Submitted:
19 March 2025
Posted:
20 March 2025
You are already at the latest version
Abstract
Pulmonary arterial hypertension is characterized by vascular and systemic inflammation. The gut microbiome influences the host immune system. Here we review the emerging preclinical and clinical evidence that strongly suggest that alterations in the gut microbiome may either initiate or facilitate progression of established pulmonary arterial hypertension by modifying systemic immune responses. We also briefly describe studies delineating the contributions of infections to pulmonary arterial hypertension pathogenesis.
Keywords:
gut microbiome
; pulmonary arterial hypertension
; inflammation
Vascular injury and inflammation are key drivers of pulmonary vascular remodeling in pulmonary arterial hypertension (PAH) [1,2,3,4,5]. Primarily known as a cardiopulmonary disease, growing evidence has implicated interorgan communication in PAH pathogenesis. We and others have observed the potential role of the intestinal microbiome in the onset and progression of PAH. The gastrointestinal system contributes to systemic inflammation as it contains about 70-80% of the body’s immune cells [6,7], along with trillions of microorganisms known as the gut microbiome [8].The composition of the gut microbiome can be influenced by many factors including environmental pollutants, medications, nutrient availability/diet, exercise, oxygen level, sex, host's developmental stage, and genetics (Figure 1) [9]. There is now evidence of the gut microbiome’s impact on many chronic diseases [10]. Clinical trials of targeted anti-inflammatory drugs to date have not clearly demonstrated benefits in PAH [11,12,13], partially due to enrollment size, trial design and patient selection. Modulating a key immune organ, such as the intestine, to alter many inflammatory pathways may be an effective adjunctive approach to treating PAH.
This review describes the experimental and clinical evidence that implicates the gut microbiota as key contributors of PAH pathogenesis. Additionally, we provide a brief overview on the effect of infections on PAH development and the gut microbiome.
1. Preclinical Studies of the Gut-Lung Axis in Pulmonary Hypertension
Multiple studies demonstrate that pulmonary hypertension (PH) rodent models have alterations in the gut microbiome commonly referred to as gut dysbiosis [14,15,16,17,18,19,20,21,22]. Monocrotaline (MCT) PAH rats have increased intestinal permeability as measured by fluorescein isothiocyanate (FITC)-dextran, lipopolysaccharide (LPS), and soluble CD14 in the blood [23] and altered intestinal morphology, with greater muscularis layer thickness and fibrosis and diminished villus length and goblet cell number [20,24]. Sugen hypoxia (SuHx) rats have an elevated fecal Firmicutes-to-Bacteroidetes ratio and less abundance of short chain fatty acid (SCFA)-producing bacteria, such as those that are acetate and butyrate-producing, with no change in lactate-producing bacteria [14]. The milder hypoxic rat PH model also has altered and distinct gut microbiota with more arginine and arginine-producing bacteria, Blautia and Bifidobacterium, and the trimethylamine N-oxide (TMAO) biosynthetic bacteria, Streptococcus [18]. Hypoxic mice have disrupted gut microbiome composition with increases of the genera Prevotella, Oscillospira, and Ruminococcus and decreases in Lactobacillus [25]. Alterations in the gut microbiome also occur in the large animal, bovine brisket disease PH model, with lower total volatile fatty acids and alpha diversity (richness and evenness of bacteria) in rumen fluid [26]. The specific intestinal microbiome differences observed in various PH rodent models are comprehensively reviewed elsewhere [27].
Several preclinical studies have modified the gut microbiome to assess its effects on PAH. Altering the microbiome with a broad-spectrum antibiotic cocktail (ampicillin, vancomycin, neomycin, and metronidazole) prior to SU5416 administration mitigates PH in SuHx rats [28]. Additionally, diet impacts PH pathogenesis and severity. Apolipoprotein E (ApoE) knockout mice given a Paigen (high fat, high cholesterol) diet develop PH [29] compared to ApoE knockout mice fed normal chow. Double ApoE/IL1R1 knockout mice consuming a Paigen diet have even worse PAH, implicating the role of IL-1 in PAH pathogenesis [29]. Western diet increases right ventricular systolic pressure (RVSP) and RV myocardial lipid deposition and reduces RV function in mice [30]. Metformin decreases RVSP and RV lipid and ceramide accumulation [30]. Moreover, high soluble-fiber diet attenuates hypoxia-induced pulmonary vascular remodeling by increasing abundance of SCFA-producing bacteria (Bacteroides, Anaerostripes, and Anaerocolumna), diminishing pro-inflammatory bacteria (Romboutsia, Mammalicoccus, Staphylococcus, Clostridioides, and Streptococcus), and reducing lung interstitial macrophages, dendritic cells, and nonclassical monocytes [31]. The serum metabolites, phosphatidylcholines, lysophosphatidylcholines, ceramides, and hexosylceramides, are lower while propionylcarnitine and probetaine are greater in high soluble-fiber fed mice compared to low soluble-fiber fed hypoxic mice [31]. Interestingly, treatment with a phosphodiesterase-5 inhibitor (tadalafil) or endothelin pathway inhibitor (macitentan) in SuHx rats reduces the plasma levels of many phosphatidylcholines [32]. However, not all suspected advantageous dietary interventions have been effective in PAH. Although intermittent fasting augments RV function and extends survival in MCT rats, there is minimal effect on PH severity [24], suggesting that more targeted dietary interventions may be needed to alter PH severity.
One of the primary ways that the gut microbiota interact with their host is via metabolites which are intermediate or end products of microbial metabolism [33]. The gut metabolites are generated from bacterial metabolism of dietary or other host substrates. Supplementation with the SCFA, butyrate, which is an endogenous histone deacetylase (HDAC) inhibitor, attenuates pulmonary vascular remodeling and accumulation of alveolar (CD68+) and interstitial (CD68+ and CD163+) lung macrophages in hypoxic PH [34]. Butyrate’s positive effects on pulmonary vascular remodeling have been corroborated in other unpublished studies [35,36] that suggest that butyrate also regulates endothelial cell inflammatory activation and migration. However, the improvement in PH with butyrate treatment appears to only be seen in prevention models but not when given after two- and four-weeks of hypoxic exposure [34].
