Submitted:
01 April 2025
Posted:
01 April 2025
You are already at the latest version
Abstract
Plastic pollution, particularly in the form of nanoplastics, has emerged as a significant global environmental concern, affecting ecosystems and human health. This review examines the complex interactions between fungi and nanoplastics, emphasizing the dual role of fungi in plastic degradation and potential pathogenicity. Fungi possess unique enzymatic machinery that allows them to degrade various plastic materials, offering promising bioremediation strategies. However, the presence of nanoplastics can impact fungal physiology, potentially altering metabolic pathways, enhancing virulence, and contributing to antifungal resistance. The interactions between nanoplastics and fungi raise critical questions about their ecological roles and implications for human health. This review underscores the need for further research into the mechanisms of nanoplastic-fungi interactions, the effects on fungal communities, and the potential for increased resistance to antifungal agents. Understanding these dynamics is essential for developing effective strategies to address the challenges posed by plastic pollution and fungal infections. The findings highlight the importance of interdisciplinary approaches in addressing the intertwined issues of environmental sustainability and public health.
Keywords:
nanoplastics
; fungi
; antifungal resistance
; plastic degradation
1. Introduction
Plastic pollution has emerged as a critical global environmental challenge, significantly affecting ecosystem health, human well-being, and long-term sustainability [1,2,3,4]. The breakdown of plastic materials occurs through various mechanisms, including UV irradiation, thermal degradation, mechanical stress, and biodegradation, ultimately leading to the formation of microplastics and nanoplastics [5,6]. Among these byproducts, nanoplastics—defined as particles smaller than 100 nanometers—have garnered considerable attention due to their pervasive presence in the environment and their potential harmful effects on living organisms. Their unique physicochemical properties enhance both reactivity and bioavailability, which raises serious environmental and health concerns [7,8,9,10].
Fungi, as vital microorganisms, play essential roles in nutrient cycling and ecosystem functioning. Certain fungal species demonstrate remarkable abilities to degrade a variety of plastics, positioning them as promising candidates for bioremediation strategies [11,12]. Their significance in plastic biodegradation is attributed to their capacity to secrete a diverse array of degrading enzymes, which catalyze the conversion of complex plastic polymers into simpler, more manageable compounds. This enzymatic activity facilitates the oxidation or hydrolysis of plastics, resulting in the formation of functional groups that increase hydrophilicity. Consequently, high molecular weight plastics can be transformed into lower molecular weight compounds that fungi can readily assimilate [13,14,15]. Some fungi exhibit the ability to effectively degrade specific plastics within just a few days, underscoring their efficiency in addressing plastic pollution [14].
Despite their beneficial potential, it is essential to recognize that fungi are not exclusively advantageous. Invasive fungal species contribute to an estimated 2.5 million human fatalities each year [16]. Among these, Cryptococcus neoformans, Aspergillus fumigatus, Candida albicans, and Candida auris have been identified by the World Health Organization (WHO) as some of the most dangerous pathogenic fungi to humans [17]. As research increasingly investigates the interactions between fungi and nanoplastics, it becomes crucial to evaluate how these plastic nanoparticles impact fungal physiology and pathogenicity. The adsorption of nanoplastics onto fungal cell surfaces may lead to alterations in physiological functions, potentially affecting ecological roles and interactions with other organisms. Furthermore, the presence of nanoplastics may influence the metabolic pathways of fungi, leading to changes in their enzymatic profiles and nutrient assimilation capabilities. This could result in both positive and negative outcomes, depending on the specific context and species involved. For example, while some fungi might enhance their plastic-degrading capabilities in response to nanoplastic exposure, others may experience detrimental effects that impair their metabolic functions and ecological interactions. The ability of fungi to adapt to the presence of nanoplastics raises important questions concerning their potential virulence and antifungal resistance, particularly in human pathogenic species. The alterations in fungal physiology due to nanoplastic interactions could lead to an increased expression of virulence factors or enhanced resistance mechanisms against antifungal agents. Understanding these dynamics is crucial for developing effective strategies to combat both plastic pollution and fungal infections.
This review aims to provide a comprehensive examination of the relationships between nanoplastics and fungi by systematically analyzing the mechanisms of plastic degradation by fungi, their metabolic pathways, and physiological adaptations. Additionally, it will address the potential implications for virulence and antifungal resistance in human pathogenic fungi. By elucidating these complex interactions, this review seeks to contribute to the broader understanding of how nanoplastics impact fungal communities and their ecological roles, as well as to inform strategies aimed at mitigating the dual threats of plastic pollution and fungal diseases.
2. The Role of Fungi in Plastic Degradation
Fungi play a pivotal role in the degradation and assimilation of various plastic materials, including polyethylene, polystyrene, and polyvinyl chloride [18,19,20]. Certain fungal species have demonstrated the remarkable capacity to degrade specific plastics within a matter of days, highlighting their potential for addressing plastic pollution [21]. The advantages of fungal degradation can be summarized into three key points:
2.1. Enhanced Substrate Penetration
The apical growth pattern of filamentous fungal mycelia enables these organisms to penetrate soil substrates more effectively than bacteria. This capability is attributed to their secretion of extracellular enzymes and hydrophobic proteins, which enhance their adhesion to hydrophobic substrates [22]. Such penetration is particularly pronounced in soil environments where saprotrophic fungi thrive [23].
2.2. Diverse Enzymatic Machinery
Fungi possess a wide range of non-specific enzyme cassettes [24,25] , which equip them to degrade and utilize hydrocarbons and aromatic compounds found in plastics as carbon and energy sources. Secreted fungal enzymes, including lignin peroxidase, manganese peroxidase, laccase, and polyesterase (Table 1), facilitate the oxidation or hydrolysis of plastic polymers, leading to the formation of functional groups that increase hydrophilicity. This enhanced hydrophilicity promotes the breakdown of high molecular weight plastics into lower molecular weight compounds that fungi can readily assimilate. Additionally, various internal factors within fungal cells can induce pro-oxidant ions, which promote degradation through oxidative reactions, particularly through the generation of reactive oxygen species. Hydroxyl radicals, recognized for their high redox potential, are particularly effective oxidants in this context [26,27]. This oxidative mechanism enables fungi to participate in the degradation of a variety of recalcitrant pollutants, including hydrocarbons, chlorophenols, and pesticides [25,28,29].
