
Submitted:
26 March 2025
Posted:
27 March 2025
You are already at the latest version
Abstract
Background: The intrauterine and early extrauterine development represents a “window of opportunity” in the immunological development. The underlying mechanisms are still poorly understood. The aim of this study was to provide reference values on B cell subpopulations in cord blood of term newborns, and peripheral venous blood of juveniles and adults to analyze the potential spectrum of their physiological age-related variation. Methods: Flow cytometry was used to evaluate human B lymphocytes and subpopulations in cord blood (n = 10), and peripheral blood from healthy children and adolescence, further called juveniles, aged 1 to 17 years (n=20) and adults aged 24 to 62 years (n = 10). Results: Our findings showed increasing frequencies of IgM memory B cells, class-switched memory B cells, marginal zone B cells and plasmablasts, from cord blood to peripheral blood of juveniles and adults. In contrast, the percentage of naïve B cells was higher in newborns than in juveniles and adults. The frequencies of immature B cells were similar in all three groups and B1 cell were similar in cord blood and peripheral blood of adults. Interestingly, transitional B cells frequencies were similar in cord blood and adults but significantly lower in juveniles. Conclusions: The frequencies of circulating B cell subpopulation are subject to considerable changes during ontogeny, reflecting overlying effects of maturation and of the acquisition of an adaptive immune memory.
Keywords:
B cells
; B1 cells
; human
; leukocytes
; cord blood
; development
; ontogeny
1. Introduction
B cells are integral to the adaptive immune response, particularly in defending against extracellular pathogens [1]. They are responsible for producing antibodies, which neutralize and eliminate invading microbes, and for forming long-lasting memory responses that provide protection upon re-exposure to the same pathogen. The development of the human immune system begins in the fetal stage but is not fully complete at birth [2], making newborns particularly vulnerable to infections. Investigating the B cell compartment in newborns contributes to a better understanding of neonatal immunity, future immune function, and helps developing targeted interventions to enhance early life immune protection.
Newborns possess an adequate number of immune cells, including B cells, but these cells require activation and further maturation through exposure to environmental antigens. This process of immune system development continues throughout childhood and adolescence, with individuals gradually acquiring the immunological capabilities of adults by puberty [3]. However, the immune system remains dynamic and undergoes further refinement and adaptation throughout life, influenced by ongoing encounters with pathogens and other environmental stimuli.
In the first few weeks following birth, B lymphocytes proliferate extensively, likely driven by exposure to a diverse array of environmental antigens. This antigenic stimulation triggers the proliferation and differentiation of antigen-specific B cells, leading to the generation of memory B cells and long-lived plasma cells [4,5]. The maturation of B lymphocytes is a complex, highly regulated, multi-phase process that begins in the bone marrow. Immature B cells, expressing IgM on their surface, exit the bone marrow and migrate to secondary lymphoid structures, such as the follicles and germinal centers of the spleen and lymph nodes [6,7]. Within these specialized microenvironments, B cells undergo further maturation, including class switching, somatic hypermutation, and affinity maturation, ultimately generating memory B cells and plasma cells capable of producing high-affinity antibodies. Memory B cells circulate continuously between the blood and lymphatic organs, providing long-term immunological memory and rapid responses upon re-exposure to the same antigen. Plasma cells, on the other hand, primarily reside in the bone marrow and secrete large quantities of antibodies to provide immediate protection against infection.
In addition to conventional B cells, a unique subset known as B1 cells plays a distinct role in immune defense. B1 cells are characterized by their ability to spontaneously secrete "natural" antibodies, primarily IgM which provides broad protection against a variety of pathogens, even in the absence of prior antigen exposure [8]. These natural antibodies contribute to innate immunity and play a key role in early defense against infection, particularly in newborns who have limited pre-existing immunity. While B1 cells are well-characterized in murine models [8,9], their identification and characterization in humans have been more challenging. The quantity and even the existence of B1 cells in humans remain a subject of debate, with multiple definitions proposed based on different sets of surface markers and functional characteristics. In mice, B1 cells are typically defined as CD5+ and/or CD11b+; however, many B1-like cells in humans do not express CD5 [10,11]. Griffin et al. [12] identified a subpopulation of B cells in both human adult peripheral blood and cord blood, characterized by the markers CD20+, CD27+, and CD43+, which exhibit functional features typical of B1 cells, such as spontaneous IgM secretion, constitutive BCR signaling, and the ability to regulate allogeneic T cell proliferation.
Unlike the mature adult immune system, the neonatal immune system is still developing, shaped by in utero experiences and poised to encounter a world teeming with new antigens. Among the key players in this developmental process B cells are the antibody-producing arm of the adaptive immune system. Neonatal cord blood, a readily accessible source of fetal blood, provides a unique insight into the composition and functionality of early B-cell populations, offering a detailed examination of the B-cell compartment in newborns. Notably, cord blood contains a greater number of B cells than adult venous blood, reflecting the active B cell lymphopoiesis occurring during fetal development. At birth, CD27+ IgM+ IgD+ memory B cells represent the dominant B cell population, but their frequency decreases during childhood and stabilizes in young adults [13,14]. In contrast, CD27+ class-switched memory B cells, which have undergone isotype switching to produce IgG, IgA, or IgE antibodies, gradually increase in frequency within peripheral blood with age, reflecting the accumulation of immunological memory following antigen encounters.
Moreover, neonatal cord blood B cells show a distinct transcriptional program and an accelerated but likely transient antibody response, with a conserved BCR repertoire and efficient IgA class switching, reflecting their unique develop-mental state [15]. Transitional B cellsare crucial for establishing self-tolerance and shaping the mature repertoire [16]. Marginal zone B cells that connect and direct innate immune reaction and are recently known to be involved in autoimmune processesare likely to contribute to early neonatal infection defense. Finally, B1-B cells, producers of natural antibodies, provide immediate, innate-like humoral protection in newborns [17,18,19]. Despite these findings, our under-standing of B cell subpopulation composition in newborns and its relationship to immune function remains incomplete. Therefore, this study aims to provide a deeper understanding of B cell subpopulation composition across different age groups, utilizing updated cell definitions and a comprehensive flow cytometry approach. We focus on cord blood as a crucial link between intrauterine and extrauterine development, providing valuable insights into the early establishment of the B cell repertoire and its potential implications for immune health in newborns and infants and children.
A deeper comprehension in the development and function of B cell subpopulations in cord blood has significant clinical implications. Variations in B cell composition and function in newborns may influence their susceptibility to infections, allergies, and autoimmune diseases [20]. Furthermore, insights gained from studying cord blood B cells could inform the development of novel immunotherapeutic strategies for neonates and infants. Understanding the mechanisms that regulate B-cell tolerance in early life could lead to new approaches for preventing or treating autoimmune diseases.