Trimethylamine N-oxide (TMAO), derived from trimethylamine (TMA), is a bacterial metabolite generated by the breakdown of dietary choline, carnitine, and betaine. It is elevated in intermediate and high risk idiopathic PAH (IPAH) patients [37]. Administering TMAO to hypoxic mice worsens PH through macrophage secretion [37]. Treating hypoxic and MCT rodents with a structural analog of choline that inhibits TMAO synthesis, 3,3-dimethyl-1-butanol (DMB), reduces RV systolic pressure and pulmonary vascular thickness/muscularization and suppresses cytokine and chemokine signaling, with the strongest associations with Cxcl6 and Il6 [37,38]. While TMAO worsens PH, long-term TMAO treatment of MCT rats may be beneficial to RV function by preserving fatty acid oxidation and decreasing pyruvate metabolism, thus preserving mitochondrial energy metabolism and mitigating the development of RV dysfunction in PAH [39]. The role of other microbial metabolites [33,39,40,41], such as amino acid metabolites and retinoic and bile acids, in PAH and RV failure is not well-defined.
There are few publications investigating the outcomes of direct manipulation of the microbiome or supplementation of specific bacterial genera/species in PAH. Fecal transplant from angiotensin-converting enzyme 2 (Ace2) overexpressing mice, which have less hypoxia-induced PH, to wild-type hypoxic mice attenuates PH development [25,42]. Administration of Lactobacillus reuteri to postnatal growth restriction pups exposed to hyperoxia mitigates PH severity and RV hypertrophy [43]. Lactobacillus rhamnosus supplementation two weeks after MCT injection does not affect pulmonary vascular remodeling but enhances RV function [44].
Other approaches to modulate the microbiome involve human umbilical cord blood-derived mesenchymal stem cells (MSCs), which are nonhematopoietic cells that can self-renew and secrete anti-bacterial peptides [45]. There is recent interest in their ability to regulate the gut microbiome and alleviate inflammatory bowel diseases [46,47,48]. Treatment with MSCs rebalances the gut microbiome (reducing the disease-associated and increasing the anti-inflammatory bacteria) and attenuates hypoxia [16]- and MCT-induced PH [49,50].
Most of the animal studies modifying the gut microbiota in PH thus far use prevention models, intervening prior to or immediately after providing a stimulus to generate PH. Promising indirect evidence supports a need to further explore the role of the gut microbiome in PH. In SuHx mice, treatment with Ang1-7 four weeks after hypoxia initiation mitigates PH, partially attenuates disease-associated changes in gut microbiota, and enhances the beneficial metabolites, butyric acid and tryptophan [51]. dAdministering irbesartan, an angiotensin II receptor blocker, 30 days after starting exposure to hypoxia mitigates PH, partially normalizes the Firmicutes-to-Bacteroidetes ratio, increases intestinal abundance of Lactobacillaceae and Lachnospiraceae, and decreases Prevotellaceae and Desulfovibrionaceae in high-altitude PH hypobaric hypoxia rats [21]. Future studies need to evaluate the efficacy of altering the gut microbiome after PAH is established. Due to the many potential environmental confounders in clinical microbiome research, animal studies are needed to establish a robust foundation for biotherapeutics targeting host microbiome composition and its systemic effects on respiratory health. Lastly, the potential gut-brain-lung axis in PAH [20,25,42] should be further explored.
2. Clinical Evidence of the Gut-Lung Axis in PAH
Clinical studies have shown that PAH patients have gut dysbiosis or a leaky gut with increased bacterial translocation from the intestinal lumen to systemic circulation [23]. Kim et al. [52] completed one of the initial clinical studies describing the distinct gut microbiome composition in PAH by studying the fecal microbiome of 18 PAH patients and 12 age- and sex-matched healthy controls. Compared to controls, PAH patients have a distinct microbiome composition with lower alpha diversity, fewer bacteria associated with polysaccharide fermentation and SCFA production (Butyrivibrio crossotus, Bacteroides cellulosilyticus, Eubacterium siraeum, Bacteroides vulgatus, Akkermansia muciniphila), and more bacteria associated with the proinflammatory metabolites, TMA and TMAO [52]. PAH patients also have a disparate intestinal virome [52].
In a subsequent single-center pilot study of 20 PAH and 20 healthy controls cohabiting with PAH patients (20 matched pairs), Jose et al. [53] observed no difference in alpha (within a specific sample) or beta diversity (between samples). In the largest PAH microbiota study to date of 72 patients, Moutsoglou et al. [54] demonstrated that the gut microbiome is less diverse in PAH patients compared to healthy controls and family members residing in the same household. Gut microbiome diversity correlates with measures of pulmonary vascular disease (mean pulmonary artery pressure, pulmonary vascular resistance, and pulmonary arterial compliance), but not RV function [54], suggesting that the alterations in the gut microbiome are not due to RV failure and intestinal congestion. PAH patients have reduced abundance of gut bacteria containing genes encoding for the production of anti-inflammatory metabolites, specifically SCFAs (Eubacterium ramulus, Firmicutes sp. coabundance gene 110, Coprococcus comes, Dorea longicatena, Bifidobacterium adolescentis, Gemmiger formicilis, Fusicatenibacter saccharivorans, Eubacterium hallii, Anaerostipes hadrus, Gordonibacter pamelaeae, Ruminococcus torques, Coprococcus catus, Coprococcus eutactus, and Blautia obeum) and secondary bile acids (Collinsella aerofaciens, Coprococcus eutactus, Anaerostipes hadrus, Eubacterium ramulus, Blautia obeum, Eubacterium hallii, Ruminococcus bicirculans, Ruminococcus torques, Eubacterium eligens, Fusicatenibacter saccharivorans, Roseburia faecis, Dorea longicatena, Coprococcus catus, and Roseburia hominis) and increased relative abundance of bacteria with genes encoding for the production of TMAO (Clostridium bolteae, Escherichia coli, and Klebsiella pneumoniae) [54]. In a study of 35 patients with IPAH [37] and another of 124 PAH patients [38], TMAO levels are elevated in higher risk patients, suggesting that TMAO is associated with worse outcomes. The role of the other intestinal microorganisms (fungi, protozoans, and archaea) in clinical PAH is not well-established.
