Submitted:
05 February 2025
Posted:
05 February 2025
You are already at the latest version
Abstract
Myocardial ischemia-reperfusion injury (MIRI) is a pathological process where the myocardium suffers further damage after blood flow is restored, posing a significant threat to patient health. Traditional treatment methods have many limitations, while innate lymphoid cells type 3 (ILC3s) and their glycolysis have shown important functions and great potential in MIRI. ILC3s, originating from common lymphoid progenitors, are widely distributed in various tissues and possess multiple functions such as anti-inflammation, immune balance regulation, promotion of cardiomyocyte regeneration, enhancement of cell survival, and activation of endogenous protective mechanisms, all of which rely on their glycolysis process. ILC3s glycolysis affects the pathological and physiological process of MIRI and its prognosis through mechanisms involving glucose uptake, metabolic pathways, metabolic sensing molecules, signal transduction pathways, cytokine secretion, metabolic product signaling, antioxidant enzyme and reactive oxygen species regulation, and apoptosis signaling pathways.
In terms of treatment, key enzymes, metabolic sensing molecules, and signaling pathways in the glycolysis pathway of ILC3s are expected to become new therapeutic targets. Activating ILC3s glycolysis can promote cardiomyocyte regeneration and repair, enhance cell survival, and improve clinical prognosis. Moreover, the regulation of ILC3s glycolysis can be combined with drug therapy, ischemic preconditioning, stem cell therapy, and other means to exert synergistic effects. By detecting the glycolysis characteristics and ILC3s functional status of patients, it is hoped to achieve precision and individualization in MIRI treatment.
However, current research still faces many challenges, including incomplete understanding of the specific mechanisms of ILC3s glycolysis, insufficient precision and effectiveness of regulatory means, and clinical transformation limited by patient individual differences and ethical considerations. In the future, it is necessary to strengthen the combination of basic research and clinical application, break through these barriers, fully explore the application value of ILC3s glycolysis in MIRI treatment, provide more effective treatment plans for patients, and improve survival quality and prognosis.
Keywords:
Myocardial Ischemia-Reperfusion Injury (MIRI)
; Innate Lymphoid Cells Type 3 (ILC3s)
; Glycolysis
; Cardiomyocyte Regeneration
; Therapeutic Targets
; Individualized Treatment
1. Introduction
MIRI refers to the further damage to myocardial cells and tissues when blood flow is restored after myocardial ischemia. This damage not only occurs during ischemia but also intensifies after blood flow is restored[1]. The pathological and physiological mechanisms of MIRI are closely related to oxidative stress[2], apoptosis[3], inflammatory response[4], metabolic disorders, and myocardial cell damage and repair,[5,6] which severely affect cardiac function and structure, leading to various cardiovascular diseases such as heart failure, arrhythmia, and cardiac arrest, posing a significant threat to patients' quality of life and life expectancy. Therefore, the treatment of MIRI is a major challenge in the medical field. The mainstream treatment plans include drug therapy (such as drugs to improve myocardial energy metabolism[7], antioxidants[8], calcium channel blockers[9]), ischemic preconditioning and postconditioning[10], and stem cell therapy (such as mesenchymal stem cells)[11]. However, these treatments have limitations such as unsatisfactory effects, large individual differences, high operational requirements, unresolved safety and stability issues, and immune rejection reactions. Despite the difficulties in research and treatment of MIRI, the medical community continues to tackle this problem. It has been found that ILC3s have anti-inflammatory[12], immune cell balance regulation[13], promotion of cardiomyocyte regeneration, enhancement of cardiomyocyte survival, and potential activation of endogenous protective mechanisms in MIRI[14]. The key to the function of ILC3s is the glycolysis process. Its effects on glucose uptake, metabolic pathways, metabolic sensing molecules, signal transduction pathways, cytokine secretion, metabolic product signaling, antioxidant enzyme regulation, reactive oxygen species generation and clearance, and apoptosis signaling pathways can influence the prognosis of MIRI. Therefore, the glycolysis of ILC3s is of great significance for the occurrence and development of MIRI and the formation of new treatment plans for MIRI.
2. ILC3s and ILC3s Glycolysis
2.1. Introduction to ILC3s
ILC3s originate from common lymphoid progenitors and are mainly enriched in the intestinal mucosa, liver, kidneys, and other areas[15]. ILC3s have unique molecular characteristics, including key transcription factors[16,17], cytokine secretion[18], receptors[19], and signaling pathways[20,21], as well as metabolic features. These characteristics enable ILC3s to play important roles in maintaining tissue homeostasis, regulating immune responses, and promoting tissue repair. ILC3s have the following functions:
Anti-inflammatory effects: During MIRI, the inflammatory response is a key factor leading to myocardial damage. ILC3s can regulate the inflammatory response by secreting anti-inflammatory cytokines. For example, ILC3s can secrete interleukin-22 (IL-22), and Jun Yan et al[22]. have proven that ILC3s and intestinal T cells secrete the cytokine IL-22 to promote epithelial integrity and protect against intestinal pathogens[23]. They can inhibit the production of pro-inflammatory cytokines such as tumor necrosis factor-alpha (TNF-α) and interleukin-1 beta (IL-1β)[24].Regulation of immune cell balance: ILC3s can interact with other immune cells to maintain immune cell balance. In the MIRI environment, they can regulate the functions of T cells and macrophages. For instance, ILC3s can inhibit over-activated macrophages from releasing inflammatory mediators while promoting the generation of macrophage subsets with repair functions, thereby reducing myocardial inflammatory damage and facilitating myocardial repair. Priscillia Bresler et al[25]. have proven that T cells regulate ILCs residing in lymph nodes in a tissue- and subset-specific manner.Promotion of cardiomyocyte regeneration: ILC3s can secrete various growth factors, such as vascular endothelial growth factor (VEGF)[26]. VEGF can stimulate myocardial angiogenesis, providing better blood supply for cardiomyocytes, which is conducive to the repair and regeneration of damaged cardiomyocytes. P.B.J Burton et al[27]. have proven that VEGF is an effective stimulant for myocardial ischemia in standard cardiopulmonary bypass coronary artery bypass grafting (CPB-CABG) and off-pump coronary artery bypass grafting (OP-CABG), which may affect postoperative graft endothelialization and cardiovascular hemodynamics.Enhancement of cardiomyocyte survival: ILC3s enhance the anti-apoptotic ability of cardiomyocytes through paracrine mechanisms by secreting cell-protective factors. In MIRI, cardiomyocytes are threatened by apoptosis due to various factors such as oxidative stress[28] and calcium overload[29]. The cell-protective factors secreted by ILC3s can help cardiomyocytes resist these harmful factors and increase the survival rate of cardiomyocytes.Activation of potential endogenous protective mechanisms: When MIRI occurs, the body attempts to initiate a series of protective programs to mitigate the damage. ILC3s may activate some endogenous protective signaling pathways, such as the PI3K-Akt pathway, through signal transmission with cardiomyocytes. The activation of these pathways can regulate the metabolism of cardiomyocytes and inhibit apoptosis, thereby exerting a protective effect in the early stage of MIRI[30,31](Figure a).
