Submitted:
20 February 2025
Posted:
21 February 2025
You are already at the latest version
Abstract
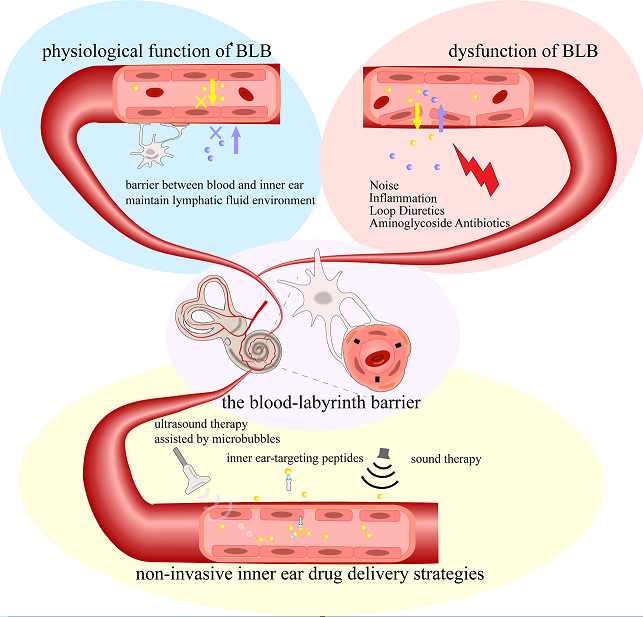
The inner ear is a relatively isolated organ, protected by the blood-labyrinth barrier (BLB). This barrier creates a unique lymphatic fluid environment within the inner ear, maintaining a stable physiological state essential for the mechano-electrical transduction process in the inner ear hair cells while simultaneously restricting most drugs from entering the lymphatic fluid. Under pathological conditions, dysfunction of the stria vascularis and disruption in barrier structure can lead to temporary or permanent hearing impairment. This review describes the structure and function of the BLB, along with recent advancements in modeling and protective studies related to the BLB. The review emphasizes some newly developed non-invasive inner ear drug delivery strategies, including sound therapy, ultrasound therapy assisted by microbubbles, inner ear-targeting peptides and route of administration cerebrospinal fluid conduit. We argue that some intrinsic properties of the BLB can be strategically utilized for effective inner ear drug delivery.
Keywords:
1. Introduction
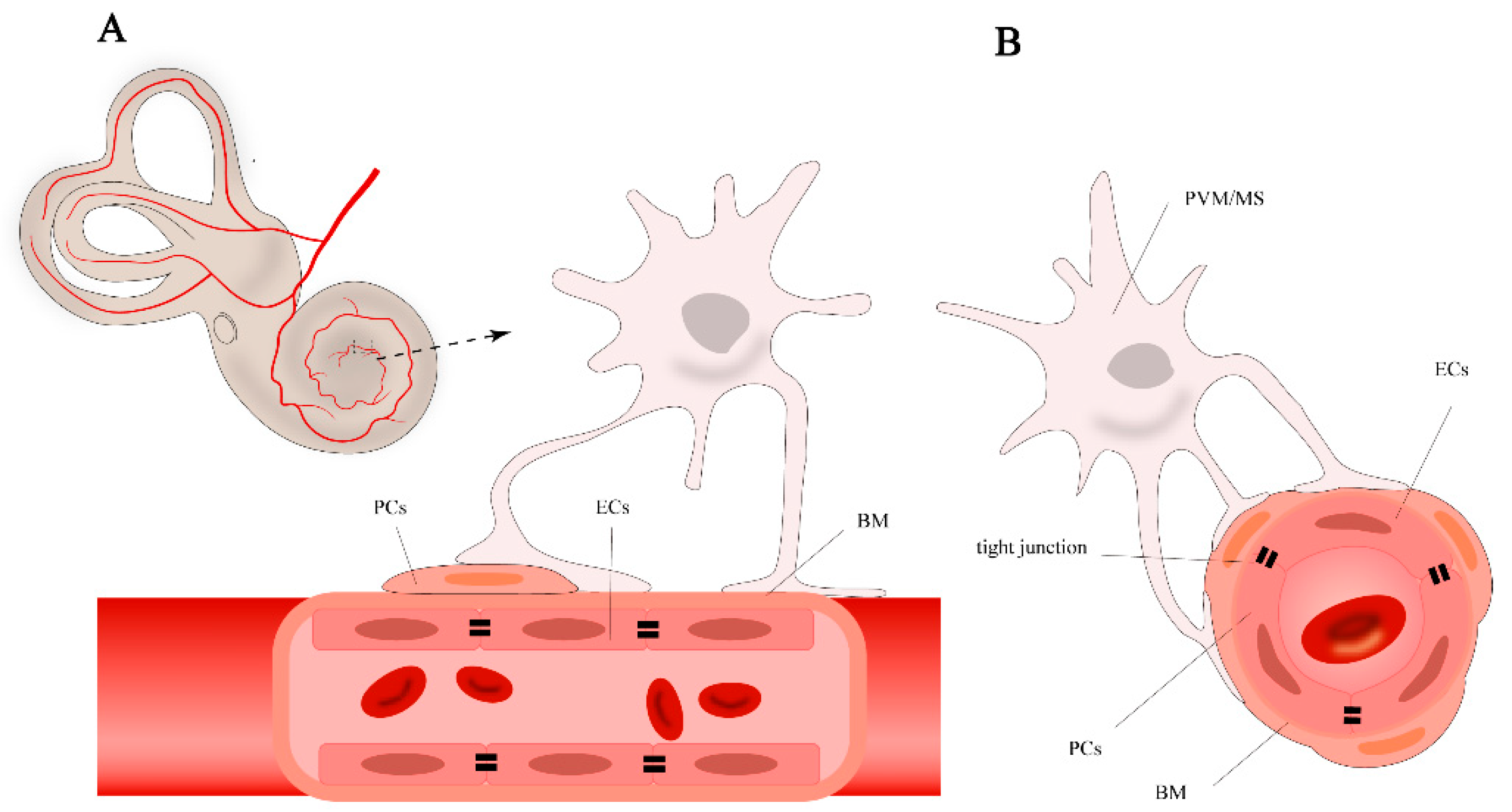
2. Stria Vascularis and Blood-Labyrinth Barrier
2.1. Cell Components of BLB
2.2. Cells in Stria Vascularis
3. Stria Vascularis in K+ Circulation and EP Generation
3.1. Pathway of Potassium Ion Circulation
3.2. Generation of Endolymphatic Potential
4. Selective Permeability of the Blood-Labyrinth Barrier
5. Inner Ear as a Former Assumed Immunoprivileged Organ
6. Pathological Mechanisms of Blood-Labyrinth Barrier Dysfunction
6.1. Loop Diuretics
6.2. Cisplatin
6.3. Aminoglycoside Antibiotics
6.4. Genetic Mutations
6.5. Acoustic Trauma
6.6. Inflammatory Responses
7. In Vivo and In Vitro Models for Studying the Blood-Labyrinth Barrier
7.1. In vivo Models
7.2. In Vitro Models
8. Current Inner Ear Drug Delivery Strategies
9. Innovative Approaches to Penetrate the BLB
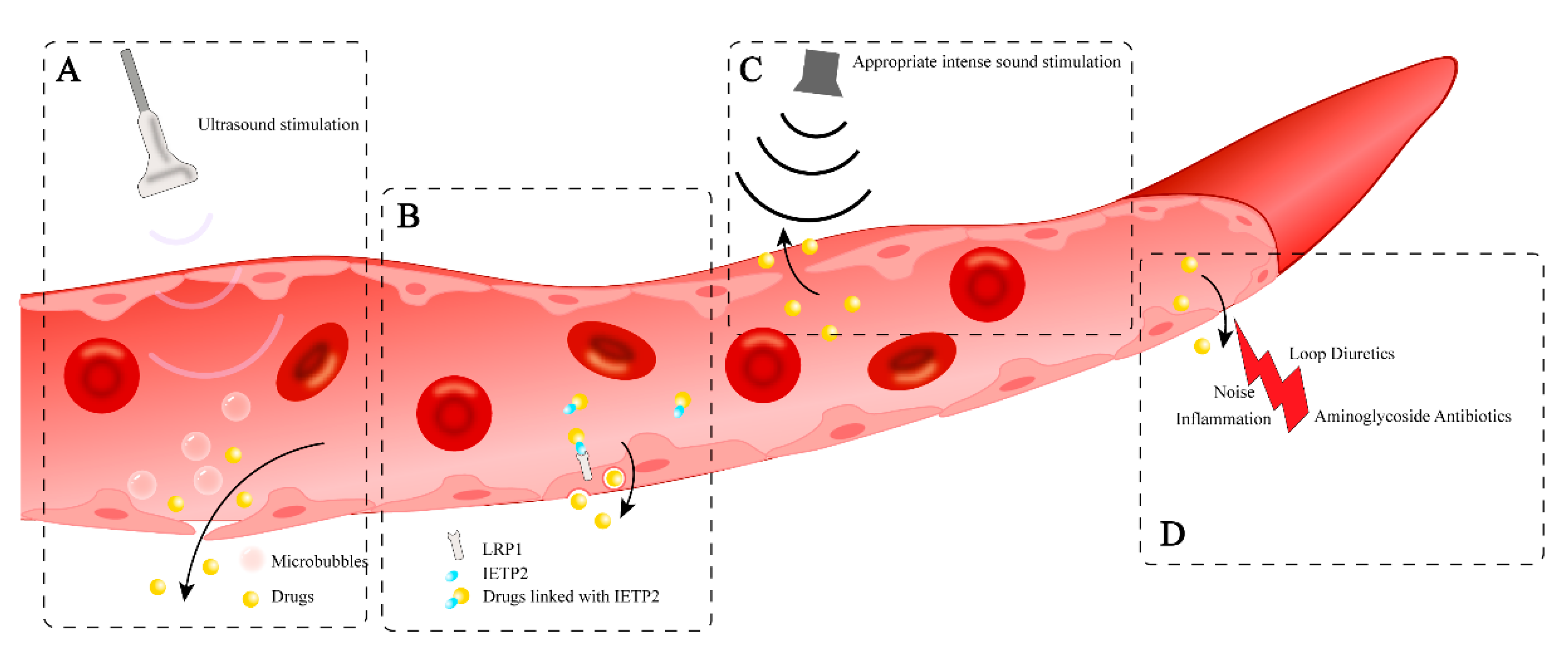
9.1. Drug-Induced Permeability Change of BLB
9.2. Ultrasound Therapy Assisted by Microbubbles
9.3. Inner Ear-Targeting Peptides
9.4. Sound Therapy
9.5. Route of Cerebrospinal Fluid Conduit
10. Strategies to Protect and Regulate Blood-Labyrinth Barrier Function
10.1. Gene Therapy
10.2. Rescue the Injured Cell of BLB
11. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| BLB | Blood-labyrinth barrier |
| BM | Basement membrane |
| CSF | Cerebrospinal fluid |
| HRP | Horseradish peroxidase |
| EP | Endocochlear potential |
| SV | Stria vascularis |
| PCs | Pericytes |
| PVM/Ms | Perivascular macrophage-like melanocytes |
| ECs | Endothelial cells |
| MCs | Marginal cells |
| ICs | Intermediate cells |
| BCs | Basal cells |
| TMPA | Trimethylphenylammonium |
| BBB | Blood-brain barrier |
| BRB | Blood-retinal barrier |
| LRP1 | Low-density lipoprotein receptor-related protein 1 |
| MSCs | Mesenchymal stem cells |
| AAV | Adeno-associated virus |
| IETP2 | Inner ear targeting peptide 2 |
| VEGF | Vascular endothelial growth factor |
| TNF-α | Tumor necrosis factor-alpha |
| HIF-1α | Hypoxia-inducible factor-1α |
References
- Schnieder, E.-A. A Contribution to the Physiology of the Perilymph Part I: The Origins of Perilymph. Ann Otol Rhinol Laryngol 1974, 83, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Jahnke, K.; Gorgas, K. The Permeability of Blood Vessels in the Guinea Pig Cochlea. I. Vessels of the Modiolus and Spiral Vessel. Anat Embryol (Berl) 1974, 146, 21–31. [Google Scholar] [CrossRef]
- Jahnke, K. The Blood-Perilymph Barrier. Arch Otorhinolaryngol 1980, 228, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Juhn, S.K.; Rybak, L.P. Labyrinthine Barriers and Cochlear Homeostasis. Acta Otolaryngol 1981, 91, 529–534. [Google Scholar] [CrossRef]
- Juhn, S.K.; Rybak, L.P.; Prado, S. Nature of Blood-Labyrinth Barrier in Experimental Conditions. Ann Otol Rhinol Laryngol 1981, 90, 135–141. [Google Scholar] [CrossRef]
- Juhn, S.K. Barrier Systems in the Inner Ear. Acta Oto-Laryngologica 1988, 105, 79–83. [Google Scholar] [CrossRef]
