Submitted:
21 February 2025
Posted:
25 February 2025
You are already at the latest version
Abstract
Type 1 diabetes (T1D) is a chronic autoimmune disease characterized by the progressive destruction of pancreatic β-cells, leading to insulin deficiency. The primary drivers of β-cell destruction in T1D involve autoimmune-mediated processes that trigger chronic inflammation and ultimately β-cell loss. Regulatory microRNAs (miRNAs) play a crucial role in modulating these processes by regulating gene expression through post-transcriptional suppression of target mRNAs. Dysregulated miRNAs have been implicated in T1D pathogenesis, serving as both potential diagnostic biomarkers and therapeutic targets. This review explores the role of miRNAs in T1D, highlighting their involvement in disease mechanisms across both rodent models and human patients. While current anti-diabetic therapies manage T1D symptoms, they do not prevent β-cell destruction, leaving patients reliant on lifelong insulin therapy. By summarizing key upregulated and downregulated miRNAs in diabetic models and patients, this review discusses the potential of miRNA-based therapies to restore β-cell function and modify disease progression.
Keywords:
Type 1 diabetes
; autoimmune
; β-cells
; apoptosis
; miRNAs
1. Introduction
Diabetes mellitus, commonly referred to as diabetes, is characterized by chronic hyperglycemia and associated with metabolic dysfunctions in carbohydrates, fats, and proteins [1]. The disease encompasses genetic, pathophysiological, and clinical factors [2]. Diabetes is categorized into several types based on its etiology and pathogenesis: type 1 diabetes (T1D), type 2 diabetes (T2D), gestational diabetes (GD), and other forms [1]. T1D, or insulin-dependent diabetes, results from dysfunction of pancreatic β-cells and accounts for about 5-10% of all diabetes cases [1]. T2D, or non-insulin-dependent diabetes, is primarily caused by insulin resistance and insulin deficiency, making up 90-95% of all cases [1]. GD is diagnosed when glucose intolerance or diabetes develops during pregnancy, typically in the second or third trimester, and affects 1-14% of pregnancies [1]. All forms of diabetes share the common feature of insulin dysregulation, either absolute or relative. Currently, there are no reliable biomarkers for diagnosing diabetes before it develops. Although various tests to assess insulin secretion, such as fasting indices, oral and intravenous glucose tolerance tests, and other provocative challenges, have been proposed, they are not widely used due to their time-consuming nature, expense, and lack of standardization [3]. Furthermore, these methods do not adequately reflect the underlying pathophysiology of β-cell dysfunction, as their correlation with β-cell mass is limited [3].
T1D, also known as one of the most common chronic diseases of childhood, affects approximately 8.4 million patients globally, according to the World Health Organization [4]. T1D results from the loss of pancreatic islet β-cells, often through autoimmune activation, β-cell autoantigen release, oxidative and endoplasmic reticulum (ER) stress, and cytokine-mediated damage, leading to β-cell destruction, insulin deficiency, and hyperglycemia through disruption of insulin signaling pathways (Figure 1) [5,6,7,8]. Although diverse causes of destruction of pancreatic islet β-cells were identified, the molecular mechanisms of the loss of pancreatic islet β-cells remains controversial. T1D is categorized into T1Da and T1Db [6]. Most patients (70-90%) fall into T1Da, an autoimmune form known as autoimmune T1D while T1Db, or idiopathic T1D, includes a small subset of patients [6].
There are a few biomarkers available to distinguish T1D from other types of diabetes. In autoimmune T1D, autoantibodies (AAbs) against β-cell antigens are produced early, often before clinical symptoms appear [9]. This early antibody production is often accompanied by genetic mutations associated with the disease, with variations in the human leukocyte antigen (HLA) class II genes being the most common [10]. The presence of two or more AAbs indicates a high risk for T1D, as it signals an active autoimmune response against β-cells [7,8]. Several AAbs targeting islet cells, insulin, tyrosine phosphatases, IA-2 and IA-2β, glutamic acid decarboxylase, and zinc transporter 8 serve as biomarkers for T1D [3,7,8,11,12]. While these AAbs and measurements of T-cell reactivity reliably identify individuals at risk of T1D by assessing β-cell dysfunction due to autoimmune activity, they lack precision in predicting T1D onset and are not effective for monitoring disease progression [3,11,12].
Furthermore, while AAbs are the most common biomarkers for T1D, they are only applicable for identifying T1D in AAbs-positive individuals. Notably, some children in the initial stage may be negative for islet AAbs and some AAbs-positive individuals may not develop T1D [7,8]. Moreover, despite extensive research into the molecular mechanisms underlying T1D pathogenesis, patients with T1D still rely on lifelong insulin therapy, which often has limited tolerance and can lead to adverse effects [13]. Thus, novel biomarkers to assess β-cell dysfunction are essential to predict T1D onset, monitor its progression, and establish effective clinical approaches in T1D therapies.
Sustained high blood glucose levels are associated with several serious diseases, including retinopathy, nephropathy, neuropathy, and cardiovascular conditions such as coronary artery disease, atherosclerosis, hypertension, and stroke [14]. Key pathological factors like hyperglycemia, hyperlipidemia, advanced glycation end products (AGEs), growth factors, and inflammatory cytokines/chemokines contribute to the increased risk of these complications [14,15,16]. Diabetic patients with these complications often require lifelong treatment, and if left unmanaged, can significantly reduce their quality of life [14]. Therefore, understanding the molecular mechanisms underlying the onset of diabetic complications is crucial for developing effective treatments.
MicroRNAs (miRNAs) can serve as biomarkers and therapeutic targets in various human diseases. miRNAs are small, noncoding RNA molecules, typically 21 to 23 nucleotides in length, that regulate gene expression. They do so by binding to the 3' untranslated regions of target messenger RNAs (mRNAs), forming the RNA-induced silencing complex [8,17]. This complex directly modulates the translation of mRNAs into proteins [8,17]. An increasing number of studies suggest miRNAs hold significant potential as biomarkers for pathogenic conditions, and as therapeutic agents for medical intervention in nearly all human disease, including T1D and T2D. Studies have shown that changes in specific miRNA expression are associated with T1D and hyperglycemia, suggesting their potential as biomarkers for T1D. miR-125b-5p and miR-365a-3p showed positive correlation with hemoglobin A1C levels [18], whereas let-7a-5p, let-7c-5p, miR-5190, and miR-770-5p exhibited negative correlations [18,19]. These miRNAs significantly influence glycosaminoglycan biosynthesis, axon guidance signaling, Rap1 signaling, focal adhesion, and neurotrophin signaling [18,20].
Recent studies have emphasized the therapeutic potential of miRNA-based strategies, such as using miRNA mimics to restore downregulated miRNAs or employing miRNA inhibitors to counteract overexpressed miRNAs, thereby achieving protein homeostasis [21,22]. The safety of prolonged treatment with specific miRNAs has also been demonstrated in preclinical models [23]. With six FDA-approved siRNA drugs (Patisiran, Givosiran, Lumasiran, Inclisiran, Vutrisiran, and Nedosiran) now in clinical use for genetic and metabolic disorders, ongoing research on miRNAs—mimicked by these siRNAs—continues to expand their applications in both preclinical and clinical settings. However, several critical challenges remain to fully harness the potential of miRNA-based therapies. These include identifying key signature miRNAs, elucidating their mechanisms of action, optimizing their use through RNAi, ensuring efficient delivery to target tissues, and validating their efficacy in vivo. Although several miRNA-based therapeutics have entered clinical trials [24], none have yet achieved FDA approval.
Numerous studies have identified dysregulated miRNAs in both T1D rodent models and patients. This review explores differentially expressed miRNAs in T1D, emphasizing their interactions with target genes involved in T1D-related pathways to uncover the molecular mechanisms underlying β-cell dysfunction in rodents and humans. Furthermore, it highlights commonly upregulated and downregulated miRNAs shared between T1D rodent models and patients. These insights aim to advance the development of diagnostic and therapeutic strategies for T1D.
2. Dysregulated miRNAs in T1D Patients
Tissue-specific gene expression patterns are crucial for tissue development, defining cell type characteristics, functions, and transcriptional regulation [25,26]. miRNAs, which exhibit both tissue- and developmental stage-specific features, play an essential role in tissue identity and function [26,27,28]. Numerous studies have demonstrated correlations between tissue-specific miRNAs and various human diseases [26]. Circulating miRNAs, influenced by tissues such as the heart, liver, pancreas, kidney, colon, and lung, have also been shown to play key roles in disease processes, with many originating from blood cells [29]. These circulating miRNAs are easily accessible, relatively stable, and exhibit disease-specific profiles [30].
Dysregulated miRNAs in Various Samples from T1D Patients
Over the past decades, numerous studies have documented the dysregulation of miRNAs in various specimens, including serum, plasma, peripheral blood, PBMCs, isolated T-cells, urine, and exosomes from T1D patients. Table 1 provides a summary of dysregulated miRNAs identified across these sample types. In T1D patients, miRNAs exhibit differential upregulation and downregulation depending on the sample type, with certain miRNAs consistently dysregulated across multiple specimens.
Recent research has identified 138 upregulated and 93 downregulated miRNAs that are consistently altered in T1D patients compared to healthy donors (Table 1). Among them, five miRNAs in various specimens were reported to be dysregulated in T1D patients: miR-21 [3,20,31,32,33,34,35] and miR-148a [3,20,31,32,33,34] were upregulated, while miR-126 [32,34,35,37] was downregulated. miR-25-3p [19,34,37,43] and miR-1275 [20,34,38] had different dysregulated patterns in various specimens. miR-21 and miR-148a regulate inflammation and apoptosis. In patients with T1D, increased expression of miR-21 and miR-148a regulates PI3K/AKT signaling and contributes to apoptosis in β-cells leading to dysregulated insulin release through impaired glucose-stimulated insulin secretion through PTEN and SOX6 [33,45,46,47]. Conversely, decreased expression of miR-25-3p, miR-126, and miR-1275 targets IL-1β and IRS-1, promoting β-cell death [48,49,50,51,52].
Notably, one interesting study [34] found that both miR-148a-3p and miR-148b-3p levels in plasma were increased in newly diagnosed T1D patients (<5 years of diagnosis) but were decreased in later-stage T1D patients (≥5 years of diagnosis). These miRNAs are regulated with the Wnt, FOXO, and insulin signaling pathway [33,34]. In the early stages of T1D, pancreatic β-cell damage progresses due to autoimmunity [5]. During this stage, the increased expression of miR-148a/b-3p show to regulate the Wnt and insulin signaling pathways, protecting the remaining pancreatic cells or modulating inflammatory responses. In the later stages, as pancreatic β-cell damage worsens, pancreatic function is lost, and chronic inflammation leads to fibrosis [1,5], the role of miR-148a/b-3p diminishes, and its expression levels decrease. These miRNAs may serve as valuable resources for identifying novel biomarkers across different stages of T1D.
Dysregulated miRNAs in Serum and Plasma
Circulating miRNAs, found in nearly all biological fluids such as serum and plasma, represent promising sensitive biomarkers for a range of conditions, as their profiles provide an accurate representation of the physiological state of the organism [53,54,55]. In particular, miRNAs expressed in serum exhibit changes in expression levels in response to disease states and physiological changes [55,56,57,58]. They exhibit altered expression profiles in various pathological conditions, including metabolic diseases (e.g., diabetes), cancer, cardiovascular diseases, and inflammatory disorders [53,54,58,59,60,61]. These changes make them valuable biomarkers for early disease diagnosis, prognosis evaluation, and monitoring therapeutic responses.
Research into the expression of these miRNA markers suggests their potential application in the early diagnosis of various diseases, frequently preceding the manifestation of clinical symptoms [57,62,63,64,65,66]. Additionally, these markers may play a crucial role in evaluating patient responses to therapeutic treatments, thus aiding in formulating personalized treatment approaches [67,68].
The analysis of miRNA profiles in T1D patients highlights distinct patterns of dysregulation in both serum and plasma, underscoring their potential as biomarkers for the disease. Specifically, 15 miRNAs were upregulated and 7 were downregulated in serum, while 18 miRNAs were upregulated and 3 miRNAs were down regulated in plasma (Table 1). Notable examples include the downregulation of miR-10b-5p, miR-146a-5p and miR-409-3p [19,33,69], and the upregulation of miR-21, miR-126, and miR-155-5p [18,70,71,72], which are associated with key pathways in T1D pathogenesis. These findings suggest that miRNAs can provide insights into T1D disease mechanisms and can serve as a foundation for developing diagnostic and therapeutic tools for T1D.
Dysregulated miRNAs in Blood Cells
Approximately 80% of all human genes are expressed in peripheral blood cells, including peripheral blood mononuclear cells (PBMCs). These cells are highly responsive to environmental changes, which can significantly impact their gene expression profiles [20,73]. In the context of T1D, hundreds of miRNAs have been reported to exhibit differential expression in PBMCs [8]. The dysfunction of PBMCs, particularly T-cells and B-cells, plays a critical role in the pathogenesis of autoimmune diseases such as T1D. Therefore, the distinct miRNA expression patterns in PBMCs hold promise as valuable biomarkers for the diagnosis and monitoring of T1D.
The evidence presented highlights the critical role of dysregulated miRNAs in the pathogenesis and progression of T1D. Studies by Takahashi et al. [20], Yang et al. [38], and Massaro et al. [41] collectively demonstrate that specific miRNA expression patterns can distinguish T1D patients, implicating these molecules in key pathways such as immune regulation, insulin signaling, and diabetic complications. Notably, miR-146a/b-5p emerge as central players in the autoimmune imbalance underlying the onset of T1D, with their persistent downregulation linked to immune dysregulation and T-cell response (Table 1). Furthermore, the target genes of miR-146a/b-5p, such as TRAF6, BCL11A, STX3 and NUMB, were upregulated in PBMCs from newly diagnosed T1D patients and those treated with insulin [38]. These target genes are involved in immune regulation and T-cell responses. For instance, TRAF6 regulates TLR signaling to maintain immunological balance [38,70], and NUMB inhibits the Notch signaling pathway, which plays a critical role in T helper cell immune responses [38,74]. STX3 involved in chemokine production by human mast cells [38,75] and BCL11A, related to plasmacytoid dendritic cells differentiation, are also key immune-related genes [38,76]. Additionally, distinct miRNA signatures associated with complications such as neuropathy, nephropathy, and retinopathy further underscore the potential of miRNAs as biomarkers for disease progression and therapeutic targets [44,52,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92]. These findings pave the way for future research into miRNA-targeted therapies aimed at mitigating both the autoimmune aspects of T1D and its associated complications.
Dysregulated miRNAs in T-Cells
T1D is characterized by the loss of functional insulin producing β-cells of pancreatic islets via the destruction of pancreatic β-cells. This phenomenon is caused by islet infiltrating self-reactive CD4+ and CD8+ T-cells [93]. Simultaneously, regulatory T-cells (Tregs) suppress these autoreactive T-cells [94]. Although, the most self-reactive T-cells are eliminated in the thymus via central tolerance induction mechanism, which strengthens immune tolerance to self-antigens under normal condition, a few remaining autoreactive T-cells are released to the peripheral circulation by avoiding this mechanism [94]. This phenomenon implies that these impaired functions of T-cells and Tregs cause the onset of autoimmune T1D through decreased tolerance to islet antigens leading to uncontrolled T-cells mediated autoimmune destruction of pancreatic β-cells [42,94].
miR-191, miR-342, and miR-510 were dysregulated in Tregs isolated from peripheral blood of T1D patients [42]. Specifically, miR-191 was downregulated in Tregs of T1D patients, contrasting with its upregulation in various cancers, including myeloid leukemia, breast cancer, and colorectal cancer [42,95,96,97]. miR-342, abundantly expressed in Tregs of healthy donors, was significantly downregulated in T1D patients [42]. This miRNA was also shown to be downregulated in human leukocytes during lipopolysaccharide-induced inflammation [42,98]. miR-342 targets key molecules such as BMPR2 and PDGFRA in cytokine signaling and EP300 in MAPK and NF-kB signaling pathways [42,98]. These observations suggest that the downregulation of miR-342 in Tregs of T1D patients may impair their functionality, contributing to autoimmune destruction in T1D [42,95,96,97].
Dysregulated miRNAs in Exosomes Derived from Plasma and Urine
Exosomes are lipid-based carriers with potential for disease-specific diagnostics, since they originate from cellular multivesicular bodies that contain specific miRNAs [43,99]. Exosomal miRNAs are tissue-specific and remain stable in plasma, whereas RNAs in blood are prone to degradation by ribonucleases [43,99,100]. miRNAs expression patterns dynamically change throughout diseases progression. Pancreatic β-cells spread exosomes containing miRNAs [101]. Thus, miRNAs derived from exosomes in plasma hold promise as candidates for early detection tools.
Dysregulated exosomal miRNAs play a critical role in T1D pathophysiology, influencing β-cell function [31,43,102], insulin resistance [43,103], and metabolic regulation [43]. Notably, miR-302d-3p, miR-378e, and miR-574-5p are downregulated, while miR-25-3p is upregulated (Table 1). Barutta et al. [44] analyzed urinary exosomal miRNA expression in T1D patients with and without early diabetic nephropathy (DN). Patients with microalbuminuria exhibited increased miR-130a and miR-145 but decreased miR-155 and miR-424 in urinary exosomes compared to non-diabetic controls and normoalbuminuric T1D patients (Table 1). In vivo, miR-145 levels were elevated in both urinary exosomes and glomeruli of STZ-induced diabetic mice compared to controls. Similarly, in vitro, miR-145 expression increased in mesangial cells and mesangial cell-derived exosomes under high-glucose conditions (Table 1) [44]. Urinary exosomal miRNAs, such as miR-145 and miR-155, show a strong association with DN, underscoring their potential as biomarkers for diabetic complications [44].
