Submitted:
14 March 2025
Posted:
17 March 2025
You are already at the latest version
Abstract
This paper considers whether forest volatile organic compounds (VOCs) and/or their metabolites have health-promoting activities, and whether this is through ‘mindfulness’ practices and/or activation of health-promoting physiological processes. The VOCs considered are the monoterpenes D-limonene, a-pinene, D3-carene and 1,8-cineole. Clearly, some of the health benefits of forest bathing come from ‘mindfulness’ practice that promote parasympathetic activities. There is also clear evidence that the 4 monoterpenes at high enough concentrations can activate physiological processes that lead to better health outcomes, particularly with decreases in anxiety and decreases in inflammation. These processes appear to involve activation of the A2A receptor and down-regulating activation of Nuclear Factor kappa B. The question is whether the monoterpenes can do so at the concentrations encountered in a forest setting. There is some research that indicates that forest atmospheric concentrations of monoterpenes can activate physiological processes that promote health but more research is required to definitively establish this. This review ends with suggestions on how to unequivocally answer the question whether the 4 monoterpenes considered at concentrations found in a forest, or their metabolites, can activate physiological processes that promote health, in particular physiological processes that decrease anxiety and inflammatory conditions and promote better endothelial function.
Keywords:
A2A receptor
; Anxiety
; Asthma
; D3-Carene
; 1
; 8-Cineole: Forest volatile organic compounds
; Health
; D-Limonene
; Monoterpenes
; NF Kappa B
; Nrf2
; a-Pinene
1. Introduction
Forest bathing for health has become popular in recent years. This paper considers the question of whether forest volatile organic compounds (VOCs), particularly monoterpenes, can activate physiological processes that promote health.
‘Forest Bathing’ for health has a long history. For example, in year 77 of the Common Era Pliny the Elder published the first ten volumes of his Natural History series and wrote in Chapter 19 of Volume V [1]: “It is a well-known fact that forests planted solely with trees from which pitch and resin are extracted, are remarkably beneficial for patients suffering from phthisis1, or who are unable to recover their strength after a long illness: indeed it is said, that in such cases to breathe the air of localities thus planted, is more beneficial even than to take a voyage to Egypt, or to go on a summer’s journey to the mountains to drink the milk there, impregnated with the perfumes of plants.”
The practice of Shinrin-Yoku (forest bathing) was made popular in Japan during the 1980s [2] and now many physicians and other healthcare providers throughout the world prescribe encounters with nature. Forest bathing does not necessarily involve exercise but is simply being in the presence of a forest for some time, although some studies on the effect of forests on health are dynamic involving activities such as walking. Today there are formal nature prescription programs in North America where healthcare providers prescribe visits to green spaces [3]. One example is the PaRx prescription program developed by Dr Melissa Lem in combination with the British Columbia Parks Foundation that allows registered healthcare providers to prescribe free access to Provincial and National parks. This program was approved by the Canadian Medical Association and has now spread across Canada.
Potential Mechanisms Involved in Forest Bathing for Health
There are many primary research articles that demonstrate there are health benefits to static forest bathing such as increasing the sense of well-being, e.g., [4] and decreasing depression, anxiety as well as increasing the anti-oxidant status of blood in a chronic stroke population e.g. [5]. There are a number of reports that indicate forest bathing by urban children with asthma improves symptoms, e.g., [6,7]. Here children are taken out of an urban environment to a forest environment. We do not know whether such improvements in asthma symptoms are due to leaving a more polluted air environment to going to a less polluted air environment, ‘mindfulness’ aspects of the forest environment, or due, in part, to the presence of monoterpenes. One study showed a positive correlation between lung function and total monoterpene exposure [7], although daily concentrations of monoterpenes measured were low and ranged from 38 to 62 ng/m3. Both static forest bathing as well as dynamic forest bathing show that a forest environment improves cardiovascular function (decreased systolic blood pressure and increased arterial compliance) while an urban environment does not, e.g., [8,9,10]. There are many reviews on this topic, e.g., [11,12].
Does forest bathing promote health through practices associated with mindfulness or do the volatile molecules emitted by the forest activate physiological processes that promote health? Forest bathing exposes one to 2 situations that can impact health through promotion of the activation of the parasympathetic nervous system: i) a calming environment that promotes activation of the parasympathetic nervous system, and ii) an atmosphere containing volatile forest molecules. The pleasant scent of many of volatile forest molecules in turn can promote activation of the parasympathetic nervous system just by delightful odours, for example, Christmas wreathes, or bring back pleasant memories. These effects may be mediated through ‘mindfulness’ practices. There is clear evidence that practicing mindfulness promotes activation of the parasympathetic nervous system and decreases blood pressure in hypertensive patients [13], increases antioxidants in blood [14] and decreases depression and anxiety [15]. There is also evidence that simply viewing forest imagery has some positive physiological and psychological effects when compared to viewing urban imagery [16].
Can one simply substitute images of forests for bathing in an actual forest? Or do VOCs activate physiological processes that promote health beyond ‘mindfulness’ practices? Can forest VOCs appear in the blood of forest bathers in sufficient concentrations that may have physiological effects? Inhalation of VOCs is an efficient way of delivering these compounds to blood. As an example, Chen and colleagues demonstrated that inhaling D-limonene by mice was 65 times more efficient in delivering the compound to blood than ingesting the same quantity [17]. In a natural setting do VOCs enter the blood stream in concentrations that may have physiological effects? For some of these molecules the answer is yes. After a 60 min walk in the Tsubetsu forest of Eastern Hokkaido, comprised mainly of spruce and fir trees, blood levels of participants’ a-pinene, the dominant monoterpene in this forest’s atmosphere, increased from 2.6±1.7 to 19.4±2.1 nM [18]. This is not that surprising since the lungs have an enormous surface area available for molecular uptake and a-pinene, like other hydrocarbon monoterpenes, is lipophilic [19] and should readily enter the blood whose lipophilic molecules should act as monoterpene sinks. Could a concentration of 19.4 nM be sufficient to activate any physiological process? Perhaps. In my own laboratory we have seen cell physiological responses to sulforaphane at 10 nM with near maximal responses seen at 50 nM [20].
Not only do monoterpenes readily enter the blood but they will readily move from blood into fatty tissues. A study in mice exposed to monoterpene vapours have shown that within 30-90 min of exposure brain levels reached over 90 µg/g for a-pinene, over 50 µg/g for D-limonene and over 70 µg/g for 1,8-cineole [21]. In these experiments 50 µL of the VOC was applied to a filter paper placed into a 5 L sealed cage where each mouse was placed. For example, with a-pinene, which has a density of 0.858 g/mL and had a 95% purity, 40.75 mg of a-pinene would have been placed on the filter. Unfortunately, the concentration of monoterpenes in the cage was not measured; however, if all, for example, the a-pinene volatilized, then the concentration would have been 8.15 mg/L, many orders of magnitude higher than would be found in a forest, but these experiments do show the lipophilicity of the monoterpenes and that fats can act as stores for monoterpenes.
Evidence That Exposure to Forest Monoterpenes Can Impact Physiological Processes
Is there any evidence that VOCs alone may positively affect health? That forest VOCs may directly affect physiological processes directly is demonstrated by experiments by Ahn and colleagues [22,23]. Mice were administered lipopolysaccharide (LPS) intraperitoneally or into the lungs. LPS is present in the membranes of gram-negative bacteria and initiates a strong systemic inflammatory response by binding to and activating the Toll-like Receptor-4 in cell membranes [24]. Following administration of LPS, mice were divided into the four groups: i) controls with no further treatment, ii) a group treated with dexamethasone, a potent anti-inflammatory steroid, iii) a group exposed to a panel of Korean pine in their cages, and iv) a group exposed to a panel of Japanese larch (or Hinoki cyprus) in their cages. The wood panels were shown to emit a number of VOCs with hexanal and the monoterpenes b-phellandrene and a-pinene being dominant (96-280 µg/m3 in the cage atmosphere). As expected, administration of dexamethasone greatly reduced a variety of inflammatory markers. Exposure to volatiles emitted by the pine, larch and cyprus panels also significantly decreased many of the inflammatory markers examined such as interleukin-1b (IL1b), IL13, tumour necrosis factor-a (TNFa) and nuclear factor kappa B (NFkB) mRNA. Although the concentrations of VOCs were high (an order or two of magnitude greater than would be encountered in a forest) in these experiments, it strongly suggests that some of the forest VOCs act directly on physiological processes that ameliorate systemic inflammation.
In conclusion, forest bathing has three potential mechanisms of action: one mediated by processes activated by ‘mindfulness’, a second by going from a more polluted environment to a less polluted environment and a third mediated by direct actions of VOCs on physiological processes.
2. What Are the Volatile Organic Compound Molecules in Northern Temperate Forests?
It is estimated that the earth’s vegetation releases about a billion tonnes of VOC carbon annually into the atmosphere [25,26]. More than half of the emissions is isoprene (~634 megatonnes), with monoterpenes at ~89 megatonnes and sesquiterpenes at ~36 megatonnes forming most of the rest. The basic building unit of the terpenes is isoprene. Isoprene has the formula of C5H8, while that of monoterpene is (C5H8)2 and sesquiterpene is (C5H8)3 – see Table 1.
The majority of the terpenes are emitted by living plants but a fraction also comes from decaying plant matter [27]. There is great variability amongst species in individual terpene chemical species emissions [28]. Even within a genus, terpene emissions can vary immensely, for example the emission rate (ng/hr/gram branch) of D3-carene for P. albicaulis is reported to be 651 and for P. longaeva it is 9.5 while for limonene it is 321 vs 12.2, respectively [29]. Coniferous forests also emit more monoterpenes and less isoprene compared to deciduous forests. This is demonstrated in a study examining the VOCs in the August air of the Tharandt forest in Germany where the dominant tree species were European spruce (72%), Scots pine (15%) European larch (10%) and 3% deciduous trees and in the Hohe Holz forest in Germany where the dominant tree species were European beech, sessile oak, Silver birch, common hornbeam, and a few newly planted European spruce and European larch [30]. The coniferous forest had about 1.8 µg/m3 of maximum isoprene concentrations forming 12% of total terpenes while the deciduous forest had a maximum isoprene level of 12 µg/m3 forming 93% of total terpenes The dominant monoterpenes in the coniferous forest was a-pinene at 6 µg/m3 while the dominant monoterpene in the deciduous forest was sabinene at 0.65 µg/m3.
Most of the studies on VOCs in forests examine terpene emissions. Because of the presence of unsaturated bonds in many terpenes, they are susceptible to oxidation and their half-lives can vary [31]; hence, it is important to analyze the composition of the atmosphere and not just pay attention to terpene synthesis and emission.
2.1. Isoprene
Although there is a large emission of isoprene by vegetation, isoprene is a very reactive chemical and its lifespan in the atmosphere is only about one hr [32]; hence, isoprene levels tend to be relatively low in the atmosphere of a forest. For example in a mixed temperate forest in Britain isoprene levels at a temperature of 25 °C was 2 ppb [33] – that is 5.57 µg/m3 at 25° C using the formula: µg/m3 = molecular weight x concentration (ppb) ÷ 24.45 (i.e., the volume of one mole in litres at standard temperature and pressure), or 0.08 nmoles/L of air. In a mixed temperate forest in Estonia, the maximum level of isoprene was 5.3 µg/m3 of air reached in summer but could be as low as 0.2 µg/m3 in winter [34].
Isoprene is also a by-product of cholesterol catabolism [35]; hence, even in the absence of isoprene in the atmosphere, isoprene is normally present in human blood and exhaled via the lungs: such human-derived blood plasma levels are in a similar concentration range as found in the atmosphere of forests, ranging from 15 to 70 nmoles per litre of blood [36]; hence, atmospheric isoprene likely will have little influence on blood levels.
2.2. Monoterpenes
There are a number of studies that have examined monoterpene concentrations in the atmosphere of forests. There are a range of concentrations of specific monoterpenes reported. Some of the differences in concentrations of monoterpenes may be due to differing heights that the samples were taken since there is increased concentrations of monoterpenes the higher into the canopy that one samples [37] while others may be due to the time of year or time of day when collected since there are seasonal [38] and diurnal changes in concentrations [39]. Some differences could be due to the amount of light or temperature [40], or possibly calculation errors.
One should keep in mind that depending upon analytical technique used, only a subset of the monoterpenes will be identified and quantitated; hence, there are differences amongst the studies on the species of monoterpenes analyzed and reported. There are also differences in reporting concentrations. A range in concentrations of monoterpenes have been reported, for example in a hemiboreal forest in Estonia, a-pinene atmospheric concentrations were reported to be 23 µg/m3 in August at ground level [34], 6 µg/m3 in August in a pine forest in Germany at 13 m above the ground [30], up to 6.5 µg/m3 above the canopy in a boreal coniferous forest in Finland [41] and up to 54 µg/m3 a few centimetres above the ground in a Scandinavian pine forest [42].
The most abundant monoterpenes in Korean cyprus and pine forests were a-pinene, b-pinene, D-limonene, myrcene and sabinene [43], with similar monoterpenes found in a beech forest, albeit at lower concentrations. In a study of a German pine forest the most abundant monoterpenes were a-pinene, b-pinene, D-limonene, sabinene, D3-carene and camphene whereas in a deciduous forest they were the same but a lower concentrations [30]. In a study of a Finnish boreal forest the major monoterpenes were a-pinene, camphene, b-pinene, myrcene, D3-carene, p-cymene, D-limonene, 1,8-cineol and terpinolene [44]. There are differences between summer and winter atmospheric concentrations with a-pinene, D3-carene and b-pinene dominating in summer while camphene, p-cymene and a-pinene dominating in winter. A Scandinavian pine forest air sampled at night had the following monoterpenes: a-pinene, b-phellandrene, camphene, b-pinene, D-limonene, D3-carene and myrcene [42].
