Submitted:
12 February 2025
Posted:
12 February 2025
You are already at the latest version
Abstract
Cysticercosis is a serious zoonotic parasitic disease caused by the larval cysts of Taenia solium parasitizing humans or pigs. The immune pathogenesis of this disease is currently unclear. Preliminary studies found that the thioredoxin peroxidase (TPx) protein in the excretory-secretory antigens (ESA) of the cysticercus cellulosae can induce T-cell immune imbalance in piglets, leading to a Th2-type immune response. In this study, flow cytometry revealed that TPx protein can induce Th1 cells differentiation within 24 hours and Th2 cells differentiation within 48 hours. Further analysis of the different transcriptomes after TPx protein treatment of Jurkat T cells suggested that TPx protein may induce host Th1/Th2 cells imbalance through the janus kinase/signal transducer and activator of transcription (JAK/STAT) signaling pathway. Western blot results indicated that treatment with TPx protein for 48 hours significantly increased the expression of p-JAK3, p-STAT6, and GATA3 proteins. Interestingly, after using a JAK3 inhibitor, the expression level of the Th1 transcription factor TBX21 significantly increased, indicating that after inhibiting Th2 cells differentiation, T cells tend to differentiate toward the Th1 cells direction. In summary, the JAK/STAT signaling pathway plays a key role in the cysticercus cellulosae TPx protein induced Th1/Th2 cells imbalance, providing important scientific evidence for elucidating the immune pathogenesis of cysticercosis and vaccine development.
Keywords:
Taenia solium
; cysticercus cellulosae
; thioredoxin peroxidase
; JAK/STAT signaling pathway
; Th1 cells differentiation
; Th2 cells differentiation
; Th1/Th2 cells imbalance
1. Introduction
Cysticercus cellulosae can parasitize various tissues in humans, including subcutaneous layers, muscles, the brain, and the eyes, causing cysticercosis. The most severe form is neurocysticercosis (NCC), which can lead to neurological symptoms such as epilepsy, headaches, cognitive decline, and increased intracranial pressure [1,2]. According to studies, 30% of epilepsy cases in endemic countries such as those in Africa, Asia, and Latin America are attributed to this disease, affecting as much as 1% of the population in these regions [3]. The World Health Organization (WHO) defines cysticercosis as a potentially eradicable tropical parasitic disease and has set goals for its prevention, elimination, and eradication [4]. Currently, the treatment of NCC mainly relies on medications and surgery, but these options have significant side effects, and the development of vaccines for humans to prevent cysticercosis is still in the exploratory stage, one of the major obstacles to vaccine development is the complex pathogenesis of cysticercosis [5,6].
Excretory-secretory antigens (ESAs) are antigenic substances secreted and excreted by parasites during their parasitic phase within the host. These antigens play essential roles in nutrient absorption, tissue invasion, immune evasion, interactions between the host and the parasite, and other lifecycle processes. Parasite ESAs can be released into the bloodstream, regulating the host's immune response, they can elicit a positive immune response to control infections or a negative immune response to evade the host's immune attacks, thereby establishing long-term chronic infections [7,8]. Cysticercus cellulosae ESAs exert strong immunoregulatory effects, causing an imbalance in the CD4+/CD8+ T ratio in peripheral blood mononuclear cells (PBMCs) of piglets, leading them toward differentiation into Th2 cells [9]. Due to the complex composition of cysticercus cellulosae ESAs, further analysis using label-free quantification (LFQ) proteomics was conducted [10], which identified and successfully expressed the thioredoxin peroxidase (TPx) protein, which plays a crucial role in regulating T-cells immune responses [11]. The TPx protein is expressed in the cyst wall, the cyst layer, and muscle cells of cysticercus cellulosae and can be specifically recognized by positive serum from cysticercosis-infected pigs [12,13,14].Cysticercus cellulosae TPx protein could increase the CD4+/CD8+ T ratio in pig PBMCs and lead to increased secretion of interleukin-4 (IL-4) and IL-10, thereby initiating a Th2-type immune response while suppressing Th1-type immune response [15].
Parasitic infections can alter the host's immune responses, involving helper CD4+ T cells (Th), B cells, and innate lymphoid cells (ILCs) that mediate adaptive immunity. Th1 cells can secrete IL-2, tumor necrosis factor (TNF), and interferon-gamma (IFN-γ), while Th2 cells can secrete IL-4, IL-5, and IL-13 [16]. Previous studies have shown that these cytokines can impact the host's gut microbiome, allergic diseases, diabetes, and the efficacy of BCG vaccination for tuberculosis [17,18,19,20]. The janus kinase/signal transducer and activator of transcription (JAK/STAT) signaling pathway is primarily regulated by cytokines and participates in the regulation of Th1/Th2 cells balance. Cytokines first bind to their corresponding receptors, causing them to dimerize, JAK kinases are then phosphorylated and coupled to the receptors, forming docking sites on phosphorylated tyrosine residues, recruiting STAT proteins with Src homology 2 (SH2) domains to phosphorylate them into dimers that translocate to the nucleus, binding to specific DNA sites to regulate gene transcription [21].After a parasitic infection, the JAK/STAT signaling pathway can modulate the host T-cells immune response, influencing T-cells proliferation and differentiation into various subgroups, playing a crucial role in the occurrence, development, and interaction between the parasite and host [22].
Parasite infections can modulate the host's metabolism and immune response. This regulation is not only crucial for understanding host-pathogen interactions and treating parasitic diseases but also valuable for developing therapeutic approaches for autoimmune and inflammatory diseases [23,24]. Experimental autoimmune encephalomyelitis (EAE) is an animal model for the human disease multiple sclerosis (MS). Taenia crassiceps excreted/secreted products (TcES) can induce Th2 cytokines while inhibiting Th1 and Th17 responses; in EAE mice treated with TcES, both the polyclonal and antigen-specific proliferative responses of lymphocytes were inhibited, demonstrating a more effective suppression of EAE progression compared to dexamethasone. This may be related to Th2 immune polarization and impaired lymphocyte activation [25]. Schistosoma soluble egg antigen (SEA) can enhance Th2 immune responses, improve type 2 diabetes in Leprdb/db mice, and reduce the severity of dextran sulfate sodium (DSS)-induced colitis [26,27]. These findings indicate that parasite excretory/secretory antigens (ESAs) hold great potential in treating autoimmune and inflammatory diseases [28].
The Jurkat cell model can simulate T lymphocyte function due to its stable cellular characteristics and straightforward culture methods. It has been widely used in vitro studies of viral diseases and atypical pathogens like Chlamydia pneumoniae and Chlamydia trachomatis, offering important references and guidance for the treatment and mechanism research of various infectious diseases [29]. Building on prior findings that the Taenia solium TPx protein can regulate the imbalance of Th1/Th2 cells in piglets, this cell model was selected to further investigate the molecular mechanisms behind cysticercosis immunity. By analyzing differential transcriptomic data, it was found that the JAK/STAT signaling pathway may be involved in TPx protein regulation of Th1/Th2 cell imbalance. This aims to lay the foundation for elucidating the molecular mechanisms by which the Taenia solium TPx protein induces Th1/Th2 cell imbalance, discovering effective multi-target new candidate vaccine molecules for cysticercosis, and exploring the applications of TPx protein in autoimmune and inflammatory diseases.
2. Materials and Methods
2.1. Preparation of Cysticercus Cellulosae TPx Protein
According to the NCBI database, the TPx gene sequence of Taenia solium (GenBank: AHZ89374.1) was PCR amplified using a whole gene synthesis method and cloned into the pcDNA3.4 vector to construct the recombinant plasmid pcDNA3.4-TPx. This plasmid was then transfected into HEK293 cells. After analysis by SDS-PAGE and Western blot, the expressed recombinant TPx protein was found to have a relative molecular mass of approximately 26 kDa, predominantly present in the supernatant of the cultured cells transfected with the recombinant plasmid. The purified recombinant TPx protein, which carries a His tag, was recognized by anti-His tag antibodies, confirming its antigenicity. The TPx protein was successfully purified and stored at -80 °C.
2.2. Cultivation and Activation of Jurkat Cells
The human leukemia T lymphocyte Jurkat cells, Clone E6-1 (referred to as “Jurkat cells” hereafter), were purchased from the Cell Bank of the Chinese Academy of Sciences, Shanghai. Jurkat cells were cultured in RPMI 1640 medium (Thermo Fisher, China) supplemented with 10% fetal bovine serum (Absin, China) and 1% antibiotics (Solarbio Sciences, China), at 37 °C with 5% CO2. They were activated with 0.5 ng/ml PMA (Solarbio Sciences, China) and 0.5 µM ionomycin (MedChemExpress, USA) for 12 hours for subsequent experiments.
