Submitted:
15 February 2025
Posted:
18 February 2025
You are already at the latest version
Abstract
Transcription factors (TFs) are critical regulators of gene expression, playing key roles in 1 various biological processes such as growth, development, and stress responses in plants. While many TF families like MYB, bZIP, and WRKY have been identified and extensively studied in model plants, a substantial number of novel TFs remain uncharacterized, particularly in non-model and economically significant plant species. The discovery of these novel TFs offers a promising avenue for advancing our understanding of plant biology and improving crop traits. Recent advancements in high-throughput sequencing, genomics, and bioinformatics have facilitated the identification of previously overlooked or newly evolved TFs, expanding the repertoire of known plant transcriptional regulators. This review highlights the emergence of novel TF families and their functional roles in regulating plant growth, adaptation to abiotic and biotic stresses, and metabolic pathways. We examine the molecular mechanisms by which these TFs govern gene expression, their interactions with other signaling pathways, and their potential applications in crop improvement. In addition, we explore the challenges and opportunities associated with the functional characterization and validation of novel TFs, as well as the technologies that are enabling these discoveries. Special attention is given to the potential of novel TFs to enhance stress tolerance, disease resistance, and overall productivity in crops, with a focus on their integration into biotechnological approaches, such as genome editing and transgenic crop development. The review concludes by outlining future research directions and the potential impact of novel TFs in addressing global agricultural challenges, including climate change, food security, and sustainability.
