Submitted:
08 February 2025
Posted:
10 February 2025
You are already at the latest version
Abstract
Muscle regulation requires both the newly discovered thick filament as well as classical thin filament regulation mechanisms. Thick filament-based regulation mechanisms are generally conceived as processes modulating the number of myosin heads capable of force generation. It has been generally assumed that the biochemical and structural states of myosin, usually correlated with each other, is the basis of this recruitment, and work jointly to regulate contractile force. This notion has been challenged recently by studies showed that the biochemical and structural states of myosin can be decoupled. Here we studied the steady state and dynamic mechanical changes in skinned porcine myocardium with and without OM or piperine to help decipher the basis of thick filament regulation and how either affects contractile force. Our study supports the notion that thick filament activation is primarily a process of myosin recruitment and that it is not necessarily coupled with chemo-cycling of crossbridges. Perturbations that result in myosin head recruitment from the thick filaments do not necessarily increase contractile force and vice versa. These new insights into thick filament activation mechanisms will be of great relevance to better designing sarcomere therapies aimed at reversing them for treatment of myopathies.
Keywords:
1. Introduction
2. Materials and Methods
2.1. Single Cardiomyocyte Mechanics
2.2. Crossbridge Kinetics
3. Results
3.1. Tension vs. Calcium Concentration Relationship before and after OM and Piperine Treatment
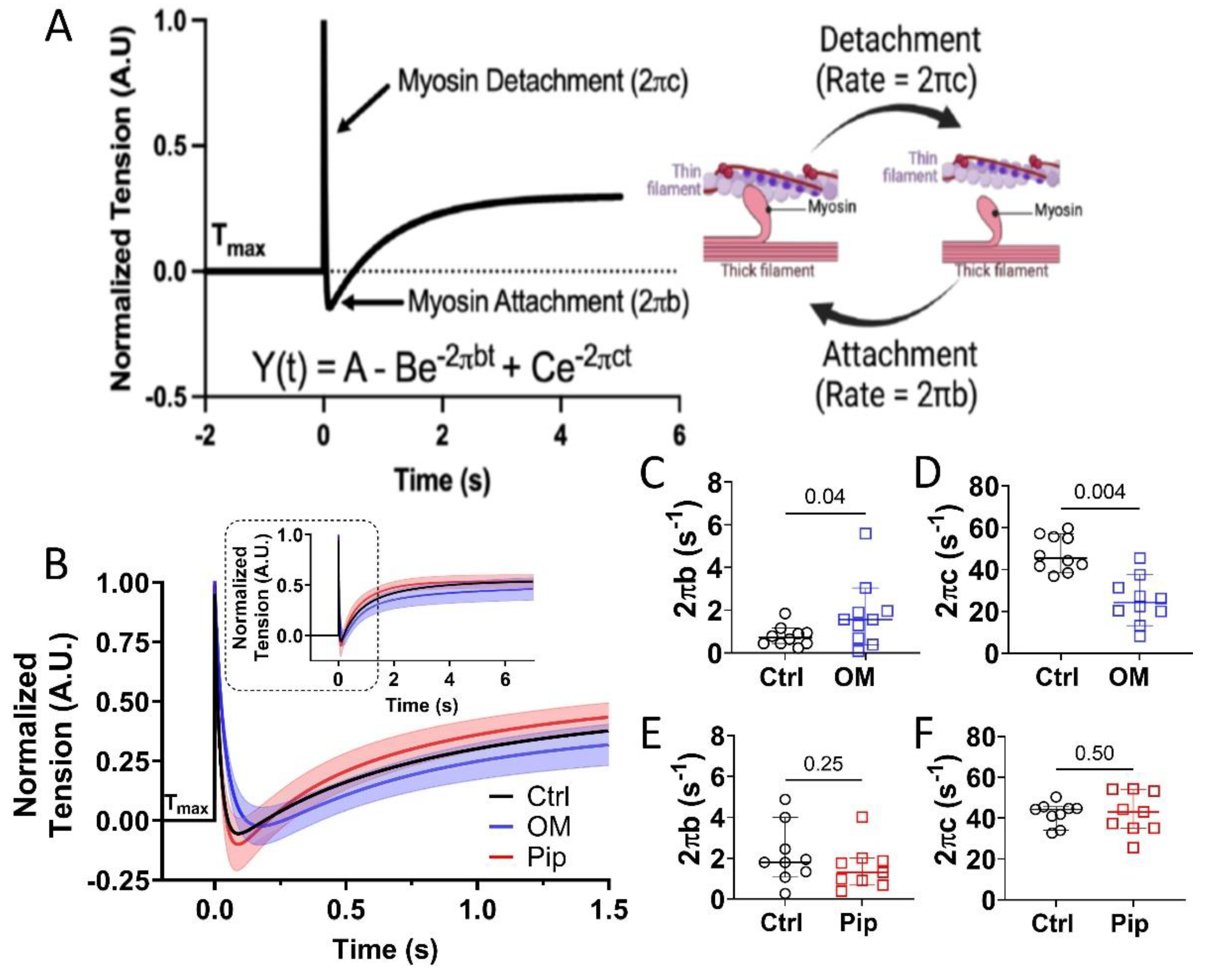
3.2. Effects of OM and Piperine on Crossbridge Attachment and Detachment from Permeabilized Porcine CMs During Maximal Activation
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Irving, M. Regulation of Contraction by the Thick Filaments in Skeletal Muscle. Biophys. J. 2017, 113, 2579–2594. [Google Scholar] [CrossRef] [PubMed]
- Spudich, J.A. Three perspectives on the molecular basis of hypercontractility caused by hypertrophic cardiomyopathy mutations. Pfl?gers Arch. Eur. J. Physiol. 2019, 471, 701–717. [Google Scholar] [CrossRef]
