Submitted:
04 February 2025
Posted:
06 February 2025
You are already at the latest version
Abstract
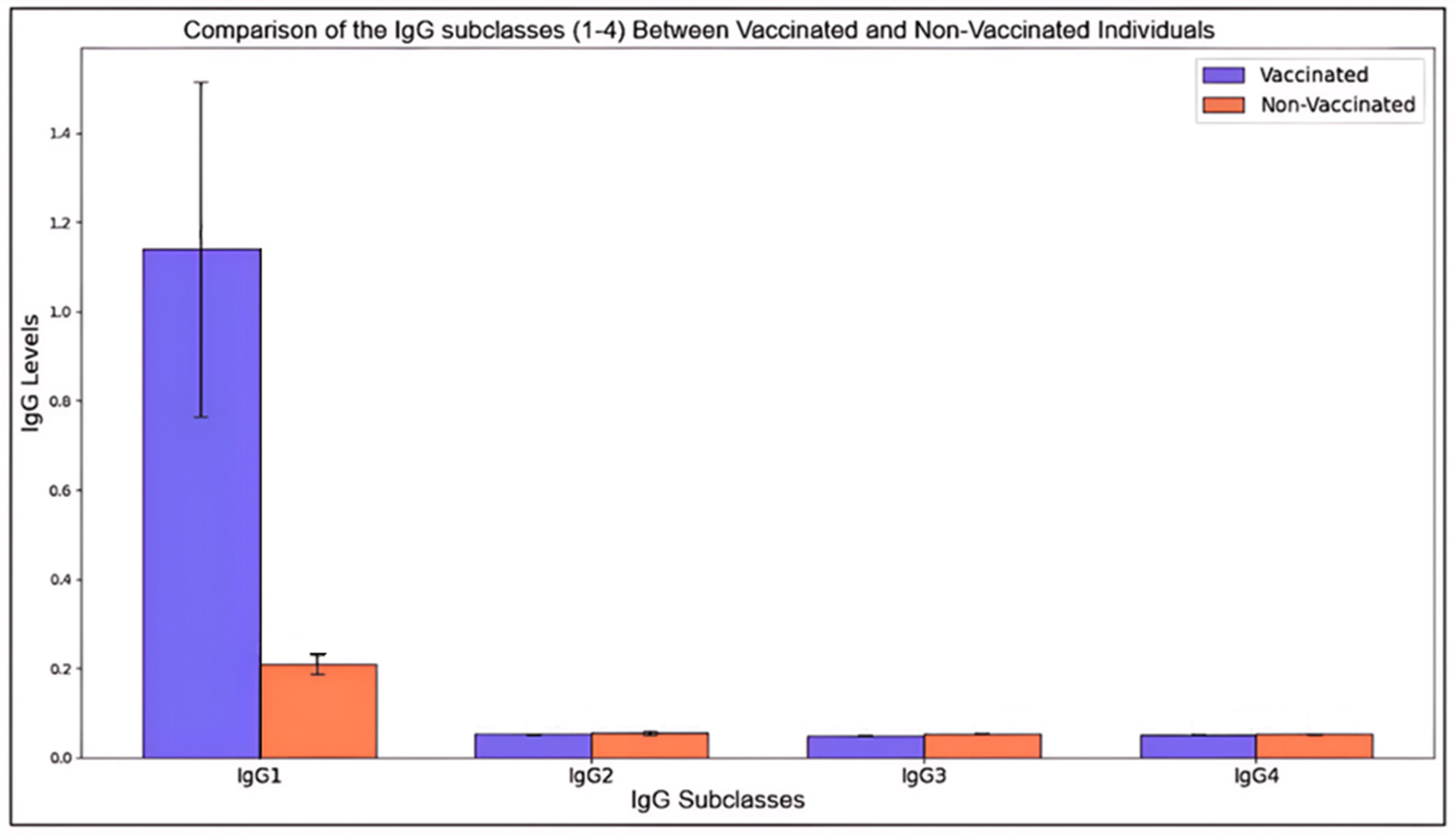
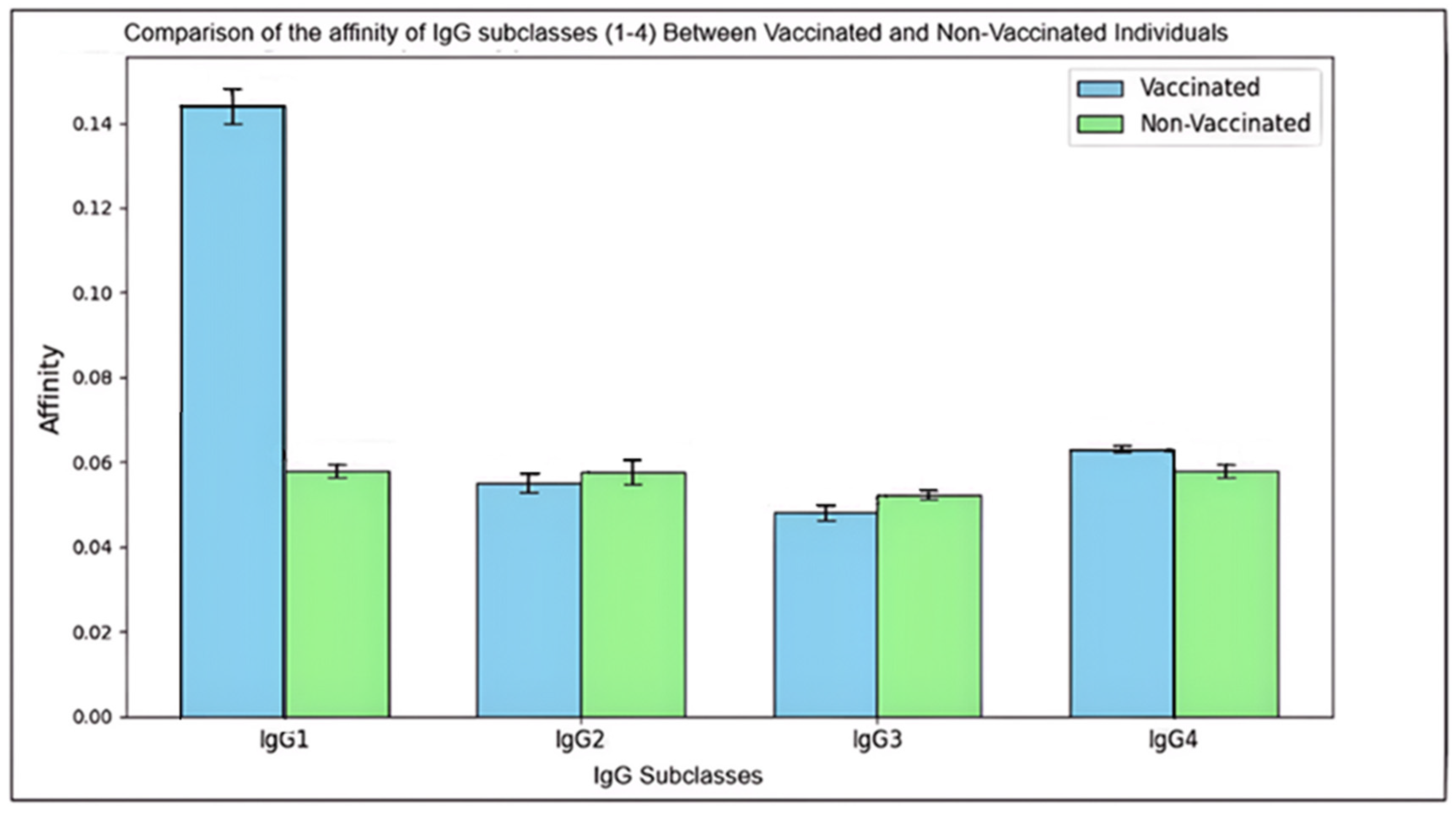
The COVID-19 pandemic has underscored the critical need to understand immune responses arising from both natural infection and vaccination. This study examines the affinity and subclass distribution of anti-nucleocapsid (anti-N) antibodies in 60 participants, comprising 30 vaccinated and 30 non-vaccinated individuals. Using enzyme-linked immunosorbent assays (ELISA), we quantified total IgG (tIgG) and its subclasses (IgG1, IgG2, IgG3, and IgG4) in serum samples. Antibody affinity was assessed using ammonium thiocyanate to evaluate the strength of antigen-antibody interactions. We observed significantly elevated levels of tIgG and its subclasses in the vaccinated participants compared to the non-vaccinated controls, indicating a more robust immune response post-vaccination. In vaccinated individuals, the mean levels of tIgG, IgG1, IgG2, IgG3, and IgG4 were 2.59, 1.02, 0.055, 0.061, and 0.095 (OD), respectively, while non-vaccinated individuals showed lower mean levels at 0.512, 0.11, 0.052, 0.051, and 0.048 (OD) for tIgG, IgG1, IgG2, IgG3, and IgG4, respectively. This notable difference suggests that vaccination promotes not only higher production of these antibody subclasses but also, as shown by the corresponding affinity tests, an increased binding strength of anti-N antibodies. These results indicate that vaccination induces a more robust and higher-quality immune response, contributing to improved immune protection against SARS-CoV-2. The study highlights the critical role of anti-N antibodies in vaccine efficacy and long-term immunity. Our findings offer valuable insights into immune dynamics, informing future vaccine development and strategies for pandemic control, potentially leading to better management of emerging variants and more effective public health interventions.