Environmental effects, such as high altitude and hypoxia, may also alter the gut microbiome. A small study of six highlander PH patients living on the Tibetan plateau range and seven lowlander PH patients (residents of Shanghai) showed that while there are overall divergent gut microbial signatures between PH patients and controls, altitude contributes to the gut microbiota differences [55]. TMA-synthesis enzymes are enriched in lowlanders with PH and there is no difference in TMA-producing microbiota between highlander controls and highlander PH patients [55].
Whether there is a causal relationship between the gut microbiome/metabolites and PAH is not yet determined in clinical studies. Mendelian randomization of data from the MiBioGen consortium, the largest genome-wide meta-analysis of intestinal microbiota [56], investigated the potential direct link between gut microbiota, metabolites, diet, and PAH [57]. The bacteria, Alistipes and Victivallis, correlate with increased PAH risk while Coprobacter, Erysipelotrichaeae, Lachnospiraceae, and Ruminococcaceae protect against PAH [57]. However, SCFAs, TMAO, and dietary patterns were not causally associated with PAH in the Mendelian randomization analysis [57]. In another recent Mendelian randomization study, Su et al. identified 11 gut microbial taxa, including Bifidobacteriaceae, Eubacterium eligens group, and Sutterella, and 24 bacterial metabolites that are linked to PAH pathogenesis by regulating the expression of ITPR2, IDE, NRIP1, and IGF1 genes in lung tissue [58]. Limitations of these studies are that the design only evaluates the impact of genetic factors on intestinal microbiota abundance and the development of PAH, small effect size with the use of single nucleotide polymorphisms (SNPs) for metabolites and bacteria, Mendelian randomization being prone to false positives, and cohorts primarily of European ethnicity, restricting the generalizability of the findings. Thus, more clinical studies are needed to determine whether there is a casual connection between the intestinal microbiota/metabolites and PAH.
There are now emerging data that not only is the fecal microbiome changed in PAH, but airway microbiome composition may also be distinct. In a study of PH patients of various etiologies (Group 1 PAH, PH due to lung disease, and chronic thromboembolic PH), Zhang et al. [59] observed higher alpha diversity (Ace and Sobs indices and Simpson index) and increased Streptococcus, Lautropia, and Ralstonia in the airways of PH patients compared to reference controls. Intratracheal instillation of Streptococcus induced PH in rats [60]. A recent study identified disparate airway mycobiomes or fungal compositions between PH and healthy controls [61]. Further research should assess the microbiome/mycobiome in other parts of the body.
3. Potential Approaches to Modulate the Gut-Lung Axis to Treat PAH
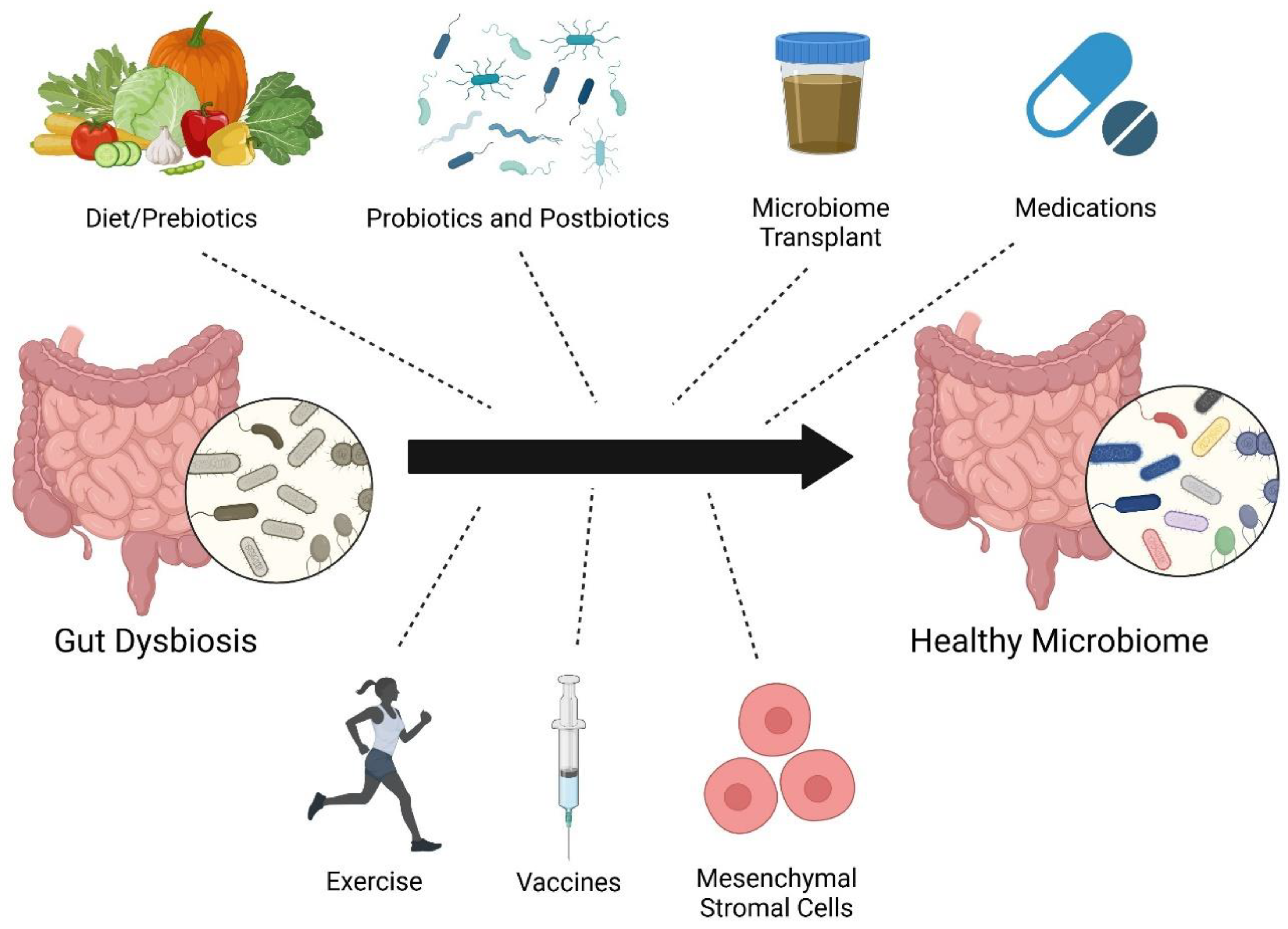
There are several different strategies to restructure the microbiome including diet/prebiotics, probiotics, postbiotics [62] (inanimate microorganisms or their components), microbiota transplant, medications, vaccines [63], exercise [64], and mesenchymal stromal cell therapy [50,65] (Figure 2). There have not yet been many clinical trials investigating how altering the gut microbiome affects PAH. There is an ongoing study evaluating the impact of microbiota transplant from healthy controls to PAH patients [66]. Unfortunately, the SARS-CoV-2 pandemic impeded the enrollment and initiation of the trial assessing the use of chlorhexidine mouthwash and oral nitrate therapy in PH patients (NCT03787082). Challenges to translating preclinical findings to PAH patients include genetic factors, age, sex, concomitant chronic diseases, and different environmental exposures [67] (medications, diet, timing of eating, toxin use, chemicals in environment/products, hygiene, air pollution, daylight exposure, etc.). Fetal/maternal or perinatal microbiota exposures may also complicate the efficacy of microbiota clinical trials. For example, supplementing omega-3 polyunsaturated fatty acids in the diet of pregnant rats improves RV systolic pressure and survival in pups exposed to hyperoxia at time of birth [68]. Additionally, the timing and duration of gut microbiome modulation needed to confer benefits in PAH are unknown. Despite these possible confounders, the vast potential health benefits of altering the microbiome in PAH should be explored to a greater extent.
