Submitted:
14 March 2025
Posted:
17 March 2025
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
SARS-CoV-2 has demonstrated a remarkable capacity for immune evasion. While initial studies focused on the Wuhan variant and adaptive immunity, later emerging strains such as Omicron exhibit mutations that may alter their immune-modulatory properties.
We performed a comprehensive review of immune evasion mechanisms associated with SARS-CoV-2 viral proteins, to focus on the evolutionary dynamics of immune modulation. We systematically analyzed and compared the impact of all currently known Wuhan and Omicron SARS-CoV-2 proteins on type I interferon (IFN) responses using a dual-luciferase reporter assay carrying an interferon-inducible promoter. Results revealed that Nsp1, Nsp5, Nsp14, and ORF6 are potent type I IFN inhibitors conserved across Wuhan and Omicron strains. Notably, we identified strain-specific differences, with Nsp6 and Spike protein exhibiting enhanced IFN suppression in Omi-cron, whereas the Envelope protein largely retained this function.
To extend these findings, we investigated selected proteins in primary human endothelial cells and also observed strain-specific differences in immune response with higher type I IFN response in cells expressing the Wuhan strain variant, suggesting that Omicron's adaptational mutations may contribute to a damped type I IFN re-sponse in the course of the pandemic's trajectory.
Keywords:
SARS-CoV-2
; interferon
; luciferase
; immune evasion
; endothelial dysfunction
; HUVEC
1. Introduction
Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) emerged in late 2019, triggering a global pandemic that profoundly impacted societies, economies, and healthcare systems. Compared to other RNA viruses coronaviruses like SARS-CoV-2 has a notably larger genome (30 kb) [1] that not only encodes essential replication machinery, but also a diverse set of accessory proteins that actively counteract host immune defenses (Table 1).
Early research prioritized adaptive immune responses [2,3,4], particularly antibody production and T cell-mediated immunity, due to the urgent need for a vaccine and therapeutic interventions. However, as our understanding of the virus evolved, increasing attention has been given to innate immune sensing pathways and their role in shaping disease outcomes. SARS-CoV-2’s ability to evade innate immunity facilitates viral replication and dissemination while contributing to severe clinical manifestations, including delayed immune responses, excessive inflammation, and endothelial dysfunction [5,6].
Despite the wealth of knowledge gained over the past few years, much of the research on SARS-CoV-2 immune evasion has been conducted using the original Wuhan strain and limited cellular models. There remains a critical gap in understanding how different SARS-CoV-2 strain proteins manipulate immune responses in endothelial cells specifically. Given the vascular complications associated with COVID-19, a detailed analysis of immune evasion mechanisms in endothelial cells is appropriate.
In this study, we conducted a comprehensive review to guide our global screening assay assessing the impact of all known SARS-CoV-2 proteins from both the Wuhan and Omicron variant on IFN induction in cell lines. Next, we selected proteins from both variants to express in primary endothelial cells to assess their effect on IFN type I response.acid-inducible gene I (RIG-I)-like receptors detect viral components like RNA species and activate downstream signaling cascades [7]. These pathways converge on mitochondrial antiviral-signaling protein (MAVS), TANK-binding kinase 1 (TBK1), and interferon regulatory factors (IRF3 and IRF7), leading to the production of type I interferons (IFN-I), such as IFN-α and IFN-β [8]. IFN-I responses initiate antiviral states in infected and neighboring cells. However, SARS-CoV-2 has evolved multiple strategies to evade IFN responses and sustain efficient replication (Figure 1). Approximately 0.5 to 1% of infected individuals succumb to COVID-19. Unlike other respiratory viruses, the SARS-CoV-2 is distinguished by its severe vascular implications [5]. In COVID-19, endothelial dysfunction and vascular inflammation are associated with thrombo-inflammatory complications, such as increased vascular permeability, thrombosis, and systemic inflammation, often referred to as the “endothelitis” observed in severe cases [9,10,11], as well as a hypercoagulable state [12] that can lead to fatal organ damage (Figure 2). While SARS-CoV-2 primarily infects respiratory epithelial cells, it has become increasingly clear that endothelial cells play a crucial role in in COVID-19 pathogenesis [13,14]. Endothelial cells (EC), which line the blood and lymphatic vessels, are critical regulators of vascular homeostasis. SARS-CoV-2 infects endothelial cells [15], initiating a cascade of events that impair EC function and exacerbate systemic inflammation.
2. Materials and Methods
2.1. Cells
HEK293T were cultivated in Iscove’s Modified Dulbecco’s Medium (IMDM) (Thermo Fisher Scientific, Merelbeke, Belgium) supplemented with 10% (v/v) heat inactivated fetal calf serum (FCS, Hyclone, Thermo Fisher Scientific), 2 mM L-glutamine (Thermo Fisher Scientific), 100 U/mL penicillin and 100 μg/mL streptomycin (Thermo Fisher Scientific).
Human umbilical cords were donated by mothers from the maternity ward after informed consent, and approved by the Ghent University Hospital ethical committee (ONZ-2022-0272). HUVEC cells were freshly isolated as previously described [178]. They were cultured in Human Large Vessel Endothelial Cell Basal Medium (formerly Medium 200) (Thermo Fisher Scientific) supplemented with Large Vessel Endothelial Supplement (LVES) (Thermo Fisher Scientific), 2 mM L-glutamine (Thermo Fisher Scientific), 100 U/mL penicillin and 100 μg/mL streptomycin (Thermo Fisher Scientific). Before cell plating, six-well plates were freshly coated with 5 μg/mL fibronectin solution (fibronectin powder, Sigma-Aldrich, Diegem, Belgium) by incubating them for 30 minutes at room temperature, followed by aspiration of the solution before cell plating. The isolated HUVECs were plated (3,5 x 105 cells/well) and kept in culture on fibronectin-coated plates for a week and passaged twice at a seeding ratio 1:2, before being used for subsequent experiments.
2.2. Expression Constructs
Codon-optimized open reading frames and proteolytically mature nonstructural proteins (Nsps) of all 29 viral proteins from SARS-CoV-2 Wuhan and Omicron BA.1 strain were inserted into the pLVX-EF1alpha-IRES-Puro expression vector, with a Kozak sequence before each start codon and a 2x-Strep tag with a linker at either the N- or C-terminus. The only exception was ORF6, which was cloned from the Omicron BA.2 strain due to the presence of an additional D61L mutation unique to this variant. Additionally, Nsp3 from the Wuhan strain was not cloned due to technical limitations. These expression constructs were kindly provided by Dr Nevan Krogan [179,180].
2.3. Dual Luciferase Reporter Assay
The assay was optimized as described previously [177]. HEK293T cells seeded in 48-well plates (0.5 x 104 cells/well) were co-transfected with 100 ng of reporter plasmids carrying an ISRE-promoter driving the Firefly luciferase activity (pISRE-F-luc), 20 ng of a pGAPDH-R-luc plasmid carrying GAPDH promoter driving Renilla luciferase activity as control of transfection and 250 ng of a viral SARS-CoV-2 protein expressing plasmid. For the latter, strep II-tagged expression constructs encoding the 30 currently known SARS-CoV-2 proteins were used (Nsp1, Nsp2, Nsp3, Nsp4, Nsp5, Nsp6, Nsp7, Nsp8, Nsp9, Nsp10, Nsp11, Nsp12, Nsp13, Nsp14, Nsp15, Nsp16, S, ORF3a, ORF3c, E, M, ORF6, ORF7a, ORF7b, ORF8, ORF9b, N, ORF9c, and ORF10), i.e., variants from the original Wuhan strain and from Omicron BA.1 (kindly provided by Dr Nevan Krogan) strain were included. FuGENE HD (Promega) was used for all of the transfections per manufacturer’s instructions. Cells were treated with 1000 U/mL IFN-β or 500 U/mL IFN-α2 and harvested and lysed 8 or 24 hours post-stimulation. Cell lysates were assayed for luciferase activity using the Dual-Glo luciferase assay system and GloMax® Explorer microplate Luminometer (Promega). Firefly luciferase activity was normalized to cell viability and to Renilla activity to account for transfection efficiency. Results were expressed as remaining percentage activity compared to fully stimulated ISRE promoter which was co-transfected with an empty control vector not carrying a viral protein. At least 3 biological replicates were included.
2.4. Cell Viability Assay
To measure metabolic activity affected by the individual SARS-CoV-2 viral proteins, transfected cells were lysed in passive lysis buffer at 8 h and 24 h post-transfection and analyzed using the CellTiter-Glo Luminescent Cell Viability Assay according to manufacturer’s instructions and GloMax® Explorer microplate Luminometer (Promega, Leiden, The Netherlands).
2.5. Production of lentiviruses Encoding Single SARS-CoV-2 Viral Proteins
Lentiviruses were produced using the Strep II-tagged expression constructs encoding the ORF9b, the Membrane and the Envelope protein both from the Wuhan and the Omicron BA.1 strain. The lentiviral titer was measured by quantification of reverse transcriptase activity (RT) via real-time PCR [181]. HEK293T cells were also transduced and stained after permeabilization with primary anti-Strep antibody (NWSHPQFEK Tag Antibody, mAb, Mouse, GenScript, Boechout, Belgium) and secondary anti-mouse APC antibody (A-865, Thermo Fisher Scientific) as controls.
2.6. Transduction of HUVEC and IFN Stimulation
250,000 cells per well were seeded in fibronectin-coated 6-well plates. A day was allowed for cell attachment before transduction. Cells were transduced with lentiviruses expressing the selected SARS-CoV-2 proteins, either from the Wuhan or the Omicron variant in the presence of 1 μg/mL polybrene was use to aid the transduction (Figure S1). The cells were stimulated with 500 IU/mL IFN-α2 or 1000 UI/mL IFN-β 24 h post-transduction. Non-stimulated transduced cells were kept for each condition and non-transduced cells were also included as controls. 24 h after the stimulation, the cells were lysed using a Qiazol lysis buffer and kept at -80 °C overnight to aid the lysis. mRNA was extracted from cell lysates the next day, using the QIAGEN RNeasy Mini kit, the mRNA samples were measured and qPCR was performed.
2.7. RT-qPCR
In brief, mRNA was extracted from HUVEC cells using the QIAGEN miRNeasy Mini kit according to the manufacturer’s instructions.. RNA (max 1 μg) was subsequently treated with amplification-grade DNAse I (Life Technologies) and used for synthesis of cDNA with Superscript III reverse transcriptase and random primers (Life Technologies). Depending on the gene to be measured, cDNA was subsequently diluted 3x (for target genes: IFNB1, IFNA2 and IFIT1) and 15x (for reference genes: ACTIN, RPL13A, YWHAZ and UBC) with Nuclease-free water (Ambion, Life Technologies). 5 μL of the diluted cDNA was then used for qPCR. For qPCR LightCycler 480 SYBR Green I Master mix (Roche Diagnostics, Vilvoorde, Belgium) was used in final reaction of 15 μL. qPCR reactions were performed in 384-well plates (LightCycler 480 Multiwell Plates 384, white, Roche Diagnostics) on the Light Cycler 480 II instrument (Roche Diagnostics).
All samples were measured in duplo. A non-template control (nuclease-free water instead of cDNA) and a serial 10-fold dilution of standard curve was used. The cDNA for the standard curve was synthesized using mRNA from poly(I:C) stimulated PBMCs and this standard curve was included for the measurement of each gene on the plate. Melting curve analysis for IFIT1, IFNA2 and IFNB1 was performed and showed a single peak. Calibrated normalized relative quantities (CNRQs) were calculated for each target genes in each sample based on the obtained Cq values, with the qBase Software (Biogazelle, CellCarta, Montreal, Quebec), using YWHAZ, ACTIN, RPL13A and UBC as reference genes and using target- and run- specific amplification efficiencies.
Primers used for qPCR were: UBC Fwd (sense) 5’- ATTTGGGTCGCGGTTCTTG -3’, UBC Rev (antisense) 5’- TGCCTTGACATTCTCGATGGT-3’, YWHAZ Fwd (sense) 5’-CTTTTGGTACATTGTGGCTTC AA -3’, YWHAZ Rev (antisense) 5’-CCGCCAGGACAAACCAGTAT -3’, ACTIN Fwd (sense) 5’- TGACCCAGATCATGTTTGAGA -3’, ACTIN Rev (antisense) 5’- AGAGGCGTACAGGGATAGCA -3’, RPL13A Fwd (sense) 5’- CCTGGAGAAGAGGAAAGAGA -3’, RPL13A Rev (antisense) 5’- TTGAGGACCTCTGTGTATTTGTCAA -3’, IFIT1 Fwd (sense) 5’- GATCTCAGAGGAGCCTGGCTAA –3’, IFIT1 Rev (antisense) 5’- TGATCATCACCATTTGTACTCATGG -3’, IFNA2 Fwd (sense) 5’- GTGAGGAAATACTTCCAAAGAATCAC –3’, IFNA2 Rev (antisense) 5’- TCTCATGATTTCTGCTCTGACAA -3’, IFNB1 Fwd (sense) 5’- GCTTCTCCACTACAGCTCTTTC -3’, IFNB1 Rev (antisense) 5’- CAGTATTCAAGCCTCCCATTCA -3’.
All primers were purchased from IDT (Integrated DNA Technologies, Europe Branch, Leuven, Belgium).
2.8. Figure Generation
3. Results
3.1. Comprehensive Review of SARS-CoV-2 Proteins and Their Immune Modulatory Effects
One of the defining features of SARS-CoV-2 pathogenesis is its ability to suppress and delay IFN responses.
To provide a comprehensive understanding of the immune modulatory effects of SARS-CoV-2 proteins, we systematically reviewed available literature to create a detailed reference (Table 1). This table catalogs the functions of all 29 currently known viral proteins on host pathways, highlighting their diverse mechanisms of action, including immune evasion, host machinery manipulation, and inflammatory response modulation. From this, we reanalyzed and organized the data into a second framework (Table 2), where viral proteins were categorized based on specific immune evasion strategies. These strategies include hijacking host protein synthesis machinery, protecting viral RNA, protecting viral proteins, facilitating immune suppression, and subverting innate immune pathways. This integrative analysis offers detailed insights into the multitiered immune evasion tactics employed by SARS-CoV-2 and sets a foundation for further exploring these viral mechanisms.
For example, within the first hour of infection, Nsp1, Nsp2, and Envelope (E), act together to suppress host mRNA translation but through distinct pathways. Nsp1 achieves this by globally shutting down host protein synthesis, facilitating rapid viral replication, Nsp2 suppresses translation by binding the GIGYF2-4EHP complex, while the Envelope (E) protein manipulates host transcription through interactions with epigenetic regulators, redirecting cellular machinery to favor viral protein production. To protect its replication machinery, it employs Nsp3 , Nsp4 and Nsp6 to form the replication-transcription complex (RTC), while many other viral proteins suppress immune signaling and antigen presentation in a coordinated manner. Plethora of proteins block the RIG-I/MAVS pathway through deubiquitinating or degrading a crucial signaling molecule or directly binding to a protein like IRF3 and preventing its nuclear translocation. SARS-CoV-2 assures its virion release by directly antagonizing tetherin via ORF7a or rerouting tetherin to endosomes and lysosomes via ORF3a.
