
Submitted:
08 February 2025
Posted:
10 February 2025
You are already at the latest version
Abstract
Antibiotic resistance is a major problem worldwide, especially with the overuse and misuse of antibiotics. This makes it more challenging to treat patients infected with antibiotic-resistant bacteria, with more costs on the health systems. This review article describes extended-spectrum β-lactam antibiotics as the mostly used antibacterial agents, and how bacteria developed beta-lactamases (ESBLs) to resist these antibiotics. The review focuses more on the problem of ESBLs in Kuwait to uncover which ESBLs are present and what ESBLs gene mutations have been found in this country. Literature review, surprisingly, revealed a limited number of studies in Kuwait on ESBLs gene mutations, published over more than 25 years. The results showed that Kuwait has an alarming number of bacterial strains with ESBLs gene mutations, but the results showed different mutations in different bacterial strains from different types of specimens collected from different hospitals in Kuwait. The data from these studies were scattered and not linked or analysed together to highlight the big picture of the problem of ESBLs in Kuwait. This review article alerts that the amount of research on ESBLs in Kuwait is not up to the expectations of a country like Kuwait, which has high-standard research facilities. Baring in mind that the geographic area of Kuwait is relatively small, the authors of this paper think that there might be more β-lactam bacteria spreading in Kuwait, which need to be explored, and that more research and more collaboration is needed among researchers in Kuwait to address this risky situation.
Keywords:
ESBL-Mutations
; SHV-112
; SHV-122
; Kuwait
1. Brief History of Antibiotics
In 1928, the Scottish bacteriologist Alexander Fleming discovered that a fungus (Penicillium notatum) could prevent the growth of Staphylococci bacteria in vitro by producing a chemical that had an antibacterial effect. That chemical was called “Penicillin”, the first scientifically identified antibiotic in history. After discovery, extensive studies were conducted to isolate and purify penicillin, and then extract adequate amounts to perform experimental animal studies and clinical trials on humans to prove the efficacy of Penicillin as a treatment for bacterial infections, which was achieved by the mid-1940s [1]. Between the 1940s and 1960s, more antibiotics were developed and used for treating patients. That period is usually referred to as the “golden era of antibiotic research”, during which more than half of the currently known antibiotics were developed and used as antibacterial treatments [1,2]. More than 20 new classes of antibiotics were introduced during that era [3]. This had an enormous positive effect in saving the lives of millions of people worldwide. For example, by the year 1944, the rate of production of penicillin was over a billion doses yearly [4]. The survival rate of bacterial pneumonia dramatically increased from 20% to 85% during that period. It is estimated that the use of antibiotics extended the average human lifespan by 23 years. Although the main purpose of antibiotics is treating patients with bacterial infections, it is also widely used as preventive medicine in cases of surgery, cancer, organ transplantation, pre-term babies, and so on [2].
2. Antibiotic Resistance
Like many great things in life, something could happen to disrupt that triumph. Several bacteria could evolve and develop mechanisms to resist the actions of antibiotics, a phenomenon known currently as “antibiotic resistance”. Surprisingly, this happened very quickly after the widespread use of penicillin, in the early 1940s, when strains of E. coli and S. aureus were found to be penicillin-resistant [1,5,6]. Still, many scientists found indications that such resistance was buried inside ancient genes in the bacterial genome but were activated later, after the exposure to antibiotics [7]. This encouraged scientists to develop new generations of antibiotics that could overcome such resistance. Still, the battle between bacteria and antibiotics continues and more bacteria are found to resist more antibiotics. Common mechanisms used by the bacteria to resist antibiotics include inactivation or breakdown of the antibiotics by bacterial enzymes, changing the bacterial components that are targeted by the antibiotics, or changing the bacterial membrane permeability of the antibiotics, by preventing intake of antibiotics or increasing antibiotic efflux out of the bacterial cell [7,8,9].
Antibiotic resistance has become a major problem worldwide, especially with the overuse and misuse of antibiotics. What makes this problem more problematic is that bacteria were found to transfer resistance genes among themselves [9]. Currently, a good number of bacteria are found to be resistant to almost all available antibiotics; such multi-drug resistant bacteria are termed “super-bugs” [1]. This puts more challenges on treating patients infected with these superbugs. For example, it was estimated that yearly more than 2 million people get infected with antibiotic-resistant bacteria in the US alone, leading to more than 23,000 deaths, with a cost of US$7,000-29,000 per patient [9]. In the European Union, more than 25,000 patients die of infections with antibiotic-resistant bacteria every year [7]. Globally, antibiotic-resistant bacterial infections caused 1.27 million deaths in 2019 [10]. In addition, the World Bank estimates that this problem could cost the health systems an additional US$ 1 trillion healthcare costs by the year 2050 [11].