Keywords:
1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. Participants and Samples

| Vaccinated | Non-vaccinated (control) | |||||
|---|---|---|---|---|---|---|
| S.# | Sex | Age | Vaccine doses | S.# | Sex | Age |
| 1 | Female | 63 | Pfizer (3 doses) | 1 | Female | 30 |
| 2 | Male | 39 | Pfizer (3 doses) | 2 | Male | 56 |
| 3 | Female | 28 | AstraZeneca (2 doses) | 3 | Female | 37 |
| 4 | Female | 26 | AstraZeneca (2 doses) + Pfizer (1 dose) | 4 | Female | 29 |
| 5 | Female | 23 | Pfizer (3 doses) | 5 | Female | 67 |
| 6 | Male | 20 | AstraZeneca (2 doses) + Pfizer (1 dose) | 6 | Female | 64 |
| 7 | Female | 27 | AstraZeneca (2 doses) + Pfizer (1 dose) | 7 | Male | 61 |
| 8 | Female | 33 | Pfizer (3 doses) | 8 | Female | 45 |
| 9 | Female | 27 | Pfizer (3 doses) | 9 | Female | 11 |
| 10 | Female | 24 | Pfizer (3 doses) | 10 | Female | 61 |
| 11 | Male | 36 | Pfizer (3 doses) | 11 | Male | 71 |
| 12 | Male | 38 | Pfizer (1 dose) + Moderna (1 dose) | 12 | Female | 51 |
| 13 | Male | 30 | AstraZeneca (2 doses) | 13 | Female | 56 |
| 14 | Female | 29 | Pfizer (3 doses) | 14 | Male | 30 |
| 15 | Female | 27 | Pfizer (3 doses) | 15 | Male | 46 |
| 16 | Female | 30 | Pfizer (3 doses) | 16 | Male | 59 |
| 17 | Male | 37 | Pfizer (3 doses) | 17 | Female | 54 |
| 18 | Male | 35 | Pfizer (3 doses) | 18 | Male | 56 |
| 19 | Male | 37 | Pfizer (2 doses) | 19 | Female | 55 |
| 20 | Male | 36 | Pfizer (1 dose) + Moderna (1 dose) | 20 | Male | 71 |
| 21 | Male | 37 | AstraZeneca (3 doses) | 21 | Female | 49 |
| 22 | Male | 40 | AstraZeneca (3 doses) | 22 | Male | 63 |
| 23 | Male | 28 | Pfizer (2 doses) | 23 | Female | 66 |
| 24 | Male | 37 | Pfizer (3 doses) | 24 | Female | 50 |
| 25 | Male | 35 | AstraZeneca (2 dose) | 25 | Male | 72 |
| 26 | Female | 18 | AstraZeneca (2 doses)+ Pfizer (1 dose) | 26 | Male | 61 |
| 27 | Female | 51 | Johnson & Johnson (1 dose) | 27 | Male | 64 |
| 28 | Female | 49 | AstraZeneca (2 doses)+ Pfizer (1 dose) | 28 | Female | 56 |
| 29 | Male | 30 | Pfizer (3 doses) | 29 | Male | 73 |
| 30 | Male | 32 | Pfizer (3 doses) | 30 | Female | 36 |
2.1.2. Anti–Human Antibodies
2.2. Methods
2.3. Statistical Analysis
3. Results
3.1. Total IgG in Vaccinated and Non-Vaccinated (X±SD)
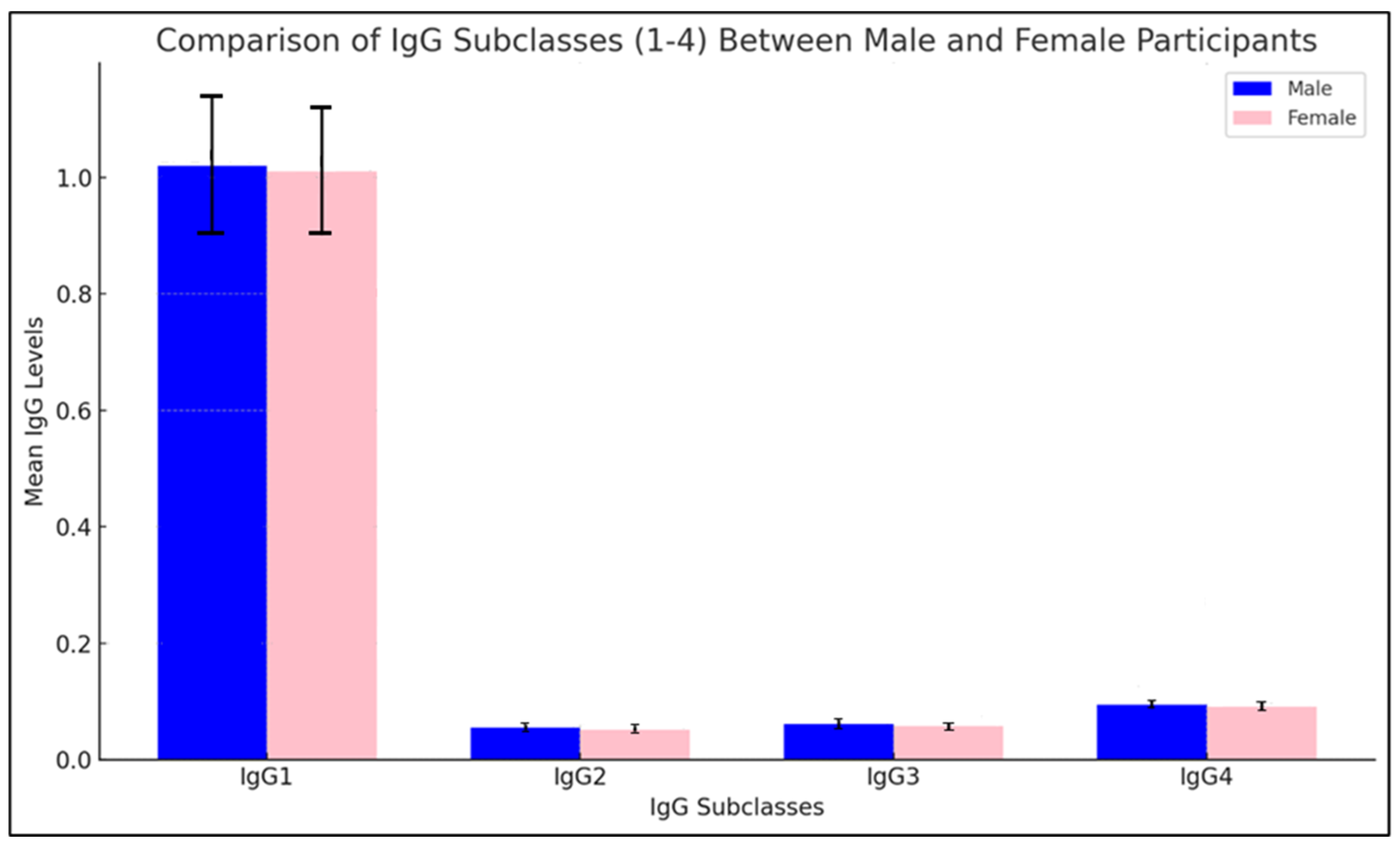
3.2. Anti-N IgG1 Among Vaccinated and Non-Vaccinated (X±SD)
3.3. Anti-N IgG2 Among Vaccinated and Non-Vaccinated (X±SD)
3.4. Anti-N IgG3 Among Vaccinated and Non-Vaccinated (X±SD)
3.5. Anti-N IgG4 Among Vaccinated and Non-Vaccinated (X±SD)
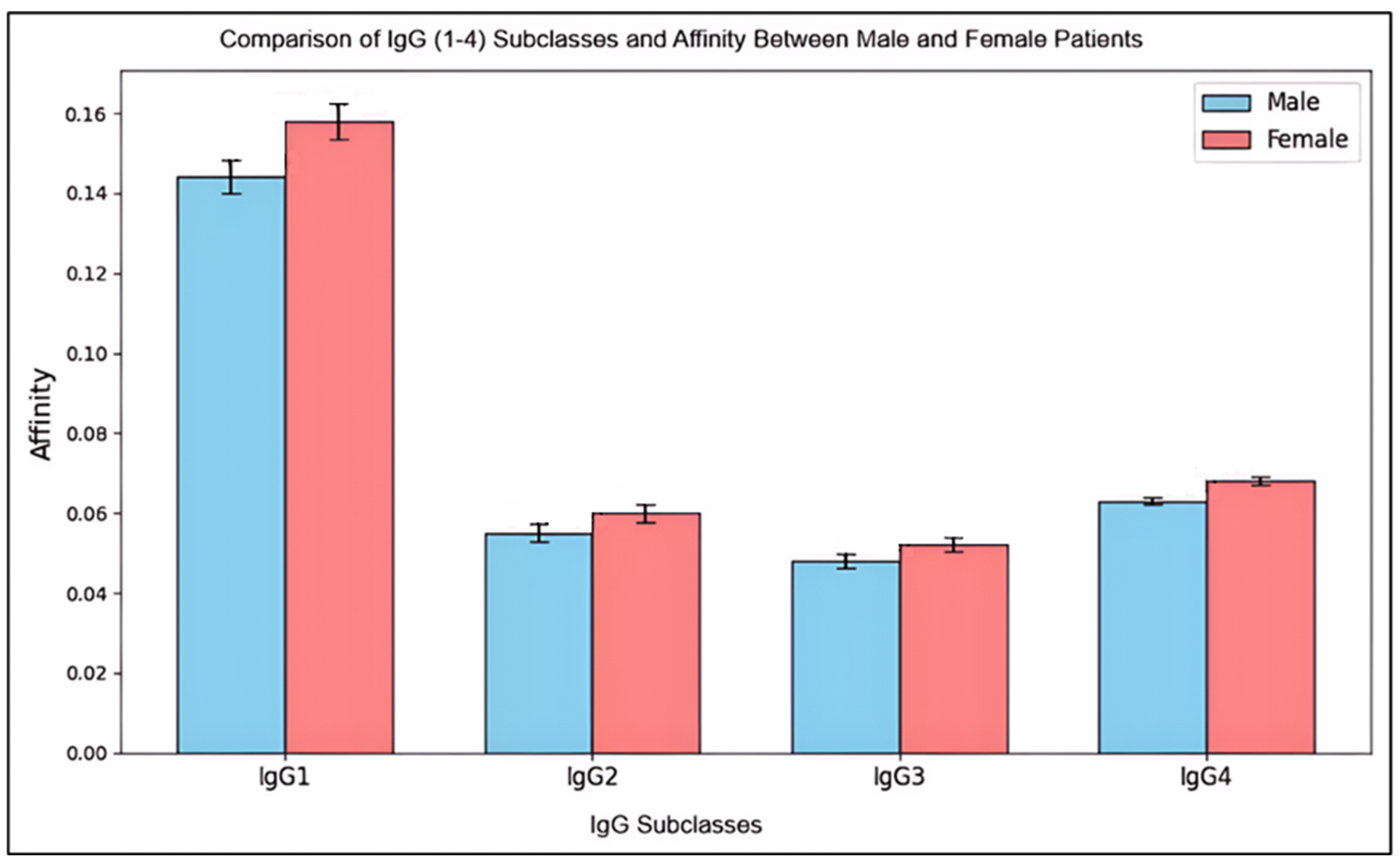
3.6. Affinity of IgG1 in Vaccinated and Non-Vaccinated (X±SD)
3.7. Affinity of IgG2 in Vaccinated and Non-Vaccinated (X±SD)
3.8. Affinity of IgG3 in Vaccinated and Non-Vaccinated (X±SD)
3.9. Affinity of IgG4 in Vaccinated and Non-Vaccinated (X±SD)
4. Discussion
5. Conclusions and Future Work
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Xu X, Chen P, Wang J, Feng J, Zhou H, Li X, et al. Evolution of the novel coronavirus from the ongoing Wuhan outbreak and modeling of its spike protein for risk of human transmission. Sci China Life Sci. 2020 Mar;63(3):457–60.