3. Dysregulated miRNAs in T1D Rodents
As previously noted, understanding tissue-specific miRNAs is essential for deciphering normal tissue development [104] and T1D progression [105]. Table 2 summarizes dysregulated miRNAs in T1D murine models across various studies. These miRNAs are categorized by species, diabetic models, tissue sources, detection methods, and expression patterns, offering valuable insights into their roles in T1D pathogenesis.
Various miRNAs have been implicated in T1D pathogenesis within pancreatic islet-related sources, including pancreatic islets, pancreatic β-cells, the whole pancreas, cultured islets, infiltrating lymphocytes, serum, and blood from T1D rodent models. Among these, 39 miRNAs were upregulated [36,106,107,108,109,113], while 47 were down regulated [36,106,107,108,109,113]. Additionally, miRNAs play a significant role in T1D-related complications. In diabetic retinopathy, for example, 13 miRNAs were upregulated in the retina, while seven were downregulated (Table 2) [77,79,80].
Among the dysregulated miRNAs implicated in T1D pathogenesis, specific miRNAs exhibit more direct roles in disease mechanisms. F[36,106–109,113or example, miR-200b, significantly upregulated in the retina of Ins2Akita mice (Table 2), targets Oxr1 and regulates oxidative stress in diabetic retinopathy [77]. In prediabetic non-obese diabetic (NOD) mice, the miR-29 family affects insulin biosynthesis, while miR-142-3p, miR-142-5p and miR-155 are linked to apoptosis and inflammation [106,107]. Additionally, miR-21, upregulated in β-cells of both NOD and streptozotocin (STZ)-induced T1D mice (Table 2), contributes to islet inflammation and β-cell dysfunction [108]. Other notable findings include the downregulation of miR-409-3p in pancreatic islets and plasma from NOD mice [36], as well as the identification of 64 upregulated and 72 downregulated miRNAs in STZ-induced T1D mice, with key roles in β-cell autoantigen release and apoptosis [109]. Human studies further demonstrate dysregulation of miR-24, miR-29a, miR-148a and miR-200a in newly diagnosed T1D patients, with miR-375 correlating with β-cell injury and miR-21 with islet inflammation [3,31,114]. Notably, overlapping miRNA patterns across diabetes subtypes, including T1D, T2D, latent autoimmune diabetes in adults (LADA), and prediabetes, suggest their potential as biomarkers for distinguishing between different forms of diabetes [3]. These findings underscore the fundamental role of miRNA dysregulation in T1D pathogenesis, influencing β-cell autoantigen release, apoptosis, and immune activation. The identification of tissue-specific and circulating miRNAs provides insights into the molecular mechanisms of T1D and highlights their potential as biomarkers for both disease progression and therapeutic intervention.
4. Dysregulated miRNAs in Diabetic Cardiomyopathy and Nephropathy
The dysregulation of miRNAs in T1D play a significant role in disease pathophysiology and its complications. Diabetes induces vascular and renal dysfunction, contributing to conditions such as diabetic cardiomyopathy (DC) [81] and DN [82], the leading cause of mortality in T1D. Microalbuminuria, the earliest clinical manifestation of DN, has a lifetime incidence of approximately 50% in T1D, with a particularly heightened risk for cardiovascular complications. Although much of the research on diabetes-related DC focuses on T2D, its impact on T1D is even more pronounced. After adjusting for age, the relative risk of cardiovascular complications in T1D remains significantly higher than in T2D [115,116,117].
Recent studies have identified altered miRNA expression patterns involved in the pathophysiology of DC and DN, where these miRNAs regulate key pathways related to cardiomyocyte hypertrophy, oxidative stress, fibrosis, and apoptosis [82,83,84]. In DC, miR-21 [85], miR-34a [86], miR-141 [78], miR-142-3p [87], miR-195 [81], and miR-199a-3p [87] are upregulated, while miR-133a [92], miR-143 [87], miR-150 [88], and miR-499 [92] are downregulated (Figure 2). Similarly, in DN, miR-21 [89], miR-130a [44], and miR-145 [44] are upregulated, while miR-15a-5p [90], miR-25 [91], miR-155 [44], and miR-424 [44] are downregulated (Figure 2). These miRNAs regulate genes and pathways implicated in diabetes-related cardiac and renal dysfunctions, highlighting their potential as diagnostic biomarkers and therapeutic targets for managing T1D complications. Their identification opens new avenues for intervention and improved patient outcomes.
5. Dysregulated miRNAs in T1D and Their Potential Targets
miRNAs play a vital role in maintaining glucose homeostasis in both humans and diabetic animal models by regulating β-cell development, apoptosis, insulin secretion, and insulin action in target tissues. T1D results from the progressive loss of pancreatic islet β-cells, driven by autoimmune activation, β-cell autoantigen release, oxidative and ER stress, and cytokine-mediated damage, ultimately leading to β-cell destruction. This loss disrupts insulin signaling pathways, causing insulin deficiency and hyperglycemia (Figure 1). Table 3 summarizes dysregulated miRNAs and their associated pathways in T1D from various studies.
β-Cell Autoantigen Release
In T1D pathogenesis, the release of β-cell autoantigens plays a pivotal role in triggering and sustaining the autoimmune response. Autoantigens such as insulin, GAD65, IA-2, and ZnT8 are released from stressed or damaged β-cells due to inflammatory insults, oxidative stress, or viral infections. Antigen-presenting cells (APCs), including dendritic cells and macrophages, process these autoantigens and present them to autoreactive T-cells in lymphoid tissues. This activation of autoreactive T-cells drives targeted β-cell destruction, creating a self-perpetuating cycle of immune-mediated damage. The continuous release of autoantigens further amplifies β-cell loss, ultimately leading to insulin deficiency and hyperglycemia.
Growing evidence suggests that dysregulated miRNAs contribute to β-cell autoantigen release and immune activation by regulating key processes such as apoptosis, inflammation, and stress responses. Research indicates significant miRNA dysregulation in T1D patients, particularly in the early stages of the disease. Notably, miR-24, miR-29a, miR-148a, and miR-200a were found to be altered in the serum of newly diagnosed T1D children, implicating their role in β-cell regulatory networks [31,114]. These findings underscore the critical role of miRNAs in modulating β-cell autoantigen release and suggest that targeting specific miRNAs may offer therapeutic strategies to preserve β-cell function and slow T1D progression. Furthermore, understanding miRNA-controlled regulatory networks could facilitate the development of precise diagnostic tools for differentiating diabetes subtypes based on miRNA expression profiles.
Autoimmune Activation
In T1D, autoimmune activation is a key driver of disease onset. Normally, the immune system distinguishes between self and non-self, protecting the body from harmful pathogens. However, in T1D, this system mistakenly targets insulin-producing β-cells in the pancreas as threats, triggering an autoimmune response. T-cells attack and destroy β-cells, progressively reducing insulin production. This immune activation is influenced by genetic predisposition and environmental factors, such as viral infections [5]. Once initiated, the autoimmune cascade leads to sustained β-cell destruction, ultimately resulting in insulin deficiency. As insulin levels decline, blood glucose rises, leading to the hallmark symptoms and complications of T1D.
At a molecular level, several miRNAs have been implicated in this autoimmune process. In recently diagnosed patients (<5 years), miR-21-5p, miR-103a-3p, miR-148b-3p, miR-155-5p, and miR-210-3p are upregulated in plasma, while miR-146a-5p is downregulated [34]. These expression patterns appear specific to the early disease stages, as no significant differences are observed beyond five years post-diagnosis [34]. Among these, miR-155-5p is notably increased in activated B and T-cells, macrophages, and dendritic cells through the NF-κB and JNK pathways, underscoring its critical role in both innate and adaptive immunity [34,140,141,142]. Upregulated miR-155-5p also modulates immune responses by reducing NF-κB activation via IKK downregulation [34,140,142]. In T1D, this regulatory function contributes to the autoimmune destruction of β-cells [34].
Additionally, miR-200 promotes a pro-apoptotic genetic signature in pancreatic islets of diabetic mice by increasing TP53 expression, which suppresses anti-apoptotic and stress-resistance networks in β-cells [34,143]. miR-210-3p, upregulated in the plasma, serum, and urine of pediatric T1D patients, downregulates FOXP3, a key regulator of Treg function, impairing immune tolerance and exacerbating autoimmunity [34,144]. This miRNA also targets genes involved in mitochondrial metabolism, angiogenesis, DNA repair, and cell survival, linking it to various cancers and cardiovascular diseases [34,69,145]. Notably, miR-21, miR-126, and miR-210 are crucial in the pathophysiology of diabetes [29]. In T1D patients, miR-21 and miR-210 levels are significantly elevated in both plasma and urine, whereas miR-126 is reduced in urine but remains unchanged in plasma [35]. Given their involvement in DN, a leading cause of ESRD, these miRNAs hold promise as noninvasive biomarkers for T1D-associated renal dysfunction [35,146,147,148,149,150].
Furthermore, research has shown that miRNA expression patterns in T1D evolve with disease progression, influencing immunological processes in the early stages. For example, miR-10b-5p, miR-17-5p, miR-30e-5p, miR-93-5p, miR-99a-5p, miR-125b-5p, miR-423-3p, and miR-497-5p exhibit significant temporal alterations based on disease duration [151]. Two distinct expression patterns were identified [151]. The first group—miR-17-5p, miR-30e-5p, miR-93-5p, and miR-423-3p—showed reduced expression within the first 12 months post-diagnosis but increased between one and five years. The second group—miR-10b-5p, miR-99a-5p, miR-125b-5p, and miR-497-5p—displayed elevated expression in the first 12 months, followed by a decline over the subsequent five years. Additionally, miR-30e-5p, miR-93-5p, and miR-423-3p maintained consistently higher expression levels throughout the study, whereas miR-10b-5p, miR-17-5p, miR-99a-5p, miR-125b-5p, and miR-497-5p exhibited persistently lower expression relative to the overall sample average at all time points [151]. These findings underscore the dynamic nature of miRNA regulation in T1D and its potential role in disease progression.
Guay et al. [107] demonstrated that rodent and human T lymphocytes release exosomes containing miR-142-5p, which can be transferred to pancreatic β-cells. Suppression of these miRNAs in recipient β-cells blocked exosome-mediated apoptosis and prevented diabetes development in NOD mice, leading to improved insulin levels, reduced insulitis scores, and diminished inflammation. Additionally, exosomes from T lymphocytes induced apoptosis and upregulated Ccl2, Ccl7, and Cxcl10 expression, activating chemokine signaling specifically in β-cells. This process recruited immune cells, potentially exacerbating β-cell destruction during the autoimmune attack [107].
Ventriglia et al. [36] demonstrated that miR-409-3p is significantly downregulated in both recently diabetic NOD mice and newly diagnosed T1D patients. Notably, miR-409-3p expression inversely correlated with insulitis severity and blood glucose levels, reflecting disease progression in NOD mice [36].
To investigate the relationship between miR-409-3p expression and immune system differentiation or metabolic status, including glucose homeostasis, its plasma levels were monitored in recently diabetic NOD mice following anti-CD3 antibody (anti-CD3 Ab) treatment [36]. Anti-CD3 Ab was known to modulate the islet immune response and slow disease progression [152,153]. In anti-CD3-treated NOD mice, plasma miR-409-3p levels were associated with T-cell dynamics and the proinflammatory environment in the islets but not with blood glucose levels [36]. These findings suggest that miR-409-3p plays a role in the primary pathogenesis of T1D by influencing immune system activity.
Taken together, autoimmune activation in T1D is a complex, multifactorial process driven by genetic predisposition and environmental triggers. Key miRNAs, including miR-155-5p, miR-200a-3p, and miR-210-3p, play pivotal roles in the autoimmune destruction of β-cells, underscoring their potential as biomarkers and therapeutic targets for disease management and progression.
Endoplasmic Reticulum and Oxidative Stress
In T1D, endoplasmic reticulum (ER) and oxidative stress are key contributors to the progressive destruction of insulin-producing β-cells. Due to their high insulin production demands and exposure to an inflammatory environment caused by autoimmune attacks, β-cells are particularly vulnerable to these stressors [154]. Oxidative stress in T1D arises from an imbalance between reactive oxygen species (ROS) production and the cell’s antioxidant defenses. Excessive ROS generation, driven by chronic inflammation, cytokine release (e.g., IL-1β, IFN-γ), and immune cell infiltration, leads to cellular damage affecting proteins, lipids, and DNA, ultimately promoting β-cell dysfunction and apoptosis [155]. The transcription factors FOXO and NRF2 play crucial roles in the antioxidant response, but their reduced activity in T1D exacerbates oxidative stress [156].
Several miRNAs regulate ER and oxidative stress mechanisms in T1D. miR-200a-3p targets KEAP1, a negative regulator of NRF2, and its downregulation in T1D impairs NRF2 activation, increasing β-cell susceptibility to oxidative damage [157]. miR-146a modulates inflammatory and stress responses by targeting TRAF6 and IRAK1, key components of the NF-κB signaling pathway [158,159]. Its dysregulation in T1D leads to increased NF-κB activity, exacerbating both ER and oxidative stress and promoting β-cell apoptosis [158]. Additionally, miR-21 influences oxidative stress by targeting PTEN, a negative regulator of the PI3K/Akt pathway. While miR-21 overexpression enhances pro-survival Akt signaling, it also contributes to oxidative stress by inhibiting genes responsible for ROS detoxification. Its dysregulation in T1D is associated with increased oxidative damage and impaired β-cell function [160,161].
These miRNAs and their target genes form critical regulatory networks that modulate ER and oxidative stress responses in β-cells. Disruptions in these pathways accelerate β-cell destruction and T1D progression. Understanding these mechanisms provides potential therapeutic targets for preserving β-cell function and delaying disease onset.
Apoptosis
Apoptosis, a tightly regulated process of programmed cell death, is essential for maintaining cellular homeostasis by eliminating compromised cells. However, in T1D, excessive apoptosis of β-cells is a key driver of disease progression. Cytotoxic T-cells, which typically target pathogens, mistakenly attack β-cells, releasing cytotoxic molecules such as granzyme B and perforin that trigger apoptosis.
Several molecular pathways mediate β-cell apoptosis in T1D. Anti-apoptotic proteins like BCL-2 and MCL-1 help preserve mitochondrial integrity under normal conditions, but the pro-inflammatory environment of T1D—dominated by cytokines such as IL-1β, TNF-α, and IFN-γ—disrupts the balance between pro- and anti-apoptotic signals, tipping the scale toward cell death. This imbalance promotes mitochondrial dysfunction, cytochrome c release, and caspase activation, culminating in β-cell apoptosis.
miRNAs play a crucial role in regulating apoptotic pathways in T1D. miR-15a-5p, miR-16-5p, miR-21-5p, miR-30e-5p, miR-34a, miR-146a influence genes involved in apoptosis [8,90,118,162,163], while miR-100-5p and miR-150-5p impact the PI3K/Akt pathway, which regulates β-cell survival and growth [8]. Chronic inflammation and oxidative stress impair PI3K/Akt signaling, weakening its protective effects on β-cells and increasing their vulnerability to apoptosis. The cyclin-dependent kinase inhibitor P21, which regulates cell cycle progression, can also induce apoptosis under inflammatory conditions. miR-21-5p, miR-100-5p, and miR-375 modulate the Cyclin-CDK complex, influencing β-cell proliferation and survival. Additionally, KLF11 and the TGF-β pathway contribute to β-cell apoptosis, with miRNAs such as miR-10b-5p, miR-21-5p, and miR-424 modulating these pathways [129,164,165,166].
Certain miRNAs are notably upregulated in T1D. For example, miR-24, which is involved in inflammation and TGF-β signaling, is implicated in both T1D and T2D pathogenesis [31,167,168,169]. miR-25, associated with apoptosis regulation and cancer pathology, is elevated in the serum of T1D children and negatively correlated with β-cell function [31,170,171,172,173,174,175]. However, in DN models, miR-25 is downregulated in the kidneys, affecting NADPH expression under hyperglycemia, underscoring its diverse role in pancreatic endocrine cell proliferation and disease pathology [31,176]. Additionally, miR-21-5p and miR-148a, which are elevated in T1D patients, contribute to apoptosis through pathways such as FoxO and TGF-β, with miR-148a specifically targeting BCL2L11 to promote β-cell death [33,177]
miRNAs in T1D also exhibit complex, context-dependent roles. For example, miR-200b promotes apoptosis by downregulating Oxr1 under oxidative stress, whereas its inhibition confers protection against apoptosis [77].The miR-29 family reduces glucose-stimulated insulin secretion and proinsulin mRNA levels but paradoxically mitigates cytokine-induced apoptosis in β-cells [106]. Similarly, miR-21, upregulated in early T1D, promotes apoptosis by suppressing BCL-2 [108]. Collectively, these findings underscore the intricate involvement of miRNAs in T1D pathophysiology, offering potential therapeutic targets for preserving β-cell function and slowing disease progression.
Insulin signaling
The pathogenesis of diabetes is closely linked to impaired insulin secretion, with both T1D and T2D involving disruptions in insulin signaling. Numerous studies have elucidated the molecular mechanisms underlying these impairments, highlighting the roles of insulin and the insulin receptor in disease progression [178].
miR-103a-3p is upregulated in both the liver of T2D patients [71] and the plasma of T1D patients [34]. This miRNA plays a critical role in insulin signaling by targeting CAV1, a key regulator of the insulin receptor in both forms of diabetes [72]. Additionally, miR-103a-3p influences miRNA biosynthesis by regulating Dicer, suggesting that its upregulation could lead to the downregulation of other miRNAs [179]. Interestingly, studies have also linked miR-103a-3p to cellular proliferation and apoptosis in cancer cell lines, indicating its broader role in cellular homeostasis [180].
