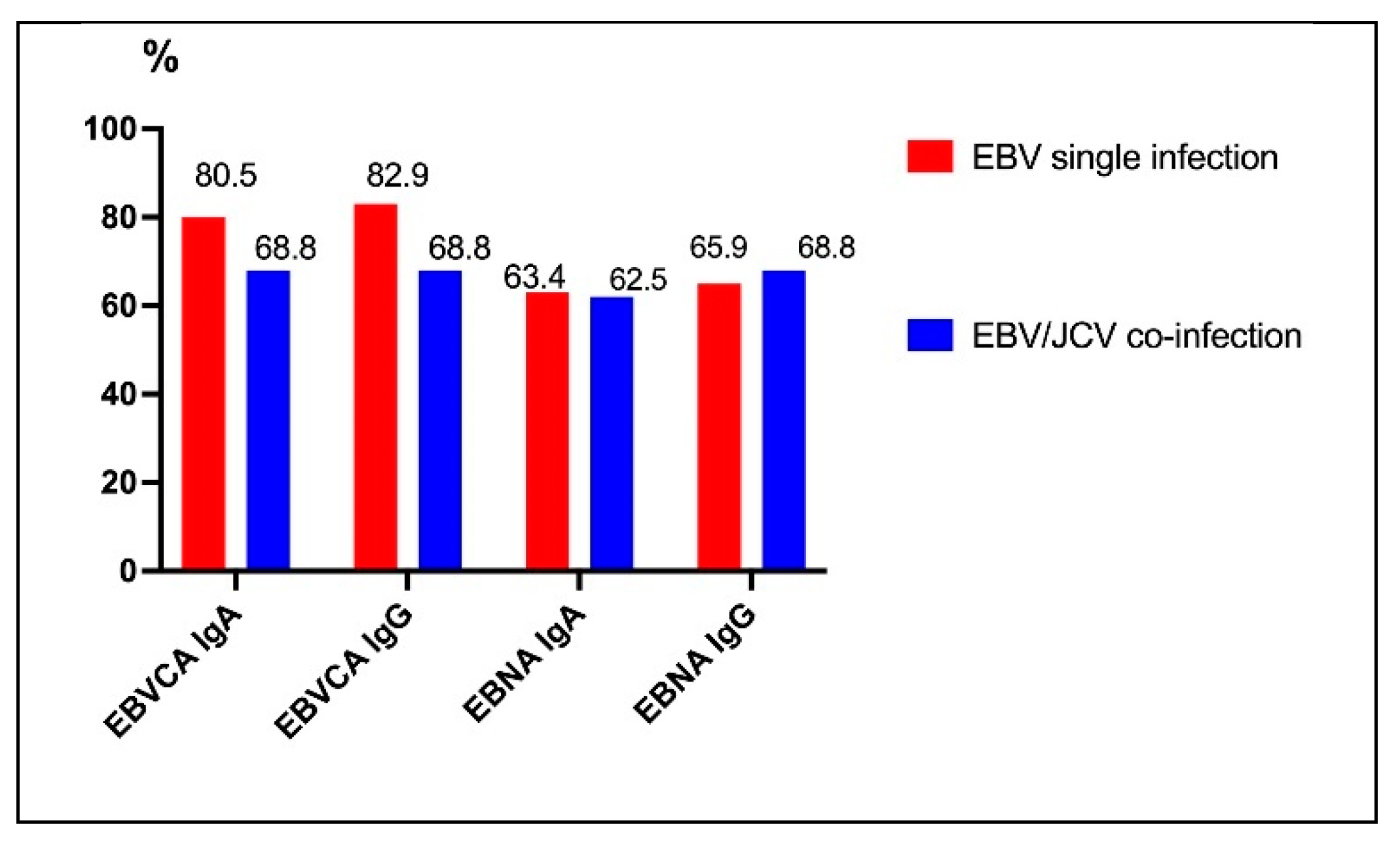
Figure 3.
Levels of EBVCA and EBNA1 antibodies in PCa patients with single EBV infection compared to patients with EBV/JCV co-infection.
Figure 3.
Levels of EBVCA and EBNA1 antibodies in PCa patients with single EBV infection compared to patients with EBV/JCV co-infection.