Beyond IFN signaling, SARS-CoV-2 modulates antigen presentation to avoid immune detection. Proteins such as ORF3a, ORF7a, and Nsp5 independently suppress MHC-I and MHC-II expression. Furthermore, SARS-CoV-2 modulates natural killer (NK) cell and T cell responses. Nsp1 downregulates NKG2D ligands, reducing NK cell recognition of infected cells, while ORF8 binds CD16a on monocytes and NK cells, impairing antibody-dependent cytotoxicity (ADCC). Nsp7 binds HLA-DRB1 and activates regulatory T cells, suppressing CD4+ and CD8+ memory responses and cloaking infected cells from adaptive immune responses. These strategies impair cytotoxic T-cell recognition of infected cells, contributing to prolonged viral persistence
3.2. Functional Screening of SARS-CoV-2 Wuhan and Omicron Strain Proteins for Impact on Innate Immune Sensing
Despite significant progress in understanding SARS-CoV-2 immune evasion strategies, most research has been focused on the original Wuhan strain (Table 1 and Table 2). Mutations in the Spike (S) protein, but also in accessory and non-structural proteins have affected the virus strain phenotype tremendously [30]. Therefore, a re-evaluation of variant protein effect on viral infectivity, immune evasion, and pathogenicity is warranted.
In this study, we performed a global screening of the SARS-CoV-2 viral proteins from both the Wuhan and Omicron strains to assess their impact on type I interferon response. In HEK293T cells, we used a dual luciferase reporter assay and Strep II-tagged expression constructs coding for the 29 currently known SARS-CoV-2 proteins (Nsp1, Nsp2, Nsp3, Nsp4, Nsp5, Nsp6, Nsp7, Nsp8, Nsp9, Nsp10, Nsp11, Nsp12, Nsp13, Nsp14, Nsp15, Nsp16, S, ORF3a, ORF3c, E, M, ORF6, ORF7a, ORF7b, ORF8, ORF9b, N, ORF9c, and ORF10) from the original Wuhan strain and from Omicron BA.1. strain (Figure 3A).
Several proteins are highly conserved and don’t have distinct variants between the two strains. Examples are the Nsp7-Nsp8 complex, the helicase (Nsp13), the endoribonuclease (Nsp15) and the 2’-O-methyltransferase (Nsp16), who are all part of the RTC. The impact of all viral proteins on a major branch of innate immunity, ISRE (IFN/pro-inflammatory cytokine induction by RIG-I-like receptors (RLRs)) was analyzed by measuring luciferase activity driven by stimulation of the ISRE promoter in a quantitative reporter assays. Stimulation by type I IFNs (IFN-α2 and IFN-β) was titrated, quantified, and optimized beforehand using quantitative firefly luciferase reporters controlled by the respective promoters.
Stimulation with IFN-α2 and IFN-β revealed that activation of the ISRE is strongly repressed by Nsp1, Nsp5, Nsp14 and ORF6 across both strains (Figure 3B-E), underscoring their conserved immune evasion functions. Additionally, proteins such as Nsp9, Nsp11, Nsp13, ORF7a, and ORF8 from the Wuhan strain (identical protein in Omicron) displayed strong IFN inhibition. Intriguingly, when stimulated by IFN-β (Figure 3 B-C) the Omicron Spike protein, carrying 32 additional mutations, exhibited enhanced suppression of IFN responses, irrespective of the stimulus either IFN-β (Figure 3B-C) or IFN-α2 (Figure 3D-E).
Additionally, we screened for cell viability at 8 and 24 hours post-transfection to evaluate the toxic effect of each viral protein on the viability of the cells (Figure 4). At 8 hours post-transfection, both Wuhan (Figure 4A) and Omicron BA.1 (Figure 4B) variants demonstrated minimal cytotoxic effects, as indicated by the near-complete preservation of ATP levels in transfected cells. This suggests that the early expression of individual viral proteins does not substantially impact cell viability under these conditions. However, by 24 hours post-transfection (Figure 4C-D), time-dependent cytotoxic effects became apparent. Certain proteins, like Nsp3, Nsp6 and ORF3a, exhibited mild to moderate reductions in cell viability, indicative of their potential to disrupt cellular processes over time.
3.3. Model to Study Vascular Impact: Immune Response in Endothelial Cells
In SARS-CoV-2’s pathology, endothelial dysfunction and coagulation disorder occurs, leading to sometimes fatal outcomes. Since ACE2 receptors are also expressed in endothelial cells (ECs), infection by SARS-CoV-2 occurs. We therefore investigated the effect of selected viral proteins on expression of type I interferon pathways in human ECs from umbilical vein (HUVECs), expressing viral proteins by lentiviral transduction.
We selected from both the Wuhan and the Omicron BA.1 strains the Envelope, the Membrane and the ORF9b protein to further compare their effect on human primary endothelial cells, because of their features (see Table 1) and a possible connection to cardiovascular complications and coagulopathies [70,151,153,169]. We could not test the ORF6 protein since expression of ORF6 from both variants was toxic and severely affected the viability of the primary endothelial cells.
Analysis of ISGs expression levels in transduced HUVEC cells revealed distinct patterns of ISG induction under non-stimulated and interferon-stimulated conditions (Figure 5).
Compared to the Wuhan strain, the Omicron variant of the Envelope protein (Figure 5A) exhibited diminished ability to induce ISG expression (IFIT1 and IFNB1) independently, as seen in the non-stimulated condition. Upon external stimulation with IFN-β or IFN-α2, the Omicron BA.1 variant did not amplify the interferon-induced signaling pathway, as evidenced by the attenuated transcription of IFIT1, IFNB1, and IFNA2, compared to the Wuhan variant of the Envelope protein. This suggests that the Envelope protein, particularly from the Omicron BA.1 variant, has evolved to avoid boosting interferon responses, which may reflect viral adaptation to evade host immune responses.
For the Membrane protein (Figure 5B), the differences between transduction and stimulations were minimal, except for a consistent reduction of IFIT1, IFNB1, and IFNA2 expression in IFN-α2 stimulated HUVECs expressing the Omicron variant. These findings suggest that the Membrane protein only plays a minor or context-dependent role in modulation of interferon responses.
Also the Omicron ORF9b protein (Figure 5C) displayed this phenotype, however both Omicron and Wuhan variants induced IFIT1 expression in unstimulated, but not in IFN-β or IFN-α2 stimulated HUVECs.
4. Discussion
The immune evasion strategies employed by SARS-CoV-2 remain one of the defining features of viral fitness. Our study provides a comprehensive review and characterization of viral proteins that contribute to this process, pointing to both conserved and variant-specific immune modulation across the Wuhan and Omicron variants. Through a systematic literature review and experimental validation using luciferase-based assays and an endothelial cell model, we identified viral proteins that modulate IFN responses.
Our global screening approach using a dual luciferase reporter assay quantitatively validated the well-characterized IFN antagonistic functions of several viral proteins while also revealing novel differences in strain-specific immune suppression. Consistent with previous reports on the Wuhan variant [39,75,104,106,170,171], Nsp1, Nsp5, Nsp14, and ORF6 emerged as potent suppressors of ISRE-driven IFN responses in both the Wuhan and Omicron variants. This supports their well-documented roles in shutting down host protein synthesis, blocking IFN-stimulated gene (ISG) expression, and preventing nuclear translocation of key transcription factors such as IRF3. Interestingly, we found that several viral proteins displayed different immune-suppressive capabilities between the two strains (Figure 3 and Figure 5), highlighting the adaptive evolution of SARS-CoV-2 in response to host immunity.
Among these, Nsp6 exhibited enhanced IFN suppression in the Omicron variant compared to the Wuhan strain (Figure 3A-D). This suggests that selective pressure on non-structural proteins has contributed to Omicron’s immune evasion capacity, beyond the extensive mutations observed in the Spike protein. In this cell line model, the Envelope protein plays a role in stringent IFN suppression in the Wuhan strain, what is pretty much conserved in Omicron. Given that the Envelope protein is implicated in viral assembly and release [47], as well as aiding in immune tolerance [48,49,164], conserved immune suppressive activity points to a selective pressure. Similarly, this pressure is also observed for the Membrane protein. In contrast, the accessory proteins ORF7a and ORF8 strongly inhibited IFN responses, and their sequences are highly conserved across the variants, suggesting a crucial role in interacting with host immune regulators and downregulating MHC-I or blocking STAT1 phosphorylation. These findings also raise an important question about whether Omicron’s reduced pathogenicity results directly from cumulative virus-host interactions contributing to an enhanced immune evasion phenotype.
The results from our cytotoxicity assay further emphasized the complexity of SARS-CoV-2-host interactions. While early expression of most viral proteins had minimal impact on cell viability at 8 hours post-transfection, a subset of proteins (Nsp3, Nsp5, Nsp6, Nsp13, Nsp14 and ORF3a) exhibited significant cytotoxicity at 24 hours. This delayed effect suggests that these proteins have a moderate effect and may disrupt host cell homeostasis over time, potentially contributing to systemic inflammation and endothelial dysfunction observed in severe COVID-19 cases.
A key feature of SARS-CoV-2 pathogenesis is its disruption of endothelial function, which leads to thrombotic complications [11]. As a respiratory virus, besides infecting ACE2-positive cells, SARS-CoV-2 appears uniquely capable of infecting human ACE2-low endothelial cells through αV/β3 integrin-mediated endocytosis [172]. This invalidates a previous hypothesis that the virus does not effectively infect the blood vessel lining due to the relatively lower ACE2 receptor density on endothelial cell surfaces, as opposed to respiratory epithelial cells [173].
Our endothelial cell model allowed us to assess the direct impact of select viral proteins (Envelope, Membrane, ORF9b) on IFN responses in primary human endothelial cells. The findings show that the Envelope protein from the Omicron variant exhibits an inhibitory effect on IFN responses compared to the Wuhan strain. In contrast, ORF9b displayed a paradoxical phenotype—inducing IFIT1 expression in unstimulated conditions while suppressing it upon IFN stimulation. This suggests that ORF9b may function as an immune modulator rather than a strict inhibitor, dynamically adjusting host responses to favor viral persistence. While it blocks TBK1 phosphorylation by targeting TOM70 at the mitochondria [174,175,176] and binds NEMO (IKKγ), thereby inhibiting NF-kB activation [149], its seems that ORF9b plays an important role in endothelial dysfunction. ORF9b is reported to upregulates fibrinogen α, β and γ – key prothrombotic factors that correlate with coagulation disorders and vascular inflammation in severe COVID-19 cases [153]. ORF9b is also reported to impair cardiomyocytes metabolism [151], similar to the Membrane protein [96] which disrupts calcium handling and compromises cardiac contractility. The inclusion of the Membrane protein in our endothelial cell studies yielded less conclusive results, as both strains showed minimal differences in ISG induction. This suggests that while the Membrane protein may contribute to immune evasion, its role in endothelial dysfunction likely occurs through alternative pathways.
While the study presents a comprehensive comparative analysis of SARS-CoV-2 immune modulation across the Wuhan and Omicron variants and is indicative of the need to study their impact on the vascular system, there are several limitations. First, the reliance on overexpression systems in transduced endothelial cells, while informative, may not fully replicate the physiological context of viral infection, where protein expression levels and timing are tightly regulated. Second, the use of ISRE-driven luciferase assays, although valuable, provides only a snapshot of IFN pathway activation and does not capture the broader landscape of immune signaling dysregulation or downstream cytokine responses. Previous studies in cell lines that tested only viral proteins from the Wuhan variant, expanded their scope by examining different steps of the immune signaling pathways, including RIG-I, MDA5, MAVS, TRIF, IRF3 or STAT1 and STAT2 [39,75,106,170,177]. Additionally, our study lacks further mechanistic validation of the effect on ISGs’ transcription in primary endothelial cells, which are physiologically relevant to SARS-CoV-2 infection and pathogenesis. However, experiments involving primary endothelial cells, such as human umbilical endothelial cells (HUVECs), are technically challenging and time-consuming, presenting an additional experimental barrier. The absence of in vivo models or patient-derived samples further restricts the translational relevance of the findings. Finally, while the evolutionary divergence of proteins such as the Envelope, the Membrane and Spike proteins is intriguing, the mechanism of their altered immune evasion capabilities remains insufficiently explored. Addressing these limitations in future studies would significantly enhance the robustness and applicability of the findings.
Nevertheless, these findings primarily aim to contribute a baseline of understanding in the SARS-CoV-2 evolution and their implications in vascular pathologies. It remains unclear whether the changes between variants reflect a trade-off between immune evasion and transmissibility, or whether additional factors such as host immune memory and vaccination have influenced the functional evolution of SARS-CoV-2 proteins. This study provides two key contributions: 1) a comprehensive assessment of SARS-CoV-2 proteins and their immune-modulatory effects across two major variants, and 2) an underexplored avenue of SARS-CoV-2 endothelial dysfunction research that warrants further investigation.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1. Lentiviral Transduction of Primary Human Umbilical Vein Endothelial Cells (HUVEC) and Interferon-Stimulated Gene Expression Analysis (A) HUVEC were freshly isolated from donated human umbilical cords, expanded on fibronectin-coated plates, and maintained in endothelial cell culture medium. (B) Lentiviruses encoding Strep II-tagged SARS-CoV-2 proteins (Wuhan and Omicron variants) were produced in HEK293T cells and titrated. HUVEC were transduced in fibronectin-coated plates using polybrene, then stimulated with IFN-α2 (500 IU/mL) or IFN-β (1000 IU/mL) 24 hours post-transduction. Non-stimulated and non-transduced controls were included. Cells were lysed and total RNA was extracted for qPCR analysis of interferon-stimulated genes (IFNβ, IFNα, IFIT1) to assess the impact of SARS-CoV-2 proteins on the IFN response. Table S1. SARS-CoV-2 viral proteins and their mechanism of action.
Author Contributions
Conceptualization, M.J. and B.V.; methodology, M.J, E.N. and B.V.; formal analysis, M.J., E.N..; investigation, M.J..; resources, M.J. and B.V..; data curation, M.J.; writing—original draft preparation, M.J.; writing—review and editing, B.V.; visualization, M.J.; supervision, B.V.; project administration, M.J.; funding acquisition, B.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Fund for Scientific Research Flanders, grant COVID-Trace.
Institutional Review Board Statement
Study was approved by the Ghent University Hospital ethical committee (ONZ-2022-0272).
Informed Consent Statement
Informed consent was obtained from all mothers who donated umbilical cords used in the study
Data Availability Statement
all data are available upon request.
Acknowledgments
We sincerely thank Dr. Nevan Krogan for providing the codon-optimized expression constructs of individual SARS-CoV-2 proteins, and the mothers willing to donate umbilical cords.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| IFN | Interferon |
| SARS-CoV | Severe Acute Respiratory Syndrome Coronavirus |
| PRRs | Pattern recognition receptors |
| TLRs | Toll-like receptors |
| RIG-I | Retinoic acid-inducible gene I |
| MAVS | Mitochondrial antiviral-signaling protein |
| TBK1 | TANK-binding kinase 1 |
| IRF | Interferon regulatory factors |
| EC | Endothelial cells |
| RTC | Replication-transcription complex |
| RLRs | RIG-I-like receptors |
| HUVEC | Human EC from umbilical vein |
| ISG | Interferon stimulated gene |
References
- Kirtipal N, Bharadwaj S, Kang SG. 2020. From SARS to SARS-CoV-2, insights on structure, pathogenicity and immunity aspects of pandemic human coronaviruses. Infect Genet Evol. 85:104502.
- Barnes CO, Jette CA, Abernathy ME, Dam K-MA, Esswein SR, Gristick HB, Malyutin AG, Sharaf NG, Huey-Tubman KE, Lee YE, Robbiani DF, Nussenzweig MC, West AP, Bjorkman PJ. 2020. SARS-CoV-2 neutralizing antibody structures inform therapeutic strategies. Nature 588:682–687.
- Cao Y, Su B, Guo X, Sun W, Deng Y, Bao L, Zhu Q, Zhang X, Zheng Y, Geng C, Chai X, He R, Li X, Lv Q, Zhu H, Deng W, Xu Y, Wang Y, Qiao L, Tan Y, Song L, Wang G, Du X, Gao N, Liu J, Xiao J, Su X, Du Z, Feng Y, Qin C, Qin C, Jin R, Xie XS. 2020. Potent Neutralizing Antibodies against SARS-CoV-2 Identified by High-Throughput Single-Cell Sequencing of Convalescent Patients’ B Cells. Cell 182:73-84.