- Cui J, Li F, Shi ZL. Origin and evolution of pathogenic coronaviruses. Nat Rev Microbiol. 2019 Mar;17(3):181–92.
- Holshue ML, DeBolt C, Lindquist S, Lofy KH, Wiesman J, Bruce H, et al. First Case of 2019 Novel Coronavirus in the United States. N Engl J Med. 2020 Mar 5;382(10):929–36.
- Lu R, Zhao X, Li J, Niu P, Yang B, Wu H, et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: implications for virus origins and receptor binding. The Lancet. 2020 Feb;395(10224):565–74.
- Narayanan K, Chen CJ, Maeda J, Makino S. Nucleocapsid-independent specific viral RNA packaging via viral envelope protein and viral RNA signal. J Virol. 2003 Mar;77(5):2922–7.
- Hurst KR, Koetzner CA, Masters PS. Identification of in vivo-interacting domains of the murine coronavirus nucleocapsid protein. J Virol. 2009 Jul;83(14):7221–34.
- Huang Q, Yu L, Petros AM, Gunasekera A, Liu Z, Xu N, et al. Structure of the N-terminal RNA-binding domain of the SARS CoV nucleocapsid protein. Biochemistry. 2004 May 25;43(20):6059–63.
- Tan YJ, Goh PY, Fielding BC, Shen S, Chou CF, Fu JL, et al. Profiles of antibody responses against severe acute respiratory syndrome coronavirus recombinant proteins and their potential use as diagnostic markers. Clin Diagn Lab Immunol. 2004 Mar;11(2):362–71.
- Shi Y, Yi Y, Li P, Kuang T, Li L, Dong M, et al. Diagnosis of severe acute respiratory syndrome (SARS) by detection of SARS coronavirus nucleocapsid antibodies in an antigen-capturing enzyme-linked immunosorbent assay. J Clin Microbiol. 2003 Dec;41(12):5781–2.
- Leung DTM, Tam FCH, Ma CH, Chan PKS, Cheung JLK, Niu H, et al. Antibody Response of Patients with Severe Acute Respiratory Syndrome (SARS) Targets the Viral Nucleocapsid. J Infect Dis. 2004 Jul 15;190(2):379–86.
- Zou L, Ruan F, Huang M, Liang L, Huang H, Hong Z, et al. SARS-CoV-2 Viral Load in Upper Respiratory Specimens of Infected Patients. N Engl J Med. 2020 Mar 19;382(12):1177–9.
- To KKW, Tsang OTY, Leung WS, Tam AR, Wu TC, Lung DC, et al. Temporal profiles of viral load in posterior oropharyngeal saliva samples and serum antibody responses during infection by SARS-CoV-2: an observational cohort study. Lancet Infect Dis. 2020 May;20(5):565–74.
- Chen W, Lan Y, Yuan X, Deng X, Li Y, Cai X, et al. Detectable 2019-nCoV viral RNA in blood is a strong indicator for the further clinical severity. Emerg Microbes Infect. 2020;9(1):469–73.
- Ong DSY, de Man SJ, Lindeboom FA, Koeleman JGM. Comparison of diagnostic accuracies of rapid serological tests and ELISA to molecular diagnostics in patients with suspected coronavirus disease 2019 presenting to the hospital. Clin Microbiol Infect. 2020 Aug;26(8):1094.e7-1094.e10.
- Long QX, Liu BZ, Deng HJ, Wu GC, Deng K, Chen YK, et al. Antibody responses to SARS-CoV-2 in patients with COVID-19. Nat Med. 2020 Jun;26(6):845–8.
- L’Huillier AG, Meyer B, Andrey DO, Arm-Vernez I, Baggio S, Didierlaurent A, et al. Antibody persistence in the first 6 months following SARS-CoV-2 infection among hospital workers: a prospective longitudinal study. Clin Microbiol Infect. 2021 Jan 20;27(5):784.e1-8.
- Lumley SF, O’Donnell D, Stoesser NE, Matthews PC, Howarth A, Hatch SB, et al. Antibody Status and Incidence of SARS-CoV-2 Infection in Health Care Workers. N Engl J Med. 2021 Feb 11;384(6):533–40.
- Jeffery-Smith A, Iyanger N, Williams S V, Chow JY, Aiano F, Hoschler K, et al. Antibodies to SARS-CoV-2 protect against re-infection during outbreaks in care homes, September and October 2020. Euro Surveill. 2021 Feb;26(5).
- Addetia A, Crawford KHD, Dingens A, Zhu H, Roychoudhury P, Huang ML, et al. Neutralizing Antibodies Correlate with Protection from SARS-CoV-2 in Humans during a Fishery Vessel Outbreak with a High Attack Rate. J Clin Microbiol. 2020 Oct 21;58(11).
- Prendecki M, Clarke C, Brown J, Cox A, Gleeson S, Guckian M, et al. Effect of previous SARS-CoV-2 infection on humoral and T-cell responses to single-dose BNT162b2 vaccine. Lancet. 2021 Mar 27;397(10280):1178–81.
- Abu Jabal K, Ben-Amram H, Beiruti K, Batheesh Y, Sussan C, Zarka S, et al. Impact of age, ethnicity, sex and prior infection status on immunogenicity following a single dose of the BNT162b2 mRNA COVID-19 vaccine: real-world evidence from healthcare workers, Israel, December 2020 to January 2021. Euro Surveill. 2021 Feb;26(6).
- Krammer F, Srivastava K, Alshammary H, Amoako AA, Awawda MH, Beach KF, et al. Antibody Responses in Seropositive Persons after a Single Dose of SARS-CoV-2 mRNA Vaccine. N Engl J Med. 2021 Apr 8;384(14):1372–4.
- Irrgang P, Gerling J, Kocher K, Lapuente D, Steininger P, Habenicht K, et al. Class switch toward noninflammatory, spike-specific IgG4 antibodies after repeated SARS-CoV-2 mRNA vaccination. Sci Immunol. 2023 Jan 27;8(79):eade2798.
- Uversky VN, Redwan EM, Makis W, Rubio-Casillas A. IgG4 Antibodies Induced by Repeated Vaccination May Generate Immune Tolerance to the SARS-CoV-2 Spike Protein. Vaccines (Basel). 2023 May 17;11(5).
- Huijbers MG, Plomp JJ, van der Maarel SM, Verschuuren JJ. IgG4-mediated autoimmune diseases: a niche of antibody-mediated disorders. Ann N Y Acad Sci. 2018 Feb;1413(1):92–103.
- Rubio-Casillas A, Redwan EM, Uversky VN. Does SARS-CoV-2 Induce IgG4 Synthesis to Evade the Immune System? Biomolecules. 2023 Sep 1;13(9).
- Asa, CS. Contraception in Dogs and Cats. Vet Clin North Am Small Anim Pract. 2018 Jul;48(4):733–42.
- Toldo S, Bussani R, Nuzzi V, Bonaventura A, Mauro AG, Cannatà A, et al. Inflammasome formation in the lungs of patients with fatal COVID-19. Inflamm Res. 2021 Jan;70(1):7–10.
- Rodrigues TS, de Sá KSG, Ishimoto AY, Becerra A, Oliveira S, Almeida L, et al. Inflammasomes are activated in response to SARS-CoV-2 infection and are associated with COVID-19 severity in patients. J Exp Med. 2021 Mar 1;218(3).
- Gudbjartsson DF, Norddahl GL, Melsted P, Gunnarsdottir K, Holm H, Eythorsson E, et al. Humoral Immune Response to SARS-CoV-2 in Iceland. N Engl J Med. 2020 Oct 29;383(18):1724–34.
- Ge XY, Li JL, Yang XL, Chmura AA, Zhu G, Epstein JH, et al. Isolation and characterization of a bat SARS-like coronavirus that uses the ACE2 receptor. Nature. 2013 Nov 28;503(7477):535–8.
- Zhang T, Wu Q, Zhang Z. Probable Pangolin Origin of SARS-CoV-2 Associated with the COVID-19 Outbreak. Curr Biol. 2020 Apr 6;30(7):1346-1351.e2.
- Wölfel R, Corman VM, Guggemos W, Seilmaier M, Zange S, Müller MA, et al. Virological assessment of hospitalized patients with COVID-2019. Nature. 2020 May;581(7809):465–9.
- Cevik M, Bamford CGG, Ho A. COVID-19 pandemic-a focused review for clinicians. Clin Microbiol Infect. 2020 Jul;26(7):842–7.
- Cevik M, Tate M, Lloyd O, Maraolo AE, Schafers J, Ho A. SARS-CoV-2, SARS-CoV, and MERS-CoV viral load dynamics, duration of viral shedding, and infectiousness: a systematic review and meta-analysis. Lancet Microbe. 2021 Jan;2(1):e13–22.
- Shereen MA, Khan S, Kazmi A, Bashir N, Siddique R. COVID-19 infection: Origin, transmission, and characteristics of human coronaviruses. J Adv Res. 2020 Jul;24:91–8.
- Lam TTY, Jia N, Zhang YW, Shum MHH, Jiang JF, Zhu HC, et al. Identifying SARS-CoV-2-related coronaviruses in Malayan pangolins. Nature. 2020 Jul;583(7815):282–5.
- Ma J, Chen Y, Wu W, Chen Z. Structure and Function of N-Terminal Zinc Finger Domain of SARS-CoV-2 NSP2. Virol Sin. 2021 Oct;36(5):1104–12.
- Sarkar S, Runge B, Russell RW, Movellan KT, Calero D, Zeinalilathori S, et al. Atomic-Resolution Structure of SARS-CoV-2 Nucleocapsid Protein N-Terminal Domain. J Am Chem Soc. 2022 Jun 15;144(23):10543–55.