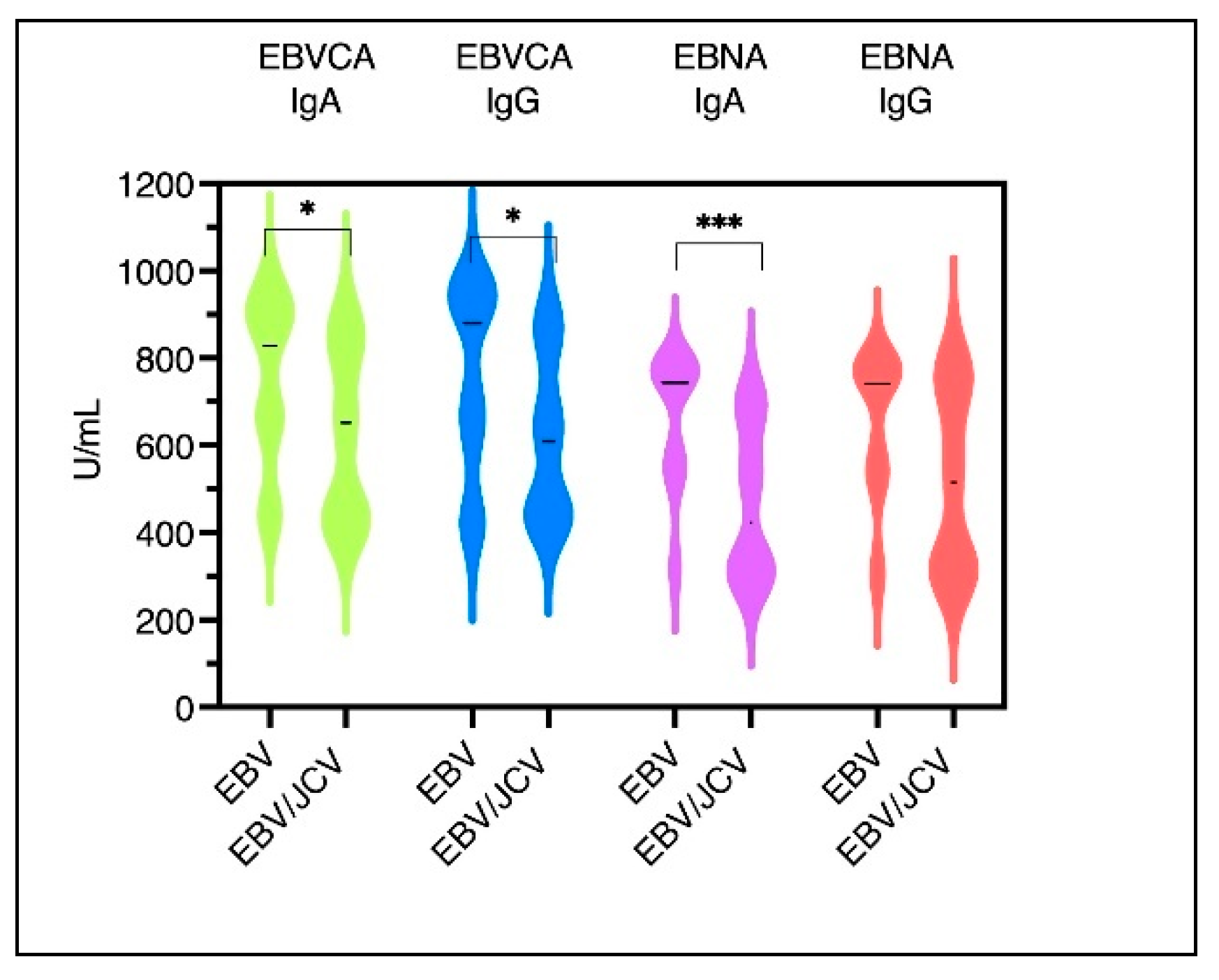
Figure 4.
The level of antibodies in relation to the risk group: (a) EBNA1 IgA, (b) EBNA1 IgG, (c) EBVCA IgA and (d) EBVCA IgG; Mann-Whitney U Test test. Red color – EBV-single infection; Blue color –EBV/JCV co-infection.
Figure 4.
The level of antibodies in relation to the risk group: (a) EBNA1 IgA, (b) EBNA1 IgG, (c) EBVCA IgA and (d) EBVCA IgG; Mann-Whitney U Test test. Red color – EBV-single infection; Blue color –EBV/JCV co-infection.

Figure 5.