Reported levels of sesquiterpenes were much lower than the common monoterpenes measured; hence, this manuscript will not deal with sesquiterpenes.
There is an enormous literature on the effect of monoterpenes on a variety of disease conditions. For example, there are studies showing that D-limonene ameliorates disease in animal models of stroke [45], Parkinson’s disease [46], cardiac infarction [47], Type 2 diabetes [48], peritoneal adhesions [49], inflammatory bowel syndrome [50], to name a few. In this manuscript only 4 monoterpenes, D-limonene, a-pinene, D3-carene and 1,8-cineole (Table 1), and two health conditions (respiratory inflammatory problems and anxiety) will be considered.
3. Pharmacokinetics and Metabolism
We should keep in mind that monoterpenes are metabolized by the various tissues and organs in the body and any therapeutic effect may be mediated by a metabolite rather than the parent monoterpene. This makes understanding the mechanisms underlying any therapeutic effect complex.
The majority of studies examining monoterpene metabolism are following administration of the monoterpene either intraperitoneally or perorally; hence, the intestine, liver and kidneys are primarily involved in the monoterpene metabolism. Lung tissue also contains xenobiotic metabolizing enzymes [51]; thus, it is very possible that the metabolites that are relevant for the therapeutic effects may be missed in the metabolic studies where the monoterpene is administered intraperitoneally, perorally or dermally. Effects via inhalation may be quite different from effects where the monoterpene is administered via other routes. For example, Heuberger and colleagues found different physiological and psychological responses whether the monoterpenes 1,8-cineole and linalool were applied dermally or via inhalation [52].
We should also keep in mind that the pharmacokinetics of inhaled volatile lipophilic compounds such as monoterpenes is complex since blood concentrations are affected by uptake into the blood, movement into lipophilic blood molecules such as low density lipoprotein, movement into fatty tissue such as brain or adipose tissue, metabolism by the liver, kidneys and lungs and exhalation of the parent compound by the lungs [53]. Following the end of exposure to a VOC such as a-pinene [53], there is a rapid (5-10 min) drop in the blood concentration followed by a slower (hours) decline. The rapid drop involves the metabolism of the compound in the aqueous phase of the blood as well as exhalation of the compound while the slower decline is due to movement of the compound from the fat molecules and fat tissue into the blood. All inhalation studies examining pharmacokinetics in humans used very high concentrations of monoterpenes, many of orders higher than found in forests.
Finally, many of the monoterpenes emitted by forests are also present in many of our food items [54] and this must be kept in mind when examining plasma levels.
D-Limonene: D-Limonene is by far the dominant monoterpene at 1.5 µg/L (i.e., 11 nM) found in several studies of blood levels of monoterpenes [55,56] – in these studies it is likely that the plasma D-limonene had a dietary origin. D-Limonene generally enters the blood either through inhalation or by ingestion. By far the majority of studies involving monoterpenes have been on D-limonene.
Inhalation of D-limonene at 10, 225 and 450 mg/m3 in human volunteers resulted in 65% of the inhaled D-limonene transferred from air to blood [57]. It should be noted that Kohlert and colleagues question whether the difference between the concentration in inhaled air versus exhaled air can be considered the amount of monoterpene transferred to blood [58]. Elimination from the blood following inhalation of the higher doses was in 3 phases with half-lives of 3, 33 and 750 min, with the slow elimination phase due to movement from adipose tissue into the blood. Following ingestion of 9.3 mg (i.e., 68 µmoles) by human volunteers there was no detectable D-limonene in the plasma at 1 hr [59], with the major metabolites detected being limonene-8,9-diol, limonene-1,2-diol followed by perillic acid. Other metabolites that have been reported in various studies include: dihydroperillic acid; dihydroperillic acid, methyl ester; perillic acid, methyl ester; perrilyl alcohol; and carveol [60,61,62]. It seems very possible that many of the therapeutic effects attributed to D-limonene may be due to one or more of its metabolites, and possibly explains why such high concentrations of D-limonene are required to observe an effect in cultured cells. Which of the metabolites may be responsible for the therapeutic effects of inhaled D-limonene is not clear, and especially so since we do not know which are the dominant metabolites formed from inhaled D-limonene. Perillic acid seems to be a compound of interest since it appears to be a Michael electron acceptor. Michael electron acceptors activate the Nrf2 signalling pathway [63,64] – see section IV: Discussion.
a-Pinene: Pharmacokinetics of inhalation of a-pinene was similar to that of D-limonene [65]. Capillary blood concentrations of a-pinene increased rapidly for the first half hour and then more slowly to reach 9 µM and 19 µM at two hr for the 225 µg/L and 450 µg/L air concentrations, respectively. At the 10 µg/L air concentration, the blood concentration stabilized at about 30 min at 0.5 µM, likely because metabolism equilibrated with transfer of a-pinene from air to blood. It was estimated that 60% of a-pinene inhaled transferred to blood. After the 2-hr exposure to the lipophilic a-pinene, elimination from the blood occurred in 3 phases, a rapid phase with a half-life of 4.8 minutes that involved, exhalation of the non-lipid-bound parent compound and metabolism by the liver, kidneys and lungs and then slower phase clearances (38 min and 695 min half-lives) as the parent compound moved from lipid molecules and adipose tissues into the blood.
Inhalation studies in rodents also looked at the accumulation of a-pinene in mammary glands, i.e., a fat rich tissue and observed an accumulation within mammary glands that accounted for the third slow phase of a-pinene elimination from the blood [66]. Mice were exposed to either 50 ppm (279 µg/m3) or double that concentration to a-pinene for 6 hr/day for seven consecutive days. Inhaled doses were calculated to be 70 and 140 mg/kg/day at the two atmospheric concentrations respectively. Assuming the transfer from air to blood is the same as in humans [65], with 60% absorption, 42 and 84 mg/kg transferred to the blood each day. a-Pinene accumulated in female mammary gland fat at 1.18 and 2.5 mg/g lipid at the two atmospheric concentrations. Even 24 hr following the 7-day exposure to a-pinene there were still 10 and 20 ng a-pinene/mL, respectively, in the blood due to the slow release from the fat stores., i.e., 73 and 147 pM.
Blood samples were obtained from human subjects before and at 1 to 5 hr after consumption of 9 mg of a-pinene [67]. Within 1 hr of consumption there was no detectable a-pinene in the blood. Four major metabolites were detected in the blood: myrtenol, cis- and trans-verbenol, myrtenic acid and possibly a hydroxylated form of myrtenic acid. Myrtenic acid has the characteristics of a Michael electron acceptor.
A study in mice observed the presence of cis- and trans-verbenol as well as myrtenol in urine following inhalation of air containing 1 ppm (5.6 mg/m3) a-pinene for a period of six hr [68].
D3-Carene: Pharmacokinetics of inhaled D3-carene was performed where human subjects were exposed to D3-carene at 10, 225 and 450 µg/L for 2 hr [69]. Maximum blood concentrations reached were ~1, 12 and 26 µmol/L. It was estimated that 70% of the D3-carene inhaled transferred to blood. The typical 3 phase elimination from blood was observed, an initial rapid phase with a half-life of 4.5 min, an intermediate phase of 35 min and a final slow phase of 30 hr. Similar results were seen by Kohlert and colleagues [58].
Following oral administration of 10 mg D3-carene the main metabolites found in urine were D3-caren-10-carboxylic acid (chaminic acid), D3-caren-9,10-dicarboxylic acid, likely a saturated version of D3-caren-10-carboxylic acid (dihydrochaminic acid), D3-caren-3,4-diol-9-al, and D3-caren-3,4,9-triol [70]. It was reported that at least one of the metabolites has a half-life of 3 hr. Of interest is that D3-caren-10-carboxylic acid, D3-caren-9,10-dicarboxylic acid and D3-caren-3,4-diol-9-al appear to be reactive molecules. Studies using human liver microsomes showed that D3-caren-10-ol and carene and D3-carene epoxide can also be formed from D3-carene [71]. D3-caren-10-carboxylic acid has the characteristic of a Michael electron acceptor.
1,8-Cineole: 1,8-Cineole is an oxidized monoterpene. In a study where human subjects inhaled air passing over 4 mL of 1,8-cineole for 20 min 1,8-cineole was rapidly transported from air to blood with plasma levels peaking at 14-19 min [72]. Peak plasma levels ranged from 3 to 7.4 nM, likely due to differing rates of metabolism amongst the individuals. The elimination of 1,8-cineole from plasma was biphasic with the first phase with a half-life of 6.68 min and the second with a half-life of 104.6 min. There were differences between the male and female subjects likely due to varying levels of fat tissue that would have absorbed 1,8-cineole. In another inhalation study where the blood analytical approach was designed to minimize volatilization of 1,8-cineole demonstrated that plasma levels of 1,8-cineole after 45 min reached anywhere from 1 to 3 µM [73]. The concentration of 1,8-cineole in the air breathed in was not determined in these studies.
Following consumption of 1 mg 1.8-cineole it was shown that the parent compound was rapidly metabolized resulting in the formation of the following metabolites found in human plasma: 2-hydroxy-1,8-cineole, 3-hydroxy-1,8-cineole, 7-hydroxy-1,8-cineole and 9-hydroxy-1,8-cineole [74]. All compounds peaked at 45 min with 2-hydroxy-1,8-cineole being the dominant metabolite at 86 nM followed by 9-hydroxy-1,8-cineole at 33 nM, with the parent compound peaking at 19 nM, indicating rapid metabolism. In a study of human milk, where participants consumed 100 mg 1,8-cineole [75], ten metabolites of 1,8-cineole were identified; a2-hydroxy-1,8-cineole, b2-hydroxy-1,8-cineole, a3-hydroxy-1,8-cineole, 7-hydroxy-1,8-cineole, 9-hydroxy-1,8-cineole, 2-oxo-1,8-cineole, 3-oxo-1,8-cineole, 2,3-dehydro-1,8- cineole, a2,3-epoxy-1,8-cineole and 4-hydroxy-1,8-cineole. There was considerable variability in the relative concentrations of metabolites found from sample to sample; however, 2-hydroxy-1,8-cineole was the dominant metabolite with at least a ten-fold higher concentration than the other metabolites [76]. Of interest is the oxidation product of 2-hydroxy-1,8-cineole, 2-oxo-1,8-cineole as well as the epoxide, may be Nrf2 activators.
Remarks: All four monoterpenes are lipophilic compounds whose inhalation pharmacokinetics are similar. Because of their lipophilicity they will absorb onto fat molecules and accumulate in fat tissue.
4. Therapeutic Effects
With forest bathing the exposure to VOCs is via the lungs. The highest concentration of VOCs would be experienced by the bronchiolar and alveolar epithelial cells followed by the endothelium of pulmonary capillaries, the pulmonary veins and then the endothelium of the systemic arterial circulation. It seems likely that the epithelium of the pulmonary system and the endothelium of the vascular systems play important roles in the potential therapeutic effects of VOCs. The importance of the endothelium to health was recognized over 70 years ago by Dr Rudi Altschul who wrote: ‘…one is as old as one’s endothelium” [77]. Modern research findings are in support of the importance of the endothelium to one’s health [78,79,80]. As noted above, the lung tissues have xenobiotic metabolizing enzymes but little attention has been paid to metabolites formed that may be unique to lung xenobiotic metabolism. Similarly, little attention has been paid to potential effects of VOCs and their metabolites on vascular function.
This will not be an exhaustive review of the literature and will focus on two health issues: inflammatory respiratory disorders and anxiety.
4.1. Anti-Inflammatory Effects: Asthma and Other Respiratory Problems
There are a number of reports that indicate forest bathing by urban children with asthma improves symptoms, e.g., [6,7]. Here, children are taken out of an urban environment to a forest environment. We do not know whether such improvements in asthma symptoms are due to leaving a more polluted air environment to going to a less polluted air environment, ‘mindfulness’ aspects of the forest environment, or due, in part the presence of monoterpenes. One study showed a positive correlation between lung function and total monoterpene exposure [7], although daily concentrations of monoterpenes measured were low and ranged from 38 to 62 ng/m3.
D-Limonene: Several studies have examined the effect of inhalation of D-limonene to ameliorate lung damage. In one, mice were sensitized to antigens from the mite Dermatophagoides farinae as well as diesel exhaust particles [81]. Here, mice were divided into 3 groups: the first group were sensitized to the mite antigens and diesel exhaust particles. In a second group, mice inhaled 0.01% (w/w) D-limonene that was nebulized using a sonicator for a 20 min period daily. Following this, the mice were administered mite antigen as well as diesel exhaust particles while a third group of mice were administered only the saline. After 48 hr the mice were administered acetylcholine to initiate airway hyperresponsiveness. The D-limonene group had greatly reduced airway hyperresponsiveness compared to the mice who received only the mite antigens and diesel exhaust particles. The D-limonene inhalation group showed greatly decreased eosinophil infiltration, IgE levels, decreased inflammatory cytokines such as IL5 and decreased tissue pathological changes.
Unfortunately, blood D-limonene was not measured in these experiments; however, the authors do report that mice were exposed over a 20 min period each day to 1 mg/kg D-limonene. If, as in humans, 65% of the D-limonene is transferred to the blood in the mice, then the mice absorbed 650 µg D-limonene/kg daily, i.e., 32.5 µg/kg/min. How does this compare to an exposure to D-limonene in a forest walk? Dudek and colleagues have reported that D-limonene may have an atmospheric concentration of 18 µg/m3 (i.e., 18 ng/L) in a mixed pine and hornbeam forest [82]. A 75 kg human walking on a flat surface breathes in 0.47 L/min/kg [83] and thus would breathe in ~8.5 ng/min/kg and absorb ~5.5 ng/min/kg, i.e., , almost 4 orders of magnitude less than the mice in the experiments by Hirota and colleagues.