Keywords:
1. Introduction
2. Novel Transcription Factor Families
2.1. GARP Family: New Insights into Photosynthesis Regulation
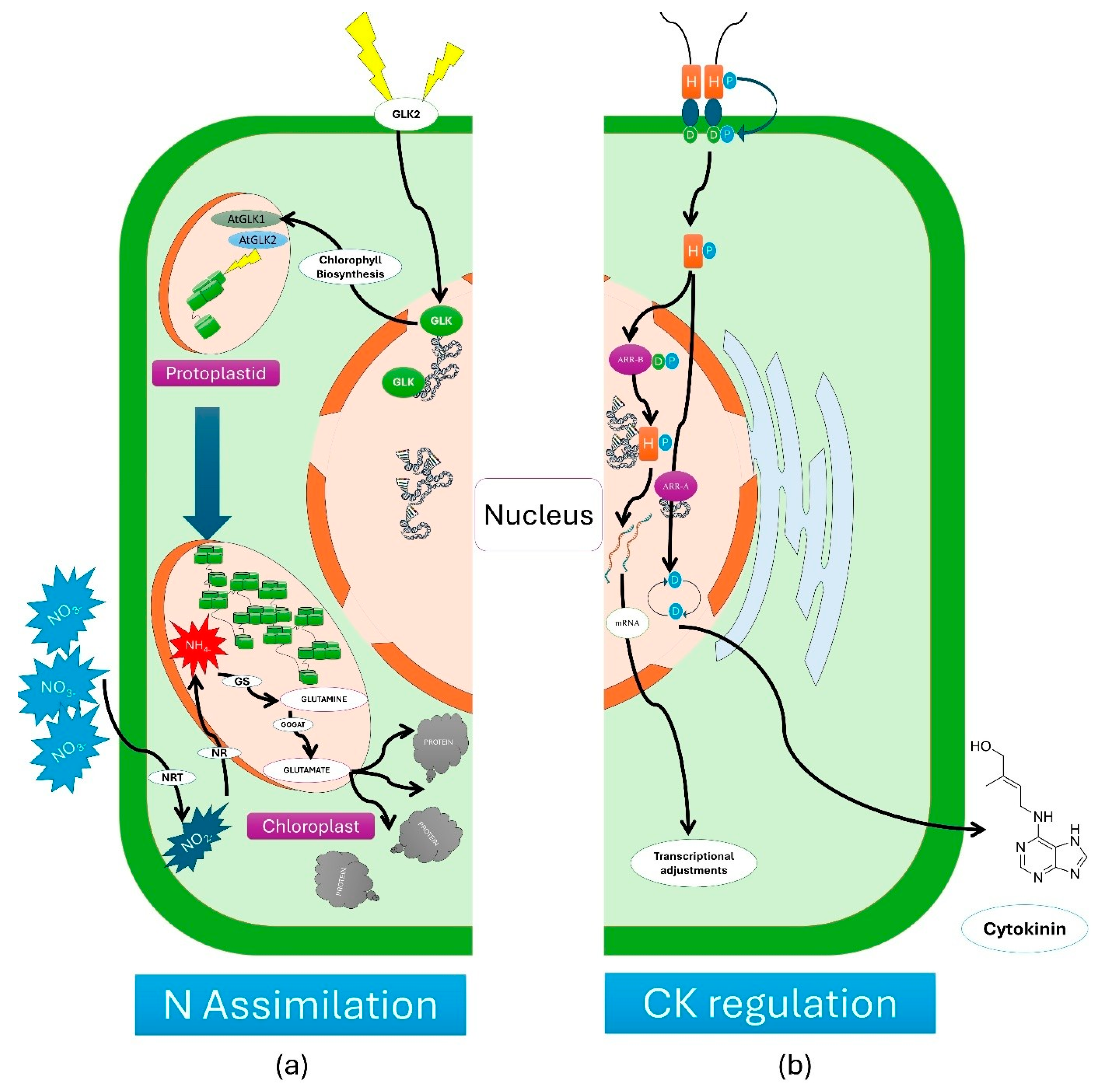
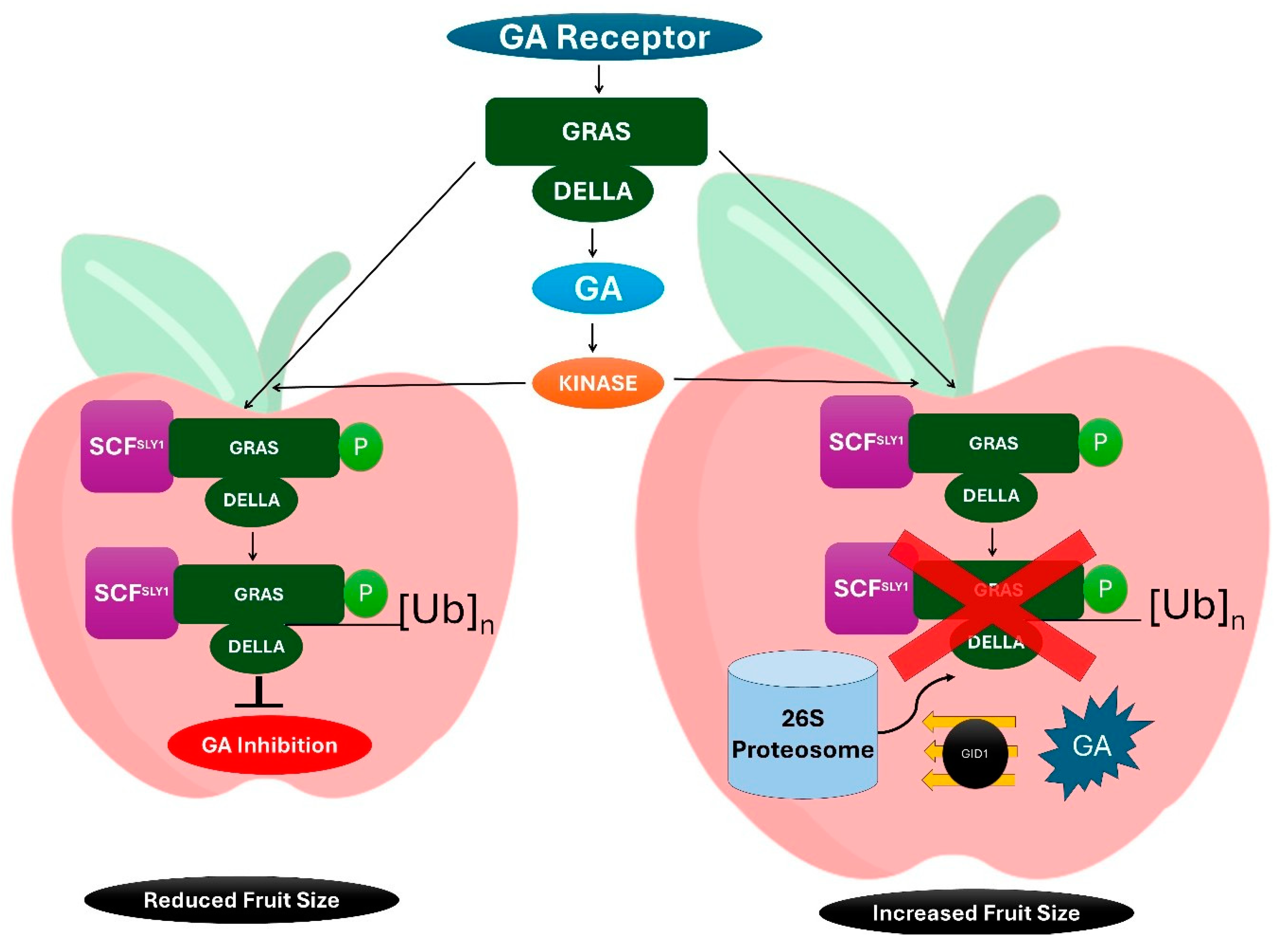
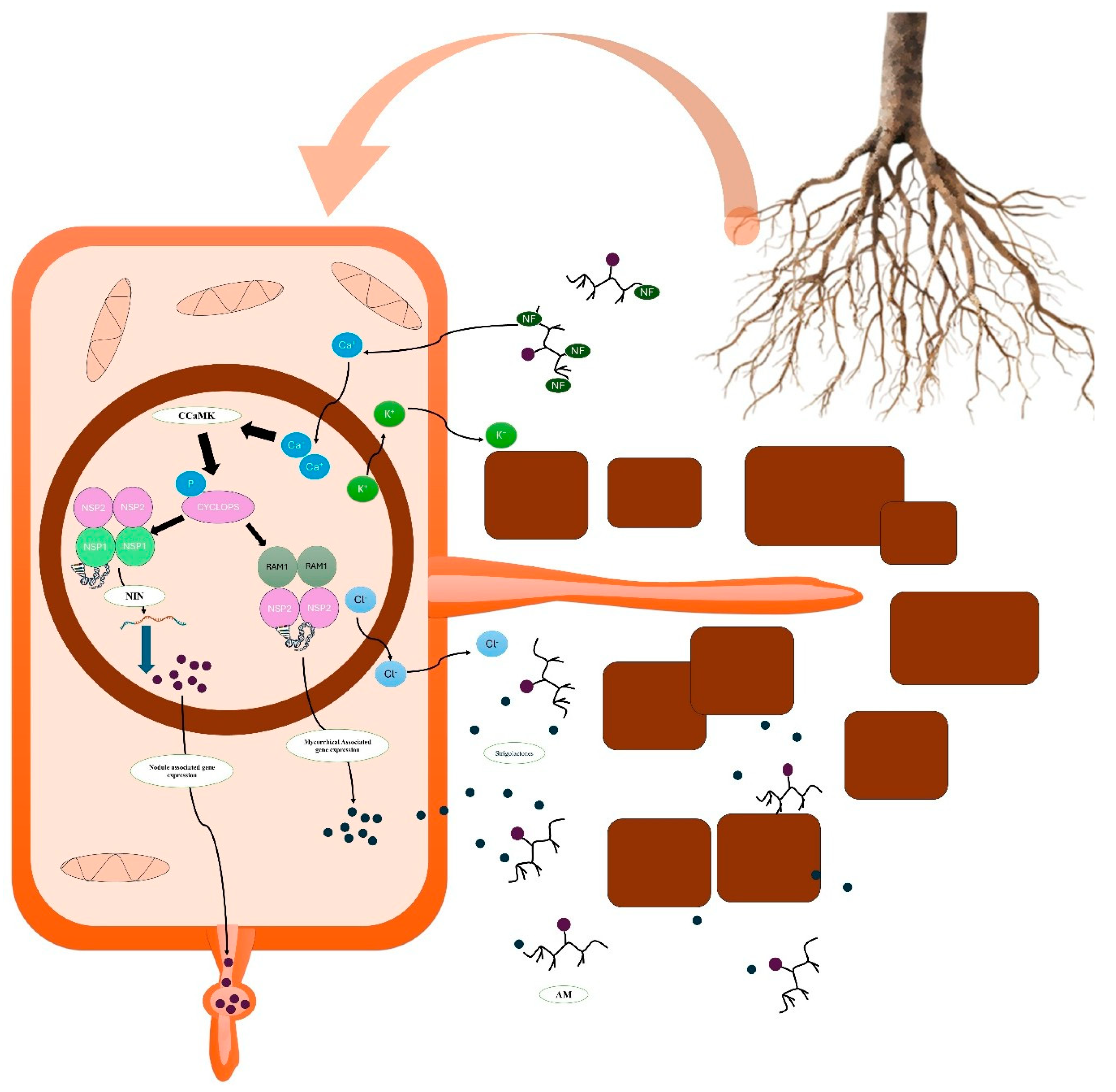
2.2. GRAS Proteins: Versatility in Symbiosis and Stress
2.3. NAC with Unusual Regulatory Mechanisms

| S.N. Tran- Scription Factor (TF) |
TF Family | Plant Species | Function | |
|---|---|---|---|---|
| SUSIBA2 | WRKY | Hordeum vulgare cv Pongo | Binds to the Sugar-Responsive Elements of the iso1 Promoter for participation in sugar signalling |
Sun et al., 2003 |
| AtVOZ1 and AtVOZ2, |
Arabidopsis | bind to the 38-bp cis-acting region of A. thaliana V-PPase gene, AVP1 | Mit- suda et al., 2004 | |
| NtWRKY12 | WRKY | Tobacco (Nicotiana tabacum ‘Samsun NN’ | NtWRKY12 and TGA1a act synergistically in PR-1a expression induced by salicylic acid and bacterial elicitors. | Van- verk et al., 2008 |
| myb52, myb-like TF, hb5 hb15 showed |
Arabidopsis | Hyper lignified SCW Ectopic lignification |
Cassan- Wang et al., 2013 |
|
| JcNAC1 | NAC | Jatropha curcas | enhanced tolerance to drought and increased susceptibility to pathogens |
Qin et al., 2014 |
| Gb- WRKY2 |
WRKY | Ginkgo biloba | flowers and strongly induced by methyl jasmonate |
Liao et al., 2015 |
| NAC050 and NAC052 |
NAC | Arabidopsis | involved in transcriptional repression and flowering time control by associating with the histone demethylase JMJ14. |
Ning et al., 2015 |
| GIP1 | bZIP | Arabidopsis | early stages of Arabidopsis development |
Shaikhali, 2015 |
| AITR | ABA- induced transcription repressors (AITRs) |
Arabidopsis | 6 Arabidopsis AITR genes are induced by exogenous ABA |
Tian et al., 2017 |
