Submitted:
28 March 2025
Posted:
31 March 2025
You are already at the latest version
Abstract
The emergence and germination of two oak species, Quercus faginea and Quercus coccifera, were evaluated in a nursery to assess their response to acorn dimensions and different substrates. Three substrates were tested: a commercial substrate, coconut fiber with ver-miculite, and olive pomace compost with two different proportions of sand. Olive pomace compost, a byproduct of olive oil production, has recently gained attention as a promising alternative substrate for nurseries, offering a sustainable solution for managing this mate-rial while promoting plant growth. Seed weight and physical characteristics were evalu-ated for both species, as well as their relationships to radicle and shoot emergence and germination. The results demonstrated a significantly higher and earlier emergence (root and shoot) and germination in the commercial substrate. Quercus faginea germination was faster than that of Quercus coccifera, seed dimension did not have a significant effect on germination time, and seedling growth was promoted in coconut fiber and vermiculite substrate.
Keywords:
seedling
; oak species
; emergence
; nursery
; olive pomace compost
1. Introduction
Species within the genus Quercus are of significant ecological, economic, and societal value, providing timber, fruit and ecosystem services such as tourism, carbon sequestration, and conservation of habitats for endemic species, while reducing wildfire and soil erosion risks [1]. Among these species are Q. faginea and Q. coccifera, which have received less research attention compared to other Quercus species. Quercus faginea (Portuguese oak) is a species within the Quercus genus and the Fagaceae family [4]. It is deciduous and is typically found in temperate regions of southern Europe and North Africa, including Portugal, Spain, Morocco, Algeria, Tunisia, and southern France [5]. In Portugal, Q. faginea occurs along the central-southern coastal regions of the Iberian Peninsula, particularly in the transitional areas of the Atlantic climate. The Portuguese oak can grow at altitudes ranging from 500 to 1500 m [2]. It is not demanding in terms of soil properties and can be found in a variety of soils, ranging from acidic siliceous soils to neutral or basic calcareous soils [3]. Q. faginea occurs both as a pure and in mixed stands with other oaks such as Q. pyrenaica, Q. suber, Q. ilex and Q. coccifera [4]. These mixed oak stands contribute to greater species richness, playing a vital role in promoting biodiversity [5]. Quercus coccifera L. (kermes oak) is an evergreen sclerophyll species native to the Mediterranean region and the North African Maghreb [6]. Its distribution extends from Portugal across southern Europe to Cyprus and Turkey, including Morocco, Libya, France, Spain, Italy, the Balkans, mainland Greece, and numerous Greek islands [7]. This species is a dominant component of semiarid shrubland ecosystems (garrigues) and is commonly used for reforestation in areas with harsh climatic conditions, poor and shallow soils, and recurrent disturbances such as fire or grazing [8].
Seed regeneration depends on successful seed production, which is influenced by years of good fruiting and by the density and spatial distribution of the trees in the forest stands [9,10]. To ensure the success of natural regeneration, controlling seed dispersal is essential, as seeds can suffer significant losses both on the tree and on the ground due to various predators, such as wild boars, deer, rodents, birds, insects, and herding [3]. As a result, seeding or planting is often required to guarantee the success of regeneration [11]. In the nursery, several factors can significantly influence seedling production, including the control of the growth environment, temperature, moisture, container type of substrate and fertilisation [12]. The type of containers used to produce forest plants in nurseries plays a critical role in the success of germination. Since the root system of the Quercus species grows extensively after germination, potentially reaching up to one meter in depth, its cultivation is typically carried out in rigid containers with internal vertical ribs to direct roots downward and prevent spiral growth [13]. Additionally, these containers are open-bottomed to promote natural root pruning [14]. Substrates are fundamental in nurseries due to their capacity to retain moisture, a critical environmental factor for recalcitrant seeds like acorns. Moisture retention directly influences tree growth, photosynthetic activity, nutrient availability, and resilience to moisture deficiency [15]. Furthermore, the type of substrate impacts respiration and transpiration in container-grown seedlings, affecting key physiological mechanisms such as photosynthetic capacity, stomatal conductivity, and water-use efficiency [16].
The seed reserves, more specifically the seed size, influence water uptake (where larger seeds are typically less permeable) and the seed coat plays an important role in regulating the water absorption process [17], the vigor of the embryo and the availability of sufficient nutrients in the endosperm ensure germination. However, for subsequent growth, the nutritional value of the substrate and its moisture content are also crucial [18]. In fact, in recent years, studies have focused on developing substrates to meet the needs of forest plants and enhance their resilience to plantation shock [19,20]. Substrate composition remains challenging in forest nurseries, with traditional soil and sand mixtures often being of poor quality [21]. A study on Q. suber by Kholkhal and Benmahioul [21] found that potting soil substrates yielded the highest germination rates, likely due to their superior water and oxygen retention, which supports enzymatic activity crucial for seed germination [22,23].
Building on the search for improved substrates, recent attention has turned to the potential of olive compost and/or olive pomace (a by-product derived from the recycling of olive oil production residues) as an alternative substrate with promising properties for plant growth. Depending on the extraction methods, olive pomace compost (OPC) can have different percentages of moisture, consisting of fragments of olive pit, skin, pulp, and residual oil [24], being rich in polyphenols, tocopherols, proteins, and squalene [25]. Moreover, these residues have a high content of cellulose, hemicellulose, and lignin, making them a valuable source of fermentable sugars and xylan, as well as a potential alternative substrate [26]. Some studies [27,28] demonstrated that OPC improves the physical and chemical characteristics of the soil, and provides nutrients such as nitrogen, potassium and phosphorus, for plant growth.
In this context, this study aims to compare the effects of three organic substrates on the development of Q. faginea and Q. coccifera seedlings in a nursery environment, with a particular focus on their influence on acorn emergence, germination and seedling development. The substrates tested include a mixture of coconut fiber and vermiculite, which is commonly used in nursery practices, alongside OPC and sand with two compositions, the primary substrate under investigation. The questions addressed in this study were: a) Do both Quercus species present the same germination and growth trends?; b) Do acorn dimensions influence germination?; c) Is seedling growth influenced by substrate?; d) Is germination in the substrate coconut fiber and vermiculite similar to that of OPC?
2. Materials and Methods
2.1. Seed Collection
Seeds of Q. faginea and Q. coccifera were collected in November 2024 from one forest stand located in Serra dos Candeeiros, Portugal, approximately 90 km northeast of Lisbon (central coordinates 39°28'50'' N, 8°53'30'' W) at an altitude of 630 m, with an annual average precipitation of 800–1200 mm and an annual average temperature of 15–16°C. The acorns were stored at a temperature of 2–4 °C until the start of the experiments.