- Klasse P, Moore JP. 2020. Antibodies to SARS-CoV-2 and their potential for therapeutic passive immunization. Elife. 9:e57877.
- Del Sole F, Farcomeni A, Loffredo L, Carnevale R, Menichelli D, Vicario T, Pignatelli P, Pastori D. 2020. Features of severe COVID-19: A systematic review and meta-analysis. Eur J Clin Invest. 50(10):e13378.
- Rossouw TM, Anderson R, Manga P, Feldman C. 2022. Emerging Role of Platelet-Endothelium Interactions in the Pathogenesis of Severe SARS-CoV-2 Infection-Associated Myocardial Injury. Front Immunol. 13:776861.
- Sievers BL, Cheng MTK, Csiba K, Meng B, Gupta RK. 2023. SARS-CoV-2 and innate immunity: the good, the bad, and the “goldilocks.” Cell Mol Immunol 21:171–183.
- Onomoto K, Onoguchi K, Yoneyama M. 2021. Regulation of RIG-I-like receptor-mediated signaling: interaction between host and viral factors. Cell Mol Immunol 18:539–555.
- Alfaro E, Díaz-García E, García-Tovar S, Galera R, Casitas R, Torres-Vargas M, López-Fernández C, Añón JM, García-Río F, Cubillos-Zapata C. 2024. Endothelial dysfunction and persistent inflammation in severe post-COVID-19 patients: implications for gas exchange. BMC Med. 22(1):242.
- Ackermann M, Kamp JC, Werlein C, Walsh CL, Stark H, Prade V, Surabattula R, Wagner WL, Disney C, Bodey AJ, Illig T, Leeming DJ, Karsdal MA, Tzankov A, Boor P, Kühnel MP, Länger FP, Verleden SE, Kvasnicka HM, Kreipe HH, Haverich A, Black SM, Walch A, Tafforeau P, Lee PD, Hoeper MM, Welte T, Seeliger B, David S, Schuppan D, Mentzer SJ, Jonigk DD. 2022. The fatal trajectory of pulmonary COVID-19 is driven by lobular ischemia and fibrotic remodelling. EBioMedicine 85:104296.
- Varga Z, Flammer AJ, Steiger P, Haberecker M, Andermatt R, Zinkernagel AS, Mehra MR, Schuepbach RA, Ruschitzka F, Moch H. 2020. Endothelial cell infection and endotheliitis in COVID-19. The Lancet. 395(10234):1417-1418.
- Becker RC, Tantry US, Khan M, Gurbel PA. 2024. The COVID-19 thrombus: distinguishing pathological, mechanistic, and phenotypic features and management. J Thromb Thrombolysis 58:15–49.
- Bai B, Yang Y, Wang Q, Li M, Tian C, Liu Y, Aung LHH, Li P, Yu T, Chu X. 2020. NLRP3 inflammasome in endothelial dysfunction. Cell Death Dis 11:776.
- Birnhuber A, Fließer E, Gorkiewicz G, Zacharias M, Seeliger B, David S, Welte T, Schmidt J, Olschewski H, Wygrecka M, Kwapiszewska G. 2021. Between inflammation and thrombosis: Endothelial cells in COVID-19. European Respiratory Journal. 58(3):2100377.
- Guney C, Akar F. 2021. Epithelial and Endothelial Expressions of ACE2: SARS-CoV-2 Entry Routes. J Pharm Pharm Sci. 24:84-93.
- Lei Y, Zhang J, Schiavon CR, He M, Chen L, Shen H, Zhang Y, Yin Q, Cho Y, Andrade L, Shadel GS, Hepokoski M, Lei T, Wang H, Zhang J, Yuan JX-J, Malhotra A, Manor U, Wang S, Yuan Z-Y, Shyy JY-J. 2021. SARS-CoV-2 Spike Protein Impairs Endothelial Function via Downregulation of ACE 2. Circ Res 128:1323–1326.
- Perico L, Morigi M, Pezzotta A, Locatelli M, Imberti B, Corna D, Cerullo D, Benigni A, Remuzzi G. 2023. SARS-CoV-2 spike protein induces lung endothelial cell dysfunction and thrombo-inflammation depending on the C3a/C3a receptor signalling. Sci Rep 13:11392.
- Kang S, Tanaka T, Inoue H, Ono C, Hashimoto S, Kioi Y, Matsumoto H, Matsuura H, Matsubara T, Shimizu K, Ogura H, Matsuura Y, Kishimoto T. 2020. IL-6 trans-signaling induces plasminogen activator inhibitor-1 from vascular endothelial cells in cytokine release syndrome. Proceedings of the National Academy of Sciences 117:22351–22356.
- Kang S, Kishimoto T. 2021. Interplay between interleukin-6 signaling and the vascular endothelium in cytokine storms. Exp Mol Med 53:1116–1123.
- Puhlmann M, Weinreich DM, Farma JM, Carroll NM, Turner EM, Alexander HR. 2005. Interleukin-1β induced vascular permeability is dependent on induction of endothelial Tissue Factor (TF) activity. J Transl Med 3:37.
- Kandhaya-Pillai R, Yang X, Tchkonia T, Martin GM, Kirkland JL, Oshima J. 2022. TNF-α/IFN-γ synergy amplifies senescence-associated inflammation and SARS-CoV-2 receptor expression via hyper-activated JAK/STAT1. Aging Cell 21(6):e13646.
- Valencia I, Lumpuy-Castillo J, Magalhaes G, Sánchez-Ferrer CF, Lorenzo Ó, Peiró C. 2024. Mechanisms of endothelial activation, hypercoagulation and thrombosis in COVID-19: a link with diabetes mellitus. Cardiovasc Diabetol 23:75.
- Won T, Wood MK, Hughes DM, Talor M V., Ma Z, Schneider J, Skinner JT, Asady B, Goerlich E, Halushka MK, Hays AG, Kim D-H, Parikh CR, Rosenberg AZ, Coppens I, Johns RA, Gilotra NA, Hooper JE, Pekosz A, Čiháková D. 2022. Endothelial thrombomodulin downregulation caused by hypoxia contributes to severe infiltration and coagulopathy in COVID-19 patient lungs. EBioMedicine 75:103812.
- Jin Y, Ji W, Yang H, Chen S, Zhang W, Duan G. 2020. Endothelial activation and dysfunction in COVID-19: from basic mechanisms to potential therapeutic approaches. Signal Transduct Target Ther 5:293.
- Lim EHT, van Amstel RBE, de Boer VV, van Vught LA, de Bruin S, Brouwer MC, Vlaar APJ, van de Beek D. 2023. Complement activation in COVID-19 and targeted therapeutic options: A scoping review. Blood Rev 57:100995.
- Conway EM, Pryzdial ELG. 2020. Is the COVID-19 thrombotic catastrophe complement-connected? Journal of Thrombosis and Haemostasis 18:2812–2822.
- Laha S, Chakraborty J, Das S, Manna SK, Biswas S, Chatterjee R. 2020. Characterizations of SARS-CoV-2 mutational profile, spike protein stability and viral transmission. Infection, Genetics and Evolution 85:104445.
- Watanabe Y, Allen JD, Wrapp D, McLellan JS, Crispin M. 2020. Site-specific glycan analysis of the SARS-CoV-2 spike. Science 369:330–333.
- Gong Y, Qin S, Dai L, Tian Z. 2021. The glycosylation in SARS-CoV-2 and its receptor ACE2. Signal Transduct Target Ther. 6(1):396.
- Harvey WT, Carabelli AM, Jackson B, Gupta RK, Thomson EC, Harrison EM, Ludden C, Reeve R, Rambaut A, Peacock SJ, Robertson DL. 2021. SARS-CoV-2 variants, spike mutations and immune escape. Nat Rev Microbiol. 19(7):409-424.
- Montezano AC, Camargo LL, Mary S, Neves KB, Rios FJ, Stein R, Lopes RA, Beattie W, Thomson J, Herder V, Szemiel AM, McFarlane S, Palmarini M, Touyz RM. 2023. SARS-CoV-2 spike protein induces endothelial inflammation via ACE2 independently of viral replication. Sci Rep 13(1):14086.
- Urata R, Ikeda K, Yamazaki E, Ueno D, Katayama A, Shin-Ya M, Ohgitani E, Mazda O, Matoba S. 2022. Senescent endothelial cells are predisposed to SARS-CoV-2 infection and subsequent endothelial dysfunction. Sci Rep 12(1):11855.
- Li F, Li J, Wang PH, Yang N, Huang J, Ou J, Xu T, Zhao X, Liu T, Huang X, Wang Q, Li M, Yang L, Lin Y, Cai Y, Chen H, Zhang Q. 2021. SARS-CoV-2 spike promotes inflammation and apoptosis through autophagy by ROS-suppressed PI3K/AKT/mTOR signaling. Biochim Biophys Acta Mol Basis Dis 1867(12):166260.
- Scheim DE, Vottero P, Santin AD, Hirsh AG. 2023. Sialylated Glycan Bindings from SARS-CoV-2 Spike Protein to Blood and Endothelial Cells Govern the Severe Morbidities of COVID-19. Int J Mol Sci. 24(23):17039.
- Savellini GG, Anichini G, Gandolfo C, Cusi MG. 2021. SARS-CoV-2 n protein targets TRIM25-mediated RIG-I activation to suppress innate immunity. Viruses 13(8):1439.
- Oh SJ, Shin OS. 2021. Sars-cov-2 nucleocapsid protein targets rig-i-like receptor pathways to inhibit the induction of interferon response. Cells 10:1–13.
- Wang W, Chen J, Yu X, Lan HY. 2022. Signaling mechanisms of SARS-CoV-2 Nucleocapsid protein in viral infection, cell death and inflammation. Int J Biol Sci. 18(12):4704-4713.
- Zhao Y, Sui L, Wu P, Wang W, Wang Z, Yu Y, Hou Z, Tan G, Liu Q, Wang G. 2021. A dual-role of SARS-CoV-2 nucleocapsid protein in regulating innate immune response. Signal Transduct Target Ther 6(1):331.
- Xia H, Cao Z, Xie X, Zhang X, Chen JYC, Wang H, Menachery VD, Rajsbaum R, Shi PY. 2020. Evasion of Type I Interferon by SARS-CoV-2. Cell Rep 33(1):108234.
- Huang C, Yin Y, Pan P, Huang Y, Chen S, Chen J, Wang J, Xu G, Tao X, Xiao X, Li J, Yang J, Jin Z, Li B, Tong Z, Du W, Liu L, Liu Z. 2023. The Interaction between SARS-CoV-2 Nucleocapsid Protein and UBC9 Inhibits MAVS Ubiquitination by Enhancing Its SUMOylation. Viruses 15(12):2304.
- Guo X, Yang S, Cai Z, Zhu S, Wang H, Liu Q, Zhang Z, Feng J, Chen X, Li Y, Deng J, Liu J, Li J, Tan X, Fu Z, Xu K, Zhou L, Chen Y. 2025. SARS-CoV-2 specific adaptations in N protein inhibit NF-κB activation and alter pathogenesis. Journal of Cell Biology 224(1):e202404131.
- Carlson CR, Asfaha JB, Ghent CM, Howard CJ, Hartooni N, Safari M, Frankel AD, Morgan DO. 2020. Phosphoregulation of Phase Separation by the SARS-CoV-2 N Protein Suggests a Biophysical Basis for its Dual Functions. Mol Cell 80:1092-1103.e4.
- Fu YZ, Wang SY, Zheng ZQ, Yi Huang, Li WW, Xu ZS, Wang YY. 2021. SARS-CoV-2 membrane glycoprotein M antagonizes the MAVS-mediated innate antiviral response. Cell Mol Immunol 18:613–620.
- Lopandić Z, Protić-Rosić I, Todorović A, Glamočlija S, Gnjatović M, Ćujic D, Gavrović-Jankulović M. 2021. IgM and IgG Immunoreactivity of SARS-CoV-2 Recombinant M Protein. Int J Mol Sci 22(9):4951.
- Liu J, Wu S, Zhang Y, Wang C, Liu S, Wan J, Yang L. 2023. SARS-CoV-2 viral genes Nsp6, Nsp8, and M compromise cellular ATP levels to impair survival and function of human pluripotent stem cell-derived cardiomyocytes. Stem Cell Res Ther 14(1):249.
- Cao Y, Yang R, Lee I, Zhang W, Sun J, Wang W, Meng X. 2021. Characterization of the SARS-CoV-2 E Protein: Sequence, Structure, Viroporin, and Inhibitors. Protein Sci. 30(6):1114-1130.
- Xia B, Shen X, He Y, Pan X, Liu FL, Wang Y, Yang F, Fang S, Wu Y, Duan Z, Zuo X, Xie Z, Jiang X, Xu L, Chi H, Li S, Meng Q, Zhou H, Zhou Y, Cheng X, Xin X, Jin L, Zhang HL, Yu DD, Li MH, Feng XL, Chen J, Jiang H, Xiao G, Zheng YT, Zhang LK, Shen J, Li J, Gao Z. 2021. SARS-CoV-2 envelope protein causes acute respiratory distress syndrome (ARDS)-like pathological damages and constitutes an antiviral target. Cell Res 31:847–860.
- Planès R, Bert JB, Tairi S, Benmohamed L, Bahraoui E. 2022. SARS-CoV-2 Envelope (E) Protein Binds and Activates TLR2 Pathway: A Novel Molecular Target for COVID-19 Interventions. Viruses 14(5):999.
- Geanes ES, McLennan R, Pierce SH, Menden HL, Paul O, Sampath V, Bradley T. 2024. SARS-CoV-2 envelope protein regulates innate immune tolerance. iScience 27(6):109975.
- Zheng M, Karki R, Williams EP, Yang D, Fitzpatrick E, Vogel P, Jonsson CB, Kanneganti TD. 2021. TLR2 senses the SARS-CoV-2 envelope protein to produce inflammatory cytokines. Nat Immunol 22:829–838.
- Lu H, Liu Z, Deng X, Chen S, Zhou R, Zhao R, Parandaman R, Thind A, Henley J, Tian L, Yu J, Comai L, Feng P, Yuan W. 2023. Potent NKT cell ligands overcome SARS-CoV-2 immune evasion to mitigate viral pathogenesis in mouse models. PLoS Pathog 19(3):e1011240.
- Bhat S, Rishi P, Chadha VD. 2022. Understanding the epigenetic mechanisms in SARS CoV-2 infection and potential therapeutic approaches. Virus Res. 318:198853.
- Schubert K, Karousis ED, Jomaa A, Scaiola A, Echeverria B, Gurzeler LA, Leibundgut M, Thiel V, Mühlemann O, Ban N. 2020. SARS-CoV-2 Nsp1 binds the ribosomal mRNA channel to inhibit translation. Nat Struct Mol Biol 27:959–966.
- Frolov I, Agback T, Palchevska O, Dominguez F, Lomzov A, Agback P, Frolova EI. 2023. All Domains of SARS-CoV-2 nsp1 Determine Translational Shutoff and Cytotoxicity of the Protein. J Virol 97(3):e0186522.
- Zhang K, Miorin L, Makio T, Dehghan I, Gao S, Xie Y, Zhong H, Esparza M, Kehrer T, Kumar A, Hobman TC, Ptak C, Gao B, Minna JD, Chen Z, García-Sastre A, Ren Y, Wozniak RW, Fontoura BMA. 2021. Nsp1 protein of SARS-CoV-2 disrupts the mRNA export machinery to inhibit host gene expression. Sci Adv. 7(6):eabe7386.