The level of anti-EBV antibodies in relation to GS: (a) EBNA1 IgA, (b) EBNA1 IgG, (c) EBVCA IgA and (d) EBVCA IgG; Mann-Whitney U Test Red color – EBV- single infection; Blue color –EBV/JCV co-infection.
Figure 5.
The level of anti-EBV antibodies in relation to GS: (a) EBNA1 IgA, (b) EBNA1 IgG, (c) EBVCA IgA and (d) EBVCA IgG; Mann-Whitney U Test Red color – EBV- single infection; Blue color –EBV/JCV co-infection.

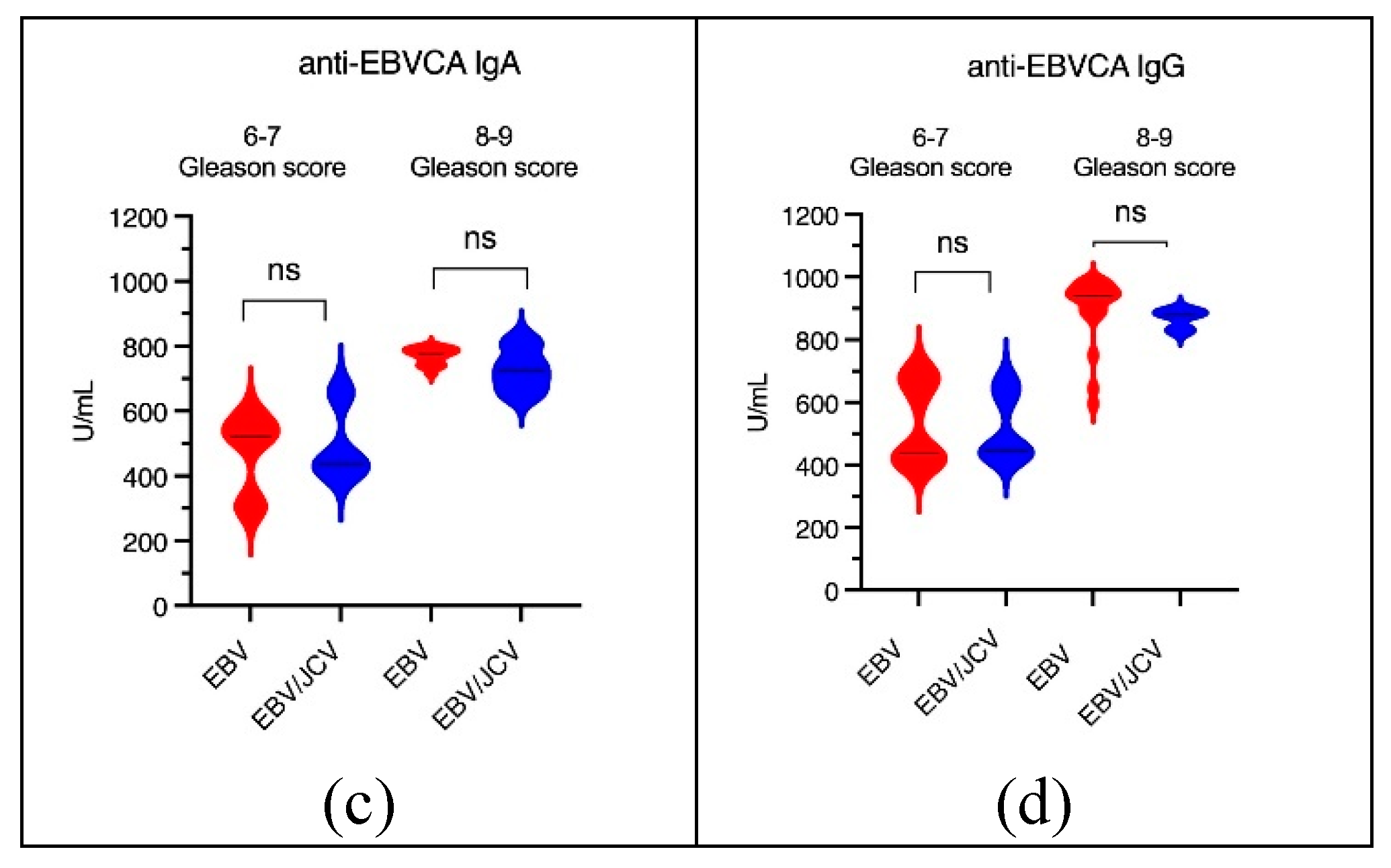
Figure 6.
