Submitted:
14 April 2025
Posted:
15 April 2025
Read the latest preprint version here
Abstract
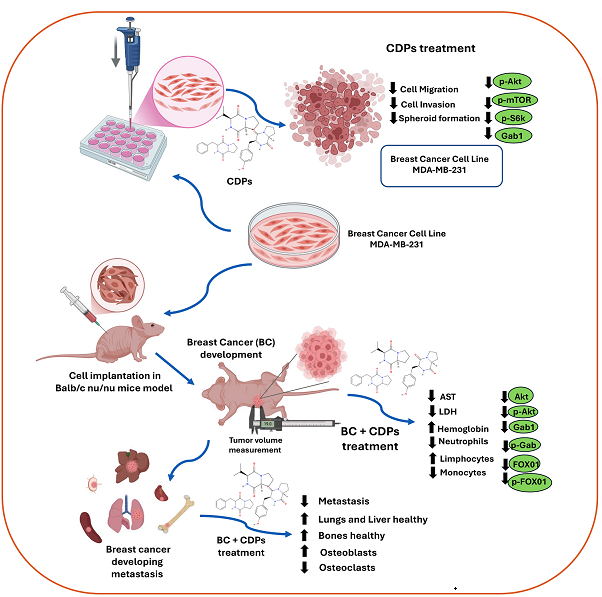
Background: Triple-negative breast cancer (TNBC) is the leading cause of death from neoplasms in women worldwide, related to a high rate of metastasis and low survival in patients. Bacterial cyclodipeptides (CDPs) have anticancer properties in several types of cancer in vitro as in vivo models, targeting several signaling pathways. Methods: The effect of the CDPs on TNBC MDA-MB-231 lines was evaluated in vitro as advanced-stage tumors developed and their ability to prevent the appearance of metastatic foci in the mammary tissue of mice model and bone structuration. Results: CDPs treatment decreased the migratory and invasive capacity of the TNBC MDA-MB-231 line in vitro more efficiently than methotrexate (MTX) (p>0.001). The anti-metastatic effect in the TNBC was associated with the down-regulation of the Akt/mTOR/S6K pathway and the metastasis markers such as Gab1 and Vimentin. MDA-MB-231 xenografted mice administrated with CDPs and combined with MTX showed a significant decrease in primary tumor development and metastatic foci (p>0.001). Likewise, the metastatic foci were observed in the lungs and liver, which were inhibited by CDPs. Metastasis markers p-Akt, Gab1, and FOX01 were significantly down-expressed in the tumor tissue of mice implanted with TNBC submitted to CDPs treatment. In addition, the metastatic behavior in the skeletal system was alleviated with the CDPs and CDPs+MTX treatments. Conclusions: The anti-metastatic effects of bacterial CDPs involve the phosphorylation inhibition of the Akt/mTOR/S6K pathway and the metastasis markers Gab1, Vimentin, and FOX01. The finding indicates that the CDPs, in combination with MTX, potentialize the anti-neoplastic property in TNBC, proposing the CDPs as a viable alternative in human breast cancer therapy.
Keywords:
Cyclodipeptides
; anti-metastatic
; Triple-negative breast cancer
; invasiveness
; metastasis
; immunosuppression mice model
1. Introduction
Breast cancer is the principal cause of death in women worldwide; it is a clinically heterogeneous disease, and around 10-15% of patients present aggressive phenotypes and develop metastasis [1]. Triple-negative breast cancer (TNBC) comprises a group of diseases with different histological, genomic, and immunological profiles, characterized by lacking hormonal estrogen and progesterone receptors, as well as human epidermal growth factor receptor 2 (HER2). According to postulate in the “soil and seed” theory, tumor cells thrive in those tissues with genetic and metabolic characteristics similar to their needs. For example, breast cancer cells tend to metastasize the bone, lung, and liver [2]. Other factors in developing breast cancer, such as obesity, represent a poor prognosis for patients since the interaction of adipocytes with tumor cells provides an optimal microenvironment that favors proliferation, invasion, metastasis, or resistance to cell death [3]. The tumor microenvironment also recruits the malignant cells with other types of cells, such as adipocytes, mesenchymal-type cells, such as fibroblasts, and the cells of the immune system, such as macrophages, T-lymphocytes, and neutrophils. These interactions are mediated by the secretion of chemokines, interleukins, or growth factors, which promote proliferation, invasion, and resistance to cell death, positioning these components as possible new therapeutic strategies [4].
Among the first-line therapeutic options for treating metastatic breast cancer are taxanes, anthracyclines, methotrexate, carboplatin, PD-1 or PD-L1 inhibitors, PARP inhibitors, and their combination with immunotherapy, radiotherapy, and surgery [5]. However, a significant percentage of patients negatively respond to these treatments, resulting in low survival and poor quality of life. Therefore, the search for new therapeutic options that improve the prognosis for patients with metastatic breast cancer is of great importance.
Bone metastasis is a common complication in advanced-stage breast and prostate cancer, disrupting the bone remodeling cycle mediated by osteoblasts and osteoclasts [6]. Currently, available T cell-based immunotherapies show promise in the treatment of cancer; they are often ineffective in patients with advanced stages of cancer that include bone metastases [2]. Metastases predominantly occur in trabecular and red marrow-rich bones, suggesting that specific bone environments favor metastatic growth [7]. Once bone metastases develop, the five-year survival rate drops to approximately 20% [8]. Bone metastases significantly impact patient health, yet current treatments, including bisphosphonates and antiresorptive antibodies, remain insufficient; therefore, develop of novel therapies that target both tumor cells and the bone microenvironment with dual antiresorptive (reducing bone remodeling) and anticancer properties are required [9].
Cyclodipeptides (CDPs) are a class of molecules synthesized by a broad type of organisms, which possess interesting biological activities such as cytotoxic effects in different cancer cell lines [10,11,12]. Previously, we described that CDPs, cyclo(L-Pro-L-Tyr), cyclo(L-Pro-L-Val), and cyclo(L-Pro-L-Phe) isolated from the Pseudomonas aeruginosa PAO1 bacterium induce apoptosis in cervical, colon, leukemia, and melanoma cancer cells by inhibiting phosphorylation of multiple kinases from the PI3K/Akt/mTOR pathway, including mTORC1/C2 complexes [11]. Furthermore, a significant decrease in tumors in mice treated with CDPs at 0.1 mg/g of weight was found in a mouse murine melanoma model. Additionally, treatment with CDPs led to the restoration of hematological parameters and a decrease in tissue damage markers AST and ALT, as well as a marked reduction in the expression of proteins that mediate pathways involved in energy metabolism, lipid synthesis, epithelial-mesenchymal transition, invasion, and metastasis [13]. Interestingly, transcriptomic analysis in HeLa cells found that CDPs inhibit gene expression in the mevalonate pathway and lipid synthesis [14]. Given this background, we sought to determine whether CDPs could have a promising effect on breast cancer and whether this effect could be observed in advanced stages of tumorigenesis [15].
In this work, we first used monolayer culture to evaluate the migratory and invasive potential of the MDA-MB-231 human triple-negative breast cancer cell line treated with CDPs and combined with Methotrexate (MTX). Further, the effect of the CDPs on advanced-stage mammary tumors developed, and their ability to prevent the appearance of metastatic foci through the implantation of the MDA-MB-231 cells in the mammary tissue of mice model was evaluated.
2. Materials and Methods
2.1. Chemicals, Reagents, and Cell Culture
Chemicals and reagents included are Dulbecco’s modified Eagle’s medium (DMEM; Sigma-Aldrich), fetal bovine serum (FBS; Gibco Life Technology), and trypsin solution (Sigma Life Science). Cyclodipeptides mixture composed mainly by cyclo(L-Pro-L-Tyr), cyclo(L-Pro-L-Val), and cyclo(L-Pro-L-Phe) are isolated from the Pseudomonas aeruginosa PAO1 bacterium cells-free supernatant with a purity > 90% was used as therapeutic bioactive compounds [16,17]. The CDPs mixture was dissolved in a DMSO-water ratio 1:3 to prepare stock solutions (100 mg/mL). The MDA-MB-231 cell line was obtained from invasive ductal carcinoma (ATCC, Manassas, VA, USA) Breast tumor cell lines from pleural effusions. This cell line is estrogen receptor (ER), progesterone receptor (PR), and E-cadherin negative, p53 mutated, lacking the growth factor receptor HER2, commonly used as a study model of triple-negative breast cancer [18]. MDA-MB-231 cells, when implanted in xenografts TNBC mouse model into lymph nodes as described below. The MCF-7 is a human metastatic breast cancer cell line (adenocarcinoma) dependent on estrogen, progesterone, and glucocorticoid receptors (ATCC, Manassas, VA, USA) [19]. MDA-MB-231 and MCF-7 cell lines were cultured in complete media [DMEM supplemented with 10% (v/v) FBS, 100 units/mL of penicillin, 40 µg/mL of streptomycin, and 1 µg/mL of amphotericin B (Sigma-Aldrich Co.), supplemented with 1.6 g/L of glucose. Cell culture media were changed twice a week, incubating at 37 °C under 80% humidity and an atmosphere of 5% CO2 to confluency. Cells were then trypsin-treated, counted using a hemocytometer chamber, and used for subsequent assays. Cell cultures and other procedures were performed in class II biological safety cabinets.
2.2. Cell Viability and Apoptosis Determination in Cell Cultures
Cell viability was determined colorimetrically with the MTT method. Briefly, cell cultures grew in 96-well flat-bottomed plates with DMEM medium with FBS for 24 h, incubated in the presence or the absence of the indicated amounts of CDPs for 24 h at 37°C with 5% CO2. To determine cell viability, MTT (50 mg/mL) in PBS was added to each well and incubated for 4 h at 37°C. Finally, 100 uL of 2-propanol/1M HCl (19:1, v/v) was added to dissolve the formazan crystals, and the absorbance was measured at 595 nm using a microplate reader (BioTek Instruments).
Necrosis and apoptosis were evaluated in cell cultures incubated in DMEM medium with FBS for 4 h with CDPs treatment. DMSO was used as a control at the same concentration to dissolve the CDPs. Following incubation, cells were collected by centrifugation at 2,000 × g for 10 min. The pellet was suspended in 20 uL and incubated with annexin V and propidium iodide (PI) (Dead Cell Apoptosis Kit; Molecular Probes, Invitrogen Life Technologies). Fluorescence was immediately quantified by FACS using an Accuri-C6 Flow Cytometer (BD Biosciences). At least 20,000 cellular events were used for calculations.
2.3. Wound Closure Migration Assay
MDA-MB-231 and MCF-7 cell lines were grown in a 95% confluent monolayer, and three wounds were made per plate with a sterile pipette tip in assays by triplicate. Plates were washed with PBS twice and fresh complete medium; the CDPs, MTX, and combined treatments were placed for 24 and 48 h, and photographs were taken every 24 h to subsequently quantify the wound area using the ImageJ software (NIH).
2.4. Invasion Assay
MDA-MB-231 or MCF-7 cell lines were cultured in a transwell chamber previously coated with Matrigel (Corning, Life Sciences) with a complete DMEM medium. Co-culture was carried out using Raw 264.7 macrophages seeded in the lower chamber with complete DMEM medium and treatments with CDPs or MTX at 0.01 and 0.005 mg/mL concentrations, respectively. The transwell insert was placed into a 24-well plate with 0.2% (w/v) crystal violet containing PBS and incubated for 5 minutes; the transwell insert was washed with any remaining crystal violet from the membrane. The membrane was dried and photographed using a microscope; multiple images were taken with a 10x or 20x objective to capture a representative field of view [20].
2.5. Multicellular Microspheroids
MDA-MB-231 or MCF-7 cell lines were cultured (2 x105 cells) under non-stick conditions (0.6% agarose) in complete DMEM medium for 14 days with periodic medium changes until the spheroids reached a size of 40-50 µm [21]. Once treatment with CDPs (0.1 mg/mL), CDPs+cd (0.1 mg/mL), or CDPs-MTX containing (0.1 mg/mL CDPs, 0.05 mg/mL MTX) was applied. Photographs were taken at 72 h of treatment, and a cell viability test was performed on the remaining cells using an MTT reduction assay. MTT, 50 mg/mL in PBS, was added to each well and incubated for 4 h at 37°C. Finally, 100 µL of 2-propanol/1M HCl (19:1, v/v) was added to dissolve the formazan crystals, and the absorbance was measured at 595 nm using a microplate reader (BioTek Instruments).
2.6. Western Blot
Confluent MCF-7 and MDA-MB-231 cells treated with 0.1 mg/ml of CDPs incubated for 15 min, 1 h, and 4 h were harvested after trypsin treatment and washed twice with PBS, subsequently centrifuged at 5000 × g at 4 °C for 10 min and resuspended in RIPA lysis buffer. The cell suspension or tumor tissue was lysed by three cycles of sonication at low intensity (20 kHz, 5 W) for 30 s each at 4 °C (Hielscher-LS24 Ultrasound Technol). The protein extract free of cellular debris was obtained by centrifugation at 7500 × g for 15 min, and the protein concentration was determined by the Bradford method (BioRad). Protein extracts were separated on 10% (SDS-PAGE) polyacrylamide gels and transferred to PVDF membranes (Millipore, Billerica, MA, USA) in a BioRad transfer chamber at 15 volts for 45 min. Briefly, PVDF membranes were incubated with TBS-T (Tris-HCL 10 mM; NaCl 0.9%; tween-20 0.1%, dry milk 5%, pH 7.8). They were washed three times with TBS-T for 6 min each and incubated with primary antibodies dissolved in TBS-T at the concentration suggested by the manufacturer. Antibodies were used: anti-AKT, anti-p-AKT (Ser 473), anti-mTOR, anti-p-mTOR, anti-S6K, anti-p-S6K, anti-Vimentin; anti-Gab1, anti-FOX01, anti-p-FOX01, and anti-b-actin (Santa Cruz Biotechnology, Santa Cruz, CA, USA and Cell Signaling Technology, MA, USA). All primary antibodies (1:5,000 dilution) were incubated for 12 h at 4°C and subsequently three washes by 6 min with TBS-T were performed and then incubated with anti-mouse secondary HRP-conjugated antibody (BioRad, CA, USA) at a concentration of 1:10,000 in TBS-T for 2 h; membranes were washed as above and developed with Supersignal West Pico Luminol (Pierce; Thermo Fisher Scientific, Waltham, MA, USA). Then, images were captured using a ChemiDoc™ MP System (Bio-Rad). Assays were conducted at least three times, and representative images are shown. The Image J software (NIH) was used to quantify image band intensities.