2.3. Adaptability to Diverse Environments
Fungi exhibit remarkable adaptability, enabling them to utilize nearly any organic carbon source and thrive in conditions characterized by low humidity, nutrient scarcity, and acidic pH levels [47]. Some species can endure dry periods through a state of cryptobiosis [48,49]. The metabolic pathways within fungi are mediated by cytochrome P450 (CYP) monooxygenases (EC:1.14.13.12), which facilitate the oxidation of substrates in microsomes [50,51]. These enzymes belong to the heme-protein superfamily and are involved in various biological processes, including adaptation to environmental stress, toxin production, and the metabolism of both endogenous and xenobiotic compounds, thereby enhancing fungal fitness [52,53,54,55]. CYP monooxygenases typically function as terminal oxidases in the electron transfer chain associated with NADPH reductase, facilitating the incorporation of an oxygen atom into the hydrocarbon chain of nanoplastics while reducing the other oxygen atom to water [56]. The number of CYP genes varies according to the lifestyle of the fungal species; yeasts and yeast-like fungi possess relatively few CYPs (e.g., 3 in Saccharomyces cerevisiae, 6 in C. neoformans, and 10 in C. albicans), while filamentous fungi typically harbor a greater number of CYP genes, as exemplified by Aspergillus spp. with 79 genes and Agaricus bisporus with 109 [57,58]. As a result, plastic-degrading fungi are predominantly filamentous species (Table 1), including Ganoderma lucidum, Pleurotus abalone, Penicillium chrysogenum, and Aspergillus niger [30,31,33,36]. Ekanayaka et al. assessed 395 filamentous fungal strains from the Ascomycota and Basidiomycota phyla, identifying over 200 species capable of degrading various plastics under diverse environmental conditions [59]. Their findings revealed that fungi such as Aspergillus tubingensis effectively disrupt the chemical bonds within plastic molecules and successfully colonize plastic surfaces [59]. Numerous plastic-degrading fungi have been isolated from both terrestrial and marine environments [60], including Trichoderma sp., Clitocybe sp., Monascus sp., and Phanerochaete sp., which enhance the degradation of polyethylene (both LDPE and HDPE), polylactic acid, polyurethanes, polyethylene terephthalate, and bisphenol A polycarbonate [61,62,63,64,65,66]. Marine environments have yielded marine yeasts such as Rhodotorula mucilaginosa, Zalerion maritimum, Alternaria alternata, Penicillium spp., and Aspergillus sp., which significantly facilitate the degradation of polyethylene and polystyrene, contributing to healthier ecosystems by reducing plastic waste [18,67,68,69,70].
Overall, the capacity of fungi to degrade plastics presents a promising avenue for bioremediation strategies aimed at mitigating the environmental impacts of plastic waste. Advances in the exploration of fungal enzymes, along with genetic engineering techniques, could enhance biodegradation processes and contribute to sustainable waste management practices.
3. The Impact of Nanoplastics on Fungal Physiology and Pathogenicity
Nanoplastics possess unique physical properties, including an increased surface area, specific transport characteristics, and distinctive interactions with light and natural colloids [7]. The larger surface area enhances the adsorption capacity of nanoplastics for natural organic matter in the environment [71]. Adsorption predominantly occurs through chemical bonding on certain types of nanoplastics, facilitated by ligand exchange mechanisms with oxide nanoplastics. This interaction reduces surface hydrophobicity, increases interactions among plastic particles, and affects their aggregate size [72]. Furthermore, the presence of electron-attracting groups within the aromatic rings of nanoplastic polymers facilitates strong π–π interactions, contributing to their exceptional ion adsorption properties [73,74]. The substances adsorbed onto nanoplastics can interact with extracellular polymers secreted by fungal cells, potentially enveloping the nanoplastics in a unique layered structure referred to as the eco-corona. This eco-corona can significantly alter the dynamics between nanoplastics and fungi [71]. Fungal cell wall thickness typically ranges from 0.1 to 1.0 micrometers, and the formation of an ecological corona layer on these walls is contingent upon the abundance and physicochemical properties of biomolecules and plastic particles. The stability of this layer is influenced by hydrogen bonds, van der Waals forces, hydrophobic interactions, and other high-energy chemical or adhesive forces [71]. Research indicates that the zeta potential of fungal cell walls is highly responsive to environmental conditions, generally fluctuating between -14 and -15 millivolts [75]. Changes in environmental pH, along with varying concentrations of ions and proteins, can promote heteroaggregation, which may consequently alter the zeta potential [76]. Exposure of fungal cells to nanoplastics may modulate the zeta potential of their cell walls, thereby affecting their functional integrity and potentially contributing to the toxicity of extracellularly secreted enzymes.
The fungal cell wall serves as the outermost layer, directly interacting with the external environment and playing a critical role in various physiological and ecological functions. It is a primary target for antimicrobial agents and the immune system, requiring a delicate balance of strength and flexibility to provide protection while facilitating nutrient uptake, membrane vesicle exchange, and external signal reception [77]. Previous studies have shown that polymeric particles ranging from 100 nm to 300 nm do not penetrate the cell walls of pathogenic fungi, such as A. fumigatus and C. albicans [78]. Thus, it can be proposed that nanoplastics exceeding 100 nm primarily interact with the surfaces of fungal cell walls, impacting the outer wall polymers and glycoproteins associated with the chitin and β-glucan-based inner wall skeleton. This interaction could disrupt spatial organization and dynamic regulatory functions, impairing the fungal ability to effectively respond to changes in growth conditions and potentially leading to toxicity. Interestingly, certain filamentous fungi may induce a “dusting effect,” wherein high concentrations of nanoplastics allow initially colonizing hyphae to adsorb or internalize these particles into vacuoles, subsequently metabolizing them into less toxic forms. This adaptive response may mitigate toxicity to later-growing hyphae, thereby promoting favorable conditions for fungal proliferation [79]. Previous research in bacterial systems has demonstrated that nanoplastics with diameters of 60 nm can penetrate cells, accumulating internally and enhancing the generation of ROS, which impose stress on bacterial cells and significantly inhibit their growth [80]. It is plausible that nanoplastics of similar sizes may also compromise pathogenic yeast cells, such as C. neoformans, given that ROS can modulate the expression of virulence factors, including capsule and melanin production [81,82]. The toxicological impact of nanoplastics on fungal cells is multifaceted, encompassing redox imbalances, membrane damage, immune responses, and genotoxic effects, which can induce various forms of cellular injury simultaneously. Collectively, nanoplastics have the potential to significantly alter the physiological states of fungi, highlighting the urgent need for further research to elucidate the complex interactions and effects of nanoplastics on fungal ecology.