2.2. ILC3s Glycolysis is Key to ILC3s Function(Figure b)
2.2.1. Glucose Uptake and Metabolic Pathways
ILC3s glycolysis plays a crucial role in the normal functioning of ILC3s, including anti-inflammatory effects, immune cell balance regulation, cardiomyocyte regeneration promotion, enhancement of cardiomyocyte survival, and activation of potential endogenous protective mechanisms[32,33,34]. Specifically, ILC3s take up glucose through glucose transporter 1 (GLUT1) and metabolize it through pathways such as glycolysis and oxidative phosphorylation. After glucose enters the cell, it is metabolized through the glycolysis process to generate adenosine triphosphate (ATP), providing energy for the cell. In myocardial ischemia-reperfusion injury, the glycolysis activity of ILC3s is enhanced, which helps maintain the energy supply of cardiomyocytes. Metabolic products can provide energy and biosynthetic precursors for ILC3s, thereby maintaining their normal cell activity and function. ILC3s take up glucose through GLUT1, and after glucose enters the cell, the glycolysis process begins, including glucose phosphorylation, G-6-P isomerization, F-6-P phosphorylation, F-1,6-2P cleavage, conversion of phosphoglycerate, and conversion of phosphoglycerate to glyceraldehyde-3-phosphate (G-3-P) under the catalysis of phosphoglycerate mutase. This step is reversible, but the continuous consumption of G-3-P drives the reaction towards G-3-P, generating ATP five times further. Pyruvate is then metabolized; under anaerobic conditions, pyruvate is converted to lactate by lactate dehydrogenase, simultaneously regenerating nicotinamide adenine dinucleotide (NAD+), to maintain the continuous glycolysis. Under aerobic conditions, pyruvate enters the mitochondria and is converted to acetyl-CoA by pyruvate dehydrogenase, entering the tricarboxylic acid (TCA) cycle for further oxidative phosphorylation. The metabolic state of ILC3s can affect their signal transduction pathways through metabolic sensing molecules such as AMP-activated protein kinase (AMPK) and mammalian target of rapamycin (mTOR). The activation of AMPK can regulate the energy metabolism and cell activity of ILC3s, while mTOR regulates their proliferation and metabolism[35,36].
2.2.2. Cytokine Secretion and Metabolic Product Signaling
ILC3s glycolysis is associated with the secretion of cytokines such as interleukin-17 (IL-17) and IL-22 and the signaling of metabolic products[31,37]. For instance, in myocardial ischemia-reperfusion injury, IL-17 mainly promotes cardiomyocyte apoptosis, neutrophil infiltration, and inflammatory response, exacerbating myocardial damage[38]. In contrast, IL-22 exerts anti-inflammatory and cardioprotective effects, promoting cardiomyocyte regeneration and repair and reducing cardiomyocyte apoptosis. Chen Yang et al[39]. found that IL-22 can prevent angiotensin II-induced cardiomyocyte apoptosis by enhancing the activity of superoxide dismutase (SOD), blocking the decrease in mitochondrial membrane potential, inhibiting reactive oxygen species (ROS) production, and preventing the release of cytochrome C. ILC3s glycolysis can also participate in the transcriptional regulation or post-translational modification of IL-17 and IL-22 genes, thereby regulating their secretion levels[40]. Metabolic products of ILC3s (such as lactate and pyruvate) can bind to receptors on the surface of cardiomyocytes, such as monocarboxylate transporters (MCT), lactate receptors (GPR81), and pyruvate transporters (MPC), to regulate the metabolism and functional state of cardiomyocytes, thereby affecting the repair process of myocardial ischemia-reperfusion injury. Han She et al[41]. found that dexmedetomidine upregulates the phosphorylation of nuclear receptor subfamily 3 group C member 1 (NR3C1), downregulates pyruvate dehydrogenase kinase 4 (PDK4), and reduces lactate production and malate dehydrogenase 2 (MDH2) lactation. Ji Hae Jun et al[42]. found that the downstream protective mechanisms of pyruvate ethyl ester in myocardial ischemia-reperfusion injury include reducing ROS-mediated inflammasome activation, upregulation of NOD-like receptor 3 (NLRP3) inflammasome and related pathways, partly by inhibiting hypoxia-induced extracellular signal-regulated kinase (ERK) and p38 phosphorylation. The transcription factor Tox2 plays an important role in maintaining the gut microbiota, and the metabolism of ILC3s is crucial for it[43].
2.2.3. Synergistic Effects of Cardiomyocyte Protection and Immune Regulation
ILC3s glycolysis interacts with cardiomyocytes and other immune cells. For example, signal molecules produced by ILC3s glycolysis can affect the metabolic state, antioxidant capacity, and apoptosis process of cardiomyocytes, thereby exerting protective or damaging effects in myocardial ischemia-reperfusion injury[44,45]. The interaction between ILC3s and other immune cells such as T cells and macrophages can also be regulated by glycolysis. Metabolic products or metabolic sensing signals can affect the signal transmission and functional synergy between ILC3s and other immune cells, thereby regulating the immune and inflammatory responses in myocardial ischemia-reperfusion injury[46,47].