The experiment was conducted in a nursery located at the University of Évora, Herdade da Mitra, Évora, Portugal. The nursery is enclosed by thermal polyethylene walls, with an east-west orientation, ventilated by windows placed laterally on both sides and equipped with cooling and heating systems, used in summer and winter, respectively.
2.2. Acorn Germination
Before the germination assay, the acorns were subjected to a visual inspection to detect any external damage, fungal infections, or deformities, followed by a flotation test to identify seeds with compromised viability. Any damaged seeds identified during these procedures were excluded from the study. Physical variables, including acorn weight (g), length (mm), and width (mm), were measured before placing the acorns on the substrates under study. Measurements were recorded using a Digital Caliper Series 1108 from INSIZE, with a resolution of 0.01 mm, and a precision scale ADP 720L for weight determination.
For this experiment, individual acorns were placed in the cells of plastic root trainer containers, measuring 50 x 29 x 16 cm. Each container was composed of 28 square cells measuring 6 × 6 x 16 cm, having a volume of 576 cm³. The containers were perforated with small holes to facilitate the removal of excess moisture from watering. The cells were filled with three different substrate compositions (cf. 2.3). The acorns were placed on top of the substrate, with the upper portion of each acorn 2-3 mm below the edge of the container cells. The acorns were exposed to natural light. Emergency (root and shoot), germination and seedling growth were recorded every seven days over 49 days. A total of 168 acorns were used in this experiment, with 28 acorns per species and per substrate. No fertilization was applied to the young seedlings.
2.3. Substrates Characterization
For this trial, three types of substrates were used: 1) olive pomace compost and sand in a ratio of 75/25% (OPC75/S), 2) olive pomace compost and sand in a ratio of 50/50% (OPC50/S); and 3) coconut fiber and vermiculite in a ratio of 80/20% (CF/V). The commercial substrate coconut (Neoquímica, Portugal) is characterized by having a pH ranging from 5.5 to 6.5.
The OPC was sourced within the PRR INOVCIRCOLIVE project, funded by the Ministry of Agriculture, and corresponds to the 2023/2024 harvest. It was produced through the composting of two-phase olive pomace (45%), cattle manure (21%), and olive leaves and twigs (33%), expressed as a percentage of dry matter. The OPC has a pH of 9 [29].
2.4. Statistical Analysis
Variance and correlation analysis were used. The normality was assessed with the Shapiro–Wilk test and the homogeneity of variance using Levene’s test. Differences between the measured variables were determined by one-way ANOVA. When the assumptions for normality and homogeneity of variance of the variables were not met, data analysis was performed on ranks using the Kruskal–Wallis one-way. Multivariate correlation analysis was conducted using the non-parametric Spearman’s rank correlation coefficient. The statistical analysis was implemented in IBM SPSS Statistics 28.0.0.0 [30] at a significance level of α = 0.05.
3. Results
3.1. Physical Characterization of Acorns
Significant differences were observed in acorn weight and acorn width between the two species analyzed (p-value = 0.001), with Q. faginea having a significantly higher mean weight and width than Q. coccifera (Figure 1, Table 1).
A wide variation in the weight and size of acorns was observed between acorns of Q. faginea and Q. coccifera (Table 1). For both species, acorn weights ranged from 1.78 to 8.34 g, indicating significant variability. The width of Q. faginea acorns exhibited significantly larger dimensions, with a minimum of 13.86 mm compared to 11.16 mm in Q. coccifera. However, in Q. coccifera, the acorn length was smaller, but no significant differences were observed, with a minimum of 25.4 mm for Q. coccifera and 27.75 mm for Q. faginea.
3.2. Root Emergence and Germination
Root emergence was observed after 7 days of placing the acorns in the substrates in both species under study. The CF/V yielded higher values in both species (Figure 2). However, in Q. faginea, no significant differences were observed between substrates. The root emergence in Q. coccifera, for the two substrates with different proportions of OPC did not show significant differences but was significantly higher in the CF/V substrate (Figure 2 top).
Regarding germination rates (Figure 2 bottom), significant differences were observed both between the substrates and between species. OPC substrates exhibited lower germination rates compared to the CF/V substrate, especially in Q. faginea. The OPC50/S substrate resulted in significantly higher germination rates in Q. coccifera (34.4%) compared to Q. faginea (17.9%). The OPC75/S substrate showed the lowest germination rates, with 19.6% for Q. faginea and 25.9% for Q. coccifera. The CF/V substrate obtained the best germination, with 53.7% for Q. faginea and 74.1% for Q. coccifera. Significant differences were also observed in germination rates between the species, with Q. coccifera achieving higher germination rates compared to Q. faginea, particularly in CF/V and OPC50/S substrates.
3.3. Shoot Emergence Time
The acorns of Q. faginea began shoot emergence on day 14 after seeding, whereas Q. coccifera only exhibited shoot emergence on day 28 after seeding (Figure 3).
Shoot emergence in Q. faginea began 14 days after seeding; however, the highest number of shoot emergences was recorded at 28 and 35 days after seeding, 31.4 e 41.2%, respectively (Figure 4). The lowest shoot emergence occurred at 21 days after seeding (7.8%). In contrast, shoot emergence in Q. coccifera started two weeks later, with the first shoots appearing 28 days after seeding (23.2%). Similar to Q. faginea, the highest shoot emergence was recorded at 35 days after seeding. By day 42 after seeding, a decrease in shoot emergence was observed in both species. It was also checked whether new shoot emergence occurred after this period, but no further emergence was observed in either species. Significant differences were observed in the number of days for shoot emergence (Figure 4). In Q. faginea, no significant differences were observed at 28 and 35 days after seeding, whereas significant differences were found at days 14, 21, and 42 after seeding, where shoot emergence was lower. For Q. coccifera, shoot emergence was significantly higher at day 35 after seeding, with no significant differences between days 28 and 42. Regarding the shoot emergence between the two species, significant differences were only observed on days 14 and 21 after seeding, where shoot emergence occurred exclusively in Q. faginea, and on day 42, where Q. coccifera exhibited significantly higher shoot emergence compared to Q. faginea.
A Spearman correlation (Figure 5) indicated that the correlation between species and substrate was very weak and not significant (ρ = -0.12; p-value = 0.885), suggesting that species had no relevant relationship with the type of substrate used for acorn germination. However, when analysing the relationship between species and the number of days required for acorn germination, a positive and significant correlation was observed (ρ = 0.352; p-value = 0.001), indicating that species influenced the time needed for shoot emergence. Specifically, Q. faginea required fewer days for germination compared to Q. coccifera. Furthermore, the correlation between species and acorn weight was significant but negative (ρ = -0.306; p-value = 0.001), suggesting an inverse relationship between species and acorn weight. This indicates that the two species exhibited distinct average acorn weights, with Q. faginea producing heavier acorns than Q. coccifera. However, when correlating acorn weight with the number of days to shoot emergence, a negative but non-significant correlation was found (ρ = -0.156; p-value = 0.06). This result suggests that, although there may be a tendency for lighter acorns to take longer to germinate, this relationship is not statistically significant.