2.7. Orthotopic Model of Metastatic Breast Cancer
Immunosuppressed Balb/c nu/nu female mice aged 8 weeks were adapted for 15 days in a pathogen-free environment with a standard diet and drinking water ad libitum. The cages, water, food, and bedding were autoclaved. Animal manipulation was authorized by the Institutional Animal and Use Committee (IACUC) from the Universidad Michoacana de San Nicolás de Hidalgo (IIIQB-UMSNH-IACUC-2022-35). Experiments complied with standard guidelines for the welfare of animals following the Institutional Committee and recommendations of the Mexican Official Regulations for the Use and Care of Animals (NOM 062-ZOO-1999; Ministry of Agriculture, Mexico). Animal handling, feeding, and care were done by trained personnel under the NIH guide for the care and use of laboratory animals.
Once the adaptation period had concluded with 100% survival of the mice, 1 x105 MDA-MB-231 metastatic breast cancer cells suspended in Matrigel were inoculated by direct injection into the nearest breast fat pad to the right armpit. Mice were sedated with ketamine (ket)/xylazine (xyl) solution intraperitoneally (IP) (80 mg/kg ket and 10-15 mg/kg xyl) and kept on a thermal mat during sedation to prevent hypothermia. The mice’s weight was monitored, and the longest diameter “a” and the shortest diameter “b” of the tumor were measured with a caliper; the tumor volume was determined using the tumor volume formula (TV) = (0.4) (ab2), “a” was the long diameter and “b” the short diameter of the visual tumor. The treatment consisted of 0.25 mg/kg of body weight of CDPs (0.1 mg/kg CDPs) via the IP or 0.05 mg/kg of MTX in physiological saline by IP via or in CDPs+MTX (0.1 mg/kg CDPs, 0.05 mg/kg MTX) combined treatment. The CDPs treatments were carried out by three rounds of administration, each consisting of three doses administered every 3 days (two days between each administration) with a week of rest between each administration round, summing nine doses per mouse. The animal groups each of five mice were evaluated as follows: healthy mice without tumor and treatment (C); healthy mice without tumor administered with the CDPs (C+CDPs); mice with TNBC tumor and without treatment (TNBC); mice with tumor and treatment with CDPs from cell inoculation in early stage [TNBC+CDPs(i)]; mice with tumor and treatment with CDPs at 35th-day post-inoculation in advanced stage [TNBC+CDPs (m)]; advanced-stage tumor-bearing and MTX-treated mice (TNBC+MTX(m)); mice with tumor and combined CDPs+MTX treatment (TNBC+CDPs+MTX(m)); each group with n=5.
2.8. Animal Euthanasia and Biological Samples
All mice were injected with a lethal dose of sodium pentobarbital intraperitoneally (100-150 mg/Kg body weight) as recommended by IIQB/UMSNH/IACUC and NOM 062-ZOO-1999. The tumors were removed, whole blood was obtained by cardiac puncture, and collected in a microtube with EDTA anticoagulant. The liver, lungs, spleen, kidneys, and femur of both legs were also removed and weighed. Blood hematocrit (Hct) was determined in a capillary tube with heparin by centrifugation at 3500 rpm for 5 min, and Hemoglobin (Hb) was determined by the Hct value divided by 3.3. The whole blood was centrifuged at 3500 rpm for 10 min to separate the blood serum. Subsequently, the enzymes aspartate aminotransferase (AST), alanine aminotransferase (ALT), and lactate dehydrogenase (LDH) were determined using a Fuji Dry-Chem NX 700 Fujifilm equipment.
2.9. Histological Analysis of Tissues and Bones
After euthanasia, the organs were dissected, and the tumors, lungs, and liver were utilized for histological analysis by observing tumor abscesses. The tumor, lungs, and liver were fixed in 4% paraformaldehyde and remained until dehydration. The samples were subjected to a dehydration process of 4 h at different concentrations of ethanol and xylene and embedded in paraffin. Tissue sections of 4 μm were cut and placed on slides for subsequent staining with H&E. Photographs were taken under the optic microscopy and recorded using an Accu-Scope EXC-120 LED microscope camera at magnifications of 4x, 10x, 40x, and 100x across tissue sections.
For bone histology, the lower limbs of the mice were dissected, removing skin and muscle tissue in the femurs. Femurs were fixed in paraformaldehyde and treated with 3 mL decalcification solution (10% EDTA, pH 7.4) for four weeks. Weight loss in femurs was determined by normalizing total animal weight over the weight of the femur during the decalcification procedure. For histology of bone slides, samples were dehydrated and underwent sequential ethanol treatments with increasing concentrations (80–100% ethanol), followed by xylene treatment and paraffin infiltration (Hycel) in two cycles before embedding in histological molds. The femurs were sectioned longitudinally to 5 μm-thick slices using a rotary HM 355S microtome (Thermo Fisher Scientific). To assess bone resorption, the presence of osteoclasts and osteoblasts in the bone sections were stained with hematoxylin and eosin (H&E).
2.10. Statistical Analysis
All data were evaluated using analysis of variance (ANOVA), and significant differences at (p< 0.05) were determined using Tuke’s, Bonferroni, or t-student posthoc tests using GraphPad Prism 6.0 software (GraphPad Software).
3. Results
3.1. Bacterial Cyclodipeptides Decrease Migration and Invasiveness in Breast Cancer Lines
Previously, the cytotoxic and apoptotic properties of the CDPs isolated from the P. aeruginosa PAO1 bacterium have been reported using human cancer cell lines such as HeLa and CaCo-2 [22] and malignant lines from animals [13]. However, the anti-metastasis capability of the bacterial CDPs is unknown. Thus, the MDA-MB-231 cell line was utilized as a model study of triple metastatic breast cancer to evaluate the anti-metastatic properties of CDPs.
The migration of the MDA-MB-231 cell line measured by injury area in monolayer plates showed a recovery of 90% of the cell monolayer in the control plate without treatment at 48 h incubation (25 µm2). Still, the CDPs treatment (0.01 mg/mL) caused significant inhibition of the MDA-MB-231 cell monolayer migration, showing ~180 µm2 of injury area after 48 h (Figure 1a). In addition, as positive control, methotrexate (MTX) at 0.005 mg/mL, showed a monolayer migration ~140 µm2 of injury area after 48 h. In contrast, the combination of CDPs+MTX treatment showed a monolayer migration of ~180 µm2 of injury area. Similar confirmative results were obtained in the MCF-7 cell line (Figure 1a).
On the other hand, chemokines favor malignancy and tumor progression, which is stimulated by seeding macrophages, promoting the invasive capacity of cancer cells [4]. The invasive phenotype of cancer cells was measured by the ability to degrade components of the extracellular matrix and filter through small porosities and by using co-culture with Raw cells to test the interaction between macrophages and tumor cells. In this test, the number of cells stained with crystal violet corresponds to the cells that were able to degrade the Matrigel and penetrate the pores of the membrane of the transwell chamber, which remained adhered to the lower face of the porous membrane. We observed first that the interaction of cancer cells with RAW macrophages co-cultured stimulated the number of invasive cells in both MDA-MB-231 and MCF-7 cell lines (Figure 1b). Secondly, the CDPs treatment (0.01 mg/mL) decreased the number of invasive cells significantly around 75% and 60% at single or co-cultured with RAW cells, respectively. In contrast to the invasive capacity of cancer cells treated with MTX (0.005 mg/mL), it was less efficient than the CDPs treatment (Figure 1b).
Additionally, microspheroids are cellular aggregates representing a model of a three-dimensional culture of cancer cells, which are used as a study model of tumorigenesis. Thus, the CDPs effect in the microspheroids structuring of the MDA-MB-231 and MCF-7 cell lines was evaluated. The results showed a significant decrease in the number and size of spheroids in both MDA-MB-231 and MCF-7 lines when they were treated with CDPs alone, combining CDPs with cyclodextrins, and in a combination of CDPs with the anti-neoplastic MTX (Figure 1c). Interestingly, the CDPs was more effective in inhibiting the cell spheroids conformation than MTX. When the viability was determined at the cell microspheroids, the CDPs treatment suspended into cyclodextrins was the better condition to cause death in both the MDA-MB-231 and MCF-7 cancer cell lines (Figure 1d-e). Additionally, the induction of apoptosis was confirmed by flow cytometry, finding that in the TNBC MDA-MB-231 line, the effective half doses (ED50) of apoptosis induction was around 0.02 mg/mL for 4 h of CDPs treatment and lethal doses (LD50) around 0.25 mg/mL for CDPs (Figure 1f).
3.2. Effect of the Bacterial CDPs in Cell Proliferation and Invasiveness Pathways
The signaling pathways involved in the anti-neoplastic effect of the bacterial CDPs in the MDA-MB-231 triple-negative breast cancer cells were studied. Results showed that the CDPs treatment did not modify the expression of the total Akt and mTOR proteins and did not function over time. Interestingly, the phosphorylation of both the p-Akt and p-mTOR proteins was significantly decreased by the CDPs treatment in a time-dependent manner (Figure 2). In addition, the p-S6K, Vimetin, and Gab1 proteins were also significantly down-regulated in their expression level in the MDA-MB-231 cell line by the CDPs treatment (Figure 2).
3.3. Effect of the Bacterial CDPs in the Metastatic MDA-MB-231 Cell Line in Breast Cancer Mice Model
The metastatic study model consisted of the implantation of the MDA-MB-231cell line (triple negative human breast cancer) in immunosuppressed mice. The metastatic MDA-MB-231 cell line was inoculated into the breasts of immunosuppressed female mice, obtaining 100% performance in generating mammary tumors.
Two treatment conditions were tested; first, the MDA-MB-231cell line inoculation joined with the CDPs treatment at the time of cell implantation, following the treatment of nine doses of CDPs (0.25 mg/kg) under 35 days of therapeutic experimental outline (Figure 3a). The second procedure consisted in the development of breast tumors by MDA-MB-231 line inoculation and development during 35 days; after that, the CDPs treatment was begun and followed by nine doses of CDPs (0.25 mg/kg) during additional 35 days of therapeutic experimental outline (Figure 3a). During the development of the study, the weight was monitored in the animals submitted to CDPs treatment. Results show that the slopes of weight gain were similar in all the mice groups, except for the mice groups with TNBC breast cancer implantation treated with CDPs (BC+CDPs) and the mice groups with TNBC implantation treated with MTX plus CDPs (TNBC+CDPs+MTX) (Figure 3b).
The tumor volume was monitored in the mice groups during the therapeutic experimental outline. Figures 4a-b show the images of tumors in the animal groups implanted with TNBC and submitted to CDPs treatment (70th day); the TNBC mice group showed means of tumor volume of ~300 mm3 (Figure 4a,b). Interestingly, the TNBC mice groups treated with CDPs since the implantation of the MDA-MB-231 cells or after the metastatic development phase (on the 35th day of the MDA-MB-231 cells implantation) showed a significant reduction of the means of tumor volume ~10 mm3 and ~3 mm3, respectively; showing major efficacy than the MTX treatment presenting ~90 mm3 tumors means (Figure 4ab).
Once the animals were euthanized, the organs and tumors were dissected and studied in detail. Tumors in the TNBC group showed weight means of ~0.9 g. In comparison, the TNBC animals treated with CDPs in both procedures showed means of ~0.15 g and ~0.05 g of weight, respectively (Figure 4c). Additionally, in the group treated with MTX, the means weight in tumors of ~0.15 g, while that in the TNBC+CDPs (m) and TNBC+CDPs+MTX(m) were significantly lesser tumor weight that 0.05 g, but notably in the 40 and 60% of animals, the tumors presence was missing, respectively (Figure 4c).
3.4. Anti-Metastatic Effect of Bacterial CDPs in the Lungs and Liver of the TNBC Model Implanted with the MDA-MB-231 Cells Line
Studies have described that the most common metastasis sites in patients with breast cancer are lung, liver, brain, and bones. In our metastatic model, the group of mice with mammary tumors without treatment (TNBC), a significant increase in the weight of the lung, kidneys, liver, and spleen was observed (Figure 5a-b), which was related to the observation of metastatic foci in these organs. Interestingly, the groups treated in combination with CDPs plus MTX showed a reversion of the weight of lungs, kidneys, liver, and spleen, also observed by reversion of the metastatic foci in organs (Figure 5a-b).
The enzymes ALT, AST, and LDH as markers of hepatocellular tissue injury were determined in the blood serum of animals implanted with breast tumors. The TNBC mouse group showed a significant increase in AST and LDH enzyme activities (Figure 5c), which were reversed in the TNBC mice treated with CDPs, MTX, and CDPs+MTX combination. The treatment with CDPs in the healthy CDPs-treated control mouse group did not alter AST and LDH levels as shown in the control mouse group. The hemoglobin parameter in the blood serum showed impairment in the groups of mice with mammary tumor TNBC and TNBC+MTX(m); however, in the TNBC+CDPs mice treated, a restoration in hemoglobin was observed, almost at the level of the control of healthy mice (Figure 5d). The cells count of leucocytes showed differences in the neutrophils, lymphocytes, and monocytes in the blood of the mice group TNBC and TNBC+MTX(m); interestingly, the groups treated with CDPs showed similar leucocytes count to the group of healthy mice. Additionally, healthy mice treated with the CDPs (C+CDPs) did not show an alteration in these hematological parameters concerning the control group (Figure 5d).