- Peng Y, Du N, Lei Y, Dorje S, Qi J, Luo T, et al. Structures of the SARS-CoV-2 nucleocapsid and their perspectives for drug design. EMBO J. 2020 Oct 15;39(20):e105938.
- Zhou P, Yang XL, Wang XG, Hu B, Zhang L, Zhang W, et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature. 2020 Mar;579(7798):270–3.
- Chang C ke, Hou MH, Chang CF, Hsiao CD, Huang T huang. The SARS coronavirus nucleocapsid protein--forms and functions. Antiviral Res. 2014 Mar;103:39–50.
- Dinesh DC, Chalupska D, Silhan J, Koutna E, Nencka R, Veverka V, et al. Structural basis of RNA recognition by the SARS-CoV-2 nucleocapsid phosphoprotein. PLoS Pathog. 2020 Dec;16(12):e1009100.
- Liu C, Ginn HM, Dejnirattisai W, Supasa P, Wang B, Tuekprakhon A, et al. Reduced neutralization of SARS-CoV-2 B.1.617 by vaccine and convalescent serum. Cell. 2021 Aug 5;184(16):4220-4236.e13.
- Zinzula L, Basquin J, Bohn S, Beck F, Klumpe S, Pfeifer G, et al. High-resolution structure and biophysical characterization of the nucleocapsid phosphoprotein dimerization domain from the Covid-19 severe acute respiratory syndrome coronavirus 2. Biochem Biophys Res Commun. 2021 Jan 29;538:54–62.
- Weisblum Y, Schmidt F, Zhang F, DaSilva J, Poston D, Lorenzi JC, et al. Escape from neutralizing antibodies by SARS-CoV-2 spike protein variants. Elife. 2020 Oct 28;9.
- Eltayeb A, Al-Sarraj F, Alharbi M, Albiheyri R, Mattar E, Abu Zeid IM, et al. Overview of the SARS-CoV-2 nucleocapsid protein. Int J Biol Macromol. 2024 Mar;260(Pt 2):129523.
- Gori Savellini G, Anichini G, Gandolfo C, Cusi MG. SARS-CoV-2 N Protein Targets TRIM25-Mediated RIG-I Activation to Suppress Innate Immunity. Viruses. 2021 Jul 23;13(8).
- Zhao Y, Sui L, Wu P, Wang W, Wang Z, Yu Y, et al. A dual-role of SARS-CoV-2 nucleocapsid protein in regulating innate immune response. Signal Transduct Target Ther. 2021 Sep 1;6(1):331.
- Oh SJ, Shin OS. SARS-CoV-2 Nucleocapsid Protein Targets RIG-I-Like Receptor Pathways to Inhibit the Induction of Interferon Response. Cells. 2021 Mar 2;10(3).
- Zheng Y, Deng J, Han L, Zhuang MW, Xu Y, Zhang J, et al. SARS-CoV-2 NSP5 and N protein counteract the RIG-I signaling pathway by suppressing the formation of stress granules. Signal Transduct Target Ther. 2022 Jan 24;7(1):22.
- Wang S, Dai T, Qin Z, Pan T, Chu F, Lou L, et al. Targeting liquid-liquid phase separation of SARS-CoV-2 nucleocapsid protein promotes innate antiviral immunity by elevating MAVS activity. Nat Cell Biol. 2021 Jul;23(7):718–32.
- Xia H, Cao Z, Xie X, Zhang X, Chen JYC, Wang H, et al. Evasion of Type I Interferon by SARS-CoV-2. Cell Rep. 2020 Oct 6;33(1):108234.
- Lei X, Dong X, Ma R, Wang W, Xiao X, Tian Z, et al. Activation and evasion of type I interferon responses by SARS-CoV-2. Nat Commun. 2020 Jul 30;11(1):3810.
- Li JY, Liao CH, Wang Q, Tan YJ, Luo R, Qiu Y, et al. The ORF6, ORF8 and nucleocapsid proteins of SARS-CoV-2 inhibit type I interferon signaling pathway. Virus Res. 2020 Sep;286:198074.
- Chen K, Xiao F, Hu D, Ge W, Tian M, Wang W, et al. SARS-CoV-2 Nucleocapsid Protein Interacts with RIG-I and Represses RIG-Mediated IFN-β Production. Viruses. 2020 Dec 30;13(1):47.
- Mu J, Fang Y, Yang Q, Shu T, Wang A, Huang M, et al. SARS-CoV-2 N protein antagonizes type I interferon signaling by suppressing phosphorylation and nuclear translocation of STAT1 and STAT2. Cell Discov. 2020 Sep 15;6(1):65.
- Cui L, Wang H, Ji Y, Yang J, Xu S, Huang X, et al. The Nucleocapsid Protein of Coronaviruses Acts as a Viral Suppressor of RNA Silencing in Mammalian Cells. J Virol. 2015 Sep;89(17):9029–43.
- Mu J, Xu J, Zhang L, Shu T, Wu D, Huang M, et al. SARS-CoV-2-encoded nucleocapsid protein acts as a viral suppressor of RNA interference in cells. Sci China Life Sci. 2020 Sep;63(9):1413–6.
- Chen YM, Wei JL, Qin RS, Hou JP, Zang GC, Zhang GY, et al. Folic acid: a potential inhibitor against SARS-CoV-2 nucleocapsid protein. Pharm Biol. 2022 Dec;60(1):862–78.
- Visvabharathy L, Hanson B, Orban Z, Lim PH, Palacio N, Jain R, et al. Neuro-COVID long-haulers exhibit broad dysfunction in T cell memory generation and responses to vaccination. medRxiv [Internet]. 2021 Aug 9. Available online: http://www.ncbi.nlm.nih.gov/pubmed/34401886.
- Yang L, Xu Q, Yang B, Li J, Dong R, Da J, et al. IgG antibody titers against SARS-CoV-2 nucleocapsid protein correlate with the severity of COVID-19 patients. BMC Microbiol. 2021 Dec 18;21(1):351.
- Feng W, Xiang Y, Wu L, Chen Z, Li Q, Chen J, et al. Nucleocapsid protein of SARS-CoV-2 is a potential target for developing new generation of vaccine. J Clin Lab Anal. 2022 Jun;36(6):e24479.
- Sen SR, Sanders EC, Gabriel KN, Miller BM, Isoda HM, Salcedo GS, et al. Predicting COVID-19 Severity with a Specific Nucleocapsid Antibody plus Disease Risk Factor Score. mSphere. 2021 Apr 28;6(2).
- Voss C, Esmail S, Liu X, Knauer MJ, Ackloo S, Kaneko T, et al. Epitope-specific antibody responses differentiate COVID-19 outcomes and variants of concern. JCI Insight. 2021 Jul 8;6(13).
- Lyudovyk O, Kim JY, Qualls D, Hwee MA, Lin YH, Boutemine SR, et al. Impaired humoral immunity is associated with prolonged COVID-19 despite robust CD8 T cell responses. Cancer Cell. 2022 Jul 11;40(7):738-753.e5.
- Domingo López-Muñoz A, Kosik I, Holly J, Yewdell JW. Cell surface SARS-CoV-2 nucleocapsid protein modulates innate and adaptive immunity. Vol. 8, Sci. Adv. 2022.
- Hartley GE, Edwards ESJ, Aui PM, Varese N, Stojanovic S, McMahon J, et al. Rapid generation of durable B cell memory to SARS-CoV-2 spike and nucleocapsid proteins in COVID-19 and convalescence. Sci Immunol. 2020 Dec 18;5(54).
- Lee CY, Shah MK, Hoyos D, Solovyov A, Douglas M, Taur Y, et al. Prolonged SARS-CoV-2 Infection in Patients with Lymphoid Malignancies. Cancer Discov. 2022 Jan 1;12(1):62–73.
- Nelson RW, Chen Y, Venezia OL, Majerus RM, Shin DS, MGH COVID-19 Collection & Processing Team, et al. SARS-CoV-2 epitope-specific CD4+ memory T cell responses across COVID-19 disease severity and antibody durability. Sci Immunol. 2022 Jul 22;7(73):eabl9464.
- Martner A, Grauers Wiktorin H, Törnell A, Ringlander J, Arabpour M, Lindh M, et al. Transient and durable T cell reactivity after COVID-19. Proc Natl Acad Sci U S A. 2022 Jul 26;119(30):e2203659119.
- Bange EM, Han NA, Wileyto P, Kim JY, Gouma S, Robinson J, et al. CD8+ T cells contribute to survival in patients with COVID-19 and hematologic cancer. Nat Med. 2021 Jul;27(7):1280–9.
- Peng Y, Felce SL, Dong D, Penkava F, Mentzer AJ, Yao X, et al. An immunodominant NP105-113-B*07:02 cytotoxic T cell response controls viral replication and is associated with less severe COVID-19 disease. Nat Immunol. 2022 Jan;23(1):50–61.
- Nguyen THO, Rowntree LC, Petersen J, Chua BY, Hensen L, Kedzierski L, et al. CD8+ T cells specific for an immunodominant SARS-CoV-2 nucleocapsid epitope display high naive precursor frequency and TCR promiscuity. Immunity. 2021 May 11;54(5):1066-1082.e5.
- Lineburg KE, Grant EJ, Swaminathan S, Chatzileontiadou DSM, Szeto C, Sloane H, et al. CD8+ T cells specific for an immunodominant SARS-CoV-2 nucleocapsid epitope cross-react with selective seasonal coronaviruses. Immunity. 2021 May 11;54(5):1055-1065.e5.
- GREY HM, KUNKEL HG. H CHAIN SUBGROUPS OF MYELOMA PROTEINS AND NORMAL 7S GAMMA-GLOBULIN. J Exp Med. 1964 Aug 1;120(2):253–66.
- TERRY WD, FAHEY JL. SUBCLASSES OF HUMAN GAMMA-2-GLOBULIN BASED ON DIFFERENCES IN THE HEAVY POLYPEPTIDE CHAINS. Science. 1964 Oct 16;146(3642):400–1.
- Mayumi M, Kuritani T, Kubagawa H, Cooper MD. IgG subclass expression by human B lymphocytes and plasma cells: B lymphocytes precommitted to IgG subclass can be preferentially induced by polyclonal mitogens with T cell help. J Immunol. 1983 Feb;130(2):671–7.
- Atanackovic D, Avila S V., Lutfi F, de Miguel-Perez D, Fan X, Sanchez-Petitto G, et al. Deep dissection of the antiviral immune profile of patients with COVID-19. Commun Biol. 2021 Dec 16;4(1):1389.