2.2.4. Antioxidant Enzyme and Reactive Oxygen Species Regulation
ILC3s glycolysis is involved in the regulation of antioxidant enzymes and the generation and clearance of reactive oxygen species. For example, ILC3s glycolysis may regulate the expression and activity of antioxidant enzymes (such as SOD, CAT) to modulate the oxidative stress levels in myocardial ischemia-reperfusion injury[48]. ILC3s glycolysis may produce a certain amount of ROS, and its metabolic products and metabolic sensing signals can also affect the clearance mechanisms of ROS. Blanda Di Luccia et al[33]. found that ILC3 activation depends on a metabolic program that combines glycolysis with ROS production. ROS has a double-edged sword effect in myocardial ischemia-reperfusion injury; moderate levels of ROS may promote cardiomyocyte protection and repair, while excessive ROS levels can lead to cardiomyocyte damage and death. Studies have shown that apoptosis and ROS are regulated by miR-182-5p through targeting STK17A to prevent myocardial ischemia-reperfusion injury[49,50].
2.2.5. Regulation of Apoptosis Signaling Pathways
ILC3s glycolysis is involved in the regulation of apoptosis signaling pathways and the control of apoptosis-related genes. For example, ILC3s glycolysis may regulate the expression and function of Bcl-2 family proteins by affecting apoptosis signaling pathways (such as the mitochondrial pathway, death receptor pathway) to regulate apoptosis in myocardial ischemia-reperfusion injury. ILC3s glycolysis can regulate apoptosis in myocardial ischemia-reperfusion injury by affecting the transcription and translation of apoptosis-related genes (such as Caspase family genes, Bcl-2 family genes)[51]. Huizi Liu et al[52]. found that in the miR-484 treatment group, the apoptosis index of cardiomyocytes in the ischemic area decreased, the membrane potential increased, and the expression of caspase-3/9 was significantly reduced.
Figure b.
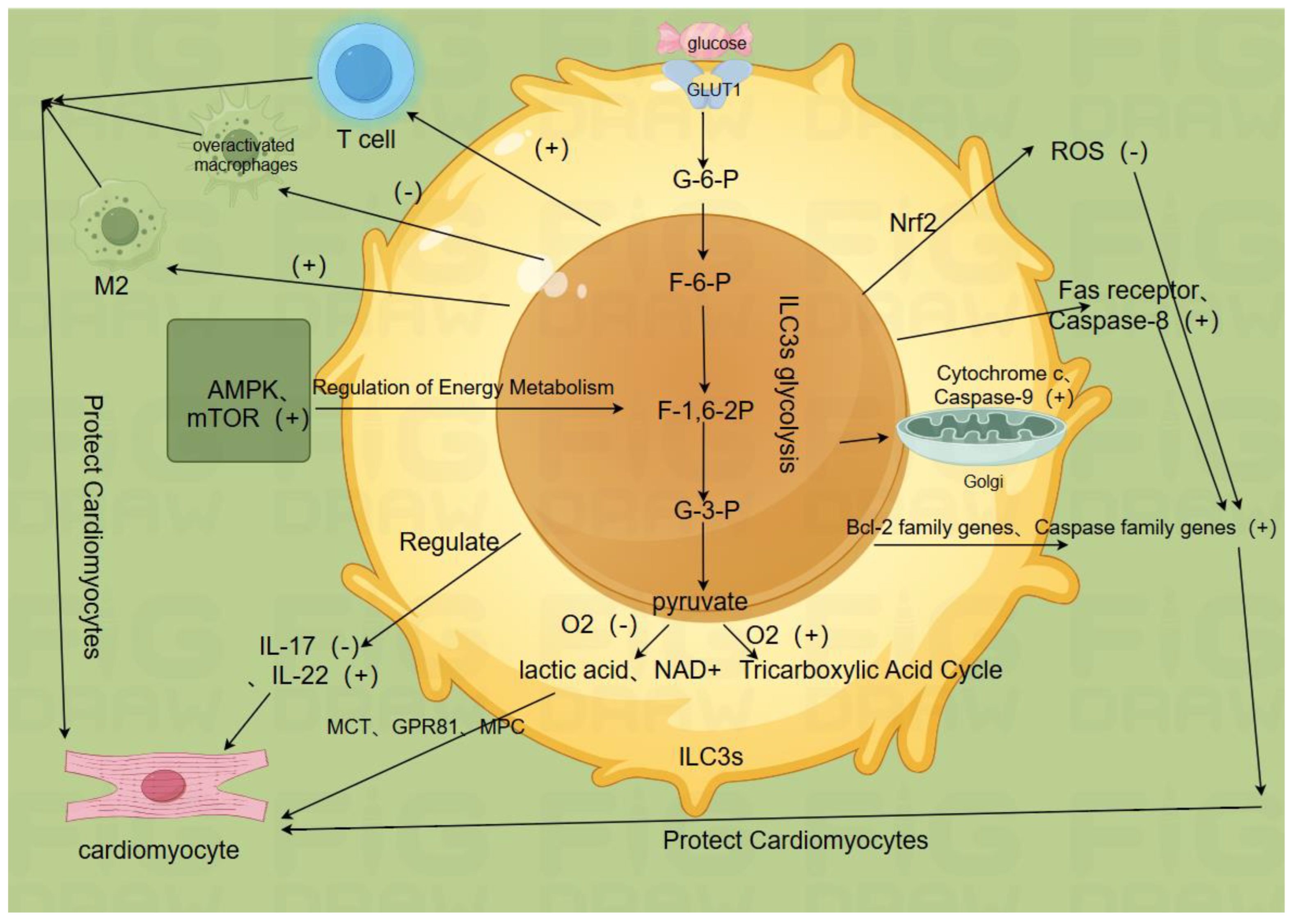
2.3. Regulation Mechanisms of ILC3s Glycolysis
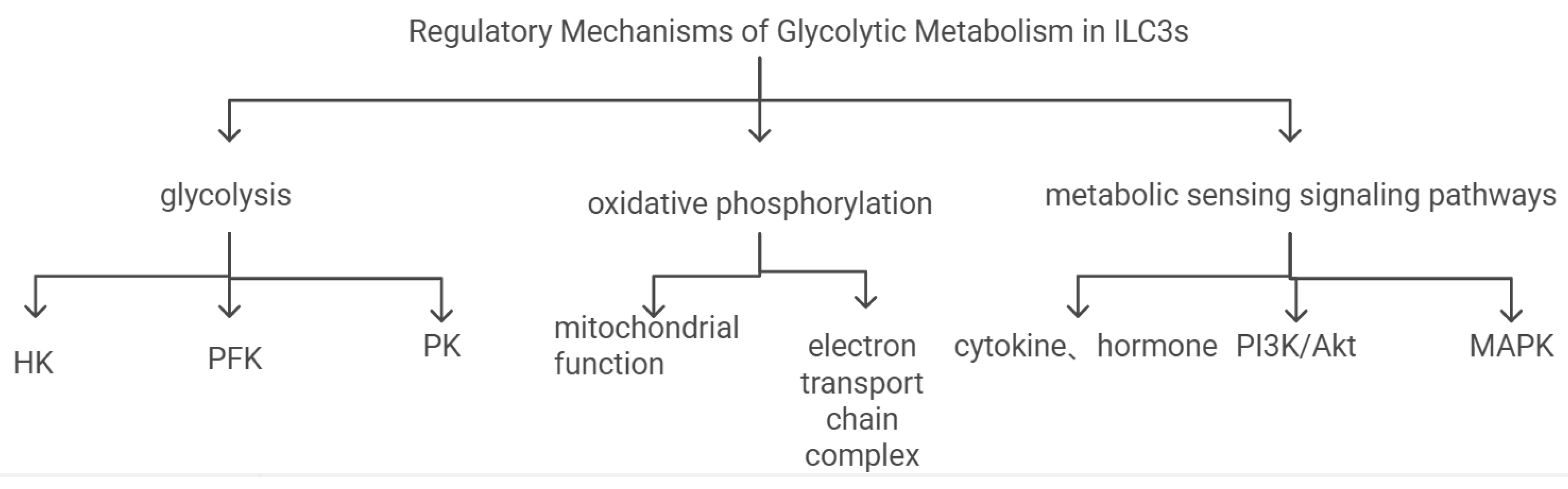