2.3. Flow Cytometry Analysis
Cells were collected and washed twice with pre-cooled PBS. An appropriate amount of allophycocyanin (APC)-labeled anti-human CD4 antibody (BD Pharmingen, USA) was added to the cell suspension and incubated in the dark at 4°C for 30 minutes. Then, 250 μl of Fixation/Permeabilization solution was added and mixed, followed by incubation in the dark for 20 minutes. After that, an appropriate amount of phycoerythrin (PE)-labeled anti-human IL-4 antibody (BD Pharmingen, USA) and fluorescein isothiocyanate (FITC)-labeled anti-human IFN-γ antibody (BD Pharmingen, USA) was added, and the mixture was incubated in the dark at 4°C for 30 minutes. Cells were washed twice with pre-cooled 1× BD Perm/Wash™ buffer and resuspended in PBS. Samples were analyzed on a BD FACSCanto II flow cytometer (BD Biosciences). All data were analyzed using FlowJo software (TreeStar, Ashland, OR, USA).
2.4. Transcriptome Sequencing and Bioinformatic Analysis
In the experiment, Jurkat cells treated with TPx protein were designated as the experimental group, while Jurkat cells without TPx protein served as the control group. Each group consisted of three samples, and cells were collected after being cultured for 0, 24, and 48 hours to extract RNA. RNA samples were tested and library construction was performed by Shanghai Ouyi Biomedical Technology, followed by RNA-seq sequencing and sequence comparison. Differential protein-coding genes were screened using DESeq software based on P < 0.05 and |log2FC| > 1.0, followed by GO and KEGG enrichment analysis.
2.5. Western Blotting
The collected cells were centrifuged at 1200 rpm for 5 minutes, washed twice with pre-chilled PBS, and lysed on ice for 30 minutes using freshly prepared lysis buffer (RIPA lysis buffer: PMSF protease inhibitor: phosphatase inhibitor = 100:1:1), followed by centrifugation at 13,000 rpm for 15 minutes at 4 °C to obtain the supernatant containing the cell protein. The protein concentration was detected using the BCA method, and the sample buffer was added before boiling in a water bath at 100 °C for 10 minutes to denature the proteins.
Protein samples were added to the sample wells. Electrophoretic at 80V for about 30 minutes, switching to 120V when markers separate, The target protein was transferred to PVDF membrane at a constant flow of 400mA for 35minutes. The PVDF membrane was blocked at room temperature with 5% non-fat milk for 2 hours. After blocking, the corresponding primary antibodies such as p-JAK3 ( Absin, China), JAK3 (Wuhan Sanying, China), p-STAT6 ( Absin, China), STAT6 (Wuhan Sanying, China), GATA3 (Wuhan Sanying, China), TBX21 ( Absin, China), and GAPDH (Wuhan Sanying, China) was separately added to the box, incubated at 4°C for over 12 h. The membrane was washed three times with TBST for 10 minutes each time. According to the antibody dilution ratio, the goat anti-rabbit (mouse) secondary antibody (Wuhan Sanying, China) was prepared with universal antibody diluent and incubated at room temperature for 1.5 h. Finally, the membrane was washed 3 times with TBST buffer for 10 minutes each time and photographed using ECL reagent (Meilunbio, China) for development. Gray value analysis was performed using lmage Lab software. As needed, we set up 100µM JAK3-IN-6 inhibitor ( Absin, China) pretreatment group for 12 h observe the effect of inhibitor on Th1 and Th2 cells differentiation.
2.6. Statistical Analysis
This study used SPSS 29.0 software for statistical analysis and GraphPad Prism 9.0 software for chart creation. Measurement data are expressed as mean ± standard deviation (x±SD, n=3) and independent samples t-tests were used for pairwise comparisons between groups. One-way ANOVA was used for comparisons among multiple groups, with P < 0.05 indicating a statistically significant difference.
3. Results
3.1. Effects of Cysticercus Cellulosae TPx Protein on Th1 and Th2 Cell Differentiation
The TPx protein was applied to Jurkat cells induced with 0.5 ng/ml PMA and a final concentration of 0.5 μM Ionomycin for 12 hours at 0h, 24h, and 48h. Jurkat cells without TPx protein served as the control group. The expression of CD4+IFN-γ+Th1 cells and CD4+IL-4+Th2 cells was detected by flow cytometry. Compared to the 0h control group, the number of CD4+IFN-γ+Th1 cells in the 24h experimental group significantly increased (Figure 1A), while in the 48h experimental group, the number of CD4+IL-4+Th2 cells significantly increased compared to the 24h group (Figure 1B). The results indicate that the TPx protein from cysticercus cellulosae induces Th1 cell differentiation in Jurkat cells at 24h and Th2 cell differentiation at 48h.
3.2. Differential Transcriptomic Analysis of TPx Protein-Induced Human Jurkat T Cell Differentiation
3.2.1. Differential Gene Analysis
Differential transcriptomic analysis was conducted on Jurkat cells treated with TPx protein for 0h, 24h, and 48h. Differentially expressed genes were screened based on P < 0.05 and |log2FC| > 1.0, resulting in a differential gene volcano plot. As shown in Table 1 and Figure 2, compared to the 0h control group, the 24h experimental group identified a total of 935 significantly differentially expressed genes, with 631 upregulated genes, including PTGDR, MS4A7, and TRPC6, and 304 downregulated genes, including CCL3, NUAK1, and PCDH1. Compared to the 24h experimental group, the 48h experimental group identified a total of 447 significantly differentially expressed genes, with 199 upregulated genes, including RFPL2, CALCB, and AGK-DT, and 248 downregulated genes, including XIRP1, RAB13, and SCNN1B.
3.2.2. Gene Ontology (GO) Functional Annotation
Further GO enrichment analysis of the differentially expressed genes was performed. As shown in Figure 3, the GO enrichment analysis indicates that compared to the 0h control group, the top 30 GO terms in the 24h experimental group were mainly involved in immune response, chemokine-mediated signaling pathway, DNA-binding transcription factor activity, and others. Compared to the 24h experimental group, the top 30 GO terms in the 48h experimental group were mainly involved in positive regulation of cell adhesion, plasma membrane, cytokine receptor activity, and others. These results suggest that the TPx protein in cysticercus cellulosae can induce changes in Jurkat cells, affecting T cell immune processes, cell differentiation, and signaling molecules.
3.2.3. KEGG Enrichment Analysis
As shown in Figure 4, KEGG enrichment analysis showed that compared to the 0h control group, the 24h experimental group was enriched in signaling pathways mainly including Cytokine-cytokine receptor interaction, Chemokine signaling pathway, MAPK signaling pathway, and others. Compared to the 24h experimental group, the 48h experimental group was enriched in signaling pathways mainly including JAK-STAT signaling pathway, PI3K-Akt signaling pathway, Hematopoietic cell lineage, and others. Based on the GO and KEGG enrichment results, it is speculated that the TPx protein from porcine cysticercus may regulate Th1/Th2 cell imbalance through the JAK-STAT signaling pathway.
Further, a heatmap of 30 differentially expressed genes related to the JAK-STAT signaling pathway and Th1/Th2 cell differentiation was constructed(Figure 5). Compared to the 0h control group, the relative expression levels of differential genes such as JAK3, STAT6, JAK1, and SOCS3 were higher in the 24h and 48h experimental groups, and increased with the duration of TPx protein treatment. Conversely, the relative expression levels of differential genes such as TBX21, STAT1, and JAK2 were lower in the 24h and 48h experimental groups compared to the 0h control group. Therefore, the TPx protein can activate the JAK/STAT signaling pathway in Jurkat cells at both 24h and 48h, and may further induce Th1/Th2 cell imbalance through signaling molecules such as JAK3, STAT6, and SOCS3.
3.3. Western Blot Assay to Detect the Expression Levels of GATA3, TBX21, JAK3, and STAT6 Proteins and Protein Phosphorylation
Based on the differential transcriptomic analysis results mentioned above, we speculate that the TPx protein from porcine cysticercus may regulate Th1/Th2 cell imbalance through the JAK/STAT signaling pathway. Therefore, we further examined the changes in the expression levels of key proteins in the JAK/STAT signaling pathway, including JAK3, p-JAK3, p-STAT6, STAT6, and the Th1/Th2 cell transcription factors TBX21 and GATA3, using Western blotting to discuss the role of the JAK/STAT signaling pathway in the TPx protein-induced Th1/Th2 cell imbalance. As shown in Figure 6, compared to the 0h control group, there were no significant differences in the expression levels of p-JAK3, p-STAT6, and GATA3 proteins in the 24h experimental group (P > 0.05), while the expression level of TBX21 protein significantly increased (P < 0.01). Compared to the 24h experimental group, the expression levels of p-JAK3, p-STAT6, and GATA3 proteins in the 48h experimental group significantly increased (P < 0.05), whereas there was no significant difference in TBX21 protein expression (P > 0.05). These results preliminarily indicate that the TPx protein can induce an increase in the expression of the Th1 cell transcription factor TBX21 at 24h and an increase in the expression levels of p-JAK3 and p-STAT6 proteins in the JAK/STAT signaling pathway at 48h, thereby further promoting the expression of the Th2 cell transcription factor GATA3. Thus, the TPx protein from porcine cysticercus can regulate Th1/Th2 cell imbalance through the JAK/STAT signaling pathway.