Another inhalation study examined the ability of D-limonene to protect lungs from smoke damage [84]. Rats were divided into three groups: Control, Vehicle and Experimental. Vehicle and Experimental rats were exposed to smoke for three 9-min periods in a closed acrylic box interrupted by 30 sec exposures to ambient air. Following this the Vehicle group was exposed to the nebulized saline Vehicle while the Experimental group was exposed to nebulized D-limonene dissolved in saline for a period of 30 minutes. The D-limonene concentration in the chamber air was 4.5 µg/L, i.e., 33 nmoles/L, which is many orders of magnitude higher than what is found in forest environments, e.g., [30]. Tracheal TNFa was significantly increased in the Vehicle group compared to the Control Group and D-limonene greatly attenuated the TNFa response to smoke. The Ferric Reducing Ability of Plasma (FRAP) was significantly reduced in the Vehicle group compared to the Control group and FRAP was significantly increased in the Experimental group compared to the Vehicle group. The Vehicle group also had significantly decreased reduced-glutathione (GSH), catalase and superoxide dismutase activities in the lung while D-limonene inhalation countered these decreases. The lung histopathology was improved in the Experimental group compared to the Vehicle group showing less inflammatory cell infiltration, less edema and fewer vessels showing hyperemia. Thus, the D-limonene reduced the oxidative stress and inflammation caused by smoke inhalation. This study used atmospheric D-limonene almost three orders of magnitude higher than what can be encountered in a forest.
In an ovalbumin model of asthma in the mouse, a 20 min exposure to aerosolized 0.01% solution of D-limonene reduced airway hyper-responsiveness in sensitized mice to methacholine [85]. D-Limonene also reduced the number of inflammatory cells in bronchoalveolar lavage fluid and reduced the concentration of ovalbumin-specific IgG. The concentration of D-limonene was not measured, but would be higher than that found in a forest environment.
Computational docking modelling showed that D-limonene can bind to adenosine receptors A2A and A2B. The researchers determined that D-limonene had no protective effects on an ovalbumin model of asthma in A2A receptor knockout mice, concluding the protective effects of D-limonene in this asthma model were mediated by activating the adenosine A2A receptor.
a-Pinene: In a mouse model of rhinitis, a-pinene ameliorated symptoms in response to the allergen, ovalbumin [86]. Experimental mice received, daily, a-pinene perorally at doses ranging from 0.1 to 10 mg/kg. All doses ameliorated the allergic responses to ovalbumin including nasal IgE, TNFa, Macrophage Inflammatory Protein-2, and decreased numbers of inflammatory and mast cells infiltrating the nasal mucosa. The increased activation of Nuclear Factor Kappa B (NFkB) in response to ovalbumin was decreased following a-pinene administration. NFkB is a transcription factor that plays a central role in inflammation and when activated promotes the expression of dozens of pro-inflammatory genes [87,88]
Are the doses used relevant to forest bathing. Even at the lowest dose each mouse would have received 100 µg a-pinene/kg daily. Is this relevant to forest bathing? It is hard to say but if the difference in efficiency of transfer is 65 times greater for inhalation than ingestion of a-pinene is similar to that found with D-limonene [17], then one can make the reasonable assumption that 100 µg/kg ingested is equivalent to breathing in 1.5 µg/kg a-pinene. Let us consider the German forest where a-pinene has been reported to be ~6 µg/m3 or 6 ng/L [30]. A mouse has a lung tidal volume of ~0.18 mL and a respiratory rate of about 255 breaths per min [89]. In 1.0 min the mouse would have breathed in and out 0.0459 L and breathed in 0.294 ng a-pinene. Assuming 60% is transferred to the blood, then each minute the mouse would absorb 0.1764 ng a-pinene. Assuming a mouse has an average weight of 25 g, this is equivalent to 7.056 ng/kg/min. A mouse would receive a total dose of 1.5 µg/kg in 3.5 hr. If the forest atmosphere contained 23 µg/m3 as reported by Noe and colleagues [34], then the mouse would have breathed in 1.5 µg a-pinene in a little under an hr. One can question whether the experiments described are relevant to forest bathing.
Inhaling the metabolite, myrtenol, was also tested in an ovalbumin model of asthma in rats and shown to improve inflammatory markers and increase anti-oxidant status in the rat hippocampus [90]. The rats were sensitized to ovalbumin and exposed to a nebulized ovalbumin for 30 min a day for a period of 28 days and then exposed to different concentrations of myrtenol in the air they breathed in. Optimal results were obtained when rats were exposed to a total of 8 mg/kg myrtenol in the 30 min period. Assuming that 60% of the myrtenol entered the blood, then rats received a dose of 4.8 mg myrtenol/kg over the 30 min exposure period, i.e., 151 µg/kg/min or 1.05 µmole/kg/min. This dosage exposure does not seem relevant to human forest bathing.
D3-Carene: In an ovalbumin model of asthma, mice were administered intranasally one of the following: saline, the 0.9 µg steroid budesonide, 120 ng D3-carene or 1200 ng D3-carene [91]. Assuming the mice were 25 g in weight and transference 1,8-cineole to blood was the same as in humans (70%) [69], then the mice received 3.36 or 33.6 µg D3-carene/kg, i.e., 24.7 or 247 nmoles/kg D3-carene. Both budesonide and D3-carene, particularly the lower dose, decreased the inflammatory markers such as bronchiolar lavage polymorphonuclear leukocytes, bronchiolar epithelial thickness, plasma IgE and supressed the increase of the cytokines IL4, IL5 and IL13. Whether such effects are mediated by D3-carene or one of its metabolites is not known; however, we do know the parent compound is rapidly metabolized [69] while some of the metabolites such as D3-caren-10-carboxylic acid have a half-life of three hr [70]. Of interest is that D3-caren-10-carboxylic acid is a likely Michael electron acceptor and activator of Nrf2.
These results show that D3-carene has promises as an inhaler to treat asthma attacks but can one get sufficient D3-carene to have therapeutic effects during a forest walk. A northern boreal forest in Norway comprised mainly of spruce and some pine has a typical D3-carene atmospheric concentration 2 m above the ground of around 4 ppb, i.e., 22 µg/m3 (22 ng/L) [92]. Petersson [42] reported that D3-carene could be as high a concentration of 39 µg/m3 (39 ng/L) on a summer night in a pine forest in southern Sweden when measured a few cm above the forest floor. A 75 kg human walking on a flat surface breathes in 0.47 L/min/kg [83] and would take 7.7 hr to breathe in 4.8 µg D3-carene/kg in the Norwegian forest to deliver 3.36 µg D3-carene/kg to the blood. Lying on the forest floor in Sweden would still take over 4 hr to breathe in 4.8 µg D3-carene/kg. Thus, D3-carene has promises as an inhaler to treat asthma but it is not likely that sufficient D3-carene can be obtained through forest bathing to affect the symptoms of asthma, if the lower dose administered to the mice is near the optimal dose.
1,8-Cineole: In an ovalbumin-sensitized form of asthma in Guinea pigs, exposure to an atmosphere containing 79 mg 1,8-cineole/m3 for 15 min ameliorated the response to an ovalbumin challenge [93]. It decreased the inflammatory cytokines TNFa and IL1b in broncho-alveolar lavages and decreased airway hyperresponsiveness to carbachol. Again, these are very high concentrations of 1,8-cineole compared to what one may encounter in a forest. Although the oxidized monoterpene 1,8-cineole is considered to be one of the major monoterpenes emitted from North American forests [94], there are few studies examining the emission rate or forest atmosphere levels of 1,8-cineole. One such study is by Hellén and colleagues who report maximal levels of 19 ppt (0.12 µg/m3) in a boreal forest from southern Finland [44] while Hakola and colleagues reported that 1,8-cineole can form up to 8% of all monoterpenes in the forest atmosphere of a northern boreal forest [95].
In a study examining the effects of protective effects of 1,8-cineole against the harmful effects of smoking, mice were exposed for 72 minutes intermittently to the smoke of 12 cigarettes each day for 5 days in the presence or absence of 1,8-cineole in the atmosphere [96]. The 1,8-cineole was administered as an aerosol into 4 L chambers in which the mice were placed for 15 min daily. The aerosolized 1,8-cineole used was 1, 3 and 10 mg, i.e., the chambers contained 250 mg/m3, 750 mg/m3 or 2,500 mg/m3, concentrations of at least 5 orders of magnitude greater than can be found in a forest. Protective effects were seen at 3 and 10 mg aerosolized concentrations as shown by reduced inflammatory cell infiltration of lungs, reduced NFkB activation, decreased levels of inflammatory cytokines such as IL1b, IL6 and less oxidative stress.
There are other animal studies showing that 1,8-cineole has positive effects on insults to the respiratory system [97]; however, these studies typically used high doses of 1,8-cineole and usually administered intraperitoneally.
A randomized, placebo-controlled, double-blinded clinical trial with asthma patients is described where 200 mg 1,8-cineole was ingested 3 times daily for a period of 6 months [98]. The average weight of the patients was ~77 kg; hence, patients consumed a dose of 16.8 µmoles/kg thrice daily. Patients who consumed 1,8-cineole showed marked improvements compared to the placebo group. But, again, such studies, although showing promising therapeutic effects of 1,8-cineole, are not relevant to forest bathing.
In vitro studies examining the response of primary cultures of human monocytes to LPS show that low micromolar concentrations of 1,8-cineole inhibit release of prom-inflammatory cytokines [99,100]. These studies also showed that 1,8-cineole at these low concentrations synergized with the effect of nebulizer steroid budesonide.
4.2. Anxiolytic Effects
Donelli and colleagues reported on a propensity score matching study of over 500 patients with anxiety that were introduced to a 3 hr period of low intensity exercise in a forest environment at several locations in Italy [101]. Extent of reductions in anxiety were related to total atmospheric monoterpene levels as well as the dominant monoterpene, a-pinene, levels. Total monoterpene concentrations ranged from 0.10 to 0.99 µg/m3.
In a controlled laboratory setting Kim and colleagues examined how a mixture of a-pinene, b-pinene and D-limonene affected stress levels in human subjects [102]. They demonstrated that 20% of the subjects could detect each monoterpene at a concentration of 3 ppb (16.7 µg/m3) and all could detect the each monoterpene at a concentration of 15 ppb (83.5 µg/m3). In subsequent experiments they chose a mixture of a-pinene:b-pinene:D-limonene at ratios of 1.0:0.5:0.5. Subjects inhaled 7 ppb (39 µg/m3), 15 (83.6 µg/m3) or 20 ppb (111.4 µg/m3) of total monoterpenes. These levels can be found in Norway spruce and Scots pine forests as measured by Hov and colleagues [92] at a level 2 m above the ground forests, where they report that the concentration sum of the a-pinene, b-pinene and D-limonene can be as high as 21 ppb (i.e., 117 µg/m3). Breathing an atmosphere of 39 µg/m3 of total monoterpenes for 2.5 min decreased subject stress as measured by brain alpha waves and heart rate variability with decreasing stress with higher levels of the monoterpene mixture. Thus, we have evidence that inhaling monoterpene mixtures at a concentration that can be found in a forest has health benefits. Whether this is due to ‘mindfulness’ mediated effects due to the pleasantness of the odours or whether these effects were mediated by direct action of the monoterpenes on physiological processes is unclear.
A study with human subjects breathing air that passed over 2 mL of 1,8-cineole for a period of 20 min, showed an with increased global cerebral blood flow [103], even in an anosmic subject. This indicated that the increased global cerebral blood flow was not mediated via olfactory perception. Thus, it is very possible that the decrease in anxiety in the studies described above was mediated by monoterpenes activating physiological processes independent of olfaction.
D-Limonene: There are a number of animal studies examining the anxiolytic effects of D-limonene but they all used high doses of D-limonene that would not be encountered in forest bathing; hence, only two studies will be considered. An elevated plus maze experiment in mice showed that inhaled D-limonene had anxiolytic effects similar to that of the GABAA diazepine receptor agonist diazepam [104]; however, the anxiolytic effect of D-limonene was not blocked by flumazenil, the GABAA diazepine receptor antagonist. This is interesting since the GABAA receptor is the target of anxiolytic drugs [105]. The concentration of D-limonene used in these experiments was enormous, since the animals were exposed to air concentrations greater than 1 µg/cm3 and this high concentration might have been able to outcompete flumazenil for the receptor binding site.
Another elevated plus maze study used 1 to 10 mg D-limonene/kg in mice showed that prior intraperitoneal injection of D-limonene significantly decreased the anxiety of mouse placed in an elevated maze at the 5 and 10 mg/kg doses administered intraperitoneally [106]. These doses are also quite high, but lower than in the inhalation experiments, being 36.7 to 73 µmoles/kg. The authors also showed that the 10 mg D-limonene/kg decreased GABA content and increased dopamine content in the striatum of the mice. In vitro experiments with PC12 cells suggested that the decrease in GABA content was due to increased release of GABA from the neurons. The administration of an A2A receptor antagonist eliminated the anxiolytic effect of D-limonene.
a-Pine: There are few studies examining the anxiolytic effects of a-pinene or its metabolites. There is one paper that examines the ability of a-pinene to ameliorate anxiety in rats and this is in Persian but has an English abstract [107]. The author states that injection of 2 mg (14.7 µmoles) and 4 mg (29.4 µmoles) a-pinene/kg significantly reduced anxiety as determined by the elevated pus maze test, comparable to that of diazepam. The administration of a-pinene also decreased malondialdehyde and increased GSH levels in the hippocampus.