4. Contribution of Infections in PAH Pathogenesis
Infectious agents, including bacteria, viruses, fungi, and parasites, can cause pulmonary arterial injury and inflammation by impacting vascular cells, leading to severe pulmonary vascular remodeling and PAH [69]. The pathophysiology and mechanisms by which infections lead to pulmonary vascular disease are more extensively reviewed elsewhere [70]. Infections in the setting of a dysregulated immune system may elevate susceptibility to PH development [71,72]. There is now growing interest in determining how host microbiota promote or resist infections [73].
A common global cause of PAH is infection by the intravascular parasite, Schistosoma mansoni. Schistosomiasis affects over 200 million people with about 1-10 million chronically infected people at risk for developing PAH [74,75]. Schistosomiasis cases occur in many regions worldwide, but the majority are observed in Africa and Asia, although longstanding epidemiological and socioeconomic challenges may underestimate its global impact. Schistosomiasis disrupts both gut and lung microbiota [76]. S. mansoni egg exposure decreases lung alpha diversity, mainly by impacting the relative abundance of the phylum, Ascomycota, while the pulmonary Firmicutes-to-Bacteroidetes ratio remains unchanged [76]. In contrast, Schistosomiasis increases gut microbiota alpha diversity and the Firmicutes-to-Bacteroidetes ratio [76], suggesting significant differences in lung and gut microbiome responses to Schistosomiasis infection.
HIV infection is also a well-recognized cause of PAH [77]. Host factors and geography contribute to the fecal microbiota disruptions that occur after HIV infection [78]. Geographic location has a greater effect on fecal microbiota composition than HIV infection status [78]. Interestingly, while HIV infection is known to disrupt gut epithelial barrier function, there are regional differences in immune activation with elevated soluble CD14 levels in HIV-infected individuals from all three regions studied (United States, Botswana, and Uganda), but there is increased intestinal fatty-acid binding protein in HIV-infected individuals from only the United States and Botswana [78]. Thus, distinct gut microbial alterations due to host region are major confounders in microbiota studies.
Unquestionably, additional work is needed to unravel the precise mechanisms contributing to the complex interactions between infections and host microbiota. Future studies should further ascertain whether microbiome alterations after infection impact susceptibility to PAH.
5. Conclusions
Animal and clinical studies reveal altered intestinal microbiota composition in PAH. Modulating the gut microbiome diminishes systemic inflammation and pulmonary immune cell infiltration. Additional research is needed to delineate whether restructuring the microbiome attenuates PAH after PAH has developed and explore the role of the gut and airway/lung microbiome in infection-associated PAH.
Acknowledgments
Figures were created with BioRender.com.
References
- Rabinovitch M, Guignabert C, Humbert M, Nicolls MR. Inflammation and immunity in the pathogenesis of pulmonary arterial hypertension. Circ Res. 2014;115:165-175. [CrossRef]
- Sweatt AJ, Hedlin HK, Balasubramanian V, Hsi A, Blum LK, Robinson WH, Haddad F, Hickey PM, Condliffe R, Lawrie A, et al. Discovery of Distinct Immune Phenotypes Using Machine Learning in Pulmonary Arterial Hypertension. Circ Res. 2019;124:904-919. [CrossRef]
- Steiner MK, Syrkina OL, Kolliputi N, Mark EJ, Hales CA, Waxman AB. Interleukin-6 overexpression induces pulmonary hypertension. Circ Res. 2009;104:236-244, 228p following 244. [CrossRef]
- Soon E, Crosby A, Southwood M, Yang P, Tajsic T, Toshner M, Appleby S, Shanahan CM, Bloch KD, Pepke-Zaba J, et al. Bone morphogenetic protein receptor type II deficiency and increased inflammatory cytokine production. A gateway to pulmonary arterial hypertension. Am J Respir Crit Care Med. 2015;192:859-872. [CrossRef]
- Savai R, Pullamsetti SS, Kolbe J, Bieniek E, Voswinckel R, Fink L, Scheed A, Ritter C, Dahal BK, Vater A, et al. Immune and inflammatory cell involvement in the pathology of idiopathic pulmonary arterial hypertension. Am J Respir Crit Care Med. 2012;186:897-908. [CrossRef]