| OsPCF2 (OsCPP5) OsNIN- like2, OsNIN- like3 and OsNIN- like4 |
TCP CPP NIN-like |
Oryza sativa L.) genotype Hasawi |
regulators of OsNHX1 gene expression in a salt tolerant rice genotype |
Almeidaet al., 2017 |
| PvBMY1 | APETALA2/Ethyle Response Factor |
Pnaenicum virgatum L | increase biomass yield in greenhouse-grown switchgrass by regulation of photosynthesis and related metabolism be-like | AmHleyvpaotelidgne- bavaram et al., 2018 |
| PvBMY3 | Nuclear- Factor Y |
|||
| Bel-like |
3. Functional Insights and Mechanisms
3.1. Unique DNA-Binding Domains
3.2. Post-Translational Modifications (PTMs)
3.3. Non-Coding RNA Interactions
4. Biotechnological Applications
4.1. Enhancing Stress Tolerance
4.2. Boosting Yield and Quality
4.3. Precision Breeding Tools
5. Challenges and Future Directions
6. Conclusions
Acknowledgments
References
- Ferrero, L.; Zhang, W.; Benhamed, M.; Crespi, M.; Ariel, F. Non-B DNA in plant genomes: prediction, mapping, and emerging roles. Trends in Plant Science 2024. [Google Scholar] [CrossRef] [PubMed]
- Udvardi, M.K.; Kakar, K.; Wandrey, M.; Montanari, O.; Murray, J.; Andriankaja, A.; Zhang, J.Y.; Benedito, V.; Hofer, J.M.; Chueng, F.; et al. Legume transcription factors: global regulators of plant development and response to the environment. Plant Physiology 2007, 144, 538–549. [Google Scholar] [CrossRef] [PubMed]
- Menconi, J.; Perata, P.; Gonzali, S. Novel R2R3 MYB transcription factors regulate anthocyanin synthesis in Aubergine tomato plants. BMC Plant Biology 2023, 23, 148–148. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Ling, J.; Song, L.; Zhao, L.; Wang, Y.; Zhao, T. Transcriptomic profiling of tomato leaves identifies novel transcriptionfactors responding to dehydration stress. International Journal of Molecular Sciences 2023, 24, 9725. [Google Scholar] [CrossRef]