Histological analyses of tumors, lungs, and liver sections stained with H&E revealed an increased number of cells of the MDA-MB-231 line and cells of inflammatory infiltrates in the TNBC mice group (Figure 6). Notably, the tumor tissue of the TNBC animals showed the presence of the MDA-MB-231 cells, large areas of fibrosis, and increased proportion of fat drops. In this sense, the CDPs and MTX treatments revert the fibrosis areas in the tumoral tissue. Still, interestingly, the MTX did not revert the content of fat drops in the tissue, which was alleviated when the CDPs were administered alone. While in the lungs and liver, treatment with CDPs, MTX, or the combination of CDPs+MTX exhibited a significantly decreased cell infiltration and foci of tumoral implantation of the MDA-MB-231 line in these tissues. As expected, there was no apparent lung and liver damage in the control groups in the CDPs or MTX administration (Figure 6).
3.5. Targets of CDPs on Proliferation and Invasiveness Signaling Pathways in Tumors Of TNBC Mice Model Implanted with the MDA-MB-231 Cell Line
Protein immunodetection analysis was conducted to study the signaling pathways involved in the anti-neoplastic and anti-metastatic effects of the bacterial CDPs in the MDA-MB-231 breast cancer cells implanted in the immunosuppressed mice model. Results showed that the expression of total Akt protein was not significantly modified by the treatments, except in the TNBC+CDPs(m) group (Figure 7). However, the CDPs and MTX treatments inhibited the expression of the total Akt phosphorylation. The Gab1 protein decreased significantly in the TNBC+MTX(m) and TNBC+CDPs+MTX(m) treatments, which was similar to the results for the p-Gab1 protein. Inhibition of the protein expression was also observed in the FOX01 protein, just like its phosphorylated protein p-FOX01 was decreased in the tumors of mice groups treated with CDPs, MTX, and in CDPs+MTX combination (Figure 7).
3.6. Anti-Metastatic Effect of Bacterial CDPs in the Bone of TNBC Mice Model Implanted with the MDA-MB-231 Cell Line
The anti-metastatic and invasiveness of the TNBC MDA-MB-231 line were determined in the exploration of bone health (Figure 8). Determination of the weight of femurs from the animals implanted with TNBC showed that the femur weight of the TNBC and TNBC+MTX(m) mice groups had the lowest weight compared to the CDPs and CDPs+MTX treated groups (Figure 8a).
Bone marrow, tumor areas, osteoclasts, and osteoblasts were identified using H&E staining. We evaluated the potential of CDPs alone and with MTX to assess bone metastasis in a xenotransplant mouse model.
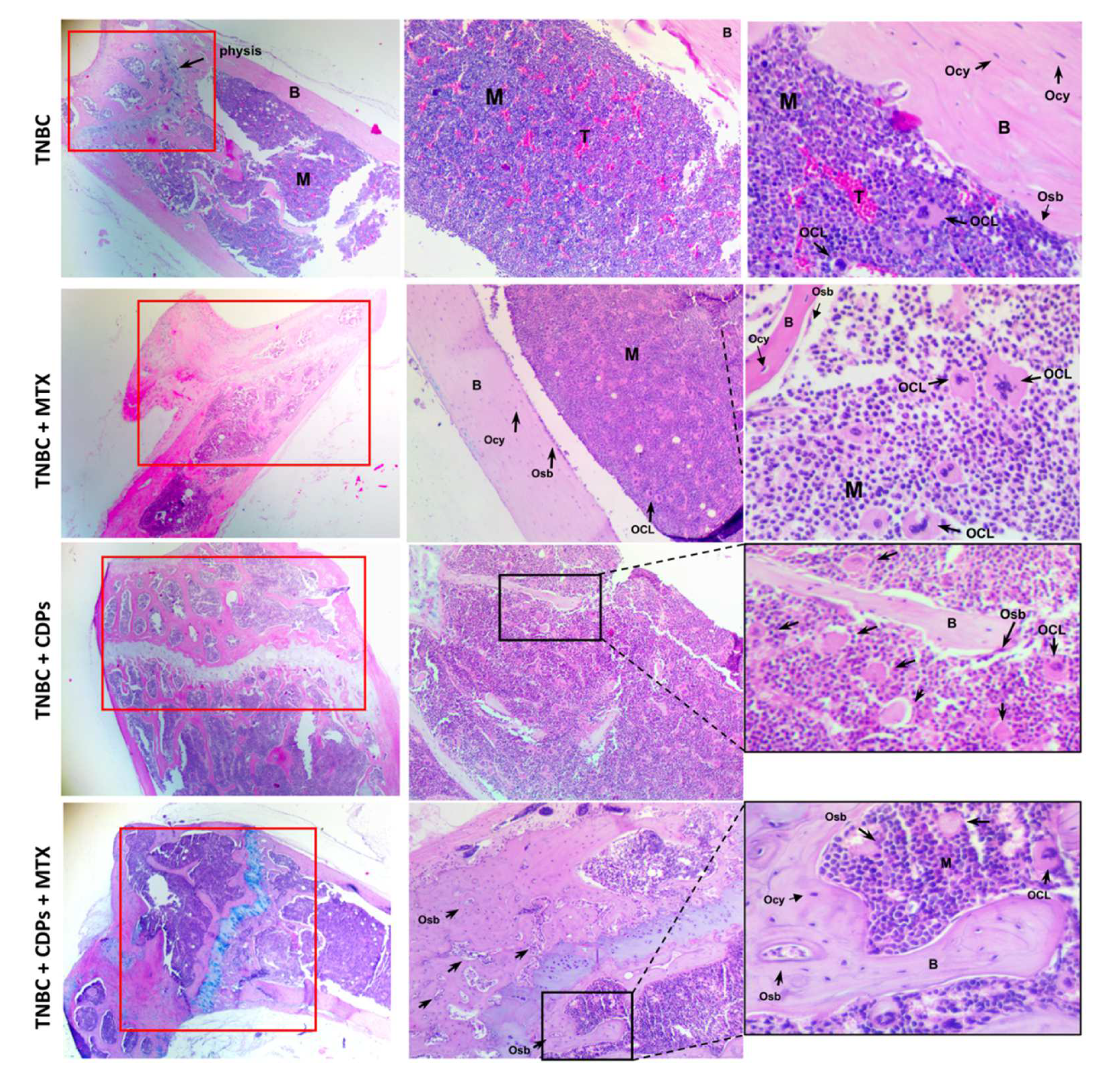
Histological analyses of femur bone sections stained with H&E revealed an increased number of cells of the MDA-MB-231 line and cells of inflammatory infiltrates in the TNBC mice group (Figure 9). Notably, the TNBC animals treated with CDPs did not exhibit cell infiltration and foci of tumoral implantation in the femur. As expected, in the TNBC group, the number of osteoclasts was significantly increased, which was reverted in the CDPs and MTX treatments; contrary, the osteoblast cells were significantly increased in the femur from animals treated with CDPs and MTX (Figure 8b; Figure 9). Quantification of osteoclasts and osteoblasts in femur sections revealed that CDPs-treated groups reduced osteoclast numbers while showing an increase in osteoblast populations (Figure 8b). The treatment with MTX was no sooner efficient than the CDPs or a combination between CDPs+MTX.
Tumor cell populations indicate regions of tumor in bone (T) (outlined with red dotted lines in Figure 9, appearing as pink areas, while the purple regions represent bone marrow (M). The results showed that in the untreated breast cancer group (TNBC), tumor infiltration in bone (T) regions was more frequent in comparison to groups treated with CDPs, either from disease onset TNBC+CDPs (i) or at advanced stages TNBC+CDPs(m), and TNBC+CDPs+MTX(m). Additionally, cortical bone areas were more extensive in the groups treated with the CDPs and MTX than in the TNBC group, suggesting a potential restoration of bone growth and integrity (Figure 9).
4. Discussion
CDPs (from the diketopiperazines family) have emerged as promising candidates for anticancer therapy due to their structural stability, high selectivity, and ability to efficiently inhibit key cancer-related pathways [23]. Previously, the cytotoxic and apoptotic effects of the bacterial CDPs have been demonstrated in various cancer cell lines [10,11,12,15,22]; in addition, their impact on breast cancer lines MCF-7 and MDA-MB-231 cells was described, triggering apoptosis in both cell lines in a concentration-dependent manner [12].
Several specific CDPs have exhibited potent anticancer activity; cyclo(Phe-Pro) inhibits the growth of HT-29, MCF-7 and HeLa cells [24], cyclo(Tyr-Cys) inhibits cervical carcinoma cells HT-29 and MCF7 [25], cyclo(L-Leu-D-Arg) shows cytotoxicity against MDA-MB-231 cells [26]. Diketopiperazines such as fumitremorgin C analogs have shown potential in overcoming multidrug resistance, including inhibiting the breast cancer resistance protein (BCRP), a key efflux transporter responsible for reducing intracellular drug accumulation. Moreover, the novel diketopiperazine-based compound HLY838 functions as an O-GlcNAc transferase (OGT) inhibitor, potentiating the anti-tumor effects of CDK9 inhibitors by downregulation c-Myc and E2F1 expression [27]. Similarly, Verticillin A, a diketopiperazine-derived compound, suppresses c-Met phosphorylation and its downstream Ras/Raf/MEK/ERK signaling pathway, significantly reducing colon cancer cell metastasis [28]. Collectively, these findings highlight the cyclopeptides such as CDPs as anticancer agents. Bacterial CDPs have demonstrated significant potential as anti-metastasic agents by targeting key proteins involved in epithelial-mesenchymal transition (EMT) and metastasis. In the murine melanoma model, CDPs derived from the P. aeruginosa bacterium significantly reduce the expression of critical EMT markers, including MMP-1, E-cadherin, N-cadherin, HIF-1a, Vimetin, and CK-1; indicating that CDPs may impact cancer cell migration and invasion [13].
In this work, functional assays further confirmed the anti-metastatic effects of P. aeruginosa CDPs were carried out.
Wound healing assays in monolayers of cell lines cultures of the triple-negative MDA-MB-231and luminal MCF7 breast cancer lines revealed that CDPs-treated cancer cells exhibited a wound area twice as large as the control and 1.5 times that of cells treated with methotrexate (MTX). MDA-MB-231 cells displayed higher migratory capacity than the MCF7 cells in all conditions (Figure 1a).
Transwell chamber assays using Matrigel and co-culture with macrophages revealed that tumor-associated macrophages (TMAs) enhanced cancer cell invasion by promoting Matrigel degradation and membrane penetration. However, treatment of the TNBC line with CDPs significantly reduced the number of invasive cells in both monoculture and co-culture conditions, suggesting their potential as metastatic agents. This inhibitory effect was comparable to MTX, which has been evaluated in combination therapies for metastatic disease, including bone metastases (Figure 1b). These findings align with previous reports indicating that TAMs play a crucial role in tumor progression by secreting chemokines and growth factors that facilitate cancer cells proliferation, migration and survival [29]. Moreover, TAMs have been implicated in modulating drug resistance, further supporting their relevance as therapeutic targets. The ability of the bacterial CDPs to disrupt TAMs cancer cell interactions highlights their potential dual role in inducing apoptosis and inhibiting metastasis.
The multicellular microspheroid model contains subpopulations of cancer stem cells (CSCs) [30] exhibiting self-renewal capacity and therapy resistance. Our microspheroids assay with MDA-MB-231 and MCF-7 cells demonstrated distinct phenotypic differences between the two lines. MCF-7 spheroids were more compact and enormous, likely due to their epithelial phenotype and high E-cadherin production [31]. At the same time, the MDA-MB-231 microspheroids were more susceptible to disintegration and more heterogeneous, reflecting their mesenchymal phenotype. Notably, the CDPs treatment significantly reduced spheroids size, number, and viability in both breast cell lines compared to untreated controls and MTX-treated, also inducing apoptosis at concentrations around 0.02 mg/mL for 4 h of CDPs treatment (Figure 1c-f). These results underscore the potential of bacterial CDPs as effective anti-metastatic agents by targeting both tumor cell invasion and CSC-associated resistance.
Previous studies from our group have demonstrated in cancer cell lines the involvement of the PI3K/AKT/mTOR pathway in the cytotoxic and proliferative effects of the bacterial CDPs [10,11,13]. Based on these findings, we analyzed key components of this pathway in the triple-negative MDA-MB-231 cell line of breast cancer, observing a significant decrease in the phosphorylation levels of AKT, mTOR, and S6K (Figure 2). This reduction in protein expression is associated with decreased cell viability, impaired tumor growth, and induction of apoptosis, corroborating previous observations [12]. Additionally, our results revealed a notable decrease in the expression of Vimentin and Gab1 following treatment with CDPs, further supporting the role of these molecules in targeting metastatic and invasive pathways (Figure 2). Gab1 overexpression has been linked to enhanced breast cancer metastasis [32]. At the same time, Vimetin is a well-established marker of EMT and is consistently upregulated during cancer progression and metastasis [33]. Therefore, the observed downregulation of Gab1 and Vimetin suggests that the CDPs reduce the invasive and migratory capacities in the MDA-MB-231 line, which may translate into decreased tumor aggressiveness.