- Furness JB, Kunze WA, Clerc N. Nutrient tasting and signaling mechanisms in the gut. II. The intestine as a sensory organ: neural, endocrine, and immune responses. Am J Physiol. 1999;277:G922-928. [CrossRef]
- Castro GA, Arntzen CJ. Immunophysiology of the gut: a research frontier for integrative studies of the common mucosal immune system. Am J Physiol. 1993;265:G599-610. [CrossRef]
- Thenappan T, Khoruts A, Chen Y, Weir EK. Can intestinal microbiota and circulating microbial products contribute to pulmonary arterial hypertension? Am J Physiol Heart Circ Physiol. 2019;317:H1093-H1101. [CrossRef]
- Van Hul M, Cani PD, Petitfils C, De Vos WM, Tilg H, El-Omar EM. What defines a healthy gut microbiome? Gut. 2024;73:1893-1908. [CrossRef]
- Lynch SV, Pedersen O. The Human Intestinal Microbiome in Health and Disease. N Engl J Med. 2016;375:2369-2379. [CrossRef]
- Trankle CR, Canada JM, Kadariya D, Markley R, De Chazal HM, Pinson J, Fox A, Van Tassell BW, Abbate A, Grinnan D. IL-1 Blockade Reduces Inflammation in Pulmonary Arterial Hypertension and Right Ventricular Failure: A Single-Arm, Open-Label, Phase IB/II Pilot Study. Am J Respir Crit Care Med. 2019;199:381-384. [CrossRef]
- Toshner M, Church C, Harbaum L, Rhodes C, Villar Moreschi SS, Liley J, Jones R, Arora A, Batai K, Desai AA, et al. Mendelian randomisation and experimental medicine approaches to interleukin-6 as a drug target in pulmonary arterial hypertension. Eur Respir J. 2022;59. [CrossRef]
- Zamanian RT, Badesch D, Chung L, Domsic RT, Medsger T, Pinckney A, Keyes-Elstein L, D'Aveta C, Spychala M, White RJ, et al. Safety and Efficacy of B-Cell Depletion with Rituximab for the Treatment of Systemic Sclerosis-associated Pulmonary Arterial Hypertension: A Multicenter, Double-Blind, Randomized, Placebo-controlled Trial. Am J Respir Crit Care Med. 2021;204:209-221. [CrossRef]
- Callejo M, Mondejar-Parreño G, Barreira B, Izquierdo-Garcia JL, Morales-Cano D, Esquivel-Ruiz S, Moreno L, Cogolludo Á, Duarte J, Perez-Vizcaino F. Pulmonary Arterial Hypertension Affects the Rat Gut Microbiome. Sci Rep. 2018;8:9681. [CrossRef]
- Hong W, Mo Q, Wang L, Peng F, Zhou Y, Zou W, Sun R, Liang C, Zheng M, Li H, et al. Changes in the gut microbiome and metabolome in a rat model of pulmonary arterial hypertension. Bioengineered. 2021;12:5173-5183. [CrossRef]
- Luo L, Chen Q, Yang L, Zhang Z, Xu J, Gou D. MSCs Therapy Reverse the Gut Microbiota in Hypoxia-Induced Pulmonary Hypertension Mice. Front Physiol. 2021;12:712139. [CrossRef]
- Cao W, Wang L, Mo Q, Peng F, Hong W, Zhou Y, Sun R, Li H, Liang C, Zhao D, et al. Disease-associated gut microbiome and metabolome changes in rats with chronic hypoxia-induced pulmonary hypertension. Front Cell Dev Biol. 2024;12:1022181. [CrossRef]
- Luo L, Yin H, Gou D. Gut Microbiota and Metabolome Changes in Three Pulmonary Hypertension Rat Models. Microorganisms. 2023;11. [CrossRef]
- Chen J, Zhou D, Miao J, Zhang C, Li X, Feng H, Xing Y, Zhang Z, Bao C, Lin Z, et al. Microbiome and metabolome dysbiosis of the gut-lung axis in pulmonary hypertension. Microbiol Res. 2022;265:127205. [CrossRef]
- Sharma RK, Oliveira AC, Yang T, Kim S, Zubcevic J, Aquino V, Lobaton GO, Goel R, Richards EM, Raizada MK. Pulmonary arterial hypertension-associated changes in gut pathology and microbiota. ERJ Open Res. 2020;6. [CrossRef]
- Nijiati Y, Maimaitiyiming D, Yang T, Li H, Aikemu A. Research on the improvement of oxidative stress in rats with high-altitude pulmonary hypertension through the participation of irbesartan in regulating intestinal flora. Eur Rev Med Pharmacol Sci. 2021;25:4540-4553. [CrossRef]
- Adak A, Maity C, Ghosh K, Mondal KC. Alteration of predominant gastrointestinal flora and oxidative damage of large intestine under simulated hypobaric hypoxia. Z Gastroenterol. 2014;52:180-186. [CrossRef]
- Ranchoux B, Bigorgne A, Hautefort A, Girerd B, Sitbon O, Montani D, Humbert M, Tcherakian C, Perros F. Gut-Lung Connection in Pulmonary Arterial Hypertension. Am J Respir Cell Mol Biol. 2017;56:402-405. [CrossRef]
- Prisco SZ, Eklund M, Moutsoglou DM, Prisco AR, Khoruts A, Weir EK, Thenappan T, Prins KW. Intermittent Fasting Enhances Right Ventricular Function in Preclinical Pulmonary Arterial Hypertension. J Am Heart Assoc. 2021;10:e022722. [CrossRef]