Ferraz et al. investigated the effects of nuclear and mitochondrial dysfunctions on T1D dysregulation, identifying 41 dysregulated miRNAs in T1D patients (Table 3) [37]. Among these, miR-21-5p had the highest number of target genes, including SOCS and AKT, which are involved in key T1D-related pathways such as insulin signaling and apoptosis [8,34] [37]. Their findings suggest that these 41 miRNAs, particularly miR-21-5p and miR-26b-5p, contribute to nuclear and mitochondrial dysfunctions, ultimately exacerbating T1D dysregulation [37].
Figure 3 illustrates dysregulated miRNAs in T1D that target key proteins, contributing to regulatory T-cell dysfunction, impaired immune tolerance, reduced β-cell proliferation, and heightened apoptosis—ultimately driving autoimmune β-cell destruction. miR-146a-5p, downregulated in T1D [181], normally inhibits STAT1 [182]. Its reduction activates STAT1 in regulatory T-cells, impairing immune tolerance [182]. miR-10b-5p, downregulated in late T1D [151], increases KLF11 expression [129], enhancing TGF-β signaling [183]. However, impaired TGF-β signaling in T1D disrupts regulatory T-cell function [184,185]. miR-210-3p, upregulated in T1D [34], suppresses FOXP3 [144], further contributing to immune dysregulation. miR-216a, crucial for pancreatic β-cells [186], is downregulated in T1D [187]. This reduction increases PTEN, inhibiting β-cell proliferation [187]. miR-21-5p, upregulated in T1D [8,118], suppresses the anti-apoptotic protein BCL2, leading to increased β-cell apoptosis [108]. miR-16-5p enhances β-cell proliferation by inhibiting apoptosis [162]. However, its downregulation in T1D upregulates CXCL10, promoting β-cell death [162]. Collectively, these dysregulated miRNA pathways drive β-cell destruction in T1D.
6. miRNA-Based Therapeutic Strategies for T1D
miRNA-based therapeutics represent an innovative approach to disease treatment by regulating gene expression at the post-transcriptional level. Given that miRNA expression is altered in various diseases, modulating their levels—either by introducing miRNAs or inhibiting their function—offers a promising therapeutic strategy. This concept parallels antisense mRNA and RNA interference (RNAi) techniques. miRNA-based therapeutics primarily follow two strategies: antisense inhibition of mature miRNAs and miRNA replacement. The choice of approach depends on whether the therapeutic goal is to suppress overexpressed miRNAs or restore downregulated miRNAs to regain lost function.
A comprehensive analysis of T1D pathogenesis and its associated complications has identified a range of consistently dysregulated miRNAs. As summarized in Figure 4, miRNA expression patterns in T1D patients and animal models show considerable overlap. In T1D patients, 138 miRNAs are upregulated, while 93 are downregulated. Similarly, T1D animal models exhibit 66 upregulated and 63 downregulated miRNAs. Notably, miR-21, miR-29a, miR-142-5p, miR-145, miR-146a, miR-186, miR-195, miR-199a-3p, miR-208, and miR-323-5p are consistently upregulated in both T1D patients and murine models. Conversely, miR-10b-5p, miR-16-5p, miR-17-5p, miR-23a, miR-126-5p, miR-143-3p, miR-150, miR-155, and miR-409-3p are consistently downregulated in both populations.
Additionally, miR-106b-5p and miR-222-3p have emerged as potential therapeutic targets due to their roles in pancreatic β-cell function. Tsukita et al. [113] found that bone marrow transplantation (BMT) restored pancreatic islets in STZ-induced diabetic mice while increasing miR-106b and miR-222 levels in serum exosomes and islets. Exosomal miRNA analysis showed elevated miR-106b-5p and miR-222-3p in the culture media of bone marrow cells from STZ-BMT mice. Notably, administering miR-106b-5p and miR-222-3p mimics enhanced β-cell proliferation and improved hyperglycemia by downregulating the Cip/Kip family, promoting β-cell regeneration [113]. A miR-216a mimic nanodrug has been also shown to enhance β-cell proliferation via PTEN inhibition, leading to increased insulin production [187]. Another study identified miR-142-3p as a key regulator of islet autoimmunity in NOD mice. Inhibiting miR-142-3p with an LNA-miRNA inhibitor enhanced regulatory T-cell stability by targeting TET2, reducing islet autoimmunity in diabetic mice [188].
Furthermore, miR-21 [33,45,46,47] and miR-146a [38,70,74,75,76,158,159]—both implicated in inflammatory and autoimmune responses—have emerged as potential therapeutic targets. Modulating these dysregulated miRNAs through inhibitors or mimics (Figure 4) may help reduce immune-mediated β-cell destruction and inflammation in T1D. Targeting these pathways could slow disease progression and potentially prevent its onset. As research advances, miRNA-based therapies offer a novel approach to preserving β-cell function and improving T1D outcomes.
7. Conclusion and Future Study
T1D results from the loss of pancreatic β-cells through autoimmune or idiopathic processes, yet the precise molecular mechanisms driving β-cell destruction remain incompletely understood. Furthermore, no therapeutics have been developed to effectively reverse disease progression in T1D.
miRNA-based therapeutics offer a unique advantage, as a single miRNA can regulate the expression of multiple genes, ranging from tens to hundreds. These therapies operate through two primary strategies: miRNA mimics, which restore deficient miRNAs to recover protein synthesis, and miRNA inhibitors, which suppress overactive miRNAs to reduce pathological gene expression. Additionally, because miRNAs are endogenous molecules, they exhibit low immunogenicity, reducing the likelihood of immune rejection.
Although no miRNA-based therapies have received FDA approval to date, numerous clinical trials, as reviewed by Singh et al. [24] are exploring their therapeutic potential. These studies demonstrate the broad applicability of miRNA therapeutics in various diseases, including diabetes and autoimmune disorders. This review highlights key preclinical studies investigating miRNA-based therapies in T1D, detailing their therapeutic efficacy, dysregulated miRNAs in T1D patients and murine models, and their associated pathways and target genes.
Despite their promise, miRNA therapeutics face several challenges. First, miRNA mimics and inhibitors are highly susceptible to degradation in circulation, limiting their stability and therapeutic efficacy. Second, intracellular delivery remains inefficient, as many miRNAs become sequestered in endosomes rather than reaching their target sites in the cytoplasm. Developing advanced delivery systems and enhancing endosomal escape mechanisms are crucial for improving gene silencing efficiency. Lastly, miRNAs can induce off-target effects by regulating multiple genes across different pathways and cell types, potentially leading to unintended gene silencing. Strategies such as sequence optimization, chemical modifications, and targeted delivery approaches can enhance specificity and minimize off-target effects, thereby improving the safety and efficacy of miRNA-based therapies.
Although current miRNA-based therapeutics face obstacles, rapidly advancing preclinical and clinical research, along with interdisciplinary innovations, may soon overcome these challenges. Given their remarkable therapeutic potential, miRNA-based treatments hold promise not only for T1D but for a broad spectrum of diseases.
Author Contributions
Writing—original draft preparation, H.C. and S.E.H.; writing—review and editing, D.K, S.E.H., R.S. and S.R.; funding acquisition, S.R. All authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The figures were created using BioRender.
Abbreviations
T1D, type 1 diabetes; T2D, type 2 diabetes; GD, gestational diabetes; ER, endoplasmic reticulum; AAbs, autoantibodies; HLA, human leukocyte antigen; AGEs, advanced glycation end products; miRNAs, microRNA; mRNAs, messenger RNAs; PBMCs, peripheral blood mononuclear cells; Treg, regulatory T-cell; NOD mice, non-obese diabetic mice; STZ, Streptozotocin; LADA, latent autoimmune diabetes in adults; DC, diabetic cardiomyopathy; DN, diabetic nephropathy; APCs, Antigen-presenting cells; ROS, reactive oxygen species; BMT, bone marrow transplantation.
References
- Banday, M.Z.; Sameer, A.S.; Nissar, S. , Pathophysiology of diabetes: An overview. Avicenna J Med 2020, 10, 174–188. [Google Scholar] [CrossRef] [PubMed]
- Tuomi, T.; Santoro, N.; Caprio, S.; Cai, M.; Weng, J.; Groop, L. , The many faces of diabetes: a disease with increasing heterogeneity. Lancet 2014, 383, 1084–94. [Google Scholar] [CrossRef] [PubMed]
- Seyhan, A.A.; Nunez Lopez, Y.O.; Xie, H.; Yi, F.; Mathews, C.; Pasarica, M.; Pratley, R.E. , Pancreas-enriched miRNAs are altered in the circulation of subjects with diabetes: a pilot cross-sectional study. Sci Rep 2016, 6, 31479. [Google Scholar] [CrossRef]
- Gregory, G.A.; Robinson, T.I.G.; Linklater, S.E.; Wang, F.; Colagiuri, S.; de Beaufort, C.; Donaghue, K.C.; International Diabetes Federation Diabetes Atlas Type 1 Diabetes in Adults Special Interest, G. ; Magliano, D.J.; Maniam, J.; Orchard, T.J.; Rai, P.; Ogle, G.D., Global incidence, prevalence, and mortality of type 1 diabetes in 2021 with projection to 2040: a modelling study. Lancet Diabetes Endocrinol 2022, 10, 741–760. [Google Scholar] [CrossRef]
- Katsarou, A.; Gudbjornsdottir, S.; Rawshani, A.; Dabelea, D.; Bonifacio, E.; Anderson, B.J.; Jacobsen, L.M.; Schatz, D.A.; Lernmark, A. , Type 1 diabetes mellitus. Nat Rev Dis Primers 2017, 3, 17016. [Google Scholar] [CrossRef] [PubMed]
- Daneman, D. , Type 1 diabetes. Lancet 2006, 367, 847–58. [Google Scholar] [CrossRef]
- Margaritis, K.; Margioula-Siarkou, G.; Giza, S.; Kotanidou, E.P.; Tsinopoulou, V.R.; Christoforidis, A.; Galli-Tsinopoulou, A. , Micro-RNA Implications in Type-1 Diabetes Mellitus: A Review of Literature. Int J Mol Sci, 2021; 22. [Google Scholar]
- Assmann, T.S.; Recamonde-Mendoza, M.; De Souza, B.M.; Crispim, D. , MicroRNA expression profiles and type 1 diabetes mellitus: systematic review and bioinformatic analysis. Endocr Connect 2017, 6, 773–790. [Google Scholar] [CrossRef]
- Pirot, P.; Cardozo, A.K.; Eizirik, D.L. , Mediators and mechanisms of pancreatic beta-cell death in type 1 diabetes. Arq Bras Endocrinol Metabol 2008, 52, 156–65. [Google Scholar] [CrossRef]
- Ounissi-Benkalha, H.; Polychronakos, C. , The molecular genetics of type 1 diabetes: new genes and emerging mechanisms. Trends Mol Med 2008, 14, 268–75. [Google Scholar] [CrossRef]
- Ziegler, A.G.; Rewers, M.; Simell, O.; Simell, T.; Lempainen, J.; Steck, A.; Winkler, C.; Ilonen, J.; Veijola, R.; Knip, M.; Bonifacio, E.; Eisenbarth, G.S. , Seroconversion to multiple islet autoantibodies and risk of progression to diabetes in children. JAMA 2013, 309, 2473–9. [Google Scholar] [CrossRef]
- Guay, C.; Regazzi, R. , Circulating microRNAs as novel biomarkers for diabetes mellitus. Nat Rev Endocrinol 2013, 9, 513–21. [Google Scholar] [CrossRef] [PubMed]
- von Scholten, B.J.; Kreiner, F.F.; Gough, S.C.L.; von Herrath, M. , Current and future therapies for type 1 diabetes. Diabetologia 2021, 64, 1037–1048. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, R.; Putta, S.; Kato, M. , MicroRNAs and diabetic complications. J Cardiovasc Transl Res 2012, 5, 413–22. [Google Scholar] [CrossRef]
- He, Z.; King, G.L. , Microvascular complications of diabetes. Endocrinol Metab Clin North Am 2004, 33, 215–38. [Google Scholar] [CrossRef]
- Beckman, J.A.; Creager, M.A.; Libby, P. , Diabetes and atherosclerosis: epidemiology, pathophysiology, and management. JAMA 2002, 287, 2570–81. [Google Scholar] [CrossRef]
- Esteller, M. , Non-coding RNAs in human disease. Nat Rev Genet 2011, 12, 861–74. [Google Scholar] [CrossRef] [PubMed]
- Satake, E.; Pezzolesi, M.G.; Md Dom, Z.I.; Smiles, A.M.; Niewczas, M.A.; Krolewski, A.S. , Circulating miRNA Profiles Associated With Hyperglycemia in Patients With Type 1 Diabetes. Diabetes 2018, 67, 1013–1023. [Google Scholar] [CrossRef]
- Erener, S.; Marwaha, A.; Tan, R.; Panagiotopoulos, C.; Kieffer, T.J. , Profiling of circulating microRNAs in children with recent onset of type 1 diabetes. JCI Insight 2017, 2, e89656. [Google Scholar] [CrossRef]
- Takahashi, P.; Xavier, D.J.; Evangelista, A.F.; Manoel-Caetano, F.S.; Macedo, C.; Collares, C.V.; Foss-Freitas, M.C.; Foss, M.C.; Rassi, D.M.; Donadi, E.A.; Passos, G.A.; Sakamoto-Hojo, E.T. , MicroRNA expression profiling and functional annotation analysis of their targets in patients with type 1 diabetes mellitus. Gene 2014, 539, 213–23. [Google Scholar] [CrossRef]
- Rupaimoole, R.; Slack, F.J. , MicroRNA therapeutics: towards a new era for the management of cancer and other diseases. Nat Rev Drug Discov 2017, 16, 203–222. [Google Scholar] [CrossRef]
- Lu, Q.; Wu, R.; Zhao, M.; Garcia-Gomez, A.; Ballestar, E. , miRNAs as Therapeutic Targets in Inflammatory Disease. Trends Pharmacol Sci 2019, 40, 853–865. [Google Scholar] [CrossRef]
- Singh, R.; Ha, S.E.; Park, H.S.; Debnath, S.; Cho, H.; Baek, G.; Yu, T.Y.; Ro, S. , Sustained Effectiveness and Safety of Therapeutic miR-10a/b in Alleviating Diabetes and Gastrointestinal Dysmotility without Inducing Cancer or Inflammation in Murine Liver and Colon. Int J Mol Sci, 2024; 25. [Google Scholar]
- Zogg, H.; Singh, R.; Ro, S. , Current Advances in RNA Therapeutics for Human Diseases. Int J Mol Sci, 2022; 23. [Google Scholar]
- Niehrs, C.; Pollet, N. , Synexpression groups in eukaryotes. Nature 1999, 402, 483–7. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Maki, M.; Ding, R.; Yang, Y.; Zhang, B.; Xiong, L. , Genome-wide survey of tissue-specific microRNA and transcription factor regulatory networks in 12 tissues. Sci Rep 2014, 4, 5150. [Google Scholar] [CrossRef]
- Ambros, V. , The functions of animal microRNAs. Nature 2004, 431, 350–5. [Google Scholar] [CrossRef] [PubMed]
- Landgraf, P.; Rusu, M.; Sheridan, R.; Sewer, A.; Iovino, N.; Aravin, A.; Pfeffer, S.; Rice, A.; Kamphorst, A.O.; Landthaler, M.; Lin, C.; Socci, N.D.; Hermida, L.; Fulci, V.; Chiaretti, S.; Foa, R.; Schliwka, J.; Fuchs, U.; Novosel, A.; Muller, R.U.; Schermer, B.; Bissels, U.; Inman, J.; Phan, Q.; Chien, M.; Weir, D.B.; Choksi, R.; De Vita, G.; Frezzetti, D.; Trompeter, H.I.; Hornung, V.; Teng, G.; Hartmann, G.; Palkovits, M.; Di Lauro, R.; Wernet, P.; Macino, G.; Rogler, C.E.; Nagle, J.W.; Ju, J.; Papavasiliou, F.N.; Benzing, T.; Lichter, P.; Tam, W.; Brownstein, M.J.; Bosio, A.; Borkhardt, A.; Russo, J.J.; Sander, C.; Zavolan, M.; Tuschl, T. , A mammalian microRNA expression atlas based on small RNA library sequencing. Cell 2007, 129, 1401–14. [Google Scholar] [CrossRef] [PubMed]