Similarly, the correlation between substrate and acorn weight was positive but very weak and not significant (ρ = 0.035; p-value = 0.672). However, when correlating substrate type with the number of days to shoot emergence, a negative and significant correlation was observed (ρ = -0.239; p-value = 0.004). This finding suggests that the type of substrate influenced the shoot emergence time of acorns. In this study, it was observed that acorns of both species germinated earlier in the CF/V substrate compared to the two substrates composed of OPC.
4. Discussion
4.1. Physical Characterization of Acorns
Intrinsic factors such as weight, length, and width of acorns can influence germination success. Among these factors, acorn weight is particularly important, as it determines the reserves available during the initial stages of seedling development [31]. The weight of acorns in Quercus species is highly variable, especially at the intraspecific level, whether individuals belong to the same or different populations [32]. In this study, the average weight and width of acorns of Q. faginea and Q. coccifera showed significant differences, with Q. faginea having a significantly higher weight and width compared to Q. coccifera. Intrapopulation variability among Quercus trees can alter the relationship between seed weight and traits related to seedling survival, as observed by González-Rodríguez et al. [3]. This variability can directly influence germination success and early seedling development, especially considering the differences in seed weight and dimensions observed for Q. faginea and Q. coccifera in this study. In Quercus species, germination success has been shown to increase with acorn weight, with this relationship being stronger in some phylogenetic groups, such as Q. robur, Q. leucotrichophora, Q. semecarpifolia, and other Quercus spp. from the Fagaceae family [33,34,35]. While these studies have reported that heavier acorns tend to exhibit higher germination rates, the results of this study suggest that this relationship is not always consistent. Although a tendency for heavier seeds to germinate faster was observed, the correlation was negative and non-significant, indicating that other factors may also influence germination dynamics. Similar results have also been reported for Q. ilex [36], and Q. faginea [2], where no correlation was found between acorn weight and germination percentage. These findings underscore the complexity of seed germination, where intrinsic traits and environmental conditions (e.g. temperature, light and humidity) [37] interact dynamically to influence germination.
4.2. Emergence and Germination
The number of acorns with radicle emergence was lower in Q. faginea than in Q. coccifera. This difference was not statistically significant, and acorn weight did not appear to influence emergence. Previous studies highlighted that pre-treatment conditions, such as cold stratification, are essential for promoting epicotyl emergence and germination [38]. For instance, in certain Quercus species, such as Q. rubra [39], Q. nigra, and Q. phellos [38], successful seedling emergence depends on the duration of cold stratification and post-stratification temperatures. Some Quercus exhibit epicotyl dormancy, with radicle emergence immediately and shoot emergence delayed to spring or summer, such as Q. vulcanica [40]. Nevertheless, Q. faginea acorns do not exhibit dormancy, and under suitable temperature and moisture conditions, germination begins within 15 days after seeding [14]. In our study, radicle emergence was observed 7 days after seeding in both species, but shoot emergence was observed in fewer individuals than the radicle emergence. These findings suggest that, under favourable conditions, radicle emergence occurs rapidly, potentially contributing to the overall success of seedling survival and development.
Regarding germination, Q. coccifera exhibited significantly higher germination rates than Q. faginea. These differences may be attributed to species-specific physiological traits, ecological adaptations, and seed structure, which determine seed viability and water absorption capacity [41]. Indeed, these factors play a crucial role in germination, with variations in seed coat thickness and chemical composition influencing imbibition rates and germination speed [17]. Germination time in this study showed that Q. coccifera acorns began to germinate on day 28 after seeding, while Q. faginea acorns started germinating 14 days after seeding, regardless of the substrate. Similar patterns have been reported in Q. ilex, which tends to germinate more slowly than Q. suber under the same conditions, likely due to differences in seed dormancy and environmental adaptability [3]. Moreover, the seed coat may contain germination inhibitors, a form of dormancy commonly referred to as chemical dormancy. The seed coat, endosperm, or embryo of many species contains phenols, aldehydes, and other chemicals that inhibit seed germination [41]. This is the case for some Q. ilex species, where proteases and other protease inhibitors in acorns have been identified as germination inhibitors [42]. Furthermore, ecological adaptation to specific environments may help explain the differences in germination rates [43] observed between Q. faginea and Q. coccifera in this study.
4.3. Effect of Substrate on Germination
In this study on Q. faginea and Q. coccifera, substrate properties had a significant impact on germination, regardless of acorn weight and size. This effect influenced both the germination rate and the time to germination. Acorns placed in CF/V germinated more readily and faster than those in OPCs. Germination rates were consistently higher in CF/V to both OPCs.
The use of composted olive pomace as a plant nursery substrate component presents several challenges due to its high organic load, elevated mineral salt content, and the presence of phytotoxic compounds [44]. Despite these potential limitations, little research has been conducted on its effectiveness as a plant substrate for the germination of Quercus species. The exception is a study by Gusmão et al. [45], which assessed Q. suber germination in a commercial substrate and olive compost. The results showed that the commercial substrate achieved a higher germination rate and promoted greater shoot height, root length, and biomass accumulation compared to OPC. In contrast, seedlings grown in OPC were smaller, likely due to the substrate’s physicochemical properties, which may have limited water absorption by the roots [45]. In this study, CF/V achieved the best results in the germination of acorns for both studied species. This may be attributed to its high-water retention capacity, which enhances seed hydration and creates optimal conditions for germination. This property is linked to its porous structure and the presence of lignin and cellulose, which improve its ability to absorb and retain moisture [46]. Furthermore, CF/V provides excellent aeration, ensuring a proper balance between water retention and oxygen availability for the seeds [47]. This may, at least partially, explain the reason why the substrates showed a weak but significant correlation with shoot emergence.
Another factor that may have influenced seed germination and subsequent plant development is the pH of the substrate, which plays a crucial role in seed germination by affecting nutrient availability and seedling health. CF/V typically has a pH range between 5 and 6, which is slightly acidic to neutral. This pH range is generally favorable for most plant species as it allows for optimal nutrient uptake and creates a balanced environment for seedling development [46]. In contrast, OPC has a much higher pH, typically ranging from acidic to values above 9 [48] making it alkaline. This high pH can limit nutrient availability, particularly for micronutrients such as iron and manganese, which become less soluble in alkaline conditions, potentially hindering seedling growth [49]. For species of the Quercus genus, the ideal pH for germination and growth is acidic to neutral [50]. Q. robur thrives in soils with a pH between 5.5 and 7.0, promoting a balanced nutrient uptake [51]. Q. faginea, in general, is considered to be a tree with low soil requirements and can be found in both siliceous (acidic) soils and calcareous (neutral to basic) soils [14]. Therefore, while CF/V provides a more neutral pH conducive to most seeds, OPC, with its higher pH, may require specific adjustments in its properties.