This study aims to outline the frequency and composition B cell subpopulations found in cord blood of newborns and venous blood of children and adolescence, further called juveniles throughout the paper, as well as adults, highlighting unique properties and potential implications for different age groups for immune development and susceptibility to disease.
2. Results
2.1. Phenotypic Characterization of Memory B Cell Subsets in Healthy Newborns, Juveniles and Adults
This study investigated the age-related differences in the distribution of B lymphocyte subpopulations across three distinct age groups: neonates, children above the age of 1 year and adults. To this end, we analyzed cord blood samples from 10 term neonates and peripheral venous blood samples from 20 juveniles and 10 adults.
Flow cytometry was used to quantify B cell subpopulations. To assess the diversity of the human B cell compartment, we defined 8 distinct B cell subpopulations based on their surface marker expression (Supplementary Table S2). The specific markers used for each subpopulation are listed in Supplementary Table S1, and the gating strategies employed for flow cytometric analysis are depicted in Figure 1.
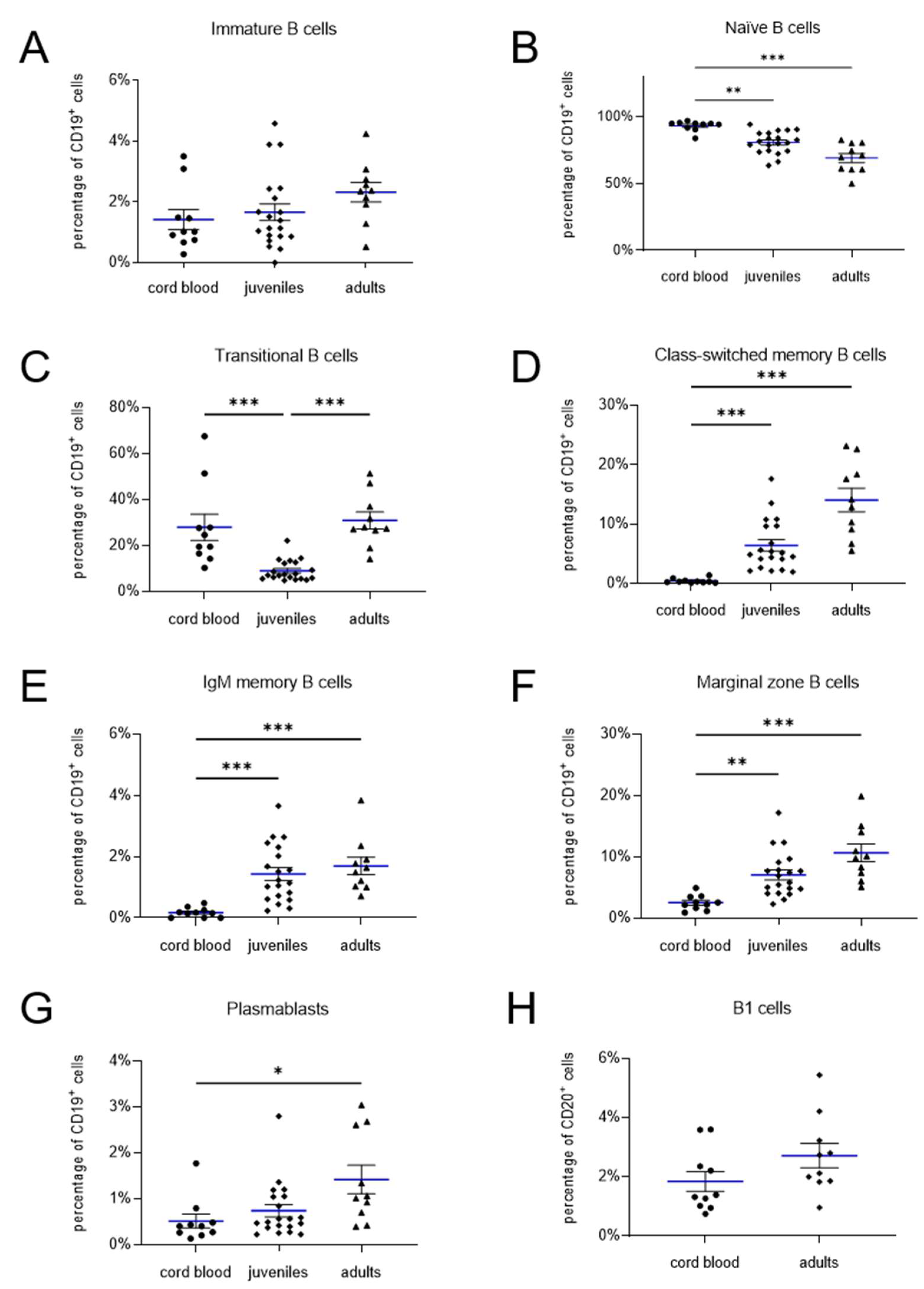
Immature B cells, defined as CD19+ CD27- IgM- IgD- (Figure 1C), were present at similar frequencies across the three age groups: neonates (1.4 ± 0.3%), juveniles (1.7 ± 0.3%), and adults (2.3 ± 0.3%) (neonates vs. juveniles: p > 0.99; neonates vs. adults: p = 0.15; juveniles vs. adults: p = 0.27) (Figure 3A). In contrast, naïve B cells (CD19+ CD27- IgD+ IgM+) were significantly more abundant in cord blood of neonates (93.2 ± 1.2%) compared to peripheral blood of juveniles (80.8 ± 1.8%; p = 0.002) and adults (69.1 ± 3.4%; p < 0.0001) (Figure 3B), indicating an age-related decline in the frequency of this subpopulation.
Transitional B cells, identified as CD19+ CD27+ CD38+ IgM+ lymphocytes (Figure 1D) [21], exhibited similar frequencies in neonates (27.9 ± 5.7%) and adults (30.9 ± 3.7%; p = 0.3527) (Figure 3C). However, a significantly lower frequency was observed in the group of juveniles (9.0 ± 1.0%; neonates vs. juveniles: p < 0.0001; children & adolescence vs. adults: p < 0.0001).