- Chen, L.; Wu, F.; Zhang, J. NAC and MYB families and lignin biosynthesis-related members identification and expression analysis in Melilotus albus. Plants 2021, 10, 303–303. [Google Scholar] [CrossRef]
- Liao, Y.L.; Shen, Y.B.; Chang, J.; Zhang, W.W.; Cheng, S.Y.; Xu, F. Isolation, expression, and promoter analysis of GbWRKY2: a novel transcription factor gene from Ginkgo biloba. International Journal of Genomics, 2015; 607185. [Google Scholar]
- Zhao, X.; Yang, J.; Li, X.; Li, G.; Sun, Z.; Chen, Y.; Chen, Y.; Xia, M.; Li, Y.; Yao, L.; et al. Identification and expression analysis of GARP superfamily genes in response to nitrogen and phosphorus stress in Spirodela polyrhiza. BMC Plant Biology 2022, 22, 308. [Google Scholar] [CrossRef]
- Alabadí, D.; Sun, T.P. Green Revolution DELLA Proteins: Functional Analysis and Regulatory Mechanisms. Annual Review of Plant Biology, 2025; 76. [Google Scholar]
- Puga, M.I.; Poza-Carrión, C.; Martinez-Hevia, I.; Perez-Liens, L.; Paz-Ares, J. Recent advances in research on phosphate starvation signaling in plants. Journal of plant research, 2024; 1–16. [Google Scholar]
- Chen, H.; Pugh, B.F. What do transcription factors interact with? Journal of molecular biology 2021, 433, 166883. [Google Scholar] [CrossRef]
- Sun, C.; Palmqvist, S.; Olsson, H.; Borén, M.; Ahlandsberg, S.; Jansson, C. A novel WRKY transcription factor, SUSIBA2, participates in sugar signaling in barley by binding to the sugar-responsive elements of the iso1 promoter. The Plant Cell 2003, 15, 2076–2092. [Google Scholar] [CrossRef]
- Sharma, M.; Sidhu, A.K.; Samota, M.K.; Gupta, M.; Koli, P.; Choudhary, M. Post-Translational Modifications in Histones and Their Role in Abiotic Stress Tolerance in Plants. Proteomes 2023, 11, 38–38. [Google Scholar] [CrossRef]
- Mitsuda, N.; Hisabori, T.; Takeyasu, K.; Sato, M.H. , 2004.