5. Conclusions
In this study, Q. coccifera showed a higher germination rate compared to Q. faginea. Although the acorns of Q. faginea were larger and heavier, seed weight did not appear to significantly influence germination. However, there was a tendency for heavier acorns to germinate in fewer days. Substrates were found to have an impact on both emergence and germination . Commercial substrate, coconut fiber and vermiculite exhibited the highest emergence and germination rates for both species, possibly due to its physicochemical properties, including high water retention capacity linked to its porous structure and the presence of lignin and cellulose, which enhance seed hydration and create optimal conditions for germination. The olive pomace compost, in both sand proportions, yielded poorer results, likely due to its physical and chemical properties that limited moisture retention during acorn germination, suggesting that modifications are needed to make it more suitable for nursery use.
Author Contributions
Conceptualization, R.P., A.D., and A.C.G.; methodology, R.P., A.D., A.R., and A.C.G.; formal analysis, R.P., A.D., and A.C.G.; investigation, R.P., A.D., A.C.G.; resources, R.P., A.D., A.R., and A.C.G.; writing—original draft preparation, R.P., A.D., and A.C.G.; writing—review and editing, R.P., A.D., A.C.G. All authors have read and agreed to the published version of the manuscript.
Acknowledgments
This work is funded by National Funds through FCT - Foundation for Science and Technology of under the Project with MED UIDB/05183 (https://doi.org/10.54499/UIDB/05183/2020) and CHANGE (https://doi.org/10.54499/LA/P/0121/2020). The authors acknowledge Microplan Lda. for their support in the establishment of the experiments at their facilities and project INOVCIRCOLIVE – “Inovação e circularidade no setor oleícola / Inovation and circularity in olive oil sector/” (PRR-C05-i03-I-000208-LA5.1).
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| OPC | Olive Pomace Compost |
| OPC75/S | Olive Pomace Compost and Sand in a ratio of 75/25% |
| OPC50/S | Olive Pomace Compost and Sand in a ratio of 50/50% |
| CF/V | Coconut Fiber and Vermiculite |
References
- Moreno, G.; S, A. ; S., B.; J, C.-D. Agroforestry systems of high nature and cultural value in Europe: provision of commercial goods and other ecosystem services. Agroforest Syst. [CrossRef]
- Alonso-Crespo, I.M.; Silla, F.; Jiménez Del Nogal, P.; Fernández, M.J.; Martínez-Ruiz, C.; Fernández-Santos, B. Effect of the Mother Tree Age and Acorn Weight in the Regenerative Characteristics of Quercus Faginea. Eur. J. Forest Res. 2020, 139, 513–523. [Google Scholar] [CrossRef]
- González-Rodríguez, V.; Villar, R.; Navarro-Cerrillo, R.M. Maternal Influences on Seed Mass Effect and Initial Seedling Growth in Four Quercus Species. Acta Oecologica 2011, 37, 1–9. [Google Scholar] [CrossRef]
- Vieira, J.; Campelo, F.; Nabais, C. Environment Controls Seasonal and Daily Cycles of Stem Diameter Variations in Portuguese Oak (Quercus Faginea Lambert). Forests 2022, 13, 170. [Google Scholar] [CrossRef]
- Maltez-Mouro, S.; García, L.V.; Freitas, H. Influence of Forest Structure and Environmental Variables on Recruit Survival and Performance of Two Mediterranean Tree Species (Quercus Faginea L. and Q. Suber Lam.). Eur.J. Forest Res. 2009, 128, 27–36. [Google Scholar] [CrossRef]
- Ozturk, M.; Altay, V. Role of Quercus Coccifera (= Q. Calliprinos) in the Light of Climate Change Scenarios in the Mediterranean Basin. Plant Fungal Res. [CrossRef]
- Stefi, A.L.; Nikou, T.; Papadopoulou, S.; Kalaboka, M.; Vassilacopoulou, D.; Halabalaki, M.; Christodoulakis, N.S. The Response of the Laboratory Cultivated Quercus Coccifera Plants to an Artificial Water Stress. Plant Stress 2022, 4. [Google Scholar] [CrossRef]
- Vilagrosa, A.; Cortina, J.; Gil-Pelegrín, E.; Bellot, J. Suitability of Drought-Preconditioning Techniques in Mediterranean Climate. Restor. Ecol. 2003, 2, 208–2016. [Google Scholar] [CrossRef]
- García, C.; Espelta, J.M.; Hampe, A. Managing Forest Regeneration and Expansion at a Time of Unprecedented Global Change. J. Appl. Ecol. 2020, 57, 2310–2315. [Google Scholar] [CrossRef]
- Gonçalves, A.C.; Fonseca, T.F. Influence Management and Disturbances on the Regeneration of Forest Stands. Front. For. Glo. Change 6. [CrossRef]
- Lázaro-González, A.; Andivia, E.; Hampe, A. Revegetation through Seeding or Planting: A Worldwide Systematic Map. Environ. Manage. 2023, 337. [Google Scholar] [CrossRef]
- Grossnickle, S.C. Why Seedlings Survive: Influence of Plant Attributes. New Forests 2012, 43, 711–738. [Google Scholar] [CrossRef]
- Mariotti, B.; Maltoni, A.; Jacobs, D.F.; Tani, A. Container Effects on Growth and Biomass Allocation in Quercus Robur and Juglans Regia Seedlings. Scand. J. For. Res. 2015, 1–15. [Google Scholar] [CrossRef]
- Oliveira, Â.; Fabião, A. ; Gonçalves, Ana Cristina; Correia, Alexandre Vaz O Carvalho-Cerquinho em Portugal, Ed.; ISA Press: Lisboa, 2001. [Google Scholar]
- Warsaw, A.L.; Fernandez, R.T.; Cregg, B.M.; Andresen, J.A. Container-Grown Ornamental Plant Growth and Water Runoff Nutrient Content and Volume Under Four Irrigation Treatments. Horts 2009, 44, 1573–1580. [Google Scholar] [CrossRef]
- Li, S.; Pezeshki, S.R.; Goodwin, S. Effects of Soil Moisture Regimes on Photosynthesis and Growth in Cattail (Typha Latifolia). Acta Oecologica 2004, 25, 17–22. [Google Scholar] [CrossRef]
- Upretee, P.; Bandara, M.S.; Tanino, K.K. The Role of Seed Characteristics on Water Uptake Preceding Germination. Seeds 2024, 3, 559–574. [Google Scholar] [CrossRef]
- Reho, M.; Vilček, J.; Torma, S.; Koco, Š.; Lisnyak, A. Influence of Soil Substrate on Success of Growing of of English Oak (Quercus Robur, L.) Seedlings: Key Study in Conditions of Forest-Steppe Ukraine. In Prime Archives in Environmental Research; Vide Leaf, Hyderabad, 2023. ISBN 978-93-92117-15-2.