Weight loss is often one of the earliest detectable symptoms of cancer disease and its progression, leading to severe muscle wasting, which significantly contributes to morbidity and mortality. This condition is primarily driven by the increased production of cytokines and other pro-inflammatory molecules secreted by the immune system to inhibit tumor progression [34,35]. In our orthotopic model of metastatic breast cancer, body weight was monitored throughout the treatment period. While the overall pattern of weight gain remained consistent across groups, a transient weight loss was observed in animals treated with CDPs during the initial weeks of administration (Figure 3b). Nevertheless, this effect did not persist; no behavioral changes or signs of deteriorating health were observed, suggesting that the temporary weight loss did not indicate toxicity or systemic distress. Although the volume and weight of the tumors generated in the TNBC group were significantly observed (Figure 4), they did not significantly modify the weight of the animals (Figure 3b). Notably, the administration of CDPs in mice prevented the generation of large and heavy tumors, observing that 40-60% of animals in the TNBC+CDPs(m) and TNBC+CDPs+MTX(m) did not show tumors (Figure 4). However, when the tumoral tissue of the mice groups was dissected and histologically analyzed, a significant presence of fibrosis and fat drops was observed in the tumors of the TNBC mice group; which was reverted in the CDPs-treated animals (Figure 6); interestingly the MTX administration, although positively contributed to diminishing fibrosis areas, this drug provokes an increased accumulation of fat drops. Developing bone marrow adipose tissue (BMAT) and fat drops in other organs is a normal physiological process; however, excessive fat accumulation may have a significant pathological implication, such as cancer [36].
In breast cancer patients, alterations in leukocyte composition, particularly in the neutrophil-to-lymphocyte ratio (NRL), have been documented. NLR is widely recognized as a prognostic biomarker in cancer, with elevated levels correlating with worse prognosis, increased tumor aggressiveness, and reduced overall survival [37]. The findings suggest that changes in leukocyte distribution may have significant clinical implications in breast cancer progression. In our study, alterations in leukocyte composition were also observed in mice with TNBC. NLR analysis revealed that the TNBC group exhibited the highest value (NLR=8).
In contrast, the groups TNBC+CDPs(i) (NLR=3) and TNBC+ CDPs+MTX(m) (NLR=3.5) showed significantly lower values, suggesting a potential reduction in tumor aggressiveness following CDPs treatment. Anemia is also common among cancer patients [38]. In our hematological analysis, hemoglobin levels showed improvement in animals treated with CDPs (Figure 5d), indicating a potential benefit of these molecules in mitigating cancer-associated hematological alterations.
Liver metastasis is common in patients with breast cancer. Previous studies have described that liver function is poor in 92% of patients diagnosed with breast cancer liver metastasis (BCLM) [39]. Moreover, ALT, AST, GGT, AP, and LDH levels are significantly higher in patients with BCLM. The determination of AST, ALT, and LDH showed a drastic increase in the group with TNBC without treatment (Figure 5c). The presence of metastatic foci was observed in the TNBC group of animals (Figure 5a); consistent with these observations, an increase in the weight of the lungs and liver organ was also observed (Figure 5b). Histological analysis of lung and liver in mice revealed that the CDPs treatment inhibited malign cells and tumor focus (Figure 6). These findings suggest that the animals treated with CDPs presented levels of AST, ALT, and LDH and organs similar to the control without disease, indicating a positive anti-tumor capability associated with the CDPs treatment.
On the other hand, critical tumorigenic markers were repressed in the TNBC CDPs-treated mice. A significant reduction in p-AKT levels was observed in tumors from rodents subjected to treatment (Figure 7). Notably, TNBC animals treated with CDPs also exhibited a decrease in total AKT protein levels, suggesting a potential disruption of the PI3K/AKT/mTOR signaling pathway in tumors associated with the CDPs administration.
AKT is pivotal in regulating FOX01 phosphorylation, a key factor in cell cycle regulation and apoptosis. Under normal conditions, AKT phosphorylates FOX01, excluding it from the nucleus and suppressing its transcriptional activity. However, when FOX01 remains dephosphorylated, it is translocated to the nucleus, activating apoptosis-related genes and cell cycle arrest [40].
FOX01, a member of the Forkhead box family of transcription factors, is known to regulate cell cycle arrest, autophagy, and apoptosis. Its tumor-suppressive role has been reported in several malignancies, including breast cancer, where its dysregulation is frequently observed [41]. In our study, tumors from TNBC animals CDPs-treated exhibited reduced phosphorylation of FOX01, supporting that CDPs inhibit tumor progression (Figure 7). These findings align with our previous transcriptomic analysis in HeLa cells treated with CDPs, where genes involved in the FOX01 signaling pathway, including GADD45A and SGK1 were upregulated [14]. Data presented here suggest a differential regulation of FOX01 by CDPs in the triple-negative breast cancer model.
Contrary to observations in the MDA-MB-231 cell line (Figure 2), Gab1 levels remained unchanged in response to CDPs treatment. However, a notable decrease in both total Gab1 and phosphorylated Gab1 (p-Gab1) levels was detected in tumors from the TNBC+CDPs+MTX(m) and TNBC+MTX(m) groups (Figure 7). Given that Gab1 acts as a key integrator of multiple signaling pathways involved in cell survival, proliferation, angiogenesis, and invasion [42]. The Gab1 downregulation suggests a potential mechanism through which CDPs, particularly in combination with MTX, inhibit tumor progression and metastasis.
Femora weight determination showed less weight in the TNBC and TNBC+MTX(m) mice groups than in the CDPs-treated (Figure 8a). Histological analysis of the femur in mice revealed that in the TNBC group zones of invasion of tumoral cells in the bone marrow (B), which were not observed in the femurs of mice submitted to treatment with CDPs or MTX (Figure 9). Additionally, the treatment with CDPs and the combination of CDPs+MTX led to decrease of osteoclasts and increase in the number of osteoblasts (Figs. 8b and 9). A similar proportion of osteoblasts and osteoclasts was observed in the group treated with MTX alone. These cells play a crucial role in osteogenesis by producing and depositing the organic bone matrix [43]. Osteoclasts are specialized cells that participate in bone modeling, calcium homeostasis, and hematopoiesis modulation bone healing; they also are essential cells related to the tumor microenvironment in bone metastases in BC [44], which inhibition can contribute to patients’ recovery and increase life span and quality of life. Another characteristic observed was the increase of the apophysis (growth bone zone) in the head of the femur in all the mice submitted to CDPs/MTX treatments. These findings suggest a potential pro-osteogenic effect of CDPs.
5. Conclusions
Data provide new insights into the molecular mechanisms involved in CDPs anti-metastatic and invasiveness effects, suggesting that the signal transduction mechanism is related to the inhibition of the phosphorylation of the Akt/mTOR/S6K pathway and the metastasis markers Vimentin, Gab1, and FOX01. Since Gab1 inhibitors have been proposed as promising anticancer agents, the observed reduction in Gab1 expression further supports the therapeutic potential. Under the mice model of TNBC, the CDPs administration reverted the development of tumors in the mammary glandules, being more effective in a combination of CDPs+MTX as a treatment. The metastasis foci in organs such as the lungs and liver, as well as in bones, were reversed by the CDPs treatment. The proportion of osteoclasts in the femoral bone marrow of TNBC animals was decreased by the CDPs administration, correlating with bone remodeling triggered by metastatic cells of the MDA-MB-231 line. The findings indicate that the CDPs, combined with MTX, potentialize the anti-neoplastic and anti-tumor effects in the TNBC model of xeno-implantation of the MDA-MB-231 line, proposing the bacterial CDPs as a viable alternative in human triple-negative breast cancer therapy.
Author Contributions
Conceptualization, J. Campos-García; Data curation, L. Martínez-Alcantar; Formal analysis, L. Martínez-Alcantar; Funding acquisition, J. Campos-García; Investigation, M. Durán-Maldonado, N. Ramírez-Gallardo, and L. Martínez-Alcantar; Methodology, M. Durán-Maldonado, N. Ramírez-Gallardo, and M. Campos-Morales; Supervision, L. Hernández-Padilla and J. Campos-García; Writing—original draft, L. Martínez-Alcantar and L. Hernández-Padilla; Writing—review & editing, L. Martínez-Alcantar and J. Campos-García.
Institutional Review Board Statement
All procedures performed in animal studies followed the ethical standards of the institution or practice at which the studies were conducted. The Institutional Animal and Use Committee (IACUC) approved the study from the Universidad Michoacana de San Nicolás de Hidalgo (protocol: IIIQB-UMSNH-IACUC-2022-33). The experiments followed the Institutional Committee and recommendations of the Mexican Official Regulations for the Use and Care of Animals (NOM 062-ZOO-1999; Ministry of Agriculture, Mexico).
Informed Consent Statement
All the authors approved the participation.
Data Availability Statement
The data supporting the findings of this study are available from the corresponding author upon request.
Acknowledgments
Thanks to the Secretaría de Ciencia, Humanidades, Tecnología e Innovación (SECIHTI, before CONAHCYT) of Mexico and the Universidad Michoacana de San Nicolás de Hidalgo by scholarship grants to M.X.D.M., N.A.R.G., and L.H.P.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the study’s design, in the collection, analyses, or interpretation of data, in the writing of the manuscript, or in the decision to publish the results.
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA: A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Huang, W.; Xiao, Z.; Wang, J.; Zhu, Y.; Zhang, F.; Lan, D.; He, F. Immunotherapy in the Fight Against Bone Metastases: A Review of Recent Developments and Challenges. Curr. Treat. Options Oncol. 2024, 25, 1–16. [Google Scholar] [CrossRef]
- Derakhshan, F. and J.S. Reis-Filho, Pathogenesis of Triple-Negative Breast Cancer. Annu Rev Pathol, 2022. 17: p. 181-204.
- Liang, Y.; Zhang, H.; Song, X.; Yang, Q. Metastatic heterogeneity of breast cancer: Molecular mechanism and potential therapeutic targets. Semin. Cancer Biol. 2020, 60, 14–27. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Wang, B.; Zhao, Y.; Tao, Z.; Wang, Y.; Chen, G.; Hu, X. Mammary adipocytes protect triple-negative breast cancer cells from ferroptosis. J. Hematol. Oncol. 2022, 15, 1–5. [Google Scholar] [CrossRef]
- Pang, L.; Gan, C.; Xu, J.; Jia, Y.; Chai, J.; Huang, R.; Li, A.; Ge, H.; Yu, S.; Cheng, H. Bone Metastasis of Breast Cancer: Molecular Mechanisms and Therapeutic Strategies. Cancers 2022, 14, 5727. [Google Scholar] [CrossRef]
- Kakhki, V.R.D.; Anvari, K.; Sadeghi, R.; Mahmoudian, A.-S.; Torabian-Kakhki, M. Pattern and distribution of bone metastases in common malignant tumors. Nucl. Med. Rev. 2013, 16, 66–69. [Google Scholar] [CrossRef]
- Ryan, C. , et al., Epidemiology of bone metastases. Bone, 2022. 158: p. 115783.
- Clézardin, P.; Coleman, R.; Puppo, M.; Ottewell, P.; Bonnelye, E.; Paycha, F.; Confavreux, C.B.; Holen, I. Bone metastasis: mechanisms, therapies, and biomarkers. Physiol. Rev. 2021, 101, 797–855. [Google Scholar] [CrossRef]
- Hernández-Padilla, L.; Vázquez-Rivera, D.; Sánchez-Briones, L.A.; Díaz-Pérez, A.L.; Moreno-Rodríguez, J.; Moreno-Eutimio, M.A.; Meza-Carmen, V.; la Cruz, H.R.-D.; Campos-García, J. The Antiproliferative Effect of Cyclodipeptides from Pseudomonas aeruginosa PAO1 on HeLa Cells Involves Inhibition of Phosphorylation of Akt and S6k Kinases. Molecules 2017, 22, 1024. [Google Scholar] [CrossRef]
- Hernández-Padilla, L.; de la Cruz, H.R.; Campos-García, J. Antiproliferative effect of bacterial cyclodipeptides in the HeLa line of human cervical cancer reveals multiple protein kinase targeting, including mTORC1/C2 complex inhibition in a TSC1/2-dependent manner. Apoptosis 2020, 25, 632–647. [Google Scholar] [CrossRef]
- Hernandez-Padilla, L.; Duran-Maldonado, M.X.; Martinez-Alcantar, L.; Rodriguez-Zavala, J.S.; Campos-Garcia, J. The HGF/Met Receptor Mediates Cytotoxic Effect of Bacterial Cyclodipeptides in Human Cervical Cancer Cells. Curr. Cancer Drug Targets 2025, 25, 230–243. [Google Scholar] [CrossRef]
- Durán-Maldonado, M.X.; Hernández-Padilla, L.; Gallardo-Pérez, J.C.; Díaz-Pérez, A.L.; Martínez-Alcantar, L.; De la Cruz, H.R.; Rodríguez-Zavala, J.S.; Pacheco-Rodríguez, G.; Moss, J.; Campos-García, J. Bacterial Cyclodipeptides Target Signal Pathways Involved in Malignant Melanoma. Front. Oncol. 2020, 10, 1111. [Google Scholar] [CrossRef] [PubMed]
- Lázaro-Mixteco, P.E.; González-Coronel, J.M.; Hernández-Padilla, L.; Martínez-Alcantar, L.; Martínez-Carranza, E.; López-Bucio, J.S.; Guevara-García, Á.A.; Campos-García, J. Transcriptomics Reveals the Mevalonate and Cholesterol Pathways Blocking as Part of the Bacterial Cyclodipeptides Cytotoxic Effects in HeLa Cells of Human Cervix Adenocarcinoma. Front. Oncol. 2022, 12, 790537. [Google Scholar] [CrossRef]
- Martínez-Alcantar, L.; Hernández-Padilla, L.; Díaz-Pérez, A.L.; Villalón-Magallán, L.G.; Durán-Maldonado, M.X.; Díaz-Pérez, C.; Campos-Morales, M.E.; Figueroa-Guzmán, C.; Campos-García, J. Cyclic Peptides as Protein Kinase Modulators and Their Involvement in the Treatment of Diverse Human Diseases. Kinases Phosphatases 2024, 2, 346–378. [Google Scholar] [CrossRef]
- González, O.; Ortíz-Castro, R.; Díaz-Pérez, C.; Díaz-Pérez, A.L.; Magaña-Dueñas, V.; López-Bucio, J.; Campos-García, J. Non-ribosomal Peptide Synthases from Pseudomonas aeruginosa Play a Role in Cyclodipeptide Biosynthesis, Quorum-Sensing Regulation, and Root Development in a Plant Host. Microb. Ecol. 2016, 73, 616–629. [Google Scholar] [CrossRef]
- Ortiz-Castro, R.; Díaz-Pérez, C.; Martínez-Trujillo, M.; del Río, R.E.; Campos-García, J.; López-Bucio, J. Transkingdom signaling based on bacterial cyclodipeptides with auxin activity in plants. Proc. Natl. Acad. Sci. 2011, 108, 7253–7258. [Google Scholar] [CrossRef]
- Amaro, A.; Angelini, G.; Mirisola, V.; Esposito, A.I.; Reverberi, D.; Matis, S.; Maffei, M.; Giaretti, W.; Viale, M.; Gangemi, R.; et al. A highly invasive subpopulation of MDA-MB-231 breast cancer cells shows accelerated growth, differential chemoresistance, features of apocrine tumors and reduced tumorigenicityin vivo. Oncotarget 2016, 7, 68803–68820. [Google Scholar] [CrossRef]
- Tian, T. MCF-7 cells lack the expression of Caspase-3. Int. J. Biol. Macromol. 2023, 231, 123310. [Google Scholar] [CrossRef]
- Justus, C.R.; Marie, M.A.; Sanderlin, E.J.; Yang, L.V. Transwell In Vitro Cell Migration and Invasion Assays. Methods Mol Biol, 2023. 2644: p. 349-359.