- Chakraborty S, Gonzalez J, Edwards K, Mallajosyula V, Buzzanco AS, Sherwood R, et al. Proinflammatory IgG Fc structures in patients with severe COVID-19. Nat Immunol. 2021 Jan;22(1):67–73.
- Yates JL, Ehrbar DJ, Hunt DT, Girardin RC, Dupuis AP, Payne AF, et al. Serological analysis reveals an imbalanced IgG subclass composition associated with COVID-19 disease severity. Cell Rep Med. 2021 Jul 20;2(7):100329.
- Korobova ZR, Zueva E V, Arsentieva NA, Batsunov OK, Liubimova NE, Khamitova I V, et al. Changes in Anti-SARS-CoV-2 IgG Subclasses over Time and in Association with Disease Severity. Viruses. 2022 Apr 29;14(5).
- Moura AD, da Costa HHM, Correa VA, de S Lima AK, Lindoso JAL, De Gaspari E, et al. Assessment of avidity related to IgG subclasses in SARS-CoV-2 Brazilian infected patients. Sci Rep. 2021 Sep 3;11(1):17642.
- Yang X, Wang F, Zhang Y, Wang L, Antonenko S, Zhang S, et al. Comprehensive Analysis of the Therapeutic IgG4 Antibody Pembrolizumab: Hinge Modification Blocks Half Molecule Exchange In Vitro and In Vivo. J Pharm Sci. 2015 Dec;104(12):4002–14.
- Schuurman J, Van Ree R, Perdok GJ, Van Doorn HR, Tan KY, Aalberse RC. Normal human immunoglobulin G4 is bispecific: it has two different antigen-combining sites. Immunology. 1999 Aug;97(4):693–8.
- Buhre JS, Pongracz T, Künsting I, Lixenfeld AS, Wang W, Nouta J, et al. mRNA vaccines against SARS-CoV-2 induce comparably low long-term IgG Fc galactosylation and sialylation levels but increasing long-term IgG4 responses compared to an adenovirus-based vaccine. Front Immunol. 2022;13:1020844.
- Hartley GE, Fryer HA, Gill PA, Boo I, Bornheimer SJ, Hogarth PM, et al. Third dose COVID-19 mRNA vaccine enhances IgG4 isotype switching and recognition of Omicron subvariants by memory B cells after mRNA but not adenovirus priming. 2023.
- Yoshimura M, Sakamoto A, Ozuru R, Kurihara Y, Itoh R, Ishii K, et al. The appearance of anti-spike receptor binding domain immunoglobulin G4 responses after repetitive immunization with messenger RNA-based COVID-19 vaccines. Int J Infect Dis. 2024 Feb;139:1–5.
- Kiszel P, Sík P, Miklós J, Kajdácsi E, Sinkovits G, Cervenak L, et al. Class switch towards spike protein-specific IgG4 antibodies after SARS-CoV-2 mRNA vaccination depends on prior infection history. Sci Rep. 2023 Aug 13;13(1):13166.
- Akhtar M, Islam MR, Khaton F, Soltana UH, Jafrin SA, Rahman SIA, et al. Appearance of tolerance-induction and non-inflammatory SARS-CoV-2 spike-specific IgG4 antibodies after COVID-19 booster vaccinations. Front Immunol. 2023;14:1309997.
- Espino AM, Armina-Rodriguez A, Alvarez L, Ocasio-Malavé C, Ramos-Nieves R, Rodriguez Martinó EI, et al. The Anti-SARS-CoV-2 IgG1 and IgG3 Antibody Isotypes with Limited Neutralizing Capacity against Omicron Elicited in a Latin Population a Switch toward IgG4 after Multiple Doses with the mRNA Pfizer-BioNTech Vaccine. Viruses. 2024 Jan 26;16(2).
- Chiang HL, Liang KH, Lu RM, Kuo TW, Lin Y, Wu HC. Broadly neutralizing human antibodies against Omicron subvariants of SARS-CoV-2. J Biomed Sci. 2023 Jul 31;30(1):59.
- Zhou H, Møhlenberg M, Thakor JC, Tuli HS, Wang P, Assaraf YG, et al. Sensitivity to Vaccines, Therapeutic Antibodies, and Viral Entry Inhibitors and Advances To Counter the SARS-CoV-2 Omicron Variant. Clin Microbiol Rev. 2022 Sep 21;35(3).
- Starr TN, Greaney AJ, Stewart CM, Walls AC, Hannon WW, Veesler D, et al. Deep mutational scans for ACE2 binding, RBD expression, and antibody escape in the SARS-CoV-2 Omicron BA.1 and BA.2 receptor-binding domains. PLoS Pathog. 2022 Nov 18;18(11):e1010951.
- Xue S, Han Y, Wu F, Wang Q. Mutations in the SARS-CoV-2 spike receptor binding domain and their delicate balance between ACE2 affinity and antibody evasion. Protein Cell. 2024 May 28;15(6):403–18.
- Geng Q, Shi K, Ye G, Zhang W, Aihara H, Li F. Structural Basis for Human Receptor Recognition by SARS-CoV-2 Omicron Variant BA.1. J Virol. 2022 Apr 27;96(8).
- Liu L, Iketani S, Guo Y, Chan JFW, Wang M, Liu L, et al. Striking antibody evasion manifested by the Omicron variant of SARS-CoV-2. Nature. 2022 Feb 24;602(7898):676–81.
- Shah M, Woo HG. Omicron: A Heavily Mutated SARS-CoV-2 Variant Exhibits Stronger Binding to ACE2 and Potently Escapes Approved COVID-19 Therapeutic Antibodies. Front Immunol. 2021;12:830527.
- Renner TM, Stuible M, Cass B, Perret S, Guimond J, Lord-Dufour S, et al. Reduced cross-protective potential of Omicron compared to ancestral SARS-CoV-2 spike vaccines against potentially zoonotic coronaviruses. npj Viruses. 2024 Nov 21;2(1):58.
- Zhang J, Cai Y, Lavine CL, Peng H, Zhu H, Anand K, et al. Structural and functional impact by SARS-CoV-2 Omicron spike mutations. Cell Rep. 2022 Apr 26;39(4):110729.
- Dhawan M, Saied AA, Mitra S, Alhumaydhi FA, Emran T Bin, Wilairatana P. Omicron variant (B.1.1.529) and its sublineages: What do we know so far amid the emergence of recombinant variants of SARS-CoV-2? Biomed Pharmacother. 2022 Oct;154:113522.
- Dejnirattisai W, Huo J, Zhou D, Zahradník J, Supasa P, Liu C, et al. SARS-CoV-2 Omicron-B.1.1.529 leads to widespread escape from neutralizing antibody responses. Cell. 2022 Feb 3;185(3):467-484.e15.
- Planas D, Saunders N, Maes P, Guivel-Benhassine F, Planchais C, Buchrieser J, et al. Considerable escape of SARS-CoV-2 Omicron to antibody neutralization. Nature. 2022 Feb;602(7898):671–5.
- Wussow F, Kha M, Faircloth K, Nguyen VH, Iniguez A, Martinez J, et al. COH04S1 and beta sequence-modified vaccine protect hamsters from SARS-CoV-2 variants. iScience. 2022 Jun 17;25(6):104457.
- Chiuppesi F, Zaia JA, Frankel PH, Stan R, Drake J, Williams B, et al. Safety and immunogenicity of a synthetic multiantigen modified vaccinia virus Ankara-based COVID-19 vaccine (COH04S1): an open-label and randomised, phase 1 trial. Lancet Microbe. 2022 Apr;3(4):e252–64.
- Jiang W, Shi L, Cai L, Wang X, Li J, Li H, et al. A two-adjuvant multiantigen candidate vaccine induces superior protective immune responses against SARS-CoV-2 challenge. Cell Rep. 2021 Dec 14;37(11):110112.
- Jia Q, Bielefeldt-Ohmann H, Maison RM, Masleša-Galić S, Cooper SK, Bowen RA, et al. Replicating bacterium-vectored vaccine expressing SARS-CoV-2 Membrane and Nucleocapsid proteins protects against severe COVID-19-like disease in hamsters. NPJ Vaccines. 2021 Mar 30;6(1):47.
- Ahn JY, Lee J, Suh YS, Song YG, Choi YJ, Lee KH, et al. Safety and immunogenicity of two recombinant DNA COVID-19 vaccines containing the coding regions of the spike or spike and nucleocapsid proteins: an interim analysis of two open-label, non-randomised, phase 1 trials in healthy adults. Lancet Microbe. 2022 Mar;3(3):e173–83.
- Hajnik RL, Plante JA, Liang Y, Alameh MG, Tang J, Bonam SR, et al. Dual spike and nucleocapsid mRNA vaccination confer protection against SARS-CoV-2 Omicron and Delta variants in preclinical models. Sci Transl Med. 2022 Sep 14;14(662):eabq1945.
- Castro JT, Azevedo P, Fumagalli MJ, Hojo-Souza NS, Salazar N, Almeida GG, et al. Promotion of neutralizing antibody-independent immunity to wild-type and SARS-CoV-2 variants of concern using an RBD-Nucleocapsid fusion protein. Nat Commun. 2022 Aug 17;13(1):4831.
- Afkhami S, D’Agostino MR, Zhang A, Stacey HD, Marzok A, Kang A, et al. Respiratory mucosal delivery of next-generation COVID-19 vaccine provides robust protection against both ancestral and variant strains of SARS-CoV-2. Cell. 2022 Mar 3;185(5):896-915.e19.
- Wang CY, Hwang KP, Kuo HK, Peng WJ, Shen YH, Kuo BS, et al. A multitope SARS-CoV-2 vaccine provides long-lasting B cell and T cell immunity against Delta and Omicron variants. J Clin Invest. 2022 May 16;132(10).
- Schur, PH. IgG subclasses--a review. Ann Allergy. 1987 Feb;58(2):89–96, 99.
- Hui, DS. Hybrid immunity and strategies for COVID-19 vaccination. Lancet Infect Dis. 2023 Jan;23(1):2–3.
- Gelderloos AT, Verheul MK, Middelhof I, de Zeeuw-Brouwer ML, van Binnendijk RS, Buisman AM, et al. Repeated COVID-19 mRNA vaccination results in IgG4 class switching and decreased NK cell activation by S1-specific antibodies in older adults. Immunity & Ageing. 2024 Sep 14;21(1):63.