Myrtenol, a metabolite of a-pinene, administered intraperitoneally at 25, 50 and 75 mg/kg significantly increased the time rats were present in the open in the elevated plus maze and Light-Dark Transition test with the lowest dose having the greatest anxiolytic effect [108]. The GABAA receptor antagonist flumazenil was able to reverse the anxiolytic effect of myrtenol. Since 25mg/kg had the greatest effect, it is unfortunate that lower doses were not tested.
D3-carene: There are no studies examining the effect of D3-carene on anxiety amelioration; however, one study examined the ability of D3-carene to enhance sleep [109]. D3-Carene was injected intraperitoneally at doses ranging from 12.5 mg (92 µmoles)/kg to 100 mg (734 µmoles)/kg, which are quite high doses. The mice were also injected with pentobarbital and time to sleep as well as sleep duration were measured. Doses of 25 mg/kg or greater significantly deceased the time to sleep while doses of 50 mg/kg or greater significantly increased the duration of sleep. This sleep-enhancing effect was mediated through the GABAA benzodiazepine receptor since it was inhibited by flumazenil.
1,8-Cineole: There are a number of papers that describe the anxiolytic effects of 1,8-cineole in rodents. I will consider only two. In the first rats were injected intraperitoneally with 1 mg 1,8-cineole (7.34 µmoles)/kg and subjected to the elevated plus maze [110]. This dose of 1,8-cineole decreased anxiety and was blocked by flumazenil, an antagonist of the GABAA benzodiazepine receptor.
A more relevant study is described by Dougnon and Ito [111]. Here, 1,8-cineole was dissolved in triethyl citrate and then placed on filter papers in closed cages with total volumes of 61.2 L [111]. Total amounts of 1,8-cineole placed in each cage ranged from 4 ng to 100 µg. Thus, if all of the cineole volatilized, mice would encounter an atmosphere containing anywhere from 65 ng/m3 to 1.63 mg/m3. After volatilization for 60 min mice were introduced into the cages and monitored every 5 min for a variety of anxiety tests including the Light-Dark box. Decreases in anxiety was seen in the Light-Dark box test at 6.5 µg 1,8-cineole/m3 with lower and higher concentrations having no effect on anxiety. Flumazenil, a GABAA benzodiazepine antagonist blocked the anxiolytic effect of 1,8-cineole. A concentration of 6.5 µg 1,8-cineole/m3 can possibly be encountered in a forest setting.
A small clinical trial demonstrated that inhaling 1,8-cineole for 5 min before selective nerve root block decreased anxiety in human patients [112]; however, a high concentration of 1,8-cineole in air was breathed in.
5. Discussion
5.1. Pharmacokinetics
The pharmacokinetics of all 4 monoterpenes is similar. The elimination of the parent compound from the blood generally occurs in 3 phases with the first phase being rapid (~5-10 min half-life) where the compound in the aqueous phase is metabolized and volatilized from the lungs, a second slower phase (~30-60 min half-life) likely involving movement of blood from lipophilic blood compounds such as low and high density lipoproteins into the aqueous phase of blood as well as some transfer from adipose tissue into blood, and a final slow phase that may have a half-life of 10 or more hr where the monoterpene is transferred from adipose tissue to the blood. The exception is the oxidized monoterpene 1,8-cineole where the slow phase has a half-life under 3 hr. This is likely due to 1,80-cineole being less soluble in lipids - it has an octanol-water partition coefficient of only 2.8 compared to 4.4 to 4.5 for D3-carene, D-limonene and a-pinene [113].
There is rapid metabolism of the parent compound with many metabolites present for a longer time in the blood. There have been few studies examining the half-lives of the metabolites. These are important studies to do since many of the therapeutic effects may be mediated by the metabolites rather than the parent compound. It should be noted that the majority of monoterpene metabolism studies used ingested monoterpene rather than inhaled monoterpene.
5.2. Mode of Administration
It is clear that the 4 monoterpenes considered have anxiolytic and anti-inflammatory effects; however, the doses examined experimentally are generally well above those that would be encountered in forest bathing. As mentioned, since the parent compounds are rapidly metabolized, it is likely that many of the therapeutic effects recorded are mediated by metabolites. The metabolites formed are derived by the Phase 1 xenobiotic metabolizing enzymes that are present in different tissues [114]. Ingested monoterpenes are primarily metabolized by the intestine, liver and kidneys [54]. However, the lung epithelium from the bronchioles to the alveoli also have xenobiotic metabolizing enzymes, including both cytochrome P450 (CYP450) and non-CYP450 oxidoreductases [51]. Although the activities of these xenobiotic metabolizing enzymes generally are lower in the lungs than other organs such as the liver, the enormous surface area of the lungs makes this a less important issue. That route of entry may make a difference in the therapeutic effects of monoterpenes is demonstrated by the experiments of Heuberger and colleagues [52]. Their experiments exposed human subjects to 1,8-cineole either by inhalation or by transdermal applications. The dermal absorption of the 1,8-cineole improved speed of attentional performance while inhalation had the opposite effect but increased the sense of well-being. Large doses were used in these experiments: 20 min inhaling an atmosphere containing 6.8 mg/L and 86.4 mg applied transdermally. If we assume that 70% of the inhaled 1,8-cineole was transferred to blood then each subject would have inhaled 45 mg of the monoterpene, quite comparable to what would have been absorbed transdermally. These experiments do point out that inhalation of a monoterpene compared to other means of administration may make a difference in physiological responses.
Most monoterpene inhalation studies are at concentrations far greater than could possibly be found in a forest environment. The exceptions are the study by Kim and colleagues who demonstrated that a mixture of a-pinene, b-pinene and D-limonene with a combined concentration of 39 µg/m3 breathed in for 2.5 min decreased anxiety in human subjects as demonstrated by heart rate variability and brain alpha waves [102] and the experiments by Dougnon and Ito [111] who showed that 6.5 µg 1,8-cineole/m3 can reduce anxiety in mice. More such inhalation studies using concentrations of monoterpenes found in forest environments need to be performed to understand how VOCs may affect health.
5.3. Possible Therapeutic Mechanisms of Action
The focus of this manuscript are the mechanisms involved in decreasing anxiety and inflammation in the respiratory system.
5.3.1. Anti-Inflammatory Mechanisms
In the studies referenced in this paper there are two possible mechanisms that should be investigated further: activation of the adenosine receptor A2A and activation of NFkB. As mentioned previously, NFkB plays a central role in promoting the expression of pro-inflammatory genes [87]. NFkB normally exists as a dimer that is bound to a member of the Inhibitor of kappa B proteins (IkBs) [115]. NFkB is activated by signalling pathways that activate IkB kinase (IKK) to phosphorylate specific serine residues on IkB that result in the degradation of IkB via the ubiquitin-proteasomal pathway, thus exposing the nuclear localization signal allowing the NFkB to translocate to the nucleus to bind to kB elements, thereby promoting the expression of dozens of pro-inflammatory genes. Factors that regulate and inhibit the activation of NFkB are complex [115].
There are also connections between the adenosine receptor, A2A and NFkB. There are a number of papers that show activating the A2A receptor decreases activation of NFkB [116,117]. Furthermore, Sands and colleagues showed that overexpressing the A2A receptor gene in C6 glioma cells decreased the inflammatory response to stimuli such as LPS by inhibiting the phosphorylation of IkB [118], perhaps through inducing the Dual Specificity Phosphatase 1 (DUSP1) [119].
5.3.2. Anxiolytic Mechanisms
The anxiolytic mechanisms appear to involve the promotion of the activation of the GABAA receptor since it is blocked by flumazenil. This may not involve monoterpenes or their metabolites directly acting on the GABAA receptor. They may act on mechanisms that ultimately result in the production of an endogenous ligand such as inosine [122] or a neurosteroid [123] for the benzodiazepine binding site of the GABAA receptor or 2-arachidonoly glycerol for the a2 GABAA receptor subunit [124].
Most studies examining the anxiolytic effects of monoterpenes use high concentrations of the monoterpene; however the studies by Kim and colleagues [102] and the experiments by Dougnon and Ito [111] show that concentrations of monoterpenes that can be found in forest environments can decrease anxiety. There are interesting observations by Hajagos-Tóth and colleagues that examined mechanisms whereby D-limonene promotes contraction of the myometrium of the pregnant rat [125]. These experiments were performed ex vivo and showed that D-limonene at 1 pM concentration promoted myometrial smooth muscle contraction and this was blocked either by a calcium L channel blocker or by a non-specific adenosine receptor blocker. Concentrations of D-limonene even as low as 0.1 pM caused an increase in cAMP in the myometrium. The researchers propose that D-limonene activated A2A receptors resulting in an increase in intracellular cAMP that in turn enabled Protein kinase A to phosphorylate L channels allowing movement of calcium ions into the cytoplasm thereby causing contraction. These observations should be investigated further since they may lead to an understanding of the mode of action of low concentrations of D-limonene or its metabolites. D-Limonene is known to bind to the adenosine A2A receptor but with half-maximal binding being around 5 µM [126] and the concentration to cause half maximal cAMP production is ~5 µM, clearly much higher than seen in the experiments by Hajagos-Toth and colleagues. It may well be possible that D-limonene binds to a yet to be discovered macromolecule that interacts with the A2A receptor such as how 2-arachidonoyl glycerol promotes activation of the GABAA receptor [124].
6. Future Directions
6.1. Determining Relevant Metabolites
The highest concentration of any monoterpene reported in a forest atmosphere is about 50 µg/m3 with most reports being under 10 µg/m3 and often below 1 µg/m3. If we are to determine whether breathing in monoterpenes at levels found in forests can affect our health by activating physiological processes, then all experiments must be conducted inhaling relevant concentrations of the monoterpenes found in a forest atmosphere. Since lung tissue can also catabolize xenobiotics and some of such metabolites may differ from that formed by the intestine, liver or kidneys, one should determine the metabolites that are formed through inhaling specific monoterpenes and determine the metabolite half-lives since the therapeutic effects may be mediated by monoterpene metabolites, particularly those with long half-lives.
6.2. Inflammation
The majority of therapeutic effects of monoterpenes reported decrease anxiety and/or inflammation. There are many reports that what underlies anxiety and depressive disorders is inflammation [127,128,129] and inflammation underlies many chronic diseases and diseases related to ageing [130].
A2A Receptor: As pointed out above, activation of the A2A adenosine receptor decreases both anxiety and inflammation. Since the A2A receptor seems to be involved in both anxiety and inflammation, actions of monoterpenes and their metabolites on the A2A receptor should investigated using several different preparations, including the ex vivo myometrium of pregnant rats and in vitro cultures of vascular endothelium and respiratory epithelium.
NFkB: NFkB is a central player in inflammation [87,88]. Activation of NFkB is a consequence of kinase cascades that result in the phosphorylation of IkB that leads to its degradation and exposure of the nuclear localization signal on NFkB allowing translocation to the nucleus and binding to kB elements, thereby promoting the expression of multiple pro-inflammatory genes [115].
Kinase activity is countered by protein phosphatase activity [131]. The catalytic centre of phosphatases are redox-sensitive and are either cysteine centred or centred on a dimetallic cluster containing iron and either zinc or manganese [132]. Protein phosphatases can be inactivated by oxidation of their catalytic centre; hence phosphorylation state of IkB can be due to either activation of protein kinase cascades or to inhibition of protein phosphatases [133]; hence, oxidative stress increases overall kinase phosphorylation status by inhibiting protein phosphatase activity. Thus, cellular redox regulates the likelihood of NFkB activation [87] and decreasing cellular oxidative stress will decrease the probability of NFkB activation [134] and, thus, inflammatory conditions [135].
Nrf2: The Nuclear factor (erythroid-derived) like-2 (Nrf2) signalling system plays a central role in decreasing oxidative stress in a cell [134,136,137]. Nrf2 is normally located in the cytosol bound to a homodimer of Kelch-like ECH-Associated Protein (Keap1). Nrf2 is continually synthesized but Nrf2 bound to Keap1 allows it to be polyubiquinated, tagging it for proteasomal degradation. Electrophiles of a particular geometric and electronic configuration can oxidize critical cysteines on Keap1, dissociating Keap1 from Nrf2, allowing Nrf2 to translocate to the nucleus where it heterodimerizes with small Maf proteins and bind to anti-oxidant (or Electrophile) Response Elements (AREs or EpREs) in the promoter regions of multiple genes, many of whose protein products decrease the probability of forming strong oxidants or scavenge strong oxidants.
Readers should be beware that I view the potential therapeutic effects of bioactive molecules through a lens that that may be skewed to Nrf2 activators. That Nrf2 activators, previously known as phase 2 enzyme inducers, could play a role in health by preventing cancer formation was first proposed by Paul Talalay of John Hopkins University [138]. He is considered to be the founder of the pharmacology of chemoprotection [139]. A review of his many accomplishments can be seen in the paper by Johnson [140]. I had been working on understanding the relationship between oxidative stress and inflammation and became intrigued with Talalay’s research and realized that the rise in cellular GSH following an oxidative insult was due to the induction of g-glutamyl-cysteine ligase, the rate limiting enzyme for GSH synthesis. Glutamyl-cysteine ligase has AREs in the promoter regions of both the regulatory and catalytic subunits [141]. I realized that phase 2 enzyme inducers could protect not only against cancer but protect against diseases that have underlying oxidative stress & inflammation components [142]. This led to research on this topic in my laboratory and publication of papers demonstrating that dietary Nrf2 activators (phase 2 enzyme inducers) could ameliorate many chronic diseases, e.g., [143,144,145,146,147,148,149]. Many laboratories are now investigating the therapeutic effects of Nrf2 activators [150].