Memory B cells can be further categorized into three subsets: class-switched memory B cells (CD19+ CD27+ IgM- IgD-), IgM memory B cells (CD19+ CD27+ IgM+ IgD-), and marginal zone B cells (CD19+ CD27+ IgM+ IgD+) (Figure 1E). All three memory B cell subsets were significantly more prevalent in the peripheral blood of juveniles and adults compared to cord blood of neonates (Figure 3D–F). Specifically, class-switched memory B cells, IgM memory B cells, and marginal zone B cells were found at frequencies of 6.4 ± 1.0%, 1.4 ± 0.2%, and 7.1 ± 0.8% in juveniles, and 14.1 ± 2.0%, 1.7 ± 0.3%, and 10.7 ± 1.4% in adults, respectively, compared to 2.5 ± 0.4%, 0.2 ± 0.1%, and 0.5 ± 0.1% in neonates (each p < 0.0001). These results indicate an age-dependent increase in the prevalence of all three memory B cell subsets.
2.2. B1 Cells Analyzed in Cord Blood and Adult Samples
B1 cells, also referred to as pre-naïve B cells, represent an intermediate stage in B cell development between transitional and naïve B cells. In humans, B1 cells are characterized by the expression of CD20, CD27, and CD43, along with the absence of CD69 [12]. Their frequency is typically reported as a percentage of the total CD20+ B cell population (Figure 2).
In our analysis, the frequency of CD20+ CD27+ CD43+ CD69- B1 cells did not significantly differ between neonates (1.9 ± 0.3%) and adults (2.7 ± 0.4%; p = 0.1431) (Figure 3H).
Figure 2.
Gating strategy and B1 cell frequency. (A) SSC vs FSC density plot. (B) Lymphocytes were gated to CD43 vs CD69 plot, (C) histogram of CD43+ CD69- cells vs CD27, (D) histogram of CD43- CD69- CD27+ vs CD 20 to identify B1 cells (CD20+ CD27+ CD43+ CD69-).
Figure 2.
Gating strategy and B1 cell frequency. (A) SSC vs FSC density plot. (B) Lymphocytes were gated to CD43 vs CD69 plot, (C) histogram of CD43+ CD69- cells vs CD27, (D) histogram of CD43- CD69- CD27+ vs CD 20 to identify B1 cells (CD20+ CD27+ CD43+ CD69-).
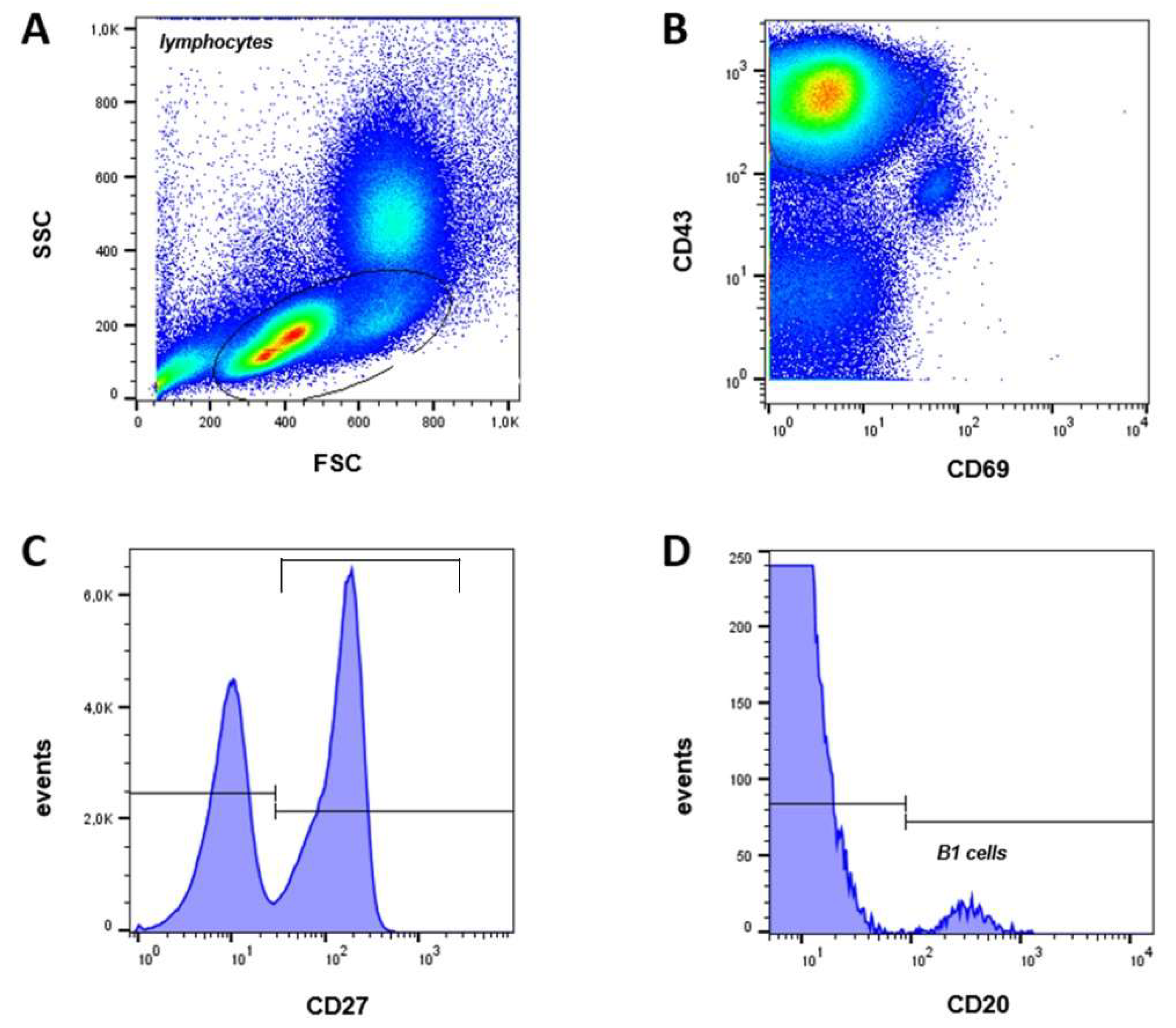
Figure 3.
Quantification of lymphocyte subpopulations in cord blood, juveniles, and adults. Mean values are indicated by blue lines, and SEM is shown as black bars. The frequency of immature and transitional B cells (A, B) as well as naive B cells (C) is higher in neonatal cord blood. In contrast, the percentage of class-switched and IgM memory B cells (D, E), marginal zone B cells (F), and plasmablasts (G) increases with age. The frequency of B1 cells (H) does not differ between neonatal cord blood and adult peripheral blood. * = p < 0.05 (statistically significant); ** = p < 0.01 (highly significant); *** = p < 0.001 (very highly significant).
Figure 3.