- Kawana, M.; Spudich, J.A.; Ruppel, K.M. Hypertrophic cardiomyopathy: Mutations to mechanisms to therapies. Front. Physiol. 2022, 13, 975076. [Google Scholar] [CrossRef]
- Sarkar, S.S.; Trivedi, D.V.; Morck, M.M.; Adhikari, A.S.; Pasha, S.N.; Ruppel, K.M.; Spudich, J.A. The hypertrophic cardiomyopathy mutations R403Q and R663H increase the number of myosin heads available to interact with actin. Sci. Adv. 2020, 6, eaax0069. [Google Scholar] [CrossRef] [PubMed]
- Powers, J.D.; Yuan, C.-C.; McCabe, K.J.; Murray, J.D.; Childers, M.C.; Flint, G.V.; Moussavi-Harami, F.; Mohran, S.; Castillo, R.; Zuzek, C.; et al. Cardiac myosin activation with 2-deoxy-ATP via increased electrostatic interactions with actin. Proc. Natl. Acad. Sci. USA 2019, 116, 11502–11507. [Google Scholar] [CrossRef]
- Ma, W.; McMillen, T.S.; Childers, M.C.; Gong, H.; Regnier, M.; Irving, T. Structural OFF/ON transitions of myosin in relaxed porcine myocardium predict calcium-activated force. Proc. Natl. Acad. Sci. USA 2023, 120. [Google Scholar] [CrossRef] [PubMed]
- Walklate, J.; Kao, K.; Regnier, M.; Geeves, M.A. Exploring the super-relaxed state of myosin in myofibrils from fast-twitch, slow-twitch, and cardiac muscle. J. Biol. Chem. 2022, 298, 101640. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Henze, M.; Anderson, R.L.; Gong, H.; Wong, F.L.; del Rio, C.L.; Irving, T. The Super-Relaxed State and Length Dependent Activation in Porcine Myocardium. Circ. Res. 2021, 129, 617–630. [Google Scholar] [CrossRef] [PubMed]
- Jani, V.P.; Song, T.; Gao, C.; Gong, H.; Sadayappan, S.; A Kass, D.; Irving, T.C.; Ma, W. The structural OFF and ON states of myosin can be decoupled from the biochemical super- and disordered-relaxed states. PNAS Nexus 2024, 3, 39. [Google Scholar] [CrossRef] [PubMed]
- Muretta, J.M.; Rohde, J.A.; Johnsrud, D.O.; Cornea, S.; Thomas, D.D. Direct real-time detection of the structural and biochemical events in the myosin power stroke. Proc. Natl. Acad. Sci. USA 2015, 112, 14272–14277. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; White, H.D.; Belknap, B.; Winkelmann, D.A.; Forgacs, E. Omecamtiv Mecarbil modulates the kinetic and motile properties of porcine beta-cardiac myosin. Biochemistry 2015, 54, 1963–1975. [Google Scholar] [CrossRef]