In this review paper, the authors focused on antibiotic resistance due to Extended-spectrum beta-lactamases (ESBLs) gene mutations, with special attention to the situation in Kuwait where the authors live and work.
3. β-Lactam Molecules
In the mid-1940s, the chemical structure of penicillin was determined [12,13] to have a core of a four-membered lactam ring (cyclic amide), with a carbonyl group, as depicted in Figure 1.
The lactam ring in penicillin is of the β type, which consists of 4 atoms: 3 carbon atoms and one nitrogen atom, forming a square-like ring. The nitrogen atom is attached to the β-carbon atom of the ring [4,14,15]. Within a few years after the discovery of penicillin, more similar molecules were found, from natural sources (bacterial or fungal) or were synthesised, that had similar chemical structures as penicillin. All these have the β-lactam ring as the essential core structure, with additional ring structures directly connected to the β-lactam ring, except for monobactams. Examples of these so called beta-lactam antibiotics include second generation penicillins (like oxacillin, methicillin, and dicloxacillin) and third generation penicillins (like Amoxicillin and ampicillin). Moreover, other classes of β-lactam molecules were discovered and introduced for clinical use, which had chemical alterations in the core beta-lactam structure, increasing the spectrum of their activity. Since the late 1970s, more of these beta-lactam antibiotics have been developed and introduced, including the penams, penems, cephems, carbapenems, and monobactams [1,4,15,16].
β-lactam antibiotics are currently the most used antibacterial agents to treat infections. For example, they account for two-thirds of all prescriptions for injectable antibiotics in the USA, half of which are cephalosporins [14].
4. How β-Lactam Antibiotics Work?
The main bactericidal effect of β-lactam antibiotics is exerted by disturbing the synthesis of bacterial cell wall. The bacterial cell wall is made up of peptidoglycans, which give the bacterial cell its shape and integrity, and protect it from outside attacks and osmotic stress [1]. Peptidoglycans are continuously made and remodelled during bacteria life and division, and these molecules should be cross-linked with peptide bonds to form a firm network of proteins that give the bacterial cell wall its strength and stability. This cross-linking is performed by specific bacterial enzymes called transpeptidases (also known as penicillin-binding proteins; PBPs) [1,14]. Different bacterial species have different numbers of transpeptidases, usually 3 to 8 enzymes [14]. Transpeptidases recognise dipeptide moieties in glycans and form peptide linkage among the peptidoglycans in the bacterial cell wall [15]. β-lactam antibiotics, through their β-lactam rings, can bind to the active site of the transpeptidases, usually a serine, preventing them from cross-linking the peptidoglycans. This disrupts the making of the bacterial cell wall, and eventually prevents bacterial growth and proliferation. In addition, the bacteria will be susceptible to lysis due to osmosis [1,15]. In short, β-lactam antibiotics prevent bacterial proliferation and kill the infective bacteria, eliminating the infection in the patient. Another advantage of β-lactam antibiotics is that they only attack the cell wall of prokaryotic cells (the bacterial cells). Therefore, the eukaryotic cells (including human cells) will not be affected [1].
5. ESBLs
The introduction of antibiotics as weapons against bacteria ignited a defensive reaction from the bacteria, which rendered some bacteria to be able to resist the bactericidal action of antibiotics. Regarding -lactam antibiotics, bacteria started producing enzymes that could inactivate the action of -lactam. As penicillin was the first antibiotic to be used in treating infections, penicillin resistance was the first reported antibiotic-resistance; as was mentioned earlier, it was reported just a few years after the widespread use of penicillin [1,5,6]. Penicillin-resistant bacteria managed to counterattack penicillin by producing penicillinases, an enzyme that can hydrolyze the -lactam of the antibiotic, leading to the inactivation of penicillin, as seen in Figure 2.
This involves the formation of an acyl-enzyme intermediate between the serine residue of the penicillinase and the carbonyl carbon of the -lactam ring, which is then hydrolyzed, and hence the antibiotic gets inactivated [17,18]. Penicillin resistance spread quickly. For example, by the late 1960s, more than 80 percent of both community and hospital-acquired strains of S. aureus were penicillin resistant [1]. An increase in penicillin-resistance has compromised the use of penicillin as a single therapeutic agent, and currently, there are limited therapeutic options that can use penicillin as monotherapy [14].