Quantification of lymphocyte subpopulations in cord blood, juveniles, and adults. Mean values are indicated by blue lines, and SEM is shown as black bars. The frequency of immature and transitional B cells (A, B) as well as naive B cells (C) is higher in neonatal cord blood. In contrast, the percentage of class-switched and IgM memory B cells (D, E), marginal zone B cells (F), and plasmablasts (G) increases with age. The frequency of B1 cells (H) does not differ between neonatal cord blood and adult peripheral blood. * = p < 0.05 (statistically significant); ** = p < 0.01 (highly significant); *** = p < 0.001 (very highly significant).
3. Discussion
We provide reference values for 8 B lymphocyte subpopulations, including B1 cells, in cord blood of healthy mature newborns. The human B1 cell population, which appears to be subject to changes during ontogenesis, has been re-defined by flow cytometric and functional analyses [22,23,24,25,26]. Our results demonstrate significant differences in the relative counts of B cell subpopulations in cord blood compared to peripheral blood of adolescence and adults. Age-dependent accumulation of immune experience from foreign antigen exposure leads to a gradual shift from naïve to mature B cells. Concurrently, the total number of B lymphocytes gradually decreases with age, while their ability to respond to foreign antigens increases [14].
B cell subpopulations are defined by characteristic expression patterns of surface antigens, cytokines, and master transcription factors. We observed that the proportion of immature B cells (CD19+ CD27- IgM- IgD-) is similar in newborns, juveniles, and adults. These immature cells migrate to the spleen, lymph nodes, or other lymphoid tissues, where they mature into long-lived, adult naïve B cells [27,28]. Naïve B cells express IgD but lack CD27, which is induced by antigen-receptor activation. In umbilical cord blood, naïve B cells (CD19+ CD27- IgM+ IgD+) constitute 93% of the total CD19+ cell population, a 25% higher frequency than in adult peripheral blood. The latter indicates a decrease in naïve B cells with age, despite our observed relative counts being higher than previously reported [14]. Piątosa et al. [14] showed that naïve CD19+ CD27- IgD+ cells increase during the first year of life and gradually decrease thereafter, reaching a plateau in children aged 5–10 years. For our flow cytometric approach, we used CD19, CD38, and IgM expression to delineate B cells. Transitional B cells (CD19+ CD27- IgM+ CD38+), an intermediate stage between immature bone marrow B cells and mature peripheral B cells [23,24,25,27,29], were found at similar frequencies in neonates and adults, contradicting previous reports of a decline with age [14,15]. Our data indicate a decrease in CD19+ CD27- IgM+ CD38+ cells from cord blood to infancy, followed by an increase in adulthood.
Mature naïve B cells differentiate into memory B cells and plasma cells [30]. CD19+ CD27+ IgM+ IgD- memory B cells were virtually absent in cord blood, likely due to limited extrauterine antigen exposure compared to adults, where this population comprises ~15% of all CD19+ B lymphocytes [10,14,31]. CD19+ CD27+ IgM+ IgG+ marginal zone B lymphocytes, scarce in cord blood, accumulate in peripheral blood after the marginal zone develops within the first two years of life [32]. The proportion of CD27+ CD38++ plasmablasts, which did not significantly differ between cord blood, juvenile, and adult blood, remained below 5% of total B cells [13].
B1 cells, distinct from conventional B cell subpopulations, are considered part of the innate immune system due to their constitutive and spontaneous secretion of "natural" IgM antibodies [8,30]. B1 cells differ from conventional B cells in their surface phenotype, function, localization, and ontogeny [33,34]. The identity and even the existence of human B1 cells have been debated due to the lack of definitive surface markers. While initially described in mice as CD5+ B cells (B1a), human B1 cells were initially defined as CD19+ CD5+ [8,35,36]. However, CD5 is expressed on other human B cell subpopulations [24]. Following Griffin et al. [12], who redefined human B1 cells based on morphological and functional studies, we defined B1 cells as CD20+ CD27+ CD43+ CD69-. CD20, unlike CD19, is lost during mature B cell differentiation [37], ensuring the exclusion of immunoglobulin-secreting plasmablasts and plasma cells. CD43 is constitutively expressed on B1 cells, while activated naïve and memory B cells, which also express CD43, co-express CD69 and CD70, which B1 cells do not [12]. Previous studies have shown a decrease in human B1 cell frequency with age [12,38,39], while murine B1 cell frequency increases [40]. We found a slight, non-significant increase in B1 cells from neonatal cord blood compared to adults, with both groups exhibiting ~1–3% of CD20+ B cells. Griffin et al. [41] described a wide range in circulating B1 cell frequency (<1% to >9%) and an age-dependent trend.
Recent studies have shown that CD20+ CD27+ CD43+ CD70- B1 cells are closely related to plasmablasts and pre-plasmablasts based on gene expression profiles and functional characteristics [42,43]. These cells were later found to include CD20+CD38hi precursors for plasmablasts and pre-plasmablasts [44]. Consequently, CD20+ CD27+ CD43+ CD38lo/int is now considered a more rigorous phenotype of human B1 cells. Prabhu et al. [45], defining B1 cells as CD19+ CD20bright CD38dull CD27+ CD43+, also found an increase in B1 cells from neonatal cord blood compared to adults, consistent with our data. Future studies should compare the various B1 cell definitions in the same samples to evaluate results in disease-associated or immunologically regulated contexts.
Overall, this study provides valuable insights into the age-related changes in human B cell subpopulations, with a particular focus on the crucial transition from intrauterine to extrauterine life. By analyzing 8 distinct B cell subsets in cord blood and adolescent and adult venous blood, comprehensive reference values have been generated that may serve as a benchmark for future research and clinical assessments. Our findings highlight the dynamic nature of B cell development and the ongoing maturation of the immune system throughout life. One notable finding is the unexpected pattern of transitional B cells, which were found at similar frequencies in neonates and adults, with a significant dip in adolescence. The latter challenges previous reports suggesting a simple age-related decline in transitional B cells [14,15] and suggests a more complex developmental trajectory. Furthermore, our confirmation of relatively stable B1 cell frequencies across age groups, consistent with Prabhu et al. [45], contributes to the ongoing debate regarding the identity and ontogeny of human B1 cells.
These findings have broader implications for understanding immune system development, optimizing vaccination strategies, and assessing disease susceptibility in newborns and infants. A deeper understanding of B cell subset distribution in early life could inform the development of targeted interventions to enhance immune protection and reduce the risk of infections and immune-mediated diseases. While this study provides valuable data on B cell subsets in cord blood and peripheral blood, it is important to acknowledge its limitations. The small sample size and the restricted number of markers used for B cell phenotyping may limit the generalizability of our findings. Additionally, peripheral blood may not fully reflect the true distribution of B cells in other immune compartments, such as the bone marrow and lymphoid tissues.