- Chen, J.; Zhao, H.-B. The Role of an Inwardly Rectifying K+ Channel (Kir4.1) in the Inner Ear and Hearing Loss. Neuroscience 2014, 265, 137–146. [Google Scholar] [CrossRef]
- Shi, X.; Nuttall, A.L. Upregulated iNOS and Oxidative Damage to the Cochlear Stria Vascularis Due to Noise Stress. Brain Res 2003, 967, 1–10. [Google Scholar] [CrossRef]
- Ohlemiller, K.K.; Rice, M.E.R.; Lett, J.M.; Gagnon, P.M. Absence of Strial Melanin Coincides with Age-Associated Marginal Cell Loss and Endocochlear Potential Decline. Hear Res 2009, 249, 1–14. [Google Scholar] [CrossRef]
- Zhou, X.-X.; Chen, S.; Xie, L.; Ji, Y.-Z.; Wu, X.; Wang, W.-W.; Yang, Q.; Yu, J.-T.; Sun, Y.; Lin, X.; et al. Reduced Connexin26 in the Mature Cochlea Increases Susceptibility to Noise-Induced Hearing Loss in Mice. Int J Mol Sci 2016, 17, 301. [Google Scholar] [CrossRef]
- Jabba, S.V.; Oelke, A.; Singh, R.; Maganti, R.J.; Fleming, S.; Wall, S.M.; Everett, L.A.; Green, E.D.; Wangemann, P. Macrophage Invasion Contributes to Degeneration of Stria Vascularis in Pendred Syndrome Mouse Model. BMC Med 2006, 4, 37. [Google Scholar] [CrossRef] [PubMed]
- Nyberg, S.; Abbott, N.J.; Shi, X.; Steyger, P.S.; Dabdoub, A. Delivery of Therapeutics to the Inner Ear: The Challenge of the Blood-Labyrinth Barrier. Science Translational Medicine 2019, 11, eaao0935. [Google Scholar] [CrossRef] [PubMed]
- Ishiyama, G.; Lopez, I.A.; Ishiyama, P.; Vinters, H.V.; Ishiyama, A. The Blood Labyrinthine Barrier in the Human Normal and Meniere’s Disease Macula Utricle. Sci Rep 2017, 7, 253. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Wang, Z.; Ren, W.; Chen, L.; Xu, C.; Li, M.; Fan, S.; Xu, Y.; Chen, M.; Zheng, F.; et al. LDL Receptor-Related Protein 1 (LRP1), a Novel Target for Opening the Blood-Labyrinth Barrier (BLB). Sig Transduct Target Ther 2022, 7, 1–12. [Google Scholar] [CrossRef]
- Sun, W.; Wang, W. Advances in Research on Labyrinth Membranous Barriers. Journal of Otology 2015, 10, 99–104. [Google Scholar] [CrossRef]
- Suzuki, M.; Yamasoba, T.; Ishibashi, T.; Miller, J.M.; Kaga, K. Effect of Noise Exposure on Blood-Labyrinth Barrier in Guinea Pigs. Hear Res 2002, 164, 12–18. [Google Scholar] [CrossRef]
- Suzuki, M.; Yamasoba, T.; Kaga, K. Development of the Blood-Labyrinth Barrier in the Rat. Hearing Research 1998, 116, 107–112. [Google Scholar] [CrossRef]
- Shi, X. Pathophysiology of the Cochlear Intrastrial Fluid-Blood Barrier (Review). Hearing Research 2016, 338, 52–63. [Google Scholar] [CrossRef]
- Zhang, F.; Dai, M.; Neng, L.; Zhang, J.H.; Zhi, Z.; Fridberger, A.; Shi, X. Perivascular Macrophage-like Melanocyte Responsiveness to Acoustic Trauma—a Salient Feature of Strial Barrier Associated Hearing Loss. FASEB J 2013, 27, 3730–3740. [Google Scholar] [CrossRef]
- Cosentino, A.; Agafonova, A.; Modafferi, S.; Salinaro, A.T.; Scuto, M.; Maiolino, L.; Fritsch, T.; Calabrese, E.J.; Lupo, G.; Anfuso, C.D.; et al. Blood–Labyrinth Barrier in Health and Diseases: Effect of Hormetic Nutrients. Antioxidants & Redox Signaling 2024. [Google Scholar] [CrossRef]
- Neng, L.; Zhang, F.; Kachelmeier, A.; Shi, X. Endothelial Cell, Pericyte, and Perivascular Resident Macrophage-Type Melanocyte Interactions Regulate Cochlear Intrastrial Fluid-Blood Barrier Permeability. J Assoc Res Otolaryngol 2013, 14, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Pardridge, W.M. The Blood-Brain Barrier: Bottleneck in Brain Drug Development. NeuroRx 2005, 2, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.-Z.; Le, Y.-Z. Significance of Outer Blood–Retina Barrier Breakdown in Diabetes and Ischemia. Investigative Ophthalmology & Visual Science 2011, 52, 2160–2164. [Google Scholar] [CrossRef]
- Shi, X.; Han, W.; Yamamoto, H.; Tang, W.; Lin, X.; Xiu, R.; Trune, D.R.; Nuttall, A.L. The Cochlear Pericytes. Microcirculation 2008, 15, 515–529. [Google Scholar] [CrossRef]
- Zhang, W.; Dai, M.; Fridberger, A.; Hassan, A.; Degagne, J.; Neng, L.; Zhang, F.; He, W.; Ren, T.; Trune, D.; et al. Perivascular-Resident Macrophage-like Melanocytes in the Inner Ear Are Essential for the Integrity of the Intrastrial Fluid-Blood Barrier. Proc Natl Acad Sci U S A 2012, 109, 10388–10393. [Google Scholar] [CrossRef] [PubMed]