With the emergence of penicillin-resistance, efforts succeeded to extract or semi-synthesize new antibiotics with modifications to penicillin, retaining the -lactam core to attack bacterial cell wall synthesis, and, at the same time, these new antibiotics were not inactivated by penicillinase [1,4,15,16]. However, with every introduction of new antibiotics, bacteria were found to develop resistance by producing enzymes that inactivate these antibiotics in a similar way employed by penicillinase [17,18]. Enzymes against -lactam antibiotics were called -lactamases. With the development of new generations of antibiotics, and new types of -lactam antibiotics against a broad spectrum of bacteria, the bacterial -lactamases are currently known as Extended-spectrum beta-lactamases (ESBLs). These hydrolytic enzymes can break down and inactivate a wide range of -lactam antibiotics, which makes these antibiotics ineffective in treating infections by ESBL-producing bacteria [17].
6. Classifications of ESBLs
Due to the large number of reported ESBLs, which are continuously increasing, there were many attempts to classify these ESBLs [19,20,21]. Ambler (1980) proposed four classes based on amino acid sequencing, which are now known as Ambler classes A, B, C, and D [22]. Another classification was made by Bush et al. (1995), which was based on the functional properties of the enzymes, and it included 4 groups and many subgroups [23]. These classifications are widely used in microbiology. On the other hand, the advancement of molecular biology techniques allowed thorough exploration of the genetics of ESBLs, and therefore it is also common to refer to different ESBLs based on their genomes, as explained next.
7. Genetics of ESBLs
Genes of ESBLs (generally referred to as bla genes) are usually encoded on plasmids, which explains the transfer of these genes among bacteria leading to the rapid widespread of antibiotics-resistance. The ESBL genes have undergone several mutations over time, resulting in a wide array of ESBL variants with different substrate specificities and hydrolytic activities against -lactam antibiotics [21]. ESBLs against most -lactam antibiotics were found to evolve. These were also found to be transferred within the bacterial species, or even across different species, conferring more bacteria to be -lactam-resistant. For example, CTX-M-15, which is a member of the CTX family of ESBLs, was initially found in E. coli; but now it has been reported in other members of the Enterobacteriaceae worldwide [24,25]. Studies have found different ESBL genes that were grouped into several families, the most famous being the TEM (Temoniera), SHV (Sulfhydryl Variable) and CTX-M (Cefotaximase-Munich) families. Others include OXA, GES, PER, VEB, TLA, and many other families [19,21,26]. The first TEM was reported in an E. coli isolate in 1965, and therefore is referred to as TEM-1 [27]. The SHV family is derived from Klebsiella species since its progenitor SHV-1 is universally present in that species [28]. CTX-M has been found mainly in isolates of Salmonella, Typhimurium, and E. coli, as well as other species of Enterobacteriaceae [19]. CTX-M is currently the most common variant of ESBLs [19,21]. Studies revealed several mutations in these genes leading to ESBL variants in different antibiotic-resistant bacterial strains. The continuous discovery of large numbers of new mutations created challenges in coordinating proper nomenclature for these mutations [20]. In the late mid-1990s, George Jacoby and Lahay Clinic created a website to name and record new ESBL gene mutations [29]. However, this website was closed in 2015 and the databases were transferred to the National Center for Biotechnology Information (NCBI), which has now become responsible for β-lactamase numerology [30].
8. ESBLs Mutations in Kuwait
The authors of the paper have contributed to the findings of mutations in ESBLs genes, namely by discovering SHV-112 and SHV-122 in bacterial strains in Kuwait [31,32]; these discoveries happened in 2010 and 2011, respectively. Kuwait is a Middle Eastern country, with a relatively small area, but a very developed healthcare system. -lactam resistance was noticed in clinical samples from different patients in Kuwaiti hospitals. The authors thought to revisit ESBLs 15 years after their discoveries of SHV-112 and SHV-122 to explore the current situation in Kuwait concerning ESBLs and -lactam resistance is, and what kind of ESBL mutations were found and may be prevalent in Kuwait.