- Gallardo-Pérez, J.C.; de Guevara, A.A.-L.; Marín-Hernández, A.; Moreno-Sánchez, R.; Rodríguez-Enríquez, S. HPI/AMF inhibition halts the development of the aggressive phenotype of breast cancer stem cells. Biochim. et Biophys. Acta (BBA) - Mol. Cell Res. 2017, 1864, 1679–1690. [Google Scholar] [CrossRef]
- Vázquez-Rivera, D.; González, O.; Guzmán-Rodríguez, J.; Díaz-Pérez, A.L.; Ochoa-Zarzosa, A.; López-Bucio, J.; Meza-Carmen, V.; Campos-García, J. Cytotoxicity of Cyclodipeptides fromPseudomonas aeruginosaPAO1 Leads to Apoptosis in Human Cancer Cell Lines. BioMed Res. Int. 2015, 2015, 1–9. [Google Scholar] [CrossRef]
- Hasan, M.; Shawon, A.R.M.; Aeyas, A.; Uddin, M.A. Cyclic peptides as an inhibitor of metastasis in breast cancer targeting MMP-1: Computational approach. Informatics Med. Unlocked 2022, 35. [Google Scholar] [CrossRef]
- Brauns, S.C.; Milne, P.; Naudé, R.; Van De Venter, M. Selected cyclic dipeptides inhibit cancer cell growth and induce apoptosis in HT-29 colon cancer cells. . 2004, 24, 1713–9. [Google Scholar] [PubMed]
- van der Merwe, E.; Huang, D.; Peterson, D.; Kilian, G.; Milne, P.; Van de Venter, M.; Frost, C. The synthesis and anticancer activity of selected diketopiperazines. Peptides 2008, 29, 1305–1311. [Google Scholar] [CrossRef] [PubMed]
- Nishanth, S.K.; Nambisan, B.; Dileep, C. Three bioactive cyclic dipeptides from the Bacillus sp. N strain associated with entomopathogenic nematode. Peptides 2014, 53, 59–69. [Google Scholar] [CrossRef]
- Wauters, I.; Goossens, H.; Delbeke, E.; Muylaert, K.; Roman, B.I.; Van Hecke, K.; Van Speybroeck, V.; Stevens, C.V. Beyond the Diketopiperazine Family with Alternatively Bridged Brevianamide F Analogues. J. Org. Chem. 2015, 80, 8046–8054. [Google Scholar] [CrossRef]
- Liu, Q.-Q.; Zeng, X.-L.; Guan, Y.-L.; Lu, J.-X.; Tu, K.; Liu, F.-Y. Verticillin A inhibits colon cancer cell migration and invasion by targeting c-Met. J. Zhejiang Univ. B 2020, 21, 779–795. [Google Scholar] [CrossRef]
- Li, C.; Xu, X.; Wei, S.; Jiang, P.; Xue, L.; Wang, J. Tumor-associated macrophages: potential therapeutic strategies and future prospects in cancer. J. Immunother. Cancer 2021, 9, e001341. [Google Scholar] [CrossRef]
- Reynolds, D.S.; Tevis, K.M.; Blessing, W.A.; Colson, Y.L.; Zaman, M.H.; Grinstaff, M.W. Breast Cancer Spheroids Reveal a Differential Cancer Stem Cell Response to Chemotherapeutic Treatment. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Amirabadi, H.E.; Tuerlings, M.; Hollestelle, A.; SahebAli, S.; Luttge, R.; van Donkelaar, C.C.; Martens, J.W.M.; Toonder, J.M.J.D. Characterizing the invasion of different breast cancer cell lines with distinct E-cadherin status in 3D using a microfluidic system. Biomed. Microdevices 2019, 21, 101. [Google Scholar] [CrossRef]
- Wang, X.; Peng, J.; Yang, Z.; Zhou, P.-J.; An, N.; Wei, L.; Zhu, H.H.; Lu, J.; Fang, Y.-X.; Gao, W.-Q. Elevated expression of Gab1 promotes breast cancer metastasis by dissociating the PAR complex. J. Exp. Clin. Cancer Res. 2019, 38, 27. [Google Scholar] [CrossRef]
- Usman, S.; Waseem, N.H.; Nguyen, T.K.N.; Mohsin, S.; Jamal, A.; Teh, M.-T.; Waseem, A. Vimentin Is at the Heart of Epithelial Mesenchymal Transition (EMT) Mediated Metastasis. Cancers 2021, 13, 4985. [Google Scholar] [CrossRef]
- Murray, S.; Schell, K.; McCarthy, D.O.; Albertini, M.R. Tumor growth, weight loss and cytokines in SCID mice. Cancer Lett. 1997, 111, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Siff, T.; Parajuli, P.; Razzaque, M.S.; Atfi, A. Cancer-Mediated Muscle Cachexia: Etiology and Clinical Management. Trends Endocrinol. Metab. 2021, 32, 382–402. [Google Scholar] [CrossRef] [PubMed]
- Aaron, N.; Costa, S.; Rosen, C.J.; Qiang, L. The Implications of Bone Marrow Adipose Tissue on Inflammaging. Front. Endocrinol. 2022, 13, 853765. [Google Scholar] [CrossRef] [PubMed]
- Chen, J. , et al., Prognostic value of neutrophil-to-lymphocyte ratio in breast cancer. FEBS Open Bio, 2015. 5: p. 502-7.
- Muthanna, F.M.; Karuppannan, M.; Abdulrahman, E.; Uitrakul, S.; Rasool, B.A.H.; Mohammed, A.H. Prevalence and Associated Factors of Anemia among Breast Cancer Patients Undergoing Chemotherapy: A Prospective Study. Adv. Pharmacol. Pharm. Sci. 2022, 2022, 1–9. [Google Scholar] [CrossRef]
- Leser, C.; Dorffner, G.; Marhold, M.; Rutter, A.; Döger, M.; Singer, C.; König-Castillo, D.M.; Deutschmann, C.; Holzer, I.; König-Castillo, D.; et al. Liver function indicators in patients with breast cancer before and after detection of hepatic metastases-a retrospective study. PLOS ONE 2023, 18, e0278454. [Google Scholar] [CrossRef]
- Jiramongkol, Y.; Lam, E.W.-F. FOXO transcription factor family in cancer and metastasis. Cancer Metastasis Rev. 2020, 39, 681–709. [Google Scholar] [CrossRef]
- Khan, M.A.; Massey, S.; Ahmad, I.; Sadaf; Akhter, N. ; Habib, M.; Mustafa, S.; Deo, S.V.S.; Husain, S.A. FOXO1 Gene Downregulation and Promoter Methylation Exhibits Significant Correlation With Clinical Parameters in Indian Breast Cancer Patients. Front. Genet. 2022, 13, 842943. [Google Scholar] [CrossRef]
- Pérez-Baena, M.J.; Cordero-Pérez, F.J.; Pérez-Losada, J.; Holgado-Madruga, M. The Role of GAB1 in Cancer. Cancers 2023, 15, 4179. [Google Scholar] [CrossRef]
- Henry, J.P. and B. Bordoni, Histology, Osteoblasts, in StatPearls. 2025, StatPearls Publishing Copyright © 2025, StatPearls Publishing LLC.: Treasure Island (FL).
- Behzatoglu, K. Osteoclasts in Tumor Biology: Metastasis and Epithelial-Mesenchymal-Myeloid Transition. Pathol. Oncol. Res. 2021, 27, 609472. [Google Scholar] [CrossRef]
Figure 1.
Effects of CDPs on migratory and invasiveness of MDA-MB-231 and MCF-7 breast cancer cells. a) Representative photographs of wound closure migration assays taken at 0 h and 48 h in breast cancer cells treated with the saline solution vehicle, CDPs, MTX, or combined CDPs+MTX treatment. b) Representative photographs of the cancer cells that managed to invade the Matrigel-covered membrane stained with crystal violet. The simple culture conditions and co-culture with Raw 264.7 macrophages are shown. (a-b) On the right, the quantification of the wound area and invasive cells from (a) and cell number from (b) is done using ImageJ Software. c) Representative photographs of multicellular microspheroids of MDA-MB-231. d) Cell viability of the microspheroids of MDA-MB-231 and MCF-7 cell cultures submitted to CDPs treatments for 72 h. e) Cell viability of the microspheroids from (d) determined by the MTT assay. f) Determination of cell viability (MTT) and apoptosis (FACS) in the MDA-MB-231 and MCF-7 cell lines treated with CDPs by 4 h. Bars represent means ± SE, n=3. Statistical analysis was performed using one-way ANOVA and Tukey’s post hoc test. SE values are shown as bars, and significant differences are indicated with different lowercase letters (p˂0.05). CDPs, cyclodipeptides; MTX, methotrexate; CDPs-cd, cyclodipeptides dissolved in β-cyclodextrins; CDPs-cd+MTX, combined treatment.
Figure 1.
Effects of CDPs on migratory and invasiveness of MDA-MB-231 and MCF-7 breast cancer cells. a) Representative photographs of wound closure migration assays taken at 0 h and 48 h in breast cancer cells treated with the saline solution vehicle, CDPs, MTX, or combined CDPs+MTX treatment. b) Representative photographs of the cancer cells that managed to invade the Matrigel-covered membrane stained with crystal violet. The simple culture conditions and co-culture with Raw 264.7 macrophages are shown. (a-b) On the right, the quantification of the wound area and invasive cells from (a) and cell number from (b) is done using ImageJ Software. c) Representative photographs of multicellular microspheroids of MDA-MB-231. d) Cell viability of the microspheroids of MDA-MB-231 and MCF-7 cell cultures submitted to CDPs treatments for 72 h. e) Cell viability of the microspheroids from (d) determined by the MTT assay. f) Determination of cell viability (MTT) and apoptosis (FACS) in the MDA-MB-231 and MCF-7 cell lines treated with CDPs by 4 h. Bars represent means ± SE, n=3. Statistical analysis was performed using one-way ANOVA and Tukey’s post hoc test. SE values are shown as bars, and significant differences are indicated with different lowercase letters (p˂0.05). CDPs, cyclodipeptides; MTX, methotrexate; CDPs-cd, cyclodipeptides dissolved in β-cyclodextrins; CDPs-cd+MTX, combined treatment.
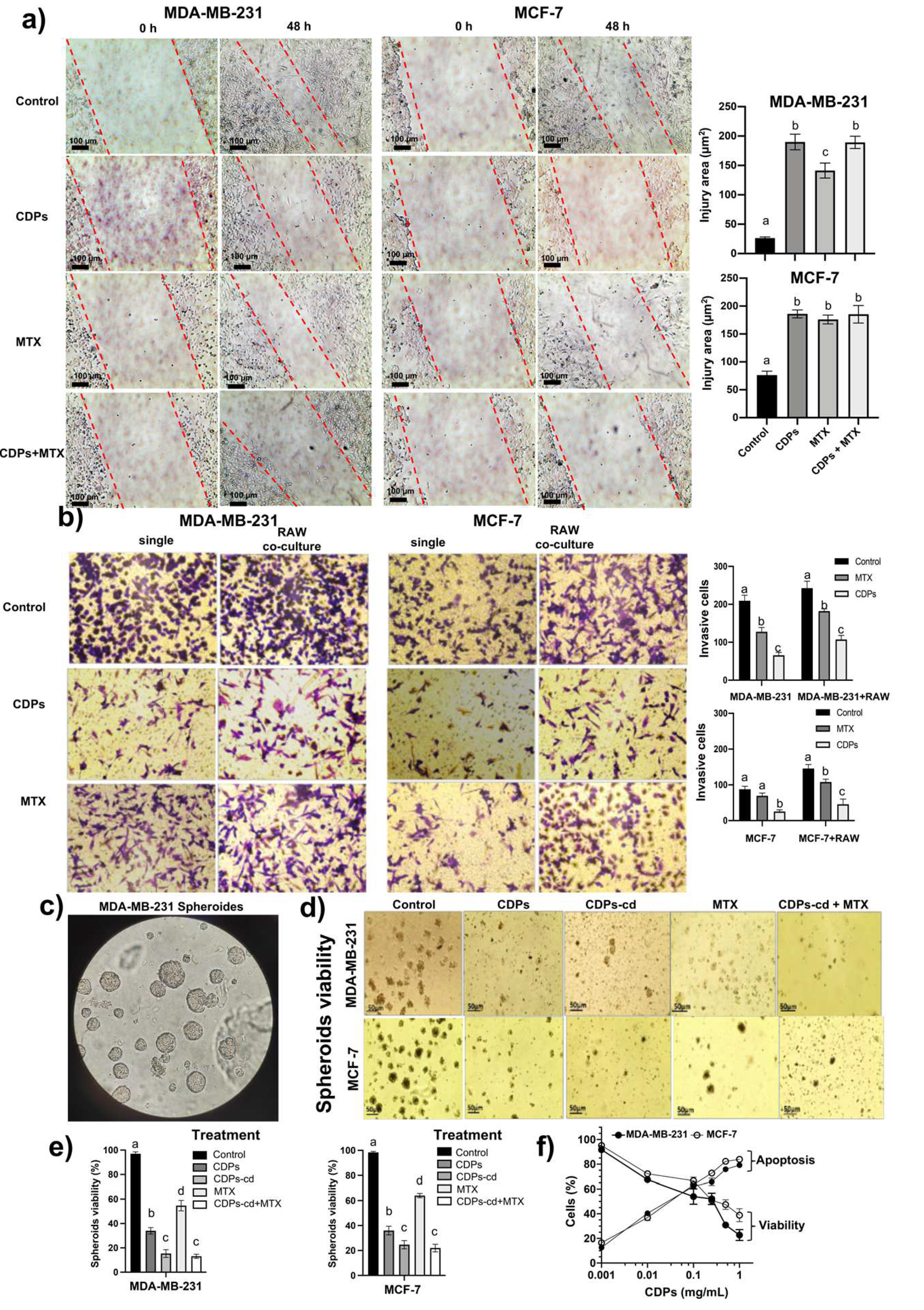
Figure 2.