The level of anti-EBV antibodies in the relation to the T stage: (a) EBNA1 IgA, (b) EBNA1 IgG, (c) EBVCA IgA, (d) EBVCA IgG; Mann-Whitney U Test. Red color – EBV-single infection; Blue color –EBV/JCV co-infection.
Figure 6.
The level of anti-EBV antibodies in the relation to the T stage: (a) EBNA1 IgA, (b) EBNA1 IgG, (c) EBVCA IgA, (d) EBVCA IgG; Mann-Whitney U Test. Red color – EBV-single infection; Blue color –EBV/JCV co-infection.
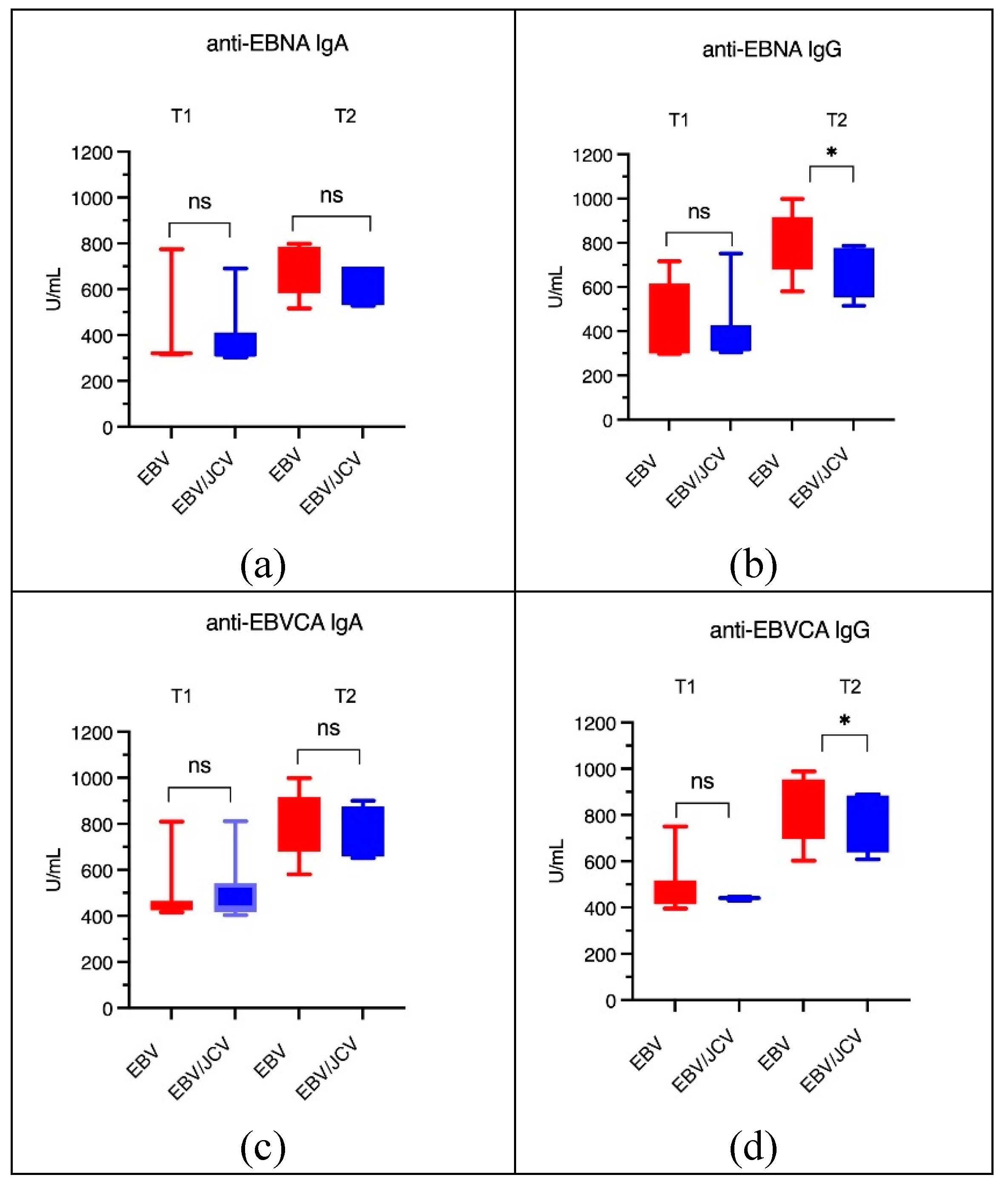
Figure 7.