- Sayed, A.S.; Xia, K.; Salma, U.; Yang, T.; Peng, J. , Diagnosis, prognosis and therapeutic role of circulating miRNAs in cardiovascular diseases. Heart Lung Circ 2014, 23, 503–10. [Google Scholar] [CrossRef]
- Zampetaki, A.; Willeit, P.; Drozdov, I.; Kiechl, S.; Mayr, M. , Profiling of circulating microRNAs: from single biomarkers to re-wired networks. Cardiovasc Res 2012, 93, 555–62. [Google Scholar] [CrossRef]
- Nielsen, L.B.; Wang, C.; Sorensen, K.; Bang-Berthelsen, C.H.; Hansen, L.; Andersen, M.L.; Hougaard, P.; Juul, A.; Zhang, C.Y.; Pociot, F.; Mortensen, H.B. , Circulating levels of microRNA from children with newly diagnosed type 1 diabetes and healthy controls: evidence that miR-25 associates to residual beta-cell function and glycaemic control during disease progression. Exp Diabetes Res 2012, 2012, 896362. [Google Scholar]
- Santos, A.S.; Ferreira, L.R.P.; da Silva, A.C.; Alves, L.I.; Damasceno, J.G.; Kulikowski, L.; Cunha-Neto, E.; da Silva, M.E.R. , Progression of Type 1 Diabetes: Circulating MicroRNA Expression Profiles Changes from Preclinical to Overt Disease. J Immunol Res 2022, 2022, 2734490. [Google Scholar] [CrossRef]
- Grieco, G.E.; Cataldo, D.; Ceccarelli, E.; Nigi, L.; Catalano, G.; Brusco, N.; Mancarella, F.; Ventriglia, G.; Fondelli, C.; Guarino, E.; Crisci, I.; Sebastiani, G.; Dotta, F. , Serum Levels of miR-148a and miR-21-5p Are Increased in Type 1 Diabetic Patients and Correlated with Markers of Bone Strength and Metabolism. Noncoding RNA, 2018; 4. [Google Scholar]
- Assmann, T.S.; Recamonde-Mendoza, M.; Punales, M.; Tschiedel, B.; Canani, L.H.; Crispim, D. , MicroRNA expression profile in plasma from type 1 diabetic patients: Case-control study and bioinformatic analysis. Diabetes Res Clin Pract 2018, 141, 35–46. [Google Scholar] [CrossRef]
- Osipova, J.; Fischer, D.C.; Dangwal, S.; Volkmann, I.; Widera, C.; Schwarz, K.; Lorenzen, J.M.; Schreiver, C.; Jacoby, U.; Heimhalt, M.; Thum, T.; Haffner, D. , Diabetes-associated microRNAs in pediatric patients with type 1 diabetes mellitus: a cross-sectional cohort study. J Clin Endocrinol Metab 2014, 99, E1661–E1665. [Google Scholar] [CrossRef] [PubMed]
- Ventriglia, G.; Mancarella, F.; Sebastiani, G.; Cook, D.P.; Mallone, R.; Mathieu, C.; Gysemans, C.; Dotta, F. , miR-409-3p is reduced in plasma and islet immune infiltrates of NOD diabetic mice and is differentially expressed in people with type 1 diabetes. Diabetologia 2020, 63, 124–136. [Google Scholar] [CrossRef]
- Ferraz, R.S.; Santos, L.C.B.; da-Silva-Cruz, R.L.; Braga-da-Silva, C.H.; Magalhaes, L.; Ribeiro-Dos-Santos, A.; Vidal, A.; Vinasco-Sandoval, T.; Reis-das-Merces, L.; Sena-Dos-Santos, C.; Pereira, A.L.; Silva, L.S.D.; de Melo, F.T.C.; de Souza, A.; Leal, V.S.G.; de Figueiredo, P.B.B.; Neto, J.F.A.; de Moraes, L.V.; de Lemos, G.N.; de Queiroz, N.N.M.; Felicio, K.M.; Cavalcante, G.C.; Ribeiro-Dos-Santos, A.; Felicio, J.S. , Global miRNA expression reveals novel nuclear and mitochondrial interactions in Type 1 diabetes mellitus. Front Endocrinol (Lausanne) 2022, 13, 1033809. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Ye, L.; Wang, B.; Gao, J.; Liu, R.; Hong, J.; Wang, W.; Gu, W.; Ning, G. , Decreased miR-146 expression in peripheral blood mononuclear cells is correlated with ongoing islet autoimmunity in type 1 diabetes patients 1miR-146. J Diabetes 2015, 7, 158–65. [Google Scholar] [CrossRef]
- Estrella, S.; Garcia-Diaz, D.F.; Codner, E.; Camacho-Guillen, P.; Perez-Bravo, F. , Expression of miR-22 and miR-150 in type 1 diabetes mellitus: Possible relationship with autoimmunity and clinical characteristics. Med Clin (Barc) 2016, 147, 245–7. [Google Scholar] [CrossRef]
- Wang, G.; Gu, Y.; Xu, N.; Zhang, M.; Yang, T. , Decreased expression of miR-150, miR146a and miR424 in type 1 diabetic patients: Association with ongoing islet autoimmunity. Biochem Biophys Res Commun 2018, 498, 382–387. [Google Scholar] [CrossRef] [PubMed]
- Massaro, J.D.; Polli, C.D.; Costa, E.S.M.; Alves, C.C.; Passos, G.A.; Sakamoto-Hojo, E.T.; Rodrigues de Holanda Miranda, W.; Bispo Cezar, N.J.; Rassi, D.M.; Crispim, F.; Dib, S.A.; Foss-Freitas, M.C.; Pinheiro, D.G.; Donadi, E.A. , Post-transcriptional markers associated with clinical complications in Type 1 and Type 2 diabetes mellitus. Mol Cell Endocrinol 2019, 490, 1–14. [Google Scholar] [CrossRef]
- Hezova, R.; Slaby, O.; Faltejskova, P.; Mikulkova, Z.; Buresova, I.; Raja, K.R.; Hodek, J.; Ovesna, J.; Michalek, J. , microRNA-342, microRNA-191 and microRNA-510 are differentially expressed in T regulatory cells of type 1 diabetic patients. Cell Immunol 2010, 260, 70–4. [Google Scholar] [CrossRef]
- Garcia-Contreras, M.; Shah, S.H.; Tamayo, A.; Robbins, P.D.; Golberg, R.B.; Mendez, A.J.; Ricordi, C. , Plasma-derived exosome characterization reveals a distinct microRNA signature in long duration Type 1 diabetes. Sci Rep 2017, 7, 5998. [Google Scholar] [CrossRef]
- Barutta, F.; Tricarico, M.; Corbelli, A.; Annaratone, L.; Pinach, S.; Grimaldi, S.; Bruno, G.; Cimino, D.; Taverna, D.; Deregibus, M.C.; Rastaldi, M.P.; Perin, P.C.; Gruden, G. , Urinary exosomal microRNAs in incipient diabetic nephropathy. PLoS One 2013, 8, e73798. [Google Scholar] [CrossRef]
- Wang, S.; Liu, Z.; Wang, J.; Ji, X.; Yao, Z.; Wang, X. , miR-21 promotes osteoclastogenesis through activation of PI3K/Akt signaling by targeting Pten in RAW264.7 cells. Mol Med Rep 2020, 21, 1125–1132. [Google Scholar] [CrossRef] [PubMed]
- Aghaei-Zarch, S.M. , Crosstalk between MiRNAs/lncRNAs and PI3K/AKT signaling pathway in diabetes mellitus: Mechanistic and therapeutic perspectives. Noncoding RNA Res 2024, 9, 486–507. [Google Scholar] [CrossRef]
- Melkman-Zehavi, T.; Oren, R.; Kredo-Russo, S.; Shapira, T.; Mandelbaum, A.D.; Rivkin, N.; Nir, T.; Lennox, K.A.; Behlke, M.A.; Dor, Y.; Hornstein, E. , miRNAs control insulin content in pancreatic beta-cells via downregulation of transcriptional repressors. EMBO J 2011, 30, 835–45. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Thielen, L.A.; Chen, J.; Grayson, T.B.; Grimes, T.; Bridges, S.L., Jr.; Tse, H.M.; Smith, B.; Patel, R.; Li, P.; Evans-Molina, C.; Ovalle, F.; Shalev, A. , Serum miR-204 is an early biomarker of type 1 diabetes-associated pancreatic beta-cell loss. Am J Physiol Endocrinol Metab 2019, 317, E723–E730. [Google Scholar] [CrossRef] [PubMed]
- Angelescu, M.A.; Andronic, O.; Dima, S.O.; Popescu, I.; Meivar-Levy, I.; Ferber, S.; Lixandru, D. , miRNAs as Biomarkers in Diabetes: Moving towards Precision Medicine. Int J Mol Sci, 2022; 23. [Google Scholar]
- Zi, Y.; Zhang, Y.; Wu, Y.; Zhang, L.; Yang, R.; Huang, Y. , Downregulation of microRNA-25-3p inhibits the proliferation and promotes the apoptosis of multiple myeloma cells via targeting the PTEN/PI3K/AKT signaling pathway. Int J Mol Med, 2021; 47. [Google Scholar]
- Kong, R.; Gao, J.; Ji, L.; Zhao, D. , MicroRNA-126 promotes proliferation, migration, invasion and endothelial differentiation while inhibits apoptosis and osteogenic differentiation of bone marrow-derived mesenchymal stem cells. Cell Cycle 2020, 19, 2119–2138. [Google Scholar] [CrossRef]
- Fang, S.; Ma, X.; Guo, S.; Lu, J. , MicroRNA-126 inhibits cell viability and invasion in a diabetic retinopathy model via targeting IRS-1. Oncol Lett 2017, 14, 4311–4318. [Google Scholar] [CrossRef]
- Rosell, R.; Wei, J.; Taron, M. , Circulating MicroRNA Signatures of Tumor-Derived Exosomes for Early Diagnosis of Non-Small-Cell Lung Cancer. Clin Lung Cancer 2009, 10, 8–9. [Google Scholar] [CrossRef]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O'Briant, K.C.; Allen, A.; Lin, D.W.; Urban, N.; Drescher, C.W.; Knudsen, B.S.; Stirewalt, D.L.; Gentleman, R.; Vessella, R.L.; Nelson, P.S.; Martin, D.B.; Tewari, M. , Circulating microRNAs as stable blood-based markers for cancer detection. Proc Natl Acad Sci U S A 2008, 105, 10513–8. [Google Scholar] [CrossRef]
- Cortez, M.A.; Calin, G.A. , MicroRNA identification in plasma and serum: a new tool to diagnose and monitor diseases. Expert Opin Biol Ther 2009, 9, 703–711. [Google Scholar] [CrossRef]
- Gilad, S.; Meiri, E.; Yogev, Y.; Benjamin, S.; Lebanony, D.; Yerushalmi, N.; Benjamin, H.; Kushnir, M.; Cholakh, H.; Melamed, N.; Bentwich, Z.; Hod, M.; Goren, Y.; Chajut, A. , Serum microRNAs are promising novel biomarkers. PLoS One 2008, 3, e3148. [Google Scholar] [CrossRef]
- Wang, C.; Hu, J.; Lu, M.; Gu, H.; Zhou, X.; Chen, X.; Zen, K.; Zhang, C.Y.; Zhang, T.; Ge, J.; Wang, J.; Zhang, C. , A panel of five serum miRNAs as a potential diagnostic tool for early-stage renal cell carcinoma. Sci Rep 2015, 5, 7610. [Google Scholar] [CrossRef] [PubMed]
- Gui, J.; Tian, Y.; Wen, X.; Zhang, W.; Zhang, P.; Gao, J.; Run, W.; Tian, L.; Jia, X.; Gao, Y. , Serum microRNA characterization identifies miR-885-5p as a potential marker for detecting liver pathologies. Clin Sci (Lond) 2011, 120, 183–93. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Chen, H.; Si, H.; Li, X.; Ding, X.; Sheng, Q.; Chen, P.; Zhang, H. , Serum miR-23a, a potential biomarker for diagnosis of pre-diabetes and type 2 diabetes. Acta Diabetol 2014, 51, 823–31. [Google Scholar] [CrossRef]
- Zhou, S.S.; Jin, J.P.; Wang, J.Q.; Zhang, Z.G.; Freedman, J.H.; Zheng, Y.; Cai, L. , miRNAS in cardiovascular diseases: potential biomarkers, therapeutic targets and challenges. Acta Pharmacol Sin 2018, 39, 1073–1084. [Google Scholar] [CrossRef]
- Olivieri, F.; Rippo, M.R.; Procopio, A.D.; Fazioli, F. , Circulating inflamma-miRs in aging and age-related diseases. Front Genet 2013, 4, 121. [Google Scholar] [CrossRef]
- Du, M.; Liu, S.; Gu, D.; Wang, Q.; Zhu, L.; Kang, M.; Shi, D.; Chu, H.; Tong, N.; Chen, J.; Adams, T.S.; Zhang, Z.; Wang, M. , Clinical potential role of circulating microRNAs in early diagnosis of colorectal cancer patients. Carcinogenesis 2014, 35, 2723–30. [Google Scholar] [CrossRef]
- Wang, W.; Sun, G.; Zhang, L.; Shi, L.; Zeng, Y. , Circulating microRNAs as novel potential biomarkers for early diagnosis of acute stroke in humans. J Stroke Cerebrovasc Dis 2014, 23, 2607–2613. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.K.; Zhu, J.Q.; Zhang, J.T.; Li, Q.; Li, Y.; He, J.; Qin, Y.W.; Jing, Q. , Circulating microRNA: a novel potential biomarker for early diagnosis of acute myocardial infarction in humans. Eur Heart J 2010, 31, 659–66. [Google Scholar] [CrossRef]
- Sohel, M.M.H. , Circulating microRNAs as biomarkers in cancer diagnosis. Life Sci 2020, 248, 117473. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, Y.; Zhao, Y.; Wang, Y.; Ding, H.; Xue, S.; Li, P. , Circulating miRNAs as biomarkers for early diagnosis of coronary artery disease. Expert Opin Ther Pat 2018, 28, 591–601. [Google Scholar] [CrossRef]
- de Gonzalo-Calvo, D.; Vea, A.; Bar, C.; Fiedler, J.; Couch, L.S.; Brotons, C.; Llorente-Cortes, V.; Thum, T. , Circulating non-coding RNAs in biomarker-guided cardiovascular therapy: a novel tool for personalized medicine? Eur Heart J 2019, 40, 1643–1650. [Google Scholar] [CrossRef]
- Gao, W.; Liu, L.; Lu, X.; Shu, Y. , Circulating microRNAs: possible prediction biomarkers for personalized therapy of non-small-cell lung carcinoma. Clin Lung Cancer 2011, 12, 14–7. [Google Scholar] [CrossRef]
- Lu, J.; Xie, F.; Geng, L.; Shen, W.; Sui, C.; Yang, J. , Potential Role of MicroRNA-210 as Biomarker in Human Cancers Detection: A Meta-Analysis. Biomed Res Int 2015, 2015, 303987. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Luo, X.; Cui, H.; Ni, X.; Yuan, M.; Guo, Y.; Huang, X.; Zhou, H.; de Vries, N.; Tak, P.P.; Chen, S.; Shen, N. , MicroRNA-146A contributes to abnormal activation of the type I interferon pathway in human lupus by targeting the key signaling proteins. Arthritis Rheum 2009, 60, 1065–75. [Google Scholar] [CrossRef]
- Zhu, H.; Leung, S.W. , Identification of microRNA biomarkers in type 2 diabetes: a meta-analysis of controlled profiling studies. Diabetologia 2015, 58, 900–11. [Google Scholar] [CrossRef]
- Trajkovski, M.; Hausser, J.; Soutschek, J.; Bhat, B.; Akin, A.; Zavolan, M.; Heim, M.H.; Stoffel, M. , MicroRNAs 103 and 107 regulate insulin sensitivity. Nature 2011, 474, 649–53. [Google Scholar] [CrossRef] [PubMed]
- Liew, C.C.; Ma, J.; Tang, H.C.; Zheng, R.; Dempsey, A.A. , The peripheral blood transcriptome dynamically reflects system wide biology: a potential diagnostic tool. J Lab Clin Med 2006, 147, 126–32. [Google Scholar] [CrossRef]
- Luty, W.H.; Rodeberg, D.; Parness, J.; Vyas, Y.M. , Antiparallel segregation of notch components in the immunological synapse directs reciprocal signaling in allogeneic Th:DC conjugates. J Immunol 2007, 179, 819–29. [Google Scholar] [CrossRef] [PubMed]
- Pattu, V.; Qu, B.; Schwarz, E.C.; Strauss, B.; Weins, L.; Bhat, S.S.; Halimani, M.; Marshall, M.; Rettig, J.; Hoth, M. , SNARE protein expression and localization in human cytotoxic T lymphocytes. Eur J Immunol 2012, 42, 470–5. [Google Scholar] [CrossRef]
- Pelayo, R.; Hirose, J.; Huang, J.; Garrett, K.P.; Delogu, A.; Busslinger, M.; Kincade, P.W. , Derivation of 2 categories of plasmacytoid dendritic cells in murine bone marrow. Blood 2005, 105, 4407–15. [Google Scholar] [CrossRef]
- Murray, A.R.; Chen, Q.; Takahashi, Y.; Zhou, K.K.; Park, K.; Ma, J.X. , MicroRNA-200b downregulates oxidation resistance 1 (Oxr1) expression in the retina of type 1 diabetes model. Invest Ophthalmol Vis Sci 2013, 54, 1689–97. [Google Scholar] [CrossRef] [PubMed]
- Baseler, W.A.; Thapa, D.; Jagannathan, R.; Dabkowski, E.R.; Croston, T.L.; Hollander, J.M. , miR-141 as a regulator of the mitochondrial phosphate carrier (Slc25a3) in the type 1 diabetic heart. Am J Physiol Cell Physiol 2012, 303, C1244–C1251. [Google Scholar] [CrossRef]
- Silva, V.A.; Polesskaya, A.; Sousa, T.A.; Correa, V.M.; Andre, N.D.; Reis, R.I.; Kettelhut, I.C.; Harel-Bellan, A.; De Lucca, F.L. , Expression and cellular localization of microRNA-29b and RAX, an activator of the RNA-dependent protein kinase (PKR), in the retina of streptozotocin-induced diabetic rats. Mol Vis 2011, 17, 2228–40. [Google Scholar] [PubMed]
- Xiong, F.; Du, X.; Hu, J.; Li, T.; Du, S.; Wu, Q. , Altered retinal microRNA expression profiles in early diabetic retinopathy: an in silico analysis. Curr Eye Res 2014, 39, 720–9. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Ma, J.; Yu, Y.; Li, M.; Ni, R.; Wang, G.; Chen, R.; Li, J.; Fan, G.C.; Lacefield, J.C.; Peng, T. , Silencing of miR-195 reduces diabetic cardiomyopathy in C57BL/6 mice. Diabetologia 2015, 58, 1949–58. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Yao, D.; Yan, H.; Chen, X.; Wang, L.; Zhan, H. , The Role of MicroRNAs in the Pathogenesis of Diabetic Nephropathy. Int J Endocrinol 2019, 2019, 8719060. [Google Scholar] [CrossRef]