- Sakagami, M.; Sano, M.; Tamaki, H.; Matsunaga, T. Ultrastructural Study of the Effect of Acute Hyper- and Hypotension on the Stria Vascularis and Spiral Ligament. Acta Oto-Laryngologica 1983, 96, 256–262. [Google Scholar] [CrossRef]
- Gow, A.; Davies, C.; Southwood, C.M.; Frolenkov, G.; Chrustowski, M.; Ng, L.; Yamauchi, D.; Marcus, D.C.; Kachar, B. Deafness in Claudin 11-Null Mice Reveals the Critical Contribution of Basal Cell Tight Junctions to Stria Vascularis Function. J Neurosci 2004, 24, 7051–7062. [Google Scholar] [CrossRef]
- Trowe, M.-O.; Maier, H.; Petry, M.; Schweizer, M.; Schuster-Gossler, K.; Kispert, A. Impaired Stria Vascularis Integrity upon Loss of E-Cadherin in Basal Cells. Developmental Biology 2011, 359, 95–107. [Google Scholar] [CrossRef]
- Steel, K.P.; Barkway, C. Another Role for Melanocytes: Their Importance for Normal Stria Vascularis Development in the Mammalian Inner Ear. Development 1989, 107, 453–463. [Google Scholar] [CrossRef]
- Wangemann, P. Comparison of Ion Transport Mechanisms between Vestibular Dark Cells and Strial Marginal Cells. Hearing Research 1995, 90, 149–157. [Google Scholar] [CrossRef]
- Patuzzi, R. Ion Flow in Stria Vascularis and the Production and Regulation of Cochlear Endolymph and the Endolymphatic Potential. Hearing Research 2011, 277, 4–19. [Google Scholar] [CrossRef] [PubMed]
- Renauld, J.M.; Khan, V.; Basch, M.L. Intermediate Cells of Dual Embryonic Origin Follow a Basal to Apical Gradient of Ingression Into the Lateral Wall of the Cochlea. Front. Cell Dev. Biol. 2022, 10, 867153. [Google Scholar] [CrossRef] [PubMed]
- Barrenäs, M.-L.; Axelsson, A. The Development of Melanin in the Stria Vascularis of the Gerbil. Acta Oto-Laryngologica 1992. [Google Scholar] [CrossRef]
- Ando, M.; Takeuchi, S. Immunological Identification of an Inward Rectifier K+ Channel (Kir4.1) in the Intermediate Cell (Melanocyte) of the Cochlear Stria Vascularis of Gerbils and Rats. Cell Tissue Res 1999, 298, 179–183. [Google Scholar] [CrossRef]
- Kikuchi, T.; Kimura, R.S.; Paul, D.L.; Takasaka, T.; Adams, J.C. Gap Junction Systems in the Mammalian Cochlea. Brain Research Reviews 2000, 32, 163–166. [Google Scholar] [CrossRef]
- Kuijpers, W.; Bonting, S.L. The Cochlear Potentials: II. The Nature of the Cochlear Endolymphatic Resting Potential. Pflugers Arch. 1970, 320, 359–372. [Google Scholar] [CrossRef]
- Nin, F.; Hibino, H.; Doi, K.; Suzuki, T.; Hisa, Y.; Kurachi, Y. The Endocochlear Potential Depends on Two K+ Diffusion Potentials and an Electrical Barrier in the Stria Vascularis of the Inner Ear. Proc Natl Acad Sci U S A 2008, 105, 1751–1756. [Google Scholar] [CrossRef]
- Patuzzi, R. Ion Flow in Cochlear Hair Cells and the Regulation of Hearing Sensitivity. Hearing Research 2011, 280, 3–20. [Google Scholar] [CrossRef]
- Wangemann, P. K+ Cycling and the Endocochlear Potential. Hearing Research 2002, 165, 1–9. [Google Scholar] [CrossRef]
- Jagger, D.J.; Nevill, G.; Forge, A. The Membrane Properties of Cochlear Root Cells Are Consistent with Roles in Potassium Recirculation and Spatial Buffering. JARO 2010, 11, 435–448. [Google Scholar] [CrossRef]
- Yoshida, T.; Sawamura, S.; Ota, T.; Higuchi, T.; Ogata, G.; Hori, K.; Nakagawa, T.; Doi, K.; Sato, M.; Nonomura, Y.; et al. Fibrocytes in the Cochlea of the Mammalian Inner Ear: Their Molecular Architecture, Physiological Properties, and Pathological Relevance. MRAJ 2017, 5. [Google Scholar] [CrossRef]
- Bovee, S.; Klump, G.M.; Köppl, C.; Pyott, S.J. The Stria Vascularis: Renewed Attention on a Key Player in Age-Related Hearing Loss. International Journal of Molecular Sciences 2024, 25, 5391. [Google Scholar] [CrossRef] [PubMed]
- Nin, F.; Hibino, H.; Doi, K.; Suzuki, T.; Hisa, Y.; Kurachi, Y. The Endocochlear Potential Depends on Two K+ Diffusion Potentials and an Electrical Barrier in the Stria Vascularis of the Inner Ear. Proceedings of the National Academy of Sciences 2008, 105, 1751–1756. [Google Scholar] [CrossRef] [PubMed]
- Wilms, V.; Köppl, C.; Söffgen, C.; Hartmann, A.-M.; Nothwang, H.G. Molecular Bases of K+ Secretory Cells in the Inner Ear: Shared and Distinct Features between Birds and Mammals. Sci Rep 2016, 6, 1–13. [Google Scholar] [CrossRef]