Many of the monoterpenes and their metabolites appear to involve activation of the Nrf2 signalling pathway: D-limonene [151], perillyl alcohol [152], carveol [153,154], a-pine [155], myrtenol [156], and 1,8-cineole [155,157], The D3-carene metabolites, D3-caren-10-carboxylic acid and D3-carene epoxide may also be Nrf2 activators.
I suggest monoterpenes and their metabolites to be screened for their ability to cause Nrf2 activation. A simple approach would be to use cell lines and screen for the ability of the compounds of interest to increase cellular GSH. The simplest way to measure GSH is to add monochorobimane to living cell cultures allowing the endogenous glutathione S-transferases to form a glutathione-monochlorobimane adduct – this adduct can be excited at 380 nm to emit at 470 nm [158]. This approach allows rapid screening of potential Nrf2 activators. I would suggest a number of different cell lines be examined, including bronchial epithelial, vascular endothelial, hepatic, etc., since there may be differences in the cells abilities to respond to monoterpenes and their metabolites.
6.3. Possibilities of Synergies
To further complicate issues there may well be synergistic interactions amongst the monoterpenes or their metabolites where each compound affects separate signalling components that activate a particular therapeutic pathway. One such example is where an Nrf2 activator phosphorylates serine residues on Keap1 but the likelihood Nrf2 translocating to the nucleus is also determined by the phosphorylation status of particular amino acid residues on Nrf2. Thus, protocatechuic acid results in Nrf2 phosphorylation [159] making it easier for Nrf2 to separate from Keap1 while sulforaphane oxidizes cysteine residues in Keap1 allowing release of Nrf2 [160]. Thus, protocatecuic acid in this example might provide the ‘tipping point’ in whether low concentrations of an Nrf2 activator may initiate the Nrf2 signalling pathway.
One example of such possible synergies with terpenes is the experiments conducted by Bai and colleagues first on an in vitro model of cardiac hypertrophy followed by an animal model [161]. Again the in vitro and in vivo doses were large (100 µM) but the results clearly showed that combining 1,8-cineole with b-caryophyllene had synergistic effects.
One can conclude that not only must one use monoterpene atmospheric concentrations that may be present in forests but one should use combinations of monoterpenes in concentrations that might be found in forests since therapeutic effects of forest bathing may be due to synergies in the actions of the various monoterpenes. An example of such a study is that by Kim and colleagues who looked at the physiological effects of a combination of b-pinene, a-pinene and D-limonene [102] using blood pressure, heart rate and heart rate variability as the outcomes.
7. Conclusions
There is clear evidence that one of the therapeutic mechanisms of forest bathing is mediated by ‘mindfulness’ promoting parasympathetic activities. Moving from a more polluted urban environment to a less polluted forest environment perhaps also accounts for some of the therapeutic effects. Clearly, monoterpenes encountered at higher concentrations than are present in forest atmospheres have effects on physiological processes that promote better health outcomes. For example, a Phase III clinical trial has been completed that shows the efficacy of 1,8-cineole in reducing cough frequency and other symptoms of bronchitis [162] and this monoterpene shows great promise as an adjunct therapy for asthma [100]. But there is enough intriguing evidence that some of the forest bathing’s therapeutic effects are mediated by the monoterpenes present in the forest atmosphere activating physiological processes that promote health, but this needs further investigation.
For therapeutic mechanisms, two avenues that seem worthwhile to explore: the activation of the adenosine A2A receptor and the activation of the Nrf2 signalling pathway. Because the vascular system is exposed to high levels of monoterpenes and their metabolites, another avenue should be investigated is the effect of these compounds on endothelial funmction.
Acknowledgments
I am retired but I thank the following funding agencies that provided grant support for my past research into bioactive molecules: National Research and Engineering Council of Canada, The Canadian Institutes of Health Research, The Christopher Reeve Paralysis Foundation and Saskatchewan’s Agriculture and Food Development Fund .
Conflicts of Interest
There are no conflicts of interest
| 1 | tuberculosis |
References
- Bostock, J.; Riley, H. T., The Natural History of Pliny. Henry G. Bohn; Project Gutenberg: https://www.gutenberg.org/cache/epub/60688/pg60688-images.html: London, 1865; Vol. V.
- Li, Q. , Effects of forest environment (Shinrin-yoku/Forest bathing) on health promotion and disease prevention -the Establishment of "Forest Medicine". Environ Health Prev Med 2022, 27, 43. [Google Scholar] [CrossRef] [PubMed]
- Victorson, D. , Cultivating Reciprocity Between People and Planet: Habit-Stacking Planetary Health Prescriptions Into Existing Nature RX Encounters During Integrative Health Visits. Glob Adv Integr Med Health 2024, 13, 27536130241245429. [Google Scholar] [PubMed]
- White, M. P.; Alcock, I.; Grellier, J.; Wheeler, B. W.; Hartig, T.; Warber, S. L.; Bone, A.; Depledge, M. H.; Fleming, L. E. , Spending at least 120 minutes a week in nature is associated with good health and wellbeing. Sci Rep 2019, 91, 7730. [Google Scholar] [CrossRef]
- Chun, M. H.; Chang, M. C.; Lee, S. J. , The effects of forest therapy on depression and anxiety in patients with chronic stroke. Int J Neurosci 2017, 1273, 199–203. [Google Scholar] [CrossRef]
- Seo, S. C.; Park, S. J.; Park, C. W.; Yoon, W. S.; Choung, J. T.; Yoo, Y. , Clinical and immunological effects of a forest trip in children with asthma and atopic dermatitis. Iran J Allergy Asthma Immunol 2015, 141, 28–36. [Google Scholar]
- Donelli, D.; Antonelli, M.; Baraldi, R.; Corli, A.; Finelli, F.; Gardini, F.; Margheritini, G.; Meneguzzo, F.; Neri, L.; Lazzerini, D.; Ardissino, D.; Piacentini, G.; Zabini, F.; Cogo, A. , Exposure to forest air monoterpenes with pulmonary function tests in adolescents with asthma: A cohort study. Forests 2012, 14. [Google Scholar] [CrossRef]
- Tsao, T. M.; Hwang, J. S.; Lin, S. T.; Wu, C.; Tsai, M. J.; Su, T. C. , Forest Bathing Is Better than Walking in Urban Park: Comparison of Cardiac and Vascular Function between Urban and Forest Parks. Int J Environ Res Public Health 2022, 196. [Google Scholar] [CrossRef]
- Mao, G. X.; Cao, Y. B.; Lan, X. G.; He, Z. H.; Chen, Z. M.; Wang, Y. Z.; Hu, X. L.; Lv, Y. D.; Wang, G. F.; Yan, J. , Therapeutic effect of forest bathing on human hypertension in the elderly. J Cardiol 2012, 606, 495–502. [Google Scholar] [CrossRef]
- Park, B. J.; Tsunetsugu, Y.; Kasetani, T.; Kagawa, T.; Miyazaki, Y. , The physiological effects of Shinrin-yoku (taking in the forest atmosphere or forest bathing): evidence from field experiments in 24 forests across Japan. Environ Health Prev Med 2010, 151, 18–26. [Google Scholar] [CrossRef]
- Vermeesch, A. L.; Ellsworth-Kopkowski, A.; Prather, J. G.; Passel, C.; Rogers, H. H.; Hansen, M. M. , Shinrin-Yoku (Forest Bathing): A Scoping Review of the Global Research on the Effects of Spending Time in Nature. Glob Adv Integr Med Health 2024, 13, 27536130241231258. [Google Scholar] [CrossRef]
- Keith, R. J.; Hart, J. L.; Bhatnagar, A. , Greenspaces And Cardiovascular Health. Circ Res 2024, 1349, 1179–1196. [Google Scholar]
- Babak, A.; Motamedi, N.; Mousavi, S. Z.; Ghasemi Darestani, N. , Effects of Mindfulness-Based Stress Reduction on Blood Pressure, Mental Health, and Quality of Life in Hypertensive Adult Women: A Randomized Clinical Trial Study. J Tehran Heart Cent 2022, 173, 127–133. [Google Scholar]
- Dada, T.; Mittal, D.; Mohanty, K.; Faiq, M. A.; Bhat, M. A.; Yadav, R. K.; Sihota, R.; Sidhu, T.; Velpandian, T.; Kalaivani, M.; Pandey, R. M.; Gao, Y.; Sabel, B. A.; Dada, R. , Mindfulness Meditation Reduces Intraocular Pressure, Lowers Stress Biomarkers and Modulates Gene Expression in Glaucoma: A Randomized Controlled Trial. J Glaucoma 2018, 2712, 1061–1067. [Google Scholar]
- Hofmann, S. G.; Gomez, A. F. , Mindfulness-Based Interventions for Anxiety and Depression. Psychiatr Clin North Am 2017, 404, 739–749. [Google Scholar]
- Song, C.; Ikei, H.; Myazaki, Y. , Pysiological effects of visual stimulation with forest imagery. Int. J. Environ. Res. Pub. Health 2018, 15, 213. [Google Scholar] [CrossRef]
- Chen, C.; Sheng, Y.; Hu, Y.; Li, W.; Feng, H.; Tang, L. , Determination of D-limonene in mice plasma and tissues by aa new GC-MS/MS method: Comparison of the pharmokinetics and tissue distribution by oral and ihalation administration in mice. Biomed Chromatogr 2019, 337, e453. [Google Scholar]
- Sumitomo, K.; Akutsu, H.; Fukuyama, S.; Minoshima, A.; Kukita, S.; Yamamura, Y.; Sato, Y.; Hayasaka, T.; Osanai, S.; Funakoshi, H.; Hasebe, N.; Nakamura, M. , Conifer-Derived Monoterpenes and Forest Walking. Mass Spectrom (Tokyo) 2015, 41, A0042. [Google Scholar]
- Li, J.; Perdue, E. M.; Paviostathis, S. G.; Auraujo, R. , Physicochemical properties of selected monoterpenes. Environ. Int. 1998, 243, 353–358. [Google Scholar]
- Wu, L.; Juurlink, B. H. , The impaired glutathione system and its up-regulation by sulforaphane in vascular smooth muscle cells from spontaneously hypertensive rats. J Hypertens 2001, 1910, 1819–25. [Google Scholar]
- Satou, T.; Hayakawa, M.; Kasuya, H.; Masuo, Y.; Koike, K. , Mouse brain concentrations of alpha-pinene, limonene, Linalool, and 1,8, cineole following inhalation. Flavour Fragrance J. 2017, 32, 36–39. [Google Scholar]
- Ahn, C.; Kim, J.-W.; Park, M.-J.; Kim, S. R.; Lee, S.-S.; Jeung, E.-B. , Anti-inflammatory effects of natural volatile organic compounds from Pinus koraiensis and Larix kaempferi in mouse model. J. Biomed. Res. 2019, 335, 343–350. [Google Scholar] [CrossRef]
- Ahn, C.; Lee, J. H.; Kim, J. W.; Park, M. J.; Lee, S. S.; Jeung, E. B. , Alleviation effects of natural volatile organic compounds from Pinus densiflora and Chamaecyparis obtusa on systemic and pulmonary inflammation. Biomed Rep 2018, 95, 405–414. [Google Scholar]
- Page, M. J.; Kell, D. B.; Pretorius, E. , The role of lipopolysaccharide-induced cell signalling in chronic inflammation. Chronic Stress 2022, 6, 1–18. [Google Scholar]
- Laothawornkitkul, J.; Taylor, J. E.; Paul, N. D.; Hewitt, C. N. , Biogenic volatile compounds in the Earth System. New Phytologist 2009, 1831, 27–51. [Google Scholar]
- Acosta Navarro, J. C.; Smolander, S.; Struthers, H.; Zorita, E.; Ekman, A. M.; Kaplan, J. O.; Guenther, A.; Arneth, A.; Riipinen, I. , Global emissions of terpenoid VOCs from terrestrial vegetation in the last millennium. J Geophys Res Atmos 2014, 11911, 6867–6885. [Google Scholar] [CrossRef]
- Mäki, M.; Aalto, J.; Hellen, H.; Pihlatie, M.; Back, J. , Interannual and Seasonal Dynamics of Volatile Organic Compound Fluxes From the Boreal Forest Floor. Front Plant Sci 2019, 10, 191. [Google Scholar]
- Geron, C.; Rasmussen, R.; Arnts, R. R.; Guenther, A. , A review and synthesis of monoterpenes speciation from forests in the United States. Atmos. Environ. 2000, 34, 1761–1781. [Google Scholar]