- Deshpande, S.; Abdollahi, M.; Wang, M.; Lanting, L.; Kato, M.; Natarajan, R. , Reduced Autophagy by a microRNA-mediated Signaling Cascade in Diabetes-induced Renal Glomerular Hypertrophy. Sci Rep 2018, 8, 6954. [Google Scholar] [CrossRef]
- Guo, R.; Nair, S. , Role of microRNA in diabetic cardiomyopathy: From mechanism to intervention. Biochim Biophys Acta Mol Basis Dis 2017, 1863, 2070–2077. [Google Scholar] [CrossRef]
- Thum, T.; Gross, C.; Fiedler, J.; Fischer, T.; Kissler, S.; Bussen, M.; Galuppo, P.; Just, S.; Rottbauer, W.; Frantz, S.; Castoldi, M.; Soutschek, J.; Koteliansky, V.; Rosenwald, A.; Basson, M.A.; Licht, J.D.; Pena, J.T.; Rouhanifard, S.H.; Muckenthaler, M.U.; Tuschl, T.; Martin, G.R.; Bauersachs, J.; Engelhardt, S. , MicroRNA-21 contributes to myocardial disease by stimulating MAP kinase signalling in fibroblasts. Nature 2008, 456, 980–4. [Google Scholar] [CrossRef]
- Fomison-Nurse, I.; Saw, E.E.L.; Gandhi, S.; Munasinghe, P.E.; Van Hout, I.; Williams, M.J.A.; Galvin, I.; Bunton, R.; Davis, P.; Cameron, V.; Katare, R. , Diabetes induces the activation of pro-ageing miR-34a in the heart, but has differential effects on cardiomyocytes and cardiac progenitor cells. Cell Death Differ 2018, 25, 1336–1349. [Google Scholar] [CrossRef]
- Shen, E.; Diao, X.; Wang, X.; Chen, R.; Hu, B. , MicroRNAs involved in the mitogen-activated protein kinase cascades pathway during glucose-induced cardiomyocyte hypertrophy. The American journal of pathology 2011, 179, 639–50. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Zhou, B.; Su, H.; Liu, Y.; Du, C. , miR-150 regulates high glucose-induced cardiomyocyte hypertrophy by targeting the transcriptional co-activator p300. Experimental cell research 2013, 319, 173–84. [Google Scholar] [CrossRef] [PubMed]
- Dhas, Y.; Arshad, N.; Biswas, N.; Jones, L.D.; Ashili, S. , MicroRNA-21 Silencing in Diabetic Nephropathy: Insights on Therapeutic Strategies. Biomedicines, 2023; 11. [Google Scholar]
- Dieter, C.; Assmann, T.S.; Costa, A.R.; Canani, L.H.; de Souza, B.M.; Bauer, A.C.; Crispim, D. , MiR-30e-5p and MiR-15a-5p Expressions in Plasma and Urine of Type 1 Diabetic Patients With Diabetic Kidney Disease. Front Genet 2019, 10, 563. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, H.; Liu, J.; Han, P.; Li, X.; Bai, H.; Zhang, C.; Sun, X.; Teng, Y.; Zhang, Y.; Yuan, X.; Chu, Y.; Zhao, B. , Variations in MicroRNA-25 Expression Influence the Severity of Diabetic Kidney Disease. J Am Soc Nephrol 2017, 28, 3627–3638. [Google Scholar] [CrossRef]
- Yildirim, S.S.; Akman, D.; Catalucci, D.; Turan, B. , Relationship between downregulation of miRNAs and increase of oxidative stress in the development of diabetic cardiac dysfunction: junctin as a target protein of miR-1. Cell biochemistry and biophysics 2013, 67, 1397–408. [Google Scholar] [CrossRef]
- Tsai, S.; Shameli, A.; Santamaria, P. , CD8+ T cells in type 1 diabetes. Adv Immunol 2008, 100, 79–124. [Google Scholar]
- ElEssawy, B.; Li, X.C. , Type 1 diabetes and T regulatory cells. Pharmacol Res 2015, 98, 22–30. [Google Scholar] [CrossRef]
- Garzon, R.; Volinia, S.; Liu, C.G.; Fernandez-Cymering, C.; Palumbo, T.; Pichiorri, F.; Fabbri, M.; Coombes, K.; Alder, H.; Nakamura, T.; Flomenberg, N.; Marcucci, G.; Calin, G.A.; Kornblau, S.M.; Kantarjian, H.; Bloomfield, C.D.; Andreeff, M.; Croce, C.M. , MicroRNA signatures associated with cytogenetics and prognosis in acute myeloid leukemia. Blood 2008, 111, 3183–9. [Google Scholar] [CrossRef]
- Hui, A.B.; Shi, W.; Boutros, P.C.; Miller, N.; Pintilie, M.; Fyles, T.; McCready, D.; Wong, D.; Gerster, K.; Waldron, L.; Jurisica, I.; Penn, L.Z.; Liu, F.F. , Robust global micro-RNA profiling with formalin-fixed paraffin-embedded breast cancer tissues. Lab Invest 2009, 89, 597–606. [Google Scholar] [CrossRef]
- Gao, S.; Zhao, Z.Y.; Wu, R.; Zhang, Y.; Zhang, Z.Y. , Prognostic value of microRNAs in colorectal cancer: a meta-analysis. Cancer Manag Res 2018, 10, 907–929. [Google Scholar] [CrossRef]
- Schmidt, W.M.; Spiel, A.O.; Jilma, B.; Wolzt, M.; Muller, M. , In vivo profile of the human leukocyte microRNA response to endotoxemia. Biochem Biophys Res Commun 2009, 380, 437–41. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Sharples, R.A.; Scicluna, B.J.; Hill, A.F. , Exosomes provide a protective and enriched source of miRNA for biomarker profiling compared to intracellular and cell-free blood. J Extracell Vesicles 2014, 3. [Google Scholar] [CrossRef]
- Yang, J.; Wei, F.; Schafer, C.; Wong, D.T. , Detection of tumor cell-specific mRNA and protein in exosome-like microvesicles from blood and saliva. PLoS One 2014, 9, e110641. [Google Scholar] [CrossRef] [PubMed]
- Guay, C.; Menoud, V.; Rome, S.; Regazzi, R. , Horizontal transfer of exosomal microRNAs transduce apoptotic signals between pancreatic beta-cells. Cell Commun Signal 2015, 13, 17. [Google Scholar] [CrossRef]
- Setyowati Karolina, D.; Sepramaniam, S.; Tan, H.Z.; Armugam, A.; Jeyaseelan, K. , miR-25 and miR-92a regulate insulin I biosynthesis in rats. RNA Biol 2013, 10, 1365–78. [Google Scholar] [CrossRef] [PubMed]
- Knezevic, I.; Patel, A.; Sundaresan, N.R.; Gupta, M.P.; Solaro, R.J.; Nagalingam, R.S.; Gupta, M. , A novel cardiomyocyte-enriched microRNA, miR-378, targets insulin-like growth factor 1 receptor: implications in postnatal cardiac remodeling and cell survival. J Biol Chem 2012, 287, 12913–26. [Google Scholar] [CrossRef]
- Ludwig, N.; Leidinger, P.; Becker, K.; Backes, C.; Fehlmann, T.; Pallasch, C.; Rheinheimer, S.; Meder, B.; Stahler, C.; Meese, E.; Keller, A. , Distribution of miRNA expression across human tissues. Nucleic Acids Res 2016, 44, 3865–77. [Google Scholar] [CrossRef]
- Cabiati, M.; Federico, G.; Del Ry, S. , Importance of Studying Non-Coding RNA in Children and Adolescents with Type 1 Diabetes. Biomedicines, 2024; 12. [Google Scholar]
- Roggli, E.; Gattesco, S.; Caille, D.; Briet, C.; Boitard, C.; Meda, P.; Regazzi, R. , Changes in microRNA expression contribute to pancreatic beta-cell dysfunction in prediabetic NOD mice. Diabetes 2012, 61, 1742–51. [Google Scholar] [CrossRef]
- Guay, C.; Kruit, J.K.; Rome, S.; Menoud, V.; Mulder, N.L.; Jurdzinski, A.; Mancarella, F.; Sebastiani, G.; Donda, A.; Gonzalez, B.J.; Jandus, C.; Bouzakri, K.; Pinget, M.; Boitard, C.; Romero, P.; Dotta, F.; Regazzi, R. , Lymphocyte-Derived Exosomal MicroRNAs Promote Pancreatic beta Cell Death and May Contribute to Type 1 Diabetes Development. Cell Metab 2019, 29, 348–361. [Google Scholar] [CrossRef]
- Sims, E.K.; Lakhter, A.J.; Anderson-Baucum, E.; Kono, T.; Tong, X.; Evans-Molina, C. , MicroRNA 21 targets BCL2 mRNA to increase apoptosis in rat and human beta cells. Diabetologia 2017, 60, 1057–1065. [Google Scholar] [CrossRef]
- Tian, C.; Ouyang, X.; Lv, Q.; Zhang, Y.; Xie, W. , Cross-talks between microRNAs and mRNAs in pancreatic tissues of streptozotocin-induced type 1 diabetic mice. Biomed Rep 2015, 3, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Ban, E.; Jeong, S.; Park, M.; Kwon, H.; Park, J.; Song, E.J.; Kim, A. , Accelerated wound healing in diabetic mice by miRNA-497 and its anti-inflammatory activity. Biomed Pharmacother 2020, 121, 109613. [Google Scholar] [CrossRef] [PubMed]
- Diao, X.; Shen, E.; Wang, X.; Hu, B. , Differentially expressed microRNAs and their target genes in the hearts of streptozotocin-induced diabetic mice. Mol Med Rep 2011, 4, 633–40. [Google Scholar]
- Alipour, M.R.; Khamaneh, A.M.; Yousefzadeh, N.; Mohammad-nejad, D.; Soufi, F.G. , Upregulation of microRNA-146a was not accompanied by downregulation of pro-inflammatory markers in diabetic kidney. Mol Biol Rep 2013, 40, 6477–83. [Google Scholar] [CrossRef] [PubMed]
- Tsukita, S.; Yamada, T.; Takahashi, K.; Munakata, Y.; Hosaka, S.; Takahashi, H.; Gao, J.; Shirai, Y.; Kodama, S.; Asai, Y.; Sugisawa, T.; Chiba, Y.; Kaneko, K.; Uno, K.; Sawada, S.; Imai, J.; Katagiri, H. , MicroRNAs 106b and 222 Improve Hyperglycemia in a Mouse Model of Insulin-Deficient Diabetes via Pancreatic beta-Cell Proliferation. EBioMedicine 2017, 15, 163–172. [Google Scholar] [CrossRef]
- Bushati, N.; Cohen, S.M. , microRNA functions. Annu Rev Cell Dev Biol 2007, 23, 175–205. [Google Scholar] [CrossRef] [PubMed]
- Rosengren, A.; Dikaiou, P. , Cardiovascular outcomes in type 1 and type 2 diabetes. Diabetologia 2023, 66, 425–437. [Google Scholar] [CrossRef]
- Lee, Y.B.; Han, K.; Kim, B.; Lee, S.E.; Jun, J.E.; Ahn, J.; Kim, G.; Jin, S.M.; Kim, J.H. , Risk of early mortality and cardiovascular disease in type 1 diabetes: a comparison with type 2 diabetes, a nationwide study. Cardiovasc Diabetol 2019, 18, 157. [Google Scholar] [CrossRef]
- Rawshani, A.; Sattar, N.; Franzen, S.; Rawshani, A.; Hattersley, A.T.; Svensson, A.M.; Eliasson, B.; Gudbjornsdottir, S. , Excess mortality and cardiovascular disease in young adults with type 1 diabetes in relation to age at onset: a nationwide, register-based cohort study. Lancet 2018, 392, 477–486. [Google Scholar] [CrossRef]
- Lakhter, A.J.; Pratt, R.E.; Moore, R.E.; Doucette, K.K.; Maier, B.F.; DiMeglio, L.A.; Sims, E.K. , Beta cell extracellular vesicle miR-21-5p cargo is increased in response to inflammatory cytokines and serves as a biomarker of type 1 diabetes. Diabetologia 2018, 61, 1124–1134. [Google Scholar] [CrossRef]
- Zhu, Y.; Sun, Y.; Zhou, Y.; Zhang, Y.; Zhang, T.; Li, Y.; You, W.; Chang, X.; Yuan, L.; Han, X. , MicroRNA-24 promotes pancreatic beta cells toward dedifferentiation to avoid endoplasmic reticulum stress-induced apoptosis. J Mol Cell Biol 2019, 11, 747–760. [Google Scholar] [CrossRef] [PubMed]
- Taheri, M.; Eghtedarian, R.; Dinger, M.E.; Ghafouri-Fard, S. , Emerging roles of non-coding RNAs in the pathogenesis of type 1 diabetes mellitus. Biomed Pharmacother 2020, 129, 110509. [Google Scholar] [CrossRef]
- Jankauskas, S.S.; Gambardella, J.; Sardu, C.; Lombardi, A.; Santulli, G. , Functional Role of miR-155 in the Pathogenesis of Diabetes Mellitus and Its Complications. Noncoding RNA, 2021; 7. [Google Scholar]
- Xue, L.; Xiong, C.; Li, J.; Ren, Y.; Zhang, L.; Jiao, K.; Chen, C.; Ding, P. , miR-200-3p suppresses cell proliferation and reduces apoptosis in diabetic retinopathy via blocking the TGF-beta2/Smad pathway. Biosci Rep, 2020; 40. [Google Scholar]
- Erener, S.; Mojibian, M.; Fox, J.K.; Denroche, H.C.; Kieffer, T.J. , Circulating miR-375 as a biomarker of beta-cell death and diabetes in mice. Endocrinology 2013, 154, 603–8. [Google Scholar] [CrossRef]
- Coulson, D.J.; Bakhashab, S.; Latief, J.S.; Weaver, J.U. , MiR-126, IL-7, CXCR1/2 receptors, inflammation and circulating endothelial progenitor cells: The study on targets for treatment pathways in a model of subclinical cardiovascular disease (type 1 diabetes mellitus). J Transl Med 2021, 19, 140. [Google Scholar] [CrossRef] [PubMed]
- Nizam, R.; Malik, M.Z.; Jacob, S.; Alsmadi, O.; Koistinen, H.A.; Tuomilehto, J.; Alkandari, H.; Al-Mulla, F.; Thanaraj, T.A. , Circulating hsa-miR-320a and its regulatory network in type 1 diabetes mellitus. Front Immunol 2024, 15, 1376416. [Google Scholar] [CrossRef]
- Rasmi, Y.; Mohamed, Y.A.; Alipour, S.; Ahmed, S.; Abdelmajed, S.S. , The role of miR-143/miR-145 in the development, diagnosis, and treatment of diabetes. J Diabetes Metab Disord 2024, 23, 39–47. [Google Scholar] [CrossRef]
- Swolin-Eide, D.; Forsander, G.; Pundziute Lycka, A.; Novak, D.; Grillari, J.; Diendorfer, A.B.; Hackl, M.; Magnusson, P. , Circulating microRNAs in young individuals with long-duration type 1 diabetes in comparison with healthy controls. Sci Rep 2023, 13, 11634. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.B.; Li, Y.; Liu, G.Y.; Li, T.H.; Li, F.; Guan, J.; Wang, H.J. , Long non-coding RNA PVT1, a molecular sponge of miR-26b, is involved in the progression of hyperglycemia-induced collagen degradation in human chondrocytes by targeting CTGF/TGF-beta signal ways. Innate Immun 2020, 26, 204–214. [Google Scholar] [CrossRef]
- Singh, R.; Ha, S.E.; Wei, L.; Jin, B.; Zogg, H.; Poudrier, S.M.; Jorgensen, B.G.; Park, C.; Ronkon, C.F.; Bartlett, A.; Cho, S.; Morales, A.; Chung, Y.H.; Lee, M.Y.; Park, J.K.; Gottfried-Blackmore, A.; Nguyen, L.; Sanders, K.M.; Ro, S. , miR-10b-5p Rescues Diabetes and Gastrointestinal Dysmotility. Gastroenterology 2021, 160, 1662–1678. [Google Scholar] [CrossRef]
- Douvris, A.; Vinas, J.; Burns, K.D. , miRNA-486-5p: signaling targets and role in non-malignant disease. Cell Mol Life Sci 2022, 79, 376. [Google Scholar] [CrossRef]
- Garavelli, S.; Bruzzaniti, S.; Tagliabue, E.; Di Silvestre, D.; Prattichizzo, F.; Mozzillo, E.; Fattorusso, V.; La Sala, L.; Ceriello, A.; Puca, A.A.; Mauri, P.; Strollo, R.; Marigliano, M.; Maffeis, C.; Petrelli, A.; Bosi, E.; Franzese, A.; Galgani, M.; Matarese, G.; de Candia, P. , Plasma circulating miR-23~27~24 clusters correlate with the immunometabolic derangement and predict C-peptide loss in children with type 1 diabetes. Diabetologia 2020, 63, 2699–2712. [Google Scholar] [CrossRef]
- Otmani, K.; Rouas, R.; Lagneaux, L.; Krayem, M.; Duvillier, H.; Berehab, M.; Lewalle, P. , Acute myeloid leukemia-derived exosomes deliver miR-24-3p to hinder the T-cell immune response through DENN/MADD targeting in the NF-kappaB signaling pathways. Cell Commun Signal 2023, 21, 253. [Google Scholar] [CrossRef] [PubMed]
- Ray, S.L.; Coulson, D.J.; Yeoh, M.L.Y.; Tamara, A.; Latief, J.S.; Bakhashab, S.; Weaver, J.U. , The Role of miR-342 in Vascular Health. Study in Subclinical Cardiovascular Disease in Mononuclear Cells, Plasma, Inflammatory Cytokines and PANX2. Int J Mol Sci, 2020; 21. [Google Scholar]
- Daamouch, S.; Bluher, M.; Vazquez, D.C.; Hackl, M.; Hofbauer, L.C.; Rauner, M. , MiR-144-5p and miR-21-5p do not drive bone disease in a mouse model of type 1 diabetes mellitus. JBMR Plus 2024, 8, ziae036. [Google Scholar] [CrossRef] [PubMed]
- Ventriglia, G.; Nigi, L.; Sebastiani, G.; Dotta, F. , MicroRNAs: Novel Players in the Dialogue between Pancreatic Islets and Immune System in Autoimmune Diabetes. Biomed Res Int 2015, 2015, 749734. [Google Scholar] [CrossRef]