The authors used PubMed to search for publications on the ESBLs in Kuwait. The search terms were “Extended-spectrum beta-lactamases in Kuwait” or “ESBLs in Kuwait”. Up to the time of the writing of this review article, a total of 41 publications were found. However, adding the word “genes” to the search bar in PubMed narrowed the results down to 20 publications. Moreover, adding the word “mutations” or even using that word without “genes” brought down the results to 4 publications only: Two of these were research papers published in 2010 by the authors of this review article [31,33]; one paper was a review article by the late Dr. Patrick West, who worked in Kuwait during the publishing of his paper in the year 2000, but the paper itself was a general review on ESBL genes, and not specifically about Kuwait [34]; the last article was a recent research paper published in 2023 [35]. It was surprising to find more than 10-year gaps in publications on ESBL gene mutations from 2000 to 2010, and then in 2023. However, by carefully checking the 41 publications found during the first search, 9 papers did mention some results on ESBLs genes, but were not picked up when the word “genes” was added to the search process. Out of the 41 papers, two were review articles, one of them being the one mentioned above which was published by Dr. West in the year 2000 [34]. The other review article was a gathering of results of publications from the 6 Arab countries present in the Arabian Gulf [36] (which is also referred to as the Persian Gulf in many geographic maps). Among the remaining 39 research publications, 8 studies were not conducted on samples from Kuwait, but some of the co-authors had affiliations with Kuwait and therefore were mentioned by the PubMed search engine. Among the remaining 31 papers, only 13 papers included some molecular techniques to confirm ESBL genes and variants in their samples, but these papers were more focused and elaborated on antibiotic resistance using conventional microbiology lab tests. Table 1 gives a summary of the findings of these 13 research articles that included data on ESBL genes & variants using molecular techniques. Figure 3 shows the timeline of published findings of ESBLs gene & variants between the years 2007 and 2023.
It may be noted from Table 1 and Figure 3 that the number of studies on ESBL gene mutations in Kuwait is not up to the expectations of a country like Kuwait, which has high-standard research facilities. The results, available from the limited number of studies published so far, show that Kuwait has an alarming number of bacterial strains with ESBL gene mutations. However, the data was scattered and not linked or analysed together to highlight the big picture of the problem of ESBLs in Kuwait. What is also alarming is the findings by a recent study which reported multidrug resistant E. coli in samples obtained from 3 sewage sites in Kuwait, many of which showed ESBLs production with presence of TEM and CTX-M [47]. What is more worrying is that some of the sewage in Kuwait is usually discharged directly into the Kuwait sea water, where a lot of fishing for food as well as recreational activities happen. This accounts for a high risk of infections with multidrug-resistant bacteria, at least E. coli according to that study, that may be difficult to treat with fewer treatment options for patients and increase morbidity and mortality rates. More elaborated studies on ESBLs and their gene mutations are needed in Kuwait, and more collaboration is needed among Kuwaiti microbiologists and clinicians to address the problem of antibiotic resistance in Kuwait to draw the appropriate health plans and strategies to deal with this problem in Kuwait. This should be done today and not tomorrow. It is also recommended to collaborate with researchers from the other Arabian Gulf countries to find any spreading of ESBLs across the borders and to unite the efforts to address the problem of antibiotic resistance in this geographic part of the world.
References
- Lobanovska, M.; Pilla, G. Penicillin’s Discovery and Antibiotic Resistance: Lessons for the Future? Yale J Biol Med. 2017, 90, 135–145. [Google Scholar]
- Hutchings, M.I.; Truman, A.W.; Wilkinson, B. Antibiotics: past, present and future. Curr Opin Microbiol. 2019, 51, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Coates, A.R.; Halls, G.; Hu, Y. Novel classes of antibiotics or more of the same? Br J Pharmacol. 2011, 163, 184–94. [Google Scholar] [CrossRef] [PubMed]
- Turner J, Muraoka A, Bedenbaugh M, Childress B, Pernot L, Wiencek M, et al. The Chemical Relationship Among Beta-Lactam Antibiotics and Potential Impacts on Reactivity and Decomposition. Front Microbiol. 2022, 13, 807955. [Google Scholar]
- Abraham, E.P.; Chain, E. An enzyme from bacteria able to destroy penicillin. 1940. Rev Infect Dis. 1988, 10, 677–678. [Google Scholar] [PubMed]
- Rammelkamp, T. Resistance of Staphylococcus aureus to the action of penicillin. Exp Biol Med. 1942, 51, 386–389. [Google Scholar] [CrossRef]
- Aminov, R.I. A brief history of the antibiotic era: lessons learned and challenges for the future. Front Microbiol. 2010, 1, 134. [Google Scholar] [CrossRef]
- Dever, L.A.; Dermody, T.S. Mechanisms of bacterial resistance to antibiotics. Arch Intern Med. 1991, 151, 886–95. [Google Scholar] [CrossRef]
- Reygaert, W.C. An overview of the antimicrobial resistance mechanisms of bacteria. AIMS Microbiol. 2018, 4, 482–501. [Google Scholar] [CrossRef]
- Antimicrobial Resistance Collaborators. Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Jonas, O.B.; Irwin, A.; Berthe, F.C.J.; Le Gall, F.G.; Marquez, P.V. Drug-resistant infections : a threat to our economic future (Vol. 2): final report (English). HNP/Agriculture Global Antimicrobial Resistance Initiative Washington, D.C.: World Bank Group. http://documents.worldbank.org/curated/en/323311493396993758/final-report.