Effect of CDPs on proteins involved in proliferation, invasion, and metastasis in MDA-MB-231 triple-negative breast cancer cells. Protein extracts obtained from the cell cultures treated with CDPs (0.1 mg/mL) in function of time were subjected to immunodetection. Protein expression for the total Akt, phosphorylated Akt, total mTOR, phosphorylated mTOR, total S6K, phosphorylated S6K, Vimentin, Gab1, and b-actin are shown. Densitometric analyses of the protein immunodetection results are presented on the right. The bars represent the means ± SE, n=3 per group. Statistical analysis was performed using one-way ANOVA, followed by Tukey’s post hoc test; SE values are shown as bars, and significant differences are indicated with different lowercase letters (p <0.05).
Figure 2.
Effect of CDPs on proteins involved in proliferation, invasion, and metastasis in MDA-MB-231 triple-negative breast cancer cells. Protein extracts obtained from the cell cultures treated with CDPs (0.1 mg/mL) in function of time were subjected to immunodetection. Protein expression for the total Akt, phosphorylated Akt, total mTOR, phosphorylated mTOR, total S6K, phosphorylated S6K, Vimentin, Gab1, and b-actin are shown. Densitometric analyses of the protein immunodetection results are presented on the right. The bars represent the means ± SE, n=3 per group. Statistical analysis was performed using one-way ANOVA, followed by Tukey’s post hoc test; SE values are shown as bars, and significant differences are indicated with different lowercase letters (p <0.05).
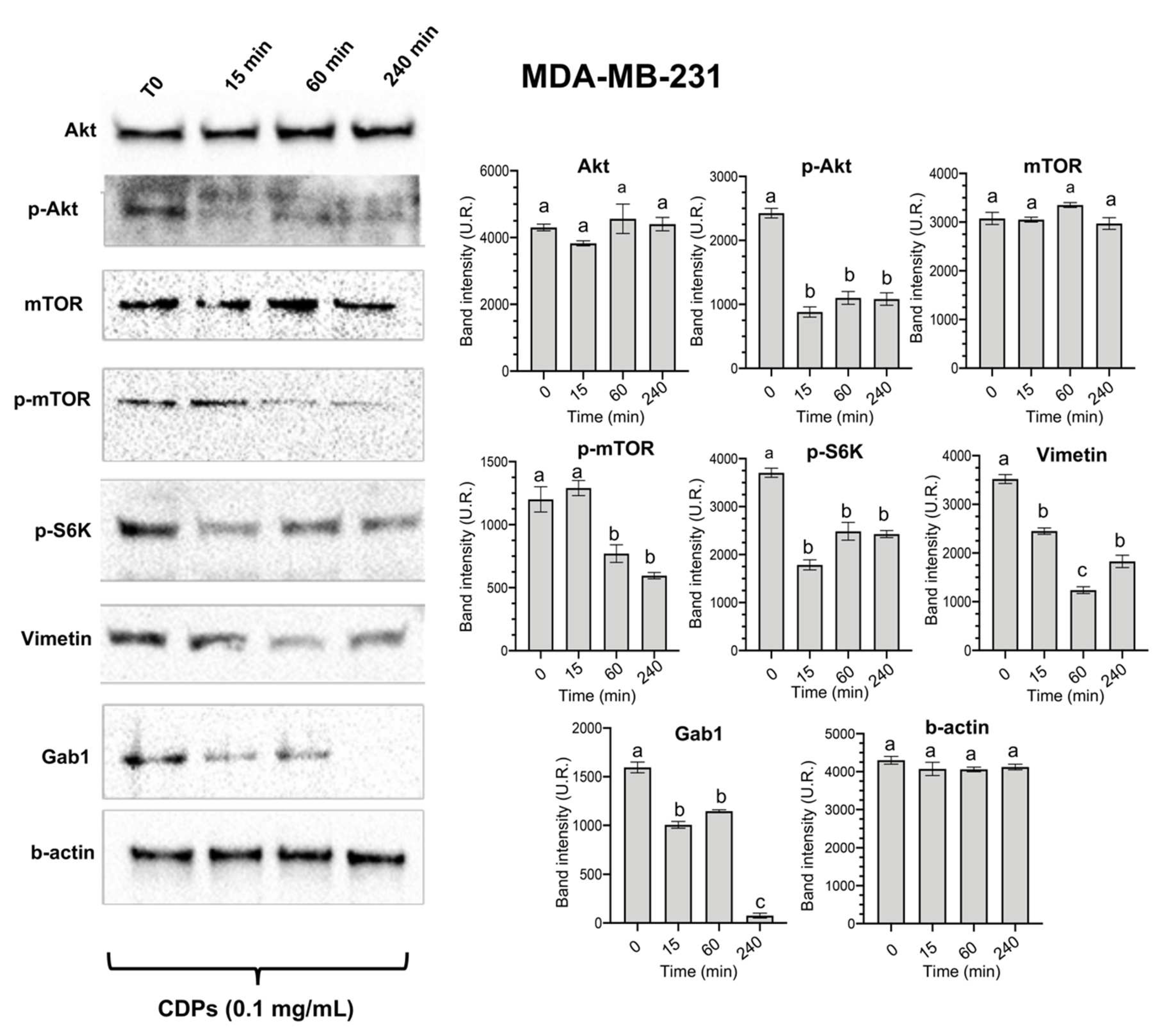
Figure 3.
Anti-tumor procedure in MDA-MB-231 immunosuppressed mice triple-negative breast cancer model by CDPs treatment. Female Balb/c nu/nu mice (8 weeks old) were submitted to an anti-tumor therapeutic study. a) Two therapeutic experimental outlines were tested: 1) the MDA-MB-231 cells line was inoculated in mice together with the CDPs administration, following nine doses of CDPs (0.25 mg/kg) under 35 days of procedure (arrows). 2) The second therapeutic experimental outline consisted of the development of mammary tumor by MDA-MB-231cells line inoculation during 35 days; after that, administration of nine doses of CDPs (0.25 mg/kg), MTX (0.05 mg/kg), or CDPs+MTX co-administrated during 35 days of procedure (arrows). b) The body weight was monitored during the therapeutic experimental outline study. The slope of weight gain values is shown in all the mice groups (right). Statistical analysis was performed using t-student; values are displayed, and significant differences are indicated with asterisks (p <0.05), n=5 per group. Mice groups: Control (C), healthy animals control without treatment; C+CDPs, healthy animals administered with CDPs; BC, animals that developed breast cancer without treatment; BC+CDPs (i), BC animals administered with CDPs as the procedure 1; BC+CDPs (m), BC animals administered with CDPs as procedure 2; BC+MTX (m), BC animals administered with methotrexate (MTX) as procedure 2; BC+CDPs+MTX (m), BC animals co-administered with CDPs + methotrexate as procedure 2.
Figure 3.
Anti-tumor procedure in MDA-MB-231 immunosuppressed mice triple-negative breast cancer model by CDPs treatment. Female Balb/c nu/nu mice (8 weeks old) were submitted to an anti-tumor therapeutic study. a) Two therapeutic experimental outlines were tested: 1) the MDA-MB-231 cells line was inoculated in mice together with the CDPs administration, following nine doses of CDPs (0.25 mg/kg) under 35 days of procedure (arrows). 2) The second therapeutic experimental outline consisted of the development of mammary tumor by MDA-MB-231cells line inoculation during 35 days; after that, administration of nine doses of CDPs (0.25 mg/kg), MTX (0.05 mg/kg), or CDPs+MTX co-administrated during 35 days of procedure (arrows). b) The body weight was monitored during the therapeutic experimental outline study. The slope of weight gain values is shown in all the mice groups (right). Statistical analysis was performed using t-student; values are displayed, and significant differences are indicated with asterisks (p <0.05), n=5 per group. Mice groups: Control (C), healthy animals control without treatment; C+CDPs, healthy animals administered with CDPs; BC, animals that developed breast cancer without treatment; BC+CDPs (i), BC animals administered with CDPs as the procedure 1; BC+CDPs (m), BC animals administered with CDPs as procedure 2; BC+MTX (m), BC animals administered with methotrexate (MTX) as procedure 2; BC+CDPs+MTX (m), BC animals co-administered with CDPs + methotrexate as procedure 2.
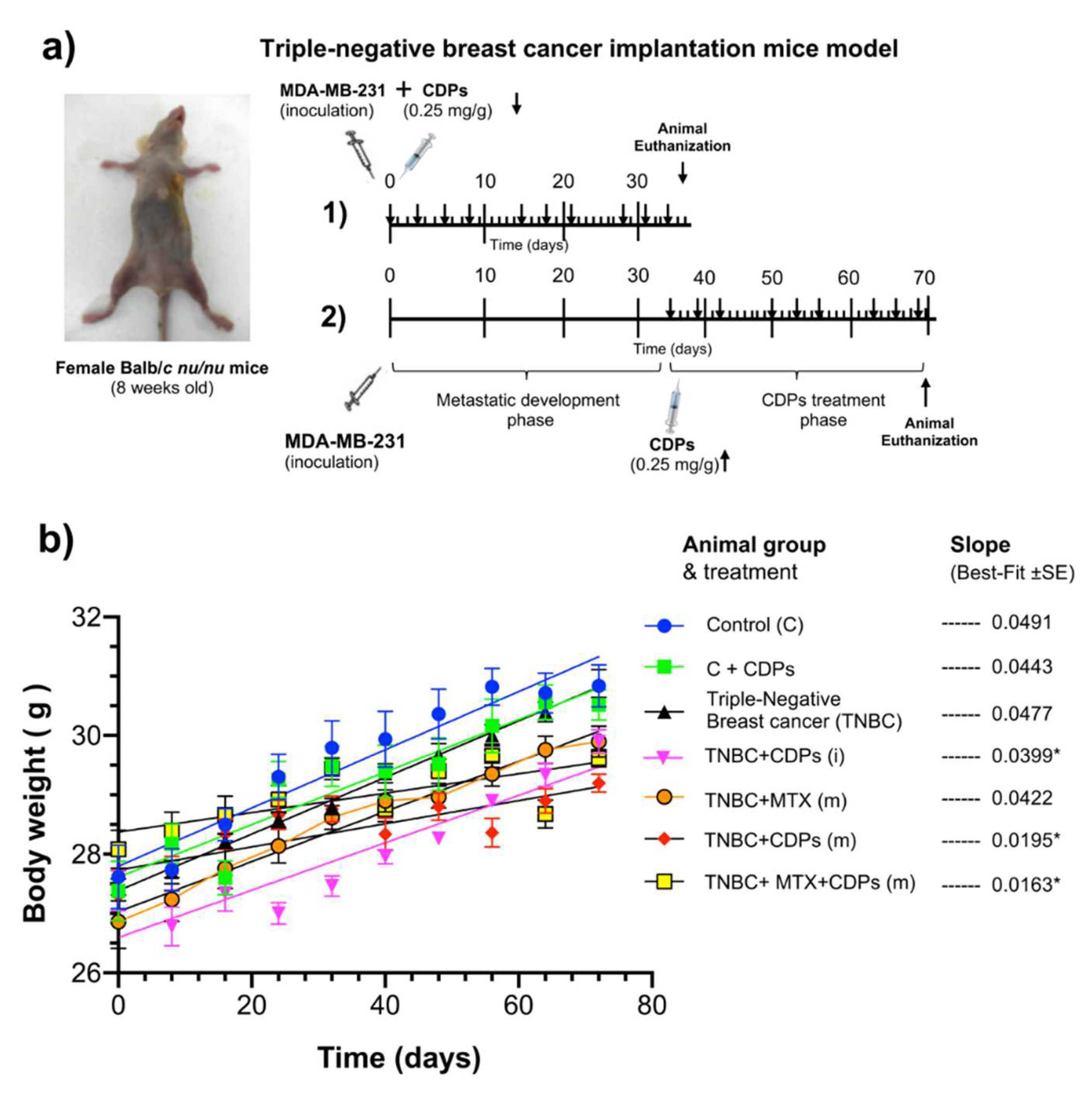
Figure 4.