4. Potential Effects of Nanoplastics on Fungal Drug Resistance
The development of antifungal agents faces significant challenges due to the shared eukaryotic structures and metabolic pathways between humans and fungi, resulting in limited therapeutic options. Fungal infections are increasingly exhibiting resistance to conventional antifungal drugs, with the efficacy of existing treatments, such as azoles and polyenes, diminishing in clinical settings. According to data from the Centers for Disease Control and Prevention, drug-resistant fungal infections were responsible for at least 35,900 deaths in the United States in 2019 [83]. This resistance typically arises through natural selection, often driven by genetic mutations or gene transfer that confer additional resistant traits. Notably, fungi are sensitive to chemical toxicity and demonstrate rapid responses to environmental changes [84]. Recent studies suggest that nanoplastics may play a critical role in promoting antifungal resistance through several mechanisms. For instance, exposure to nanoplastics can trigger stress responses and adaptive mechanisms in fungi. In response to nanoplastic exposure, fungi may activate defense pathways, including oxidative stress responses and efflux pumps, which enhance their resistance to antifungal drugs. For example, Lactarius deliciosus exhibits oxidative stress in the presence of polystyrene, leading to increased secretion of organic acids and enhanced absorption of phosphorus and potassium, although growth is inhibited at high concentrations [85]. Moreover, exposure to nanoplastics has been shown to augment the secretion of extracellular enzymes in fungi, including β-glucosidase, glycine aminopeptidase, and phenol oxidase, thereby altering the community structure [86]. Similar observations in bacterial studies indicate that polystyrene exposure induces oxidative stress, leading to increased synthesis of glutathione and enhanced activity of the tricarboxylic acid (TCA) cycle, as well as of efflux pumps, which subsequently promote growth and resistance in Escherichia coli [87,88]. Nanoplastics may also induce oxidative damage within fungal cells. For instance, low-density polystyrene has been shown to alter the membrane composition of Trichoderma harzianum, resulting in increased membrane permeability and enhanced activities of ROS, superoxide dismutase (SOD), and catalase (CAT) [89]. Recent findings from our research indicate that exposure to nanoplastics can induce ROS production in C. neoformans, disrupting normal cellular functions (unpublished data). Such oxidative damage may drive fungi to develop resistance through mutations or other adaptive changes that enhance their chances of survival. Overall, the interaction of nanoplastics with fungi has the potential to significantly impact drug resistance. The multifaceted stress responses triggered by nanoplastics may not only enhance the ability of fungi to withstand antifungal agents but also promote the evolution of resistance mechanisms. These findings underscore the need for further research to elucidate the complexities of nanoplastic interactions and their implications for fungal pathogenicity and treatment strategies.
5. Conclusions and Future Perspectives
The intricate interactions between nanoplastics and fungi present a dual-edged sword for environmental science and public health. As emerging pollutants, nanoplastics have demonstrated significant effects on fungal physiology, including alterations in metabolic pathways, physiological responses, and even virulence factors. The ability of certain fungal species to degrade plastics offers promising avenues for bioremediation, yet the presence of nanoplastics complicates these interactions by influencing fungal physiological functions and potentially enhancing antifungal resistance mechanisms. This review highlights the necessity for a deeper understanding of the multifaceted relationships between nanoplastics and fungi. The evidence suggests that while fungi can leverage nanoplastics to improve their plastic-degrading capacities, exposure to these pollutants may simultaneously drive adaptations that bolster their resistance to antifungal agents (Figure 1). This paradox emphasizes the urgency of investigating the mechanisms underlying these interactions, as they could have far-reaching implications for both ecological health and clinical outcomes in fungal infections. Future research should focus on several key areas to elucidate the complex dynamics of nanoplastic-fungi interactions. Firstly, studies should aim to identify the specific molecular pathways activated in fungi upon exposure to nanoplastics, particularly regarding oxidative stress responses and enzymatic adaptations. Secondly, the ecological impact of these interactions on fungal communities in various environments, including terrestrial and aquatic ecosystems, warrants further exploration. Long-term studies are essential to assess the implications of chronic nanoplastic exposure on fungal diversity and function. Additionally, advancing our understanding of the link between nanoplastic exposure and antifungal resistance mechanisms is critical. Investigating the potential for genetic mutations and horizontal gene transfer in fungi exposed to nanoplastics could provide insights into the development of resistance traits. This knowledge is particularly vital given the rising incidence of drug-resistant fungal infections that pose substantial public health threats. In conclusion, while the potential of fungi in bioremediation strategies remains promising, the challenges posed by nanoplastics necessitate a comprehensive investigation into their effects on fungal physiology and ecology. By addressing these research gaps, we can develop more effective strategies for managing plastic pollution and mitigating the associated risks to human health and the environment. Continued interdisciplinary collaboration will be crucial in paving the way for innovative solutions to combat the dual challenges of plastic pollution and fungal diseases.