- Fisher T, Gluck A, Narayanan K, Kuroda M, Nachshon A, Hsu JC, Halfmann PJ, Yahalom-Ronen Y, Tamir H, Finkel Y, Schwartz M, Weiss S, Tseng CTK, Israely T, Paran N, Kawaoka Y, Makino S, Stern-Ginossar N. 2022. Parsing the role of NSP1 in SARS-CoV-2 infection. Cell Rep 39(11):110954.
- Lokugamage KG, Narayanan K, Huang C, Makino S. 2012. Severe Acute Respiratory Syndrome Coronavirus Protein nsp1 Is a Novel Eukaryotic Translation Inhibitor That Represses Multiple Steps of Translation Initiation. J Virol 86:13598–13608.
- Vazquez C, Swanson SE, Negatu SG, Dittmar M, Miller J, Ramage HR, Cherry S, Jurado KA. 2021. SARS-CoV-2 viral proteins NSP1 and NSP13 inhibit interferon activation through distinct mechanisms. PLoS One 16(6):e0253089.
- Lui WY, Ong CP, Cheung PHH, Ye ZW, Chan CP, To KKW, Yuen KS, Jin DY. 2024. Nsp1 facilitates SARS-CoV-2 replication through calcineurin-NFAT signaling. mBio 15(4):e0039224.
- Lee MJ, Leong MW, Rustagi A, Beck A, Zeng L, Holmes S, Qi LS, Blish CA. 2022. SARS-CoV-2 escapes direct NK cell killing through Nsp1-mediated downregulation of ligands for NKG2D. Cell Rep 41(13):111892.
- Xu Z, Choi J-H, Dai DL, Luo J, Ladak RJ, Li Q, Wang Y, Zhang C, Wiebe S, Liu ACH, Ran X, Yang J, Naeli P, Garzia A, Zhou L, Mahmood N, Deng Q, Elaish M, Lin R, Mahal LK, Hobman TC, Pelletier J, Alain T, Vidal SM, Duchaine T, Mazhab-Jafari MT, Mao X, Jafarnejad SM, Sonenberg N. 2022. SARS-CoV-2 impairs interferon production via NSP2-induced repression of mRNA translation. Proc Natl Acad Sci U S A. 119(32):e2204539119.
- Zou L, Moch C, Graille M, Chapat C. 2022. The SARS-CoV-2 protein NSP2 impairs the silencing capacity of the human 4EHP-GIGYF2 complex. iScience 25(7):104646.
- Lacasse É, Gudimard L, Dubuc I, Gravel A, Allaeys I, Boilard É, Flamand L. 2023. SARS-CoV-2 Nsp2 Contributes to Inflammation by Activating NF-κB. Viruses 15(2):334.
- Khan MT, Zeb MT, Ahsan H, Ahmed A, Ali A, Akhtar K, Malik SI, Cui Z, Ali S, Khan AS, Ahmad M, Wei DQ, Irfan M. 2021. SARS-CoV-2 nucleocapsid and Nsp3 binding: an in silico study. Arch Microbiol 203:59–66.
- Fu Z, Huang B, Tang J, Liu S, Liu M, Ye Y, Liu Z, Xiong Y, Zhu W, Cao D, Li J, Niu X, Zhou H, Zhao YJ, Zhang G, Huang H. 2021. The complex structure of GRL0617 and SARS-CoV-2 PLpro reveals a hot spot for antiviral drug discovery. Nat Commun 12(1):488.
- Clemente V, D’arcy P, Bazzaro M. 2020. Deubiquitinating enzymes in coronaviruses and possible therapeutic opportunities for COVID-19. Int J Mol Sci. 21(10):3492.
- Russo LC, Tomasin R, Matos IA, Manucci AC, Sowa ST, Dale K, Caldecott KW, Lehtiö L, Schechtman D, Meotti FC, Bruni-Cardoso A, Hoch NC. 2021. The SARS-CoV-2 Nsp3 macrodomain reverses PARP9/DTX3L-dependent ADP-ribosylation induced by interferon signaling. Journal of Biological Chemistry 297(3):101041.
- Garvanska DH, Alvarado RE, Mundt FO, Lindqvist R, Duel JK, Coscia F, Nilsson E, Lokugamage K, Johnson BA, Plante JA, Morris DR, Vu MN, Estes LK, McLeland AM, Walker J, Crocquet-Valdes PA, Mendez BL, Plante KS, Walker DH, Weisser MB, Överby AK, Mann M, Menachery VD, Nilsson J. 2024. The NSP3 protein of SARS-CoV-2 binds fragile X mental retardation proteins to disrupt UBAP2L interactions. EMBO Rep 25:902–926.
- Lavigne M, Helynck O, Rigolet P, Boudria-Souilah R, Nowakowski M, Baron B, Brülé S, Hoos S, Raynal B, Guittat L, Beauvineau C, Petres S, Granzhan A, Guillon J, Pratviel G, Teulade-Fichou MP, England P, Mergny JL, Munier-Lehmann H. 2021. SARS-CoV-2 Nsp3 unique domain SUD interacts with guanine quadruplexes and G4-ligands inhibit this interaction. Nucleic Acids Res 49:7695–7712.
- Faizan MI, Chaudhuri R, Sagar S, Albogami S, Chaudhary N, Azmi I, Akhtar A, Ali SM, Kumar R, Iqbal J, Joshi MC, Kharya G, Seth P, Roy SS, Ahmad T. 2022. NSP4 and ORF9b of SARS-CoV-2 Induce Pro-Inflammatory Mitochondrial DNA Release in Inner Membrane-Derived Vesicles. Cells 11(19):2969.
- Zimmermann L, Zhao X, Makroczyova J, Wachsmuth-Melm M, Prasad V, Hensel Z, Bartenschlager R, Chlanda P. 2023. SARS-CoV-2 nsp3 and nsp4 are minimal constituents of a pore spanning replication organelle. Nat Commun 14(1):7894.
- Li W, Qiao J, You Q, Zong S, Peng Q, Liu Y, Hu S, Liu W, Li S, Shu X, Sun B. 2021. SARS-CoV-2 Nsp5 Activates NF-κB Pathway by Upregulating SUMOylation of MAVS. Front Immunol 12:750969.
- Liu Y, Qin C, Rao Y, Ngo C, Feng JJ, Zhao J, Zhang S, Wang T-Y, Carriere E, Savas AC, Zarinfar M, Rice S, Yang H, Yuan W, Camarero JA, Yu J, Chen XS, Zhang C, Feng P. 2021. SARS-CoV-2 Nsp5 Demonstrates Two Distinct Mechanisms Targeting RIG-I and MAVS To Evade the Innate Immune Response. mBio. 12(5):e0233521.
- Zheng Y, Deng J, Han L, Zhuang MW, Xu Y, Zhang J, Nan ML, Xiao Y, Zhan P, Liu X, Gao C, Wang PH. 2022. SARS-CoV-2 NSP5 and N protein counteract the RIG-I signaling pathway by suppressing the formation of stress granules. Signal Transduct Target Ther 7(1):22.
- Shemesh M, Aktepe TE, Deerain JM, McAuley JL, Audsley MD, David CT, Purcell DFJ, Urin V, Hartmann R, Moseley GW, Mackenzie JM, Schreiber G, Harari D. 2021. SARS-CoV-2 suppresses IFNβ production mediated by NSP1, 5, 6, 15, ORF6 and ORF7b but does not suppress the effects of added interferon. PLoS Pathog 17(8):e1009800.
- Chen J, Li Z, Guo J, Xu S, Zhou J, Chen Q, Tong X, Wang D, Peng G, Fang L, Xiao S. 2022. SARS-CoV-2 nsp5 Exhibits Stronger Catalytic Activity and Interferon Antagonism than Its SARS-CoV Ortholog. J Virol 96(8):e0003722.
- Lu JL, Zhou XL. 2023. SARS-CoV-2 main protease Nsp5 cleaves and inactivates human tRNA methyltransferase TRMT1. J Mol Cell Biol. 15(4):mjad024.
- Ju X, Wang Z, Wang P, Ren W, Yu Y, Yu Y, Yuan B, Song J, Zhang X, Zhang Y, Xu C, Tian B, Shi Y, Zhang R, Ding Q. 2023. SARS-CoV-2 main protease cleaves MAGED2 to antagonize host antiviral defense. mBio 14:e0137323.
- Naik NG, Lee S-C, Veronese BHS, Ma Z, Toth Z. 2022. Interaction of HDAC2 with SARS-CoV-2 NSP5 and IRF3 Is Not Required for NSP5-Mediated Inhibition of Type I Interferon Signaling Pathway. Microbiol Spectr 10(5):e0232222.
- Taefehshokr N, Lac A, Vrieze AM, Dickson BH, Guo PN, Jung C, Blythe EN, Fink C, Aktar A, Dikeakos JD, Dekaban GA, Heit B. 2024. SARS-CoV-2 NSP5 antagonizes MHC II expression by subverting histone deacetylase 2. J Cell Sci 137(10):jcs262172.
- Bhat S, Rishi P, Chadha VD. 2022. Understanding the epigenetic mechanisms in SARS CoV-2 infection and potential therapeutic approaches. Virus Res. 318:198853.
- Li Y, Yu Q, Huang R, Chen H, Ren H, Ma L, He Y, Li W. 2023. SARS-CoV-2 SUD2 and Nsp5 Conspire to Boost Apoptosis of Respiratory Epithelial Cells via an Augmented Interaction with the G-Quadruplex of BclII. mBio 14(2):e0335922.
- Zhang C, Jiang Q, Liu Z, Li N, Hao Z, Song G, Li D, Chen M, Lin L, Liu Y, Li X, Shang C, Li Y. 2024. SARS-CoV-2 NSP6 reduces autophagosome size and affects viral replication via sigma-1 receptor. J Virol 98(11):e0075424.
- Jiao P, Fan W, Ma X, Lin R, Zhao Y, Li Y, Zhang H, Jia X, Bi Y, Feng X, Li M, Liu W, Zhang K, Sun L. 2023. SARS-CoV-2 nonstructural protein 6 triggers endoplasmic reticulum stress-induced autophagy to degrade STING1. Autophagy 19:3113–3131.
- Benvenuto D, Angeletti S, Giovanetti M, Bianchi M, Pascarella S, Cauda R, Ciccozzi M, Cassone A. 2020. Evolutionary analysis of SARS-CoV-2: how mutation of Non-Structural Protein 6 (NSP6) could affect viral autophagy. Journal of Infection 81:e24–e27.
- Nishitsuji H, Iwahori S, Ohmori M, Shimotohno K, Murata T. 2022. Ubiquitination of SARS-CoV-2 NSP6 and ORF7a Facilitates NF-κB Activation. mBio 13(4):e0097122.
- Bills CJ, Xia H, Chen JYC, Yeung J, Kalveram BK, Walker D, Xie X, Shi PY. 2023. Mutations in SARS-CoV-2 variant nsp6 enhance type-I interferon antagonism. Emerg Microbes Infect 12(1):2209208.
- Sun X, Liu Y, Huang Z, Xu W, Hu W, Yi L, Liu Z, Chan H, Zeng J, Liu X, Chen H, Yu J, Chan FKL, Ng SC, Wong SH, Wang MH, Gin T, Joynt GM, Hui DSC, Zou X, Shu Y, Cheng CHK, Fang S, Luo H, Lu J, Chan MTV, Zhang L, Wu WKK. 2022. SARS-CoV-2 non-structural protein 6 triggers NLRP3-dependent pyroptosis by targeting ATP6AP1. Cell Death Differ 29:1240–1254.
- Zhu J yi, Wang G, Huang X, Lee H, Lee JG, Yang P, van de Leemput J, Huang W, Kane MA, Yang P, Han Z. 2022. SARS-CoV-2 Nsp6 damages Drosophila heart and mouse cardiomyocytes through MGA/MAX complex-mediated increased glycolysis. Commun Biol 5(1):1039.
- Deng J, Zheng Y, Zheng SN, Nan ML, Han L, Zhang J, Jin Y, Pan JA, Gao C, Wang PH. 2023. SARS-CoV-2 NSP7 inhibits type I and III IFN production by targeting the RIG-I/MDA5, TRIF, and STING signaling pathways. J Med Virol 95(3):e28561.
- Ghelichkhani F, Gonzalez FA, Kapitonova MA, Rozovsky S. 2023. Selenoprotein S Interacts with the Replication and Transcription Complex of SARS-CoV-2 by Binding nsp7. J Mol Biol 435(8):168008.
- Miah SMS, Lelias S, Gutierrez AH, McAllister M, Boyle CM, Moise L, De Groot AS. 2023. A SARS-CoV-2 NSP7 homolog of a Treg epitope suppresses CD4+ and CD8+ T cell memory responses. Front Immunol 14:1290688.
- Guo J, Li WL, Huang M, Qiao J, Wan P, Yao Y, Ye L, Ding Y, Wang J, Peng Q, Liu W, Xia Y, Shu X, Sun B. 2024. SARS-CoV-2 Nsp7 plays a role in cognitive dysfunction by impairing synaptic plasticity. Front Neurosci 18:1490099.
- Yang Z, Zhang X, Wang F, Wang P, Kuang E, Li X. 2020. Suppression of MDA5-mediated antiviral immune responses by NSP8 of SARS-CoV-2 Emerg Microbes Infect. 81:e24.
- Zhang X, Yang Z, Pan T, Sun Q, Chen Q, Wang PH, Li X, Kuang E. 2023. SARS-CoV-2 Nsp8 suppresses MDA5 antiviral immune responses by impairing TRIM4-mediated K63-linked polyubiquitination. PLoS Pathog 19(11):e1011792.
- Liu J, Wu S, Zhang Y, Wang C, Liu S, Wan J, Yang L. 2023. SARS-CoV-2 viral genes Nsp6, Nsp8, and M compromise cellular ATP levels to impair survival and function of human pluripotent stem cell-derived cardiomyocytes. Stem Cell Res Ther 14(1):249.
- Zong S, Wu Y, Li W, You Q, Peng Q, Wang C, Wan P, Bai T, Ma Y, Sun B, Qiao J. 2023. SARS-CoV-2 Nsp8 induces mitophagy by damaging mitochondria. Virol Sin 38:520–530.
- Makiyama K, Hazawa M, Kobayashi A, Lim K, Voon DC, Wong RW. 2022. NSP9 of SARS-CoV-2 attenuates nuclear transport by hampering nucleoporin 62 dynamics and functions in host cells. Biochem Biophys Res Commun 586:137–142.
- Zhang Y, Xin B, Liu Y, Jiang W, Han W, Deng J, Wang P, Hong X, Yan D. 2023. SARS-COV-2 protein NSP9 promotes cytokine production by targeting TBK1. Front Immunol 14:1211816.
- Lundrigan E, Toudic C, Pennock E, Pezacki JP. 2024. SARS-CoV-2 Protein Nsp9 Is Involved in Viral Evasion through Interactions with Innate Immune Pathways. ACS Omega 9:26428–26438.
- Benoni R, Krafcikova P, Baranowski MR, Kowalska J, Boura E, Cahová H. 2021. Substrate specificity of sars-cov-2 nsp10-nsp16 methyltransferase. Viruses 13(9):1722.
- Wang H, Rizvi SR, Dong D, Lou J, Wang Q, Sopipong W, Su Y, Najar F, Agarwal PK, Kozielski F, Haider S. 2023. Emerging variants of SARS-CoV-2 NSP10 highlight strong functional conservation of its binding to two non-structural proteins, NSP14 and NSP16. Elife 12:RP87884.