- Runyon, J. B.; Gray, C. A.; Jenkins, M. J. , Volatiles of High-Elevation Five-Needle Pines: Chemical Signatures through Ratios and Insight into Insect and Pathogen Resistance. J Chem Ecol 2020, 463, 264–274. [Google Scholar]
- Borsdorf, H.; Bentele, M.; Müller, M.; Rebmann, C.; Mayer, T. , Comparison of seasonal and diurnal concentration profiles of BVOCs in coniferous and deciduous forests. Atmosphere 2023, 14. [Google Scholar]
- Calogirou, A.; Larsen, B. R.; Kotzias, D. , Gas-phase terpene oxidation products: a review. Atmos. Envirn. 1999, 33, 1423–1439. [Google Scholar]
- Bates, K. H.; Jacob, D. J. , A new model mechanism for atmospheric oxidation of isoprene: global effects on oxidants, nitrogen oxides, organic products, and secondary organic aerosol. Atmos. Chem. Phys. 2019, 19. [Google Scholar] [CrossRef]
- Ferracci, V.; Bolas, C. G.; Freshwater, R. A.; Staniaszek, Z.; King, T.; Jaars, K.; Otu-Larbi, F.; Beale, J.; Malhi, Y.; Waine, T. W.; Jones, R. L.; Ashworth, K.; Harris, N. R. P. , Continuous isoprene measurments in a UK Temperate Forest for a whole growing season: Effects of drought stress during the 2018 heatwave. Geophys. Res. Lett. 2020, 47, e2020GL088885. [Google Scholar] [CrossRef]
- Noe, S. M.; Copolovici, L.; Niinemtes, Ü. , Seasonal variation in vertical volatile compounds air concentration within a remote hemiboreal mixed forest. Atmos. Chem. Phys. 2012, 12, 3909–3926. [Google Scholar]
- Sukul, P.; Richter, A.; Junghanss, C.; Schubert, J. K.; Miekisch, W. , Origin of breath isoprene in humans is revealed via multi-omic investigations. Commun Biol 2023, 61, 999. [Google Scholar]
- Cailleux, A.; Cogny, M.; Allain, P. , Blood isoprene concentrations in humans and in some animal species. Biochem Med Metab Biol 1992, 472, 157–60. [Google Scholar]
- Wang, L.; Lun, X.; Wang, Q.; Wu, J. , Biogenic volatile organic compounds emissions, atmospheric chemistry, and environmental implications: a review. Environ. Chem. Lett. 2024, 22, 3033–3058. [Google Scholar]
- Hakola, H.; Tarvainen, V.; Laurila, T.; Hiltunen, V.; Hellén, H.; Keronen, P. , Seasonal variation of VOC concentrations above a boreal coniferous forest. Atmos. Environ. 2003, 37, 1623–1634. [Google Scholar]
- Riba, M. L.; Tathy, J. P.; Tsiropouos, N.; Monsarat, B.; Torres, L. , Diurnal variation in the concentration of a-pinene and b-pinene in the Landes Forest (France). Atmos. Environ. 1987, 211, 191–193. [Google Scholar]
- Staudt, M.; Lhoutellier, L. , Monoterpene and sesquiterpene emissions from Quercus coccifera exhibit interacting response to light and temperature. Biogeosci. 2011, 8, 2757–2771. [Google Scholar]
- Hakola, H.; Hellén, H.; Tarvainen, V.; Bãck, J.; Patokoski, P. , Annual variations of atmospheric VOC concentrations in a boreal forest. Boreal Environ. Res. 2009, 14, 722–730. [Google Scholar]
- Petersson, G. , High ambient concentrations of monoterpenes in a Scandanavian pine forest. Atmos. Environ. 1988, 22, 2617–2619. [Google Scholar]
- Oh, G.-Y.; Park, G.-H.; Kim, I.-S.; Bae, J.-S.; Park, H.-Y.; Seo, Y.-G.; Yang, S.-I.; Lee, J.-K.; S-H. , J.; Lee, W.-J., Comparison of major monoterpene concentrations in the ambient air of South Korea forests. J. Korean For. Soc. 2010, 995, 698–705. [Google Scholar]
- Hakola, H.; Hellén, H.; Hemmilä, M.; Rinne, J.; Kulmala, M. , In situ measurement of volatile organic compounds in a boreal forest. Atmos. Chem. Phys. 2012, 12, 11665–11678. [Google Scholar]
- Wang, X.; Li, G.; Shen, W. , Protective effects of D-Limonene against transient cerebral ischemia in stroke-prone spontaneously hypertensive rats. Exp Ther Med 2018, 151, 699–706. [Google Scholar]
- Eddin, L. B.; Azimullah, S.; Jha, N. K.; Nagoor Meeran, M. F.; Beiram, R.; Ojha, S. , Limonene, a Monoterpene, Mitigates Rotenone-Induced Dopaminergic Neurodegeneration by Modulating Neuroinflammation, Hippo Signaling and Apoptosis in Rats. Int J Mol Sci 2023, 246. [Google Scholar]
- Saifullah, M.; Mahmood, T.; Ahsan, F.; Bano, S.; Zaidi, S. M. H.; Khan, M. M. U. , Cardioprotective Potential of d-limonene against Isoproterenol induced Myocardial Infarction in Rats. Cell Biochem Biophys 2024. [Google Scholar]
- Victor Antony Santiago, J.; Jayachitra, J.; Shenbagam, M.; Nalini, N. , Dietary d-limonene alleviates insulin resistance and oxidative stress-induced liver injury in high-fat diet and L-NAME-treated rats. Eur J Nutr 2012, 511, 57–68. [Google Scholar]
- Razazi, A.; Kakanezhadi, A.; Raisi, A.; Pedram, B.; Dezfoulian, O.; Davoodi, F. , D-limonene inhibits peritoneal adhesion formation in rats via anti-inflammatory, anti-angiogenic, and antioxidative effects. Inflammopharmacology 2024, 322, 1077–1089. [Google Scholar]
- Kathem, S. H.; Nasrawi, Y. S.; Mutlag, S. H.; Nauli, S. M. , Limonene Exerts Anti-Inflammatory Effect on LPS-Induced Jejunal Injury in Mice by Inhibiting NF-kappaB/AP-1 Pathway. Biomolecules 2024, 143. [Google Scholar]
- Oesch, F.; Fabian, E.; Landsiedel, R. , Xenobiotica-metabolizing enzymes in the lung of experimental animals, man and in human lung models. Arch Toxicol 2019, 9312, 3419–3489. [Google Scholar]
- Heuberger, E.; Ilmberger, J.; Hartter, E.; Buchbauer, G. , Physiological and behavioral effects of 1,8-cineole and (+)-Linalool: Comparison of inhalation and massage aromatherapy. Natural `prod. Comm. 2008; 3, 1103–1110. [Google Scholar]
- Emond, C.; Krishnan, K. , A physiological pharmacokinetic model based on tissue lipid content for simulating inhalation pharmacokinetics of highly lipophilic volatile organic chemicals. Toxicol Mech Methods 2006, 168, 395–403. [Google Scholar]
- de Alvarenga, J. F. R.; Genaro, B.; Costa, B. L.; Purgatto, E.; Manach, C.; Fiamoncini, J. , Monoterpenes: current knowledge on food source, metabolism, and health effects. Crit Rev Food Sci Nutr 2023, 6310, 1352–1389. [Google Scholar]
- Silva, L. K.; Espenship, M. F.; Newman, C. A.; Blount, B. C.; De Jesus, V. R. , Quantification of Seven Terpenes in Human Serum by Headspace Solid-Phase Microextraction-Gas Chromatography-Tandem Mass Spectrometry. Environ Sci Technol 2020, 5421, 13861–13867. [Google Scholar]
- Fang, Y.-W.; Wang, C.; Lin, C.-Y., Biomonitoring serum levels of three monoterpenes and their association with liver function tests in US adults: NHANES 2013-2014. Research Square 2024, 2024, https://www.researchsquare.com/article/rs-4152134/v1.
- Falk-Filipsson, A.; Lof, A.; M. , H.; Hjelm, E. W.; Wang, Z., D-Limonene exposure to humans by inhalation: Uptake, distribution, elimination, and effects on the pulmonary function. J. Tox.Environ. Health 1993, 38, 77–88. [Google Scholar]
- Kohlert, C.; van Rensen, I.; Marz, R.; Schindler, G.; Graefe, E. U.; Veit, M. , Bioavailability and pharmacokinetics of natural volatile terpenes in animals and humans. Planta Med 2000, 666, 495–505. [Google Scholar]
- Schmidt, L.; Goen, T. , R-Limonene metabolism in humans and metabolite kinetics after oral administration. Arch Toxicol 2017, 913, 1175–1185. [Google Scholar]
- Crowell, P. L.; Lin, S.; Vedejs, E.; Gould, M. N. , Identification of metabolites of the antitumor agent d-limonene capable of inhibiting protein isoprenylation and cell growth. Cancer Chemother Pharmacol 1992, 313, 205–12. [Google Scholar]
- Crowell, P. L.; Elson, C. E.; Bailey, H. H.; Elegbede, A.; Haag, J. D.; Gould, M. N. , Human metabolism of the experimental cancer therapeutic agent d-limonene. Cancer Chemother Pharmacol 1994, 351, 31–7. [Google Scholar]
- Shimada, T.; Shindo, M.; Miyazawa, M. , Species differences in the metabolism of (+)- and (-)-limonenes and their metabolites, carveols and carvones, by cytochrome P450 enzymes in liver microsomes of mice, rats, Guinea pigs, rabbits, dogs, monkeys, and humans. Dru Metab. Pharmacokin. 2002, 507–513. [Google Scholar]
- Dinkova-Kostova, A. T.; Massiah, M. A.; Bozak, R. E.; Hicks, R. J.; Talalay, P. , Potency of Michael reaction acceptors as inducers of enzymes that protect against carcinogenesis depends on their reactivity with sulfhydryl groups. Proc Natl Acad Sci U S A 2001, 986, 3404–9. [Google Scholar]
- Liang, S. T.; Chen, C.; Chen, R. X.; Li, R.; Chen, W. L.; Jiang, G. H.; Du, L. L. , Michael acceptor molecules in natural products and their mechanism of action. Front Pharmacol 2022, 13, 1033003. [Google Scholar] [CrossRef] [PubMed]
- Falk, A. A.; Hagberg, M. T.; Lof, A. E.; Wigaeus-Hjelm, E. M.; Wang, Z. P. , Uptake, distribution and elimination of alpha-pinene in man after exposure by inhalation. Scand J Work Environ Health 1990, 165, 372–8. [Google Scholar] [CrossRef] [PubMed]
- Waidyanatha, S.; Hackett, M.; Black, S. R.; Stout, M. D.; Fennell, T. R.; Silinski, M. R.; Watson, S. L.; Licause, J.; Robinson, V. G.; Sparrow, B.; Fernando, R. A.; Cooper, S.; Rider, C. V. , Toxicokinetic evaluation of the common indoor air pollutant, alpha-pinene, and its potential reactive metabolite, alpha-pinene oxide, following inhalation exposure in rodents. Toxicol Appl Pharmacol 2021, 418, 115496. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, L.; Goen, T. , Human metabolism of alpha-pinene and metabolite kinetics after oral administration. Arch Toxicol 2017, 912, 677–687. [Google Scholar] [CrossRef]
- Jin, L.; Xie, Z.; Lorkiewicz, P.; Srivastava, S.; Bhatnagar, A.; Conklin, D. J. , Endothelial-dependent relaxation of alpha-pinene and two metabolites, myrtenol and verbenol, in isolated murine blood vessels. Am J Physiol Heart Circ Physiol 2023, 3256, H1446–H1460. [Google Scholar] [CrossRef]
- Falk, A.; Lof, A.; Hagberg, M.; Hjelm, E. W.; Wang, Z. , Human exposure to 3-carene by inhalation: toxicokinetics, effects on pulmonary function and occurrence of irritative and CNS symptoms. Toxicol Appl Pharmacol 1991, 1102, 198–205. [Google Scholar] [CrossRef]
- Schmidt, L.; Belov, V. N.; Goen, T. , Human metabolism of Delta3-carene and renal elimination of Delta3-caren-10-carboxylic acid (chaminic acid) after oral administration. Arch Toxicol 2015, 893, 381–92. [Google Scholar] [CrossRef]
- Duisken, M.; Benz, D.; Peiffer, T. H.; Blomeke, B.; Hollender, J. , Metabolism of Delta(3)-carene by human cytochrome p450 enzymes: identification and characterization of two new metabolites. Curr Drug Metab 2005, 66, 593–601. [Google Scholar] [CrossRef]
- Jāger, K.; Našel, C.; Binder, R.; Stimpfl, T.; Vycudilik, W.; Buchbauer, G. , Pharmaokinetic studies of the frangrance compound 1,8-cineole in humans during inhalation. Chem. Senses 1996, 21, 477–480. [Google Scholar] [CrossRef]
- Friedl, S. M.; Heuberger, E.; Oedendorfer, K.; Kitzer, S.; Jaganjac, L.; Stappen, I.; Reznicek, G. , Quantitification of 1,8-cineole in himan blood and plasma and the impact of liner choice in heaf-space chromatography. Ciurr. Bioactive Compounds 2015, 11, 49–55. [Google Scholar] [CrossRef]
- Horst, K.; Rychlik, M. , Quantification of 1,8-cineole and of its metabolites in humans using stable isotope dilution assays. Mol Nutr Food Res 2010, 5410, 1515–29. [Google Scholar]
- Kirsch, F.; Horst, K.; Röhrig, W.; Rychlik, M.; Buettner, A. , Tracing metabolite profiles in human milk: studies on the odorant 1,8-cineole transferred into breast milk after oral intake. Metabolomics 2013, 9, 483–486. [Google Scholar] [CrossRef]
- Kirsch, F.; Buettner, A. , Characterisation of the metabolites of 1,8-cineole transferred into human milk: concentrations and ratio of enantiomers. Metabolites 2013, 31, 47–71. [Google Scholar] [CrossRef] [PubMed]
- Altschul, R. , Endothelium. The MacMillan Co.,: New York, 1954; p 157.