3.4. Effects of JAK3-IN-6 Inhibitor on Cell Differentiation and the Expression Levels of JAK3, STAT6, GATA3, and TBX21 Proteins and Protein Phosphorylation
After pretreatment with the JAK3-IN-6 inhibitor in combination with TPx to induce cells for 12 hours, a Western blot was performed to detect changes in the levels of JAK3, p-JAK3, STAT6, p-STAT6, GATA3, and TBX21 proteins. This section discusses the effects of the JAK3-IN-6 inhibitor on TPx protein-induced Th1/Th2 cell imbalance. As shown in Figure 7, compared to the 0h control group, the JAK3-IN-6 inhibitor significantly suppressed the expression of p-JAK3, p-STAT6, and GATA3 proteins in the 24h and 48h experimental groups (P < 0.05). Additionally, in the 48h JAK3-IN-6 + TPx group compared to the TPx group, the JAK3-IN-6 inhibitor further suppressed the expression of p-JAK3, p-STAT6, and GATA3 proteins induced by TPx (P < 0.05). Compared to the 0h control group, the expression levels of TBX21 protein in the JAK3-IN-6 groups at 24h and 48h significantly increased (P < 0.05), and in the 48h JAK3-IN-6 + TPx group, the expression level of TBX21 protein significantly increased compared to the TPx group (P < 0.05). These results further indicate that the TPx protein may induce Th1/Th2 cell imbalance by activating the JAK/STAT signaling pathway.
4. Discussion
Taenia solium cysticercosis and Taeniasis (TSCT), caused by the tapeworm T. solium, is a foodborne and zoonotic disease classified since 2010 by WHO as a neglected tropical isease [30]. Research has shown that the PBMCs of symptomatic NCC patients exhibit a higher Th1 cytokine response, characterized by IL-1β, TNF-α and IFN-γ, while asymptomatic patients display a Th2 cytokine response with significantly elevated levels of IL-4 and IL-10. A shift from Th1 to Th2 cytokine response may favor the establishment of multicystic infection [31,32]. However, current studies have only focused on this phenomenon, and the molecular mechanisms by which the TPx protein regulates Th1/Th2 cell imbalance in cysticercosis patients have not been fully elucidated. Therefore, this study aims to further investigate the molecular mechanisms by which the TPx protein induces host Th1/Th2 cell imbalance through differential transcriptomic analysis.
Th1 cells are defined by the expression of the lineage cytokine IFN-γ and the master transcription factor T-bet, playing a role in early anti-infection responses. Th2 cells, defined by the expression of lineage cytokines IL-4/IL-5/IL-13 and the master transcription factor GATA3, enable the parasite to evade the host's immune attack, leading to a state of persistent chronic infection [33,34]. The TPx protein from Schistosoma mansoni can induce a significant proliferative response in T cells, along with the secretion of IFN-γ and IL-2 [35]. The TPx protein from Fasciola gigantica can stimulate healthy goat PBMCs to secrete IFN-γ, IL-2, IL-4, and IL-10, resulting in a mixed Th1/Th2 immune response [36]. The TPx protein from Trichinella spiralis can increase levels of Th2 cytokines (IL-4 and IL-10) while decreasing levels of Th1 cytokines (IFN-γ, IL-12, and TNF-α), resulting in a Th2-type immune response [37]. The results of this study indicate that the TPx protein from cysticercus cellulosae can induce T cell differentiation towards Th1 cells at 24h, generating a Th1-type immune response, while at 48h, it can induce T cell differentiation towards Th2 cells, producing a Th2-type immune response. This phenomenon is similar to the immune response observed in mouse models of cysticercosis. During early infection (2 weeks), the host develops a non-permissive Th1 response, whereas during late infection (8 weeks), molecules from the cysticerci induce a Th2 response that is permissive to parasite growth [38].
This study performed GO and KEGG enrichment analysis on differentially expressed genes after treatment with the TPx protein in Jurkat cells at 0h, 24h, and 48h. The results showed that these differentially expressed genes were enriched in signaling pathways, including the JAK-STAT signaling pathway, cytokine-cytokine receptor interaction, MAPK signaling pathway, and PI3K-Akt signaling pathway, which are involved in cellular immune processes, cell differentiation, and changes in signaling molecules in Jurkat T cells. Further analysis of the differentially expressed genes related to the JAK/STAT signaling pathway revealed that as the duration of TPx protein treatment increased, the expression levels of genes associated with the JAK/STAT signaling pathway, such as JAK3, STAT6, SOCS3, and JAK1, were higher, while the expression levels of JAK2, STAT1, TBX21, and IL-2 were lower. Suppressors of cytokine signaling (SOCS) are a family of negative regulatory molecules in the JAK/STAT signaling pathway, consisting of eight members (CIS and SOCS1-SOCS7), which participate in the proliferation and differentiation of T helper lymphocyte subpopulations by regulating cytokine signaling pathways [39]. Among them, SOCS3 plays a crucial role in Th1 and Th2 cell differentiation by inhibiting STAT4 signaling through binding to Tyr-800 on the IL-12 receptor beta 2 chain, thereby promoting Th2 cell differentiation [40]. The TPS protein from Haemonchus contortus can significantly inhibit the proliferation of goat PBMCs and the expression of INF-γ, IL-9, and IL-2 in a dose-dependent manner, while promoting IL-10 transcription. Further research found that rHc-TPS can significantly activate STAT3, thereby promoting the expression of SOCS3. Thus, the IL-10/STAT3/SOCS3 axis may be an important mechanism by which Haemonchus contortus evades host immune attack [41]. Infection with Trichuris muris in mice leads to increased expression of SOCS3, and a lack of SOCS3 results in a lower worm burden [42]. In patients infected with filarial parasites, levels of IFN-γ significantly decrease, while the expression of IL-4, IL-5, and IL-10 increases. In infected patients, the expression of SOCS-1, SOCS-5, and SOCS-7 in T cells is significantly reduced, while SOCS-3 expression increases. Therefore, the impaired Th1 response observed in lymphatic filariasis is closely related to the decreased expression of T-bet, SOCS-1, SOCS-5, and SOCS-7, along with the increased expression of SOCS-3 in T cells [43]. This suggests that SOCS3 may play an important role in regulating the balance between Th1 and Th2 cells, aiding in the understanding of the immune evasion mechanisms of parasites during host-parasite interactions, and providing insights for discovering new drug targets.
The JAK/STAT signaling pathway is mainly regulated by cytokines, which participate in the regulation of intracellular signal transduction and are crucial for modulating adaptive immune responses. This pathway is related to biological processes such as cell differentiation, proliferation, apoptosis, and immune regulation [44,45]. For example, studies have shown that IL-4 promotes the expression of GATA3 by activating STAT6, thereby modulating T cell proliferation and Th2 cell differentiation [46]. After infection with Trichinella spiralis, IL-4 levels in the mesenteric lymph nodes of STAT6-/- mice were significantly reduced [47]. Similarly, in the culture supernatant of splenocytes from STAT6-/- mice infected with Schistosoma mansoni cercariae, a similar phenomenon was observed, indicating a significant suppression of the Th2-type immune response [48]. The transthyretin (TTR) protein found in the excretory/secretory products of the Haemonchus contortus can block IL-4 induced proliferation of goat PBMCs. In addition, HcTTR can also significantly downregulate the mRNA transcription levels of JAK2 and STAT6 in the JAK/STAT signaling pathway [49]. This indicates that STAT6 plays an important role in the Th2-type immune response.Our study revealed similar findings, compared to the 0h control group, TBX21 protein expression was significantly increased in the 24h experimental group, while levels of p-JAK3, p-STAT6, and GATA3 protein expression were notably elevated in the 48h experimental group, promoting the differentiation of T cells towards Th2 cells. This phenomenon suggests that cestode infection may activate the JAK/STAT signaling pathway, prompting the host to produce IFN-γ and generate Th1-type immune responses in the early stages of infection. However, during long-term infection, the activation of STAT6 leads to the secretion of large amounts of IL-4 and IL-10, further promoting the proliferation and differentiation of Th2 cells. This results in the excessive activation of Th2 responses and the weakening of Th1 responses, causing an imbalance in the Th1/Th2 cell ratio and a decrease in host immune function, allowing cysticerci to evade immune defenses and persistently survive within the host, maintaining a state of chronic infection.
5. Conclusion
In summary, this study found that the TPx protein from porcine cysticerci can induce Jurkat cells to differentiate into Th1 cells at 24h and into Th2 cells at 48h. Further analysis of the differential transcriptomic data regarding the effects of TPx protein on Jurkat cells revealed that the JAK/STAT signaling pathway may play an important role in the regulation of Th1/Th2 cell imbalance. Western blot assays indicated that the TPx protein can upregulate the expression of p-JAK3 and p-STAT6 at 48h, promoting the expression of the Th2 transcription factor GATA3, thereby inducing Th2 cell differentiation. This suggests that the JAK/STAT signaling pathway plays a key role in the TPx protein-induced Th1/Th2 cell imbalance, paving the way for further research in this field.