- Inamura, N.; Salt, A.N. Permeability Changes of the Blood-Labyrinth Barrier Measured in Vivo during Experimental Treatments. Hearing Research 1992, 61, 12–18. [Google Scholar] [CrossRef]
- Laurell, G.F.E.; Teixeira, M.; Duan, M.; Sterkers, O.; Ferrary, E. Intact Blood–Perilymph Barrier in the Rat after Impulse Noise Trauma. Acta Oto-Laryngologica 2008, 128, 608–612. [Google Scholar] [CrossRef]
- Schmitz, H.M.; Johnson, S.B.; Santi, P.A. Kanamycin-Furosemide Ototoxicity in the Mouse Cochlea: A 3-Dimensional Analysis. Otolaryngology–Head and Neck Surgery 2014, 150, 666–672. [Google Scholar] [CrossRef]
- Angelini, E.; Teixeira, M.; Aran, J.-M.; Ferrary, E. Taurine Entry into Perilymph of the Guinea Pig. European Archives of Oto-Rhino-Laryngology 1998, 255, 331–333. [Google Scholar] [CrossRef]
- Suzuki, M.; Kitamura, K.; Nomura, Y. Anionic Sites of the Basement Membrane of the Labyrinth. Acta Oto-Laryngologica 1991, 111, 112–115. [Google Scholar] [CrossRef]
- Suzuki, M.; Kitamura, K.; Nomura, Y. Influence of Changed Blood pH on Anionic Sites in the Labyrinth. Acta Oto-Laryngologica 1995, 115, 747–753. [Google Scholar] [CrossRef]
- Suzuki, M.; Kaga, K. Effect of Cisplatin on the Negative Charge Barrier in Strial Vessels of the Guinea Pig A Transmission Electron Microscopic Study Using Polyethyleneimine Molecules. Eur Arch Otorhinolaryngol 1996, 253, 351–355. [Google Scholar] [CrossRef]
- Spadoni, I.; Fornasa, G.; Rescigno, M. Organ-Specific Protection Mediated by Cooperation between Vascular and Epithelial Barriers. Nat Rev Immunol 2017, 17, 761–773. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.P. Immunology of the Inner Ear: Evidence of Local Antibody Production. Ann Otol Rhinol Laryngol 1984, 93, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.P. Immunology of the Inner Ear: Response of the Inner Ear to Antigen Challenge. Otolaryngol Head Neck Surg 1983, 91, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.; Ryan, A. Fundamental Immune Mechanisms of the Brain and Inner Ear. Head and Neck Surgery 1995. [Google Scholar]
- Woolf, N.K.; Harris, J.P. Cochlear Pathophysiology Associated with Inner Ear Immune Responses. Acta Oto-Laryngologica 1986. [Google Scholar] [CrossRef]
- Fujioka, M.; Okano, H.; Ogawa, K. Inflammatory and Immune Responses in the Cochlea: Potential Therapeutic Targets for Sensorineural Hearing Loss. Front. Pharmacol. 2014, 5, 128290. [Google Scholar] [CrossRef]
- Okano, T.; Nakagawa, T.; Kita, T.; Kada, S.; Yoshimoto, M.; Nakahata, T.; Ito, J. Bone Marrow-Derived Cells Expressing Iba1 Are Constitutively Present as Resident Tissue Macrophages in the Mouse Cochlea. Journal of Neuroscience Research 2008, 86, 1758–1767. [Google Scholar] [CrossRef]
- Hirose, K.; Discolo, C.M.; Keasler, J.R.; Ransohoff, R. Mononuclear Phagocytes Migrate into the Murine Cochlea after Acoustic Trauma. Journal of Comparative Neurology 2005, 489, 180–194. [Google Scholar] [CrossRef]
- Hu, B.H.; Zhang, C.; Frye, M.D. Immune Cells and Non-Immune Cells with Immune Function in Mammalian Cochleae. Hearing Research 2018, 362, 14–24. [Google Scholar] [CrossRef]
- Du, X.; Choi, C.-H.; Chen, K.; Cheng, W.; Floyd, R.A.; Kopke, R.D. Reduced Formation of Oxidative Stress Biomarkers and Migration of Mononuclear Phagocytes in the Cochleae of Chinchilla after Antioxidant Treatment in Acute Acoustic Trauma. International Journal of Otolaryngology 2011, 2011, 1–13. [Google Scholar] [CrossRef]
- Fredelius, L.; Rask-Andersen, H. The Role of Macrophages in the Disposal of Degeneration Products within the Organ of Corti after Acoustic Overstimulation. Acta Oto-Laryngologica 1990, 109, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Corey, D.P.; Akyuz, N.; Holt, J.R. Function and Dysfunction of TMC Channels in Inner Ear Hair Cells. Csh Perspect Med 2019, 9, a033506. [Google Scholar] [CrossRef] [PubMed]
- Reynard, P.; Thai-Van, H. Drug-Induced Hearing Loss: Listening to the Latest Advances. Therapies 2024, 79, 283–295. [Google Scholar] [CrossRef]
- Tn, L.; Lv, S.; J, L.; B, W. Current Insights in Noise-Induced Hearing Loss: A Literature Review of the Underlying Mechanism, Pathophysiology, Asymmetry, and Management Options. Journal of otolaryngology - head & neck surgery = Le Journal d’oto-rhino-laryngologie et de chirurgie cervico-faciale 2017, 46. [Google Scholar] [CrossRef]