Tumor tissue EBV load in single EBV infection and in EBV/JCV co-infection. ** p = 0.0068; Mann-
Whitney U Test.
Figure 7.
Tumor tissue EBV load in single EBV infection and in EBV/JCV co-infection. ** p = 0.0068; Mann-
Whitney U Test.
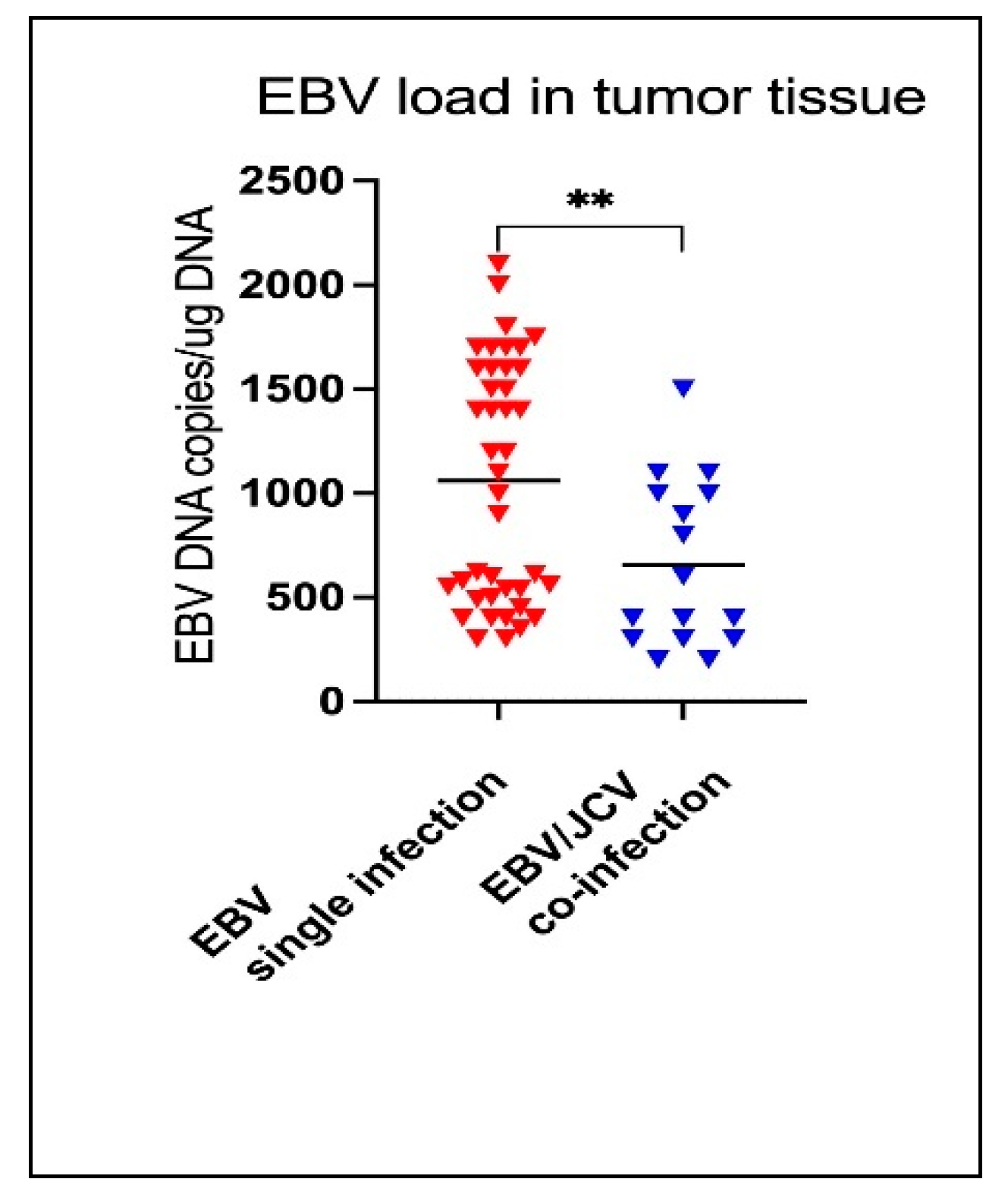
Figure 8.