- Cann, J.; Tang, E.; Thomas, S.C. Biochar and Deactivated Yeast as Seed Coatings for Restoration: Performance on Alternative Substrates. Seeds 2024, 3, 544–558. [Google Scholar] [CrossRef]
- Clausing, R.; Falace, A.; De la Fuente, G.; Della Torre, C. Ex-Situ Restoration of the Mediterranean Forest-Forming Macroalga Ericaria Amentacea: Optimizing Growth in Culture May Not Be the Key to Growth in the Field. Mar. Environ. Res. 2024, 202. [Google Scholar] [CrossRef]
- Kholkhal, D.; Benmahioul, B. Effects of Substrate on the Germination and Seedling Growth of Quercus Suber L. Biodiv. Res. Conserv. 2021, 64, 7–14. [Google Scholar] [CrossRef]
- Alvino, F. de O.; Rayol, B.P. Different substrata effects in the germination of Ochroma Pyramidale (Cav. ex Lam.) urb. (Bombacaceae). Ciência Florestal 17. [CrossRef]
- Piva, A.L.; Mezzalira, E.J.; Santin, A.; Schwantes, D.; Klein, J.; Villa, F.; Tsutsumi, C.Y.; Nava, G.A. Mergence and Initial Development of Cape Gooseberry (Physalis Peruviana) Seedlings with Different Substrates Compositions. Afr. J. Agric. Res. 2013, 6579–6584. [Google Scholar] [CrossRef]
- Ayadi, M.; Awad, S.; Villot, A.; Abderrabba, M.; Tazerout, M. Heterogeneous Acid Catalyst Preparation from Olive Pomace and Its Use for Olive Pomace Oil Esterification. Renew. Energy. 2021, 165, 1–13. [Google Scholar] [CrossRef]
- Ribeiro, T.B.; Oliveira, A.L.; Costa, C.; Nunes, J.; Vicente, A.A.; Pintado, M. Total and Sustainable Valorisation of Olive Pomace Using a Fractionation Approach. Applied Sci. 2020, 10, 6785. [Google Scholar] [CrossRef]
- Dermeche, S.; Nadour, M.; Larroche, C.; Moulti-Mati, F.; Michaud, P. Olive Mill Wastes: Biochemical Characterizations and Valorization Strategies. Process Biochem. 2013, 48, 1532–1552. [Google Scholar] [CrossRef]
- Buono, D.D.; Said-Pullicino, D.; Proietti, P.; Nasini, L.; Gigliotti, G. Utilization of Olive Husks as Plant Growing Substrates: Phytotoxicity and Plant Biochemical Responses. Compost Sci. Util. 2011, 19, 52–60. [Google Scholar] [CrossRef]
- Ameziane, H.; Nounah, A.; Khamar, M.; Zouahri, A. Composting Olive Pomace: Evolution of Organic Matter and Compost Quality. Agron. Res. [CrossRef]
- Sempiterno, C.; Fernandes, R.; Dias, A.; Fitas, V. Valorização Agronómica de Bagaço Húmido Compostado e não Compostado – Impacto no Desenvolvimento de Lolium perenne L. In Proceedings of the X National Symposium on Olive Growing, Bragança, Portugal, 2024., 23–25 October.
- Afonso, A.; Nunes, C. Aplicações e Soluções Em SPSS. Universidade de Évora, 2019. [Google Scholar]
- Tilki, F. Influence of Acorn Size and Storage Duration on Moisture Content, Germination and Survival of Quercus Petraea (Mattuschka). New For. 2010, 18, 5–17. [Google Scholar]
- Gómez, J.M. Bigger is not always better: conflicting selective pressures on seed size in Quercus ilex. Evolution 2004, 58, 71–80. [Google Scholar] [CrossRef]
- Purohit, V.K.; Tamta, S.; Nandi, S.K.; Rikhari, H.C.; Palni, L.M.S. Does acorn weight influence germination and subsequent seedling growth of central himalayan oaks? J. Trop. For. Sci. 2003, 15, 483–492. [Google Scholar]
- Sánchez-Montes De Oca, E.J.; Badano, E.I.; Silva-Alvarado, L.E.; Flores, J.; Barragán-Torres, F.; Flores-Cano, J.A. Acorn Weight as Determinant of Germination in Red and White Oaks: Evidences from a Common-Garden Greenhouse Experiment. Ann. For. Sci. 2018, 75, 12. [Google Scholar] [CrossRef]
- Bogdan, S.; Ivanković, M.; Vujnović, Z.; Lanšćak, M.; Gradečki Poštenjak, M.; Bogunović, S.; Gavranović Markić, A. Acorn Yields and Seed Viability of Pedunculate Oak in a 10-Year Period in Forest Seed Objects across Croatia. SEEFOR 2022, 13, 2201. [Google Scholar] [CrossRef]
- Fernández-Santos, B.; Moro, D.; Martínez-Ruiz, C.; Fernández, M.J. Efectos del peso de la bellota y de la edad del árbol productor en las características regenerativas de Quercus ilex. Avances en la restauración de sistemas forestales. Técnicas de implantación. University of Salamanca, 2013, 197-202.