- Xu, K.; Chen, S.; Xie, L.; Qiu, Y.; Bai, X.; Liu, X.-Z.; Zhang, H.-M.; Wang, X.-H.; Jin, Y.; Sun, Y.; et al. Local Macrophage-Related Immune Response Is Involved in Cochlear Epithelial Damage in Distinct Gjb2-Related Hereditary Deafness Models. Front Cell Dev Biol 2021, 8. [Google Scholar] [CrossRef]
- Cannon, P.J.; Kilcoyne, M.M. Ethacrynic Acid and Furosemide: Renal Pharmacology and Clinical Use. Progress in Cardiovascular Diseases 1969, 12, 99–118. [Google Scholar] [CrossRef]
- Ding, D.; Liu, H.; Qi, W.; Jiang, H.; Li, Y.; Wu, X.; Sun, H.; Gross, K.; Salvi, R. Ototoxic Effects and Mechanisms of Loop Diuretics. Journal of Otology 2016, 11, 145–156. [Google Scholar] [CrossRef]
- Tang, Q.; Wang, X.; Jin, H.; Mi, Y.; Liu, L.; Dong, M.; Chen, Y.; Zou, Z. Cisplatin-Induced Ototoxicity: Updates on Molecular Mechanisms and Otoprotective Strategies. European Journal of Pharmaceutics and Biopharmaceutics 2021, 163, 60–71. [Google Scholar] [CrossRef]
- Tan, W.J.T.; Vlajkovic, S.M. Molecular Characteristics of Cisplatin-Induced Ototoxicity and Therapeutic Interventions. International Journal of Molecular Sciences 2023, 24, 16545. [Google Scholar] [CrossRef]
- So, H.; Kim, H.; Lee, J.-H.; Park, C.; Kim, Y.; Kim, E.; Kim, J.-K.; Yun, K.-J.; Lee, K.-M.; Lee, H.-Y.; et al. Cisplatin Cytotoxicity of Auditory Cells Requires Secretions of Proinflammatory Cytokines via Activation of ERK and NF-kappaB. J Assoc Res Otolaryngol 2007, 8, 338–355. [Google Scholar] [CrossRef]
- Zhang, N.; Cai, J.; Xu, L.; Wang, H.; Liu, W. Cisplatin-Induced Stria Vascularis Damage Is Associated with Inflammation and Fibrosis. Neural Plasticity 2020, 2020, 8851525. [Google Scholar] [CrossRef] [PubMed]
- Anfuso, C.D.; Cosentino, A.; Agafonova, A.; Zappalà, A.; Giurdanella, G.; Trovato Salinaro, A.; Calabrese, V.; Lupo, G. Pericytes of Stria Vascularis Are Targets of Cisplatin-Induced Ototoxicity: New Insights into the Molecular Mechanisms Involved in Blood-Labyrinth Barrier Breakdown. International Journal of Molecular Sciences 2022, 23, 15790. [Google Scholar] [CrossRef] [PubMed]
- Guthrie, O.W. Aminoglycoside Induced Ototoxicity. Toxicology 2008, 249, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Forge, A.; Fradis, M. Structural Abnormalities in the Stria Vascularis Following Chronic Gentamicin Treatment. Hearing Research 1985, 20, 233–244. [Google Scholar] [CrossRef]
- Wang, Q.; Steyger, P.S. Trafficking of Systemic Fluorescent Gentamicin into the Cochlea and Hair Cells. JARO: Journal of the Association for Research in Otolaryngology 2009, 10, 205. [Google Scholar] [CrossRef]
- Stepanyan, R.S.; Indzhykulian, A.A.; Vélez-Ortega, A.C.; Boger, E.T.; Steyger, P.S.; Friedman, T.B.; Frolenkov, G.I. TRPA1-Mediated Accumulation of Aminoglycosides in Mouse Cochlear Outer Hair Cells. JARO 2011, 12, 729–740. [Google Scholar] [CrossRef]
- Ding, D.; McFadden, S.L.; Browne, R.W.; Salvi, R.J. Late Dosing with Ethacrynic Acid Can Reduce Gentamicin Concentration in Perilymph and Protect Cochlear Hair Cells. Hearing Res 2003, 185, 90–96. [Google Scholar] [CrossRef]
- Didier, A.; Miller, J.M.; Nuttall, A.L. The Vascular Component of Sodium Salicylate Ototoxicity in the Guinea Pig. Hearing Research 1993, 69, 199–206. [Google Scholar] [CrossRef]
- Smith, D.I.; lawrence, M.; Hawkins, J.E. Effects of Noise and Quinine on the Vessels of the Stria Vascularis: An Image Analysis Study. American Journal of Otolaryngology 1985, 6, 280–289. [Google Scholar] [CrossRef]
- Cohen-Salmon, M.; Regnault, B.; Cayet, N.; Caille, D.; Demuth, K.; Hardelin, J.-P.; Janel, N.; Meda, P.; Petit, C. Connexin30 Deficiency Causes Instrastrial Fluid-Blood Barrier Disruption within the Cochlear Stria Vascularis. Proc Natl Acad Sci U S A 2007, 104, 6229–6234. [Google Scholar] [CrossRef]
- Zhou, X.-X.; Chen, S.; Xie, L.; Ji, Y.-Z.; Wu, X.; Wang, W.-W.; Yang, Q.; Yu, J.-T.; Sun, Y.; Lin, X.; et al. Reduced Connexin26 in the Mature Cochlea Increases Susceptibility to Noise-Induced Hearing Loss in Mice. International Journal of Molecular Sciences 2016, 17, 301. [Google Scholar] [CrossRef] [PubMed]