Despite these limitations, our findings are largely consistent with previous research on B cell development. The higher frequency of naïve B cells and lower frequencies of memory B cells in cord blood compared to adult blood align with previous reports [14]. However, the observed pattern of transitional B cells differs from some studies [14,15], highlighting the need for further research clarifying the potential dynamic distribution of B cell subsets. The observed age-related changes in B cell subpopulations can be explained by a combination of factors, including antigen exposure, B cell maturation processes, and the development of immunological memory. The high proportion of naïve B cells in cord blood reflects the limited antigen exposure in utero. As newborns encounter environmental antigens, the adaptive immune system is activated, leading to the generation of memory B cells and a gradual shift towards a more mature B cell repertoire.
The dynamic changes in transitional B cells may reflect intervals of B cell development and differentiation occurring at different ages. The initial interval in neonates could represent the transition of immature B cells from the bone marrow to the periphery. The subsequent dip in children and adolescents, within the group called juveniles, might be due to the maturation of these cells into naïve B cells, followed by a potential increase again in adults, possibly due to ongoing B cell development or recruitment from other compartments.
In conclusion, we found that cell subpopulations differ considerably between cord blood, juveniles, and adult blood. These changes likely reflect a combination of developmental maturation and the establishment of adaptive immunological memory. Limitations of this study include the small sample size, and the limited number of markers used for B cell phenotyping. Additionally, peripheral blood may not accurately reflect the true quantity of B cells, as plasmablasts tend to localize in the bone marrow. Given the scarcity of studies on B cell subsets in cord blood, the strength of our study lies in its focus on cord blood as a crucial link between intrauterine and extrauterine development. Further research focusing on the longitudinal changes in these B-cell populations and their functional capabilities is warranted to fully elucidate the development of a competent adaptive immune system in early life
4. Materials and Methods
4.1. Patient Samples
Umbilical cord blood samples were obtained from term neonates (gestational age: 37-42 weeks) following uncomplicated deliveries. Peripheral blood samples were collected from healthy infants, children and adolescence, called juvenils throughout the paper (age: 1-17 years; mean: 4 years) and adults (age: 24-62 years; mean: 37.5 years). All participants were of Caucasian ethnicity, without any history of infectious, immunological, hematological, or other chronic diseases, nor were they receiving any treatment that could potentially affect the immune system.
Cord blood was collected via venipuncture of the umbilical veine into vacutainer tubes containing ethylenediaminetetraacetic acid (EDTA) immediately after delivery. Peripheral venous blood samples from juvenils and adult donors were similarly obtained in EDTA tubes.
This study was conducted in accordance with the ethical principles outlined in the World Medical Association's Declaration of Helsinki. Ethical approval was granted by the Institutional Review Board of Philipps-University Marburg, and written informed consent was obtained from the parents of neonates and from adult participants.
4.2. Cell Isolation and B Cell Enrichment
For cell isolation, we used 8-10 ml of blood. Erythrocytes were lysed and peripheral blood mononuclear cells (PBMCs) or cord blood mononuclear cells (CBMC) were isolated by Ficoll-Hypaque gradient (PAA, Linz, Austria). 1 x 107 cells untouched B cells were enriched by negative selection using Magnetic Bead-Activated Cell Sorting (MACS) with B cell (B-CLL) isolation kit (MiltenyiBiotec, Bergisch Gladbach, Germany) according to the manufacturer`s instructions.
4.3. Flow Cytometry
To quantify the B lymphocyte subsets, they were labelled according to Supplementary Table S1 with anti-CD19 (PE/APC), anti-CD20-FITC, anti-CD27 (FITC/PE/PerCP), anti-CD38 (APC), anti-CD43 (APC), anti-CD69 (PE), anti-IgD (FITC) (all from BD Biosciences, Heidelberg, Germany) and anti-IgM (PerCP) (BioLegend, Fell, Germany). All staining steps were performed in CellWASH buffer (BD Biosciences, Heidelberg, Germany). Cells were stained using a master mix of antibodies to maintain consistency among the samples.
Flow cytometry was directly performed on a four-colour FACS Calibur flow cytometer (BD Biosciences, Heidelberg, Germany) using Summit 4.3 software (Beckman Coulter, Krefeld, Germany). Cells were gated into a physical lymphocyte gate in the forward scatter/side scatter plot. Gates were preset and the measurements were performed blinded for sample identity. Live/dead cell discrimination was carried out by staining with propidium iodide (PI).
4.4. Statistical Analysis
Statistical analyses were performed using Prism 5.0 (GraphPad Software, La Jolla, California) and SPSS 28.0.0.(IBM, Chicago, IL,USA). Group differences were tested using three types of statistical tests. For comparsion of two different groups the two-tailed Student t-test was used. To determine if there is a significant difference between three groups Kruskal-Wallis test was used, if normal distribution was not given. Ordinary one-way ANOVA test was used for normal distribution. The p-values were two sided and subjected to a significance level of 0,05. Due to the explorative nature of the investigation we did not account for multiple statistical testing, and therefore reported raw, unadjusted p-values. Means are given with standard error (SEM, Table 1).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
SGF: data analysis and interpretation, manuscript writing, final approval of manuscript. MB: data interpretation, manuscript writing, final approval of manuscript. NNT, CS, TS: manuscript writing, final approval of manuscript. SM: collection and assembly of data, data analysis and interpretation, manuscript writing, final approval of manuscript. TR, SK, RFM, MZ: conception and design, manuscript writing, final approval of manuscript.
Funding
Funded by a grant from the “von Behring-Roentgen Foundation” and by a grant from the “Rhoen Klinikum AG”.
Institutional Review Board Statement
The Ethics Committee of the Philipps-University Marburg (AZ 20/07) approved this study.
Informed Consent Statement
The written informed consent of parents and adult donors was obtained
Data Availability Statement
The datasets supporting the conclusions of this article are included within the article or are available from the authors upon request.
Acknowledgments
We thank Sabine Jennemann and Regina Stoehr for excellent technical help. We would also like to thank Regine Weber and Elisabeth Kaiser for their support with the manuscript.
Conflicts of Interest
Tobias Rogosch is an employee of CSL Behring. The remaining authors declare no conflict of interest.