Anti-tumor effect of CDPs on xenografted triple-negative breast cancer MDA-MB-231 cells in immunosuppressed mice. a) Photograph of mouse groups on the 70th day of the therapeutic experimental procedure before euthanizing. Photographs of dissected tumors from each mouse group are shown, n=5 per group; nomenclature is indicated in Figure 3. The scale of the ruler corresponds to centimeters. b) Determination of tumor volume expressed in mm3. c) Tumor weight expressed in grams. The results are presented as the means ± SE, n = 5. One-way ANOVA with Bonferroni post-hoc test was used to compare treatments with the untreated group (TNBC); significant differences (p< 0.01) are indicated with an asterisk (*).
Figure 4.
Anti-tumor effect of CDPs on xenografted triple-negative breast cancer MDA-MB-231 cells in immunosuppressed mice. a) Photograph of mouse groups on the 70th day of the therapeutic experimental procedure before euthanizing. Photographs of dissected tumors from each mouse group are shown, n=5 per group; nomenclature is indicated in Figure 3. The scale of the ruler corresponds to centimeters. b) Determination of tumor volume expressed in mm3. c) Tumor weight expressed in grams. The results are presented as the means ± SE, n = 5. One-way ANOVA with Bonferroni post-hoc test was used to compare treatments with the untreated group (TNBC); significant differences (p< 0.01) are indicated with an asterisk (*).
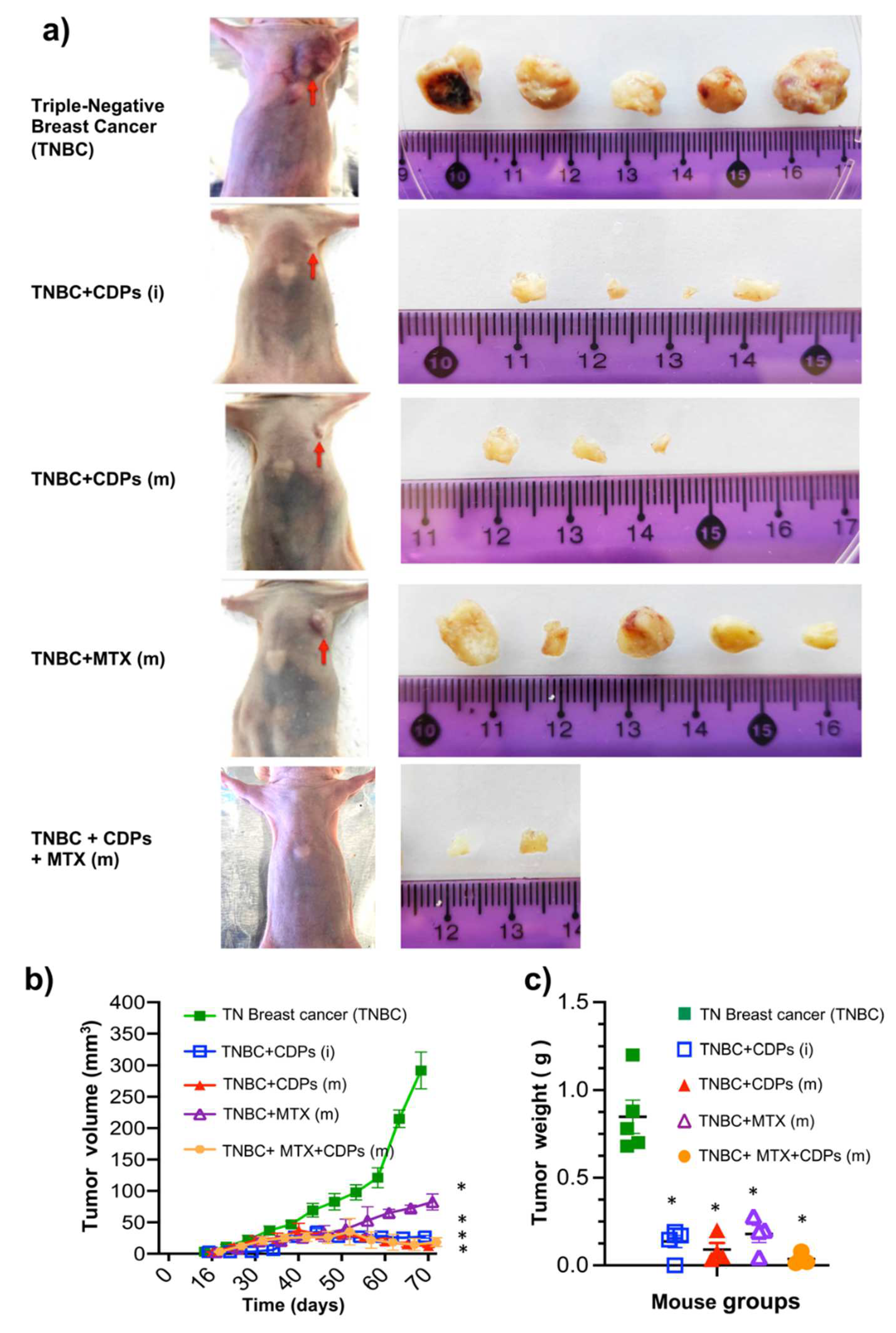
Figure 5.
Anti-tumor effect of CDPs on organs of xenografted triple-negative breast cancer MDA-MB-231 cells in immunosuppressed mice. a) Photograph of lungs and liver of mice dissected on the 70th day of the therapeutic experimental procedure. Arrows indicate metastatic foci in organs. Representative photographs of organs from each mouse group are shown, n=5 per group; nomenclature is shown in Figure 3. b) Determination of organ weight expressed in grams. c) Determination of enzyme activity of AST, ALT, and LDH in serum of mice groups. d) Hematological profile, hemoglobin (g/dL), and leucocyte counts in mice with TNBC implantation. Graphs represent the means ± SE, n = 5. One-way ANOVA with Bonferroni post-hoc test was used to compare treatments with the control group (C); significant differences (p< 0.05) are indicated with asterisks (*) with different lowercase letters (p <0.05) showed Tukey’s test.
Figure 5.
Anti-tumor effect of CDPs on organs of xenografted triple-negative breast cancer MDA-MB-231 cells in immunosuppressed mice. a) Photograph of lungs and liver of mice dissected on the 70th day of the therapeutic experimental procedure. Arrows indicate metastatic foci in organs. Representative photographs of organs from each mouse group are shown, n=5 per group; nomenclature is shown in Figure 3. b) Determination of organ weight expressed in grams. c) Determination of enzyme activity of AST, ALT, and LDH in serum of mice groups. d) Hematological profile, hemoglobin (g/dL), and leucocyte counts in mice with TNBC implantation. Graphs represent the means ± SE, n = 5. One-way ANOVA with Bonferroni post-hoc test was used to compare treatments with the control group (C); significant differences (p< 0.05) are indicated with asterisks (*) with different lowercase letters (p <0.05) showed Tukey’s test.
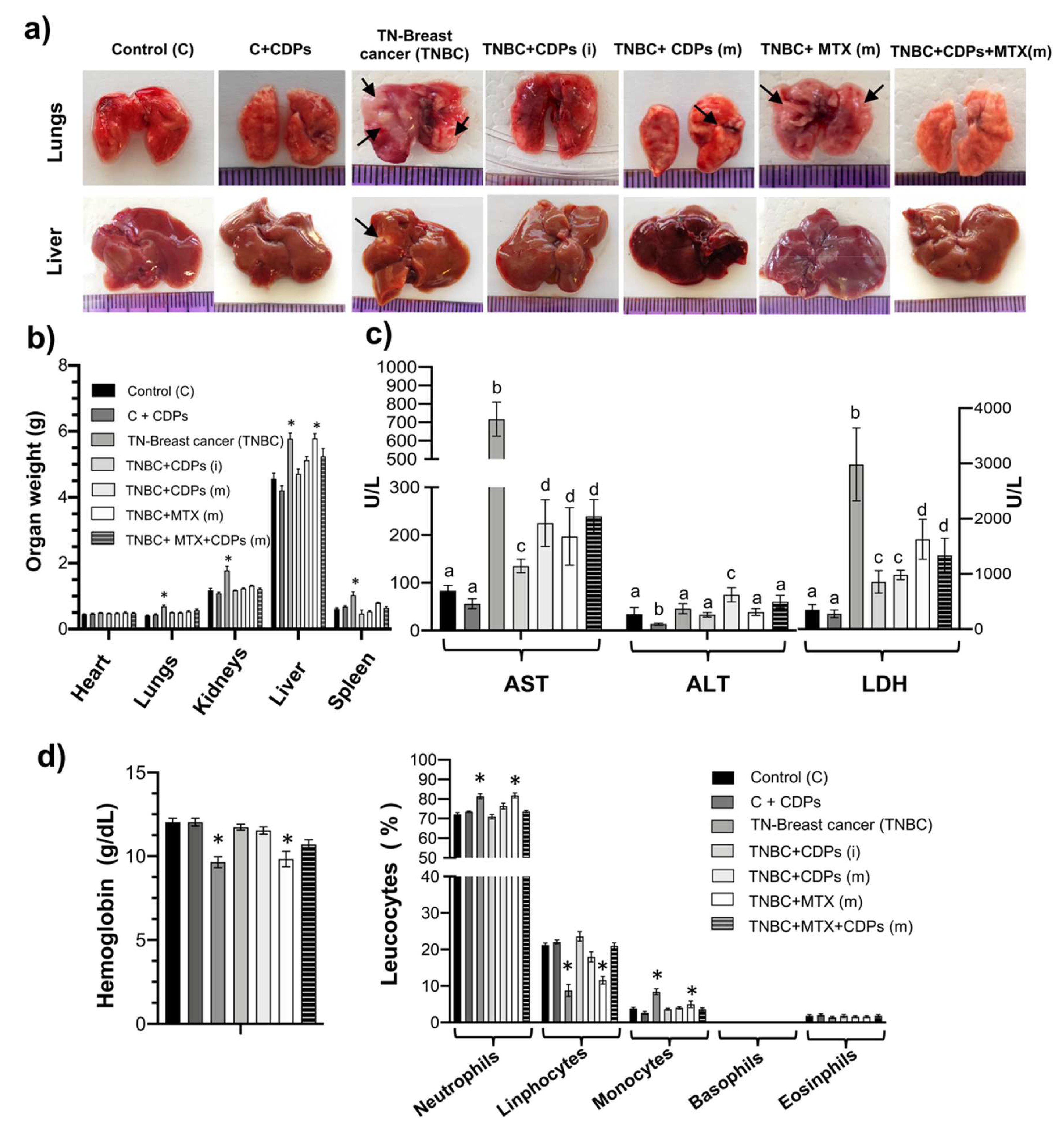
Figure 6.
Histological features of tumors, lungs, and liver of xenografted triple-negative breast cancer MDA-MB-231 cells in immunosuppressed mice treated with CDPs. Female Balb/c nu/nu mice (8 weeks old) were submitted to an anti-tumor study by 70 days, n=5 per group. Tumors, lungs, and liver tissue sections were stained with hematoxylin and eosin (H&E) and observed using microscopy; representative photographs of tissues dissected from the animal groups are shown and named in Figure 3. The fibrosis areas (yellow arrows) and fat drops (black arrows) are shown in tumoral histology pictures. In lung histology pictures, the MDA-MB-231 cell infiltration is circled with a red dot line, while the leukocyte infiltration in liver tissue is shown with red arrowheads.
Figure 6.
Histological features of tumors, lungs, and liver of xenografted triple-negative breast cancer MDA-MB-231 cells in immunosuppressed mice treated with CDPs. Female Balb/c nu/nu mice (8 weeks old) were submitted to an anti-tumor study by 70 days, n=5 per group. Tumors, lungs, and liver tissue sections were stained with hematoxylin and eosin (H&E) and observed using microscopy; representative photographs of tissues dissected from the animal groups are shown and named in Figure 3. The fibrosis areas (yellow arrows) and fat drops (black arrows) are shown in tumoral histology pictures. In lung histology pictures, the MDA-MB-231 cell infiltration is circled with a red dot line, while the leukocyte infiltration in liver tissue is shown with red arrowheads.