- Yang L, Zeng XT, Luo RH, Ren SX, Liang LL, Huang QX, Tang Y, Fan H, Ren HY, Zhang WJ, Zheng YT, Cheng W. 2024. SARS-CoV-2 NSP12 utilizes various host splicing factors for replication and splicing regulation. J Med Virol. 96(1):e29396.
- Wang W, Zhou Z, Xiao X, Tian Z, Dong X, Wang C, Li L, Ren L, Lei X, Xiang Z, Wang J. 2021. SARS-CoV-2 nsp12 attenuates type I interferon production by inhibiting IRF3 nuclear translocation. Cell Mol Immunol 18:945–953.
- Vazquez C, Swanson SE, Negatu SG, Dittmar M, Miller J, Ramage HR, Cherry S, Jurado KA. 2021. SARS-CoV-2 viral proteins NSP1 and NSP13 inhibit interferon activation through distinct mechanisms. PLoS One 16(6):e0253089.
- Yuen CK, Lam JY, Wong WM, Mak LF, Wang X, Chu H, Cai JP, Jin DY, To KKW, Chan JFW, Yuen KY, Kok KH. 2020. SARS-CoV-2 nsp13, nsp14, nsp15 and orf6 function as potent interferon antagonists. Emerg Microbes Infect 9:1418–1428.
- Feng K, Zhang HJ, Min YQ, Zhou M, Deng F, Wang HL, Li PQ, Ning YJ. 2023. SARS-CoV-2 NSP13 interacts with host IRF3, blocking antiviral immune responses. J Med Virol 95(6):e28881.
- Fung SY, Siu KL, Lin H, Chan CP, Yeung ML, Jin DY. 2022. SARS-CoV-2 NSP13 helicase suppresses interferon signaling by perturbing JAK1 phosphorylation of STAT1. Cell Biosci 12(1):36.
- Li TW, Kenney AD, Park JG, Fiches GN, Liu H, Zhou D, Biswas A, Zhao W, Que J, Santoso N, Martinez-Sobrido L, Yount JS, Zhu J. 2022. SARS-CoV-2 Nsp14 protein associates with IMPDH2 and activates NF-κB signaling. Front Immunol 13:1007089.
- Zaffagni M, Harris JM, Patop IL, Reddy Pamudurti N, Nguyen S, Kadener S. SARS-CoV-2 Nsp14 mediates the effects of viral infection on the host cell Elife. 11:e71945.
- Tofaute MJ, Weller B, Graß C, Halder H, Dohai B, Falter-Braun P, Krappmann D. 2024. SARS-CoV-2 NSP14 MTase activity is critical for inducing canonical NF-κB activation. Biosci Rep 44(1):BSR20231418.
- Moeller NH, Passow KT, Harki DA, Aihara H. 2022. SARS-CoV-2 nsp14 Exoribonuclease Removes the Natural Antiviral 3′-Deoxy-3′,4′-didehydro-cytidine Nucleotide from RNA. Viruses 14(8):1790.
- Walter M, Chen IP, Vallejo-Gracia A, Kim I-J, Bielska O, Lam VL, Hayashi JM, Cruz A, Shah S, Soveg FW, Gross JD, Krogan NJ, Jerome KR, Schilling B, Ott M, Verdin E. 2022. SIRT5 is a proviral factor that interacts with SARS-CoV-2 Nsp14 protein. PLoS Pathog 18:e1010811.
- Zhang D, Ji L, Chen X, He Y, Sun Y, Ji L, Zhang T, Shen Q, Wang X, Wang Y, Yang S, Zhang W, Zhou C. 2023. SARS-CoV-2 Nsp15 suppresses type I interferon production by inhibiting IRF3 phosphorylation and nuclear translocation. iScience 26(9):107705.
- Otter CJ, Bracci N, Parenti NA, Ye C, Asthana A, Blomqvist EK, Tan LH, Pfannenstiel JJ, Jackson N, Fehr AR, Silverman RH, Burke JM, Cohen NA, Martinez-Sobrido L, Weiss SR. 2024. SARS-CoV-2 nsp15 endoribonuclease antagonizes dsRNA-induced antiviral signaling. Proc Natl Acad Sci U S A. 121(15):e2320194121.
- Wang X, Zhu B. 2024. SARS-CoV-2 nsp15 preferentially degrades AU-rich dsRNA via its dsRNA nickase activity. Nucleic Acids Res 52:5257–5272.
- Vithani N, Ward MD, Zimmerman MI, Novak B, Borowsky JH, Singh S, Bowman GR. 2021. SARS-CoV-2 Nsp16 activation mechanism and a cryptic pocket with pan-coronavirus antiviral potential. Biophys J 120:2880–2889.
- Russ A, Wittmann S, Tsukamoto Y, Herrmann A, Deutschmann J, Lagisquet J, Ensser A, Kato H, Gramberg T. 2022. Nsp16 shields SARS–CoV -2 from efficient MDA5 sensing and IFIT1 -mediated restriction . EMBO Rep. 23(12):e55648.
- Xu H, Akinyemi IA, Chitre SA, Loeb JC, Lednicky JA, McIntosh MT, Bhaduri-McIntosh S. 2022. SARS-CoV-2 viroporin encoded by ORF3a triggers the NLRP3 inflammatory pathway. Virology 568:13–22.
- Nie Y, Mou L, Long Q, Deng D, Hu R, Cheng J, Wu J. 2023. SARS-CoV-2 ORF3a positively regulates NF-κB activity by enhancing IKKβ-NEMO interaction. Virus Res 328:199086.
- Arshad N, Laurent-Rolle M, Ahmed WS, Hsu JCC, Mitchell SM, Pawlak J, Sengupta D, Biswas KH, Cresswell P. 2023. SARS-CoV-2 accessory proteins ORF7a and ORF3a use distinct mechanisms to down-regulate MHC-I surface expression. Proc Natl Acad Sci U S A 120(1):e2208525120.
- Ren Y, Shu T, Wu D, Mu J, Wang C, Huang M, Han Y, Zhang XY, Zhou W, Qiu Y, Zhou X. 2020. The ORF3a protein of SARS-CoV-2 induces apoptosis in cells. Cell Mol Immunol. 17(8):881-883.
- Stewart H, Palmulli R, Johansen KH, McGovern N, Shehata OM, Carnell GW, Jackson HK, Lee JS, Brown JC, Burgoyne T, Heeney JL, Okkenhaug K, Firth AE, Peden AA, Edgar JR. 2023. Tetherin antagonism by SARS-CoV -2 ORF3a and spike protein enhances virus release . EMBO Rep 24(12):e57224.
- Chen D, Zheng Q, Sun L, Ji M, Li Y, Deng H, Zhang H. 2021. ORF3a of SARS-CoV-2 promotes lysosomal exocytosis-mediated viral egress. Dev Cell 56:3250-3263.e5.
- Miao G, Zhao H, Li Y, Ji M, Chen Y, Shi Y, Bi Y, Wang P, Zhang H. 2021. ORF3a of the COVID-19 virus SARS-CoV-2 blocks HOPS complex-mediated assembly of the SNARE complex required for autolysosome formation. Dev Cell 56:427-442.e5.
- Zhang Y, Sun H, Pei R, Mao B, Zhao Z, Li H, Lin Y, Lu K. 2021. The SARS-CoV-2 protein ORF3a inhibits fusion of autophagosomes with lysosomes. Cell Discov 7(1):31.
- Walia K, Sharma A, Paul S, Chouhan P, Kumar G, Ringe R, Sharma M, Tuli A. 2024. SARS-CoV-2 virulence factor ORF3a blocks lysosome function by modulating TBC1D5-dependent Rab7 GTPase cycle. Nat Commun. 15(1):2053.
- Suleman M, Said A, Khan H, Rehman SU, Alshammari A, Crovella S, Yassine HM. 2024. Mutational analysis of SARS-CoV-2 ORF6-KPNA2 binding interface and identification of potent small molecule inhibitors to recuse the host immune system. Front Immunol 14:1266776.
- Addetia A, Lieberman NAP, Phung Q, Hsiang T-Y, Xie H, Roychoudhury P, Shrestha L, Loprieno MA, Huang M-L, Gale M, Jerome KR, Greninger AL. 2021. SARS-CoV-2 ORF6 Disrupts Bidirectional Nucleocytoplasmic Transport through Interactions with Rae1 and Nup98. mBio. 12(2):e00065-21.
- Miorin L, Kehrer T, Teresa Sanchez-Aparicio M, Zhang K, Cohen P, Patel RS, Cupic A, Makio T, Mei M, Moreno E, Danziger O, White KM, Rathnasinghe R, Uccellini M, Gao S, Aydillo T, Mena I, Yin X, Martin-Sancho L, Krogan NJ, Chanda SK, Schotsaert M, Wozniak RW, Ren Y, Rosenberg BR, A Fontoura BM, García-Sastre A. SARS-CoV-2 Orf6 hijacks Nup98 to block STAT nuclear import and antagonize interferon signaling. Proc Natl Acad Sci U S A. 117(45):28344-28354.
- Kehrer T, Cupic A, Ye C, Yildiz S, Bouhaddou M, Crossland NA, Barrall EA, Cohen P, Tseng A, Çağatay T, Rathnasinghe R, Flores D, Jangra S, Alam F, Mena I, Aslam S, Saqi A, Rutkowska M, Ummadi MR, Pisanelli G, Richardson RB, Veit EC, Fabius JM, Soucheray M, Polacco BJ, Ak B, Marin A, Evans MJ, Swaney DL, Gonzalez-Reiche AS, Sordillo EM, van Bakel H, Simon V, Zuliani-Alvarez L, Fontoura BMA, Rosenberg BR, Krogan NJ, Martinez-Sobrido L, García-Sastre A, Miorin L. 2023. Impact of SARS-CoV-2 ORF6 and its variant polymorphisms on host responses and viral pathogenesis. Cell Host Microbe 31:1668-1684.e12.
- Hall R, Guedan A, Yap MW, Young GR, Harvey R, Stoye JP, Bishop KN. 2022. SARS-CoV-2 ORF6 disrupts innate immune signalling by inhibiting cellular mRNA export. PLoS Pathog 18(8):e1010349.
- Miyamoto Y, Itoh Y, Suzuki T, Tanaka T, Sakai Y, Koido M, Hata C, Wang CX, Otani M, Moriishi K, Tachibana T, Kamatani Y, Yoneda Y, Okamoto T, Oka M. 2022. SARS-CoV-2 ORF6 disrupts nucleocytoplasmic trafficking to advance viral replication. Commun Biol 5(1):483.
- Khatun O, Sharma M, Narayan R, Tripathi S. 2023. SARS-CoV-2 ORF6 protein targets TRIM25 for proteasomal degradation to diminish K63-linked RIG-I ubiquitination and type-I interferon induction. Cellular and Molecular Life Sciences 80(12):364.
- López-Ayllón BD, de Lucas-Rius A, Mendoza-García L, García-García T, Fernández-Rodríguez R, Suárez-Cárdenas JM, Santos FM, Corrales F, Redondo N, Pedrucci F, Zaldívar-López S, Jiménez-Marín Á, Garrido JJ, Montoya M. 2023. SARS-CoV-2 accessory proteins involvement in inflammatory and profibrotic processes through IL11 signaling. Front Immunol 14:1220306.
- Cao Z, Xia H, Rajsbaum R, Xia X, Wang H, Shi PY. 2021. Ubiquitination of SARS-CoV-2 ORF7a promotes antagonism of interferon response. Cell Mol Immunol. 18(3):746-748.
- Liu Z, Fu Y, Huang Y, Zeng F, Rao J, Xiao X, Sun X, Jin H, Li J, Yang J, Du W, Liu L. 2022. Ubiquitination of SARS-CoV-2 ORF7a Prevents Cell Death Induced by Recruiting BclXL To Activate ER Stress. Microbiol Spectr 10(6):e0150922.
- Hou P, Wang X, Wang H, Wang T, Yu Z, Xu C, Zhao Y, Wang W, Zhao Y, Chu F, Chang H, Zhu H, Lu J, Zhang F, Liang X, Li X, Wang S, Gao Y, He H. 2023. The ORF7a protein of SARS-CoV-2 initiates autophagy and limits autophagosome-lysosome fusion via degradation of SNAP29 to promote virus replication. Autophagy 19:551–569.
- Timilsina U, Umthong S, Ivey EB, Waxman B, Stavrou S. 2022. SARS-CoV-2 ORF7a potently inhibits the antiviral effect of the host factor SERINC5. Nat Commun 13(1):2935.
- Yang R, Zhao Q, Rao J, Zeng F, Yuan S, Ji M, Sun X, Li J, Yang J, Cui J, Jin Z, Liu L, Liu Z. 2021. SARS-CoV-2 Accessory Protein ORF7b Mediates Tumor Necrosis Factor-α-Induced Apoptosis in Cells. Front Microbiol 12:654709.
- García-García T, Fernández-Rodríguez R, Redondo N, de Lucas-Rius A, Zaldívar-López S, López-Ayllón BD, Suárez-Cárdenas JM, Jiménez-Marín Á, Montoya M, Garrido JJ. 2022. Impairment of antiviral immune response and disruption of cellular functions by SARS-CoV-2 ORF7a and ORF7b. iScience 25(11):105444.
- Xiao X, Fu Y, You W, Huang C, Zeng F, Gu X, Sun X, Li J, Zhang Q, Du W, Cheng G, Liu Z, Liu L. 2024. Inhibition of the RLR signaling pathway by SARS-CoV-2 ORF7b is mediated by MAVS and abrogated by ORF7b-homologous interfering peptide. J Virol 98(5):e0157323.
- Zhang Y, Chen Y, Li Y, Huang F, Luo B, Yuan Y, Xia B, Ma X, Yang T, Yu F, Liu J, Liu B, Song Z, Chen J, Yan S, Wu L, Pan T, Zhang X, Li R, Huang W, He X, Xiao F, Zhang J, Zhang H. The ORF8 protein of SARS-CoV-2 mediates immune evasion through down-regulating MHC-Ι. Proc Natl Acad Sci U S A. 118(23):e2024202118.
- Beaudoin-Bussières G, Arduini A, Bourassa C, Medjahed H, Gendron-Lepage G, Richard J, Pan Q, Wang Z, Liang C, Finzi A. 2022. SARS-CoV-2 Accessory Protein ORF8 Decreases Antibody-Dependent Cellular Cytotoxicity. Viruses 14(6):1237.
- Móvio MI, Almeida GWC de, Martines I das GL, Barros de Lima G, Sasaki SD, Kihara AH, Poole E, Nevels M, Carlan da Silva MC. 2024. SARS-CoV-2 ORF8 as a Modulator of Cytokine Induction: Evidence and Search for Molecular Mechanisms. Viruses. 16(1):161.
- Kumar J, Dhyani S, Kumar P, Sharma NR, Ganguly S. 2023. SARS-CoV-2–encoded ORF8 protein possesses complement inhibitory properties. J Biol Chem. 299(3):102930.
- Arduini A, Laprise F, Liang C. 2023. SARS-CoV-2 ORF8: A Rapidly Evolving Immune and Viral Modulator in COVID-19. Viruses. 15(4):871.
- Gao X, Zhu K, Qin B, Olieric V, Wang M, Cui S. 2021. Crystal structure of SARS-CoV-2 Orf9b in complex with human TOM70 suggests unusual virus-host interactions. Nat Commun 12(1):2843.
- Wu J, Shi Y, Pan X, Wu S, Hou R, Zhang Y, Zhong T, Tang H, Du W, Wang L, Wo J, Mu J, Qiu Y, Yang K, Zhang LK, Ye BC, Qi N. 2021. SARS-CoV-2 ORF9b inhibits RIG-I-MAVS antiviral signaling by interrupting K63-linked ubiquitination of NEMO. Cell Rep 34(7):108761.