- Wangemann, P.; Itza, E.M.; Albrecht, B.; Wu, T.; Jabba, S.V.; Maganti, R.J.; Lee, J.H.; Everett, L.A.; Wall, S.M.; Royaux, I.E.; et al. Loss of KCNJ10 Protein Expression Abolishes Endocochlear Potential and Causes Deafness in Pendred Syndrome Mouse Model. BMC Med 2004, 2, 30. [Google Scholar] [CrossRef] [PubMed]
- Rehm, H.L.; Zhang, D.-S.; Brown, M.C.; Burgess, B.; Halpin, C.; Berger, W.; Morton, C.C.; Corey, D.P.; Chen, Z.-Y. Vascular Defects and Sensorineural Deafness in a Mouse Model of Norrie Disease. J. Neurosci. 2002, 22, 4286–4292. [Google Scholar] [CrossRef] [PubMed]
- Kruegel, J.; Rubel, D.; Gross, O. Alport Syndrome—Insights from Basic and Clinical Research. Nat Rev Nephrol 2013, 9, 170–178. [Google Scholar] [CrossRef]
- Cable, J.; Jackson, I.J.; Steel, K.P. Light (B), a Mutation That Causes Melanocyte Death, Affects Stria Vascularis Function in the Mouse Inner Ear. Pigment Cell Research 1993, 6, 215–225. [Google Scholar] [CrossRef]
- Kitamura, K.; Sakagami, M.; Umemoto, M.; Takeda, N.; Doi, K.; Kasugai, T.; Kitamura, Y. Strial Dysfunction in a Melanocyte Deficient Mutant Rat (Ws/Ws Rat). Acta Oto-Laryngologica 1994. [Google Scholar] [CrossRef]
- Chen, J.; Nathans, J. Estrogen-Related Receptor β/NR3B2 Controls Epithelial Cell Fate and Endolymph Production by the Stria Vascularis. Developmental Cell 2007, 13, 325–337. [Google Scholar] [CrossRef]
- Zong, Y.-J.; Liu, X.-Z.; Tu, L.; Sun, Y. Cytomembrane Trafficking Pathways of Connexin 26, 30, and 43. Int J Mol Sci 2023, 24, 10349. [Google Scholar] [CrossRef]
- Zhao, H.-B.; Kikuchi, T.; Ngezahayo, A.; White, T.W. Gap Junctions and Cochlear Homeostasis. J Membr Biol 2006, 209, 177–186. [Google Scholar] [CrossRef]
- Huang, L.; Bitner-Glindzicz, M.; Tranebjærg, L.; Tinker, A. A Spectrum of Functional Effects for Disease Causing Mutations in the Jervell and Lange-Nielsen Syndrome. Cardiovascular Research 2001, 51, 670–680. [Google Scholar] [CrossRef]
- Chang, Q.; Wang, J.; Li, Q.; Kim, Y.; Zhou, B.; Wang, Y.; Li, H.; Lin, X. Virally Mediated Kcnq1 Gene Replacement Therapy in the Immature Scala Media Restores Hearing in a Mouse Model of Human Jervell and Lange-Nielsen Deafness Syndrome. EMBO Molecular Medicine 2015. [Google Scholar] [CrossRef] [PubMed]
- Seidman, M.D.; Quirk, W.S.; Shirwany, N.A. Mechanisms of Alterations in the Microcirculation of the Cochlea. Annals of the New York Academy of Sciences 1999, 884, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Shi, X. Physiopathology of the Cochlear Microcirculation. Hearing Research 2011, 282, 10–24. [Google Scholar] [CrossRef]
- Shin, S.-A.; Lyu, A.-R.; Jeong, S.-H.; Kim, T.H.; Park, M.J.; Park, Y.-H. Acoustic Trauma Modulates Cochlear Blood Flow and Vasoactive Factors in a Rodent Model of Noise-Induced Hearing Loss. International Journal of Molecular Sciences 2019, 20, 5316. [Google Scholar] [CrossRef]
- Yang, Y.; Dai, M.; Wilson, T.M.; Omelchenko, I.; Klimek, J.E.; Wilmarth, P.A.; David, L.L.; Nuttall, A.L.; Gillespie, P.G.; Shi, X. Na+/K+-ATPase A1 Identified as an Abundant Protein in the Blood-Labyrinth Barrier That Plays an Essential Role in the Barrier Integrity. PLoS One 2011, 6, e16547. [Google Scholar] [CrossRef]
- Shi, X. Cochlear Pericyte Responses to Acoustic Trauma and the Involvement of Hypoxia-Inducible Factor-1α and Vascular Endothelial Growth Factor. The American Journal of Pathology 2009, 174, 1692–1704. [Google Scholar] [CrossRef]
- Wu, J.; Han, W.; Chen, X.; Guo, W.; Liu, K.; Wang, R.; Zhang, J.; Sai, N. Matrix Metalloproteinase-2 and -9 Contribute to Functional Integrity and Noise-induced Damage to the Blood-Labyrinth-Barrier. Molecular Medicine Reports 2017, 16, 1731–1738. [Google Scholar] [CrossRef]
- Hirose, K.; Liberman, M.C. Lateral Wall Histopathology and Endocochlear Potential in the Noise-Damaged Mouse Cochlea. JARO 2003, 4, 339–352. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, S.; Hou, Z.; Cai, J.; Dong, M.; Shi, X. Lipopolysaccharide-Induced Middle Ear Inflammation Disrupts the Cochlear Intra-Strial Fluid–Blood Barrier through Down-Regulation of Tight Junction Proteins. PLOS ONE 2015, 10, e0122572. [Google Scholar] [CrossRef]