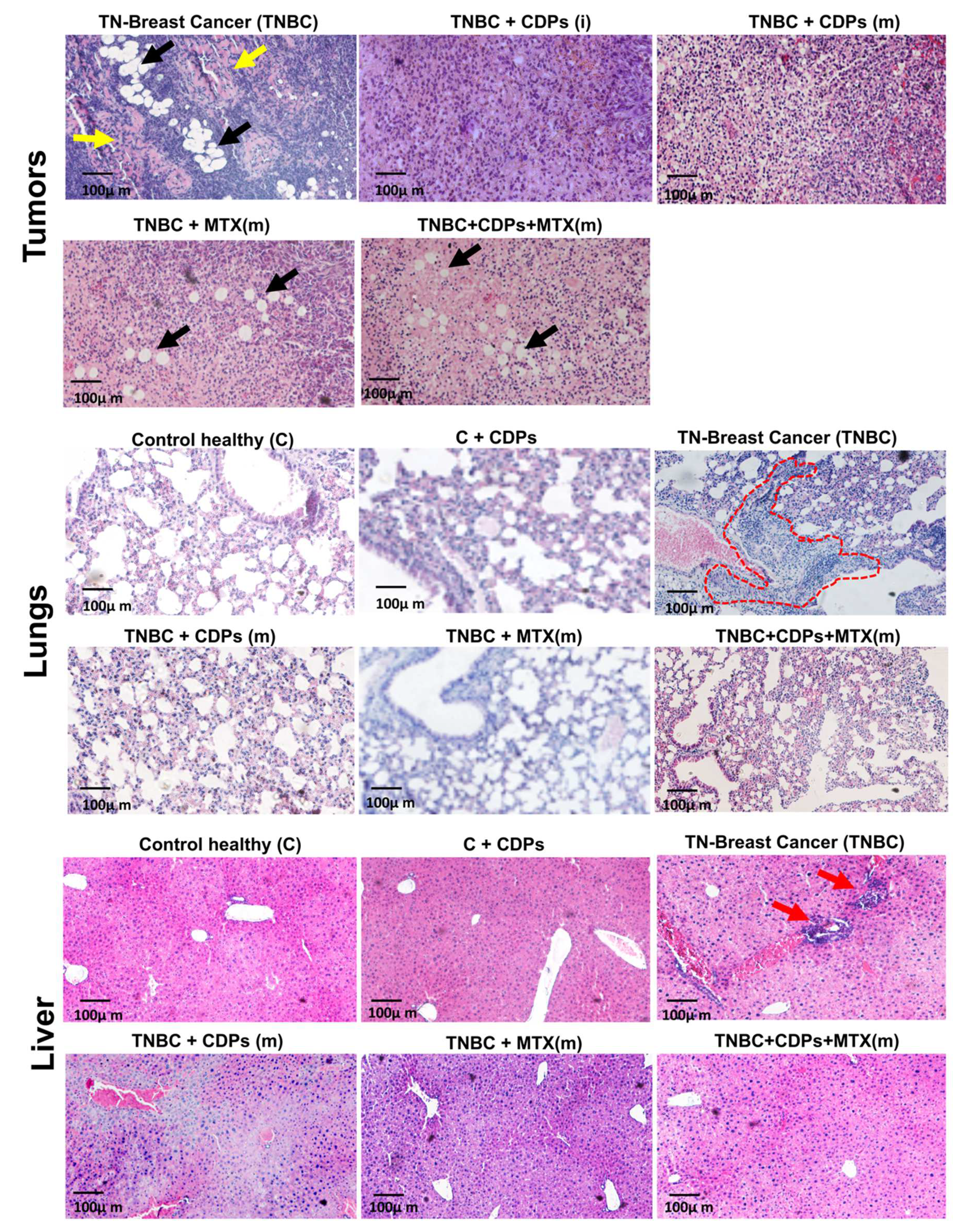
Figure 7.
Effect of CDPs on signaling proteins in tumor xenografted with MDA-MB-231 triple-negative breast cancer cells in immunosuppressed mice treated with CDPs. Protein extracts from the tumor dissected from the mice groups (administered with CDPs, MTX, and co-administered CDPs+MTX) were subjected to immunodetection. The animal group’s nomenclature is indicated in Figure 3. Protein expression for the total Akt, phosphorylated Akt, total Gab1, phosphorylated p-Gab1, total FOX01, and phosphorylates p-FOX01 are shown. Densitometric analyses of the protein immunodetection results are presented below. The bars represent the means ± SE, n=3 per group. Statistical analysis was performed using one-way ANOVA, followed by Tukey’s posthoc test; significant differences are indicated with different lowercase letters (p <0.05).
Figure 7.
Effect of CDPs on signaling proteins in tumor xenografted with MDA-MB-231 triple-negative breast cancer cells in immunosuppressed mice treated with CDPs. Protein extracts from the tumor dissected from the mice groups (administered with CDPs, MTX, and co-administered CDPs+MTX) were subjected to immunodetection. The animal group’s nomenclature is indicated in Figure 3. Protein expression for the total Akt, phosphorylated Akt, total Gab1, phosphorylated p-Gab1, total FOX01, and phosphorylates p-FOX01 are shown. Densitometric analyses of the protein immunodetection results are presented below. The bars represent the means ± SE, n=3 per group. Statistical analysis was performed using one-way ANOVA, followed by Tukey’s posthoc test; significant differences are indicated with different lowercase letters (p <0.05).
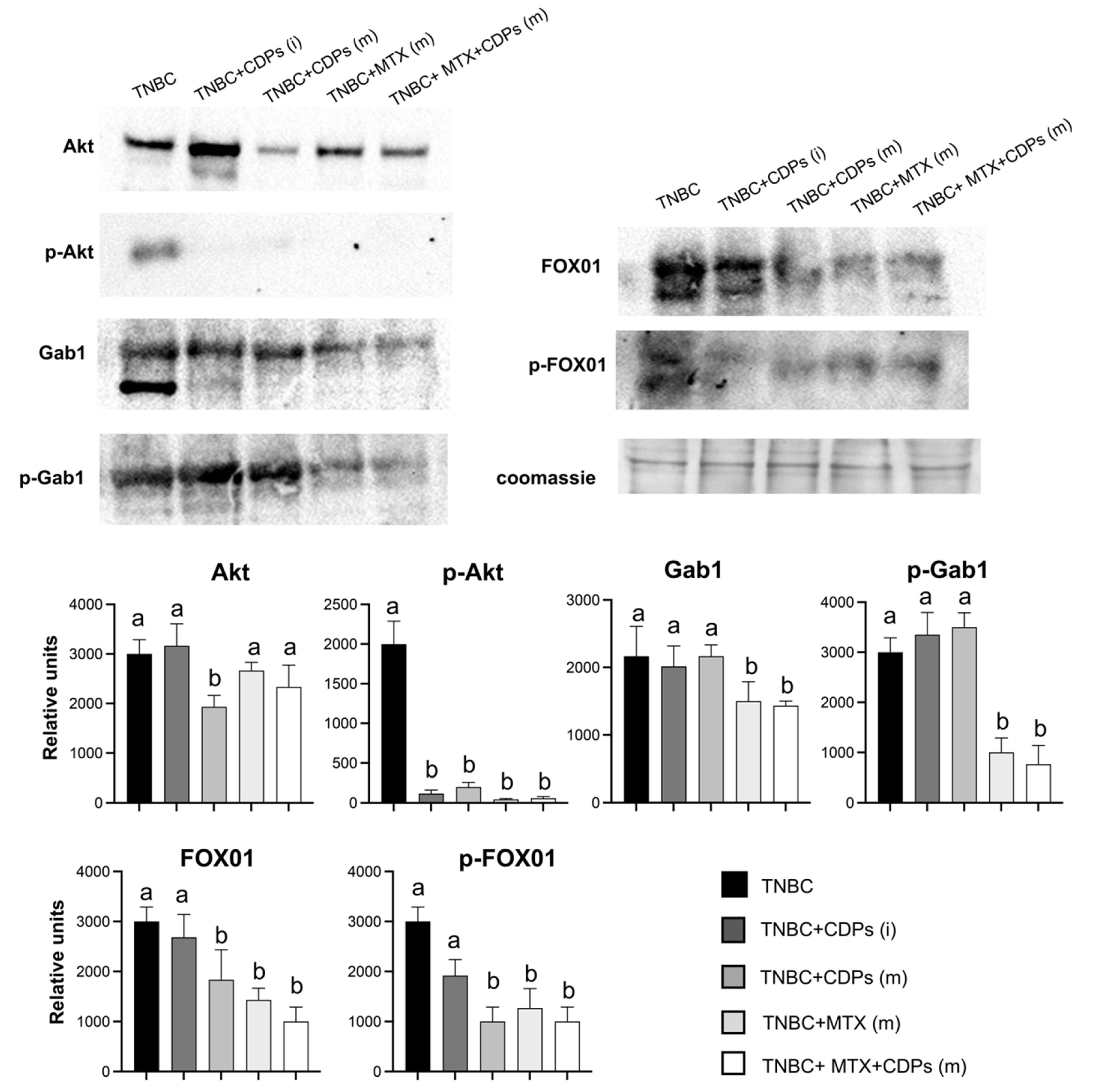
Figure 8.
Analysis of bone cancer in xenografted breast cancer MDA-MB-231 cells in immunosuppressed mice treated with CDPs. Female Balb/c nu/nu mice (8 weeks old) were submitted to an anti-tumor study by 70 days, n=5 per group. Animal groups are named as in Figure 3. a) Determination of weight in the femurs from animals after the therapeutic experimental procedure. b) Quantification of osteoclasts and osteoblasts in H&E-stained femur tissue dissections. Bars represent the means ± SE, n = 5. One-way ANOVA using Tukey’s post-hoc test to compare treatments; significant differences (p< 0.05) are indicated as different lowercase letters.
Figure 8.
Analysis of bone cancer in xenografted breast cancer MDA-MB-231 cells in immunosuppressed mice treated with CDPs. Female Balb/c nu/nu mice (8 weeks old) were submitted to an anti-tumor study by 70 days, n=5 per group. Animal groups are named as in Figure 3. a) Determination of weight in the femurs from animals after the therapeutic experimental procedure. b) Quantification of osteoclasts and osteoblasts in H&E-stained femur tissue dissections. Bars represent the means ± SE, n = 5. One-way ANOVA using Tukey’s post-hoc test to compare treatments; significant differences (p< 0.05) are indicated as different lowercase letters.
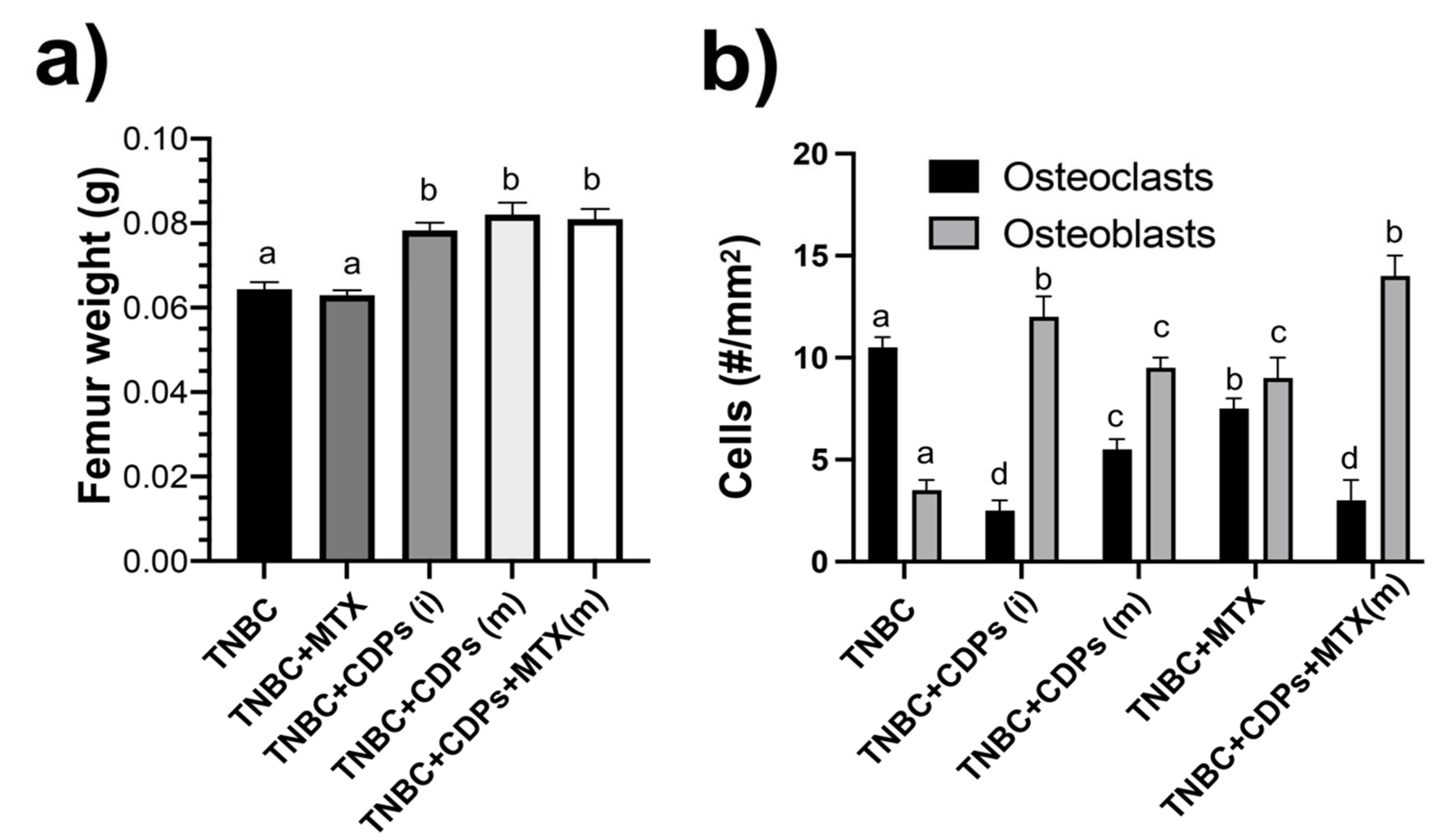
Figure 9.
Histological features of femurs from xenografted TNBC animals with the MDA-MB-231 cells treated with CDPs. Female Balb/c nu/nu mice (8 weeks old) were submitted to an anti-tumor study by 70 days, n=5 per group. Femur tissue sections were stained with H&E and observed using microscopy; representative photographs of tissues dissected from the animal groups are shown and named as in Figure 3. Red dotted lines outline tumor cell populations indicating tumor bone (T), which are observed as pink areas. The purple region represents the bone marrow (M), osteoblasts (Osb), osteoclasts (OCL), osteocytes (Ocy), and cortical bone (B). Scale bar =100 μm and ×4, ×10, or ×40X magnifications are shown.
Figure 9.
Histological features of femurs from xenografted TNBC animals with the MDA-MB-231 cells treated with CDPs. Female Balb/c nu/nu mice (8 weeks old) were submitted to an anti-tumor study by 70 days, n=5 per group. Femur tissue sections were stained with H&E and observed using microscopy; representative photographs of tissues dissected from the animal groups are shown and named as in Figure 3. Red dotted lines outline tumor cell populations indicating tumor bone (T), which are observed as pink areas. The purple region represents the bone marrow (M), osteoblasts (Osb), osteoclasts (OCL), osteocytes (Ocy), and cortical bone (B). Scale bar =100 μm and ×4, ×10, or ×40X magnifications are shown.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