- Hodgkin, D.C. The X-ray analysis of the structure of penicillin. Adv Sci. 1949, 6, 85–9. [Google Scholar]
- Abraham EP, Chain E, Fletcher CM, Gardner AD, Heatley NG, Jennings MA, et al. Further observations on penicillin. Lancet 1941, 238, 177–189. [Google Scholar] [CrossRef]
- Bush, K.; Bradford, P.A. beta-lactams and beta-lactamase inhibitors: an overview. Cold Spring Harb Perspect Med, 2016; 6, a025247. [Google Scholar]
- Kim D, Kim S, Kwon Y, Kim Y, Park H, Kwak K, et al. Structural Insights for β-Lactam Antibiotics. Biomol Ther 2023, 31, 141–147. [Google Scholar] [CrossRef]
- Fernandes R, Amador P, Prudêncio C. β-Lactams chemical structure, mode of action and mechanisms of resistance. Rev Med Microbiol. 2013, 24, 7–17. [Google Scholar] [CrossRef]
- Bush, K.; Jacoby, G.A. Updated functional classification of β-lactamases. Antimicrob Agents Chemother. 2010, 54, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Drawz, S.M.; Bonomo, R.A. Three decades of beta-lactamase inhibitors. Clin Microbiol Rev. 2010, 23, 160–201. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, S.; Fatima, J.; Shakil, S.; Rizvi, S.M.; Kamal, M.A. Antibiotic resistance and extended spectrum beta-lactamases: Types, epidemiology and treatment. Saudi J Biol Sci. 2015, 22, 90–101. [Google Scholar] [CrossRef]
- Bush, K. Classification for β-lactamases: historical perspectives. Expert Rev Anti Infect Ther. 2023, 21, 513–522. [Google Scholar] [CrossRef]
- Paterson, D.L.; Bonomo, R.A. Extended-spectrum beta-lactamases: a clinical update. Clin Microbiol Rev. 2005, 18, 657–686. [Google Scholar] [CrossRef] [PubMed]
- Ambler, R.P. The structure of beta-lactamases. Philos Trans R Soc Lond B Biol Sci. 1980, 289, 321–31. [Google Scholar] [PubMed]
- Bush, K.; Jacoby, G.; Medeiros, A. A functional classification scheme for b-lactamases and its correlation with molecular structure. Antimicrob Agents Chemother. 1995, 39, 1211–1233. [Google Scholar] [CrossRef]
- Bush, K.; Fisher, J.F. Epidemiological expansion, structural studies, and clinical challenges of new b-lactamases from gram-negative bacteria. Annu Rev Microbiol. 2011, 65, 455–478. [Google Scholar] [CrossRef]
- Woodford, N.; Turton, J.F.; Livermore, D.M. Multi resistant gram negative bacteria: the role of high-risk clones in the dissemination of antibiotic resistance. FEMS Microbiol. Rev. 2011, 35, 736–755. [Google Scholar] [CrossRef] [PubMed]
- Castanheira, M.; Simner, P.J.; Bradford, P.A. Extended-spectrum β-lactamases: an update on their characteristics, epidemiology and detection. JAC Antimicrob Resist. 2021, 3, dlab092. [Google Scholar] [CrossRef] [PubMed]
- Datta, N.; Kontomichalou, P. Penicillinase synthesis controlled by infectious R factors in Enterobacteriaceae. Nature. 1965, 208, 239–241. [Google Scholar] [CrossRef] [PubMed]
- Pitton, J.S. Mechanisms of bacterial resistance to antibiotics. Ergeb Physiol. 1972, 65, 15–93. [Google Scholar]
- Jacoby, G.A.; Bush, K. Amino acid sequences for TEM, SHV and OXA extended-spectrum and inhibitor resistant β-lactamases: Lahey Clinic; 1997. Available from: http://www.lahey.org/Studies/.
- NCBI National center for biotechnology information. Pathogen Detection Reference Gene Catalog. cited 2023 Feb 24. https://www.ncbi.nlm.nih.gov/pathogens/refgene/#.
- Dashti, A.A.; Jadaon, M.M.; Amyes, S.G. Retrospective study of an outbreak in a Kuwaiti hospital of multidrug-resistant Klebsiella pneumoniae possessing the new SHV-112 extended-spectrum beta-lactamase. J Chemother. 2010, 22, 335–8. [Google Scholar] [CrossRef] [PubMed]
- Dashti, A.A.; Vali, L.; Jadaon, M.M.; El-Shazly, S. The emergence of a multidrug-resistant Escherichia coli isolate harboring a novel SHV-122 enzyme is a serious threat for hospitalised patients. 16th HSC Poster Conference 2011, Faculty of Medicine, Kuwait University, Kuwait, May 3-5, 2011. 3 May.