References
- LeBien, T.W.; Tedder, T.F. B lymphocytes: how they develop and function. Blood 2008, 112, 1570–1580. [Google Scholar] [CrossRef]
- Comans-Bitter, W.M.; de Groot, R.; van den Beemd, R.; Neijens, H.J.; Hop, W.C.; Groeneveld, K.; Hooijkaas, H.; van Dongen, J.J. Immunophenotyping of blood lymphocytes in childhood. Reference values for lymphocyte subpopulations. J Pediatr 1997, 130, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Weiskopf, D.; Weinberger, B.; Grubeck-Loebenstein, B. The aging of the immune system. Transpl Int 2009, 22, 1041–1050. [Google Scholar] [CrossRef] [PubMed]
- Cerutti, A.; Cols, M.; Puga, I. Marginal zone B cells: virtues of innate-like antibody-producing lymphocytes. Nat Rev Immunol 2013, 13, 118–132. [Google Scholar] [CrossRef] [PubMed]
- Berrón-Ruíz, L.; López-Herrera, G.; Ávalos-Martínez, C.E.; Valenzuela-Ponce, C.; Ramírez-SanJuan, E.; Santoyo-Sánchez, G.; Mújica Guzmán, F.; Espinosa-Rosales, F.J.; Santos-Argumedo, L. Variations of B cell subpopulations in peripheral blood of healthy Mexican population according to age: Relevance for diagnosis of primary immunodeficiencies. Allergol Immunopathol (Madr) 2016, 44, 571–579. [Google Scholar] [CrossRef]
- Yoshida, T.; Mei, H.; Dörner, T.; Hiepe, F.; Radbruch, A.; Fillatreau, S.; Hoyer, B.F. Memory B and memory plasma cells. Immunol Rev 2010, 237, 117–139. [Google Scholar] [CrossRef]
- Tarlinton, D.; Good-Jacobson, K. Diversity among memory B cells: origin, consequences, and utility. Science 2013, 341, 1205–1211. [Google Scholar] [CrossRef]
- Baumgarth, N. The double life of a B-1 cell: self-reactivity selects for protective effector functions. Nat Rev Immunol 2011, 11, 34–46. [Google Scholar] [CrossRef]
- Hardy, R.R. B-1 B cell development. J Immunol 2006, 177, 2749–2754. [Google Scholar] [CrossRef]
- Klein, U.; Rajewsky, K.; Küppers, R. Human immunoglobulin (Ig)M+IgD+ peripheral blood B cells expressing the CD27 cell surface antigen carry somatically mutated variable region genes: CD27 as a general marker for somatically mutated (memory) B cells. J Exp Med 1998, 188, 1679–1689. [Google Scholar] [CrossRef]
- Tangye, S.G.; Liu, Y.J.; Aversa, G.; Phillips, J.H.; de Vries, J.E. Identification of functional human splenic memory B cells by expression of CD148 and CD27. J Exp Med 1998, 188, 1691–1703. [Google Scholar] [CrossRef] [PubMed]
- Griffin, D.O.; Holodick, N.E.; Rothstein, T.L. Human B1 cells in umbilical cord and adult peripheral blood express the novel phenotype CD20+ CD27+ CD43+ CD70. J Exp Med 2011, 208, 67–80. [Google Scholar] [CrossRef]
- Morbach, H.; Eichhorn, E.M.; Liese, J.G.; Girschick, H.J. Reference values for B cell subpopulations from infancy to adulthood. Clin Exp Immunol 2010, 162, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Piątosa, B.; Wolska-Kuśnierz, B.; Pac, M.; Siewiera, K.; Gałkowska, E.; Bernatowska, E. B cell subsets in healthy children: reference values for evaluation of B cell maturation process in peripheral blood. Cytometry B Clin Cytom 2010, 78, 372–381. [Google Scholar] [CrossRef]
- Budeus, B.; Kibler, A.; Brauser, M.; Homp, E.; Bronischewski, K.; Ross, J.A.; Görgens, A.; Weniger, M.A.; Dunst, J.; Kreslavsky, T. , et al. Human Cord Blood B Cells Differ from the Adult Counterpart by Conserved Ig Repertoires and Accelerated Response Dynamics. J Immunol 2021, 206, 2839–2851. [Google Scholar] [CrossRef]
- Borriello, F.; Pasquarelli, N.; Law, L.; Rand, K.; Raposo, C.; Wei, W.; Craveiro, L.; Derfuss, T. Normal B-cell ranges in infants: A systematic review and meta-analysis. Journal of Allergy and Clinical Immunology 2022, 150, 1216–1224. [Google Scholar] [CrossRef]
- Marinkovic, D.; Marinkovic, T. Putative role of marginal zone B cells in pathophysiological processes. Scand J Immunol 2020, 92, e12920. [Google Scholar] [CrossRef] [PubMed]
- Montecino-Rodriguez, E.; Dorshkind, K. B-1 B Cell Development in the Fetus and Adult. Immunity 2012, 36, 13–21. [Google Scholar] [CrossRef]
- Hong, H.; Lee, J.; Park, G.-Y.; Kim, S.; Park, J.; Park, J.S.; Song, Y.; Lee, S.; Kim, T.J.; Lee, Y.J. , et al. Postnatal regulation of B-1a cell development and survival by the CIC-PER2-BHLHE41 axis. Cell Reports 2022, 38, 110386. [Google Scholar] [CrossRef]
- Merino-Vico, A.; Frazzei, G.; van Hamburg, J.P.; Tas, S.W. Targeting B cells and plasma cells in autoimmune diseases: From established treatments to novel therapeutic approaches. Eur J Immunol 2023, 53, e2149675. [Google Scholar] [CrossRef]
- Marie-Cardine, A.; Divay, F.; Dutot, I.; Green, A.; Perdrix, A.; Boyer, O.; Contentin, N.; Tilly, H.; Tron, F.; Vannier, J.P. , et al. Transitional B cells in humans: characterization and insight from B lymphocyte reconstitution after hematopoietic stem cell transplantation. Clin Immunol 2008, 127, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Weller, S.; Braun, M.C.; Tan, B.K.; Rosenwald, A.; Cordier, C.; Conley, M.E.; Plebani, A.; Kumararatne, D.S.; Bonnet, D.; Tournilhac, O. , et al. Human blood IgM "memory" B cells are circulating splenic marginal zone B cells harboring a prediversified immunoglobulin repertoire. Blood 2004, 104, 3647–3654. [Google Scholar] [CrossRef]
- Lee, J.; Kuchen, S.; Fischer, R.; Chang, S.; Lipsky, P.E. Identification and characterization of a human CD5+ pre-naive B cell population. J Immunol 2009, 182, 4116–4126. [Google Scholar] [CrossRef]