- Ou, X.; Sun, L.; Chen, Y.; Zhao, Z.; Jian, W. Characteristics of NAC transcription factors in Solanaceae crops and their roles in responding to abiotic and biotic stresses. Biochemical and Biophysical Research Communications, 2024; 149840. [Google Scholar]
- Bo, C.; Liu, D.; Yang, J.; Ji, M.; Li, Z.; Zhu, Y.; Duan, Y.; Xue, J.; Xue, T. Comprehensive in silico characterization of NAC transcription factor family of Pinellia ternata and functional analysis of PtNAC66 under high-temperature tolerance in transgenic Arabidopsis thaliana. Plant Physiology and Biochemistry 2024, 208, 108539. [Google Scholar] [CrossRef]
- Waseem, M.; Nkurikiyimfura, O.; Niyitanga, S.; Jakada, B.H.; Shaheen, I.; Aslam, M.M. GRAS transcription factors emerging regulator in plants growth, development, and multiple stresses. Molecular Biology Reports 2022, 49, 9673–9685. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Sun, G.; Xiao, J.; He, X.; Jiang, H.; Zhang, Z.; Zhang, Q.; Li, K.; Zhang, S.; Shi, X.; et al. AlphaFold-guided redesign of a plant pectin methylesterase inhibitor for broad-spectrum disease resistance. Molecular Plant 2024, 17, 1344–1368. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.S.; Ali, A.; Abbas, M.; Sun, Y.; Li, Y.; Li, Q.; Ragauskas, A.J. Harnessing miRNA156: A molecular Toolkit for reshaping plant development and achieving ideal architecture. Plant Physiology and Biochemistry 2024, 215, 109071. [Google Scholar] [CrossRef] [PubMed]
- Suhorukova, A.V.; Sobolev, D.S.; Milovskaya, I.G.; Fadeev, V.S.; Goldenkova-Pavlova, I.V.; Tyurin, A.A. A Molecular Orchestration of Plant Translation under Abiotic Stress. Cells 2023, 12, 2445. [Google Scholar] [CrossRef]
- Kumari, P.; Ojha, R.; Varshney, V.; Gupta, V.; Salvi, P. Transcription Factors and Their Regulatory Role in Plant Defence Response. In Proceedings of the Biotechnological Advances for Disease Tolerance in Plants. Springer Nature; 2024; pp. 337–362. [Google Scholar]
- Dutta, M.; Saha, A.; Moin, M.; Kirti, P.B. Genome-wide identification, transcript profiling and bioinformatic analyses of GRAS transcription factor genes in rice. Frontiers in Plant Science 2021, 12, 777285. [Google Scholar] [CrossRef]
- Qin, X.; Zheng, X.; Huang, X.; Lii, Y.; Shao, C.; Xu, Y.; Chen, F. A novel transcription factor JcNAC1 response to stress in newmodel woody plant Jatropha curcas. Planta 2014, 239, 511–520. [Google Scholar] [CrossRef]
- Méndez, T.; Guajardo, J.; Cruz, N.; Gutiérrez, R.A.; Norambuena, L.; Vega, A.; Moya-León, M.A.; Herrera, R. The Characterization of a Novel PrMADS11 Transcription Factor from Pinus radiata Induced Early in Bent Pine Stem. International Journal of Molecular Sciences 2024, 25, 7245–7245. [Google Scholar] [CrossRef]
- Hartmann, R.M.; Schaepe, S.; Nübel, D.; Petersen, A.C.; Bertolini, M.; Vasilev, J.; Küster, H.; Hohnjec, N. Insights into the complex role of GRAS transcription factors in the arbuscular mycorrhiza symbiosis. Scientific Reports 2019, 9, 3360. [Google Scholar] [CrossRef]
- Ning, Y.Q.; Ma, Z.Y.; Huang, H.W.; Mo, H.; Zhao, T.T.; Li, L.; Cai, T.; Chen, S.; Ma, L.; He, X.J. Two novel NAC transcription factors regulate gene expression and flowering time by associating with the histone demethylase JMJ14. Nucleic acids research 2015, 43, 1469–1484. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, Z.; Yu, X.; Zhao, L.; Zhao, M.; Han, X.; Qi, S. Identification of two novel R2R3-MYB transcription factors, 333 PsMYB114L and PsMYB12L, related to anthocyanin biosynthesis in Paeonia suffruticosa. International journal of molecular sciences 334 2019, 20, 1055. [Google Scholar] [CrossRef]
- Mishra, S.; Roychowdhury, R.; Ray, S.; Hada, A.; Kumar, A.; Sarker, U.; Aftab, T.; Das, R. , 2024.