- Han L, Zhuang MW, Deng J, Zheng Y, Zhang J, Nan ML, Zhang XJ, Gao C, Wang PH. 2021. SARS-CoV-2 ORF9b antagonizes type I and III interferons by targeting multiple components of the RIG-I/MDA-5–MAVS, TLR3–TRIF, and cGAS–STING signaling pathways. J Med Virol 93:5376–5389.
- Zhang P, Liu Y, Li C, Stine LD, Wang PH, Turnbull MW, Wu H, Liu Q. 2023. Ectopic expression of SARS-CoV-2 S and ORF-9B proteins alters metabolic profiles and impairs contractile function in cardiomyocytes. Front Cell Dev Biol 11:1110271.
- Homma D, Limlingan SJM, Saito T, Ando K. 2024. SARS-CoV-2-derived protein Orf9b enhances MARK2 activity via interaction with the autoinhibitory KA1 domain. FEBS Lett 598(19):2385-2393.
- Sarvari J, Jalili S, Mohammad S, Hashemi A. 2024. SARS-COV-2 ORF9b Dysregulate Fibrinogen and Albumin Genes in a Liver Cell Line. Rep Biochem Mol Biol. 13(1):51-58.
- Li X, Hou P, Ma W, Wang X, Wang H, Yu Z, Chang H, Wang T, Jin S, Wang X, Wang W, Zhao Y, Zhao Y, Xu C, Ma X, Gao Y, He H. 2022. SARS-CoV-2 ORF10 suppresses the antiviral innate immune response by degrading MAVS through mitophagy. Cell Mol Immunol 19:67–78.
- Han L, Zheng Y, Deng J, Nan ML, Xiao Y, Zhuang MW, Zhang J, Wang W, Gao C, Wang PH. 2022. SARS-CoV-2 ORF10 antagonizes STING-dependent interferon activation and autophagy. J Med Virol 94:5174–5188.
- Schubert K, Karousis ED, Jomaa A, Scaiola A, Echeverria B, Gurzeler LA, Leibundgut M, Thiel V, Mühlemann O, Ban N. 2020. SARS-CoV-2 Nsp1 binds the ribosomal mRNA channel to inhibit translation. Nat Struct Mol Biol 27:959–966.
- Korneeva N, Khalil MI, Ghosh I, Fan R, Arnold T, De Benedetti A. 2023. SARS-CoV-2 viral protein Nsp2 stimulates translation under normal and hypoxic conditions. Virol J 20(1):55.
- Thorne LG, Bouhaddou M, Reuschl AK, Zuliani-Alvarez L, Polacco B, Pelin A, Batra J, Whelan MVX, Hosmillo M, Fossati A, Ragazzini R, Jungreis I, Ummadi M, Rojc A, Turner J, Bischof ML, Obernier K, Braberg H, Soucheray M, Richards A, Chen KH, Harjai B, Memon D, Hiatt J, Rosales R, McGovern BL, Jahun A, Fabius JM, White K, Goodfellow IG, Takeuchi Y, Bonfanti P, Shokat K, Jura N, Verba K, Noursadeghi M, Beltrao P, Kellis M, Swaney DL, García-Sastre A, Jolly C, Towers GJ, Krogan NJ. 2022. Evolution of enhanced innate immune evasion by SARS-CoV-2. Nature 602:487–495.
- Takata MA, Gonçalves-Carneiro D, Zang TM, Soll SJ, York A, Blanco-Melo D, Bieniasz PD. 2017. CG dinucleotide suppression enables antiviral defence targeting non-self RNA. Nature 550:124–127.
- Malone B, Urakova N, Snijder EJ, Campbell EA. 2022. Structures and functions of coronavirus replication–transcription complexes and their relevance for SARS-CoV-2 drug design. Nat Rev Mol Cell Biol 23:21–39.
- Hagelauer E, Lotke R, Kmiec D, Hu D, Hohner M, Stopper S, Nchioua R, Kirchhoff F, Sauter D, Schindler M. 2023. Tetherin Restricts SARS-CoV-2 despite the Presence of Multiple Viral Antagonists. Viruses 15:2364.
- Bills C, Xie X, Shi PY. 2023. The multiple roles of nsp6 in the molecular pathogenesis of SARS-CoV-2. Antiviral Res. 213:105590.
- Krachmarova E, Petkov P, Lilkova E, Ilieva N, Rangelov M, Todorova N, Malinova K, Hristova R, Nacheva G, Gospodinov A, Litov L. 2023. Insights into the SARS-CoV-2 ORF6 Mechanism of Action. Int J Mol Sci 24(14):11589.
- Zheng M, Karki R, Williams EP, Yang D, Fitzpatrick E, Vogel P, Jonsson CB, Kanneganti TD. 2021. TLR2 senses the SARS-CoV-2 envelope protein to produce inflammatory cytokines. Nat Immunol 22:829–838.
- Yu H, Yang L, Han Z, Zhou X, Zhang Z, Sun T, Zheng F, Yang J, Guan F, Xie J, Liu C. 2023. SARS-CoV-2 nucleocapsid protein enhances the level of mitochondrial reactive oxygen species. J Med Virol 95(12):e29270.
- Kohyama M, Suzuki T, Nakai W, Ono C, Matsuoka S, Iwatani K, Liu Y, Sakai Y, Nakagawa A, Tomii K, Ohmura K, Okada M, Matsuura Y, Ohshima S, Maeda Y, Okamoto T, Arase H. SARS-CoV-2 ORF8 is a viral cytokine regulating immune responses. Int Immunol. 35(1):43-52.
- Fajgenbaum DC, June CH. 2020. Cytokine Storm. New England Journal of Medicine 383:2255–2273.
- Lee MJ, Leong MW, Rustagi A, Beck A, Zeng L, Holmes S, Qi LS, Blish CA. 2022. SARS-CoV-2 escapes direct NK cell killing through Nsp1-mediated downregulation of ligands for NKG2D. Cell Rep 41(13):111892.
- Freda CT, Yin W, Ghebrehiwet B, Rubenstein DA. 2022. SARS-CoV-2 Structural Proteins Exposure Alter Thrombotic and Inflammatory Responses in Human Endothelial Cells. Cell Mol Bioeng 15:43–53.
- Lei X, Dong X, Ma R, Wang W, Xiao X, Tian Z, Wang C, Wang Y, Li L, Ren L, Guo F, Zhao Z, Zhou Z, Xiang Z, Wang J. 2020. Activation and evasion of type I interferon responses by SARS-CoV-2. Nat Commun 11(1):3810.
- Jeong H. 2023. Ion channels activity of SARS-CoV-2 envelope protein through calcium influx assay using large unilamellar vesicles. Biophys J 122:372a.
- Bugatti A, Filippini F, Bardelli M, Zani A, Chiodelli P, Messali S, Caruso A, Caccuri F. 2022. SARS-CoV-2 Infects Human ACE2-Negative Endothelial Cells through an αvβ3 Integrin-Mediated Endocytosis Even in the Presence of Vaccine-Elicited Neutralizing Antibodies. Viruses 14:705.
- Schimmel L, Chew KY, Stocks CJ, Yordanov TE, Essebier P, Kulasinghe A, Monkman J, dos Santos Miggiolaro AFR, Cooper C, de Noronha L, Schroder K, Lagendijk AK, Labzin LI, Short KR, Gordon EJ. 2021. Endothelial cells are not productively infected by SARS-CoV-2. Clin Transl Immunology 10(10):e1350.
- Jiang H wei, Zhang H nan, Meng Q feng, Xie J, Li Y, Chen H, Zheng Y xiao, Wang X ning, Qi H, Zhang J, Wang PH, Han ZG, Tao S ce. 2020. SARS-CoV-2 Orf9b suppresses type I interferon responses by targeting TOM70. Cell Mol Immunol. 17(9):998-1000.
- Gao W, Wang L, Ju X, Zhao S, Li Z, Su M, Xu J, Wang P, Ding Q, Lv G, Zhang W. 2022. The Deubiquitinase USP29 Promotes SARS-CoV-2 Virulence by Preventing Proteasome Degradation of ORF9b. mBio 13(3):e0130022.
- Lenhard S, Gerlich S, Khan A, Rödl S, Bökenkamp JE, Peker E, Zarges C, Faust J, Storchova Z, Räschle M, Riemer J, Herrmann JM. 2023. The Orf9b protein of SARS-CoV-2 modulates mitochondrial protein biogenesis. J Cell Biol. 222(10):e202303002.
- Hirschenberger M, Hayn M, Laliberté A, Koepke L, Kirchhoff F, Sparrer KMJ. 2021. Luciferase reporter assays to monitor interferon signaling modulation by SARS-CoV-2 proteins. STAR Protoc 2(4):100781.
- Baudin B, Bruneel A, Bosselut N, Vaubourdolle M. 2007. A protocol for isolation and culture of human umbilical vein endothelial cells. Nat Protoc 2:481–485.
- Gordon DE, Jang GM, Bouhaddou M, Xu J, Obernier K, White KM, O’Meara MJ, Rezelj V V., Guo JZ, Swaney DL, et al.. 2020. A SARS-CoV-2 protein interaction map reveals targets for drug repurposing. Nature 583:459–468.
- Bouhaddou M, Memon D, Meyer B, White KM, Rezelj V V., Correa Marrero M, Polacco BJ, Melnyk JE, Ulferts S, Kaake RM, et al.. 2020. The Global Phosphorylation Landscape of SARS-CoV-2 Infection. Cell 182:685-712.e19.
- Vermeire J, Naessens E, Vanderstraeten H, Landi A, Iannucci V, van Nuffel A, Taghon T, Pizzato M, Verhasselt B. 2012. Quantification of Reverse Transcriptase Activity by Real-Time PCR as a Fast and Accurate Method for Titration of HIV, Lenti- and Retroviral Vectors. PLoS One. 7(12):e50859.
Figure 1.
Affected steps of immune sensing by SARS-CoV-2 proteins (Wuhan strain) based on literature search (Table 1 and S1). SARS-CoV-2 proteins inhibit type I interferon (IFN-I) responses at multiple stages. Left panel: IFN-I induction inhibition, including sensing (Nsp14, Nsp15, Nsp16, N, Nsp3), IRF3 activation (Nsp15), downstream signaling (ORF9b, M, ORF6, ORF3b), and IFN-I expression (Nsp1). Right panel: IFN-I signaling inhibition, targeting IFNAR binding (ORF3a), STAT1 activation (Nsp1), and nuclear translocation of STAT1 (ORF6). These mechanisms highlight the multi-faceted immune evasion strategies of SARS-CoV-2. For references see Table 1.
Figure 1.
Affected steps of immune sensing by SARS-CoV-2 proteins (Wuhan strain) based on literature search (Table 1 and S1). SARS-CoV-2 proteins inhibit type I interferon (IFN-I) responses at multiple stages. Left panel: IFN-I induction inhibition, including sensing (Nsp14, Nsp15, Nsp16, N, Nsp3), IRF3 activation (Nsp15), downstream signaling (ORF9b, M, ORF6, ORF3b), and IFN-I expression (Nsp1). Right panel: IFN-I signaling inhibition, targeting IFNAR binding (ORF3a), STAT1 activation (Nsp1), and nuclear translocation of STAT1 (ORF6). These mechanisms highlight the multi-faceted immune evasion strategies of SARS-CoV-2. For references see Table 1.
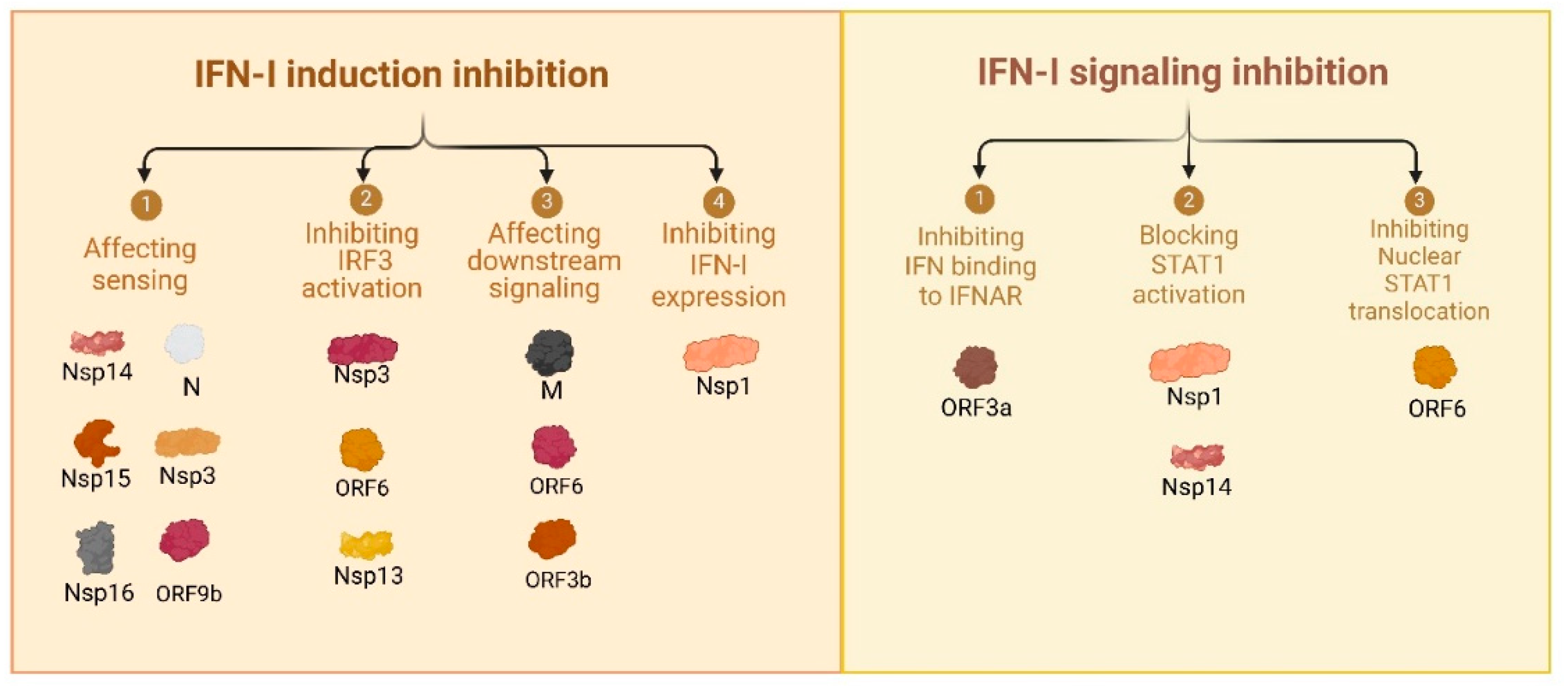
Figure 2.
Mechanisms of Endothelial Dysfunction in COVID-19. The figure illustrates the key mechanisms driving endothelial dysfunction in COVID-19 and their downstream effects on the vascular system that can lead to fatal organ damage. A. Direct SARS-CoV-2 infection: The viral spike protein binds to ACE2 receptors expressed on endothelial cells, facilitating viral entry and replication, leading to cellular damage [15,16,17]. B. Systemic inflammation: Elevated pro-inflammatory cytokines, including IL-6, IL-1β, and TNF-α, activate endothelial cells, inducing an amplified inflammatory response [18,19,20,21]. C. Hypercoagulable state: Endothelial injury promotes thrombin generation and platelet aggregation, resulting in the formation of thrombi and widespread vascular occlusion [12,22]. D. Hypoxia: Reduced oxygen delivery due to severe respiratory distress exacerbates endothelial dysfunction, further impairing tissue oxygenation [23,24]. E. Complement activation: Overactivation of the complement cascade causes endothelial damage and contributes to pro-thrombotic states through the generation of C3 and C5 convertases [17,25,26].