Author Contributions
Q.M. and H.C conducted the experiments. Q.M, H.C, H.X and B.Z designed the experiments. Q.M analyzed the data. H.C and H.X assisted with the experiments. Q.M wrote the manuscript. B.Z revised the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This study was funded by National Natural Science Foundation of China (No.82460402); National Natural Science Foundation of China (No. 81960378); Project of Basic Research Program of Guizhou Provincial Department of Science and Technology ( [2023]-516); Project of Guizhou Province Graduate Education Innovation (2024YJSKYJJ312); Project of Zunyi Medical University Graduate Education Innovation (ZYK208).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors without undue reservation.
Acknowledgments
We thank Shanghai OE Biotech Co., Ltd. (Shanghai, China) for supporting differential transcriptomics analysis.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Del Brutto, O.H. Human neurocysticercosis: an overview. Pathogens 2022, 11, 1212. [Google Scholar] [CrossRef] [PubMed]
- Pujari, A.; Bhaskaran, K.; Modaboyina, S.; et al. Cysticercosis in ophthalmology. Surv Ophthalmol 2022, 67, 544–569. [Google Scholar] [CrossRef]
- Ta, R.; Blond, B.N. The prevalence of and contributors to neurocysticercosis in endemic regions. J Neurol Sci 2022, 441, 120393. [Google Scholar] [CrossRef] [PubMed]
- Butala, C.; Brook, T.M.; Majekodunmi, A.O.; et al. Neurocysticercosis: current perspectives on diagnosis and management. Front Vet Sci 2021, 8, 615703. [Google Scholar] [CrossRef] [PubMed]
- Kaur, R.; Arora, N.; Rawat, S.S.; et al. Vaccine for a neglected tropical disease Taenia solium cysticercosis: fight for eradication against all odds. Expert Rev Vaccines 2021, 20, 1447–1458. [Google Scholar] [CrossRef] [PubMed]
- Sergio, R.M.; Elizabeth, M.E.; Omar, G.O.; et al. Transplastomic plants yield a multicomponent vaccine against cysticercosis. J Biotechnol 2018, 266, 124–132. [Google Scholar]
- Nuamtanong, S.; Reamtong, O.; Phuphisut, O.; et al. Transcriptome and excretory-secretory proteome of infective-stage larvae of the nematode Gnathostoma spinigerum reveal potential immunodiagnostic targets for development. Parasite 2019, 26, 34. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.M.; Zhou, B.Y. Research progress on the relationship between tapeworm excretion secretions and host immune effect. Chin J Parasitol Parasit Dis 2020, 38, 128–133. [Google Scholar]
- Fan, X.; Zhang, Y.; Ouyang, R.; et al. Cysticercus cellulosae regulates T-cell responses and interacts with the host immune system by excreting and secreting antigens. Front Cell Infect Microbiol 2021, 11, 728222. [Google Scholar] [CrossRef] [PubMed]
- Li, L.Z. Effect of proteomics-based cysticercus cellulosae excretory secretory antigen LRRC15 protein on T-cell immune response in piglets. Zunyi: Zunyi Medical University, 2022.
- He, W.; Li, L.Z.; Sun, X.Q.; et al. Screening, validation, T-cellantigenicepitopes prediction and eukaryoticexpression of cysticercus cellulosae excretory-secretory antigen thioredoxin peroxidase protein. Journal of Pathogen Biology 2023, 18, 174–179. [Google Scholar]
- Li, L.; He, W.; Fan, X.; et al. Proteomic analysis of Taenia solium cysticercus and adult stages. Front Vet Sci 2023, 9, 934197. [Google Scholar] [CrossRef]
- Molina-López, J.; Jiménez, L.; Ochoa-Sánchez, A.; et al. Molecular cloning and characterization of a 2-Cys peroxiredoxin from Taenia solium. J Parasitol 2006, 92, 796–802. [Google Scholar] [CrossRef]
- Yin, C.; Luo, X.N.; Wang, S.; et al. Prokaryotic expression and biological properties of thioredoxin peroxidase from Taenia solium. Acta Veterinaria et Zootechnica Sinica 2014, 45, 1512–1517. [Google Scholar]
- He, W.; Sun, X.; Luo, B.; et al. Regulation of piglet T-cell immune responses by thioredoxin peroxidase from cysticercus cellulosae excretory-secretory antigens. Front Microbiol 2022, 13, 1019810. [Google Scholar] [CrossRef] [PubMed]
- Gause, W.C.; Rothlin, C.; Loke, P. Heterogeneity in the initiation, development and function of type 2 immunity. Nat Rev Immunol 2020, 20, 603–614. [Google Scholar] [CrossRef] [PubMed]
- Loke, P.; Harris, N.L. Networking between helminths, microbes, and mammals. Cell Host Microbe 2023, 31, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Haddad, E.B.; Cyr, S.L.; Arima, K.; et al. Current and emerging strategies to inhibit type 2 inflammation in atopic dermatitis. Dermatol Ther (Heidelb) 2022, 12, 1501–1533. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.R.; Zhang, R.H.; Li, R.; et al. The effects of helminth infections against type 2 diabetes. Parasitol Res 2021, 120, 1935–1942. [Google Scholar] [CrossRef] [PubMed]
- Bhengu, K.N.; Naidoo, P.; Singh, R.; et al. Immunological interactions between intestinal helminth infections and tuberculosis. Diagnostics (Basel) 2022, 12, 2676. [Google Scholar] [CrossRef] [PubMed]
- Mu, Q.Q.; Zhou, B.Y. Progress of researches on molecular mechanisms underlying helminth infection-mediated type 1/2 host immune responses. Chinese Journal of Schistosomiasis Control 2023, 35, 534–538. [Google Scholar] [PubMed]
- Wen, T.H.; Tsai, K.W.; Wu, Y.J.; et al. The framework for human host immune responses to four types of parasitic infections and relevant key JAK/STAT signaling. Int J Mol Sci 2021, 22, 13310. [Google Scholar] [CrossRef]
- Janssen, L.; Silva Santos, G.L.; Muller, H.S.; et al. Schistosome-derived molecules as modulating actors of the immune system and promising candidates to treat autoimmune and inflammatory diseases. J Immunol Res 2016, 5267485. [Google Scholar] [CrossRef]
- McSorley, H.J.; Hewitson, J.P.; Maizels, R.M. Immunomodulation by helminth parasites: defining mechanisms and mediators. Int J Parasitol 2013, 43, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Peón, A.N.; Ledesma-Soto, Y.; Olguín, J.E.; et al. Helminth products potently modulate experimental autoimmune encephalomyelitis by downregulating neuroinflammation and promoting a suppressive microenvironment. Mediators Inflamm 2017, 8494572. [Google Scholar] [CrossRef]
- Tang, C.L.; Yu, X.H.; Li, Y.; et al. Schistosoma japonicum soluble egg antigen protects against type 2 diabetes in Lepr db/db mice by enhancing regulatory T cells and Th2 cytokines. Front Immunol 2019, 10, 1471. [Google Scholar] [CrossRef] [PubMed]