The rational regulation of ILC3s glycolysis is crucial for ensuring that the energy metabolism and function of ILC3s are in an appropriate state. It is regulated by the following aspects:1.Regulation of Glycolysis Pathway: It is regulated by hexokinase (HK)[53], phosphofructokinase (PFK)[54], and pyruvate kinase (PK)[55]. The role of metabolic sensors: Metabolic sensors such as AMPK and mTOR can affect the metabolic state of ILC3s by regulating the activity of key enzymes in the glycolysis pathway[56,57].2.Regulation of Oxidative Phosphorylation Pathway: Regulation of mitochondrial function[58]: The mitochondrial function of ILC3s can affect their energy supply by regulating the oxidative phosphorylation pathway. Jingzeng Cai et al[59]. found that di(2-ethylhexyl) phthalate (DEHP) can impair mitochondrial function and damage cardiomyocytes through the interleukin-6 (IL-6), Janus kinase (JAK) signal transduction, and signal transducer and activator of transcription 3 (STAT3) pathway. Changes in the morphology, quantity, and functional state of mitochondria can affect the metabolism and immune function of ILC3s[33,60]. Regulation of the electron transport chain: Changes in the activity of complexes in the electron transport chain can regulate the energy metabolism and oxidative stress levels of ILC3s, thereby affecting their role in myocardial ischemia-reperfusion injury[61].3.Regulation of Metabolic Sensing Signaling Pathways: Regulation by cytokines and hormones: Hormones such as insulin and glucagon can affect the function of ILC3s by regulating their glycolysis pathway[62]. These hormones can activate or inhibit specific signaling pathways, such as glucagon regulating transcription factors nuclear factor erythroid 2-related factor 2 (Nrf2) and Kelch-like ECH-associated protein 1 (Keap1), and insulin regulating the PI3K/AKT signaling pathway to regulate the metabolic state of ILC3s. Interaction of intracellular signaling pathways: The interaction of signaling pathways such as PI3K/Akt and MAPK can regulate the glycolysis and immune function of ILC3s, thereby affecting their regulatory role in myocardial ischemia-reperfusion injury[63,64,65] (Figure c).