- García-De La Cruz, Y.; López-Barrera, F.; Ramos-Prado, J.M. Germinación y Emergencia de Plántulas de Cuatro Especies de Encino Amenazadas. MYB 2016, 22, 77–87. [Google Scholar] [CrossRef]
- Hawkins, T.S. Dormancy Break and Germination Requirements in Acorns of Two Bottomland Quercus Species (Sect. Lobatae ) of the Eastern United States with References to Ecology and Phylogeny. Seed Sci. Res. 2020, 30, 199–205. [Google Scholar] [CrossRef]
- Jastrzębowski, S.; Ukalska, J.; Walck, J.L. Does the Lag Time between Radicle and Epicotyl Emergences in Acorns of Pedunculate Oak (Quercus Robur L.) Depend on the Duration of Cold Stratification and Poststratification Temperatures? Modelling with the Sigmoidal Growth Curves Approach. Seed Sci Res. 2021, 1–11. [Google Scholar]
- Tilki, F.; Alptekin, C.U. Germination and Seedling Growth of Quercus Vulcanica: Effects of Stratification, Desiccation, Radicle Pruning, and Season of Sowing. New For. 2006, 32, 243–251. [Google Scholar] [CrossRef]
- Wang, N.; Zhang, Z.; Xu, W.; Zhou, H.; Ning, R. Research Progress on Dormancy Mechanism and Germination Technology of Kobresia Seeds. Plants 2022, 11, 3192. [Google Scholar] [CrossRef]
- Escandón, M.; Bigatton, E.D.; Guerrero-Sánchez, V.M.; Hernández-Lao, T.; Rey, M.-D.; Jorrín-Novo, J.V.; Castillejo, M.A. Identification of Proteases and Protease Inhibitors in Seeds of the Recalcitrant Forest Tree Species Quercus Ilex. Front. Plant Sci. 2022, 13, 907042. [Google Scholar] [CrossRef]
- Amimi, N.; Dussert, S.; Vaissayre, V.; Ghouil, H.; Doulbeau, S.; Costantini, C.; Ammari, Y.; Joët, T. Variation in Seed Traits among Mediterranean Oaks in Tunisia and Their Ecological Significance. Ann. Bot. 2020, 125, 891–904. [Google Scholar] [CrossRef] [PubMed]
- Canet, R.; Pomares, F.; Cabot, B.; Chaves, C.; Ferrer, E.; Ribó, M.; Albiach, M.R. Composting Olive Mill Pomace and Other Residues from Rural Southeastern Spain. Waste Manag. 2008, 28, 2585–2592. [Google Scholar] [CrossRef]
- Gusmão, A.G.; Toloto, M.; Segatelli, A.B.; Fonseca, F.; Cortez, P.; Figueiredo, T.D.; Hernández, Z. Efeito de diferentes substratos na taxa de crescimento de Quercus suber L. Revista de Ciências Agrárias. [CrossRef]
- Arenas, M.; Vavrina, C.S.; Cornell, J.A.; Hanlon, E.A.; Hochmuth, G.J. Coir as an Alternative to Peat in Media for Tomato Transplant Production. HortSci. 2002, 37, 309–312. [Google Scholar] [CrossRef]
- Prasad, M. Physical, Chemical and Biological Properties of Coir Dust. Acta Hortic. 1997, 21–27. [Google Scholar] [CrossRef]
- P. Rueda, M.; Domínguez-Vidal, A.; Aranda, V.; Ayora-Cañada, M.J. Monitoring the Composting Process of Olive Oil Industry Waste: Benchtop FT-NIR vs. Miniaturized NIR Spectrometer. Agronomy 2024, 14, 3061. [Google Scholar] [CrossRef]
- Ferrarezi, R.S.; Lin, X.; Gonzalez Neira, A.C.; Tabay Zambon, F.; Hu, H.; Wang, X.; Huang, J.-H.; Fan, G. Substrate pH Influences the Nutrient Absorption and Rhizosphere Microbiome of Huanglongbing-Affected Grapefruit Plants. Front. Plant Sci. 2022, 13, 856937. [Google Scholar] [CrossRef]
- Kühne, C.; Bartsch, N. Germination of Acorns and Development of Oak Seedlings (Quercus Robur L.) Following Flooding. J. For. Sci. 2007, 53, 391–399. [Google Scholar] [CrossRef]
- Oneț, A.; Teușdea, A.; Boja, N.; Domuța, C.; Oneț, C. Effects of Common Oak (Quercus Robur L.) Defolition on the Soil Properties of an Oak Forest in Western Plain of Romania. Ann. For. Res. 2016, 59, 1. [Google Scholar] [CrossRef]
Figure 1.
Example of acorn sizes (width and length) of Q. faginea and Q. coccifera.
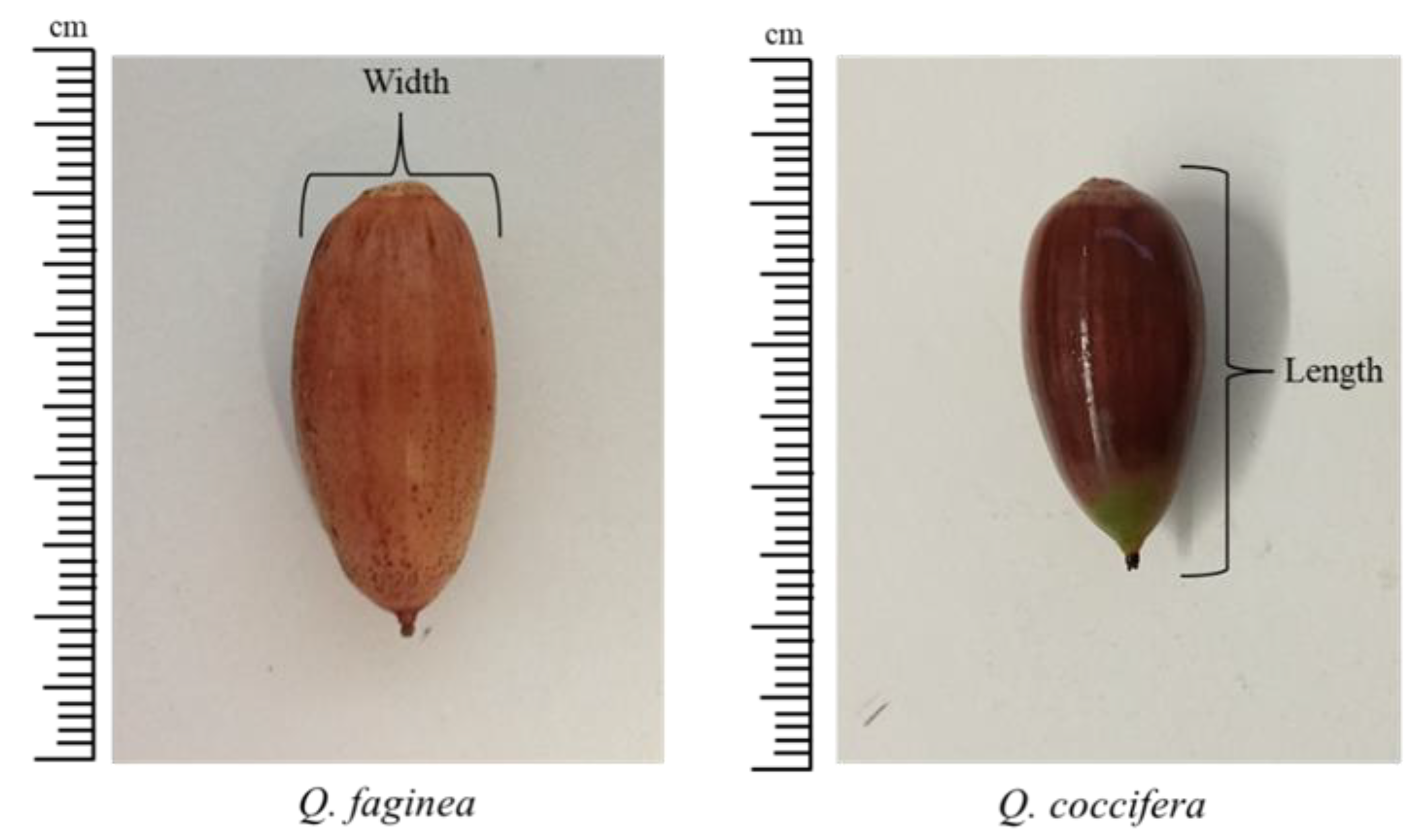
Figure 2.