Funding
The work was supported by the grants Natural Science Research of Jiangsu Higher Education Institutions of China (No. 24KJD430010), Natural Science Foundation of Jiangsu Province of China (No. BK20240948), Nantong Jiangsu Scientific Research Project of China (No. JC2023043), and Postgraduate Research & Practice Innovation Program of Jiangsu Province (KYCX24_3537, KYCX24_3538).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Wilcox, C.; Van Sebille, E.; Hardesty, B. D. Threat of plastic pollution to seabirds is global, pervasive, and increasing. Proceedings of the National Academy of Sciences of the United States of America 2015, 12(38), 11899–904. [Google Scholar] [CrossRef] [PubMed]
- Geyer, R.; Jambeck, J. R.; Law, K. L. Production, use, and fate of all plastics ever made. Science advances 2017, 3(7), e1700782. [Google Scholar] [CrossRef] [PubMed]
- Hoseini, M.; Bond, T. Predicting the global environmental distribution of plastic polymers. Environmental pollution (Barking, Essex : 1987) 2022, 300, 118966. [Google Scholar] [CrossRef] [PubMed]
- Cowger, W.; Willis, K. A.; Bullock, S.; Conlon, K.; Emmanuel, J.; Erdle, L. M.; Eriksen, M.; Farrelly, T. A.; Hardesty, B. D.; Kerge, K. Global producer responsibility for plastic pollution. Science advances 2024, 10(17), eadj8275. [Google Scholar] [CrossRef]
- Fotopoulou, K. N.; Karapanagioti, H. K. Degradation of various plastics in the environment. Hazardous chemicals associated with plastics in the marine environment 2019, 71–92. [Google Scholar]
- Zhang, K.; Hamidian, A. H.; Tubić, A.; Zhang, Y.; Fang, J. K.; Wu, C.; Lam, P. K. Understanding plastic degradation and microplastic formation in the environment: A review. Environmental Pollution 2021, 274, 116554. [Google Scholar] [CrossRef]
- Gigault, J.; El Hadri, H.; Nguyen, B.; Grassl, B.; Rowenczyk, L.; Tufenkji, N.; Feng, S.; Wiesner, M. Nanoplastics are neither microplastics nor engineered nanoparticles. Nature nanotechnology 2021, 16(5), 501–507. [Google Scholar] [CrossRef]
- Mitrano, D. M.; Wick, P.; Nowack, B. Placing nanoplastics in the context of global plastic pollution. Nature Nanotechnology 2021, 16(5), 491–500. [Google Scholar] [CrossRef]
- Sui, A.; Yao, C.; Chen, Y.; Li, Y.; Yu, S.; Qu, J.; Wei, H.; Tang, J.; Chen, G. Polystyrene nanoplastics inhibit StAR expression by activating HIF-1α via ERK1/2 MAPK and AKT pathways in TM3 Leydig cells and testicular tissues of mice. Food and chemical toxicology : an international journal published for the British Industrial Biological Research Association 2023, 173, 113634. [Google Scholar] [CrossRef]
- Bu, W.; Yu, M.; Ma, X.; Shen, Z.; Ruan, J.; Qu, Y.; Huang, R.; Xue, P.; Ma, Y.; Tang, J.; Zhao, X. Gender-specific effects of prenatal polystyrene nanoparticle exposure on offspring lung development. Toxicology letters 2025, 407, 1–16. [Google Scholar] [CrossRef]
- Rodrigues, M. O.; Abrantes, N.; Gonçalves, F. J. M.; Nogueira, H.; Marques, J. C.; Gonçalves, A. M. M. Impacts of plastic products used in daily life on the environment and human health: What is known? Environmental toxicology and pharmacology 2019, 72, 103239. [Google Scholar] [CrossRef] [PubMed]
- Ryu, H. W.; Kim, D. H.; Jae, J.; Lam, S. S.; Park, E. D.; Park, Y. K. Recent advances in catalytic co-pyrolysis of biomass and plastic waste for the production of petroleum-like hydrocarbons. Bioresource technology 2020, 310, 123473. [Google Scholar] [CrossRef] [PubMed]
- Temporiti, M. E. E.; Nicola, L.; Nielsen, E.; Tosi, S. Fungal enzymes involved in plastics biodegradation. Microorganisms 2022, 10(6), 1180. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S. S.; Ionescu, D.; Grossart, H.-P. Tapping into fungal potential: Biodegradation of plastic and rubber by potent Fungi. Science of The Total Environment 2024, 934, 173188. [Google Scholar] [CrossRef]
- Yang, W.-K.; Gong, Z.; Wang, B.-T.; Hu, S.; Zhuo, Y.; Jin, C.-Z.; Jin, L.; Lee, H.-G.; Jin, F.-J. Biodegradation of low-density polyethylene by mixed fungi composed of Alternaria sp. and Trametes sp. isolated from landfill sites. BMC microbiology 2024, 24(1), 321. [Google Scholar] [CrossRef]
- Denning, D. W. Global incidence and mortality of severe fungal disease. The Lancet Infectious Diseases 2024. [Google Scholar] [CrossRef]
- Kriegl, L.; Egger, M.; Boyer, J.; Hoenigl, M.; Krause, R. New treatment options for critically important WHO fungal priority pathogens. Clinical Microbiology and Infection 2024. [Google Scholar] [CrossRef]
- Vaksmaa, A.; Polerecky, L.; Dombrowski, N.; Kienhuis, M. V.; Posthuma, I.; Gerritse, J.; Boekhout, T.; Niemann, H. Polyethylene degradation and assimilation by the marine yeast Rhodotorula mucilaginosa. ISME communications 2023, 3(1), 68. [Google Scholar] [CrossRef]
- Cowan, A. R.; Costanzo, C. M.; Benham, R.; Loveridge, E. J.; Moody, S. C. Fungal bioremediation of polyethylene: Challenges and perspectives. Journal of Applied Microbiology 2022, 132(1), 78–89. [Google Scholar] [CrossRef]
- Maity, S.; Banerjee, S.; Biswas, C.; Guchhait, R.; Chatterjee, A.; Pramanick, K. Functional interplay between plastic polymers and microbes: a comprehensive review. Biodegradation 2021, 32(5), 487–510. [Google Scholar] [CrossRef]