- Dashti, A.A.; Jadaon, M.M.; Gomaa, H.H.; Noronha, B.; Udo, E.E. Transmission of a Klebsiella pneumoniae clone harbouring genes for CTX-M-15-like and SHV-112 enzymes in a neonatal intensive care unit of a Kuwaiti hospital. J Med Microbiol. 2010, 59 Pt 6, 687–692. [Google Scholar] [CrossRef] [PubMed]
- West, P.W. Extended-spectrum beta-lactamase-producing Klebsiella spp. Br J Biomed Sci. 2000, 57, 226–33. [Google Scholar] [PubMed]
- Alajmi, R.Z.; Alfouzan, W.A.; Mustafa, A.S. The Prevalence of Multidrug-Resistant Enterobacteriaceae among Neonates in Kuwait. Diagnostics 2023, 13, 1505. [Google Scholar] [CrossRef]
- Zowawi HM, Balkhy HH, Walsh TR, Paterson DL. β-Lactamase production in key gram-negative pathogen isolates from the Arabian Peninsula. Clin Microbiol Rev. 2013, 26, 361–80. [Google Scholar] [CrossRef] [PubMed]
- Dashti, A.A.; West, P.W. Extended-spectrum beta-lactamase-producing Escherichia coli isolated in the Al-Amiri Hospital in 2003 and compared with isolates from the Farwania hospital outbreak in 1994-96 in Kuwait. J Chemother. 2007, 19, 271–6. [Google Scholar] [CrossRef] [PubMed]
- Cattoir, V.; Poirel, L.; Rotimi, V.; Soussy, C.J.; Nordmann, P. Multiplex PCR for detection of plasmid-mediated quinolone resistance qnr genes in ESBL-producing enterobacterial isolates. J Antimicrob Chemother. 2007, 60, 394–7. [Google Scholar] [CrossRef] [PubMed]
- Rotimi, V.O.; Jamal, W.; Pal, T.; Sovenned, A.; Albert, M.J. Emergence of CTX-M-15 type extended-spectrum beta-lactamase-producing Salmonella spp. in Kuwait and the United Arab Emirates. J Med Microbiol. 2008, 57 Pt 7, 881–886. [Google Scholar] [CrossRef] [PubMed]
- Al Sweih, N.; Salama, M.F.; Jamal, W.; Al Hashem, G.; Rotimi, V.O. An outbreak of CTX-M-15-producing Klebsiella pneumoniae isolates in an intensive care unit of a teaching hospital in Kuwait. Indian J Med Microbiol. 2011, 29, 130–5. [Google Scholar] [CrossRef] [PubMed]
- Al Hashem, G.; Al Sweih, N.; Jamal, W.; Rotimi, V.O. Sequence analysis of bla(CTX-M) genes carried by clinically significant Escherichia coli isolates in Kuwait hospitals. Med Princ Pract. 2011, 20, 213–9. [Google Scholar] [CrossRef] [PubMed]
- Bonnin RA, Rotimi VO, Al Hubail M, Gasiorowski E, Al Sweih N, Nordmann P, et al. Wide dissemination of GES-type carbapenemases in Acinetobacter baumannii isolates in Kuwait. Antimicrob Agents Chemother. 2013, 57, 183–8. [Google Scholar] [CrossRef] [PubMed]
- Vali, L.; Dashti, A.A.; Jadaon, M.M.; El-Shazly, S. The emergence of plasmid mediated quinolone resistance qnrA2 in extended spectrum β-lactamase producing Klebsiella pneumoniae in the Middle East. Daru. 2015, 23, 34. [Google Scholar] [CrossRef]
- Al-Sweih, N.; Jamal, W.; Mokaddas, E.; Habashy, N.; Kurdi, A.; Mohamed, N. Evaluation of the in vitro activity of ceftaroline, ceftazidime/avibactam and comparator antimicrobial agents against clinical isolates from paediatric patients in Kuwait: ATLAS data 2012-19. JAC Antimicrob Resist. 2021, 3, dlab159. [Google Scholar] [CrossRef]
- Moghnia, O.H.; Al-Sweih, N.A. Whole Genome Sequence Analysis of Multidrug Resistant Escherichia coli and Klebsiella pneumoniae Strains in Kuwait. Microorganisms. 2022, 10, 507. [Google Scholar] [CrossRef] [PubMed]
- Findlay, J.; Sierra, R.; Raro, O.H.F.; Aires-de-Sousa, M.; Andrey, D.O.; Nordmann, P. Plasmid-mediated fosfomycin resistance in Escherichia coli isolates of worldwide origin. J Glob Antimicrob Resist. 2023, 35, 137–142. [Google Scholar] [CrossRef]
- Redha, M.A.; Al Sweih, N.; Albert, M.J. Multidrug-Resistant and Extensively Drug-Resistant Escherichia coli in Sewage in Kuwait: Their Implications. Microorganisms. 2023, 11, 2610. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Structure of -lactam molecule. (a) general structure showing the -lactam ring, (b) structure of 2-Azetidinone as the simplest -lactam, (c) structure of penicillin as an example of -lactams. Illustration is drawn by the authors of this paper.