- Hasby, E.A.; Hasby Saad, M.A.; Shohieb, Z.; et al. FoxP3+ T regulatory cells and immunomodulation after Schistosoma mansoni egg antigen immunization in experimental model of inflammatory bowel disease. Cell Immunol 2015, 295, 67–76. [Google Scholar] [CrossRef]
- Smallwood, T.B.; Giacomin, P.R.; Loukas, A.; et al. Helminth immunomodulation in autoimmune disease. Front Immunol 2017, 8, 453. [Google Scholar] [CrossRef]
- Chen, J.L.; Nong, G.M. Advances in the application of Jurkat cell models in infectious disease research. Chinese Journal of Contemporary Pediatrics 2018, 20, 236–242. [Google Scholar] [PubMed]
- Prodjinotho, U.F.; Lema, J.; Lacorcia, M.; et al. Host immune responses during Taenia solium neurocysticercosis infection and treatment. PLoS Negl Trop Dis 2020, 14, e0008005. [Google Scholar] [CrossRef]
- Prasad, K.N. My experience on taeniasis and neurocysticercosis. Trop Parasitol 2021, 11, 71–77. [Google Scholar] [CrossRef]
- Verma, A.; Prasad, K.N.; Cheekatla, S.S.; et al. Immune response in symptomatic and asymptomatic neurocysticercosis. Med Microbiol Immunol 2011, 200, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Zhu, J. CD4 T helper cell subsets and related human immunological disorders. Int J Mol Sci 2020, 21, 8011. [Google Scholar] [CrossRef]
- Fan, X.M.; Zhou, B.Y. Research progress of immunoregulation of T lymphocytes in cysticercosis. Chin J Endemiol 2021, 40, 164–168. [Google Scholar]
- Williams, D.L.; Asahi, H.; Botkin, D.J.; et al. Schistosome infection stimulates host CD4(+) T helper cell and B-cell responses against a novel egg antigen, thioredoxin peroxidase. Infect Immun 2001, 69, 1134–1141. [Google Scholar] [CrossRef] [PubMed]
- Tian, A.L.; Tian, X.; Chen, D.; et al. Modulation of the functions of goat peripheral blood mononuclear cells by fasciola gigantica thioredoxin peroxidase In Vitro. Pathogens 2020, 9, 758. [Google Scholar] [CrossRef] [PubMed]
- Jin, Q.W.; Zhang, N.Z.; Li, W.H.; et al. Trichinella spiralis thioredoxin peroxidase 2 regulates protective Th2 immune response in mice by directly inducing alternatively activated macrophages. Front Immunol 2020, 11, 2015. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Zaragoza, M.; Jiménez, L.; Hernández, M.; et al. Protein expression profile of Taenia crassiceps cysticerci related to Th1- and Th2-type responses in the mouse cysticercosis model. Acta Trop 2020, 212, 105696. [Google Scholar] [CrossRef]
- Ge, G.; Yang, Z.Y.; Zhang, X.Y.; et al. Research progress on the effect of SOCS on Th cell differentiation in infectious diseases by regulating JAK/STAT pathway. Chinese Journal of Mycology 2021, 16, 51–55. [Google Scholar]
- Palmer, D.C.; Restifo, N.P. Suppressors of cytokine signaling (SOCS) in T cell differentiation, maturation, and function. Trends Immunol 2009, 30, 592–602. [Google Scholar] [CrossRef]
- Wen, Z.; Aleem, M.T.; Aimulajiang, K.; et al. The GT1-TPS structural domain protein from Haemonchus contortus could be suppressive antigen of goat PBMCs. Front Immunol 2022, 12, 787091. [Google Scholar] [CrossRef] [PubMed]
- Shaw, E.J.; Smith, E.E.; Whittingham-Dowd, J.; et al. Intestinal epithelial suppressor of cytokine signaling 3 (SOCS3) impacts on mucosal homeostasis in a model of chronic inflammation. Immun Inflamm Dis 2017, 5, 336–345. [Google Scholar] [CrossRef]
- Babu, S.; Kumaraswami, V.; Nutman, T.B. Transcriptional control of impaired Th1 responses in patent lymphatic filariasis by T-box expressed in T cells and suppressor of cytokine signaling genes. Infect Immun 2005, 73, 3394–3401. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Liang, X.; Shaikh, A.S.; et al. JAK/STAT signal transduction: promising attractive targets for immune, inflammatory and hematopoietic diseases. Curr Drug Targets 2018, 19, 487–500. [Google Scholar] [CrossRef] [PubMed]
- Stark, G.R.; Cheon, H.; Wang, Y. Responses to cytokines and interferons that depend upon JAKs and STATs. Cold Spring Harb Perspect Biol 2018, 10, a028555. [Google Scholar] [CrossRef] [PubMed]
- Karpathiou, G.; Papoudou-Bai, A., Ferrand E; et al. STAT6: a review of a signaling pathway implicated in various diseases with a special emphasis in its usefulness in pathology. Pathol Res Pract 2021, 223, 153477. [Google Scholar] [CrossRef] [PubMed]
- Li, W.L.; Cao, J.; Lan, P.; et al. Effect of Trichinella spiralis infection on expression and distribution of colonic epithelial E-cadherin in mice and its mechanism. Chinese Journal of Gastrointestinal Surgery 2013, 16, 386–389. [Google Scholar]
- Osada, Y.; Horie, Y.; Nakae, S.; et al. STAT6 and IL-10 are required for the anti-arthritic effects of Schistosoma mansoni via different mechanisms. Clin Exp Immunol 2019, 195, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Lu, M.; Wang, W.; et al. HcTTR: a novel antagonist against goat interleukin 4 derived from the excretory and secretory products of Haemonchus contortus. Vet Res 2019, 50, 42. [Google Scholar] [CrossRef]
Figure 1.
Effect of TPx protein on the differentiation of Th1 and Th2 cells (n=3, x±S, %). Note: A: Effect of TPx protein on Th1 cell differentiation; B: Effect of TPx protein on Th2 cell differentiation; ***means P < 0.0001; ns means no significant difference.
Figure 1.
Effect of TPx protein on the differentiation of Th1 and Th2 cells (n=3, x±S, %). Note: A: Effect of TPx protein on Th1 cell differentiation; B: Effect of TPx protein on Th2 cell differentiation; ***means P < 0.0001; ns means no significant difference.
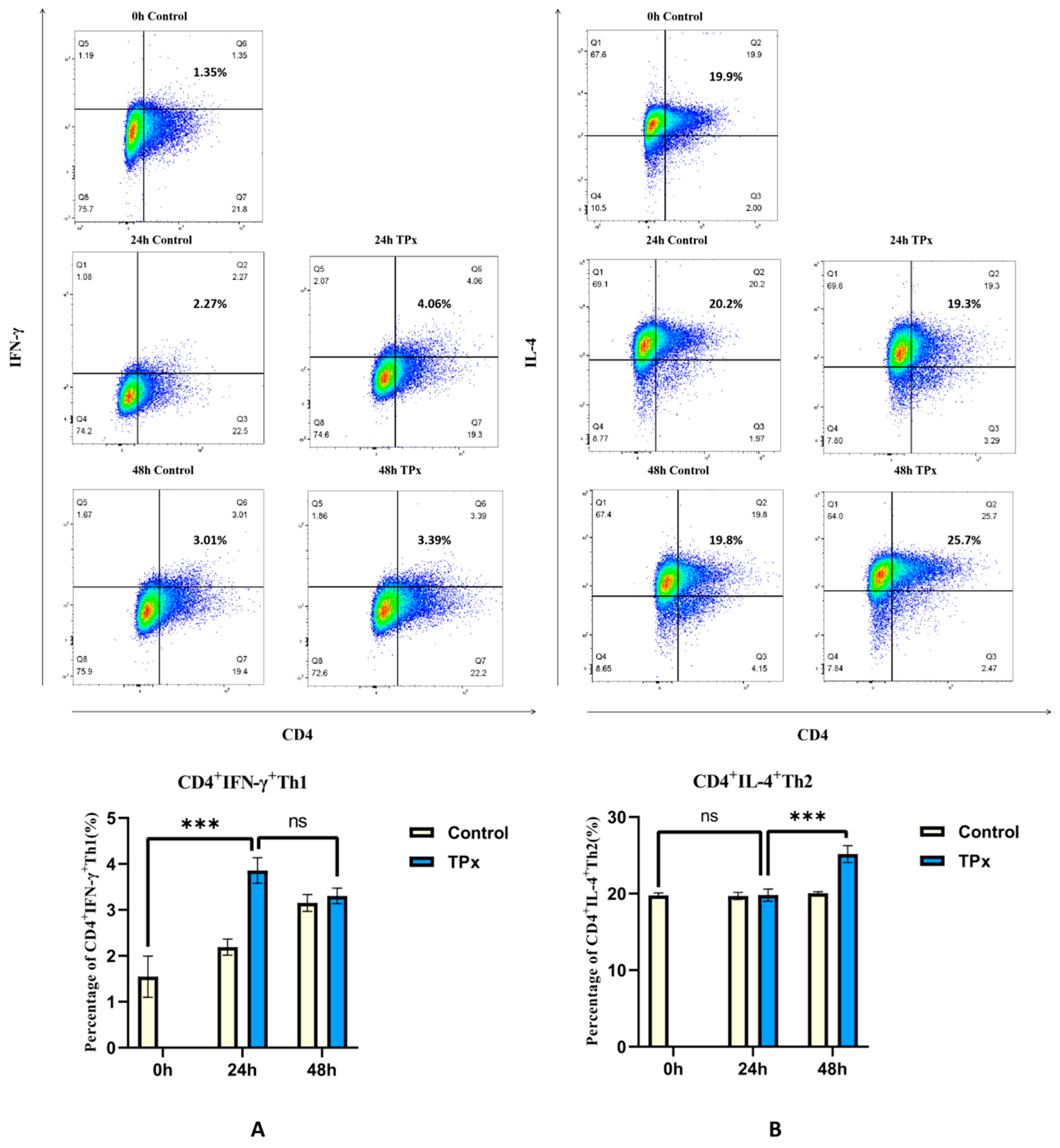
Figure 2.
Differential gene volcano plot. Note: A: Volcano plot of the 24-hour experimentalgroup compared with the 0-hour control group; B: Volcano plot of the 48-hour experimentalgroup compared with the 24-hour experimental group; Each point in the plot represents a detected gene probe, reflecting the differential expression generated by the comparison on the volcano plot.
Figure 2.
Differential gene volcano plot. Note: A: Volcano plot of the 24-hour experimentalgroup compared with the 0-hour control group; B: Volcano plot of the 48-hour experimentalgroup compared with the 24-hour experimental group; Each point in the plot represents a detected gene probe, reflecting the differential expression generated by the comparison on the volcano plot.