- Huertas, A.; Guignabert, C.; Barbera, J. A.; Bartsch, P.; Bhattacharya, J.; Bhattacharya, S.; Bonsignore, M. R.; Dewachter, L.; Dinh-Xuan, A. T.; Dorfmuller, P.; Gladwin, M. T.; Humbert, M.; Kotsimbos, T.; Vassilakopoulos, T.; Sanchez, O.; Savale, L.; Testa, U.; Wilkins, M. R. , Pulmonary vascular endothelium: the orchestra conductor in respiratory diseases: Highlights from basic research to therapy. Eur Respir J 2018, 514. [Google Scholar]
- Pi, X.; Xie, L.; Patterson, C. , Emerging Roles of Vascular Endothelium in Metabolic Homeostasis. Circ Res 2018, 1234, 477–494. [Google Scholar]
- Augustin, H. G.; Koh, G. Y. , A systems view of the vascular endothelium in health and disease. Cell 2024, 18718, 4833–4858. [Google Scholar]
- Hirota, R.; Nakamura, H.; Bhatti, S. A.; Ngatu, N. R.; Muzembo, B. A.; Dumavibhat, N.; Eitoku, M.; Sawamura, M.; Suganuma, N. , Limonene inhalation reduces allergic airway inflammation in Dermatophagoides farinae-treated mice. Inhal Toxicol 2012, 246, 373–81. [Google Scholar]
- Dudek, T.; Marc, M.; Zabiegala, B. , Chemical Composition of Atmospheric Air in Nemoral Scots Pine Forests and Submountainous Beech Forests: The Potential Region for the Introduction of Forest Therapy. Int J Environ Res Public Health 2022, 1923. [Google Scholar] [CrossRef]
- Fukuoka, Y.; Iihoshi, M.; Nazunin, J. T.; Abe, D.; Fukuba, Y. , Dynamic Characteristics of Ventilatory and Gas Exchange during Sinusoidal Walking in Humans. PLoS One 2017, 121, e0168517. [Google Scholar]
- Santana, H. S. R.; de Carvalho, F. O.; dos Santos, D. M.; da Silva, E. A. P.; Silva, E. R.; Shanmugam, S.; Heimfarth, L.; Nunes, P. S.; de Oliveira e Silva, A. M.; de Souza Araújo, A. A.; de Albuquerque, R. L. C.; dos Santos, M. R. V. , Inhaled D-limonene minimizes acute lung injury and reduces oxidative stress induced by smoke in rats. Phytomed. Plus 2022, 23, 100308. [Google Scholar]
- Patel, M.; Narke, D.; Kurade, M.; Frey, K. M.; Rajalingam, S.; Siddiquee, A.; Mustafa, S. J.; Ledent, C.; Ponnoth, D. S. , Limonene-induced activation of A(2A) adenosine receptors reduces airway inflammation and reactivity in a mouse model of asthma. Purinergic Signal 2020, 163, 415–426. [Google Scholar]
- Nam, S. Y.; Chung, C. K.; Seo, J. H.; Rah, S. Y.; Kim, H. M.; Jeong, H. J. , The therapeutic efficacy of alpha-pinene in an experimental mouse model of allergic rhinitis. Int Immunopharmacol 2014, 231, 273–82. [Google Scholar]
- Christman, J. W.; Blackwell, T. S.; Juurlink, B. H. , Redox regulation of nuclear factor kappa B: therapeutic potential for attenuating inflammatory responses. Brain Pathol 2000, 101, 153–62. [Google Scholar]
- Guo, Q.; Jin, Y.; Chen, X. X.; Ye, X.; Shen, X.; Lin, M.; Zeng, C.; Zhou, T.; Zhang, J. , NF kappa B in biology and targetted therapy: New insights and translational implications. Signal Transduct. Target. Therap. 2024, 9, 53. [Google Scholar]
- Fairchild, G. A. , Measurement of respiratory volume for virus retention studies in mice. Appl Microbiol 1972, 245, 812–8. [Google Scholar]
- Bejeshk, M. A.; Aminizadeh, A. H.; Jafari, E.; Motamed, S.; Zangiabadi, I.; Ghasemi, A.; Fathi, M.; Nexhadi, A.; Akhgarandouz, F.; Bejeshk, F.; Mohamedi, L.; Mohamedi, F.; Rajizadeh, M. A. , Myrtenol ameliorates recognition memories’ impairment and anxiety-like behaviors induced by asthma mitigating hippocampal inflammation and oxidative stress in rats. Neuroimmunomod. 2023, 30, 42–54. [Google Scholar]
- Kim, K.; Kim, Y.; Lee, J.; Lee, M.; Ahn, C.; Park, M. J.; Na, H.; Jeung, E. B. , 3-carene supresses inflammatory cytokine interleukin-4, interleukin-5 and interleukin-13 in a murine model of asthma. J Physiol Pharmacol 2024, 752, 195–203. [Google Scholar]
- Hov, Ø.; Schjoldager, J.; Wathne, M. , Measurement and modeling of the concentrations of terpenes in coniferous forest air. J. Geophys. Res. 1983, 88, 10679–10688. [Google Scholar]
- Bastos, V. P.; Gomes, A. S.; Lima, F. J.; Brito, T. S.; Soares, P. M.; Pinho, J. P.; Silva, C. S.; Santos, A. A.; Souza, M. H.; Magalhaes, P. J. , Inhaled 1,8-cineole reduces inflammatory parameters in airways of ovalbumin-challenged Guinea pigs. Basic Clin Pharmacol Toxicol 2011, 1081, 34–9. [Google Scholar]
- Guenther, A.; Zimmerman, P.; Wildermuth, M. , Natural volatile compound emission rate estimates for US woodland landscapes. Atmos. Environ. 1994, 28, 1197–1210. [Google Scholar]
- Hakola, H.; Laurila, T.; Rinne, J.; Puhto, K. , The ambient concentrations of biogenic hydrocarbons at a northern European, boreal site. Atmos. Environ. 2000, 34, 4971–4982. [Google Scholar] [CrossRef]
- Kennedy-Feitosa, E.; Okuro, R. T.; Pinho Ribeiro, V.; Lanzetti, M.; Barroso, M. V.; Zin, W. A.; Porto, L. C.; Brito-Gitirana, L.; Valenca, S. S. , Eucalyptol attenuates cigarette smoke-induced acute lung inflammation and oxidative stress in the mouse. Pulm Pharmacol Ther 2016, 41, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liu, X.; Zheng, Y.; Rao, R.; Li, W. , Protective effects of 1,8-cineole (eucalyptol) on respiratory system: A systemic review and meta-analysis from animal studies. Pharmacon. Mag. 2020, 37, 1737–1753. [Google Scholar] [CrossRef]
- Worth, H.; Dethlefsen, U. , Patients with asthma benefit from concomitant therapy with cineole: a placebo-controlled, double-blind trial. J Asthma 2012, 498, 849–53. [Google Scholar] [CrossRef]
- Juergens, L. J.; Racké, K.; Tuleta, I.; Stoeber, M.; Juergens, U. W. , Anti-inflammatory effects of 1,8-cineole (eucalyptol) improve glucocorticoid effects in vitro: A novel approach of steroid-sparing dd-on therapy for COPD and asthma. Synergy 2017, 5, 1–8. [Google Scholar] [CrossRef]
- Juergens, L. J.; Worth, H.; Juergens, U. R. , New Perspectives for mucolytic, anti-inflammatory and adjunctive therapy with 1,8-Cineole in COPD and asthma: Review on the new therapeutic approach. Adv Ther 2020, 375, 1737–1753. [Google Scholar] [CrossRef]
- Donelli, D.; Meneguzzo, F.; Antonelli, M.; Aedissino, D.; Niccoli, G.; Gronchi, G.; Baraldi, R.; Neri, L.; Zabini, F. , Effects of plant-emitted monoterpenes on anxiety symptoms: A propensity-matched obsevational cohort study. J. Environ. Res. Oublic Health 2023, 20, 2773. [Google Scholar] [CrossRef]
- Kim, J.-C.; Dinh, T.-V.; Oh, H.-K.; Son, Y.-S.; Ahn, J.-W.; Song, K.-Y.; Choi, I.-Y.; Park, C.-R.; Szulejko, J. E.; Kim, J.-H. , The potential benefits of therapeutic treatment using gaseous terpenes at ambient low levels. Appl. Sci. 2019, 9, 4507. [Google Scholar] [CrossRef]
- Našel, C.; Naše, B.; Samec, P.; Schindler, E.; Buchbauer, G. , Functional imaging effects of fragrances on the human brain after prolonged inhalation. Chem. Senses 1994, 19, 350–364. [Google Scholar] [CrossRef]
- Lima, N. G.; De Sousa, D. P.; Pimenta, F. C.; Alves, M. F.; De Souza, F. S.; Macedo, R. O.; Cardoso, R. B.; de Morais, L. C.; Melo Diniz Mde, F.; de Almeida, R. N. , Anxiolytic-like activity and GC-MS analysis of (R)-(+)-limonene fragrance, a natural compound found in foods and plants. Pharmacol Biochem Behav 2013, 1033, 450–4. [Google Scholar]
- Witkin, J. M.; Barrett, J. E. , ANXIOLYTICS: Origins, drug discovery, and mechanisms. Pharmacol Biochem Behav 2024, 245, 173858. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Seo, S.; Lamichhane, S.; Seo, J.; Hong, J. T.; Cha, H. J.; Yun, J. , Limonene has anti-anxiety activity via adenosine A2A receptor-mediated regulation of dopaminergic and GABAergic neuronal function in the striatum. Phytomedicine 2021, 83, 153474. [Google Scholar] [CrossRef] [PubMed]
- Saeedi, R.-R. M. , Anti-anxiety effect of alpha-pinene in comparison with Diazepam in adult male rats. J. Kasham Un. Med. Sci. 2020, 24, 245–253. [Google Scholar]
- Moreira, M. R.; Salvadori, M. G.; de Almeida, A. A.; de Sousa, D. P.; Jordan, J.; Satyal, P.; de Freitas, R. M.; de Almeida, R. N. , Anxiolytic-like effects and mechanism of (-)-myrtenol: a monoterpene alcohol. Neurosci Lett 2014, 579, 119–24. [Google Scholar] [CrossRef]
- Woo, J.; Yang, H.; Yoon, M.; Gadhe, C. G.; Pae, A. N.; Cho, S.; Lee, C. J. , 3-Carene, a Phytoncide from Pine Tree Has a Sleep-enhancing Effect by Targeting the GABA(A)-benzodiazepine Receptors. Exp Neurobiol 2019, 285, 593–601. [Google Scholar] [CrossRef]
- Ceremuga, T. E.; McClellan, C. B.; Green, X. C.; Heber, B. E.; Jolly, M. L.; Malone, T. B.; Schaaf, J. L.; Isaacs, A. P. , Investigation of the Anxiolytic and Antidepressant Effects of Eucalyptol (1,8-Cineole), a Compound From Eucalyptus, in the Adult Male Sprague-Dawley Rat. AANA J 2017, 854, 277–284. [Google Scholar]
- Dougnon, G.; Ito, M. , Inhalation Administration of the Bicyclic Ethers 1,8- and 1,4-cineole Prevent Anxiety and Depressive-Like Behaviours in Mice. Molecules 2020, 258. [Google Scholar] [CrossRef]
- Kim, K. Y.; Seo, H. J.; Min, S. S.; Park, M.; Seol, G. H. , The effect of 1,8-cineole inhalation on preoperative anxiety: a randomized clinical trial. Evid Based Complement Alternat Med 2014, 2014, 820126. [Google Scholar] [CrossRef]
- Griffen, S.; Wyllie, S. G.; Markham, J. , Determination of octanol-water partition coefficients for terpenoids using reversed-phase high-performance liquid chromatography. J. Chromatogr. A 1999, 864, 221–228. [Google Scholar] [CrossRef]
- Alcorn, J. , The xenobiotic elimination system: An overview. Nova Publishers: Broccoli: Cultivation, Nutritional Properties and Effects on Health, 2016; p 342.