- Nagy, L.; Kovács, Á.; Bódi, B.; Pásztor, E.T.; Fülöp, G.Á.; Tóth, A.; Édes, I.; Papp, Z. The novel cardiac myosin activator omecamtiv mecarbil increases the calcium sensitivity of force production in isolated cardiomyocytes and skeletal muscle fibres of the rat. Br. J. Pharmacol. 2015, 172, 4506–4518. [Google Scholar] [CrossRef]
- Nakanishi, T.; Oyama, K.; Tanaka, H.; Kobirumaki-Shimozawa, F.; Ishii, S.; Terui, T.; Ishiwata, S.; Fukuda, N. Effects of omecamtiv mecarbil on the contractile properties of skinned porcine left atrial and ventricular muscles. Front. Physiol. 2022, 13, 947206. [Google Scholar] [CrossRef] [PubMed]
- Jani, V.; Qian, W.; Yuan, S.; Irving, T.; Ma, W. EMD-57033 Augments the Contractility in Porcine Myocardium by Promoting the Activation of Myosin in Thick Filaments. Int. J. Mol. Sci. 2022, 23, 14517. [Google Scholar] [CrossRef] [PubMed]
- Palmer, B.M.; Suzuki, T.; Wang, Y.; Barnes, W.D.; Miller, M.S.; Maughan, D.W. Two-State Model of Acto-Myosin Attachment-Detachment Predicts C-Process of Sinusoidal Analysis. Biophys. J. 2007, 93, 760–769. [Google Scholar] [CrossRef]
- Lindqvist, J.; Lee, E.J.; Karimi, E.; Kolb, J.; Granzier, H. Omecamtiv mecarbil lowers the contractile deficit in a mouse model of nebulin-based nemaline myopathy. PloS ONE 2019, 14, e0224467. [Google Scholar] [CrossRef] [PubMed]
- Swenson, A.M.; Tang, W.; Blair, C.A.; Fetrow, C.M.; Unrath, W.C.; Previs, M.J.; Campbell, K.S.; Yengo, C.M. Omecamtiv Mecarbil Enhances the Duty Ratio of Human beta-Cardiac Myosin Resulting in Increased Calcium Sensitivity and Slowed Force Development in Cardiac Muscle. J. Biol. Chem. 2017, 292, 3768–3778. [Google Scholar] [CrossRef]
- Mamidi, R.; Li, J.; Gresham, K.S.; Verma, S.; Doh, C.Y.; Li, A.; Lal, S.; dos Remedios, C.G.; Stelzer, J.E. Dose-Dependent Effects of the Myosin Activator Omecamtiv Mecarbil on Cross-Bridge Behavior and Force Generation in Failing Human Myocardium. Circ. Hear. Fail. 2017, 10. [Google Scholar] [CrossRef] [PubMed]