- Okal, E. J.; Heng, G.; Magige, E. A.; Khan, S.; Wu, S.; Ge, Z.; Zhang, T.; Mortimer, P. E.; Xu, J. Insights into the mechanisms involved in the fungal degradation of plastics. Ecotoxicology and environmental safety 2023, 262, 115202. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, C. Fungal potential for the degradation of petroleum-based polymers: An overview of macro-and microplastics biodegradation. Biotechnology advances 2020, 40, 107501. [Google Scholar] [CrossRef] [PubMed]
- Dix, N. J.; Webster, J.; Dix, N. J.; Webster, J. Fungi of extreme environments. Fungal ecology 1995, 322–340. [Google Scholar]
- Aranda, E. Promising approaches towards biotransformation of polycyclic aromatic hydrocarbons with Ascomycota fungi. Current opinion in biotechnology 2016, 38, 1–8. [Google Scholar] [CrossRef]
- Deshmukh, R.; Khardenavis, A. A.; Purohit, H. J. Diverse metabolic capacities of fungi for bioremediation. Indian journal of microbiology 2016, 56(3), 247–264. [Google Scholar] [CrossRef]
- Backa, S.; Gierer, J.; Reitberger, T.; Nilsson, T. Hydroxyl radical activity associated with the growth of white-rot fungi. 1993. [Google Scholar] [CrossRef]
- Lawton, L. A.; Robertson, P. K. Physico-chemical treatment methods for the removal of microcystins (cyanobacterial hepatotoxins) from potable waters. Chemical Society Reviews 1999, 28(4), 217–224. [Google Scholar] [CrossRef]
- Young, D.; Rice, J.; Martin, R.; Lindquist, E.; Lipzen, A.; Grigoriev, I.; Hibbett, D. Degradation of bunker C fuel oil by white-rot fungi in sawdust cultures suggests potential applications in bioremediation. PloS one 2015, 10(6), e0130381. [Google Scholar] [CrossRef]
- Hasan, I. F. Role of filamentous fungi to remove petroleum hydrocarbons from the environment. Microbial Action on Hydrocarbons 2018, 567–580. [Google Scholar]
- Mir-Tutusaus, J. A.; Baccar, R.; Caminal, G.; Sarrà, M. Can white-rot fungi be a real wastewater treatment alternative for organic micropollutants removal? A review. Water research 2018, 138, 137–151. [Google Scholar] [CrossRef]
- Odigbo, C.; Adenipekun, C.; Oladosu, I.; Ogunjobi, A. Polyethylene terephthalate (PET) biodegradation by Pleurotus ostreatus and Pleurotus pulmonarius. Environmental Monitoring and Assessment 2023, 195(5), 585. [Google Scholar] [CrossRef] [PubMed]
- Espinosa-Valdemar, R. M.; Turpin-Marion, S.; Delfín-Alcalá, I.; Vázquez-Morillas, A. Disposable diapers biodegradation by the fungus Pleurotus ostreatus. Waste management (New York, N.Y.) 2011, 31(8), 1683-8. [Google Scholar] [CrossRef] [PubMed]
- Wolski, E. A.; Durruty, I.; Haure, P. M.; González, J. F. Penicillium chrysogenum: phenol degradation abilities and kinetic model. Water, Air, & Soil Pollution 2012, 223, 2323–2332. [Google Scholar]
- Ogunmolu, F. E.; Kaur, I.; Gupta, M.; Bashir, Z.; Pasari, N.; Yazdani, S. S. Proteomics Insights into the Biomass Hydrolysis Potentials of a Hypercellulolytic Fungus Penicillium funiculosum. Journal of proteome research 2015, 14(10), 4342–58. [Google Scholar] [CrossRef]
- Sowmya, H.V.; Ramalingappa; Krishnappa, M.; Thippeswamy, B. Degradation of polyethylene by Penicillium simplicissimum isolated from local dumpsite of Shivamogga district. Environment, Development and Sustainability 2015, 17(4), 731–745. [Google Scholar] [CrossRef]
- Safdar, A.; Ismail, F.; Imran, M. Biodegradation of synthetic plastics by the extracellular lipase of Aspergillus niger. Environmental Advances 2024, 17, 100563. [Google Scholar] [CrossRef]
- Maeda, H.; Yamagata, Y.; Abe, K.; Hasegawa, F.; Machida, M.; Ishioka, R.; Gomi, K.; Nakajima, T. Purification and characterization of a biodegradable plastic-degrading enzyme from Aspergillus oryzae. Applied microbiology and biotechnology 2005, 67(6), 778–88. [Google Scholar] [CrossRef]
- El-Dash, H. A.; Yousef, N. E.; Aboelazm, A. A.; Awan, Z. A. Optimizing Eco-Friendly Degradation of Polyvinyl Chloride (PVC) Plastic Using Environmental Strains of Malassezia Species and Aspergillus fumigatus. 2023, 24(20). [Google Scholar] [CrossRef]
- Marchut-Mikolajczyk, O.; Kwapisz, E.; Wieczorek, D.; Antczak, T. Biodegradation of diesel oil hydrocarbons enhanced with Mucor circinelloides enzyme preparation. International Biodeterioration & Biodegradation 2015, 104, 142–148. [Google Scholar]
- Al Mousa, A. A.; Hassane, A. M. A.; Gomaa, A. E.-R. F.; Aljuriss, J. A.; Dahmash, N. D.; Abo-Dahab, N. F. Response-Surface Statistical Optimization of Submerged Fermentation for Pectinase and Cellulase Production by Mucor circinelloides and M. hiemalis. Fermentation 2022, 8(5), 205. [Google Scholar] [CrossRef]
- Vršanská, M.; Voběrková, S.; Jiménez Jiménez, A. M.; Strmiska, V.; Adam, V. Preparation and Optimisation of Cross-Linked Enzyme Aggregates Using Native Isolate White Rot Fungi Trametes versicolor and Fomes fomentarius for the Decolourisation of Synthetic Dyes. International journal of environmental research and public health 2017, 15(1). [Google Scholar] [CrossRef] [PubMed]
- Arora, D. S.; Sandhu, D. K. , Laccase production and wood degradation byTrametes hirsuta. Folia Microbiologica 1984, 29(4), 310–315. [Google Scholar] [CrossRef]
- Kersten, P.; Cullen, D. Extracellular oxidative systems of the lignin-degrading Basidiomycete Phanerochaete chrysosporium. Fungal genetics and biology : FG & B 2007, 44(2), 77–87. [Google Scholar]
- Kobayashi, N.; Wada, N.; Yokoyama, H.; Tanaka, Y.; Suzuki, T.; Habu, N.; Konno, N. Extracellular enzymes secreted in the mycelial block of Lentinula edodes during hyphal growth. 2023, 13(1), 36. [Google Scholar] [CrossRef]