3. Multiple Regulatory Mechanisms of ILC3s Glycolysis in Myocardial Ischemia-Reperfusion Injury
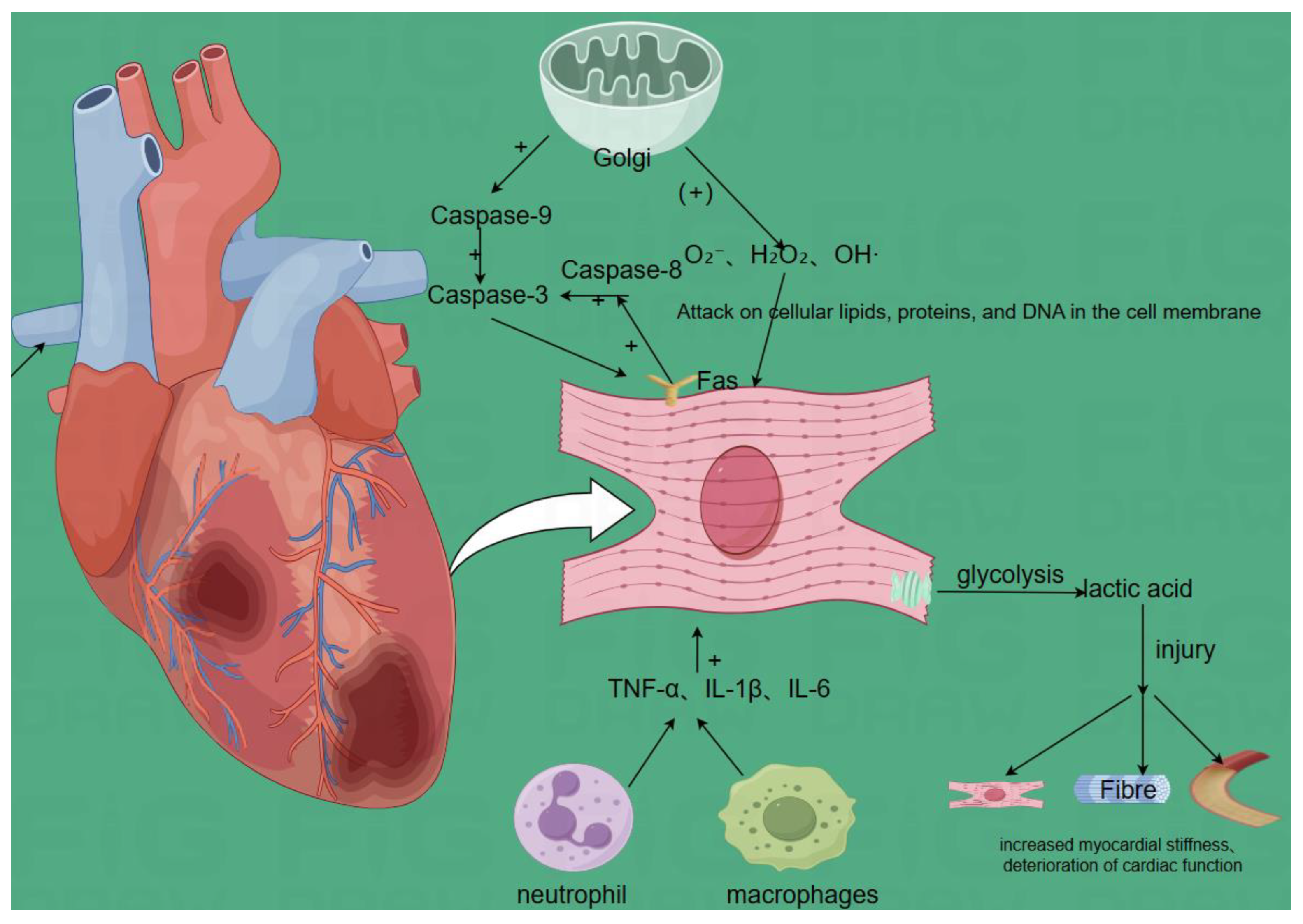
3.1. Pathophysiology of Myocardial Ischemia-Reperfusion Injury(Figure d)
When blood flow is restored after myocardial ischemia, further damage occurs to myocardial cells and tissues, mainly including the following changes:
Oxidative Stress: During reperfusion, the sudden increase in oxygen leads to the production of a large amount of reactive oxygen species (ROS) by mitochondria and other organelles, such as superoxide anions, hydrogen peroxide, and hydroxyl radicals[66,67]. Excessive ROS can attack the lipids, proteins, and DNA of cell membranes, leading to the destruction of cell structure and function, increasing cell membrane permeability, reducing enzyme activity, and causing DNA damage, ultimately leading to myocardial cell damage and death. Peiying Pai et al[66]. found that superoxide anion scavengers reduced hypoxia-induced Fas-related death domain, such as Fas ligand, Fas death receptor, Fas-dependent apoptotic pathway, such as activated caspase-8 and activated caspase-3, and mitochondrial-dependent apoptotic pathway, such as Bad, activated caspase-9 and activated caspase-3, endonuclease G (EndoG), apoptosis-inducing factor (AIF), and TUNING positive cell apoptosis, while enhancing cardiac survival pathways. Soochan Bae et al. found that reducing the expression levels of NADPH oxidase (NOX) 2 and 4 is beneficial for reducing hydrogen peroxide generation after MIRI and has protective significance for cardiomyocytes.
Apoptosis: Myocardial ischemia-reperfusion injury activates various apoptosis signaling pathways, such as the mitochondrial pathway and death receptor pathway. In the mitochondrial pathway, the mitochondrial membrane potential decreases, cytochrome c is released, and caspase-9 is activated; in the death receptor pathway, the binding of death receptors (such as Fas receptors) on the cell surface with ligands activates caspase-8. The activation of these signaling pathways leads to a cascade reaction of caspase family proteins, ultimately activating caspase-3 and leading to cardiomyocyte apoptosis[68,69].
Inflammatory Response: Myocardial ischemia-reperfusion injury induces the infiltration of inflammatory cells (such as neutrophils and macrophages). These inflammatory cells release a large amount of pro-inflammatory cytokines, such as TNF-α, IL-1β, and IL-6, which not only damage cardiomyocytes but also affect myocardial structure and function through processes such as promoting fibrosis and vascular remodeling[70,71].
Metabolic Disorders: During ischemia, the energy metabolism of cardiomyocytes mainly relies on glycolysis. After reperfusion, due to impaired mitochondrial function, the oxidative phosphorylation pathway cannot be restored in time, leading to insufficient energy supply of cardiomyocytes. Metabolic products (such as lactate) produced during ischemia cannot be cleared in time after reperfusion, further affecting the metabolism and function of cardiomyocytes[72].
Cardiomyocyte Damage and Repair: Myocardial ischemia-reperfusion injury leads to necrosis and apoptosis of cardiomyocytes, destruction of myocardial tissue structure, and decreased myocardial contractility. After injury, the body initiates a series of repair processes, including cardiomyocyte regeneration, fibrosis, and angiogenesis. However, due to the limited regenerative capacity of cardiomyocytes, the fibrosis process often dominates, leading to increased myocardial stiffness and further deterioration of cardiac function[73].
Figure d.