- Sharma RK, Oliveira AC, Yang T, Karas MM, Li J, Lobaton GO, Aquino VP, Robles-Vera I, de Kloet AD, Krause EG, et al. Gut Pathology and Its Rescue by ACE2 (Angiotensin-Converting Enzyme 2) in Hypoxia-Induced Pulmonary Hypertension. Hypertension. 2020;76:206-216. [CrossRef]
- Gaowa N, Panke-Buisse K, Wang S, Wang H, Cao Z, Wang Y, Yao K, Li S. Brisket Disease Is Associated with Lower Volatile Fatty Acid Production and Altered Rumen Microbiome in Holstein Heifers. Animals (Basel). 2020;10. [CrossRef]
- Huang L, Zhang H, Liu Y, Long Y. The Role of Gut and Airway Microbiota in Pulmonary Arterial Hypertension. Front Microbiol. 2022;13:929752. [CrossRef]
- Sanada TJ, Hosomi K, Shoji H, Park J, Naito A, Ikubo Y, Yanagisawa A, Kobayashi T, Miwa H, Suda R, et al. Gut microbiota modification suppresses the development of pulmonary arterial hypertension in an SU5416/hypoxia rat model. Pulm Circ. 2020;10:2045894020929147. [CrossRef]
- Lawrie A, Hameed AG, Chamberlain J, Arnold N, Kennerley A, Hopkinson K, Pickworth J, Kiely DG, Crossman DC, Francis SE. Paigen diet-fed apolipoprotein E knockout mice develop severe pulmonary hypertension in an interleukin-1-dependent manner. Am J Pathol. 2011;179:1693-1705. [CrossRef]
- Brittain EL, Talati M, Fortune N, Agrawal V, Meoli DF, West J, Hemnes AR. Adverse physiologic effects of Western diet on right ventricular structure and function: role of lipid accumulation and metabolic therapy. Pulm Circ. 2019;9:2045894018817741. [CrossRef]
- Pakhomov NV, Kostyunina DS, Macori G, Dillon E, Brady T, Sundaramoorthy G, Connolly C, Blanco A, Fanning S, Brennan L, et al. High-Soluble-Fiber Diet Attenuates Hypoxia-Induced Vascular Remodeling and the Development of Hypoxic Pulmonary Hypertension. Hypertension. 2023;80:2372-2385. [CrossRef]
- Mamazhakypov A, Weiß A, Zukunft S, Sydykov A, Kojonazarov B, Wilhelm J, Vroom C, Petrovic A, Kosanovic D, Weissmann N, et al. Effects of macitentan and tadalafil monotherapy or their combination on the right ventricle and plasma metabolites in pulmonary hypertensive rats. Pulm Circ. 2020;10:2045894020947283. [CrossRef]
- Krautkramer KA, Fan J, Bäckhed F. Gut microbial metabolites as multi-kingdom intermediates. Nat Rev Microbiol. 2021;19:77-94. [CrossRef]
- Karoor V, Strassheim D, Sullivan T, Verin A, Umapathy NS, Dempsey EC, Frank DN, Stenmark KR, Gerasimovskaya E. The Short-Chain Fatty Acid Butyrate Attenuates Pulmonary Vascular Remodeling and Inflammation in Hypoxia-Induced Pulmonary Hypertension. Int J Mol Sci. 2021;22. [CrossRef]
- 2022 Annual World Congress of the Pulmonary Vascular Research Institute. In: Pulm Circ. © 2022 The Authors. Pulmonary Circulation published by Wiley Periodicals LLC on behalf of the Pulmonary Vascular Research Institute.; 2022.
- Pulgarin A, Alabdallat M, Methe B, Morris A, Al Ghouleh I. Abstract 18631: Mechanistic Insight on the Protective Role of Microbiome-Derived Butyrate in Pulmonary Hypertension. Circulation. 2023;148:A18631-A18631. doi: doi:10.1161/circ.148.suppl_1.18631.
- Huang Y, Lin F, Tang R, Bao C, Zhou Q, Ye K, Shen Y, Liu C, Hong C, Yang K, et al. Gut Microbial Metabolite Trimethylamine. Am J Respir Cell Mol Biol. 2022;66:452-460. [CrossRef]
- Yang Y, Zeng Q, Gao J, Yang B, Zhou J, Li K, Li L, Wang A, Li X, Liu Z, et al. High-circulating gut microbiota-dependent metabolite trimethylamine N-oxide is associated with poor prognosis in pulmonary arterial hypertension. Eur Heart J Open. 2022;2:oeac021. [CrossRef]
- Videja M, Vilskersts R, Korzh S, Cirule H, Sevostjanovs E, Dambrova M, Makrecka-Kuka M. Microbiota-Derived Metabolite Trimethylamine N-Oxide Protects Mitochondrial Energy Metabolism and Cardiac Functionality in a Rat Model of Right Ventricle Heart Failure. Front Cell Dev Biol. 2020;8:622741. [CrossRef]
- Levy M, Thaiss CA, Elinav E. Metabolites: messengers between the microbiota and the immune system. Genes Dev. 2016;30:1589-1597. [CrossRef]
- Roager HM, Licht TR. Microbial tryptophan catabolites in health and disease. Nat Commun. 2018;9:3294. [CrossRef]
- Oliveira AC, Yang T, Li J, Sharma RK, Karas MK, Bryant AJ, de Kloet AD, Krause EG, Joe B, Richards EM, et al. Fecal matter transplant from Ace2 overexpressing mice counteracts chronic hypoxia-induced pulmonary hypertension. Pulm Circ. 2022;12:e12015. [CrossRef]