Figure 2.
Mechanisms of Endothelial Dysfunction in COVID-19. The figure illustrates the key mechanisms driving endothelial dysfunction in COVID-19 and their downstream effects on the vascular system that can lead to fatal organ damage. A. Direct SARS-CoV-2 infection: The viral spike protein binds to ACE2 receptors expressed on endothelial cells, facilitating viral entry and replication, leading to cellular damage [15,16,17]. B. Systemic inflammation: Elevated pro-inflammatory cytokines, including IL-6, IL-1β, and TNF-α, activate endothelial cells, inducing an amplified inflammatory response [18,19,20,21]. C. Hypercoagulable state: Endothelial injury promotes thrombin generation and platelet aggregation, resulting in the formation of thrombi and widespread vascular occlusion [12,22]. D. Hypoxia: Reduced oxygen delivery due to severe respiratory distress exacerbates endothelial dysfunction, further impairing tissue oxygenation [23,24]. E. Complement activation: Overactivation of the complement cascade causes endothelial damage and contributes to pro-thrombotic states through the generation of C3 and C5 convertases [17,25,26].
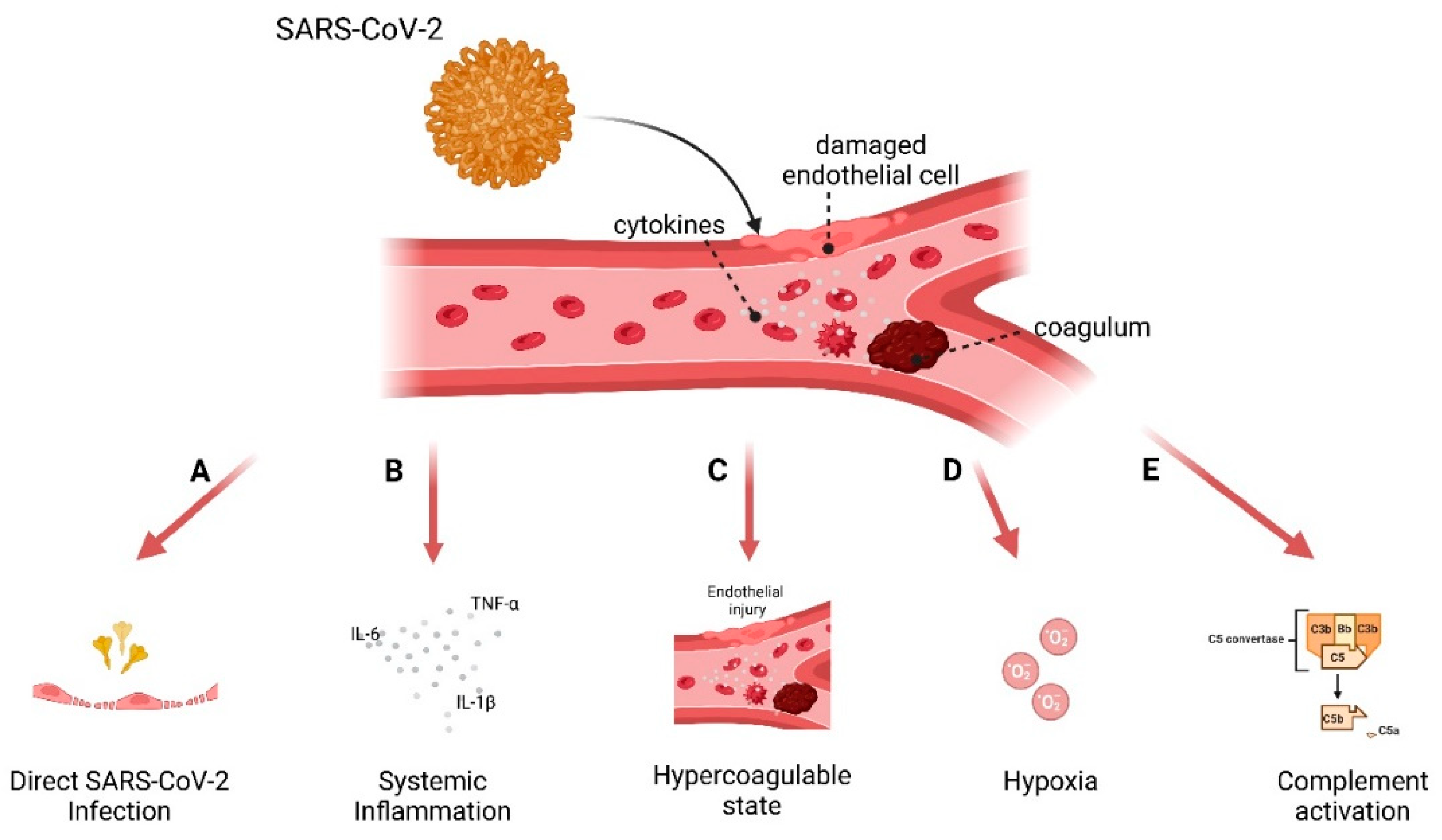
Figure 3.
Functional Screening of SARS-CoV-2 Proteins from Wuhan and Omicron BA.1 variants on their ability to suppress or induce immune response. A Schematic representation of the experimental workflow. HEK293T cells were co-transfected with Firefly luciferase reporter plasmid under the control of an ISRE promoter (pISRE-F-luc), a Renilla luciferase plasmid (pGAPDH-R-luc) as a control for transfection efficiency, SARS-CoV-2 protein expression plasmids encoding individual SARS-CoV-2 proteins from the Wuhan (B, D) or Omicron BA.1 (C, E) variants, and stimulated with IFN-β or IFN-α2. Firefly luciferase activity was measured using a Dual Luciferase Reporter Assay and normalized to Renilla activity and cell viability. Data represent the mean ± SEM from three independent experiments, each including two technical replicates. Cells were stimulated with IFN-β (B, C) or IFN-α2 (D, E). Results are presented as fold-change in relative luminescence units (RLU) compared to fully stimulated ISRE promoter activity in cells co-transfected with an empty control vector (dotted lines). Panels B and C: IFN-β-stimulated ISRE promoter activity. Both Wuhan and Omicron BA.1 proteins show a range of inhibitory effects, with certain proteins (e.g., Nsp1, Nsp14) inducing significant suppression of promoter activity. Panels D and E: IFN-α2-stimulated ISRE promoter activity. Similar patterns of suppression are observed for both variants, with Nsp1 and ORF9b showing the strongest inhibition of ISRE-driven luciferase expression.
Figure 3.
Functional Screening of SARS-CoV-2 Proteins from Wuhan and Omicron BA.1 variants on their ability to suppress or induce immune response. A Schematic representation of the experimental workflow. HEK293T cells were co-transfected with Firefly luciferase reporter plasmid under the control of an ISRE promoter (pISRE-F-luc), a Renilla luciferase plasmid (pGAPDH-R-luc) as a control for transfection efficiency, SARS-CoV-2 protein expression plasmids encoding individual SARS-CoV-2 proteins from the Wuhan (B, D) or Omicron BA.1 (C, E) variants, and stimulated with IFN-β or IFN-α2. Firefly luciferase activity was measured using a Dual Luciferase Reporter Assay and normalized to Renilla activity and cell viability. Data represent the mean ± SEM from three independent experiments, each including two technical replicates. Cells were stimulated with IFN-β (B, C) or IFN-α2 (D, E). Results are presented as fold-change in relative luminescence units (RLU) compared to fully stimulated ISRE promoter activity in cells co-transfected with an empty control vector (dotted lines). Panels B and C: IFN-β-stimulated ISRE promoter activity. Both Wuhan and Omicron BA.1 proteins show a range of inhibitory effects, with certain proteins (e.g., Nsp1, Nsp14) inducing significant suppression of promoter activity. Panels D and E: IFN-α2-stimulated ISRE promoter activity. Similar patterns of suppression are observed for both variants, with Nsp1 and ORF9b showing the strongest inhibition of ISRE-driven luciferase expression.
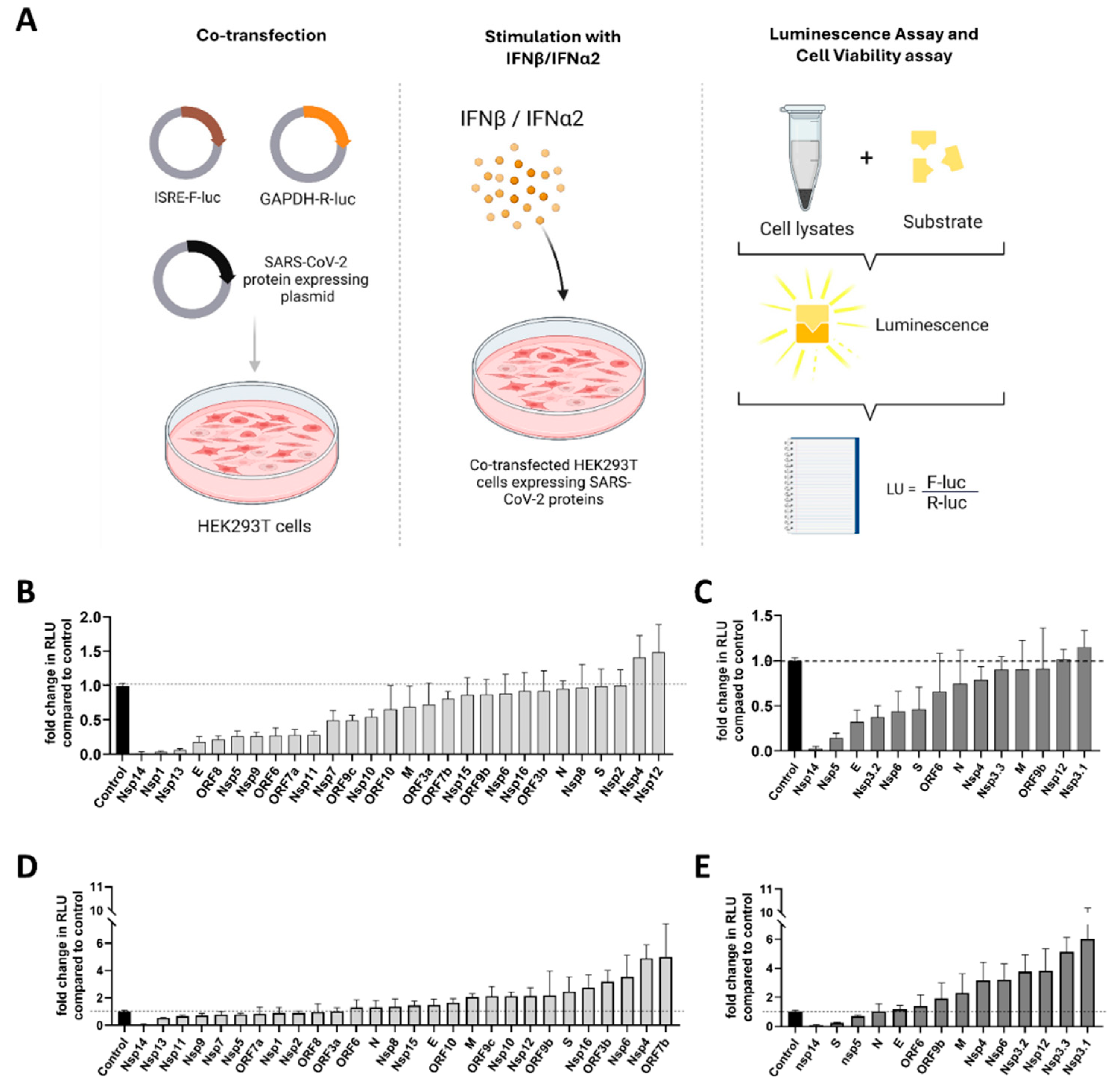
Figure 4.
Cytotoxicity of individual SARS-CoV-2 proteins from Wuhan and Omicron BA.1 variants. HEK293T cells were transfected with expression vectors encoding individual SARS-CoV-2 proteins from the Wuhan (A, C) or Omicron BA.1 (B, D) variants. Cell viability was measured at 8 hours (A, B) and 24 hours (C, D) post-transfection using the CellTiter-Glo® Luminescent Cell Viability Assay. The luminescent signal reflects ATP levels, proportional to viable cells (Supplemental material). Fold-change in luminescence activity (absolute luminescent units (ALU), see Figure S1) was calculated compared to an empty vector controls (n=2 for each experiment), included to account for transfection effects. Panels A and C show cytotoxicity profiles of individual Wuhan variant proteins. Minimal reductions in cell viability were observed at 8 hours post-transfection (A), while some proteins exhibited mild to moderate toxicity by 24 hours (C). Panels B and D show cytotoxicity profiles of individual Omicron BA.1 variant proteins. Similar to the Wuhan variant, most proteins showed minimal toxicity at 8 hours (B). At 24 hours (D), a subset of proteins induced moderate reductions in cell viability compared to the empty vector control. Data represent the mean ± SEM of fold-change in luminescence activity compared to control from three independent experiments, each with two technical replicates.
Figure 4.
Cytotoxicity of individual SARS-CoV-2 proteins from Wuhan and Omicron BA.1 variants. HEK293T cells were transfected with expression vectors encoding individual SARS-CoV-2 proteins from the Wuhan (A, C) or Omicron BA.1 (B, D) variants. Cell viability was measured at 8 hours (A, B) and 24 hours (C, D) post-transfection using the CellTiter-Glo® Luminescent Cell Viability Assay. The luminescent signal reflects ATP levels, proportional to viable cells (Supplemental material). Fold-change in luminescence activity (absolute luminescent units (ALU), see Figure S1) was calculated compared to an empty vector controls (n=2 for each experiment), included to account for transfection effects. Panels A and C show cytotoxicity profiles of individual Wuhan variant proteins. Minimal reductions in cell viability were observed at 8 hours post-transfection (A), while some proteins exhibited mild to moderate toxicity by 24 hours (C). Panels B and D show cytotoxicity profiles of individual Omicron BA.1 variant proteins. Similar to the Wuhan variant, most proteins showed minimal toxicity at 8 hours (B). At 24 hours (D), a subset of proteins induced moderate reductions in cell viability compared to the empty vector control. Data represent the mean ± SEM of fold-change in luminescence activity compared to control from three independent experiments, each with two technical replicates.