- Abbott, N.J. Inflammatory Mediators and Modulation of Blood–Brain Barrier Permeability. Cell Mol Neurobiol 2000, 20, 131–147. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhang, J.; Rao, Y.; Chen, J.; Chen, K.; Tang, Y. Lipopolysaccharide Disrupts the Cochlear Blood-Labyrinth Barrier by Activating Perivascular Resident Macrophages and up-Regulating MMP-9. International Journal of Pediatric Otorhinolaryngology 2019, 127, 109656. [Google Scholar] [CrossRef]
- Ichimiya, I.; Yoshida, K.; Hirano, T.; Suzuki, M.; Mogi, G. Significance of Spiral Ligament Fibrocytes with Cochlear Inflammation. International Journal of Pediatric Otorhinolaryngology 2000, 56, 45–51. [Google Scholar] [CrossRef]
- Hirose, K.; Hartsock, J.J.; Johnson, S.; Santi, P.; Salt, A.N. Systemic Lipopolysaccharide Compromises the Blood-Labyrinth Barrier and Increases Entry of Serum Fluorescein into the Perilymph. J Assoc Res Otolaryngol 2014, 15, 707–719. [Google Scholar] [CrossRef]
- Soltesz, P.; Der, H.; Veres, K.; Laczik, R.; Sipka, S.; Szegedi, G.; Szodoray, P. Immunological Features of Primary Anti-Phospholipid Syndrome in Connection with Endothelial Dysfunction. Rheumatology 2008, 47, 1628–1634. [Google Scholar] [CrossRef] [PubMed]
- Thin and Open Vessel Windows for Intra-Vital Fluorescence Imaging of Murine Cochlear Blood Flow. Hearing Research 2014, 313, 38–46. [CrossRef]
- F, Z.; J, Z.; L, N.; X, S. Characterization and Inflammatory Response of Perivascular-Resident Macrophage-like Melanocytes in the Vestibular System. Journal of the Association for Research in Otolaryngology : JARO 2013, 14. [Google Scholar] [CrossRef]
- Neng, L.; Zhang, J.; Yang, J.; Zhang, F.; Lopez, I.A.; Dong, M.; Shi, X. Structural Changes in Thestrial Blood–Labyrinth Barrier of Aged C57BL/6 Mice. Cell Tissue Res 2015, 361, 685–696. [Google Scholar] [CrossRef]
- Z, H.; L, N.; J, Z.; J, C.; X, W.; Y, Z.; Ia, L.; X, S. Acoustic Trauma Causes Cochlear Pericyte-to-Myofibroblast-Like Cell Transformation and Vascular Degeneration, and Transplantation of New Pericytes Prevents Vascular Atrophy. The American journal of pathology 2020, 190. [Google Scholar] [CrossRef]
- Hou, Z.; Wang, X.; Cai, J.; Zhang, J.; Hassan, A.; Auer, M.; Shi, X. Platelet-Derived Growth Factor Subunit B Signaling Promotes Pericyte Migration in Response to Loud Sound in the Cochlear Stria Vascularis. JARO 2018, 19, 363–379. [Google Scholar] [CrossRef]
- S, D.; F, R.; D, M.; G, G.; L, P.; L, S. An in Vitro Blood-Brain Barrier Model: Cocultures between Endothelial Cells and Organotypic Brain Slice Cultures. Proceedings of the National Academy of Sciences of the United States of America 1998, 95. [Google Scholar] [CrossRef]
- Neng, L.; Zhang, W.; Hassan, A.; Zemla, M.; Kachelmeier, A.; Fridberger, A.; Auer, M.; Shi, X. Isolation and Culture of Endothelial Cells, Pericytes and Perivascular Resident Macrophage-like Melanocytes from the Young Mouse Ear. Nat Protoc 2013, 8, 709–720. [Google Scholar] [CrossRef] [PubMed]
- Sekulic-Jablanovic, M.; Paproth, J.; Sgambato, C.; Albano, G.; Fuster, D.G.; Bodmer, D.; Petkovic, V. Lack of NHE6 and Inhibition of NKCC1 Associated With Increased Permeability in Blood Labyrinth Barrier-Derived Endothelial Cell Layer. Front. Cell. Neurosci. 2022, 16, 862119. [Google Scholar] [CrossRef] [PubMed]
- Tm, C.; Eb, B.; J, R. Toward Three-Dimensional in Vitro Models to Study Neurovascular Unit Functions in Health and Disease. Neural regeneration research 2021, 16. [Google Scholar] [CrossRef]
- Neng, L.; Zhang, J.; Yang, J.; Zhang, F.; Lopez, I.A.; Dong, M.; Shi, X. Structural Changes in Thestrial Blood–Labyrinth Barrier of Aged C57BL/6 Mice. Cell Tissue Res 2015, 361, 685–696. [Google Scholar] [CrossRef]
- Sekulic, M.; Abdollahi, N.; Graf, L.; Deigendesch, N.; Puche, R.; Bodmer, D.; Petkovic, V. Human Blood-Labyrinth Barrier on a Chip: A Unique in Vitro Tool for Investigation of BLB Properties. RSC Adv. 2023, 13, 25508–25517. [Google Scholar] [CrossRef]
- Sekulic, M.; Puche, R.; Bodmer, D.; Petkovic, V. Human Blood-Labyrinth Barrier Model to Study the Effects of Cytokines and Inflammation. Front. Mol. Neurosci. 2023, 16. [Google Scholar] [CrossRef]
- Liu, H.; Hao, J.; Li, K.S. Current Strategies for Drug Delivery to the Inner Ear. Acta Pharmaceutica Sinica B 2013, 3, 86–96. [Google Scholar] [CrossRef]