- Wedgwood S, Warford C, Agvatisiri SR, Thai PN, Chiamvimonvat N, Kalanetra KM, Lakshminrusimha S, Steinhorn RH, Mills DA, Underwood MA. The developing gut-lung axis: postnatal growth restriction, intestinal dysbiosis, and pulmonary hypertension in a rodent model. Pediatr Res. 2020;87:472-479. [CrossRef]
- Prisco SZ, Blake M, Kazmirczak F, Moon R, Vogel N, Moutsoglou D, Thenappan T, Prins KW. Restructures the Micro/Mycobiome to Combat Glycoprotein-130 Associated Microtubule Remodeling and Right Ventricular Dysfunction in Pulmonary Arterial Hypertension. bioRxiv. 2024. [CrossRef]
- Alcayaga-Miranda F, Cuenca J, Khoury M. Antimicrobial Activity of Mesenchymal Stem Cells: Current Status and New Perspectives of Antimicrobial Peptide-Based Therapies. Front Immunol. 2017;8:339. [CrossRef]
- Yang F, Ni B, Liu Q, He F, Li L, Zhong X, Zheng X, Lu J, Chen X, Lin H, et al. Human umbilical cord-derived mesenchymal stem cells ameliorate experimental colitis by normalizing the gut microbiota. Stem Cell Res Ther. 2022;13:475. [CrossRef]
- Ocansey DKW, Wang L, Wang J, Yan Y, Qian H, Zhang X, Xu W, Mao F. Mesenchymal stem cell-gut microbiota interaction in the repair of inflammatory bowel disease: an enhanced therapeutic effect. Clin Transl Med. 2019;8:31. [CrossRef]
- Liu A, Li C, Wang C, Liang X, Zhang X. Impact of Mesenchymal Stem Cells on the Gut Microbiota and Microbiota Associated Functions in Inflammatory Bowel Disease: A Systematic Review of Preclinical Evidence on Animal Models. Curr Stem Cell Res Ther. 2024;19:981-992. [CrossRef]
- Kim KC, Lee JC, Lee H, Cho MS, Choi SJ, Hong YM. Changes in Caspase-3, B Cell Leukemia/Lymphoma-2, Interleukin-6, Tumor Necrosis Factor-α and Vascular Endothelial Growth Factor Gene Expression after Human Umbilical Cord Blood Derived Mesenchymal Stem Cells Transfusion in Pulmonary Hypertension Rat Models. Korean Circ J. 2016;46:79-92. [CrossRef]
- Muhammad SA, Abbas AY, Saidu Y, Fakurazi S, Bilbis LS. Therapeutic efficacy of mesenchymal stromal cells and secretome in pulmonary arterial hypertension: A systematic review and meta-analysis. Biochimie. 2020;168:156-168. [CrossRef]
- Abudukeremu A, Aikemu A, Yang T, Fang L, Aihemaitituoheti A, Zhang Y, Shanahaiti D, Nijiati Y. Effects of the ACE2-Ang-(1-7)-Mas axis on gut flora diversity and intestinal metabolites in SuHx mice. Front Microbiol. 2024;15:1412502. [CrossRef]
- Kim S, Rigatto K, Gazzana MB, Knorst MM, Richards EM, Pepine CJ, Raizada MK. Altered Gut Microbiome Profile in Patients With Pulmonary Arterial Hypertension. Hypertension. 2020;75:1063-1071. [CrossRef]
- Jose A, Apewokin S, Hussein WE, Ollberding NJ, Elwing JM, Haslam DB. A unique gut microbiota signature in pulmonary arterial hypertension: A pilot study. Pulm Circ. 2022;12:e12051. [CrossRef]
- Moutsoglou DM, Tatah J, Prisco SZ, Prins KW, Staley C, Lopez S, Blake M, Teigen L, Kazmirczak F, Weir EK, et al. Pulmonary Arterial Hypertension Patients Have a Proinflammatory Gut Microbiome and Altered Circulating Microbial Metabolites. Am J Respir Crit Care Med. 2023;207:740-756. [CrossRef]
- Dong W, Ma L, Huang Q, Yang X, Mei Z, Kong M, Sun Z, Zhang Z, Li J, Zou J, et al. Gut microbiome alterations in pulmonary hypertension in highlanders and lowlanders. ERJ Open Res. 2023;9. [CrossRef]
- Kurilshikov A, Medina-Gomez C, Bacigalupe R, Radjabzadeh D, Wang J, Demirkan A, Le Roy CI, Raygoza Garay JA, Finnicum CT, Liu X, et al. Large-scale association analyses identify host factors influencing human gut microbiome composition. Nat Genet. 2021;53:156-165. [CrossRef]
- Li X, Tan JS, Xu J, Zhao Z, Zhao Q, Zhang Y, Duan A, Huang Z, Zhang S, Gao L, et al. Causal impact of gut microbiota and associated metabolites on pulmonary arterial hypertension: a bidirectional Mendelian randomization study. BMC Pulm Med. 2024;24:185. [CrossRef]
- Su L, Wang X, Lin Y, Zhang Y, Yao D, Pan T, Huang X. Exploring the Causal Relationship Between Gut Microbiota and Pulmonary Artery Hypertension: Insights From Mendelian Randomization. J Am Heart Assoc. 2025:e038150. [CrossRef]
- Zhang C, Zhang T, Lu W, Duan X, Luo X, Liu S, Chen Y, Li Y, Chen J, Liao J, et al. Altered Airway Microbiota Composition in Patients With Pulmonary Hypertension. Hypertension. 2020;76:1589-1599. [CrossRef]