- Dooley, J.; Garcia-Perez, J.E.; Sreenivasan, J.; Schlenner, S.M.; Vangoitsenhoven, R.; Papadopoulou, A.S.; Tian, L.; Schonefeldt, S.; Serneels, L.; Deroose, C.; Staats, K.A.; Van der Schueren, B.; De Strooper, B.; McGuinness, O.P.; Mathieu, C.; Liston, A. , The microRNA-29 Family Dictates the Balance Between Homeostatic and Pathological Glucose Handling in Diabetes and Obesity. Diabetes 2016, 65, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.Y.; Yang, C.Y.; Rui, Z.L. , MicroRNA-125b-5p improves pancreatic beta-cell function through inhibiting JNK signaling pathway by targeting DACT1 in mice with type 2 diabetes mellitus. Life Sci 2019, 224, 67–75. [Google Scholar] [CrossRef]
- Liu, L.; Yan, J.; Xu, H.; Zhu, Y.; Liang, H.; Pan, W.; Yao, B.; Han, X.; Ye, J.; Weng, J. , Two Novel MicroRNA Biomarkers Related to beta-Cell Damage and Their Potential Values for Early Diagnosis of Type 1 Diabetes. J Clin Endocrinol Metab 2018, 103, 1320–1329. [Google Scholar] [CrossRef]
- Bagge, A.; Dahmcke, C.M.; Dalgaard, L.T. , Syntaxin-1a is a direct target of miR-29a in insulin-producing beta-cells. Horm Metab Res 2013, 45, 463–6. [Google Scholar]
- Ma, X.; Becker Buscaglia, L.E.; Barker, J.R.; Li, Y. , MicroRNAs in NF-kappaB signaling. J Mol Cell Biol 2011, 3, 159–66. [Google Scholar] [CrossRef]
- Wang, P.; Hou, J.; Lin, L.; Wang, C.; Liu, X.; Li, D.; Ma, F.; Wang, Z.; Cao, X. , Inducible microRNA-155 feedback promotes type I IFN signaling in antiviral innate immunity by targeting suppressor of cytokine signaling 1. J Immunol 2010, 185, 6226–33. [Google Scholar] [CrossRef]
- Faraoni, I.; Antonetti, F.R.; Cardone, J.; Bonmassar, E. , miR-155 gene: a typical multifunctional microRNA. Biochim Biophys Acta 2009, 1792, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Belgardt, B.F.; Ahmed, K.; Spranger, M.; Latreille, M.; Denzler, R.; Kondratiuk, N.; von Meyenn, F.; Villena, F.N.; Herrmanns, K.; Bosco, D.; Kerr-Conte, J.; Pattou, F.; Rulicke, T.; Stoffel, M. , The microRNA-200 family regulates pancreatic beta cell survival in type 2 diabetes. Nat Med 2015, 21, 619–27. [Google Scholar] [CrossRef] [PubMed]
- Fayyad-Kazan, H.; Rouas, R.; Fayyad-Kazan, M.; Badran, R.; El Zein, N.; Lewalle, P.; Najar, M.; Hamade, E.; Jebbawi, F.; Merimi, M.; Romero, P.; Burny, A.; Badran, B.; Martiat, P. , MicroRNA profile of circulating CD4-positive regulatory T cells in human adults and impact of differentially expressed microRNAs on expression of two genes essential to their function. J Biol Chem 2012, 287, 9910–9922. [Google Scholar] [CrossRef]
- Devlin, C.; Greco, S.; Martelli, F.; Ivan, M. , miR-210: More than a silent player in hypoxia. IUBMB Life 2011, 63, 94–100. [Google Scholar] [CrossRef]
- Molitch, M.E.; DeFronzo, R.A.; Franz, M.J.; Keane, W.F.; Mogensen, C.E.; Parving, H.H.; Steffes, M.W.; American Diabetes, A. , Nephropathy in diabetes. Diabetes Care 2004, 27 Suppl 1, S79–83. [Google Scholar]
- Argyropoulos, C.; Wang, K.; McClarty, S.; Huang, D.; Bernardo, J.; Ellis, D.; Orchard, T.; Galas, D.; Johnson, J. , Urinary microRNA profiling in the nephropathy of type 1 diabetes. PLoS One 2013, 8, e54662. [Google Scholar] [CrossRef]
- Wang, G.; Kwan, B.C.; Lai, F.M.; Chow, K.M.; Li, P.K.; Szeto, C.C. , Urinary sediment miRNA levels in adult nephrotic syndrome. Clin Chim Acta 2013, 418, 5–11. [Google Scholar] [CrossRef]
- Wang, S.; Aurora, A.B.; Johnson, B.A.; Qi, X.; McAnally, J.; Hill, J.A.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. , The endothelial-specific microRNA miR-126 governs vascular integrity and angiogenesis. Dev Cell 2008, 15, 261–71. [Google Scholar] [CrossRef]
- Long, J.; Wang, Y.; Wang, W.; Chang, B.H.; Danesh, F.R. , Identification of microRNA-93 as a novel regulator of vascular endothelial growth factor in hyperglycemic conditions. J Biol Chem 2010, 285, 23457–65. [Google Scholar] [CrossRef]
- Samandari, N.; Mirza, A.H.; Kaur, S.; Hougaard, P.; Nielsen, L.B.; Fredheim, S.; Mortensen, H.B.; Pociot, F. , Influence of Disease Duration on Circulating Levels of miRNAs in Children and Adolescents with New Onset Type 1 Diabetes. Noncoding RNA, 2018; 4. [Google Scholar]
- Wallberg, M.; Recino, A.; Phillips, J.; Howie, D.; Vienne, M.; Paluch, C.; Azuma, M.; Wong, F.S.; Waldmann, H.; Cooke, A. , Anti-CD3 treatment up-regulates programmed cell death protein-1 expression on activated effector T cells and severely impairs their inflammatory capacity. Immunology 2017, 151, 248–260. [Google Scholar] [CrossRef]
- Benson, R.A.; Garcon, F.; Recino, A.; Ferdinand, J.R.; Clatworthy, M.R.; Waldmann, H.; Brewer, J.M.; Okkenhaug, K.; Cooke, A.; Garside, P.; Wallberg, M. , Non-Invasive Multiphoton Imaging of Islets Transplanted Into the Pinna of the NOD Mouse Ear Reveals the Immediate Effect of Anti-CD3 Treatment in Autoimmune Diabetes. Front Immunol 2018, 9, 1006. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.H.; Wu, Z.; Hu, C.; Zhang, M.; Wang, W.Z.; Hu, X.B. , Endoplasmic reticulum stress and destruction of pancreatic beta cells in type 1 diabetes. Chin Med J (Engl) 2020, 133, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Padgett, L.E.; Broniowska, K.A.; Hansen, P.A.; Corbett, J.A.; Tse, H.M. , The role of reactive oxygen species and proinflammatory cytokines in type 1 diabetes pathogenesis. Ann N Y Acad Sci 2013, 1281, 16–35. [Google Scholar] [CrossRef] [PubMed]
- Leenders, F.; Groen, N.; de Graaf, N.; Engelse, M.A.; Rabelink, T.J.; de Koning, E.J.P.; Carlotti, F. , Oxidative Stress Leads to beta-Cell Dysfunction Through Loss of beta-Cell Identity. Front Immunol 2021, 12, 690379. [Google Scholar] [CrossRef]
- Wei, J.; Zhang, Y.; Luo, Y.; Wang, Z.; Bi, S.; Song, D.; Dai, Y.; Wang, T.; Qiu, L.; Wen, L.; Yuan, L.; Yang, J.Y. , Aldose reductase regulates miR-200a-3p/141-3p to coordinate Keap1-Nrf2, Tgfbeta1/2, and Zeb1/2 signaling in renal mesangial cells and the renal cortex of diabetic mice. Free Radic Biol Med 2014, 67, 91–102. [Google Scholar] [CrossRef]
- Ghaffari, M.; Razi, S.; Zalpoor, H.; Nabi-Afjadi, M.; Mohebichamkhorami, F.; Zali, H. , Association of MicroRNA-146a with Type 1 and 2 Diabetes and their Related Complications. J Diabetes Res 2023, 2023, 2587104. [Google Scholar] [CrossRef]
- Velosa, A.P.; Teodoro, W.R.; dos Anjos, D.M.; Konno, R.; Oliveira, C.C.; Katayama, M.L.; Parra, E.R.; Capelozzi, V.L.; Yoshinari, N.H. , Collagen V-induced nasal tolerance downregulates pulmonary collagen mRNA gene and TGF-beta expression in experimental systemic sclerosis. Respir Res 2010, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Salas-Perez, F.; Codner, E.; Valencia, E.; Pizarro, C.; Carrasco, E.; Perez-Bravo, F. , MicroRNAs miR-21a and miR-93 are down regulated in peripheral blood mononuclear cells (PBMCs) from patients with type 1 diabetes. Immunobiology 2013, 218, 733–7. [Google Scholar] [CrossRef]
- Qadir, M.M.F.; Klein, D.; Alvarez-Cubela, S.; Dominguez-Bendala, J.; Pastori, R.L. , The Role of MicroRNAs in Diabetes-Related Oxidative Stress. Int J Mol Sci, 2019; 20. [Google Scholar]
- Gao, X.; Zhao, S. , miRNA-16-5p inhibits the apoptosis of high glucose-induced pancreatic beta cells via targeting of CXCL10: potential biomarkers in type 1 diabetes mellitus. Endokrynol Pol 2020, 71, 404–410. [Google Scholar] [CrossRef]
- Roggli, E.; Britan, A.; Gattesco, S.; Lin-Marq, N.; Abderrahmani, A.; Meda, P.; Regazzi, R. , Involvement of microRNAs in the cytotoxic effects exerted by proinflammatory cytokines on pancreatic beta-cells. Diabetes 2010, 59, 978–86. [Google Scholar] [CrossRef]
- Lin, L.; Mahner, S.; Jeschke, U.; Hester, A. , The Distinct Roles of Transcriptional Factor KLF11 in Normal Cell Growth Regulation and Cancer as a Mediator of TGF-beta Signaling Pathway. Int J Mol Sci, 2020; 21. [Google Scholar]
- Pezzolesi, M.G.; Satake, E.; McDonnell, K.P.; Major, M.; Smiles, A.M.; Krolewski, A.S. , Circulating TGF-beta1-Regulated miRNAs and the Risk of Rapid Progression to ESRD in Type 1 Diabetes. Diabetes 2015, 64, 3285–93. [Google Scholar] [CrossRef] [PubMed]
- Pooja Rathan, V.; Bhuvaneshwari, K.; Nideesh Adit, G.; Kavyashree, S.; Thulasi, N.; Geetha, A.V.S.; Milan, K.L.; Ramkumar, K.M. , Therapeutic potential of SMAD7 targeting miRNA in the pathogenesis of diabetic nephropathy. Arch Biochem Biophys 2024, 764, 110265. [Google Scholar] [CrossRef] [PubMed]
- Qin, W.; Shi, Y.; Zhao, B.; Yao, C.; Jin, L.; Ma, J.; Jin, Y. , miR-24 regulates apoptosis by targeting the open reading frame (ORF) region of FAF1 in cancer cells. PLoS One 2010, 5, e9429. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.C.; Hilyard, A.C.; Wu, C.; Davis, B.N.; Hill, N.S.; Lal, A.; Lieberman, J.; Lagna, G.; Hata, A. , Molecular basis for antagonism between PDGF and the TGFbeta family of signalling pathways by control of miR-24 expression. EMBO J 2010, 29, 559–73. [Google Scholar] [CrossRef]
- Donath, M.Y.; Storling, J.; Maedler, K.; Mandrup-Poulsen, T. , Inflammatory mediators and islet beta-cell failure: a link between type 1 and type 2 diabetes. J Mol Med (Berl) 2003, 81, 455–70. [Google Scholar] [CrossRef]
- Chen, X.; Ba, Y.; Ma, L.; Cai, X.; Yin, Y.; Wang, K.; Guo, J.; Zhang, Y.; Chen, J.; Guo, X.; Li, Q.; Li, X.; Wang, W.; Zhang, Y.; Wang, J.; Jiang, X.; Xiang, Y.; Xu, C.; Zheng, P.; Zhang, J.; Li, R.; Zhang, H.; Shang, X.; Gong, T.; Ning, G.; Wang, J.; Zen, K.; Zhang, J.; Zhang, C.Y. , Characterization of microRNAs in serum: a novel class of biomarkers for diagnosis of cancer and other diseases. Cell Res 2008, 18, 997–1006. [Google Scholar] [CrossRef]
- Hu, Z.; Dong, J.; Wang, L.E.; Ma, H.; Liu, J.; Zhao, Y.; Tang, J.; Chen, X.; Dai, J.; Wei, Q.; Zhang, C.; Shen, H. , Serum microRNA profiling and breast cancer risk: the use of miR-484/191 as endogenous controls. Carcinogenesis 2012, 33, 828–34. [Google Scholar] [CrossRef]
- Wu, Q.; Wang, C.; Lu, Z.; Guo, L.; Ge, Q. , Analysis of serum genome-wide microRNAs for breast cancer detection. Clin Chim Acta, 2012; 413, 1058–1065. [Google Scholar]
- Li, L.M.; Hu, Z.B.; Zhou, Z.X.; Chen, X.; Liu, F.Y.; Zhang, J.F.; Shen, H.B.; Zhang, C.Y.; Zen, K. , Serum microRNA profiles serve as novel biomarkers for HBV infection and diagnosis of HBV-positive hepatocarcinoma. Cancer research 2010, 70, 9798–807. [Google Scholar] [CrossRef]
- Razumilava, N.; Bronk, S.F.; Smoot, R.L.; Fingas, C.D.; Werneburg, N.W.; Roberts, L.R.; Mott, J.L. , miR-25 targets TNF-related apoptosis inducing ligand (TRAIL) death receptor-4 and promotes apoptosis resistance in cholangiocarcinoma. Hepatology 2012, 55, 465–75. [Google Scholar] [CrossRef]
- Zhang, H.; Zuo, Z.; Lu, X.; Wang, L.; Wang, H.; Zhu, Z. , MiR-25 regulates apoptosis by targeting Bim in human ovarian cancer. Oncol Rep 2012, 27, 594–8. [Google Scholar]
- Fu, Y.; Zhang, Y.; Wang, Z.; Wang, L.; Wei, X.; Zhang, B.; Wen, Z.; Fang, H.; Pang, Q.; Yi, F. , Regulation of NADPH oxidase activity is associated with miRNA-25-mediated NOX4 expression in experimental diabetic nephropathy. Am J Nephrol 2010, 32, 581–9. [Google Scholar] [CrossRef]
- Gonzalez-Martin, A.; Adams, B.D.; Lai, M.; Shepherd, J.; Salvador-Bernaldez, M.; Salvador, J.M.; Lu, J.; Nemazee, D.; Xiao, C. , The microRNA miR-148a functions as a critical regulator of B cell tolerance and autoimmunity. Nat Immunol 2016, 17, 433–40. [Google Scholar] [CrossRef] [PubMed]
- Ha, S.E.; Singh, R.; Jin, B.; Baek, G.; Jorgensen, B.G.; Zogg, H.; Debnath, S.; Park, H.S.; Cho, H.; Watkins, C.M.; Cho, S.; Kim, M.S.; Lee, M.Y.; Yu, T.Y.; Jeong, J.W.; Ro, S. , miR-10a/b-5p-NCOR2 Regulates Insulin-Resistant Diabetes in Female Mice. Int J Mol Sci, 2024; 25. [Google Scholar]
- Martello, G.; Rosato, A.; Ferrari, F.; Manfrin, A.; Cordenonsi, M.; Dupont, S.; Enzo, E.; Guzzardo, V.; Rondina, M.; Spruce, T.; Parenti, A.R.; Daidone, M.G.; Bicciato, S.; Piccolo, S. , A MicroRNA targeting dicer for metastasis control. Cell 2010, 141, 1195–207. [Google Scholar] [CrossRef] [PubMed]
- Kfir-Erenfeld, S.; Haggiag, N.; Biton, M.; Stepensky, P.; Assayag-Asherie, N.; Yefenof, E. , miR-103 inhibits proliferation and sensitizes hemopoietic tumor cells for glucocorticoid-induced apoptosis. Oncotarget 2017, 8, 472–489. [Google Scholar] [CrossRef]
- Barutta, F.; Corbetta, B.; Bellini, S.; Guarrera, S.; Matullo, G.; Scandella, M.; Schalkwijk, C.; Stehouwer, C.D.; Chaturvedi, N.; Soedamah-Muthu, S.S.; Durazzo, M.; Gruden, G. , MicroRNA 146a is associated with diabetic complications in type 1 diabetic patients from the EURODIAB PCS. J Transl Med 2021, 19, 475. [Google Scholar] [CrossRef]
- Lu, L.F.; Boldin, M.P.; Chaudhry, A.; Lin, L.L.; Taganov, K.D.; Hanada, T.; Yoshimura, A.; Baltimore, D.; Rudensky, A.Y. , Function of miR-146a in controlling Treg cell-mediated regulation of Th1 responses. Cell 2010, 142, 914–29. [Google Scholar] [CrossRef]
- Ellenrieder, V.; Buck, A.; Harth, A.; Jungert, K.; Buchholz, M.; Adler, G.; Urrutia, R.; Gress, T.M. , KLF11 mediates a critical mechanism in TGF-beta signaling that is inactivated by Erk-MAPK in pancreatic cancer cells. Gastroenterology 2004, 127, 607–20. [Google Scholar] [CrossRef]
- Guasparri, I.; Keller, S.A.; Cesarman, E. , KSHV vFLIP is essential for the survival of infected lymphoma cells. J Exp Med 2004, 199, 993–1003. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Jin, W.; Hardegen, N.; Lei, K.J.; Li, L.; Marinos, N.; McGrady, G.; Wahl, S.M. , Conversion of peripheral CD4+CD25- naive T cells to CD4+CD25+ regulatory T cells by TGF-beta induction of transcription factor Foxp3. J Exp Med 2003, 198, 1875–86. [Google Scholar] [CrossRef]
- Erener, S.; Ellis, C.E.; Ramzy, A.; Glavas, M.M.; O'Dwyer, S.; Pereira, S.; Wang, T.; Pang, J.; Bruin, J.E.; Riedel, M.J.; Baker, R.K.; Webber, T.D.; Lesina, M.; Bluher, M.; Algul, H.; Kopp, J.L.; Herzig, S.; Kieffer, T.J. , Deletion of pancreas-specific miR-216a reduces beta-cell mass and inhibits pancreatic cancer progression in mice. Cell Rep Med 2021, 2, 100434. [Google Scholar] [CrossRef]
- Wang, P.; Liu, Q.; Zhao, H.; Bishop, J.O.; Zhou, G.; Olson, L.K.; Moore, A. , miR-216a-targeting theranostic nanoparticles promote proliferation of insulin-secreting cells in type 1 diabetes animal model. Sci Rep 2020, 10, 5302. [Google Scholar] [CrossRef] [PubMed]
- Scherm, M.G.; Serr, I.; Zahm, A.M.; Schug, J.; Bellusci, S.; Manfredini, R.; Salb, V.K.; Gerlach, K.; Weigmann, B.; Ziegler, A.G.; Kaestner, K.H.; Daniel, C. , miRNA142-3p targets Tet2 and impairs Treg differentiation and stability in models of type 1 diabetes. Nature communications 2019, 10, 5697. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Pathophysiological mechanisms underlying Type 1 diabetes. Type 1 diabetes (T1D) results from the progressive destruction of pancreatic islet β-cells, driven by autoantigen release, autoimmune activation, oxidative and ER stress, cytokine-induced damage, and apoptosis. This cascade of pathological events leads to β-cell loss, insulin deficiency, and hyperglycemia, ultimately disrupting insulin signaling pathways.