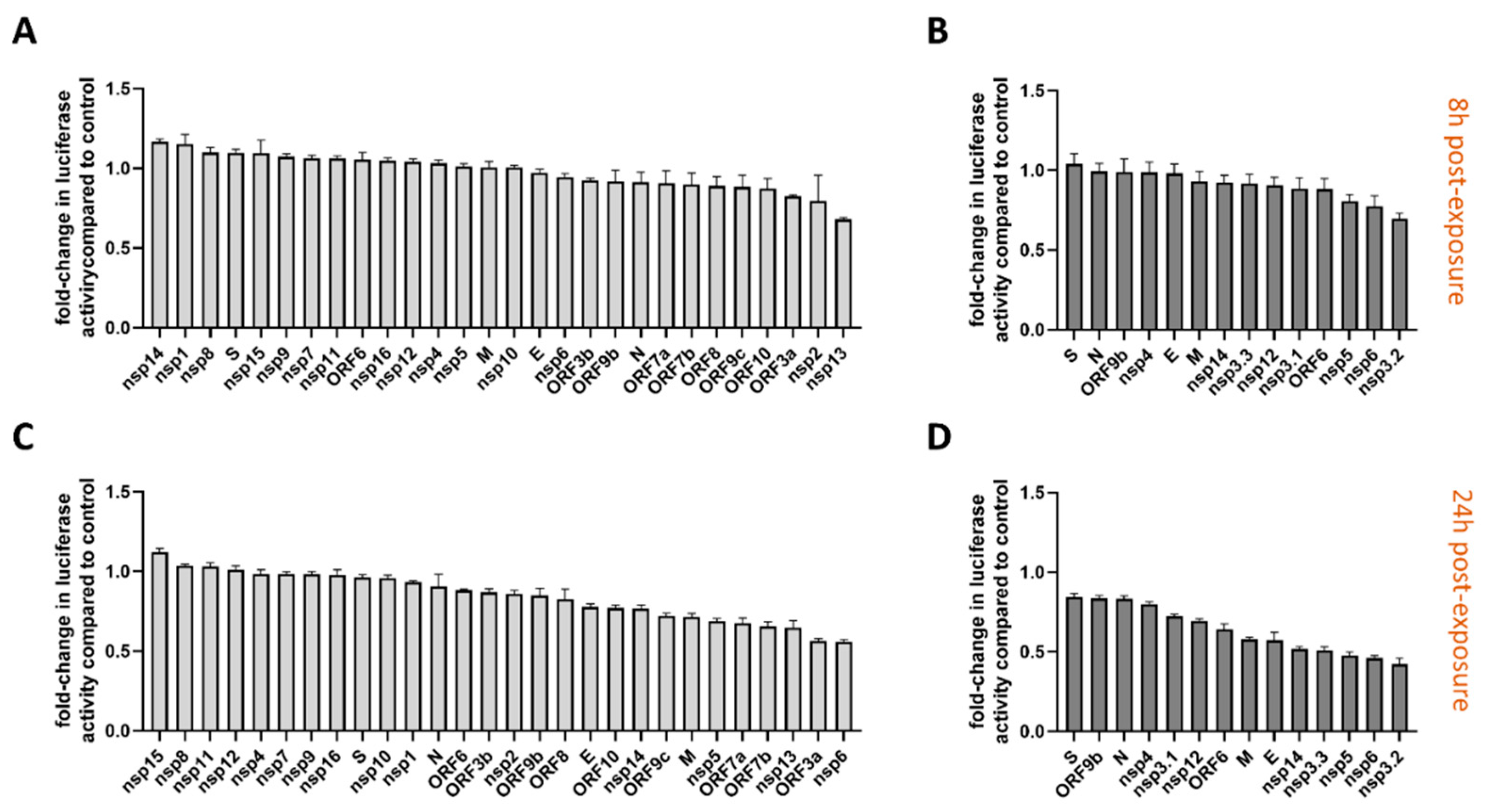
Figure 5.
The effect of SARS-CoV-2 Envelope, Membrane, and ORF9b proteins from Wuhan and Omicron BA.1 variants on IFN-I response in HUVEC cells, with or without IFN stimulation. HUVEC cells were transduced with lentiviruses encoding the Envelope (A), Membrane (B), or ORF9b (C) proteins from the SARS-CoV-2 Wuhan (light grey bars) or Omicron BA.1 variants (dark grey bars). Non-transduced (NTD) cells were used as controls (black bars). Cells were either non-stimulated or stimulated with IFN-β or IFN-α2 for 24 hours. Expression of IFIT1 (left column), IFNB1 (middle column), and IFNA2 (right column) was measured by qPCR. Fold-change in gene expression levels was calculated relative to non-transduced, non-stimulated cells. Where visible, error bars represent mean ± SEM from two independent experiments. The data are obtained with six independent biological donors.
Figure 5.
The effect of SARS-CoV-2 Envelope, Membrane, and ORF9b proteins from Wuhan and Omicron BA.1 variants on IFN-I response in HUVEC cells, with or without IFN stimulation. HUVEC cells were transduced with lentiviruses encoding the Envelope (A), Membrane (B), or ORF9b (C) proteins from the SARS-CoV-2 Wuhan (light grey bars) or Omicron BA.1 variants (dark grey bars). Non-transduced (NTD) cells were used as controls (black bars). Cells were either non-stimulated or stimulated with IFN-β or IFN-α2 for 24 hours. Expression of IFIT1 (left column), IFNB1 (middle column), and IFNA2 (right column) was measured by qPCR. Fold-change in gene expression levels was calculated relative to non-transduced, non-stimulated cells. Where visible, error bars represent mean ± SEM from two independent experiments. The data are obtained with six independent biological donors.
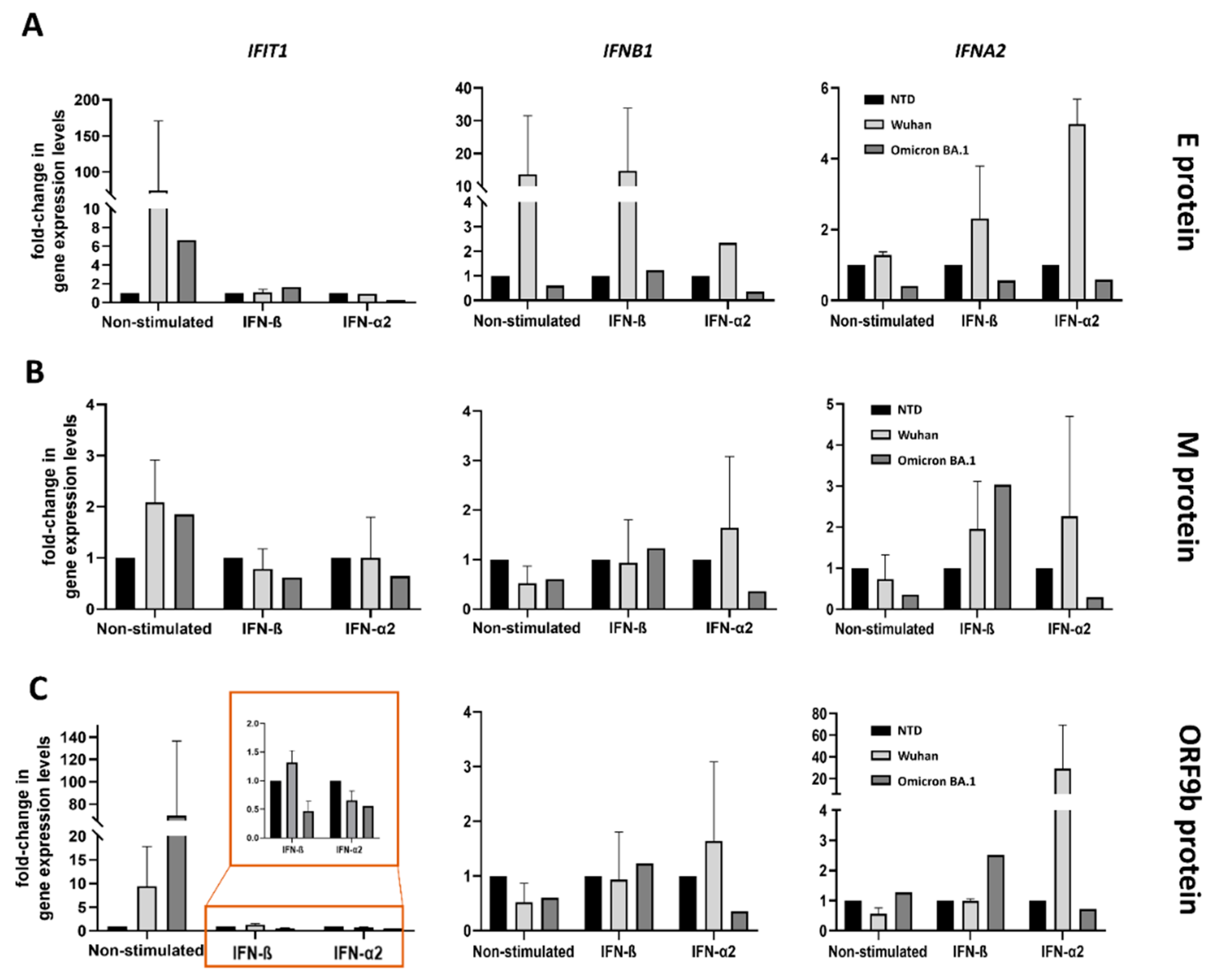

Table 1.
SARS-CoV-2 Wuhan strain viral protein’s effect on host.
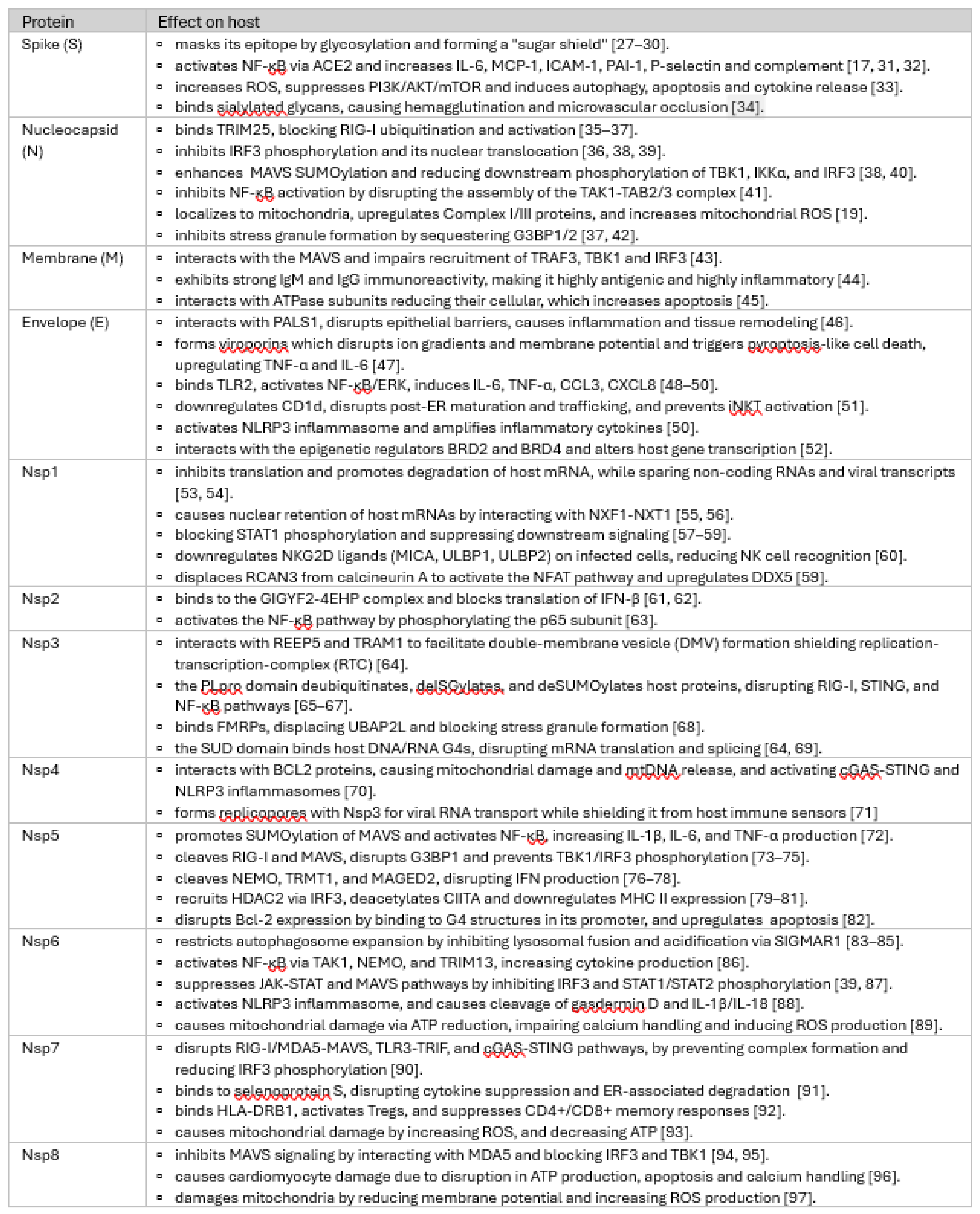 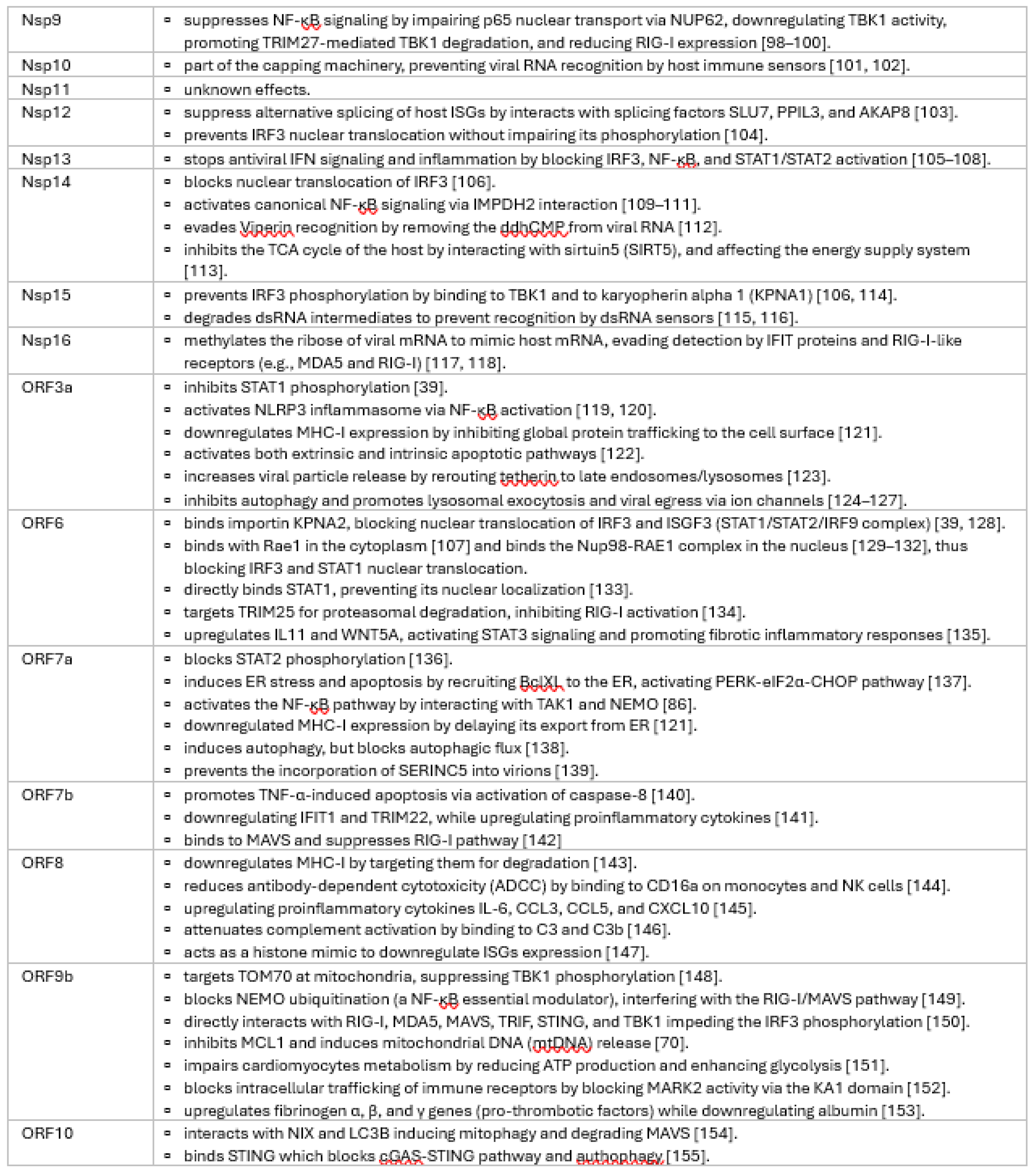
|
Table 2.
Immune evasion strategies by SARS-CoV-2 Wuhan strain.
 
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