- Zhang C, Zhang T, Xing Y, Lu W, Chen J, Luo X, Wu X, Liu S, Chen L, Zhang Z, et al. Airway delivery of Streptococcus salivarius is sufficient to induce experimental pulmonary hypertension in rats. Br J Pharmacol. 2023;180:2102-2119. [CrossRef]
- Zhang C, Zhong B, Jiang Q, Lu W, Wu H, Xing Y, Wu X, Zhang Z, Zheng Y, Li P, et al. Distinct airway mycobiome signature in patients with pulmonary hypertension and subgroups. BMC Med. 2025;23:148. [CrossRef]
- Salminen S, Collado MC, Endo A, Hill C, Lebeer S, Quigley EMM, Sanders ME, Shamir R, Swann JR, Szajewska H, et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat Rev Gastroenterol Hepatol. 2021;18:649-667. [CrossRef]
- Zimmermann P. The immunological interplay between vaccination and the intestinal microbiota. NPJ Vaccines. 2023;8:24. [CrossRef]
- Buys R, Avila A, Cornelissen VA. Exercise training improves physical fitness in patients with pulmonary arterial hypertension: a systematic review and meta-analysis of controlled trials. BMC Pulm Med. 2015;15:40. [CrossRef]
- Castillo-Galán S, Parra V, Cuenca J. Unraveling the pathogenesis of viral-induced pulmonary arterial hypertension: Possible new therapeutic avenues with mesenchymal stromal cells and their derivatives. Biochim Biophys Acta Mol Basis Dis. 2025;1871:167519. [CrossRef]
- Moutsoglou DM. 2021 American Thoracic Society BEAR Cage Winning Proposal: Microbiome Transplant in Pulmonary Arterial Hypertension. Am J Respir Crit Care Med. 2022;205:13-16. [CrossRef]
- Ahn J, Hayes RB. Environmental Influences on the Human Microbiome and Implications for Noncommunicable Disease. Annu Rev Public Health. 2021;42:277-292. [CrossRef]
- Zhong Y, Catheline D, Houeijeh A, Sharma D, Du L, Besengez C, Deruelle P, Legrand P, Storme L. Maternal omega-3 PUFA supplementation prevents hyperoxia-induced pulmonary hypertension in the offspring. Am J Physiol Lung Cell Mol Physiol. 2018;315:L116-L132. [CrossRef]
- Marinho Y, Villarreal ES, Loya O, Oliveira SD. Mechanisms of lung endothelial cell injury and survival in pulmonary arterial hypertension. Am J Physiol Lung Cell Mol Physiol. 2024;327:L972-L983. [CrossRef]
- Oliveira SD, Almodóvar S, Butrous G, De Jesus Perez V, Fabro A, Graham BB, Mocumbi A, Nyasulu PS, Tura-Ceide O, Oliveira RKF, et al. Infection and pulmonary vascular diseases consortium: United against a global health challenge. Pulm Circ. 2024;14:e70003. [CrossRef]
- Cool CD, Voelkel NF, Bull T. Viral infection and pulmonary hypertension: is there an association? Expert Rev Respir Med. 2011;5:207-216. [CrossRef]
- Pullamsetti SS, Savai R, Janssen W, Dahal BK, Seeger W, Grimminger F, Ghofrani HA, Weissmann N, Schermuly RT. Inflammation, immunological reaction and role of infection in pulmonary hypertension. Clin Microbiol Infect. 2011;17:7-14. [CrossRef]
- Mizutani T, Ishizaka A, Koga M, Tsutsumi T, Yotsuyanagi H. Role of Microbiota in Viral Infections and Pathological Progression. Viruses. 2022;14. [CrossRef]
- Alves JL, Gavilanes F, Jardim C, Fernandes CJCD, Morinaga LTK, Dias B, Hoette S, Humbert M, Souza R. Pulmonary arterial hypertension in the southern hemisphere: results from a registry of incident Brazilian cases. Chest. 2015;147:495-501. [CrossRef]
- Knafl D, Gerges C, King CH, Humbert M, Bustinduy AL. Schistosomiasis-associated pulmonary arterial hypertension: a systematic review. Eur Respir Rev. 2020;29. [CrossRef]
- Marinho Y, Villarreal ES, Aboagye SY, Williams DL, Sun J, Silva CLM, Lutz SE, Oliveira SD. Schistosomiasis-associated pulmonary hypertension unveils disrupted murine gut-lung microbiome and reduced endoprotective Caveolin-1/BMPR2 expression. Front Immunol. 2023;14:1254762. [CrossRef]
- Almodovar S, Cicalini S, Petrosillo N, Flores SC. Pulmonary hypertension associated with HIV infection: pulmonary vascular disease: the global perspective. Chest. 2010;137:6S-12S. [CrossRef]
- Rocafort M, Gootenberg DB, Luévano JM, Paer JM, Hayward MR, Bramante JT, Ghebremichael MS, Xu J, Rogers ZH, Munoz AR, et al. HIV-associated gut microbial alterations are dependent on host and geographic context. Nat Commun. 2024;15:1055. [CrossRef]
Figure 1.
Factors influencing intestinal microbiome composition.
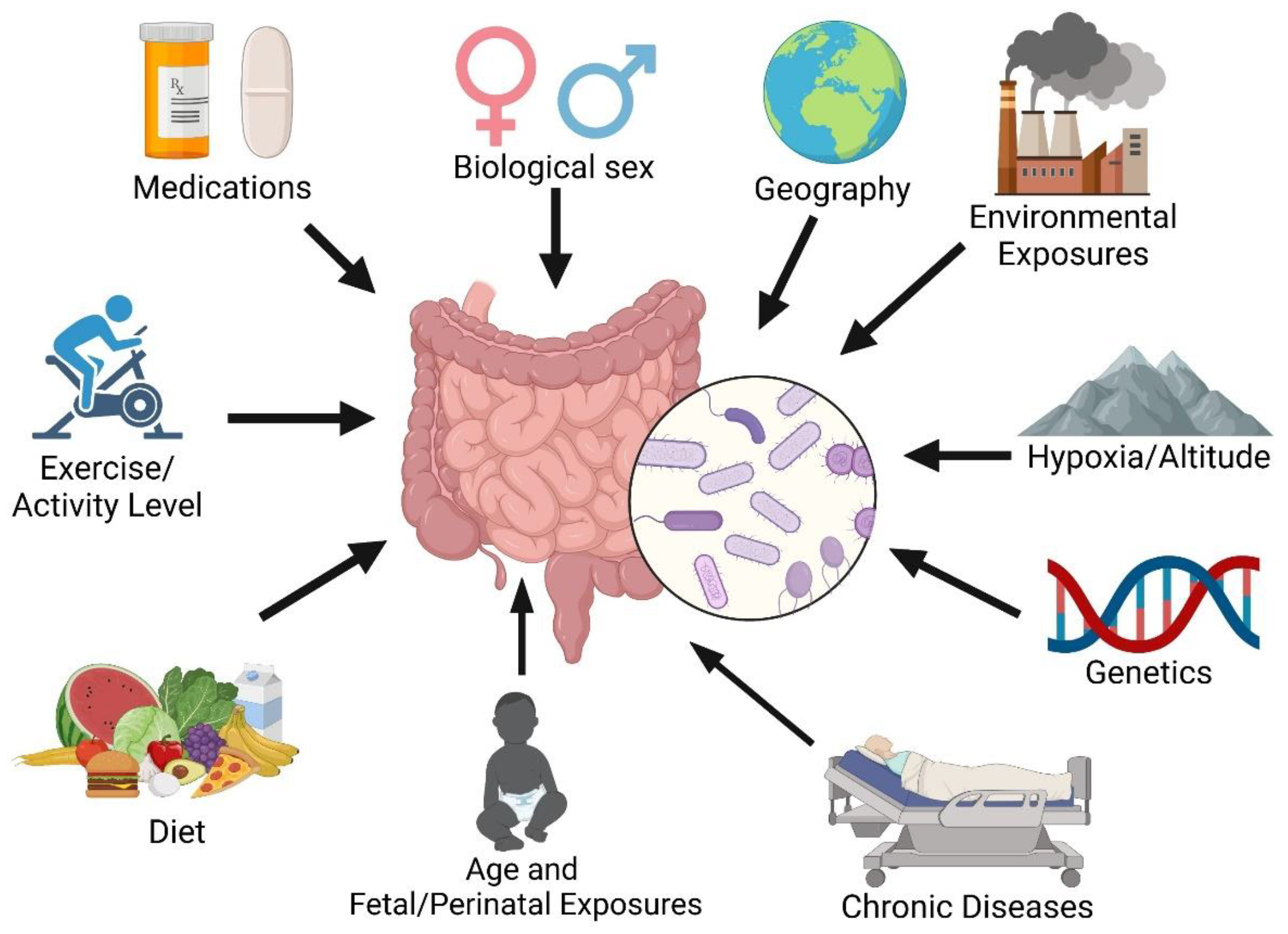
Figure 2.
Potential therapeutic approaches to restructure the gut microbiome.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