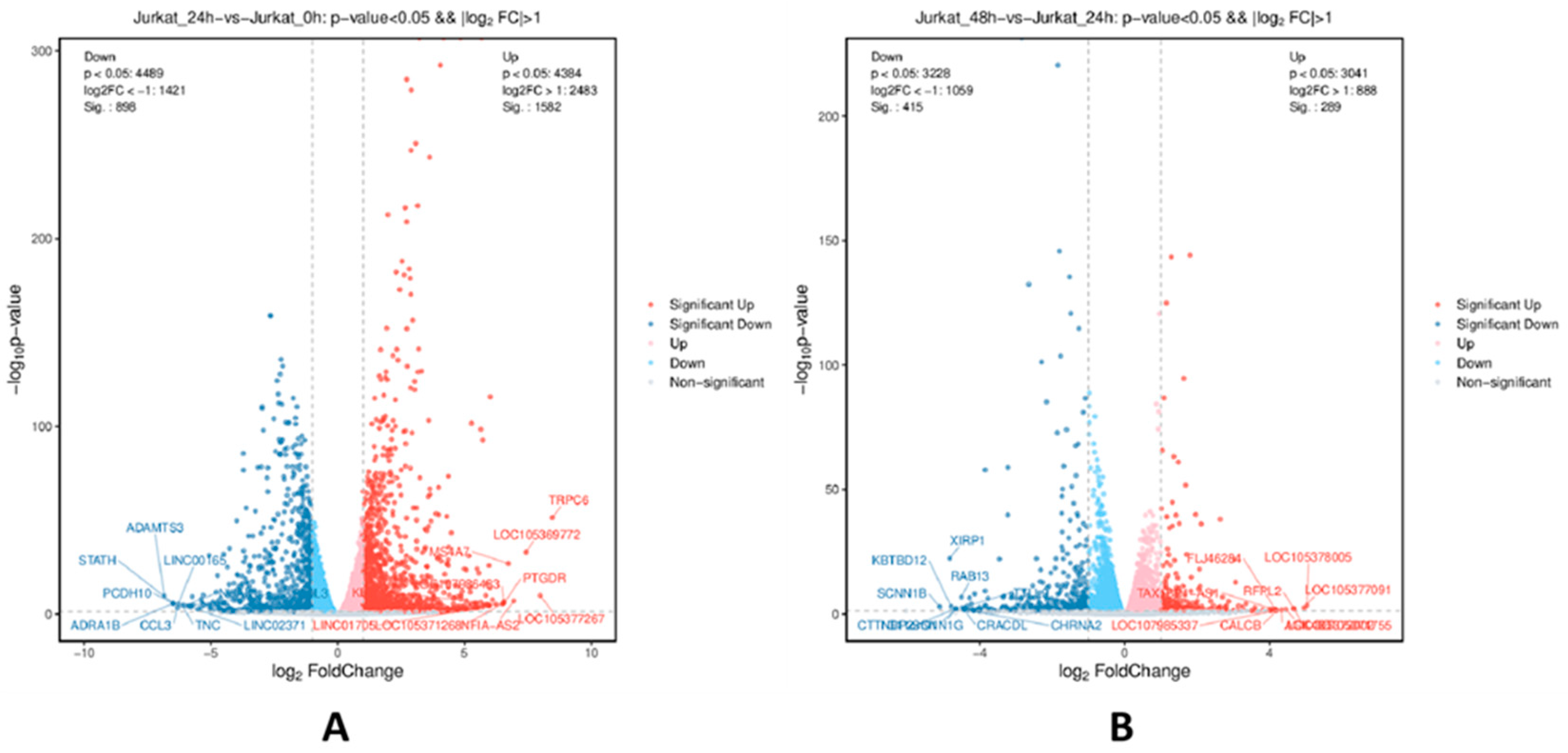
Figure 3.
Top 30 enriched GO terms results of differentially expressed genes. Note: A: Top 30 enriched GO terms of the 24-hour experimental group compared with the 0-hour control group; B: Top 30 enriched GO terms of the 48-hour experimental group compared with the 24-hour experimental group; The horizontal axis represents -log10 p-values, and the vertical axis represents enriched GO terms.
Figure 3.
Top 30 enriched GO terms results of differentially expressed genes. Note: A: Top 30 enriched GO terms of the 24-hour experimental group compared with the 0-hour control group; B: Top 30 enriched GO terms of the 48-hour experimental group compared with the 24-hour experimental group; The horizontal axis represents -log10 p-values, and the vertical axis represents enriched GO terms.
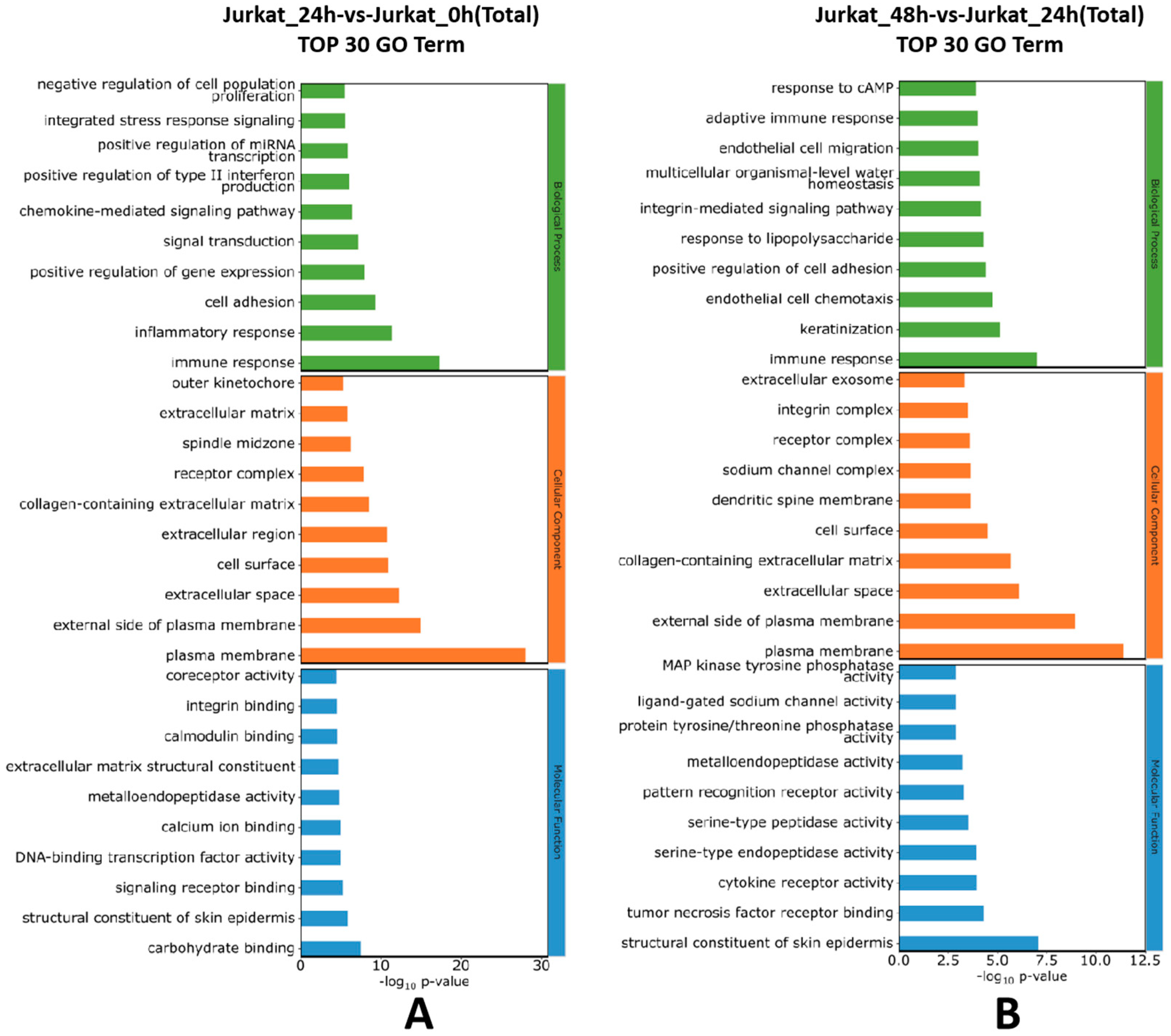
Figure 4.
Bubble plots of the top 20 KEGG enrichment results of differentially expressed genes. Note: A: Bubble plot comparing KEGG pathways between the 24-hour experimental group and the 0-hour control group; B: Bubble plot comparing KEGG pathways between the 48-hour experimental group and the 24-hour experimental group.
Figure 4.
Bubble plots of the top 20 KEGG enrichment results of differentially expressed genes. Note: A: Bubble plot comparing KEGG pathways between the 24-hour experimental group and the 0-hour control group; B: Bubble plot comparing KEGG pathways between the 48-hour experimental group and the 24-hour experimental group.