- Kooiker, K.B.; Mohran, S.; Turner, K.L.; Ma, W.; Martinson, A.; Flint, G.; Qi, L.; Gao, C.; Zheng, Y.; McMillen, T.S.; et al. Danicamtiv Increases Myosin Recruitment and Alters Cross-Bridge Cycling in Cardiac Muscle. Circ. Res. 2023, 133, 430–443. [Google Scholar] [CrossRef]
- Ma, W.; Nag, S.; Gong, H.; Qi, L.; Irving, T.C. Cardiac myosin filaments are directly regulated by calcium. J. Gen. Physiol. 2022, 154. [Google Scholar] [CrossRef]
- Anderson, R.L.; Trivedi, D.V.; Sarkar, S.S.; Henze, M.; Ma, W.; Gong, H.; Rogers, C.S.; Gorham, J.M.; Wong, F.L.; Morck, M.M.; et al. Deciphering the super relaxed state of human beta-cardiac myosin and the mode of action of mavacamten from myosin molecules to muscle fibers. Proc. Natl. Acad. Sci. USA 2018, 115, E8143–E8152. [Google Scholar] [CrossRef] [PubMed]
- Nag, S.; Gollapudi, S.K.; del Rio, C.L.; Spudich, J.A.; McDowell, R. Mavacamten, a precision medicine for hypertrophic cardiomyopathy: From a motor protein to patients. Sci. Adv. 2023, 9, eabo7622. [Google Scholar] [CrossRef]
- Ho, C.Y.; Mealiffe, M.E.; Bach, R.G.; Bhattacharya, M.; Choudhury, L.; Edelberg, J.M.; Hegde, S.M.; Jacoby, D.; Lakdawala, N.K.; Lester, S.J.; et al. Evaluation of Mavacamten in Symptomatic Patients With Nonobstructive Hypertrophic Cardiomyopathy. Circ. 2020, 75, 2649–2660. [Google Scholar] [CrossRef] [PubMed]
- Jani, V.; Aslam, M.I.; Fenwick, A.J.; Ma, W.; Gong, H.; Milburn, G.; Nissen, D.; Salazar, I.M.C.; Hanselman, O.; Mukherjee, M.; et al. Right Ventricular Sarcomere Contractile Depression and the Role of Thick Filament Activation in Human Heart Failure With Pulmonary Hypertension. Circulation 2023, 147, 1919–1932. [Google Scholar] [CrossRef]
- Ma, W.; Gong, H.; Irving, T. Myosin Head Configurations in Resting and Contracting Murine Skeletal Muscle. Int. J. Mol. Sci. 2018, 19, 2643. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; White, H.D.; Offer, G.W.; Yu, L.C. Stabilization of Helical Order in the Thick Filaments by Blebbistatin: Further Evidence of Coexisting Multiple Conformations of Myosin. Biophys. J. 2009, 96, 3673–3681. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.-Q.; Padrón, R.; Craig, R. Blebbistatin Stabilizes the Helical Order of Myosin Filaments by Promoting the Switch 2 Closed State. Biophys. J. 2008, 95, 3322–3329. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Offer, G.; Gu, J.; White, H.D.; Yu, L.C. Temperature and ligand dependence of conformation and helical order in myosin filaments. Biochemistry 2003, 42, 390–401. [Google Scholar] [CrossRef] [PubMed]
- Caremani, M.; Brunello, E.; Linari, M.; Fusi, L.; Irving, T.C.; Gore, D.; Piazzesi, G.; Irving, M.; Lombardi, V.; Reconditi, M. Low temperature traps myosin motors of mammalian muscle in a refractory state that prevents activation. J. Gen. Physiol. 2019, 151, 1272–1286. [Google Scholar] [CrossRef]
- Ma, W.; Duno-Miranda, S.; Irving, T.; Craig, R.; Padrón, R. Relaxed tarantula skeletal muscle has two ATP energy-saving mechanisms. J. Gen. Physiol. 2021, 153. [Google Scholar] [CrossRef]
- Cheung, A.; Dantzig, J.A.; Hollingworth, S.; Baylor, S.M.; Goldman, Y.; Mitchison, T.J.; Straight, A.F. A small-molecule inhibitor of skeletal muscle myosin II. Nat. Cell Biol. 2001, 4, 83–88. [Google Scholar] [CrossRef] [PubMed]
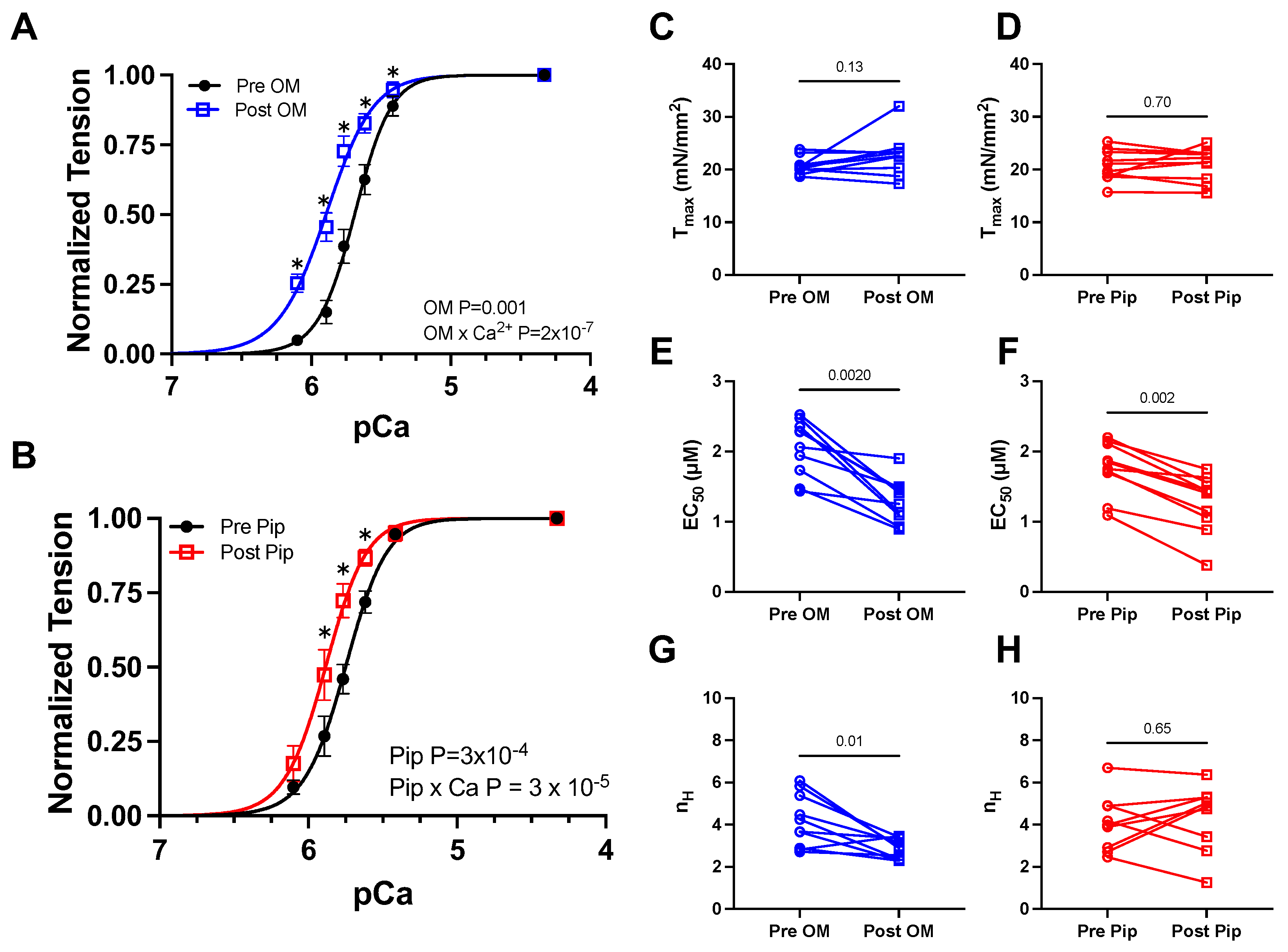

| Ctrl | OM | p values | Ctrl | PIP | p values | |
|---|---|---|---|---|---|---|
| pCa 8 | 0 | 0 | ns | 0 | 0 | ns |
| pCa 6.10 | 1.04 ± 0.26 | 5.73 ± 0.78 | **** | 1.92 ± 0.45 | 3.68 ± 1.28 | ns |
| pCa 5.89 | 3.18 ± 1.93 | 10.32 ± 1.26 | **** | 5.34 ± 1.23 | 10.35 ± 1.95 | **** |
| pCa 5.77 | 8.15 ±1.32 | 16.49 ± 1.44 | **** | 9.50 ± 1.03 | 15.41 ± 1.17 | **** |
| pCa 5.62 | 13.17 ± 1.21 | 18.58 ± 0.89 | **** | 15.15 ± 1.28 | 18.57 ± 0.76 | ** |
| pCa 5.42 | 18.58 ± 0.73 | 21.52 ± 1.39 | * | 19.76 ± 0.98 | 20.40 ± 0.98 | ns |
| pCa 4.33 | 20.98 ± 0.54 | 22.71 ± 1.20 | Ns | 20.85 ± 0.93 | 21.62 ± 1.16 | ns |
| Tmax | 20.70 ± 0.52 | 22.70 ± 1.25 | Ns | 20.72 ± 0.92 | 20.98 ± 0.97 | ns |
| EC50 (μM) | 2.06 ± 0.13 | 1.34 ± 0.31 | ** | 1.76 ± 0.12 | 1.27 ± 0.13 | ** |
| nh | 4.18 ± 0.40 | 2.88 ± 0.15 | * | 4.07 ± 0.44 | 4.34 ± 0.52 | ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