- Zavarzina, A. G.; Lisov, A. V.; Leontievsky, A. The Role of Ligninolytic Enzymes Laccase and a Versatile Peroxidase of the White-Rot Fungus Lentinus tigrinus in Biotransformation of Soil Humic Matter: Comparative In Vivo Study. Journal of Geophysical Research: Biogeosciences 2018, 123, 2727–2742. [Google Scholar] [CrossRef]
- Covino, S.; Cvancarová, M.; Muzikár, M.; Svobodová, K.; D’Annibale, A.; Petruccioli, M.; Federici, F.; Kresinová, Z.; Cajthaml, T. An efficient PAH-degrading Lentinus (Panus) tigrinus strain: effect of inoculum formulation and pollutant bioavailability in solid matrices. Journal of hazardous materials 2010, 183(1-3), 669-76. [Google Scholar] [CrossRef]
- Case, N. T.; Gurr, S. J.; Fisher, M. C.; Blehert, D. S.; Boone, C.; Casadevall, A.; Chowdhary, A.; Cuomo, C. A.; Currie, C. R.; Denning, D. W. Fungal impacts on Earth’s ecosystems. Nature 2025, 638(8049), 49–57. [Google Scholar] [CrossRef]
- Magan, N. Fungi in extreme environments. The Mycota 2007, 4, 85–103. [Google Scholar]
- Mancera-López, M.; Esparza-García, F.; Chávez-Gómez, B.; Rodríguez-Vázquez, R.; Saucedo-Castañeda, G.; Barrera-Cortés, J. Bioremediation of an aged hydrocarbon-contaminated soil by a combined system of biostimulation–bioaugmentation with filamentous fungi. International Biodeterioration & Biodegradation 2008, 61(2), 151–160. [Google Scholar]
- Durairaj, P.; Malla, S.; Nadarajan, S. P.; Lee, P.-G.; Jung, E.; Park, H. H.; Kim, B.-G.; Yun, H. Fungal cytochrome P450 monooxygenases of Fusarium oxysporum for the synthesis of ω-hydroxy fatty acids in engineered Saccharomyces cerevisiae. Microbial Cell Factories 2015, 14, 1–16. [Google Scholar] [CrossRef]
- Asemoloye, M. D.; Jonathan, S. G.; Ahmad, R. Synergistic plant-microbes interactions in the rhizosphere: a potential headway for the remediation of hydrocarbon polluted soils. International journal of phytoremediation 2019, 21(2), 71–83. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, R. Cytochromes P450 as versatile biocatalysts. Journal of biotechnology 2006, 124(1), 128–145. [Google Scholar] [CrossRef] [PubMed]
- Van Beilen, J. B.; Funhoff, E. G. Alkane hydroxylases involved in microbial alkane degradation. Applied microbiology and biotechnology 2007, 74, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D. E.; Kraševec, N.; Mullins, J.; Nelson, D. R. The CYPome (cytochrome P450 complement) of Aspergillus nidulans. Fungal genetics and biology 2009, 46(1), S53–S61. [Google Scholar] [CrossRef]
- Moktali, V.; Park, J.; Fedorova-Abrams, N. D.; Park, B.; Choi, J.; Lee, Y.-H.; Kang, S. Systematic and searchable classification of cytochrome P450 proteins encoded by fungal and oomycete genomes. BMC genomics 2012, 13, 1–13. [Google Scholar] [CrossRef]
- Chen, W.; Lee, M.-K.; Jefcoate, C.; Kim, S.-C.; Chen, F.; Yu, J.-H. Fungal cytochrome p450 monooxygenases: their distribution, structure, functions, family expansion, and evolutionary origin. Genome biology and evolution 2014, 6(7), 1620–1634. [Google Scholar] [CrossRef]
- Dauda, W. P.; Abraham, P.; Glen, E.; Adetunji, C. O.; Ghazanfar, S.; Ali, S.; Al-Zahrani, M.; Azameti, M. K.; Alao, S. E. L.; Zarafi, A. B. Robust Profiling of Cytochrome P450s (P450ome) in Notable Aspergillus spp. Life 2022, 12(3), 451. [Google Scholar] [CrossRef]
- Doddapaneni, H.; Subramanian, V.; Fu, B.; Cullen, D. A comparative genomic analysis of the oxidative enzymes potentially involved in lignin degradation by Agaricus bisporus. Fungal genetics and biology 2013, 55, 22–31. [Google Scholar] [CrossRef]
- Ekanayaka, A. H.; Tibpromma, S.; Dai, D.; Xu, R.; Suwannarach, N.; Stephenson, S. L.; Dao, C.; Karunarathna, S. C. A review of the fungi that degrade plastic. Journal of fungi 2022, 8(8), 772. [Google Scholar] [CrossRef]
- Viel, T.; Manfra, L.; Zupo, V.; Libralato, G.; Cocca, M.; Costantini, M. Biodegradation of plastics induced by marine organisms: future perspectives for bioremediation approaches. Polymers 2023, 15(12), 2673. [Google Scholar] [CrossRef]
- Munir, E.; Harefa, R.; Priyani, N.; Suryanto, D. In Plastic degrading fungi Trichoderma viride and Aspergillus nomius isolated from local landfill soil in Medan, IOP conference series: Earth and environmental science, 2018; IOP Publishing: 2018; p 012145.
- Satti, S. M.; Shah, A. A.; Auras, R.; Marsh, T. L. Isolation and characterization of bacteria capable of degrading poly (lactic acid) at ambient temperature. Polymer Degradation and Stability 2017, 144, 392–400. [Google Scholar] [CrossRef]
- El-Morsy, E.; Hassan, H.; Ahmed, E. Biodegradative activities of fungal isolates from plastic contaminated soils. Mycosphere 2017, 8(8), 1071–1087. [Google Scholar] [CrossRef]
- Janczak, K.; Hrynkiewicz, K.; Znajewska, Z.; Dąbrowska, G. Use of rhizosphere microorganisms in the biodegradation of PLA and PET polymers in compost soil. International Biodeterioration & Biodegradation 2018, 130, 65–75. [Google Scholar]
- Ojha, N.; Pradhan, N.; Singh, S.; Barla, A.; Shrivastava, A.; Khatua, P.; Rai, V.; Bose, S. Evaluation of HDPE and LDPE degradation by fungus, implemented by statistical optimization. Scientific Reports 2017, 7(1), 39515. [Google Scholar] [CrossRef]
- Artham, T.; Doble, M. Biodegradation of physicochemically treated polycarbonate by fungi. Biomacromolecules 2010, 11(1), 20–28. [Google Scholar] [CrossRef]
- do Paço, A. M. S. Biotechnological Tools for Plastic Waste Remediation Using Marine Fungi. Universidade de Aveiro (Portugal), 2024.