3.2. Effects of ILC3s Glycolysis on Myocardial Ischemia-Reperfusion Injury
3.2.1. Glucose Uptake and Metabolic Pathways
Glucose Uptake: ILC3s take up glucose through glucose transporter GLUT1. In myocardial ischemia-reperfusion injury, glucose uptake is crucial for maintaining the energy supply of cardiomyocytes. The increased expression of GLUT1 in cardiomyocytes can promote glucose uptake and glycolysis, thereby enhancing the regenerative capacity of cardiomyocytes and the cardiac repair response. During ischemia-reperfusion injury, the increased glucose uptake by ILC3s can provide more energy for cardiomyocytes, helping them resist damage and promote repair[74].Metabolic Pathways: Glycolysis is one of the main metabolic pathways of ILC3s glycolysis. In myocardial ischemia-reperfusion injury, the activation of the glycolysis pathway can rapidly produce ATP, providing immediate energy for cardiomyocytes. The activity regulation of key enzymes such as HK and PFK is crucial for the glycolysis process[75]. The enhancement of glycolysis can promote the proliferation and regeneration of cardiomyocytes. In addition to glycolysis, oxidative phosphorylation is also an important metabolic pathway for ILC3s glycolysis. During the reperfusion stage, the recovery of mitochondrial function and the activation of the oxidative phosphorylation pathway are important for maintaining the long-term energy supply and functional recovery of cardiomyocytes. However, the recovery of mitochondrial function takes time, so the rapid energy supply of glycolysis is particularly important in the early stage of reperfusion[76,77]
3.2.2. Metabolic Sensing Molecules and Signal Transduction
Metabolic Sensing Molecules: AMPK is activated when cellular energy is insufficient and regulates various metabolic pathways to maintain cellular energy homeostasis. There are certain differences in the functions and regulatory mechanisms of AMPK in different cell types, and these differences have important specific effects in the process of MIRI. AMPK is activated when energy is insufficient. The glycolysis state of ILC3s can be regulated through the activation of AMPK. In myocardial ischemia-reperfusion injury, the activation of AMPK can promote glucose uptake and glycolysis, enhancing the energy supply of cardiomyocytes. Additionally, AMPK can reduce ROS production by regulating the expression of antioxidant enzymes, thereby alleviating oxidative stress damage. The glycolysis state of ILC3s can affect through the regulation of the mTOR signaling pathway. In myocardial ischemia-reperfusion injury, the activity regulation of mTOR can affect the proliferation and autophagy processes of cardiomyocytes, thereby influencing the survival and functional recovery of cardiomyocytes[78,79].Signal Transduction Pathways: The glycolysis of ILC3s can affect the function of cardiomyocytes by regulating signaling pathways such as PI3K/Akt, JAK2/STAT3, and Nrf2/HO-1. Xiaqiong Wang et al[80]. found that the specific deletion of Gpr34 encoding lysophosphatidylserine (LysoPS) receptor in ILC3s or the inhibition of downstream PI3K-AKT or ERK inhibited IL-22 production in response to apoptotic neutrophils. Zhou Liang et al[81]. found that ILC3s increased the expression of programmed cell death 1 (PD-1) and subsequent interleukin-17A (IL-17A) production, directly activating myofibroblasts and fibrotic niche formation. In myocardial ischemia-reperfusion injury, the activation of the above signaling pathways can promote the survival of cardiomyocytes, reduce inflammatory responses, reduce cell apoptosis, and improve the energy supply of cardiomyocytes by regulating glycolysis pathways, thereby affecting the regeneration and repair processes of cardiomyocytes.
3.2.3. Cytokine Secretion and Metabolic Product Signaling
Cytokine Secretion: IL-17 can promote inflammatory responses and cardiomyocyte apoptosis, while IL-22 has anti-inflammatory and cardioprotective effects. The glycolysis state of ILC3s can affect the transcriptional regulation or post-translational modification of IL-17 and IL-22 genes to regulate their secretion levels. By regulating the secretion of these two cytokines, the glycolysis of ILC3s can regulate inflammatory responses and the protection and repair of cardiomyocytes. Other cytokines affected: The glycolysis of ILC3s may also affect the secretion of other cytokines, such as TNF-α, IL-6, etc. These cytokines also have important regulatory roles in myocardial ischemia-reperfusion injury, regulating inflammatory responses, cell proliferation, and apoptosis processes, thereby affecting the damage and repair of cardiomyocytes. Metabolic Product Signaling: Metabolic products of ILC3s glycolysis, such as lactate and pyruvate, can regulate the metabolism and functional state of cardiomyocytes by binding to receptors on the surface of cardiomyocytes. For example, lactate can act as a signaling molecule and regulate the energy metabolism and antioxidant capacity of cardiomyocytes by binding to monocarboxylate transporters (MCT) on the surface of cardiomyocytes, thereby affecting the repair process of myocardial ischemia-reperfusion injury. Additionally, during ischemia, the reverse activity of succinate dehydrogenase (SDH) leads to the accumulation of succinate. During reperfusion, succinate is rapidly oxidized, producing a large amount of ROS, causing oxidative stress damage to cardiomyocytes[82]. The glycolysis of ILC3s may regulate the metabolism and clearance of succinate, affecting the oxidative stress levels of cardiomyocytes, thereby exerting a protective role in myocardial ischemia-reperfusion injury. Moreover, metabolic products of ILC3s glycolysis can not only directly affect cardiomyocytes but also regulate immune and inflammatory responses by interacting with other immune cells (such as T cells, macrophages). For example, metabolic products can regulate the polarization state of macrophages, promoting their shift to an anti-inflammatory phenotype (M2 type), thereby exerting a protective role in myocardial ischemia-reperfusion injury[83].