- Chiuppesi F, Nguyen VH, Park Y, Contreras H, Karpinski V, Faircloth K, et al. Synthetic multiantigen MVA vaccine COH04S1 protects against SARS-CoV-2 in Syrian hamsters and non-human primates. NPJ Vaccines. 2022 Jan 21;7(1):7.
| Vaccinated | Non-vaccinated (control) | ||||||
|---|---|---|---|---|---|---|---|
| Sample number | Mean (X) | Standard Deviation (SD) | Gender | Sample number | Mean (X) | Standard Deviation (SD) | Gender |
| #1 | 2.250 | 0.042 | Female | #1 | 0.512 | 0.167 | Female |
| #2 | 2.336 | 0.089 | Male | #2 | 0.752 | 0.116 | Male |
| #3 | 2.319 | 0.062 | Female | #3 | 0.775 | 0.139 | Female |
| #4 | 2.229 | 0.030 | Female | #4 | 0.586 | 0.178 | Female |
| #5 | 2.258 | 0.011 | Female | #5 | 0.819 | 0.130 | Female |
| #6 | 2.183 | 0.080 | Male | #6 | 0.708 | 0.083 | Female |
| #7 | 2.10 | 0.25 | Female | #7 | 0.549 | 0.116 | Male |
| #8 | 2.01 | 0.35 | Female | #8 | 0.812 | 0.113 | Female |
| #9 | 2.31 | 0.10 | Female | #9 | 0.715 | 0.116 | Female |
| #10 | 2.28 | 0.15 | Female | #10 | 0.586 | 0.178 | Female |
| #11 | 2.178 | 0.096 | Male | #11 | 0.756 | 0.160 | Male |
| #12 | 2.306 | 0.011 | Male | #12 | 0.660 | 0.195 | Female |
| #13 | 2.10 | 0.23 | Male | #13 | 0.745 | 0.214 | Female |
| #14 | 2.16 | 0.15 | Female | #14 | 0.727 | 0.112 | Male |
| #15 | 1.88 | 0.26 | Female | #15 | 0.518 | 0.001 | Male |
| #16 | 0.84 | 0.56 | Female | #16 | 0.512 | 0.167 | Male |
| #17 | 2.451 | 0.014 | Male | #17 | 0.752 | 0.116 | Female |
| #18 | 1.70 | 0.12 | Male | #18 | 0.775 | 0.139 | Male |
| #19 | 2.412 | 0.048 | Male | #19 | 0.586 | 0.178 | Female |
| #20 | 2.592 | 0.016 | Male | #20 | 0.819 | 0.130 | Male |
| #21 | 2.434 | 0.100 | Male | #21 | 0.708 | 0.083 | Female |
| #22 | 2.486 | 0.023 | Male | #22 | 0.549 | 0.116 | Male |
| #23 | 2.47 | 0.11 | Male | #23 | 0.812 | 0.113 | Female |
| #24 | 2.441 | 0.032 | Male | #24 | 0.715 | 0.116 | Female |
| #25 | 2.45 | 0.10 | Male | #25 | 0.586 | 0.178 | Male |
| #26 | 2.363 | 0.034 | Female | #26 | 0.756 | 0.160 | Male |
| #27 | 2.501 | 0.056 | Female | #27 | 0.660 | 0.195 | Male |
| #28 | 1.71 | 0.42 | Female | #28 | 0.745 | 0.214 | Female |
| #29 | 1.4 | 1.2 | Male | #29 | 0.727 | 0.112 | Male |
| #30 | 1.88 | 0.26 | Male | #30 | 0.624 | 0.001 | Female |
| Vaccinated | Non-vaccinated (control) | ||||||
|---|---|---|---|---|---|---|---|
| Sample number | Mean (X) | Standard Deviation (SD) | Gender | Sample number | Mean (X) | Standard Deviation (SD) | Gender |
| #1 | 0.1443 | 0.0083 | Female | #1 | 0.0663 | 0.0021 | Female |
| #2 | 0.2797 | 0.0108 | Male | #2 | 0.1203 | 0.0065 | Male |
| #3 | 1.0117 | 0.0175 | Female | #3 | 0.1120 | 0.0312 | Female |
| #4 | 1.5940 | 0.0035 | Female | #4 | 0.0653 | 0.0035 | Female |
| #5 | 1.2063 | 0.0172 | Female | #5 | 0.0547 | 0.0021 | Female |
| #6 | 0.8613 | 0.0266 | Male | #6 | 0.0753 | 0.0023 | Female |
| #7 | 0.4510 | 0.0061 | Female | #7 | 0.0530 | 0.0010 | Male |
| #8 | 0.6750 | 0.0115 | Female | #8 | 0.1450 | 0.0092 | Female |
| #9 | 0.1003 | 0.0029 | Female | #9 | 0.0963 | 0.0042 | Female |
| #10 | 1.1467 | 0.0382 | Female | #10 | 0.0887 | 0.0029 | Female |
| #11 | 1.7470 | 0.0682 | Male | #11 | 0.1003 | 0.0251 | Male |
| #12 | 0.9363 | 0.0064 | Male | #12 | 0.0677 | 0.0038 | Female |
| #13 | 0.5203 | 0.0210 | Male | #13 | 0.3127 | 0.0129 | Female |
| #14 | 0.8817 | 0.0278 | Female | #14 | 0.0673 | 0.0029 | Male |
| #15 | 0.5375 | 0.0078 | Female | #15 | 0.2680 | 0.0212 | Male |
| #16 | 1.4877 | 0.0083 | Female | #16 | 0.2603 | 0.0285 | Male |
| #17 | 1.3603 | 0.0641 | Male | #17 | 0.0613 | 0.0023 | Female |
| #18 | 0.5377 | 0.0818 | Male | #18 | 0.3517 | 0.0619 | Male |
| #19 | 0.0500 | 0.0028 | Male | #19 | 0.0773 | 0.0021 | Female |
| #20 | 2.3477 | 0.3050 | Male | #20 | 0.0853 | 0.0040 | Male |
| #21 | 1.1397 | 0.3751 | Male | #21 | 0.2090 | 0.0215 | Female |
| #22 | 1.7917 | 0.1093 | Male | #22 | 0.1383 | 0.0169 | Male |
| #23 | 1.7467 | 0.0587 | Male | #23 | 0.0783 | 0.0114 | Female |
| #24 | 0.7747 | 0.5534 | Male | #24 | 0.2033 | 0.1409 | Female |
| #25 | 1.4160 | 0.1129 | Male | #25 | 0.125 | 0.065 | Male |
| #26 | 1.8209 | 0.0665 | Female | #26 | 0.128 | 0.030 | Male |
| #27 | 0.8823 | 0.4975 | Female | #27 | 0.073 | 0.005 | Male |
| #28 | 1.2237 | 0.4916 | Female | #28 | 0.064 | 0.0 | Female |
| #29 | 0.3290 | 0.3741 | Male | #29 | 0.373 | 0.048 | Male |
| #30 | 0.5275 | 0.0035 | Male | #30 | 0.061 | 0.004 | Female |
| Vaccinated | Non-vaccinated (control) | ||||||
|---|---|---|---|---|---|---|---|
| Sample number | Mean (X) | Standard Deviation (SD) | Gender | Sample number | Mean (X) | Standard Deviation (SD) | Gender |
| #1 | 0.0583 | 0.0012 | Female | #1 | 0.062 | 0.0028 | Female |
| #2 | 0.059 | 0.0016 | Male | #2 | 0.0537 | 0.0005 | Male |
| #3 | 0.0593 | 0.0017 | Female | #3 | 0.0533 | 0.0026 | Female |
| #4 | 0.0553 | 0.0034 | Female | #4 | 0.0507 | 0.0005 | Female |
| #5 | 0.0537 | 0.0005 | Female | #5 | 0.0523 | 0.0029 | Female |
| #6 | 0.0563 | 0.0050 | Male | #6 | 0.0543 | 0.0012 | Female |
| #7 | 0.0597 | 0.0123 | Female | #7 | 0.0503 | 0.0029 | Male |
| #8 | 0.0583 | 0.0060 | Female | #8 | 0.0517 | 0.0033 | Female |
| #9 | 0.0547 | 0.0054 | Female | #9 | 0.0613 | 0.0069 | Female |
| #10 | 0.0503 | 0.0024 | Female | #10 | 0.0513 | 0.0021 | Female |
| #11 | 0.0533 | 0.0061 | Male | #11 | 0.0507 | 0.0025 | Male |
| #12 | 0.053 | 0.0029 | Male | #12 | 0.078 | 0.0439 | Female |
| #13 | 0.0500 | 0.0014 | Male | #13 | 0.0497 | 0.0005 | Female |
| #14 | 0.0507 | 0.0025 | Female | #14 | 0.074 | 0.0325 | Male |
| #15 | 0.0525 | 0.0005 | Female | #15 | 0.054 | 0.0030 | Male |
| #16 | 0.0513 | 0.0017 | Female | #16 | 0.0527 | 0.0017 | Male |
| #17 | 0.0510 | 0.0022 | Male | #17 | 0.07 | 0.0283 | Female |
| #18 | 0.0527 | 0.0033 | Male | #18 | 0.1997 | 0.2124 | Male |
| #19 | 0.0503 | 0.0012 | Male | #19 | 0.2060 | 0.2221 | Female |
| #20 | 0.0527 | 0.0066 | Male | #20 | 0.0497 | 0.0012 | Male |
| #21 | 0.0500 | 0.0014 | Male | #21 | 0.061 | 0.0136 | Female |
| #22 | 0.0517 | 0.0012 | Male | #22 | 0.055 | 0.0043 | Male |
| #23 | 0.062 | 0.0008 | Male | #23 | 0.0527 | 0.0034 | Female |
| #24 | 0.0527 | 0.0017 | Male | #24 | 0.0503 | 0.0029 | Female |
| #25 | 0.0603 | 0.0111 | Male | #25 | 0.0527 | 0.0029 | Male |
| #26 | 0.0570 | 0.0042 | Female | #26 | 0.053 | 0.0022 | Male |
| #27 | 0.0550 | 0.0036 | Female | #27 | 0.051 | 0.0008 | Male |
| #28 | 0.0587 | 0.0012 | Female | #28 | 0.0537 | 0.0025 | Female |
| #29 | 0.0597 | 0.0019 | Male | #29 | 0.0553 | 0.0019 | Male |
| #30 | 0.059 | 0.0010 | Male | #30 | 0.063 | 0.0010 | Female |
| Vaccinated | Non-vaccinated (control) | ||||||