Figure 1.
Pathophysiological mechanisms underlying Type 1 diabetes. Type 1 diabetes (T1D) results from the progressive destruction of pancreatic islet β-cells, driven by autoantigen release, autoimmune activation, oxidative and ER stress, cytokine-induced damage, and apoptosis. This cascade of pathological events leads to β-cell loss, insulin deficiency, and hyperglycemia, ultimately disrupting insulin signaling pathways.
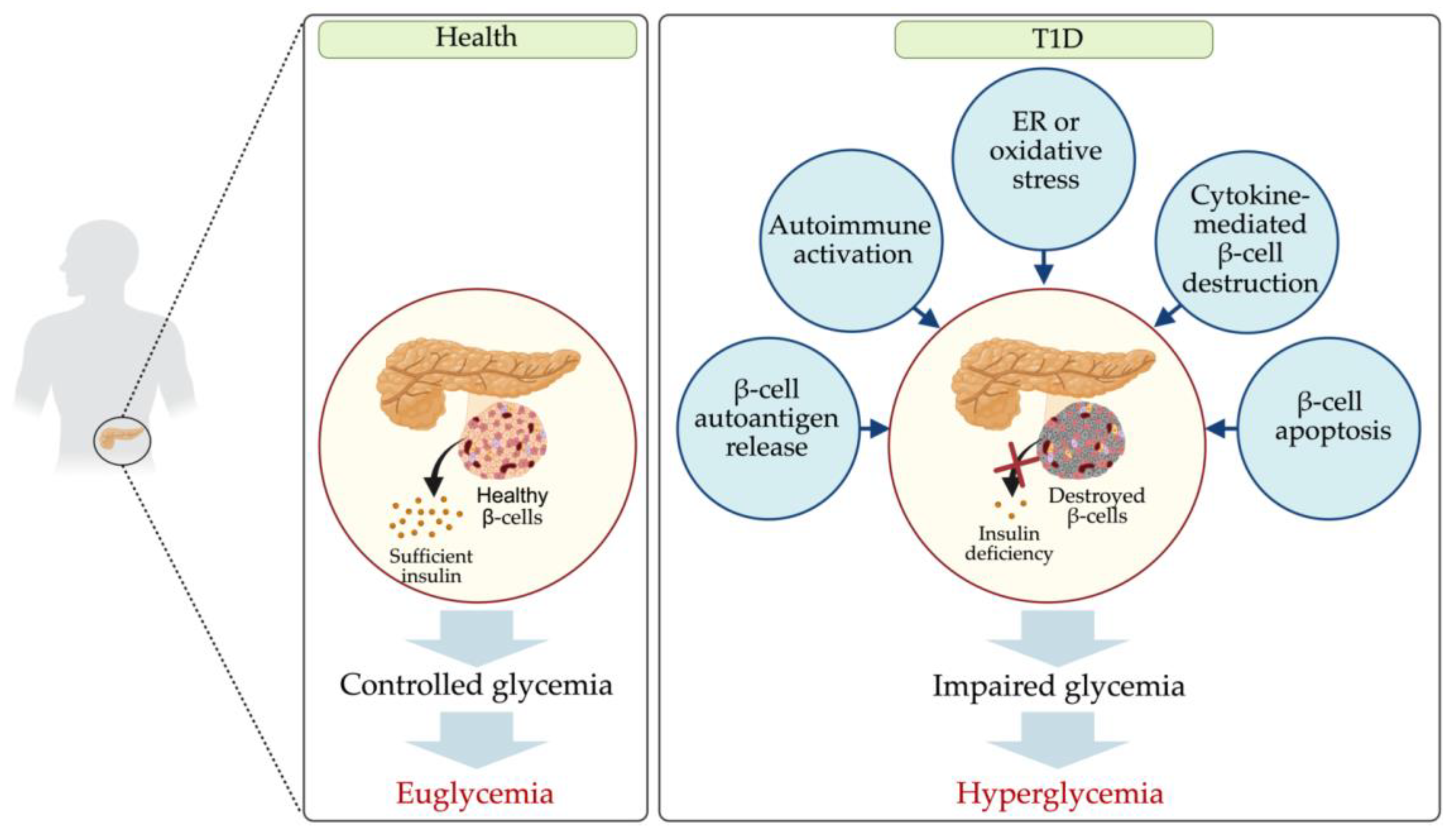
Figure 2.
Dysregulated miRNAs in diabetes-related cardiomyopathy and nephropathy. .In T1D patients, diabetic cardiomyopathy and diabetic nephropathy are closely associated. Dysregulated miRNAs contribute to disease progression, with downregulated miRNAs shown in blue and upregulated miRNAs in red.
Figure 2.
Dysregulated miRNAs in diabetes-related cardiomyopathy and nephropathy. .In T1D patients, diabetic cardiomyopathy and diabetic nephropathy are closely associated. Dysregulated miRNAs contribute to disease progression, with downregulated miRNAs shown in blue and upregulated miRNAs in red.
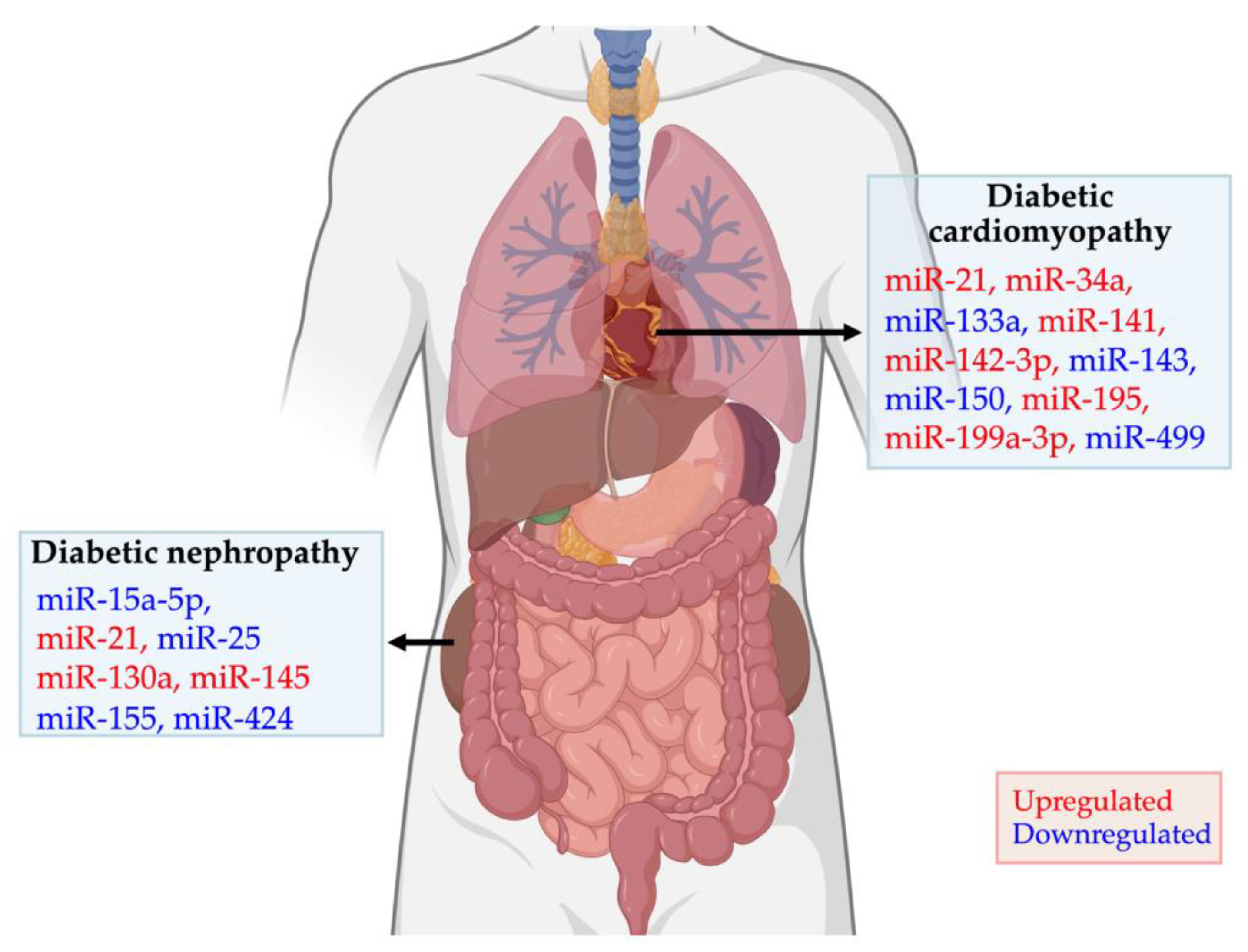
Figure 3.
miRNA regulation in T1D pathogenesis and their target pathways. Dysregulated miRNAs play a pivotal role in T1D pathogenesis by modulating key immune and apoptotic processes. Upregulated miRNAs and their target genes are highlighted in red, while downregulated ones are shown in blue. These miRNAs contribute to regulatory T-cell dysfunction, impaired immune tolerance, inhibition of β-cell proliferation, and apoptosis, ultimately driving autoimmune β-cell destruction.
Figure 3.
miRNA regulation in T1D pathogenesis and their target pathways. Dysregulated miRNAs play a pivotal role in T1D pathogenesis by modulating key immune and apoptotic processes. Upregulated miRNAs and their target genes are highlighted in red, while downregulated ones are shown in blue. These miRNAs contribute to regulatory T-cell dysfunction, impaired immune tolerance, inhibition of β-cell proliferation, and apoptosis, ultimately driving autoimmune β-cell destruction.
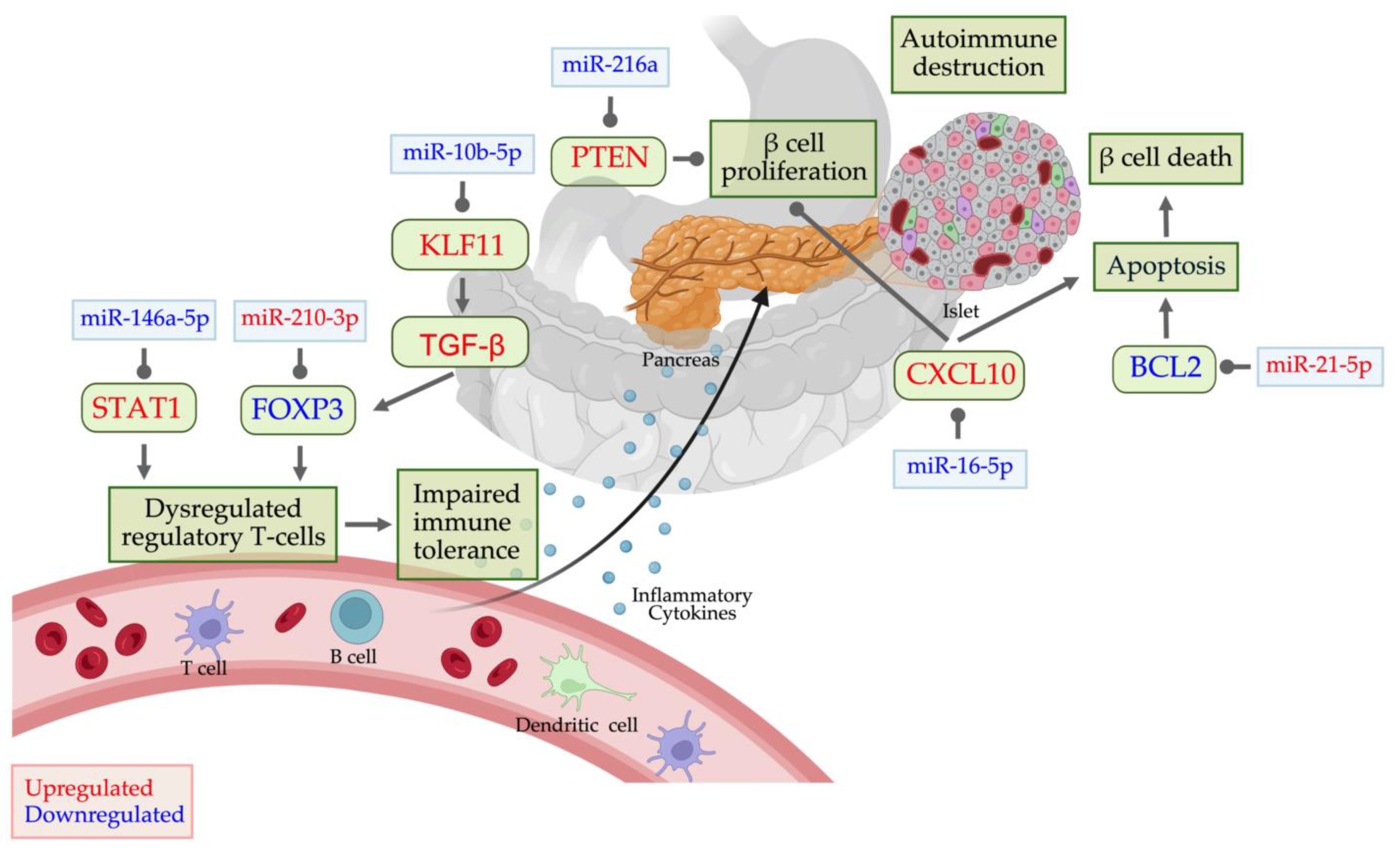
Figure 4.
Conserved dysregulated miRNAs in T1D patients and murine models, and therapeutic strategies.In T1D patients, 134 miRNAs are upregulated and 96 downregulated, while T1D animal models exhibit 67 upregulated and 77 downregulated miRNAs. Ten miRNAs (miR-21, miR-29a, miR-142-5p, miR-145, miR-146a, miR-186, miR-195, miR-199a-3p, miR-208, and miR-323-3p) are consistently upregulated in both humans and animal models. Conversely, nine miRNAs (miR-10b-5p, miR-16-5p, miR-17-5p, miR-23a, miR-106b, miR-126-5p, miR-143-3p, miR-150, miR-155, miR-222-3p, and miR-409-3p) are consistently downregulated in both. miR-106b-5p and miR-222-3p currently under investigation in preclinical and clinical trials for their therapeutic potential in T1D. As therapeutic strategies, ungulated miRNAs may be targeted with inhibitors, while downregulated miRNAs can be restored using mimics.
Figure 4.
Conserved dysregulated miRNAs in T1D patients and murine models, and therapeutic strategies.In T1D patients, 134 miRNAs are upregulated and 96 downregulated, while T1D animal models exhibit 67 upregulated and 77 downregulated miRNAs. Ten miRNAs (miR-21, miR-29a, miR-142-5p, miR-145, miR-146a, miR-186, miR-195, miR-199a-3p, miR-208, and miR-323-3p) are consistently upregulated in both humans and animal models. Conversely, nine miRNAs (miR-10b-5p, miR-16-5p, miR-17-5p, miR-23a, miR-106b, miR-126-5p, miR-143-3p, miR-150, miR-155, miR-222-3p, and miR-409-3p) are consistently downregulated in both. miR-106b-5p and miR-222-3p currently under investigation in preclinical and clinical trials for their therapeutic potential in T1D. As therapeutic strategies, ungulated miRNAs may be targeted with inhibitors, while downregulated miRNAs can be restored using mimics.