- Gao, R.; Liu, R.; Sun, C. A marine fungus Alternaria alternata FB1 efficiently degrades polyethylene. Journal of Hazardous Materials 2022, 431, 128617. [Google Scholar] [CrossRef]
- Ameen, F.; Moslem, M.; Hadi, S.; Al-Sabri, A. E. Biodegradation of Low Density Polyethylene (LDPE) by Mangrove fungi from the red sea coast. Progress in Rubber Plastics and Recycling Technology 2015, 31(2), 125–143. [Google Scholar] [CrossRef]
- Sarkhel, R.; Sengupta, S.; Das, P.; Bhowal, A. Comparative biodegradation study of polymer from plastic bottle waste using novel isolated bacteria and fungi from marine source. Journal of Polymer Research 2020, 27, 1–8. [Google Scholar] [CrossRef]
- Liu, S.; Junaid, M.; Liao, H.; Liu, X.; Wu, Y.; Wang, J. Eco-corona formation and associated ecotoxicological impacts of nanoplastics in the environment. The Science of the total environment 2022, 836, 155703. [Google Scholar] [CrossRef]
- Junaid, M.; Wang, J. Interaction of nanoplastics with extracellular polymeric substances (EPS) in the aquatic environment: A special reference to eco-corona formation and associated impacts. Water research 2021, 201, 117319. [Google Scholar] [CrossRef]
- Hüffer, T.; Hofmann, T. Sorption of non-polar organic compounds by micro-sized plastic particles in aqueous solution. Environmental pollution (Barking, Essex : 1987) 2016, 214, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zhang, M.; Sha, W.; Wang, Y. Sorption Behavior and Mechanisms of Organic Contaminants to Nano and Microplastics. 2020, 25(8). [Google Scholar] [CrossRef] [PubMed]
- Ramos, M. M.; Dos, S. M. E.; da, S. S. I.; Lima, A. L.; de Oliveira, F. R.; de Freitas, C. M.; Fernandes, C. P.; de Carvalho, J. C. T.; Ferreira, I. M. Silver nanoparticle from whole cells of the fungi Trichoderma spp. isolated from Brazilian Amazon. 2020, 42(5), 833–843. [Google Scholar]
- Mikolajczyk, A.; Gajewicz, A.; Rasulev, B.; Schaeublin, N.; Maurer-Gardner, E.; Hussain, S.; Leszczynski, J.; Puzyn, T. Zeta Potential for Metal Oxide Nanoparticles: A Predictive Model Developed by a Nano-Quantitative Structure–Property Relationship Approach. Chemistry of Materials 2015, 27(7), 2400–2407. [Google Scholar] [CrossRef]
- Gow, N. A. R.; Lenardon, M. D. Architecture of the dynamic fungal cell wall. 2023, 21(4), 248–259. [Google Scholar] [CrossRef]
- Orasch, T.; Gangapurwala, G.; Vollrath, A.; González, K.; Alex, J.; De San Luis, A.; Weber, C.; Hoeppener, S.; Cseresnyés, Z.; Figge, M. T.; Guerrero-Sanchez, C.; Schubert, U. S.; Brakhage, A. A. Polymer-based particles against pathogenic fungi: A non-uptake delivery of compounds. Biomaterials advances 2023, 146, 213300. [Google Scholar] [CrossRef]
- Mafla-Endara, P. M.; Meklesh, V.; Beech, J. P.; Ohlsson, P.; Pucetaite, M.; Hammer, E. C. Exposure to polystyrene nanoplastics reduces bacterial and fungal biomass in microfabricated soil models. The Science of the total environment 2023, 904, 166503. [Google Scholar] [CrossRef]
- Dai, S.; Ye, R.; Huang, J.; Wang, B.; Xie, Z.; Ou, X.; Yu, N.; Huang, C.; Hua, Y.; Zhou, R.; Tian, B. Distinct lipid membrane interaction and uptake of differentially charged nanoplastics in bacteria. 2022, 20(1), 191. [Google Scholar] [CrossRef]
- Zaragoza, O.; Chrisman, C. J.; Castelli, M. V.; Frases, S.; Cuenca-Estrella, M.; Rodríguez-Tudela, J. L.; Casadevall, A. Capsule enlargement in Cryptococcus neoformans confers resistance to oxidative stress suggesting a mechanism for intracellular survival. Cellular microbiology 2008, 10(10), 2043–57. [Google Scholar] [CrossRef]
- Momin, M.; Webb, G. The Environmental Effects on Virulence Factors and the Antifungal Susceptibility of Cryptococcus neoformans. Int J Mol Sci 2021, 22(12). [Google Scholar] [CrossRef]
- Prevention, U. S. C. f. D. C. a. Antibiotic resistance threats in the United States. 2019. [Google Scholar]
- Shruti, V. C.; Kutralam-Muniasamy, G.; Pérez-Guevara, F. Do microbial decomposers find micro- and nanoplastics to be harmful stressors in the aquatic environment? A systematic review of in vitro toxicological research. Science of The Total Environment 2023, 903, 166561. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Gao, B. Physiological response of ectomycorrhizal fungi (Lactarius delicious) to microplastics stress. Global Nest Journal 2023, 25(3), 115–124. [Google Scholar]
- Du, J.; Qv, W.; Niu, Y.; Qv, M.; Jin, K.; Xie, J.; Li, Z. Nanoplastic pollution inhibits stream leaf decomposition through modulating microbial metabolic activity and fungal community structure. Journal of hazardous materials 2022, 424 Pt A, 127392. [Google Scholar] [CrossRef]
- Fang, S.; Huang, Y.; Xiang, Z.; Zeng, R.; Zeng, S.; Liu, S. Polystyrene nanoplastics foster Escherichia coli O157:H7 growth and antibiotic resistance with a stimulating effect on metabolism. Environmental Science: Nano 2023, 10(5), 1341–1351. [Google Scholar] [CrossRef]
- Liu, X.; Xu, R.; Wu, H.; Xu, K.; Zhang, W.; Wang, Z.; Gong, W. Nanoplastics promote the dissemination of antibiotic resistance through conjugative gene transfer: implications from oxidative stress and gene expression. Environmental Science: Nano 2023, 10(5), 1329–1340. [Google Scholar] [CrossRef]
- Jasińska, A.; Różalska, S. Microplastic-Induced Oxidative Stress in Metolachlor-Degrading Filamentous Fungus Trichoderma harzianum. 2022, 23(21). [Google Scholar] [CrossRef]
Figure 1.