- Prescott, J. A.; Mitchell, J. P.; Cook, S. J. , Inhibitory feed-back control of NF-kB signalling in health and disease. Biochem. J. 2021, 478, 2619–2664. [Google Scholar] [CrossRef]
- Bshesh, K.; Zhao, B.; Spight, D.; Biaggioni, I.; Feokistov, I.; Denenberg, A.; Wong, H. R.; Shanley, T. P. , The A2A receptor mediates an endogenous regulatory pathway of cytokine expression in THP-1 cells. J Leukoc Biol 2002, 725, 1027–36. [Google Scholar]
- Liu, W.; Li, Q.; Fng, W.; Cai, L.; Wang, Z.; Kou, B.; Zhou, C.; Zhou, Y.; Yao, Z.; Wei, M.; Zhang, S. , A2AR regulate inflammation through PKA/NF-kB signaling pathways in invertebral disc degeneration. Eur. J. Med. Res. 2024, 29, 13. [Google Scholar]
- Sands, W. A.; Martin, A. F.; Strong, E. W.; Palmer, T. M. , Specific inhibition of nuclear factor-kappaB-dependent inflammatory responses by cell type-specific mechanisms upon A2A adenosine receptor gene transfer. Mol Pharmacol 2004, 665, 1147–59. [Google Scholar]
- Koroskenyi, K.; Kiss, B.; Szondy, Z. , Adenosine A2A receptor signaling attenuates LPS-induced pro-inflammatory cytokine formation of mouse macrophages by inducing the expression of DUSP1. Biochim Biophys Acta 2016, (7 Pt A), 1461–71. [Google Scholar]
- Quintans, J. S. S.; Shanmugam, S.; Heimfarth, L.; Araujo, A. A. S.; Almeida, J.; Picot, L.; Quintans-Junior, L. J. , Monoterpenes modulating cytokines - A review. Food Chem Toxicol 2019, 123, 233–257. [Google Scholar]
- Salminen, A.; Lehtonen, M.; Suuronen, T.; Kaarniranta, K.; Huuskonen, J. , Terpenoids: natural inhibitors of NF-kappaB signaling with anti-inflammatory and anticancer potential. Cell Mol Life Sci 2008, 6519, 2979–99. [Google Scholar]
- Yarom, M.; Tang, X. W.; Wu, E.; Carlson, R. G.; Vander Velde, D.; Lee, X.; Wu, J. , Identification of inosine as an endogenous modulator for the benzodiazepine binding site of the GABAA receptors. J Biomed Sci 1998, 54, 274–80. [Google Scholar]
- Mortensen, M.; Bright, D. P.; Fagotti, J.; Dorovykh, V.; Cerna, B.; Smart, T. G. , Forty Years Searching for Neurosteroid Binding Sites on GABA(A) Receptors. Neuroscience 2024. [Google Scholar]
- Bakas, T.; van Nieuwenhuijzen, P. S.; Devenish, S. O.; McGregor, I. S.; Arnold, J. C.; Chebib, M. , The direct actions of cannabidiol and 2-arachidonoyl glycerol at GABA(A) receptors. Pharmacol Res 2017, 119, 358–370. [Google Scholar]
- Hajagos-Tóth, J.; Hódi, A.; Seres, A. B.; Gáspár, R. , Effects of d- and l-limonene on the pregrnant rat myometrium in vivo. Coat. Med. J. 2015, 56, 431–438. [Google Scholar]
- Park, H. M.; Lee, J. H.; Yaoyao, J.; Jun, H. J.; Lee, S. J. , Limonene, a natural cyclic terpene, is an agonistic ligand for adenosine A(2A) receptors. Biochem Biophys Res Commun 2011, 4041, 345–8. [Google Scholar]
- Felger, J. C. , Imaging the Role of Inflammation in Mood and Anxiety-related Disorders. Curr Neuropharmacol 2018, 165, 533–558. [Google Scholar]
- Vogelzangs, N.; Beekman, A. T.; de Jonge, P.; Penninx, B. W. , Anxiety disorders and inflammation in a large adult cohort. Transl Psychiatry 2013, 34, e249. [Google Scholar]
- Milaneschi, Y.; Kappelmann, N.; Ye, Z.; Lamers, F.; Moser, S.; Jones, P. B.; Burgess, S.; Penninx, B.; Khandaker, G. M. , Association of inflammation with depression and anxiety: evidence for symptom-specificity and potential causality from UK Biobank and NESDA cohorts. Mol Psychiatry 2021, 2612, 7393–7402. [Google Scholar]
- Kunnumakkara, A. B.; Shabnam, B.; Girisa, S.; Harsha, C.; Banik, K.; Devi, T. B.; Choudhury, R.; Sahu, H.; Parama, D.; Sailo, B. L.; Thakur, K. K.; Gupta, S. C.; Aggarwal, B. B. , Inflammation, NF-kappaB, and Chronic Diseases: How are They Linked? Crit Rev Immunol 2020, 401, 1–39. [Google Scholar]
- Rusnak, F.; Reiter, T. , Sensing electrons: protein phosphatase redox regulation. Trends Biochem Sci 2000, 2511, 527–9. [Google Scholar]
- Hunter, T. , Protein kinases and phosphatases: the yin and yang of protein phosphorylation and signaling. Cell 1995, 802, 225–36. [Google Scholar]
- Sadeghinejad, L.; Noyan, H.; Juurlink, B. H. J. , Cellular Redox: Aging and Diet. In Broccoli: Cultivation, Nutritional Properties and Effects on Health, Juurlink, B. H. J., Ed. Nova Publishers: New York, 2016; pp 87-110.
- Saha, S.; Buttari, B.; Panieri, E.; Profumo, E.; Saso, L. , An Overview of Nrf2 Signaling Pathway and Its Role in Inflammation. Molecules 2020, 2522. [Google Scholar]
- Chiurchiu, V.; Maccarrone, M. , Chronic inflammatory disorders and their redox control: from molecular mechanisms to therapeutic opportunities. Antioxid Redox Signal 2011, 159, 2605–41. [Google Scholar]
- Juurlink, B. H. J. , The Nrf2 Signalling System. In Broccoli: Cultivation, Nutritional Properties and Effects on Health, Juurlink, B. H. J., Ed. Nova Publishers: New York, 2016; Vol. 1, pp 111-119.
- Ma, Q. , Role of Nrf2 in oxidative stress and toxicity. Annu. Rev. Pharmacol. Toxicol. 2015, 53, 401–426. [Google Scholar]
- Talalay, P.; Fahey, J. W.; Holtzclaw, W. D.; Prestera, T.; Zhang, Y. , Chemoprotection against cancer by phase 2 enzyme induction. Toxicol Lett 1995, 82-83, 173–9. [Google Scholar] [PubMed]
- Fahey, J. W.; Shapiro, T. A. , In Memoriam. Inimitable Paul Talalay (1923-2019). Trends Pharmacol. 2019, 40, 359–361. [Google Scholar]
- Johnson, T. L. , Paul Talalaty: The Catalyst. In Broccoli: Cultivation, Nutritional `properties and Effects on Health, Juurlink, B. H. J., Ed. Nova Publishers: New York, 2016; Vol. 1, pp 1-8.
- Wild, A. C.; Mulcahy, R. T. , Regulation of gamma-glutamylcysteine synthetase subunit gene expression: insights into transcriptional control of antioxidant defenses. Free Radic Res 2000, 324, 281–301. [Google Scholar]
- Juurlink, B. H. , Therapeutic potential of dietary phase 2 enzyme inducers in ameliorating diseases that have an underlying inflammatory component. Can J Physiol Pharmacol 2001, 793, 266–82. [Google Scholar]
- Mohamed, A. A.; Avila, J. G.; Schultke, E.; Kamencic, H.; Skihar, V.; Obayan, A.; Juurlink, B. H. , Amelioration of experimental allergic encephalitis (EAE) through phase 2 enzyme induction. Biomed Sci Instrum 2002, 38, 9–13. [Google Scholar]
- Wu, L.; Noyan Ashraf, M. H.; Facci, M.; Wang, R.; Paterson, P. G.; Ferrie, A.; Juurlink, B. H. , Dietary approach to attenuate oxidative stress, hypertension, and inflammation in the cardiovascular system. Proc Natl Acad Sci U S A 2004, 10118, 7094–9. [Google Scholar]
- Noyan-Ashraf, M. H.; Sadeghinejad, Z.; Juurlink, B. H. , Dietary approach to decrease aging-related CNS inflammation. Nutr Neurosci 2005, 82, 101–10. [Google Scholar]
- Noyan-Ashraf, M. H.; Wu, L.; Wang, R.; Juurlink, B. H. , Dietary approaches to positively influence fetal determinants of adult health. FASEB J 2006, 202, 371–3. [Google Scholar]
- Noyan-Ashraf, M. H.; Sadeghinejad, Z.; Davies, G. F.; Ross, A. R.; Saucier, D.; Harkness, T. A.; Juurlink, B. H. , Phase 2 protein inducers in the diet promote healthier aging. J Gerontol A Biol Sci Med Sci 2008, 6311, 1168–76. [Google Scholar]
- Senanayake, G. V.; Banigesh, A.; Wu, L.; Lee, P.; Juurlink, B. H. , The dietary phase 2 protein inducer sulforaphane can normalize the kidney epigenome and improve blood pressure in hypertensive rats. Am J Hypertens 2012, 252, 229–35. [Google Scholar]
- Black, A. M.; Armstrong, E. A.; Scott, O.; Juurlink, B. J. H.; Yager, J. Y. , Broccoli sprout supplementation during pregnancy prevents brain injury in the newborn rat following placental insufficiency. Behav Brain Res 2015, 291, 289–298. [Google Scholar] [PubMed]
- Dinkova-Kostova, A. T.; Copple, I. M. , Advances and challenges in therapeutic targeting of Nrf2. Trends Pharmacol. Sci. 2022, 44, 137–149. [Google Scholar]
- Kumar, K. J. S.; Vani, M. G.; Wang, S. Y. , Limonene protects human skin keratinocytes against UVB-induced photodamage and photoaging by activating the Nrf2-dependent antioxidant defense system. Environ Toxicol 2022, 3712, 2897–2909. [Google Scholar]
- Fang, Y.; Zheng, Y.; Gao, Q.; Pang, M.; Wu, Y.; Feng, X.; Tao, X.; Hu, Y.; Lin, Z.; Lin, W. , Activation of the Nrf2/Keap1 signaling pathway mediates the neuroprotective effect of Perillyl alcohol against cerebral hypoxic-ischemic damage in neonatal rats. Redox Rep 2024, 291, 2394714. [Google Scholar]
- Alvi, A. M.; Al Kury, L. T.; Alattar, A.; Ullah, I.; Muhammad, A. J.; Alshaman, R.; Shah, F. A.; Khan, A. U.; Feng, J.; Li, S. , Carveol Attenuates Seizure Severity and Neuroinflammation in Pentylenetetrazole-Kindled Epileptic Rats by Regulating the Nrf2 Signaling Pathway. Oxid Med Cell Longev 2021, 2021, 9966663. [Google Scholar]
- Rahman, Z. U.; Al Kury, L. T.; Alattar, A.; Tan, Z.; Alshaman, R.; Malik, I.; Badshah, H.; Uddin, Z.; Khan Khalil, A. A.; Muhammad, N.; Khan, S.; Ali, A.; Shah, F. A.; Li, J. B.; Li, S. , Carveol a Naturally-Derived Potent and Emerging Nrf2 Activator Protects Against Acetaminophen-Induced Hepatotoxicity. Front Pharmacol 2020, 11, 621538. [Google Scholar]
- Porres-Martinez, M.; Gonzalez-Burgos, E.; Carretero, M. E.; Gomez-Serranillos, M. P. , In vitro neuroprotective potential of the monoterpenes alpha-pinene and 1,8-cineole against H2O2-induced oxidative stress in PC12 cells. Z Naturforsch C J Biosci 2016, 71, 191–9. [Google Scholar]
- Bejeshk, M. A.; Najafipour, H.; Khaksari, M.; Nematollahi, M. H.; Rajizadeh, M. A.; Dehesh, T.; Bagheri, F.; Sepehri, G. , Myrtenol-loaded niosomes can prevent lung ischemia-reperfusion injury model in rats by balancing the Nrf2/Keap1 and NF-kappaB signaling pathways. Naunyn Schmiedebergs Arch Pharmacol 2025. [Google Scholar]
- Liu, Z.; Wang, J.; Jin, X.; Gao, P.; Zhao, Y.; Yin, M.; Ma, X.; Xin, Z.; Zhao, Y.; Zhou, X.; Gao, W. , 1,8-Cineole Alleviates OGD/R-Induced Oxidative Damage and Restores Mitochondrial Function by Promoting the Nrf2 Pathway. Biol Pharm Bull 2023, 4610, 1371–1384. [Google Scholar]
- Kamencic, H.; Lyon, A.; Paterson, P. G.; Juurlink, B. H. , Monochlorobimane fluorometric method to measure tissue glutathione. Anal Biochem 2000, 2861, 35–7. [Google Scholar]
- Vari, R.; D'Archivio, M.; Filesi, C.; Carotenuto, S.; Scazzocchio, B.; Santangelo, C.; Giovannini, C.; Masella, R. , Protocatechuic acid induces antioxidant/detoxifying enzyme expression through JNK-mediated Nrf2 activation in murine macrophages. J Nutr Biochem 2011, 225, 409–17. [Google Scholar]
- Dinkova-Kostova, A. T.; Holtzclaw, W. D.; Cole, R. N.; Itoh, K.; Wakabayashi, N.; Katoh, Y.; Yamamoto, M.; Talalay, P. , Direct evidence that sulfhydryl groups of Keap1 are the sensors regulating induction of phase 2 enzymes that protect against carcinogens and oxidants. Proc Natl Acad Sci U S A 2002, 9918, 11908–13. [Google Scholar]
- Bai, C.; Ma, Q.; Li, Q.; Yu, L.; Zhen, D.; Liu, M.; Wei, C. , Combination of 1,8-cineole and beta-caryophyllene synergistically reverses cardiac hypertrophy in isoprenaline-induced mice and H9c2 cells. Bioorg Chem 2022, 124, 105823. [Google Scholar]
- Kardos, P.; Khaletskaya, O.; Kropova, O. , Efficacy and safety of cineole (Soledum) in the treatment of patients with acute bronchitis: Results of an open-label randomized clinical phase III study) Clin. Phytosci. 2021, 7, 83. [Google Scholar] [CrossRef]

Table 1.
Terpenoids, including the 4 monoterpenes and some of their metabolites considered in this paper. Chemical structures obtained either from Chemspider or Phytohub.
Table 1.
Terpenoids, including the 4 monoterpenes and some of their metabolites considered in this paper. Chemical structures obtained either from Chemspider or Phytohub.
| Terpene | Parent Compound | Metabolite | Metabolite |
|---|---|---|---|
| Isoprene 63.12 daltons |
 |
||
| Monoterpene D-Limonene 136.238 daltons |
 |
 Carveol 152.237 daltons |
 Perillic acid 166.22 daltons |
| Monoterpene a-Pinene 136.238 daltons |
 |
 Myrtenol 152.237 daltons |
 Myrtenic acid 166.220 daltons |
| Monoterpene D3-Carene 136.238 daltons |
 |
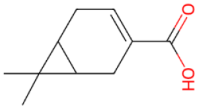 D3-caren-10-carboxylic acid 180.247 daltons |
 D3-Carene epoxide D3-Carene epoxide152.237 daltons |
| Monoterpene 1,8-Cineol 154.252 daltons |
 |
 2-hydroxy1,8-cineole |
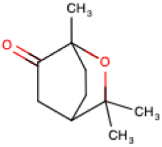 2-oxo-1,8-cineole |
| Sesquiterpene b-Caryophyllene 204.357 daltons |
 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