Figure 1.
Structure of -lactam molecule. (a) general structure showing the -lactam ring, (b) structure of 2-Azetidinone as the simplest -lactam, (c) structure of penicillin as an example of -lactams. Illustration is drawn by the authors of this paper.
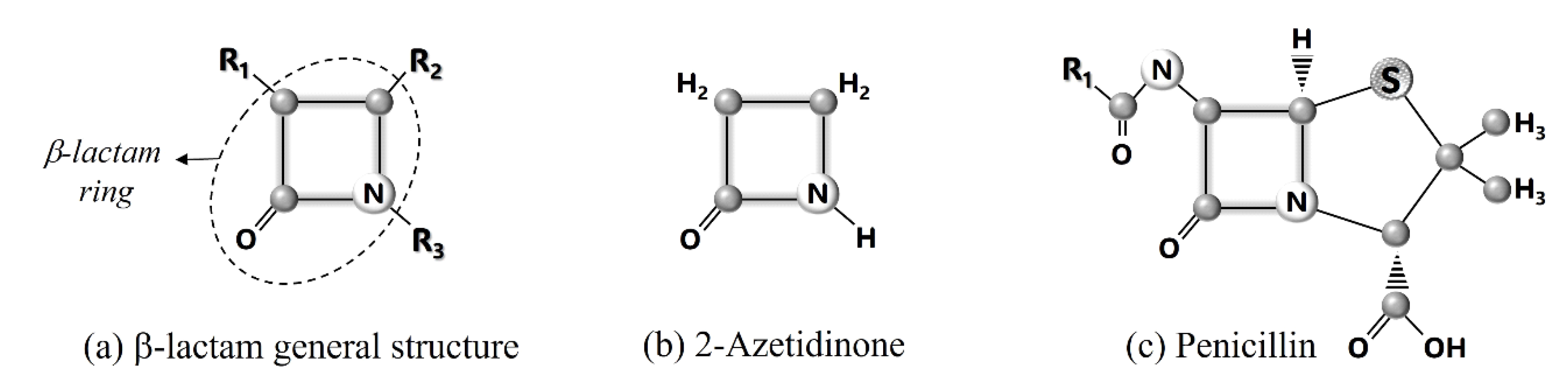
Figure 2.
Hydrolysis of -lactam ring as the method of penicillin-resistance exerted by the bacteria. Illustration is drawn by the authors of this paper.
Figure 2.
Hydrolysis of -lactam ring as the method of penicillin-resistance exerted by the bacteria. Illustration is drawn by the authors of this paper.
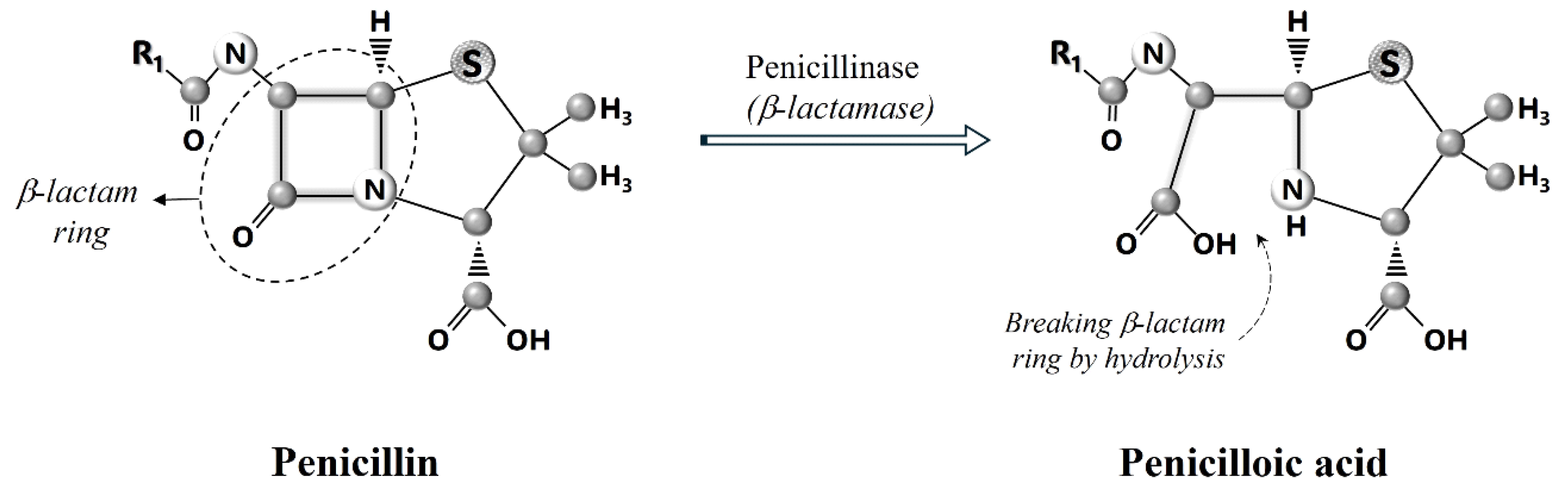
Figure 3.