- Khan, Y.; Xiong, Z.; Zhang, H.; Liu, S.; Yaseen, T.; Hui, T. Expression and roles of GRAS gene family in plant growth, signal transduction, biotic and abiotic stress resistance and symbiosis formation-A review. Plant Biology 2022, 24, 404–416. [Google Scholar] [CrossRef] [PubMed]
- Duan, B.; Xie, X.; Jiang, Y.; Zhu, N.; Zheng, H.; Liu, Y.; Hua, X.; Zhao, Y.; Sun, Y. GhMYB44 enhances stomatal closure to confer drought stress tolerance in cotton and Arabidopsis. Plant Physiology and Biochemistry 2023, 198, 107692. [Google Scholar] [CrossRef]
- Kim, H.J.; Hong, S.H.; Kim, Y.W.; Lee, I.H.; Jun, J.H.; Phee, B.K.; Rupak, T.; Jeong, H.; Lee, Y.; Hong, B.S.; et al. Gene regulatory cascade of senescence-associated NAC transcription factors activated by ETHYLENE-INSENSITIVE2-mediated leaf senescence signalling in Arabidopsis. Journal of experimental botany 2014, 65, 4023–4036. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Tan, N.W.K.; Chung, F.Y.; Yamaguchi, N.; Gan, E.S.; Ito, T. Transcriptional regulators of plant adaptation to heat stress. International Journal of Molecular Sciences 2023, 24, 13297. [Google Scholar] [CrossRef]
- Xia, L.; Sun, S.; Han, B.; Yang, X. NAC domain transcription factor gene GhNAC3 confers drought tolerance in plants. Plant Physiology and Biochemistry 2023, 195, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Cui, Y.; Feng, Y.; Hu, Y.; Liu, L.; Duan, L. Long non-coding RNAs of plants in response to abiotic stresses and their regulating roles in promoting environmental adaption. Cells 2023, 12, 729. [Google Scholar] [CrossRef]
- Singh, P.; Mukhopadhyay, K. Comprehensive molecular dissection of TIFY Transcription factors reveal their dynamic responses to biotic and abiotic stress in wheat. Triticum aestivum L.). Scientific Reports 2021, 11, 9739. [Google Scholar] [CrossRef]
- Rushton, P.J.; Bokowiec, M.T.; Laudeman, T.W.; Brannock, J.F.; Chen, X.; Timko, M.P. TOBFAC: the database of tobacco transcription factors. BMC bioinformatics 2008, 9, 1–7. [Google Scholar] [CrossRef]
- Ho-Plágaro, T.; García-Garrido, J.M. Multifarious and interactive roles of GRAS transcription factors during arbuscular mycorrhiza development. Frontiers in Plant Science 2022, 13, 836213. [Google Scholar] [CrossRef]
- Chorostecki, U.; Bologna, N.G.; Ariel, F. The plant noncoding transcriptome: a versatile environmental sensor. The EMBO Journal 2023, 42, 114400. [Google Scholar] [CrossRef]
- Deng, C.; Wang, J.; Lu, C.; Li, Y.; Kong, D.; Hong, Y.; Huang, H.; Dai, S. CcMYB6-1 and CcbHLH1, two novel transcription factors synergistically involved in regulating anthocyanin biosynthesis in cornflower. Plant physiology and biochemistry 2020, 151, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.P.; Kim, C. Photosynthetic ROS and retrograde signaling pathways. New Phytologist 2024, 244, 1183–1198. [Google Scholar] [CrossRef]
- Ambavaram, M.M.; Ali, A.; Ryan, K.P.; Peoples, O.; Snell, K.D.; Somleva, M.N. Novel transcription factors PvBMY1 and PvBMY3increase biomass yield in greenhouse-grown switchgrass. Panicum virgatum L.). Plant Science 2018, 273, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Pooam, M.; El-Ballat, E.M.; Jourdan, N.; Ali, H.M.; Hano, C.; Ahmad, M.; El-Esawi, M.A. SNAC3 transcription factor enhances arsenic stress tolerance and grain yield in rice (Oryza sativa L.) through regulating physio-biochemical mechanisms, stress responsive genes, and cryptochrome 1b. Plants 2023, 12, 2731. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Zhang, L.; Yu, Q.; Zhang, J.; Li, P.; Zhang, Y.; Xing, X.; Ding, L.; Fang, W.; Chen, F.; et al. Integrated signals of jasmonates, sugars, cytokinins and auxin influence the initial growth of the second buds of chrysanthemum after decapitation. Biology 2021, 10, 440–440. [Google Scholar] [CrossRef]