- ERSNER, M.S.; SPIEGEL, E.A.; ALEXANDER, M.H. TRANSTYMPANIC INJECTION OF ANESTHETICS FOR THE TREATMENT OF MÉNIÈRE’S SYNDROME. A.M.A. Archives of Otolaryngology 1951, 54, 43–52. [Google Scholar] [CrossRef]
- Bowe, S.N.; Jacob, A. Round Window Perfusion Dynamics: Implications for Intracochlear Therapy. Current Opinion in Otolaryngology & Head and Neck Surgery 2010, 18, 377. [Google Scholar] [CrossRef]
- Sewell, W.F.; Borenstein, J.T.; Chen, Z.; Fiering, J.; Handzel, O.; Holmboe, M.; Kim, E.S.; Kujawa, S.G.; McKenna, M.J.; Mescher, M.M.; et al. Development of a Microfluidics-Based Intracochlear Drug Delivery Device. Audiology and Neurotology 2009, 14, 411–422. [Google Scholar] [CrossRef]
- Lv, J.; Wang, H.; Cheng, X.; Chen, Y.; Wang, D.; Zhang, L.; Cao, Q.; Tang, H.; Hu, S.; Gao, K.; et al. AAV1-hOTOF Gene Therapy for Autosomal Recessive Deafness 9: A Single-Arm Trial. The Lancet 2024, 403, 2317–2325. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, Y.; Currie, J.-C.; Demeule, M.; Régina, A.; Ché, C.; Abulrob, A.; Fatehi, D.; Sartelet, H.; Gabathuler, R.; Castaigne, J.-P.; et al. Transport Characteristics of a Novel Peptide Platform for CNS Therapeutics. Journal of Cellular and Molecular Medicine 2010, 14, 2827–2839. [Google Scholar] [CrossRef] [PubMed]
- Jh, P.; Y, K.; J, Y.; Jw, C. Antioxidant Therapy against Oxidative Damage of the Inner Ear: Protection and Preconditioning. Antioxidants (Basel, Switzerland) 2020, 9. [Google Scholar] [CrossRef]
- Sound Conditioning Reduces Noise-Induced Permanent Threshold Shift in Mice. Hearing Research 2000, 148, 213–219. [CrossRef]
- Wang, X.; Gu, J.; Xu, K.; Xu, B.; Yu, D.; Wu, H. Sound Conditioning Strategy Promoting Paracellular Permeability of the Blood-labyrinth-barrier Benefits Inner Ear Drug Delivery. Bioengineering & Transla Med 2024, 9, e10596. [Google Scholar] [CrossRef]
- He, Y.; Chen, Z.; Liu, Q.; Li, Z.; Wen, D.; Zhang, H.; Zhang, M.; Jiang, D.; Li, H.; Wen, L.; et al. Reversible Opening of the Blood-Labyrinth Barrier by Low-Pressure Pulsed Ultrasound and Microbubbles for the Treatment of Inner Ear Diseases. Journal of Controlled Release 2024, 372, 318–330. [Google Scholar] [CrossRef]
- Zhao, B.; Chen, Y.; Liu, J.; Zhang, L.; Wang, J.; Yang, Y.; Lv, Q.; Xie, M. Blood-Brain Barrier Disruption Induced by Diagnostic Ultrasound Combined with Microbubbles in Mice. Oncotarget 2017, 9, 4897–4914. [Google Scholar] [CrossRef]
- McMahon, D.; Poon, C.; Hynynen, K. Evaluating the Safety Profile of Focused Ultrasound and Microbubble-Mediated Treatments to Increase Blood-Brain Barrier Permeability. Expert Opinion on Drug Delivery 2019. [Google Scholar] [CrossRef]
- K. Mathiesen, B.; M. Miyakoshi, L.; R. Cederroth, C.; Tserga, E.; Versteegh, C.; A. R. Bork, P.; L. Hauglund, N.; Stefan Gomolka, R.; Mori, Y.; K. Edvall, N.; et al. Delivery of Gene Therapy through a Cerebrospinal Fluid Conduit to Rescue Hearing in Adult Mice. Science Translational Medicine 2023. [Google Scholar] [CrossRef]
- Valentini, C.; Szeto, B.; Kysar, J.W.; Lalwani, A.K. Inner Ear Gene Delivery: Vectors and Routes. Hearing, Balance and Communication 2020, 18, 278–285. [Google Scholar] [CrossRef]
- Zhang, J.; Hou, Z.; Wang, X.; Jiang, H.; Neng, L.; Zhang, Y.; Yu, Q.; Burwood, G.; Song, J.; Auer, M.; et al. VEGFA165 Gene Therapy Ameliorates Blood-Labyrinth Barrier Breakdown and Hearing Loss. JCI Insight 2021, 6. [Google Scholar] [CrossRef] [PubMed]
- O, M.; K, I.; Y, S.; T, O.; S, N.; Y, K.; M, S.; M, F.; T, K.; Y, Z.; et al. Altered Cochlear Fibrocytes in a Mouse Model of DFN3 Nonsyndromic Deafness. Science (New York, N.Y.) 1999, 285. [Google Scholar] [CrossRef]
- Lang, H.; Ebihara, Y.; Schmiedt, R.A.; Minamiguchi, H.; Zhou, D.; Smythe, N.; Liu, L.; Ogawa, M.; Schulte, B.A. Contribution of Bone Marrow Hematopoietic Stem Cells to Adult Mouse Inner Ear: Mesenchymal Cells and Fibrocytes. Journal of Comparative Neurology 2006, 496, 187–201. [Google Scholar] [CrossRef] [PubMed]
- K, K.; Y, F.; N, H.; Y, O.; H, K.; R, K.; R, K.; S, N.; H, S.; M, F.; et al. Mesenchymal Stem Cell Transplantation Accelerates Hearing Recovery through the Repair of Injured Cochlear Fibrocytes. The American journal of pathology 2007, 171. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