3.2.4. Immune Regulation and Antioxidant Effects
Interaction with Other Immune Cells: Signal molecules produced by ILC3s glycolysis can affect the metabolic state, antioxidant capacity, and apoptosis process of cardiomyocytes. For example, metabolic products and cytokines secreted by ILC3s can regulate the mitochondrial function and energy metabolism of cardiomyocytes, enhance the antioxidant capacity of cardiomyocytes, reduce oxidative stress damage, and may also regulate the apoptosis process of cardiomyocytes by regulating apoptosis signaling pathways in cardiomyocytes, thereby affecting the repair of myocardial ischemia-reperfusion injury. Additionally, the interaction between ILC3s and other immune cells such as T cells and macrophages may also be regulated by glycolysis. For example, metabolic products and cytokines secreted by ILC3s can regulate the proliferation and differentiation of T cells, promote the generation of anti-inflammatory T cells (such as Treg cells), inhibit inflammatory responses[84]; at the same time, ILC3s can also regulate the polarization state of macrophages, promoting the shift of macrophages to an anti-inflammatory phenotype (M2 type), thereby exerting a protective role in myocardial ischemia-reperfusion injury[70].Regulation of Antioxidant Enzyme Expression and Activity: Metabolic products and metabolic sensing signals in the glycolysis process of ILC3s can activate the Nrf2 signaling pathway, thereby promoting the expression of antioxidant enzymes (such as superoxide dismutase SOD, glutathione peroxidase GPx, catalase CAT, etc.). By clearing reactive ROS, oxidative stress damage is alleviated, and cardiomyocytes are protected[85]. Antioxidant enzymes and cell protection: The enhanced activity of antioxidant enzymes can effectively clear ROS in cardiomyocytes, prevent oxidative damage of ROS to cell membrane lipids, proteins, and DNA, and maintain the integrity of cell structure and function. Generation of reactive oxygen species: ILC3s glycolysis can promote the generation of ROS by affecting mitochondrial function and the electron transport chain, etc. For example, during reperfusion, mitochondria produce a large amount of ROS due to the activation of the oxidative phosphorylation pathway, such as superoxide anions, hydrogen peroxide, and hydroxyl radicals. Clearance of reactive oxygen species: ILC3s glycolysis can enhance the ability of cardiomyocytes to clear ROS by regulating the expression and activity of antioxidant enzymes. For example, by activating the Nrf2 signaling pathway to promote the expression of antioxidant enzymes, the excessive ROS can be effectively cleared, preventing damage to cardiomyocytes.
3.2.5. Regulation of Apoptosis Signaling Pathways
ILC3s glycolysis can regulate the mitochondrial pathway of apoptosis signaling pathways by affecting mitochondrial function. For example, metabolic products and metabolic sensing signals can regulate mitochondrial membrane potential and the opening of mitochondrial permeability transition pores (PTP), thereby affecting the release of cytochrome c. After cytochrome c is released, it can activate caspase-9, triggering a caspase cascade reaction, leading to cardiomyocyte apoptosis. By regulating the mitochondrial pathway, ILC3s glycolysis plays a role in myocardial ischemia-reperfusion injury, affecting the survival and repair of cardiomyocytes. Regulation of the death receptor pathway: Glycolysis can also regulate the death receptor pathway of apoptosis signaling pathways by affecting the interaction between death receptors (such as Fas receptors) on the cell surface and their ligands. For example, the glycolysis state of ILC3s may affect the expression and activity of Fas ligands, thereby regulating the activation of Fas receptors and the activation of caspase-8, and further affecting the apoptosis process of cardiomyocytes. Regulation of apoptosis-related genes: ILC3s glycolysis can regulate apoptosis in cardiomyocytes by affecting the expression of Bcl-2 family genes and Caspase family genes. Metabolic products and metabolic sensing signals can regulate the transcription and translation of the above gene families' anti-apoptotic genes and pro-apoptotic genes, thereby affecting the expression and function of gene family proteins, and further regulating apoptosis in cardiomyocytes.
4. Potential Applications of ILC3s Glycolysis in MIRI Treatment
4.1. Key Enzymes, Metabolic Sensing Molecules, and Signal Pathways in ILC3s Glycolysis Pathway
4.1.1. Hexokinase, Phosphofructokinase, and Pyruvate Kinase
Key enzymes such as HK, PFK, and PK in the glycolysis pathway of ILC3s, metabolic sensing molecules (such as AMPK, mTOR), and signaling pathways (such as PI3K/Akt, JAK2/STAT3, Nrf2/HO-1) play important roles in MIRI and can be used as novel therapeutic targets in MIRI treatment. By regulating these targets, new therapeutic strategies can be developed to provide new ideas and methods for MIRI treatment. In ILC3s, the activity of HK and PFK directly affects the energy metabolism and function of cells. By regulating the activity of these enzymes, the metabolic state of ILC3s can be changed, thereby affecting their anti-inflammatory and protective functions in MIRI. Currently, inhibitors and activators of HK and PFK are being developed. For example, 2-deoxy-D-glucose (2-DG) is an HK inhibitor that inhibits glycolysis, reduces lactate production, and thereby reduces oxidative stress and inflammatory responses. Additionally, some small molecule compounds such as phosphofructokinase inhibitor (PFK-158) have been shown to activate PFK and enhance cellular energy metabolism. Currently, activators of PK such as pyruvate kinase activator (TEPP-46) are being studied. TEPP-46 activates PK, enhances cellular energy metabolism, and improves ischemia-reperfusion injury[86,87].
4.1.2. AMPK and mTOR
After activation, AMPK can promote glycolysis and fatty acid oxidation, enhancing cellular energy supply. The activity of mTOR is closely related to cell proliferation and metabolism. In ILC3s, the activity regulation of AMPK and mTOR can affect the metabolic state and function of cells, thereby exerting a protective role in MIRI. Currently, activators of AMPK such as AMP-activated protein kinase (AICAR) and Metformin have been widely studied[88,89]. AICAR activates AMPK, promotes cellular energy metabolism, reduces oxidative stress and inflammatory responses. Metformin activates AMPK, improves cellular metabolic state, and reduces myocardial ischemia-reperfusion injury. Additionally, mTOR inhibitors such as Rapamycin have been studied to regulate cell metabolism and function.
4.1.3. PI3K/Akt, JAK2/STAT3, and Nrf2/HO-1 Pathways
The PI3K/Akt signaling pathway plays an important role in cell survival and metabolic regulation, promoting cellular energy metabolism and anti-apoptotic functions by activating Akt. The JAK2/STAT3 signaling pathway plays a key role in cell proliferation and immune regulation. The Nrf2/HO-1 signaling pathway regulates the expression of antioxidant enzymes to reduce oxidative stress damage. Currently, activators of the PI3K/Akt pathway such as Insulin and insulin-like growth factor-1 (IGF-1) have been widely studied. Insulin activates the PI3K/Akt pathway, promotes cellular energy metabolism and survival. Additionally, JAK2 inhibitors such as Tofacitinib have been studied to regulate immune responses. Nrf2 activators such as Sulforaphane activate the Nrf2/HO-1 pathway, enhancing cellular antioxidant capacity.