- Panchal, A.; Maurya, J.; Seni, S.; Singh, R.K.; Prasad, M. An insight into the roles of regulatory ncRNAs in plants: An abiotic stress and developmental perspective. Plant Physiology and Biochemistry 2023, 201, 107823. [Google Scholar] [CrossRef]
- Chakraborty, S.; Harris, J.M. At the crossroads of salinity and rhizobium-legume symbiosis. Molecular Plant-Microbe Interactions 2022, 35, 540–553. [Google Scholar] [CrossRef]
- Liu, Z.; Xiong, T.; Zhao, Y.; Qiu, B.; Chen, H.; Kang, X.; Yang, J. Genome-wide characterization and analysis of golden 2-like transcription factors related to leaf chlorophyll synthesis in diploid and triploid Eucalyptus urophylla. Frontiers in Plant Science 2022, 13, 952877. [Google Scholar] [CrossRef]
- Verk, M.C.V.; Pappaioannou, D.; Neeleman, L.; Bol, J.F.; Linthorst, H.J. A novel WRKY transcription factor is required for induction of PR-1a gene expression by salicylic acid and bacterial elicitors. Plant Physiology 2008, 146, 1983–1995. [Google Scholar] [CrossRef]
- Chen, H.; Li, H.; Lu, X.; Chen, L.; Liu, J.; Wu, H. Identification and expression analysis of GRAS transcription factors to elucidatecandidate genes related to stolons, fruit ripening and abiotic stresses in woodland strawberry (Fragaria vesca). International journal of molecular sciences 2019, 20, 4593. [Google Scholar] [CrossRef]
- Lehti-Shiu, M.D.; Panchy, N.; Wang, P.; Uygun, S.; Shiu, S.H. Diversity, expansion, and evolutionary novelty of plant DNA-binding transcription factor families. Biochimica et Biophysica Acta (BBA)-Gene Regulatory Mechanisms 2017, 1860, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, F.; Shah, S.H.; Jan, A. Overexpression of the DREB1A gene under stress-inducible promoter delays leaf senescence and improves drought tolerance in rice. Cereal Research Communications 2023, 51, 851–857. [Google Scholar] [CrossRef]
- Cai, L.; Guo, Z.; Ding, J.; Gai, Z.; Liu, J.; Meng, Q.; Yang, X.; Zhang, N.; Wang, Q. Genome-Wide Identification and Exogenous Hormone and Stress Response Expression Analysis of the GARP Gene Family in Soybean (Glycine max). Agriculture, 2024; 2109. [Google Scholar]
- Shaikhali, J. GIP1 protein is a novel cofactor that regulates DNA-binding affinity of redox-regulated members of bZIP transcription factors involved in the early stages of Arabidopsis development. Protoplasma 2015, 252, 867–883. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Yang, Y.Y.; Chen, J.Y.; Lakshmanan, P.; Kuang, J.F.; Lu, W.J.; Shan, W. MaNAC029 modulates ethylene biosynthesis and 390 fruit quality and undergoes MaXB3-mediated proteasomal degradation during banana ripening. Journal of Advanced Research 2023, 53, 33–47. [Google Scholar] [CrossRef]
- Sutradhar, M.; Singh, B.K.; Samanta, S.; Ali, M.N.; Mandal, N. The overexpression of OsMed 37_6, a mediator complex subunit enhances salt stress tolerance in rice. Biocatalysis and Agricultural Biotechnology 2024, 58, 103212. [Google Scholar] [CrossRef]