|---|---|---|---|---|---|---|---|
| Sample number | Mean (X) | Standard Deviation (SD) | Gender | Sample number | Mean (X) | Standard Deviation (SD) | Gender |
| #1 | 0.0527 | 0.0019 | Female | #1 | 0.052 | 0.00163 | Female |
| #2 | 0.0477 | 0.0012 | Male | #2 | 0.05067 | 0.00047 | Male |
| #3 | 0.0493 | 0.0040 | Female | #3 | 0.158 | 0.03590 | Female |
| #4 | 0.0500 | 0.0022 | Female | #4 | 0.04933 | 0.00170 | Female |
| #5 | 0.0507 | 0.0012 | Female | #5 | 0.045 | 0.00082 | Female |
| #6 | 0.0550 | 0.0024 | Male | #6 | 0.09833 | 0.00974 | Female |
| #7 | 0.0557 | 0.0005 | Female | #7 | 0.04767 | 0.00189 | Male |
| #8 | 0.0520 | 0.0022 | Female | #8 | 0.054 | 0.00432 | Female |
| #9 | 0.0507 | 0.0012 | Female | #9 | 0.07633 | 0.00386 | Female |
| #10 | 0.0477 | 0.0005 | Female | #10 | 0.05133 | 0.00047 | Female |
| #11 | 0.0547 | 0.0038 | Male | #11 | 0.04733 | 0.00170 | Male |
| #12 | 0.0473 | 0.0019 | Male | #12 | 0.046 | 0.00082 | Female |
| #13 | 0.0603 | 0.0005 | Male | #13 | 0.139 | 0.01431 | Female |
| #14 | 0.0470 | 0.0014 | Female | #14 | 0.04733 | 0.00094 | Male |
| #15 | 0.0480 | 0.0010 | Female | #15 | 0.0485 | 0.00050 | Male |
| #16 | 0.0690 | 0.0065 | Female | #16 | 0.061 | 0.00374 | Male |
| #17 | 0.0643 | 0.0026 | Male | #17 | 0.06133 | 0.00544 | Female |
| #18 | 0.0753 | 0.0070 | Male | #18 | 0.06533 | 0.00411 | Male |
| #19 | 0.0680 | 0.0102 | Male | #19 | 0.054 | 0.00163 | Female |
| #20 | 0.0703 | 0.0097 | Male | #20 | 0.06033 | 0.00170 | Male |
| #21 | 0.0577 | 0.0049 | Male | #21 | 0.059 | 0.00216 | Female |
| #22 | 0.0537 | 0.0033 | Male | #22 | 0.056 | 0.00141 | Male |
| #23 | 0.0570 | 0.0059 | Male | #23 | 0.05533 | 0.00249 | Female |
| #24 | 0.0500 | 0.0036 | Male | #24 | 0.06367 | 0.00713 | Female |
| #25 | 0.0633 | 0.0062 | Male | #25 | 0.05767 | 0.00236 | Male |
| #26 | 0.0557 | 0.0033 | Female | #26 | 0.052 | 0.0057 | Male |
| #27 | 0.0660 | 0.0029 | Female | #27 | 0.0553 | 0.0050 | Male |
| #28 | 0.0537 | 0.0005 | Female | #28 | 0.052 | 0.0022 | Female |
| #29 | 0.0540 | 0.0008 | Male | #29 | 0.0507 | 0.0029 | Male |
| #30 | 0.0620 | 0.0090 | Male | #30 | 0.0515 | 0.0045 | Female |
| Vaccinated | Non-vaccinated (control) | ||||||
|---|---|---|---|---|---|---|---|
| Sample number | Mean (X) | Standard Deviation (SD) | Gender | Sample number | Mean (X) | Standard Deviation (SD) | Gender |
| #1 | 0.049 | 0.00082 | Female | #1 | 0.046 | 0.00082 | Female |
| #2 | 0.050 | 0.00283 | Male | #2 | 0.04467 | 0.00125 | Male |
| #3 | 0.048 | 0.00082 | Female | #3 | 0.04567 | 0.00386 | Female |
| #4 | 0.048 | 0.00216 | Female | #4 | 0.04267 | 0.00309 | Female |
| #5 | 0.04833 | 0.00094 | Female | #5 | 0.04333 | 0.00125 | Female |
| #6 | 0.05067 | 0.00125 | Male | #6 | 0.05233 | 0.00189 | Female |
| #7 | 0.05133 | 0.00205 | Female | #7 | 0.04467 | 0.00189 | Male |
| #8 | 0.04667 | 0.00262 | Female | #8 | 0.046 | 0.00163 | Female |
| #9 | 0.04833 | 0.00613 | Female | #9 | 0.04767 | 0.00330 | Female |
| #10 | 0.046 | 0.00141 | Female | #10 | 0.04467 | 0.00205 | Female |
| #11 | 0.050 | 0.00356 | Male | #11 | 0.043 | 0.00141 | Male |
| #12 | 0.04633 | 0.00170 | Male | #12 | 0.04433 | 0.00125 | Female |
| #13 | 0.05133 | 0.00236 | Male | #13 | 0.06167 | 0.00450 | Female |
| #14 | 0.050 | 0.00455 | Female | #14 | 0.04833 | 0.00249 | Male |
| #15 | 0.047 | 0.001 | Female | #15 | 0.05 | 0.002 | Male |
| #16 | 0.04767 | 0.00047 | Female | #16 | 0.054 | 0.00163 | Male |
| #17 | 0.10133 | 0.00309 | Male | #17 | 0.049 | 0.00141 | Female |
| #18 | 0.04733 | 0.00189 | Male | #18 | 0.04533 | 0.00125 | Male |
| #19 | 0.05967 | 0.00047 | Male | #19 | 0.04533 | 0.00205 | Female |
| #20 | 0.051 | 0.00082 | Male | #20 | 0.04367 | 0.00094 | Male |
| #21 | 0.048 | 0.00163 | Male | #21 | 0.04567 | 0.00094 | Female |
| #22 | 0.04467 | 0.00047 | Male | #22 | 0.04433 | 0.00125 | Male |
| #23 | 0.04767 | 0.00094 | Male | #23 | 0.04467 | 0.00094 | Female |
| #24 | 0.04567 | 0.00125 | Male | #24 | 0.048 | 0.00374 | Female |
| #25 | 0.050 | 0.00141 | Male | #25 | 0.04667 | 0.00047 | Male |
| #26 | 0.04633 | 0.00094 | Female | #26 | 0.043 | 0.00082 | Male |
| #27 | 0.05867 | 0.00047 | Female | #27 | 0.04967 | 0.00309 | Male |
| #28 | 0.04633 | 0.00047 | Female | #28 | 0.04733 | 0.00125 | Female |
| #29 | 0.04467 | 0.00125 | Male | #29 | 0.04667 | 0.00047 | Male |
| #30 | 0.046 | 0.001 | Male | #30 | 0.0455 | 0.0005 | Female |
| Vaccinated | Non-vaccinated (control) | ||||||
|---|---|---|---|---|---|---|---|
| Sample number | Mean (X) | Standard Deviation (SD) | Gender | Sample number | Mean (X) | Standard Deviation (SD) | Gender |
| #1 | 0.0633 | 0.0021 | Female | #1 | 0.073 | 0.0028 | Female |
| #2 | 0.1177 | 0.0050 | Male | #2 | 0.2763 | 0.0109 | Male |
| #3 | 0.1873 | 0.0005 | Female | #3 | 0.0883 | 0.0025 | Female |
| #4 | 0.1987 | 0.0049 | Female | #4 | 0.1123 | 0.0012 | Female |
| #5 | 0.0503 | 0.0012 | Female | #5 | 0.0713 | 0.0012 | Female |
| #6 | 0.055 | 0.0008 | Male | #6 | 0.0457 | 0.0033 | Female |
| #7 | 0.0963 | 0.0041 | Female | #7 | 0.1743 | 0.0033 | Male |
| #8 | 0.1623 | 0.0024 | Female | #8 | 0.1457 | 0.0026 | Female |
| #9 | 0.0477 | 0.0009 | Female | #9 | 0.1467 | 0.0042 | Female |
| #10 | 0.0953 | 0.0048 | Female | #10 | 0.125 | 0.0071 | Female |
| #11 | 0.1203 | 0.0025 | Male | #11 | 0.052 | 0.0016 | Male |
| #12 | 0.2263 | 0.0066 | Male | #12 | 0.0937 | 0.0017 | Female |
| #13 | 0.0527 | 0.0005 | Male | #13 | 0.0970 | 0.0029 | Female |
| #14 | 0.1403 | 0.0005 | Female | #14 | 0.1023 | 0.0062 | Male |
| #15 | 0.2040 | 0.0010 | Female | #15 | 0.087 | 0.0 | Male |
| #16 | 0.3347 | 0.0100 | Female | #16 | 0.2747 | 0.0144 | Male |
| #17 | 0.2223 | 0.0165 | Male | #17 | 0.084 | 0.0029 | Female |
| #18 | 0.108 | 0.0062 | Male | #18 | 0.1393 | 0.0059 | Male |
| #19 | 0.108 | 0.0057 | Male | #19 | 0.0633 | 0.0012 | Female |
| #20 | 0.1993 | 0.0198 | Male | #20 | 0.075 | 0.0033 | Male |
| #21 | 0.2063 | 0.0009 | Male | #21 | 0.0663 | 0.0021 | Female |
| #22 | 0.139 | 0.0067 | Male | #22 | 0.066 | 0.0029 | Male |
| #23 | 0.1303 | 0.0012 | Male | #23 | 0.0457 | 0.0012 | Female |
| #24 | 0.0777 | 0.0017 | Male | #24 | 0.123 | 0.0024 | Female |
| #25 | 0.1837 | 0.0029 | Male | #25 | 0.1467 | 0.0045 | Male |
| #26 | 0.0757 | 0.0009 | Female | #26 | 0.1547 | 0.0054 | Male |
| #27 | 0.3237 | 0.0039 | Female | #27 | 0.0833 | 0.0046 | Male |
| #28 | 0.1863 | 0.0060 | Female | #28 | 0.0553 | 0.0019 | Female |
| #29 | 0.0507 | 0.0029 | Male | #29 | 0.3257 | 0.0017 | Male |
| #30 | 0.0685 | 0.0015 | Male | #30 | 0.046 | 0.001 | Female |
| Vaccinated | Non-vaccinated (control) | ||||||