- Sims, G.P.; Ettinger, R.; Shirota, Y.; Yarboro, C.H.; Illei, G.G.; Lipsky, P.E. Identification and characterization of circulating human transitional B cells. Blood 2005, 105, 4390–4398. [Google Scholar] [CrossRef] [PubMed]
- Palanichamy, A.; Barnard, J.; Zheng, B.; Owen, T.; Quach, T.; Wei, C.; Looney, R.J.; Sanz, I.; Anolik, J.H. Novel human transitional B cell populations revealed by B cell depletion therapy. J Immunol 2009, 182, 5982–5993. [Google Scholar] [CrossRef]
- Huck, K.; Feyen, O.; Ghosh, S.; Beltz, K.; Bellert, S.; Niehues, T. Memory B-cells in healthy and antibody-deficient children. Clin Immunol 2009, 131, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Viau, M.; Zouali, M. B-lymphocytes, innate immunity, and autoimmunity. Clin Immunol 2005, 114, 17–26. [Google Scholar] [CrossRef]
- Warnatz, K.; Schlesier, M. Flowcytometric phenotyping of common variable immunodeficiency. Cytometry B Clin Cytom 2008, 74, 261–271. [Google Scholar] [CrossRef]
- Wehr, C.; Kivioja, T.; Schmitt, C.; Ferry, B.; Witte, T.; Eren, E.; Vlkova, M.; Hernandez, M.; Detkova, D.; Bos, P.R. , et al. The EUROclass trial: defining subgroups in common variable immunodeficiency. Blood 2008, 111, 77–85. [Google Scholar] [CrossRef]
- Hayakawa, K.; Hardy, R.R.; Parks, D.R.; Herzenberg, L.A. The "Ly-1 B" cell subpopulation in normal immunodefective, and autoimmune mice. J Exp Med 1983, 157, 202–218. [Google Scholar] [CrossRef]
- Banchereau, J.; Rousset, F. Human B lymphocytes: phenotype, proliferation, and differentiation. Adv Immunol 1992, 52, 125–262. [Google Scholar] [CrossRef]
- Sanz, I.; Wei, C.; Lee, F.E.; Anolik, J. Phenotypic and functional heterogeneity of human memory B cells. Semin Immunol 2008, 20, 67–82. [Google Scholar] [CrossRef]
- Hayakawa, K.; Hardy, R.R.; Stall, A.M.; Herzenberg, L.A.; Herzenberg, L.A. Immunoglobulin-bearing B cells reconstitute and maintain the murine Ly-1 B cell lineage. Eur J Immunol 1986, 16, 1313–1316. [Google Scholar] [CrossRef] [PubMed]
- Sidman, C.L.; Shultz, L.D.; Hardy, R.R.; Hayakawa, K.; Herzenberg, L.A. Production of immunoglobulin isotypes by Ly-1+ B cells in viable motheaten and normal mice. Science 1986, 232, 1423–1425. [Google Scholar] [CrossRef]
- Gupta, R.; Jain, P.; Deo, S.V.; Sharma, A. Flow cytometric analysis of CD5+ B cells: a frame of reference for minimal residual disease analysis in chronic lymphocytic leukemia. Am J Clin Pathol 2004, 121, 368–372. [Google Scholar] [CrossRef] [PubMed]
- Wuttke, N.J.; Macardle, P.J.; Zola, H. Blood group antibodies are made by CD5+ and by CD5- B cells. Immunol Cell Biol 1997, 75, 478–483. [Google Scholar] [CrossRef] [PubMed]
- Jego, G.; Bataille, R.; Pellat-Deceunynck, C. Interleukin-6 is a growth factor for nonmalignant human plasmablasts. Blood 2001, 97, 1817–1822. [Google Scholar] [CrossRef]
- Descatoire, M.; Weill, J.C.; Reynaud, C.A.; Weller, S. A human equivalent of mouse B-1 cells? J Exp Med 2011, 208, 2563–2564. [Google Scholar] [CrossRef]
- Perez-Andres, M.; Grosserichter-Wagener, C.; Teodosio, C.; van Dongen, J.J.; Orfao, A.; van Zelm, M.C. The nature of circulating CD27+CD43+ B cells. J Exp Med 2011, 208, 2565–2566. [Google Scholar] [CrossRef]
- Haas, K.M.; Blevins, M.W.; High, K.P.; Pang, B.; Swords, W.E.; Yammani, R.D. Aging promotes B-1b cell responses to native, but not protein-conjugated, pneumococcal polysaccharides: implications for vaccine protection in older adults. J Infect Dis 2014, 209, 87–97. [Google Scholar] [CrossRef]
- Griffin, D.O.; Rothstein, T.L. Human b1 cell frequency: isolation and analysis of human b1 cells. Front Immunol 2012, 3, 122. [Google Scholar] [CrossRef] [PubMed]
- Covens, K.; Verbinnen, B.; Geukens, N.; Meyts, I.; Schuit, F.; Van Lommel, L.; Jacquemin, M.; Bossuyt, X. Characterization of proposed human B-1 cells reveals pre-plasmablast phenotype. Blood 2013, 121, 5176–5183. [Google Scholar] [CrossRef] [PubMed]
- Inui, M.; Hirota, S.; Hirano, K.; Fujii, H.; Sugahara-Tobinai, A.; Ishii, T.; Harigae, H.; Takai, T. Human CD43+ B cells are closely related not only to memory B cells phenotypically but also to plasmablasts developmentally in healthy individuals. Int Immunol 2015, 27, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Quách, T.D.; Rodríguez-Zhurbenko, N.; Hopkins, T.J.; Guo, X.; Hernández, A.M.; Li, W.; Rothstein, T.L. Distinctions among Circulating Antibody-Secreting Cell Populations, Including B-1 Cells, in Human Adult Peripheral Blood. J Immunol 2016, 196, 1060–1069. [Google Scholar] [CrossRef]
- Prabhu, S.B.; Rathore, D.K.; Nair, D.; Chaudhary, A.; Raza, S.; Kanodia, P.; Sopory, S.; George, A.; Rath, S.; Bal, V. , et al. Comparison of Human Neonatal and Adult Blood Leukocyte Subset Composition Phenotypes. PLoS One 2016, 11, e0162242. [Google Scholar] [CrossRef]
Figure 1.
Gating strategy and B cells subpopulations frequency. (A) SSC vs FSC density plot. (B) B cells: CD19+ (C), immature B cells (CD19+ CD27- IgM- IgD-), naive B cells (CD19+ CD27- IgM+ IgD+), (D) transitional B cells (CD19+ CD27- IgM+ CD38+), (E) marginal zone B cells (CD19+ CD27+ IgM+ IgD+), IgM memory B cells (CD19+ CD27+ IgM+ IgD-), class-switched memory B cells (CD19+ CD27+ IgM- IgD-), (F) plasmablasts (CD19+ CD27+ IgM- CD38++).