- Liu, Y.; Zhu, P.; Cai, S.; Haughn, G.; Page, J.E. Three novel transcription factors involved in cannabinoid biosynthesis in Cannabis sativa L. Plant Molecular Biology 2021, 106, 49–65. [Google Scholar] [CrossRef]
- Almeida, D.M.; Gregorio, G.B.; Oliveira, M.M.; Saibo, N.J. Five novel transcription factors as potential regulators of OsNHX1 gene expression in a salt tolerant rice genotype. Plant molecular biology 2017, 93, 61–77. [Google Scholar] [CrossRef]
- Hecker, D.; Lauber, M.; Ardakani, F.B.; Ashrafiyan, S.; Manz, Q.; Kersting, J.; Hoffmann, M.; Schulz, M.H.; List, M. Computational tools for inferring transcription factor activity. Proteomics 2023, 23, 2200462. [Google Scholar] [CrossRef]
- Rahman, H.; Ramanathan, V.; Nallathambi, J.; Duraialagaraja, S.; Muthurajan, R. Over-expression of a NAC 67 transcription factor from finger millet (Eleusine coracana L.) confers tolerance against salinity and drought stress in rice. BMC biotechnology 2016, 16, 7–20. [Google Scholar] [CrossRef]
- Baoxiang, W.; Zhiguang, S.; Yan, L.; Bo, X.; Jingfang, L.; Ming, C.; Yungao, X.; Bo, Y.; Jian, L.; Jinbo, L.; et al. A pervasive phosphorylation cascade modulation of plant transcription factors in response to abiotic stress. Planta 2023, 258, 73. [Google Scholar] [CrossRef]
- Iannelli, M.A.; Nicolodi, C.; Coraggio, I.; Fabriani, M.; Baldoni, E.; Frugis, G. A Novel Role of Medicago truncatula KNAT3/4/5- like Class 2 KNOX Transcription Factors in Drought Stress Tolerance. International Journal of Molecular Sciences 2023, 24, 12668. [Google Scholar] [CrossRef] [PubMed]
- Cassan-Wang, H.; Goué, N.; Saidi, M.N.; Legay, S.; Sivadon, P.; Goffner, D.; Grima-Pettenati, J. , 2013.
- Batool, A.; Zahra, N.; Naseer, R.; Shahzad, S.; Iqbal, S.; Kausar, A.; Raza, A. Harnessing the role of genes involved in plant architectural changes. Plant Growth Regulation 2023, 101, 15–34. [Google Scholar] [CrossRef]
- Vranic´, M.; Perochon, A.; Doohan, F.M. Transcriptional Profiling Reveals the Wheat Defences against Fusarium Head Blight Disease Regulated by a NAC Transcription Factor. Plants 2023, 12, 2708. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Lv, F.; Gao, L.; Gu, J.; Yang, R.; Li, S.; Li, Y.; Li, S.; Wang, P. Novel R2R3-MYB transcription factor LiMYB75 enhances leaf callus regeneration efficiency in Lagerstroemia indica. Forests 2023, 14, 517. [Google Scholar] [CrossRef]
- Tian, H.; Chen, S.; Yang, W.; Wang, T.; Zheng, K.; Wang, Y.; Cheng, Y.; Zhang, N.; Liu, S.; Li, D.; et al. A novel family of transcription factors conserved in angiosperms is required for ABA signalling. Plant, cell & environment 2017, 40, 2958–2971. [Google Scholar]
- Cahn, J.; Regulski, M.; Lynn, J.; Ernst, E.; Alves, C.D.S.; Ramakrishnan, S.; Chougule, K.; Wei, S.; Lu, Z.; Xu, X.; et al. MaizeCODE reveals bi-directionally expressed enhancers that harbor molecular signatures of maize domestication. Nature Communications 2024, 15, 10854. [Google Scholar] [CrossRef]
- Singh, M.; Singh, A.; Yadav, N.; Yadav, D.K. Current perspectives of ubiquitination and SUMOylation in abiotic stress tolerance in plants. Frontiers in Plant Science 2022, 13, 993194. [Google Scholar] [CrossRef]
- Zhang, D.; Ma, Y.; Naz, M.; Ahmed, N.; Zhang, L.; Zhou, J.J.; Yang, D.; Chen, Z. Advances in CircRNAs in the Past Decade: Review of CircRNAs Biogenesis, Regulatory Mechanisms, and Functions in Plants 2024, 15, 958–958.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).