Schematic summary model illustrating the intricate interactions between fungi and nanoplastics, focusing on biodegradation, pathogenicity, and resistance.
Figure 1.
Schematic summary model illustrating the intricate interactions between fungi and nanoplastics, focusing on biodegradation, pathogenicity, and resistance.
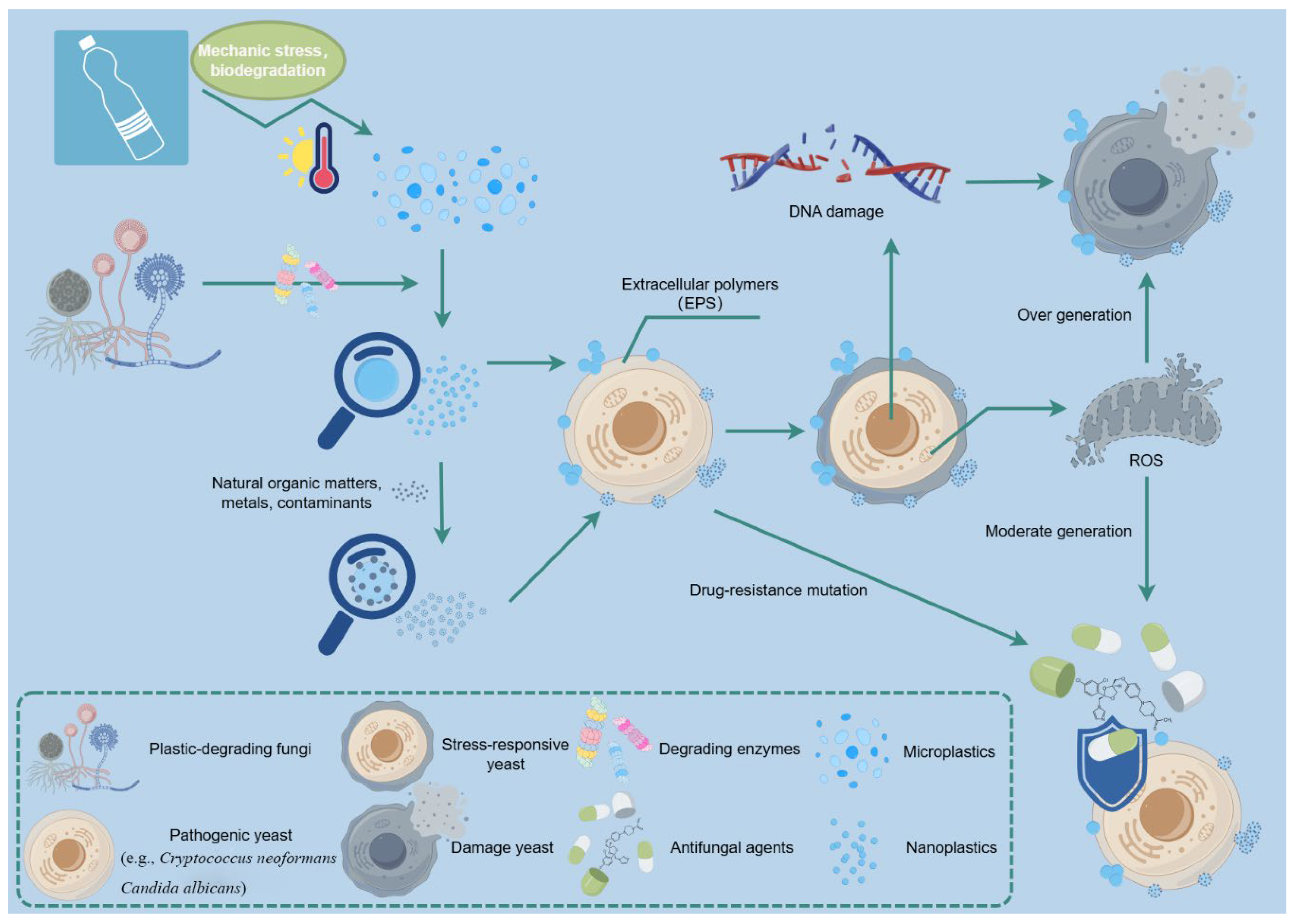

Table 1.
Key fungal groups and their enzymatic roles in plastic degradation.
| Genus | Representative Species | Enzymes | Features | References |
|---|---|---|---|---|
| Ganoderma | Ganoderma lucidum; G. applanatum | Lignin peroxidase, manganese peroxidase, laccase, polyesterase | Degradation of plastics and other organic materials | [30] |
| Pleurotus | Pleurotus abalones; P. ostreatus | Cellulases, lignin peroxidase, manganese peroxidase, laccase | Capable of degrading polylactic acid (PLA), polyethylene (PE), polypropylene (PP), and polyester plastics such as PET | [31,32] |
| Penicillium | Penicillium chrysogenum; P. funiculosum; P. simplicissimum | Polyesterases, esterases, ligninases, cellulases, phenol oxidase | Strains of Penicillium capable of degrading plastics such as polyurethane and polyethylene | [33,34,35] |
| Aspergillus | Aspergillus niger; A. oryzae; A. fumigatus | Esterases, polyesterases, cellulases, phenol oxidase, ligninases | Capable of degrading polyester plastics, such as PET | [36,37,38] |
| Mucor | Mucor circinelloides; M. hiemalis | Polyesterases, esterases, oxidases, cellulases | Capable of degrading polyester plastics | [39,40] |
| Trametes | Trametes versicolor; T. hirsuta | Lignin peroxidase, manganese peroxidase, laccase, polyesterase | Effective in plastic degradation, especially for polymers like polyurethane and polyethylene | [41,42] |
| Phanerochaete | Phanerochaete chrysosporium | Lignin peroxidase, manganese peroxidase, laccase, polyesterase | Degradation of lignin and certain plastics | [43] |
| Lentinus | Lentinus edodes; L. tigrinus | Lignin peroxidase, manganese peroxidase, laccase, cellulase | Effective in degrading lignin and some synthetic materials | [44,45,46] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