Figure 1.
Gating strategy and B cells subpopulations frequency. (A) SSC vs FSC density plot. (B) B cells: CD19+ (C), immature B cells (CD19+ CD27- IgM- IgD-), naive B cells (CD19+ CD27- IgM+ IgD+), (D) transitional B cells (CD19+ CD27- IgM+ CD38+), (E) marginal zone B cells (CD19+ CD27+ IgM+ IgD+), IgM memory B cells (CD19+ CD27+ IgM+ IgD-), class-switched memory B cells (CD19+ CD27+ IgM- IgD-), (F) plasmablasts (CD19+ CD27+ IgM- CD38++).
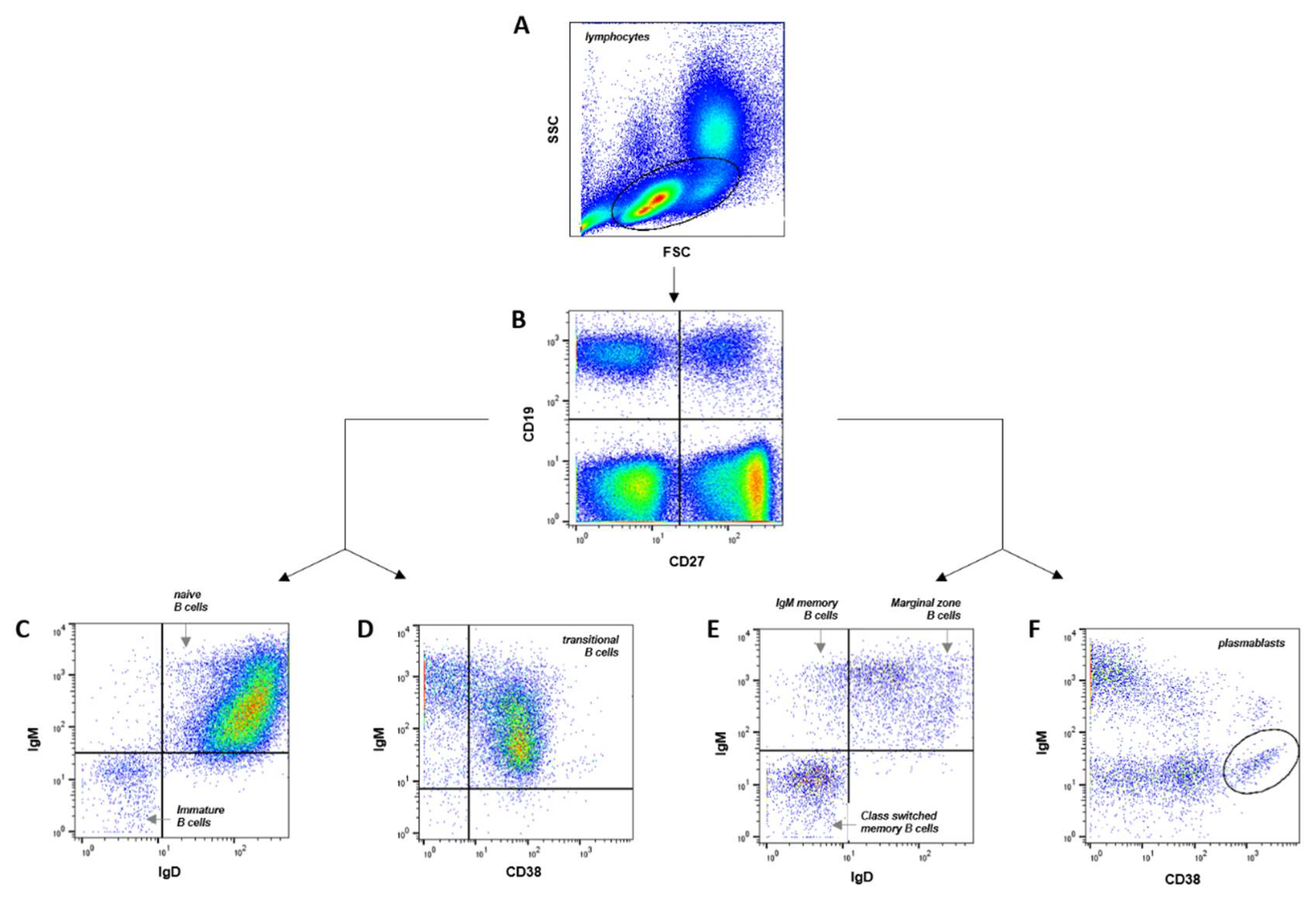

Table 1.
B cell populations in cord blood, juveniles and adult venous blood. Immature B cells, naïve B cells, transitional B cells, marginal zone B cells, IgM memory B cells, class switched memory B cells and plasmablasts values refer to percentage of CD19+ cells, B1 cells values refer to percentage of CD20+ cells.
Table 1.
B cell populations in cord blood, juveniles and adult venous blood. Immature B cells, naïve B cells, transitional B cells, marginal zone B cells, IgM memory B cells, class switched memory B cells and plasmablasts values refer to percentage of CD19+ cells, B1 cells values refer to percentage of CD20+ cells.
| Lymphocyte subset | Neonates (Cord blood) |
Juveniles (Peripheral blood) |
Adults (Peripheral blood) |
|---|---|---|---|
| immature B cells | 1.4 ± 0.3 % | 1.7 ± 0.3 % | 2.3 ± 0.3 % |
| naive B cells | 93.2 ± 1.2 % | 80.8 ± 1.8 % | 69.1 ± 3.4 % |
| transitional B cells | 27.9 ± 5.7 % | 9.0 ± 1.0 % | 30.9 ± 3.7 % |
| marginal zone B cells | 2.5 ± 0.4 % | 7.1 ± 0.8 % | 10.7 ± 1.4 % |
| IgM memory B cells | 0.2 ± 0.1 % | 1.4 ± 0.2 % | 1.7 ± 0.3 % |
| class switched memory B cells | 0.5 ± 0.1 % | 6.4 ± 1.0 % | 14.1 ± 2.0 % |
| plasmablasts | 0.5 ± 0.2 % | 0.7 ± 0.1 % | 1.4 ± 0.3 % |
| B1 cells (% of CD20+ cells) | 1.9 ± 0.3 % | - | 2.7 ± 0.4 % |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