Effects of viral co-infection (by Du et al. [47]).
Figure 8.
Effects of viral co-infection (by Du et al. [47]).
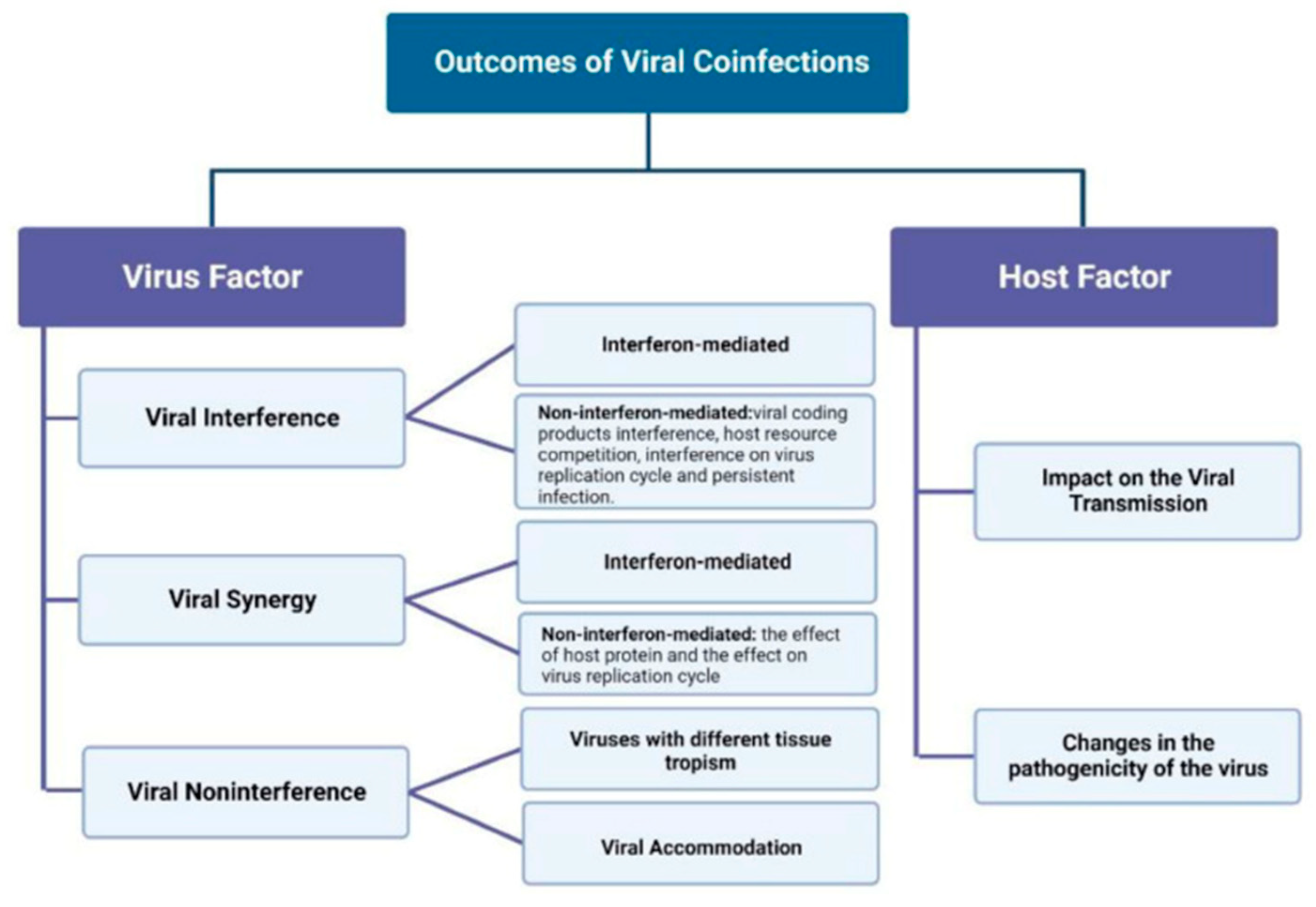
Table 2.
Clinical characteristics of PCa patients with EBV single infection and EBV/JCV co-infection.
Table 2.