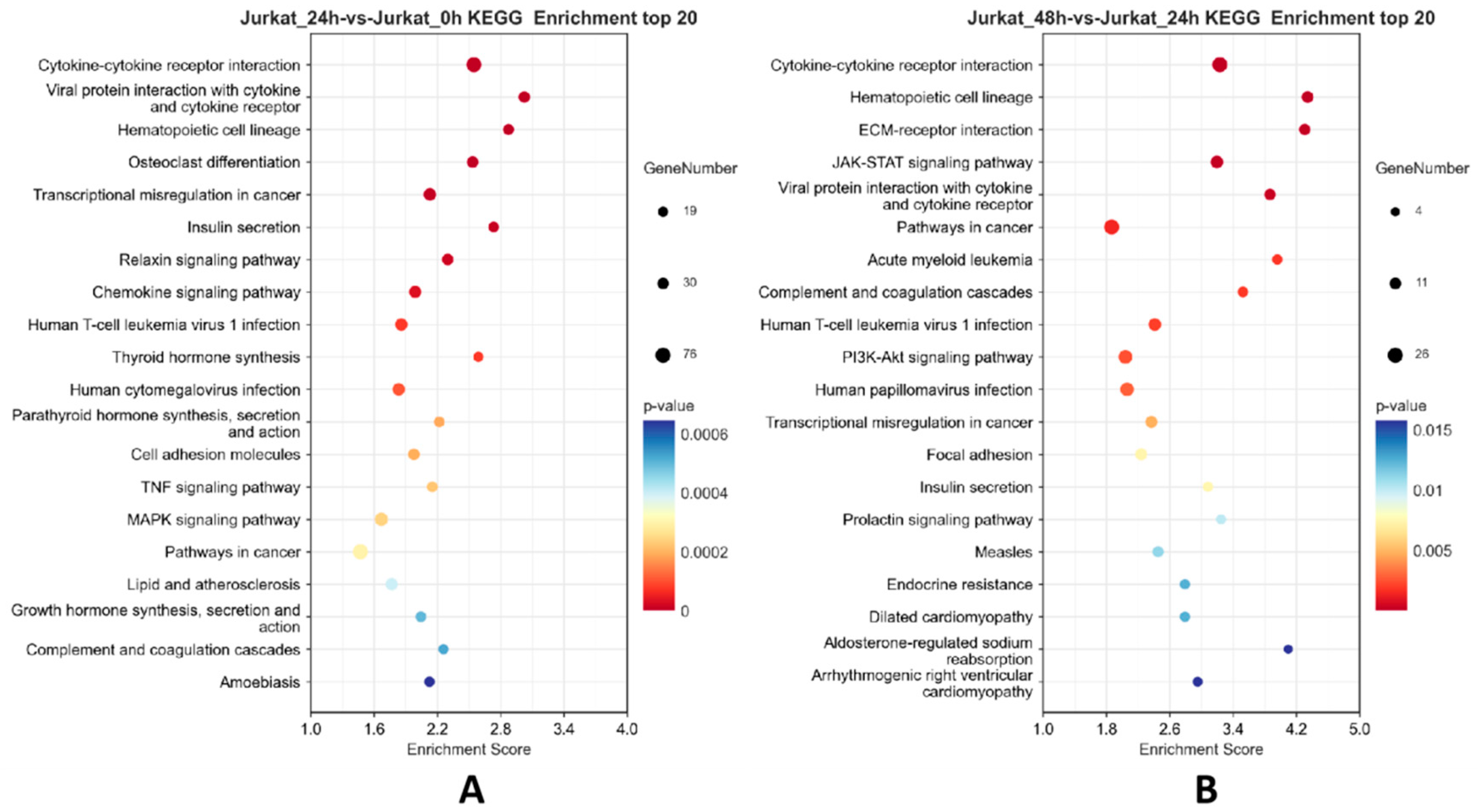
Figure 5.
Heatmap of differential genes related to the JAK/STAT signaling pathway and Th1, Th2 cell differentiation. Note: “Jurkat_0h” represents the 0-hour control group, “Jurkat_24h” represents the 24-hour experimental group, and “Jurkat_48h” represents the 48-hour experimental group. In the image, red color indicates genes encoding proteins with relatively high expression, while blue color indicates genes encoding proteins with relatively low expression.
Figure 5.
Heatmap of differential genes related to the JAK/STAT signaling pathway and Th1, Th2 cell differentiation. Note: “Jurkat_0h” represents the 0-hour control group, “Jurkat_24h” represents the 24-hour experimental group, and “Jurkat_48h” represents the 48-hour experimental group. In the image, red color indicates genes encoding proteins with relatively high expression, while blue color indicates genes encoding proteins with relatively low expression.
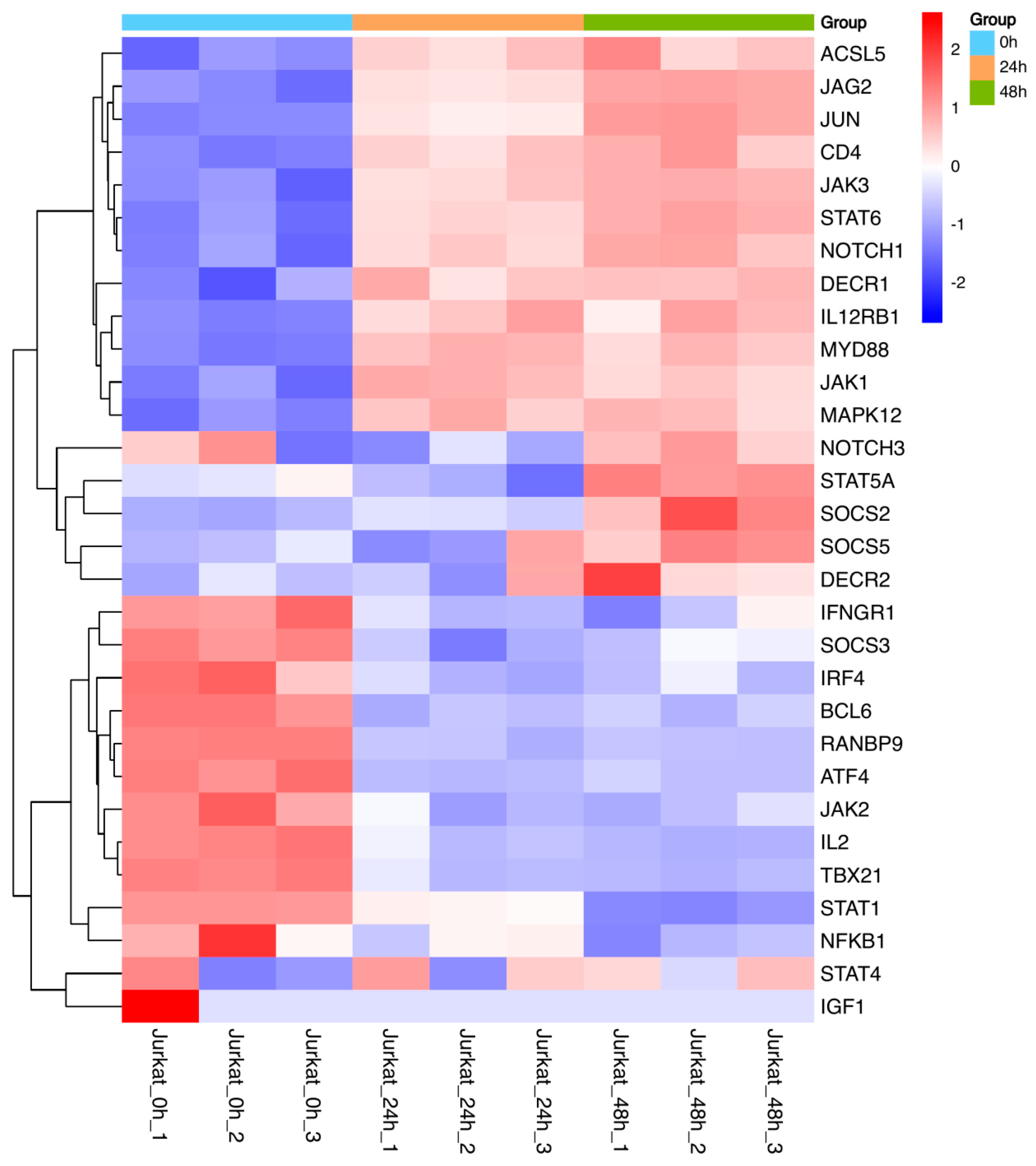
Figure 6.
Effect of TPx protein on the expression of proteins related to the JAK/STAT signaling pathway (n=3, x±S). Note:A: p-JAK3/JAK3 ratio; B: p-STAT6/STAT6 ratio; C: GATA3/GAPDH ratio; D:TBX21/GAPDH ratio; * means P < 0.05; ** means P < 0.01;**** means P < 0.0001;ns means no significant difference.
Figure 6.
Effect of TPx protein on the expression of proteins related to the JAK/STAT signaling pathway (n=3, x±S). Note:A: p-JAK3/JAK3 ratio; B: p-STAT6/STAT6 ratio; C: GATA3/GAPDH ratio; D:TBX21/GAPDH ratio; * means P < 0.05; ** means P < 0.01;**** means P < 0.0001;ns means no significant difference.