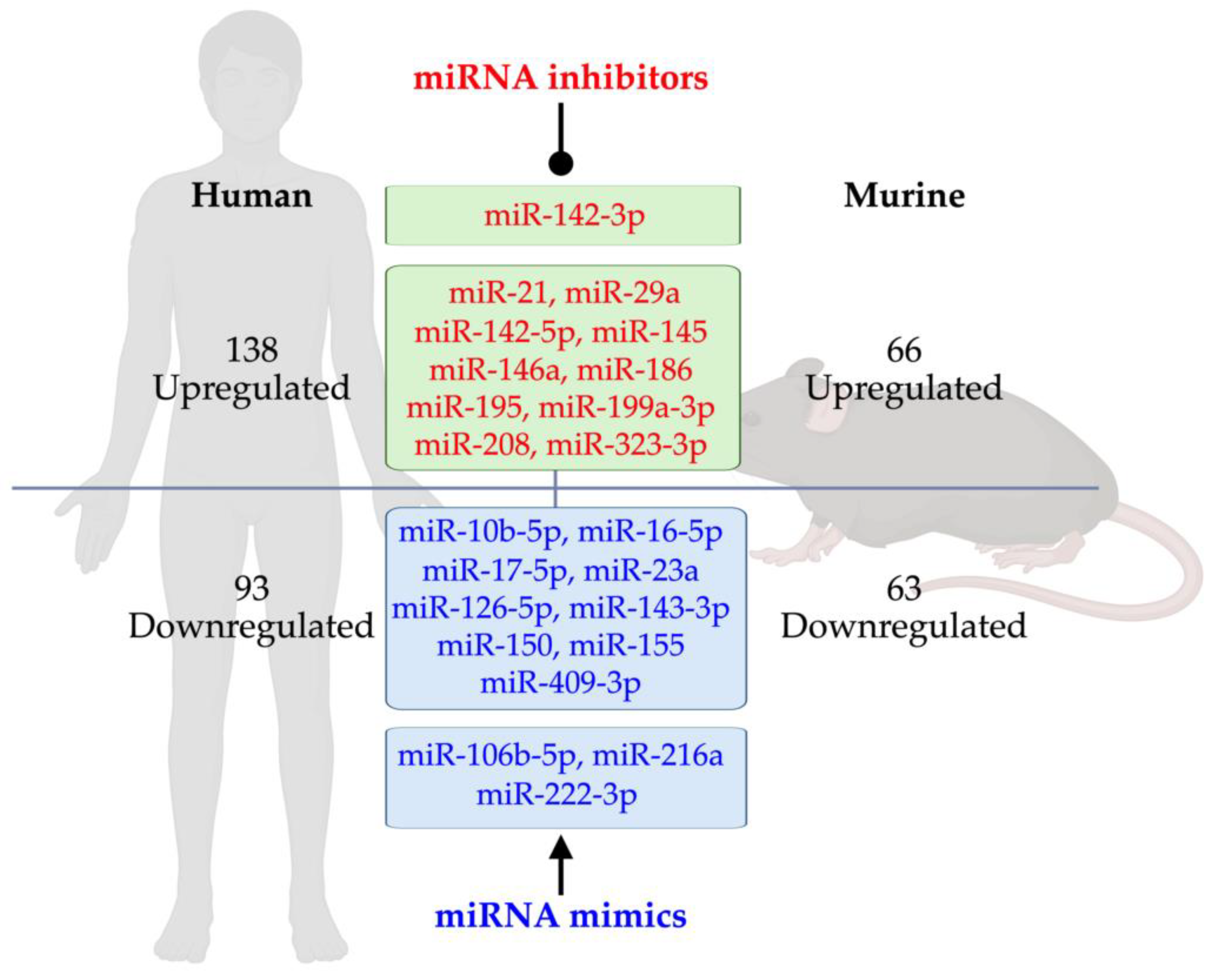

Table 1.
Altered miRNA profiles in various blood and urine sample types from T1D patients.
| Source | Detection method | miRNA profile | Ref. | ||||
|---|---|---|---|---|---|---|---|
| Sample | T1D patient characteristic | Expression | miRNA | ||||
| Diagnosis | Age (years) | n | n | ||||
| Source | Detection method | miRNA profile | Ref. | ||||
| Serum | 21 to 42 days | 9.0 ± 1.8 | 10 | qPCR | Up | let-7e/g-5p miR-18a-5p miR-23b-3p miR-25-3p miR-30e-5p miR-93-5p miR-103a-2-5p miR-125a-3p miR-140-5p miR-144-5p miR-182-5p miR-183-5p miR-192-5p miR-214-5p miR-221-3p miR-222-3p miR-324-3p/5p miR-331-3p miR-345-5p miR-377-3p miR-454-3p miR-500a-5p miR-502-3p miR-1468 |
[19] |
| Down | miR-100-5p miR-154-3p miR-490-5p miR-630 miR-636 miR-639 miR-675-3p miR-720 |
||||||
| 12 months |
404 | Sequencing qPCR |
Up | miR-10a miR-21 miR-24 miR-25 miR-26a/b miR-27a/b miR-29a/b miR-30a-5p miR-103 miR-125b miR-148a miR-152 miR-181a miR-199a miR-200a/c miR-210 miR-222 miR-320a miR-340 |
[31] | ||
| <6 months or 2 to 5 yrs |
26 | qPCR | Up | miR-10a miR-21 miR-27a miR-92a miR-100 miR-148a miR-200a miR-208 miR-212 miR-323-3p miR-346 miR-451 miR-886-3p |
[32] | ||
| Down | miR-16-5p miR-125a-5p miR-126 miR-146a miR-155 miR-197 miR-342-3p miR-374 miR-454 miR-518d |
||||||
| 15.71 ± 11.33 yrs | 33.57 ± 8.17 | 15 | qPCR | Up | miR-21-5p miR-148a |
[33] | |
| Plasma | <5 yrs | 19.2 ± 6.4 | 16 | Microarray qPCR | Up | miR-15b-5p miR-21-3p/5p miR-25-3p miR-29a-3p miR-101-3p miR-103a-3p miR-133a-5p miR-148a-3p miR-148b-3p miR-155-5p miR-200a/c-3p miR-210-3p miR-222-3p miR-320 miR-342 miR-1275 |
[34] |
| Down | miR-29b-3p miR-146a-5p miR-181a-5p miR-338-3p |
||||||
| ≥5 yrs | 19.9 ± 4.6 | 17 | Microarray qPCR | Up | miR-26b-5p miR-146a-5p miR-148b-3p miR-338-3p miR-340-5p miR-1275 |
[34] | |
| Down | miR-15b-5p miR-103a-3p miR-126-3p miR-148a-3p miR-155-5p miR-181a-5p miR-200a/c-3p miR-210-3p miR-222-3p |
||||||
| < 1 yr | 12.93 ± 3.34 | 16 | qPCR |
Up | miR-21 miR-210 |
[35] | |
| - | 25.9 ± 5.7 | 16 | qPCR | Up | miR-21 miR-24 miR-29a miR-30d miR-34a miR-126 miR-146a miR-148a miR-375 miR-376a |
[3] | |
| - | 37 | 95 | qPCR | Up | miR-125b-5p miR-365a-3p miR-770-5p |
[18] | |
| Down | miR-5190 | ||||||
| < 1 yr | 31.0±10.2 | 34 | qPCR | Down | miR-409-3p | [36] | |
| Peripheral blood | - | 26.93 ± 9.62 | 12 | Sequencing qPCR |
Up | let-7i-5p miR-26b-5p miR-99b-5p miR-342-3p miR-501-3p miR-652-3p |
[37] |
| Down | miR-15a-5p miR-15b-3p/5p miR-16-5p miR-16-2-3p miR-17-5p miR-21-5p miR-25-3p miR-26a/b-5p miR-27b-3p miR-30e-5p miR-93-5p miR-98-5p miR-100-5p miR-101-3p miR-103a-5p miR-107 miR-106b-3p/5p miR-126-3p/5p miR-130a/b-3p miR-143-3p miR-144-3p/5p miR-181a-5p miR-185-5p miR-221-3p miR-363-3p miR-532-5p |
||||||
| PBMCs | - | 23.5 ± 3.9 | 11 | Microarray | Up | let-7f/g miR-7 miR-10a miR-15b miR-16 miR-18b miR-19a miR-20a miR-20b miR-21 miR-26b miR-27b miR-30e miR-32 miR-33a miR-98 miR-101 miR-126 miR-148a/b miR-186 miR-195 miR-199a-3p miR-301a miR-335 miR-338-3p miR-340 miR-424 miR-450a miR-454 miR-542-3p miR-548c-3p |
[20] |
| Down | miR-140-3p miR-324-5p miR-342-3p/5p miR-423-5p miR-720 miR-766 miR-940 miR-1275 |
||||||
| Newly diagnosed T1D patients | 17.50 ± 3.68 | 60 | Microarray | Up | miR-320b miR-486-5p miR-652 miR-1275 miR-1301 |
[38] | |
| Down | miR-15b miR-19b miR-22 miR-23a miR-25 miR-28-5p miR-29a miR-30b/c miR-146a miR-146b-5p miR-200c miR-221 miR-342-5p |
||||||
| - | - | 20 | qPCR | Up | miR-22 | [39] | |
| Down | miR-150 | ||||||
| - | - | 78 | qPCR | Down | miR-146a miR-150 miR-424 |
[40] | |
| ≥10 yrs | 37.35 ± 12.82 | 31 | Sequencing qPCR |
Up | miR-133a-3p miR-142-5p miR-144-5p miR-145-3p miR-193a-5p miR-199a-5p miR-382-5p miR-409-5p miR-486-5p miR-543 miR-873-5p miR-1249-3p miR-1299 miR-3150b-3p miR-4531 |
[41] | |
| Down | miR-16-5p miR-144-3p miR-409-5p miR-501-3p miR-1271-5p miR-4485-3p |
||||||
| T-cells and Tregs isolated from peripheral blood | - | - | 5 | qPCR | Up | miR-510 | [42] |
| Down | miR-191 miR-342 |
||||||
| Urine | < 1 yrs | 12.93 ± 3.34 | 68 | qPCR | Up | miR-21 miR-210 |
[35] |
| Down | miR-126 | ||||||
| Plasma-derived exosome | 25.3 ± 15.9 yrs | 46.1 ± 14.4 | 12 | Microarray qPCR |
Up | miR-25-3p | [43] |
| Down | miR-16 miR-302d-3p miR-378e miR-570-3p miR-574-5p miR-579 |
||||||
| Urine-derived exosome | 30.7 ± 6.3 yrs | 57.9 ± 3.7 |
12 | qPCR | Up | miR-130a miR-145 |
[44] |
| Down | miR-155 miR-424 |
||||||
Table 2.
Altered miRNA profiles across various cells and tissues in T1D rodent models.
| Species | Diabetic Animal Models |
Source | Detection Methods | miRNA Expression Alteration | Ref. | |
|---|---|---|---|---|---|---|
| Expression | miRNAs | |||||
| Mice | Akita spontaneous mutation (Ins2Akita) mice | Retina |
Microarray qPCR |
Up | miR-7a miR-28 miR-124 miR-186 miR-199a-3p miR-200b miR-369-5p miR-410 miR-429 |
[77] |
| Down | miR-184 miR-296-5p miR-467b miR-539 miR-1196 miR-1224 |
|||||
| Pre-diabetic non-obese diabetic (NOD) mice | Pancreatic islet, cultured islet, infiltrating lymphocytes | qPCR | Up | miR-29a miR-29b miR-29c miR-142-5p miR-155 |
[106,107] | |
| Down | miR-142-3p miR-150 |
|||||
| Pancreatic β-cells | qPCR | Up | miR-142-5p | [107] |
||
| Down | miR-150 miR-155 |
|||||
| Diabetic NOD mice | Pancreatic islet/plasma | qPCR | Up | miR-21 | [108] | |
| Microarray qPCR | Down | miR-126a-3p miR-126a-5p miR-155 miR-188-3p miR-204 miR-218 miR-409-3p |
[36] | |||
| C57BL/6J mice induced with Streptozotocin (STZ) | Pancreatic tissue | Microarray | Up | miR-154-3p miR-296-3p miR-323-3p miR-491-5p miR-669m-3p miR-670-5p miR-697 miR-881-3p miR-3058-3p miR-5119 miR-5130 miR-5622-3p |
[109] | |
| Down | miR-7a-5p miR-7b-5p let-7a-5p let-7f-5p miR-10b-5p miR-16-5p miR-17-5p miR-26b-5p miR-28a-5p miR-28c miR-101a-3p miR-101b-3p miR-101c miR-126-3p miR-126-5p miR-143-3p miR-151-5p miR-184-3p miR-410-5p miR-451a miR-466 miR-467c-3p miR-467f miR-467g miR-467h miR-669 miR-1187 miR-3086-3p miR-5625-5p |
|||||
| Pancreatic islet | qPCR | Up | miR-21 | [108] | ||
| Full-thickness skin lesion | qPCR | Up | miR-29 | [110] | ||
| Down | miR-16 miR-21 miR-23a miR-24 miR-27b miR-31 miR-132 miR-195 miR-497 |
|||||
| Glomeruli | qPCR | Up | miR-145 | [44] | ||
| Heart | Microarray | Up | miR-21 miR-24 miR-142-3p miR-195 miR-199a-3p miR-208 miR-221 miR-499-3p miR-700 miR-705 |
[111] | ||
| Down | miR-1 miR-20a miR-29a miR-143 miR-220b miR-373 |
|||||
| FVB mice induced with STZ |
Heart | qPCR | Up | miR-107 miR-122 miR-125b-3p miR-134 miR-139-5p miR-141 miR-185 miR-193b miR-197 miR-200c miR-208-b miR-221 miR-222 miR-295 miR-298 miR-329 miR-346 miR-409-3p miR-431 miR-466g miR-467a miR-541 miR-542-5p miR-666-3p miR-702 miR-770-3p |
[78] | |
| Down | miR-302a miR-882 miR-883-5p |
|||||
| Rats | Sprague-Dawley or Wistar rats induced with STZ | Retina |
Microarray qPCR | Up | miR-29b miR-34 miR-203 miR-216 miR-410 |
[79] [80] |
| Down | miR-212 | |||||
| Kidney | qPCR | Up | miR-146a | [112] | ||
Table 3.
Altered miRNAs and their associated pathways in T1D.
| Group | Pathway | Upregulated miRNAs |
Downregulated miRNAs |
|---|---|---|---|
| Apoptosis | β-cell apoptosis | miR-21-5p [33,118] miR-24 [119] miR-34a [120] miR-155 [34,121] miR-181a-5p [8] miR-195 [81] miR-200a-3p [34,122] miR-375 [123] miR-424 [20] |
miR-100-5p [42] miR-126 [124] miR-146a-5p [34,40] miR-150-5p [40] miR-320a-3p [125] miR-324-5p [20,109] miR-342-3p [42] miR-424 [40] |
| TP53 signaling | miR-145 [126] |
miR-324-5p [20] miR-342-3p [20] miR-423-5p [20] |
|
| Wnt signaling | miR-143-3p [127] miR-144-5p [19] miR-148a-3p [33,34] miR-365a-3p [18] |
miR-140-3p [20] miR-324-5p [20] miR-342-3p [20] miR-766 [20] miR-940 [20] |
|
| TGF-β signaling | miR-26b [34,128] miR-382-5p [41] |
miR-10b-5p [129] | |
| mTOR signaling | miR-323-3p [109] | ||
| ER or oxidative stress |
FOXO signaling | miR-21-5p [33] miR-148a-3p [34] miR-323-3p [109] miR-486-5p [32,130] |
|
| NF-κB signaling | miR-24-3p [131,132] miR-155-5p [34] |
miR-146a-5p [34] miR-342 [133] |
|
| NRF2 | miR-21-5p [134] miR-144-5p [134] |
||
| Notch signaling | miR-140-3p [20] miR-146a-5p [34] miR-324-5p [20] miR-423-5p [20] miR-1275 [20] |
||
| Endocytosis | miR-21-5p [118,135] miR-29a-3p [34,136] |
miR-324-5p [20] miR-342-3p [20,42] |
|
| Immune system activation |
Immune system-related | miR-103a-3p [34] miR-155-5p [34] miR-200a-3p [34] |
|
| T-cell regulation | miR-31 [42] miR-342 [42] |
||
| Chemokine signaling [20] | miR-18b miR-20b miR-33a miR-101 miR-186 miR-338-3p |
miR-940 | |
| β-cell autoantigen release |
Jak-STAT signaling | miR-21-5p [8] miR-24-3p [8,105] miR-125b-5p [18,137] miR-181-5p [8] miR-323-3p [109] miR-210-5p [8] |
|
| MAPK signaling | miR-199a [20] miR-342 [34] miR-450a [20] miR-548c-3p [20] |
miR-100-5p [8] miR-150-5p [8] |
|
| β-cell insulin release |
Insulin signaling | miR-21 [20] miR-26b [37] miR-32 [20] miR-103a-3p [34] miR-143-3p [37] miR-148a [20] miR-155-5p [34] miR-200a-3p [34] miR-210-3p [34] miR-320c [138] miR-424 [20] miR-1225-5p [138] |
miR-29a-3p [139] miR-146a-5p [34] miR-324-5p [20] miR-342-3p [20] miR-423-5p [20] |
| Axon guidance [20] | miR-21 miR-26b miR-32 miR-126 miR-424 |
miR-766 miR-940 |
|
| Focal adhesion [20] | miR-7 miR-19a miR-27b miR-98 miR-148b miR-195 miR-301a miR-335 miR-454 miR-542-3p |
miR-1275 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