Percentage of emergence (top) and germination (bottom) for Q. faginea and Q. coccifera across all substrates (mean ± standard error). Different lowercase letters indicate statistically significant differences among the three substrates, and uppercase letters denote differences between species (p ≤ 0.05).
Figure 2.
Percentage of emergence (top) and germination (bottom) for Q. faginea and Q. coccifera across all substrates (mean ± standard error). Different lowercase letters indicate statistically significant differences among the three substrates, and uppercase letters denote differences between species (p ≤ 0.05).
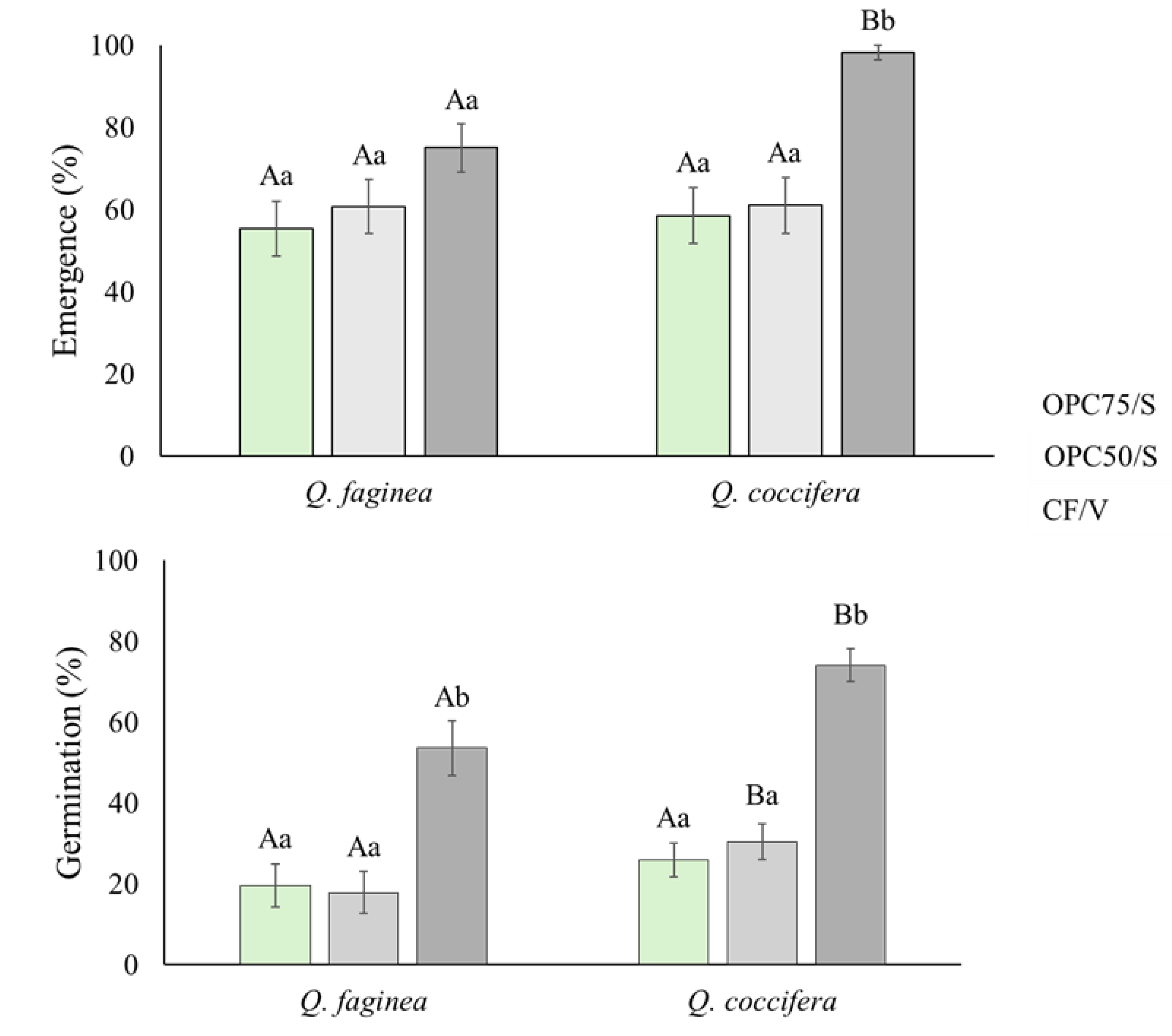
Figure 3.
Illustration of the aerial growth of Q. faginea and Q. coccifera seedlings during the experimental period.
Figure 3.
Illustration of the aerial growth of Q. faginea and Q. coccifera seedlings during the experimental period.

Figure 4.
Shoot emergence (in days) and germination (in percentage) of the acorns of Q. faginea and Q. coccifera.
Figure 4.
Shoot emergence (in days) and germination (in percentage) of the acorns of Q. faginea and Q. coccifera.