Clinical characteristics of PCa patients with EBV single infection and EBV/JCV co-infection.
| PCa Patients | |||||
| EBV single infection N = 41 |
EBV/JCV co-infection N = 16 |
||||
| n | % | n | % | ||
| Risk | Low | 10 | 24.4 | 9 | 56.2 |
| Intermediate/ high | 31 | 75.6 | 7 | 43.8 | |
| p | 0.0307* | ||||
| Gleason score | 6-7 | 18 | 43.9 | 12 | 75.0 |
| 8-9 | 23 | 56.1 | 4 | 25.0 | |
| p | 0.0430* | ||||
| T | T1 | 11 | 26.8 | 10 | 62.5 |
| T2 | 30 | 73.2 | 6 | 37.5 | |
| T3 | 0 | 0.0 | 0 | 0.0 | |
| T4 | 0 | 0.0 | 0 | 0.0 | |
| p | 0.0166* | ||||
| N | N0 | 41 | 100.0 | 16 | 100.0 |
| M | M0 | 41 | 100.0 | 16 | 100.0 |
* Statistically significant; Pearson’s chi-squared test.
Table 3.
Prevalence of EBVCA and EBNA1 antibodies in PCa patients according to the risk group.
| EBV-positive | EBV/JCV coinfection | |||||||||
| Low risk | Intermediate/ high risk |
p | Low risk | Intermediate/ high risk |
p | |||||
| n | (%) | n | (%) | n | (%) | n | (%) | |||
| n=10 | n=30 | n=9 | n=7 | |||||||
| EBVCA IgA |
7 | 70.00 | 26 | 86.67 | 0.2297 | 5 | 55.56 | 6 | 85.71 | 0.1967 |
| EBVCA IgG |
9 | 90.00 | 25 | 83.33 | 0.9090 | 5 | 55.56 | 6 | 85.71 | 0.1967 |
| EBNA IgA |
3 | 30.00 | 23 | 83.33 | 0.0036* | 5 | 55.56 | 5 | 71.43 | 0.5153 |
| EBNA IgG |
4 | 40.00 | 23 | 83.33 | 0.0159* | 5 | 55.56 | 6 | 85.71 | 0.1967 |
* Statistically significant; Pearson’s chi-squared test.
Table 4.
Prevalence of EBVCA and EBNA1 antibodies in PCa patients according to GS.
| EBV- single infection | EBV/JCV co-infection | |||||||||
| 6-7 Gleason score |
8-9 Gleason score |
p | 6-7 Gleason score |
8-9 Gleason score |
p | |||||
| n | (%) | n | (%) | n | (%) | n | (%) | |||
| n=18 | n=23 | n=12 | n=4 | |||||||
| EBVCA IgA | 12 | 66.67 | 13 | 56.52 | 0.5087 | 7 | 58.33 | 4 | 100.00 | 0.1195 |
| EBVCA IgG | 13 | 72.22 | 21 | 91.30 | 0.1071 | 8 | 66.67 | 3 | 75.00 | 0.7555 |
| EBNA IgA | 9 | 50.00 | 17 | 73.91 | 0.0217* | 7 | 58.33 | 3 | 75.00 | 0.5510 |
| EBNA IgG | 10 | 55.56 | 17 | 73.91 | 0.2186 | 7 | 58.33 | 4 | 100.00 | 0.1195 |
* Statistically significant; Pearson’s chi-squared test.
Table 5.
Prevalence of EBVCA and EBNA1 antibodies in patients with PCa ac-cording to T stage.
| EBV single infection | EBV/JCV co-infection | |||||||||
| T1 | T2 | p | T1 | T2 | p | |||||
| n | (%) | n | (%) | n | (%) | n (%) | (%) | |||
| n=11 | n=30 | n=10 | n=6 | |||||||
| EBVCA IgA | 7 | 63.64 | 26 | 86.67 | 0.0992 | 6 | 60.00 | 5 | 83.33 | 0.3296 |
| EBVCA IgG | 9 | 81.82 | 25 | 83.33 | 0.9090 | 5 | 50.00 | 6 | 100.00 | 0.0367* |
| EBNA IgA | 3 | 27.27 | 23 | 76.67 | 0.0036* | 6 | 60.00 | 4 | 66.67 | 0.7897 |
| EBNA IgG | 4 | 36.36 | 23 | 76.67 | 0.0159* | 6 | 60.00 | 5 | 83.33 | 0.3296 |
* Statistically significant; Pearson’s chi-squared test.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