The timeline of published findings of ESBLs gene & variants in Kuwait between the years 2007 and 2023, showing the years of publications and the genes variants reported by 13 studies, which were described in Table 1.
Figure 3.
The timeline of published findings of ESBLs gene & variants in Kuwait between the years 2007 and 2023, showing the years of publications and the genes variants reported by 13 studies, which were described in Table 1.


Table 1.
Summary of the findings of 13 research articles that included data on ESBLs genes & variants using molecular techniques in Kuwait, as found by searching the PubMed engine. * This was an international study from different countries. Only one case was included from Kuwait.
Table 1.
Summary of the findings of 13 research articles that included data on ESBLs genes & variants using molecular techniques in Kuwait, as found by searching the PubMed engine. * This was an international study from different countries. Only one case was included from Kuwait.
| Years of testing | Bacteria | Hospital in Kuwait | ESBL genes reported (percentage not shown) | Reference (publication year) |
| 2003 1994 to 1996 |
E. coli | Al-Amiri Al-Farwania |
TEM & SHV (general) | 37 (2007) |
| 2002 to 2004 | E. cloacae, C. freundii | Mubarak Al-Kabir |
SHV-12, VEB-1b | 38 (2007) |
| 2003 to 2006 | Salmonella spp. | Mubarak Al-Kabir, Infectious Diseases H. |
TEM, CTX-M-15 | 39 (2008) |
| 2005 to 2006 | K. pneumoniae | Al-Jahra | TEM-1, SHV-112, CTX-M-15-like |
33 (2010) |
| 2010 | K. pneumoniae | Al-Amiri | SHV-112 | 31 (2010) |
| 2008 | K. pneumoniae | Mubarak Al-Kabeer | TEM-1, CTX-M-15 | 40 (2011) |
| 2008 | E. coli | Mubarak Al-Kabeer, Al-Amiri, Al-Sabah, Al-Adan, Al-Jahra, Ibn Sina, Al-Farwaniya, Maternity |
TEM, SHV (general), CTX-M (-15, -14, -14b, TOHO-1) |
41 (2011) |
| 2007 to 2008 | A. baumannii | Mubarak Al-Kabeer, Al-Sabah, Al-Adan, Al-Jahra, Al-Babtain, Al- Razi |
GES-11, GES-14, OXA-23 OXA-51-like (-74, -66, -71, -98) |
42 (2013) |
| Non-specified | K. pneumoniae | Al-Amiri, Al-Adan, Al-Ahmadi (Kuwait Oil Company H.) |
TEM-1, SHV, CTX-M (-2, -15) |
43 (2015) |
| 2012 to 2019 | E. coli, K. pneumoniae | 3 non-specified hospitals | CTX-M-15 | 44 (2021) |
| 2020 to 2021 | E. coli, K. pneumoniae | Mubarak Al-Kabeer, Ibn Sina, Al-Babtain |
TEM-1, SHV-11/-12, CTX-M-15, OXA-1/-48, KPC-2/-29, CMY-4/-6, OKP-B, ACT, EC. |
45 (2022) |
| 2020 | Enterobacteriaceae spp. (mostly E. coli, Klebsiella spp., Enterobacter spp.) | Al-Farwaniya | TEM, SHV & CTX-M (general) | 35 (2023) |
| 2017 to 2022 | E. coli | Non-specified | CMY-2 | 46 (2023) * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