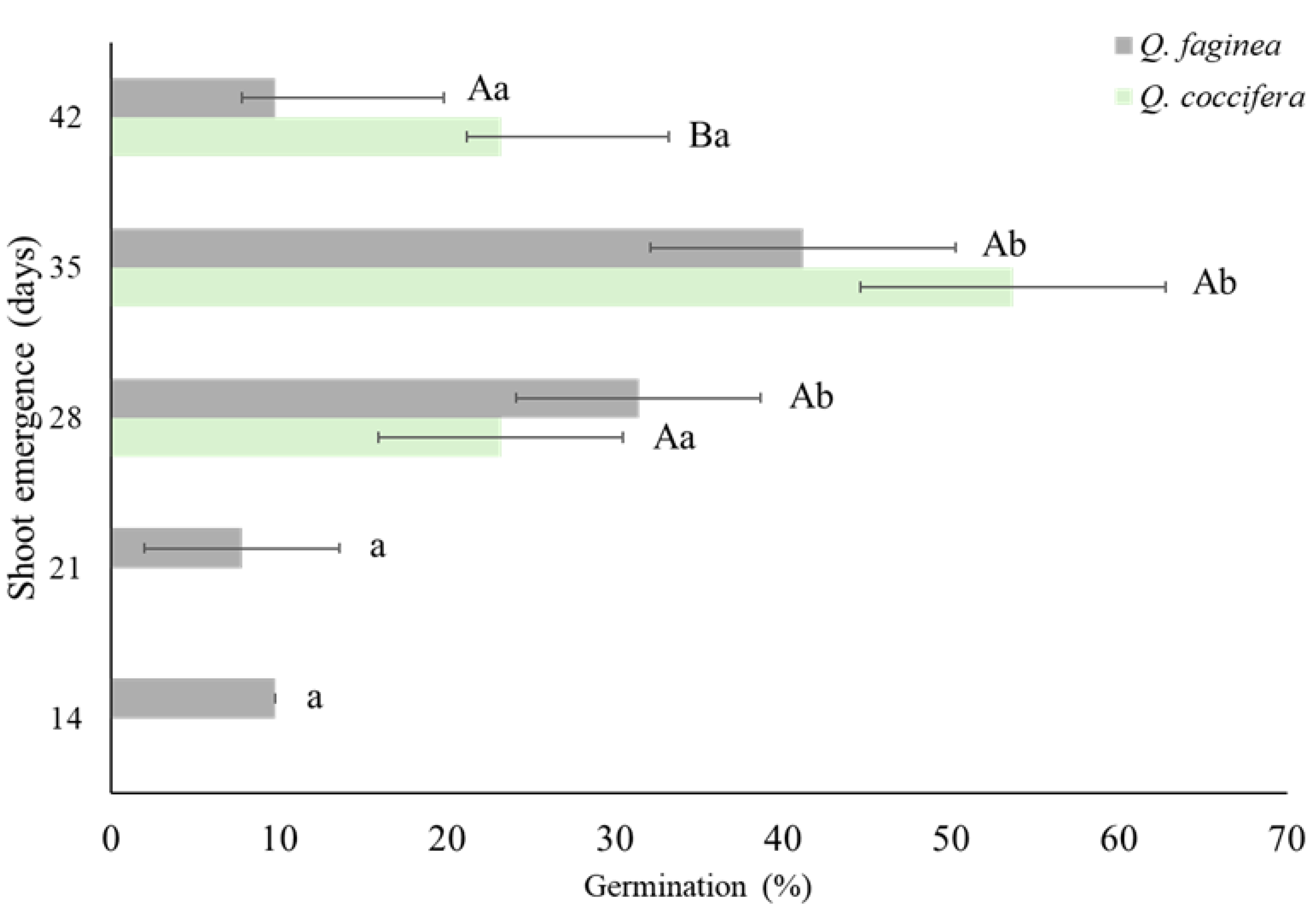
Figure 5.
Heat map of Spearman correlation coefficients.
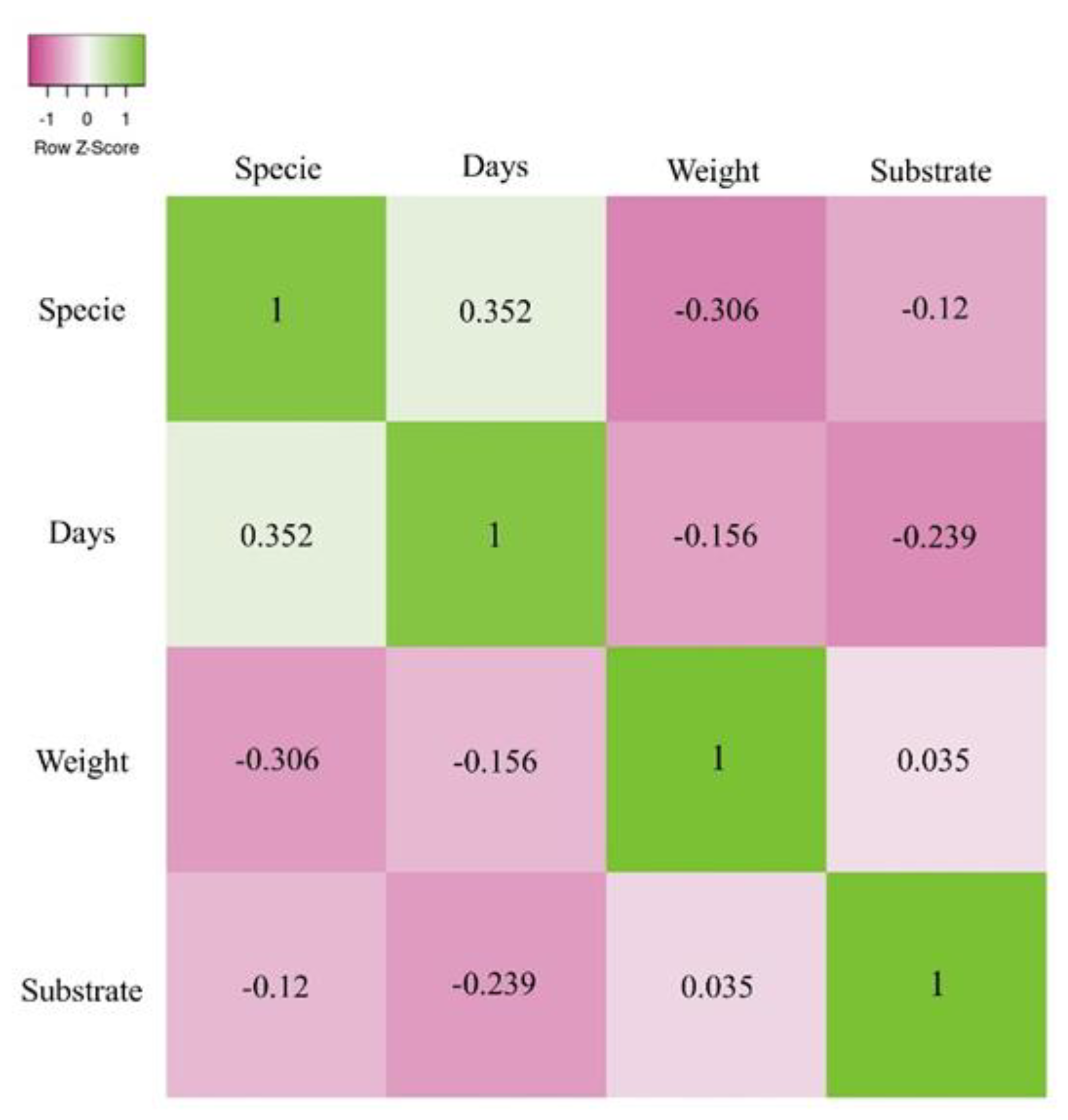

Table 1.
Physical characteristics (weight, length, and width) of Q. faginea and Q. coccifera. The data represent the average measurements with the associated standard error for each parameter.
Table 1.
Physical characteristics (weight, length, and width) of Q. faginea and Q. coccifera. The data represent the average measurements with the associated standard error for each parameter.
| Parameter | N | Q. faginea | Q. coccifera |
|---|---|---|---|
| Weight | 168 | 5.18 ± 0.08 b | 4.28 ± 0.07 a |
| Lenght | 168 | 32.25 ± 0.18 b | 31.74 ± 0.15 b |
| Width | 168 | 16.06 ± 0.09 b | 14.09 ± 0.09 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