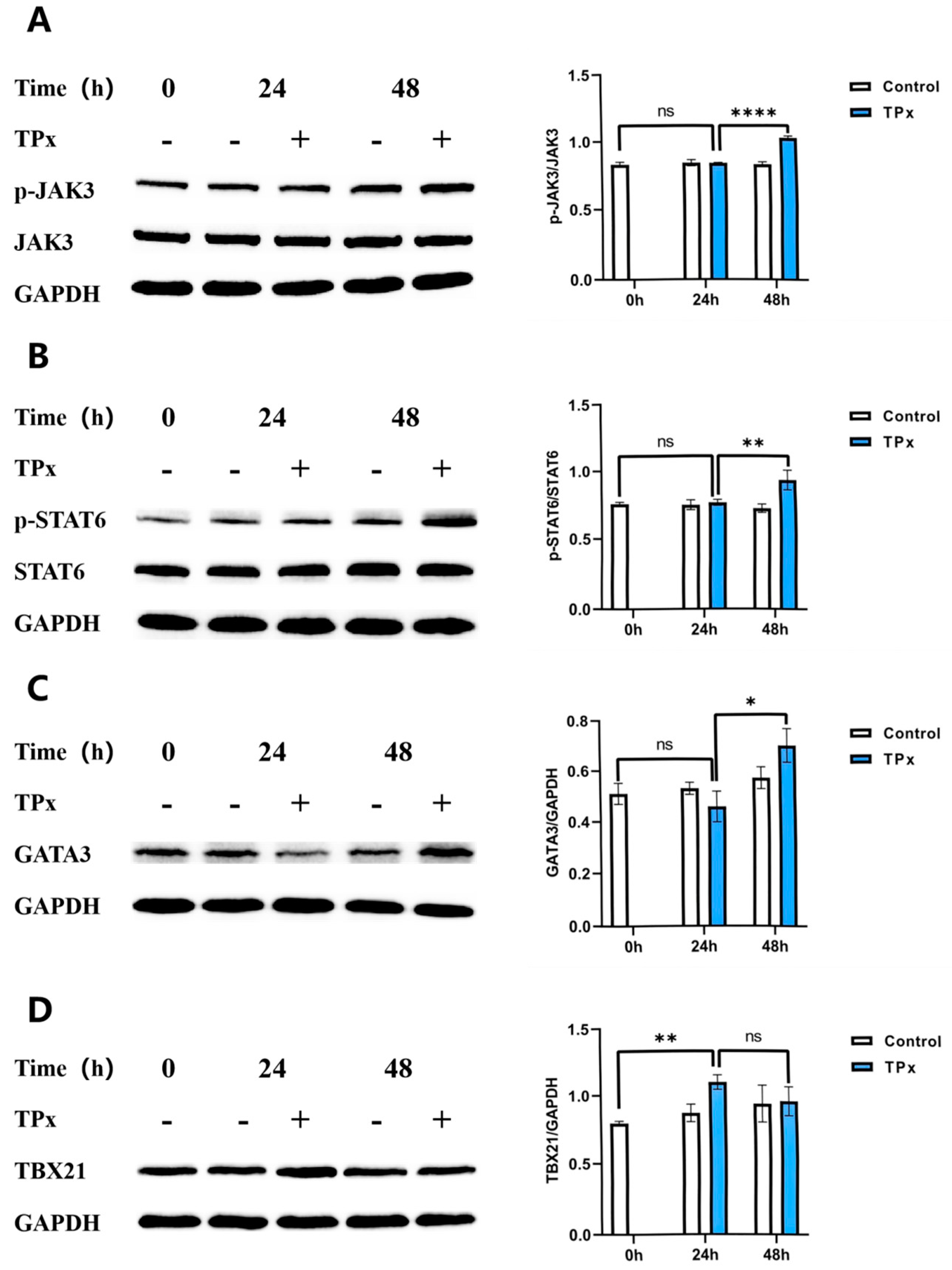
Figure 7.
The effect JAK3-IN-6 inhibitors on expression of JAK/STAT signaling pathway-related proteins (n=3,x±S) Note:A: p-JAK3/JAK3 ratio; B: p-STAT6/STAT6 ratio; C: GATA3/GAPDH ratio; D:TBX21/GAPDH ratio; * means P < 0.05; ** means P < 0.01;*** means P < 0.001.
Figure 7.
The effect JAK3-IN-6 inhibitors on expression of JAK/STAT signaling pathway-related proteins (n=3,x±S) Note:A: p-JAK3/JAK3 ratio; B: p-STAT6/STAT6 ratio; C: GATA3/GAPDH ratio; D:TBX21/GAPDH ratio; * means P < 0.05; ** means P < 0.01;*** means P < 0.001.
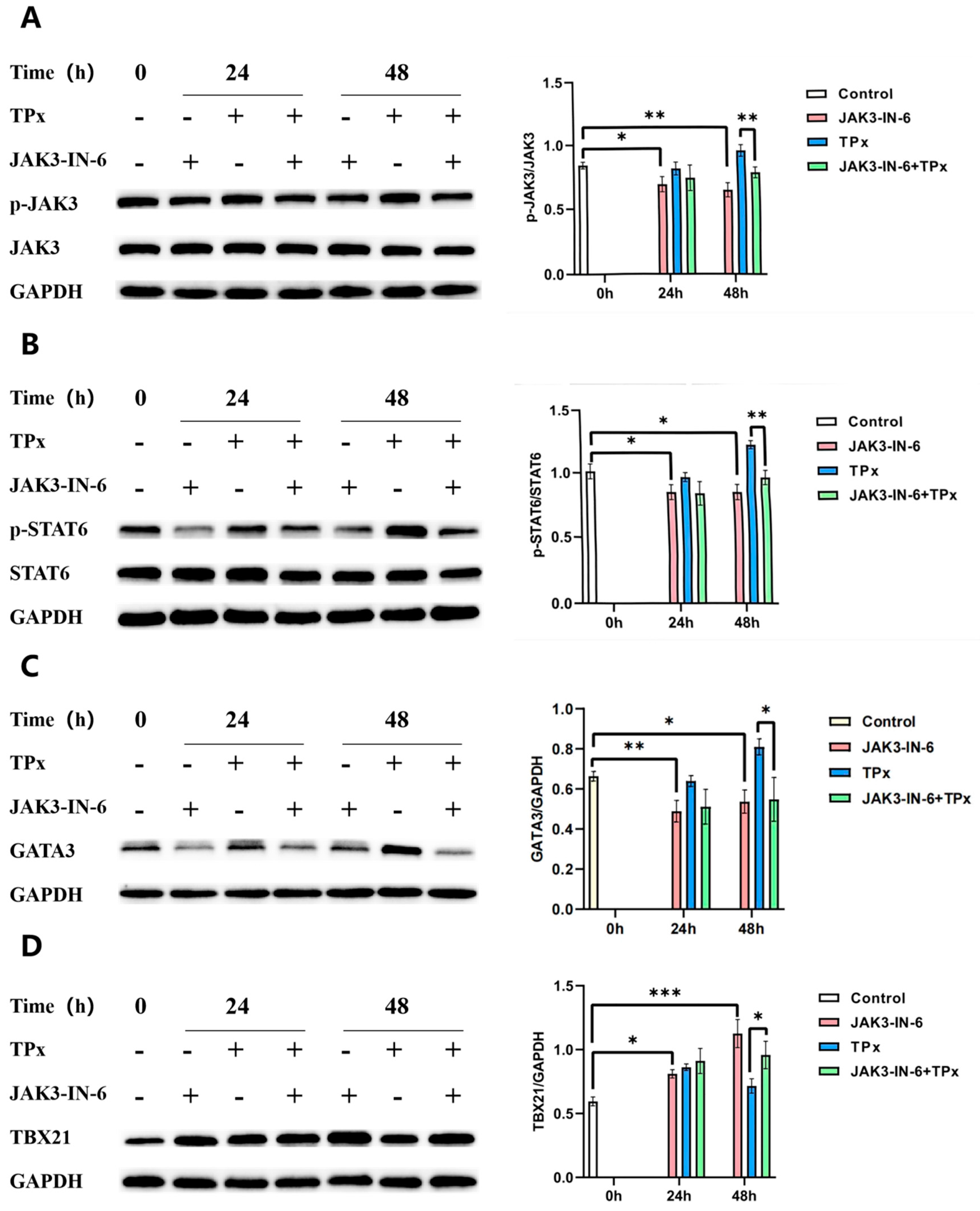

Table 1.
Number of differentially expressed genes.
| Compared groups | Regulated type | q-value<0.05&|log2FC|>1.0 |
|---|---|---|
| Jurkat_24h-vs-Jurakt_0h | Up-regulated | 631 |
| Down-regulated | 304 | |
| Jurkat_48h-vs-Jurakt_24h | Up-regulated | 199 |
| Down-regulated | 248 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