|---|---|---|---|---|---|---|---|
| Sample number | Mean (X) | Standard Deviation (SD) | Gender | Sample number | Mean (X) | Standard Deviation (SD) | Gender |
| #1 | 0.055 | 0.00216 | Female | #1 | 0.05767 | 0.00287 | Female |
| #2 | 0.0523 | 0.00249 | Male | #2 | 0.050 | 0.00082 | Male |
| #3 | 0.0543 | 0.00125 | Female | #3 | 0.05433 | 0.00047 | Female |
| #4 | 0.0513 | 0.00047 | Female | #4 | 0.054 | 0.00163 | Female |
| #5 | 0.0480 | 0.00141 | Female | #5 | 0.04933 | 0.00047 | Female |
| #6 | 0.0503 | 0.00125 | Male | #6 | 0.05067 | 0.00047 | Female |
| #7 | 0.0587 | 0.00125 | Female | #7 | 0.04767 | 0.00170 | Male |
| #8 | 0.0663 | 0.00236 | Female | #8 | 0.05067 | 0.00125 | Female |
| #9 | 0.0517 | 0.00125 | Female | #9 | 0.049 | 0.00082 | Female |
| #10 | 0.0497 | 0.00094 | Female | #10 | 0.04833 | 0.00094 | Female |
| #11 | 0.0483 | 0.00094 | Male | #11 | 0.051 | 0.00163 | Male |
| #12 | 0.0537 | 0.00094 | Male | #12 | 0.04767 | 0.00205 | Female |
| #13 | 0.048 | 0.0 | Male | #13 | 0.04833 | 0.00170 | Female |
| #14 | 0.04767 | 0.00047 | Female | #14 | 0.0483 | 0.0025 | Male |
| #15 | 0.053 | 0.002 | Female | #15 | 0.0465 | 0.0007 | Male |
| #16 | 0.04933 | 0.00125 | Female | #16 | 0.0617 | 0.0021 | Male |
| #17 | 0.05267 | 0.00094 | Male | #17 | 0.0487 | 0.0012 | Female |
| #18 | 0.052 | 0.00082 | Male | #18 | 0.052 | 0.0017 | Male |
| #19 | 0.04767 | 0.00125 | Male | #19 | 0.056 | 0.0010 | Female |
| #20 | 0.04933 | 0.00205 | Male | #20 | 0.0473 | 0.0012 | Male |
| #21 | 0.051 | 0.00163 | Male | #21 | 0.0517 | 0.0025 | Female |
| #22 | 0.04767 | 0.00125 | Male | #22 | 0.0447 | 0.0015 | Male |
| #23 | 0.047 | 0.00245 | Male | #23 | 0.062 | 0.0036 | Female |
| #24 | 0.04733 | 0.00170 | Male | #24 | 0.0517 | 0.0029 | Female |
| #25 | 0.04867 | 0.00125 | Male | #25 | 0.0527 | 0.0015 | Male |
| #26 | 0.04667 | 0.00125 | Female | #26 | 0.0467 | 0.0006 | Male |
| #27 | 0.05033 | 0.00170 | Female | #27 | 0.050 | 0.0020 | Male |
| #28 | 0.051 | 0.00082 | Female | #28 | 0.0577 | 0.0021 | Female |
| #29 | 0.047 | 0.00082 | Male | #29 | 0.0500 | 0.0016 | Male |
| #30 | 0.0515 | 0.00050 | Male | #30 | 0.0605 | 0.0005 | Female |
| Vaccinated | Non-vaccinated (control) | ||||||
|---|---|---|---|---|---|---|---|
| Sample number | Mean (X) | Standard Deviation (SD) | Gender | Sample number | Mean (X) | Standard Deviation (SD) | Gender |
| #1 | 0.04767 | 0.00170 | Female | #1 | 0.0523 | 0.00094 | Female |
| #2 | 0.048 | 0.00141 | Male | #2 | 0.053 | 0.00082 | Male |
| #3 | 0.04833 | 0.00094 | Female | #3 | 0.050 | 0.0 | Female |
| #4 | 0.052 | 0.00408 | Female | #4 | 0.0633 | 0.00047 | Female |
| #5 | 0.04733 | 0.00249 | Female | #5 | 0.0453 | 0.00094 | Female |
| #6 | 0.047 | 0.00163 | Male | #6 | 0.049 | 0.00163 | Female |
| #7 | 0.06767 | 0.00386 | Female | #7 | 0.0447 | 0.00047 | Male |
| #8 | 0.04667 | 0.00047 | Female | #8 | 0.0467 | 0.00125 | Female |
| #9 | 0.045 | 0.00082 | Female | #9 | 0.0587 | 0.00125 | Female |
| #10 | 0.04533 | 0.00125 | Female | #10 | 0.046 | 0.0 | Female |
| #11 | 0.05833 | 0.00125 | Male | #11 | 0.044 | 0.00163 | Male |
| #12 | 0.06533 | 0.00125 | Male | #12 | 0.0433 | 0.00094 | Female |
| #13 | 0.04567 | 0.00125 | Male | #13 | 0.0443 | 0.00170 | Female |
| #14 | 0.04533 | 0.00047 | Female | #14 | 0.0453 | 0.00094 | Male |
| #15 | 0.049 | 0.0 | Female | #15 | 0.047 | 0.0 | Male |
| #16 | 0.04867 | 0.00047 | Female | #16 | 0.0527 | 0.00125 | Male |
| #17 | 0.04833 | 0.00047 | Male | #17 | 0.0513 | 0.00047 | Female |
| #18 | 0.04733 | 0.00125 | Male | #18 | 0.0603 | 0.00125 | Male |
| #19 | 0.04867 | 0.00125 | Male | #19 | 0.068 | 0.00163 | Female |
| #20 | 0.04933 | 0.00170 | Male | #20 | 0.0473 | 0.00125 | Male |
| #21 | 0.04767 | 0.00047 | Male | #21 | 0.0477 | 0.00094 | Female |
| #22 | 0.04833 | 0.00094 | Male | #22 | 0.047 | 0.00082 | Male |
| #23 | 0.04867 | 0.00249 | Male | #23 | 0.0527 | 0.00125 | Female |
| #24 | 0.046 | 0.00082 | Male | #24 | 0.0497 | 0.00125 | Female |
| #25 | 0.048 | 0.00141 | Male | #25 | 0.0477 | 0.00094 | Male |
| #26 | 0.047 | 0.00163 | Female | #26 | 0.045 | 0.00082 | Male |
| #27 | 0.04767 | 0.00094 | Female | #27 | 0.0447 | 0.00047 | Male |
| #28 | 0.04733 | 0.00125 | Female | #28 | 0.0463 | 0.00094 | Female |
| #29 | 0.04733 | 0.00094 | Male | #29 | 0.04767 | 0.00125 | Male |
| #30 | 0.046 | 0.0 | Male | #30 | 0.05 | 0.0 | Female |
| Vaccinated | Non-vaccinated (control) | ||||||
|---|---|---|---|---|---|---|---|
| Sample number | Mean (X) | Standard Deviation (SD) | Gender | Sample number | Mean (X) | Standard Deviation (SD) | Gender |
| #1 | 0.063 | 0.00082 | Female | #1 | 0.058 | 0.00163 | Female |
| #2 | 0.05367 | 0.00047 | Male | #2 | 0.05533 | 0.00125 | Male |
| #3 | 0.0553 | 0.00094 | Female | #3 | 0.058 | 0.00141 | Female |
| #4 | 0.0573 | 0.0017 | Female | #4 | 0.054 | 0.00141 | Female |
| #5 | 0.0547 | 0.0019 | Female | #5 | 0.0547 | 0.0012 | Female |
| #6 | 0.0647 | 0.00047 | Male | #6 | 0.0533 | 0.0009 | Female |
| #7 | 0.0567 | 0.0017 | Female | #7 | 0.0517 | 0.0005 | Male |
| #8 | 0.053 | 0.0014 | Female | #8 | 0.055 | 0.0008 | Female |
| #9 | 0.0533 | 0.0012 | Female | #9 | 0.0523 | 0.0017 | Female |
| #10 | 0.0553 | 0.0012 | Female | #10 | 0.0513 | 0.0012 | Female |
| #11 | 0.059 | 0.0014 | Male | #11 | 0.0490 | 0.0008 | Male |
| #12 | 0.0527 | 0.00094 | Male | #12 | 0.0473 | 0.0021 | Female |
| #13 | 0.0507 | 0.00047 | Male | #13 | 0.5057 | 0.0082 | Female |
| #14 | 0.054 | 0.0022 | Female | #14 | 0.0477 | 0.0009 | Male |
| #15 | 0.052 | 0.001 | Female | #15 | 0.049 | 0.0 | Male |
| #16 | 0.0553 | 0.0012 | Female | #16 | 0.0597 | 0.0005 | Male |
| #17 | 0.144 | 0.0041 | Male | #17 | 0.0537 | 0.0005 | Female |
| #18 | 0.0543 | 0.00047 | Male | #18 | 0.0517 | 0.0005 | Male |
| #19 | 0.06467 | 0.00047 | Male | #19 | 0.052 | 0.0024 | Female |
| #20 | 0.05767 | 0.00170 | Male | #20 | 0.0507 | 0.0012 | Male |
| #21 | 0.05567 | 0.00094 | Male | #21 | 0.0513 | 0.0012 | Female |
| #22 | 0.05233 | 0.00125 | Male | #22 | 0.0513 | 0.0005 | Male |
| #23 | 0.054 | 0.00141 | Male | #23 | 0.0507 | 0.0019 | Female |
| #24 | 0.053 | 0.00141 | Male | #24 | 0.0527 | 0.0012 | Female |
| #25 | 0.05833 | 0.00205 | Male | #25 | 0.0517 | 0.0009 | Male |
| #26 | 0.05367 | 0.00047 | Female | #26 | 0.0493 | 0.0012 | Male |
| #27 | 0.06433 | 0.00047 | Female | #27 | 0.0493 | 0.00047 | Male |
| #28 | 0.05233 | 0.00047 | Female | #28 | 0.0500 | 0.00141 | Female |
| #29 | 0.052 | 0.00141 | Male | #29 | 0.0520 | 0.00082 | Male |
| #30 | 0.052 | 0.001 | Male | #30 | 0.0495 | 0.0005 | Female |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).