4.2. Cardiomyocytes and Vasculature in ILC3s Glycolysis Pathway
4.2.1. Cardiomyocyte Proliferation
ILC3s glycolysis can promote the regeneration and repair of cardiomyocytes, enhancing the survival capacity of cardiomyocytes. In clinical treatment, activating the glycolysis pathway of ILC3s can provide more energy and biosynthetic precursors for cardiomyocytes, promote the proliferation and regeneration of cardiomyocytes, improve the prognosis after MIRI, reduce the area of myocardial infarction, and improve cardiac function. Cardiomyocyte proliferation can be detected by detecting the expression levels of proliferation markers such as nuclear protein (Ki-67) and proliferating cell nuclear antigen (PCNA) through immunohistochemistry or flow cytometry.
4.2.2. Angiogenesis
ILC3s glycolysis can promote angiogenesis by detecting microvascular density (MVD) or the expression levels of endothelial cell markers such as platelet endothelial cell adhesion molecule-1 (CD31) to assess the promoting effect of ILC3s-secreted VEGF and other factors on angiogenesis. VEGF is a key factor in promoting angiogenesis, which can stimulate myocardial angiogenesis, improve myocardial microcirculation, and thereby promote the regeneration and repair of cardiomyocytes. Recombinant VEGF and its analogs (such as VEGF-A, VEGF-C) have been widely studied for the treatment of myocardial ischemia-reperfusion injury[90]. Preclinical studies have shown that VEGF can significantly increase myocardial microvascular density and improve cardiac function.
4.3. Combined Applications of ILC3s Glycolysis in MIRI Treatment
4.3.1. Combination of ILC3s Glycolysis with Drug Therapy in MIRI Treatment
The regulation of ILC3s glycolysis can be used in combination with other treatments (such as drug therapy, ischemic preconditioning and postconditioning, stem cell therapy, etc.) to exert synergistic effects and improve treatment outcomes. Combination of ILC3s glycolysis with drug therapy in MIRI treatment: Drug therapy is usually used as a basic treatment method and can be performed before or simultaneously with the regulation of ILC3s glycolysis. For example, antioxidants such as N-acetylcysteine (NAC) [91]or anti-inflammatory drugs such as aspirin can be used before regulating ILC3s glycolysis to reduce initial inflammatory responses and oxidative stress, creating a more favorable environment for subsequent treatment. Antioxidants clear ROS to reduce oxidative stress damage. The regulation of ILC3s glycolysis activates the Nrf2/HO-1 signaling pathway to further enhance the expression of antioxidant enzymes, and the two work synergistically to more effectively protect cardiomyocytes. Aspirin inhibits the production of inflammatory mediators to reduce inflammatory responses. The regulation of ILC3s glycolysis secretes anti-inflammatory cytokines (such as IL-22) and inhibits pro-inflammatory cytokines (such as IL-1β) to further regulate immune responses.[39,92]
4.3.2. Combination of ILC3s Glycolysis with Ischemic Preconditioning and Postconditioning in MIRI Treatment
Combination of ILC3s glycolysis with ischemic preconditioning and postconditioning in MIRI treatment: Ischemic preconditioning (IPC) is usually performed before surgery or interventional treatment, while ischemic postconditioning (IPostC) is immediately implemented after blood flow is restored. The regulation of ILC3s glycolysis can be performed before or simultaneously with IPC or IPostC to enhance its protective effects. IPC activates endogenous protective mechanisms (such as the PI3K/Akt signaling pathway) to enhance the tolerance of cardiomyocytes. The regulation of ILC3s glycolysis activates similar signaling pathways to further enhance the energy metabolism and anti-apoptotic capacity of cardiomyocytes[93]. IPostC reduces oxidative stress and inflammatory responses during reperfusion to protect cardiomyocytes[94]. The regulation of ILC3s glycolysis secretes antioxidant enzymes and anti-inflammatory cytokines to further reduce oxidative stress and inflammatory damage.
4.3.3. Combination of ILC3s Glycolysis with Stem Cell Therapy in MIRI Treatment
Combination of ILC3s glycolysis with stem cell therapy in MIRI treatment: Stem cell therapy is usually performed as early as possible after MIRI to promote the regeneration and repair of cardiomyocytes. The regulation of ILC3s glycolysis can be performed before or simultaneously with stem cell transplantation to optimize the myocardial microenvironment and improve the survival rate and function of stem cells. Stem cells (such as mesenchymal stem cells) secrete growth factors (such as VEGF) to promote myocardial angiogenesis. The regulation of ILC3s glycolysis secretes similar factors to further enhance angiogenesis and cardiomyocyte repair capacity. The regulation of ILC3s glycolysis improves the energy metabolism of cardiomyocytes, providing a more favorable metabolic environment for stem cells, enhancing the survival rate and function of stem cells.
4.4. Clinical Detection and Individualized Adjustment Strategies for ILC3s Glycolysis in MIRI Treatment
4.4.1. Clinical Detection
The metabolic state of ILC3s glycolysis and ILC3s function may vary among different patients with MIRI. By evaluating the glycolysis characteristics and ILC3s status of patients, individualized treatment can be provided to develop more precise treatment plans. ILC3s function detection: Isolate ILC3s from peripheral blood or tissues, and detect surface markers such as CD45, CD127, CD3, CD117, etc. by flow cytometry. Cytokine secretion detection: Detect cytokines secreted by ILC3s such as IL-22, IL-17 using enzyme-linked immunosorbent assay (ELISA) or flow cytometry. Metabolic activity detection: Detect the glycolysis activity (such as glucose uptake, lactate production) and oxidative phosphorylation levels (such as ATP generation) of ILC3s to assess their metabolic state. In clinical practice, the functional state of ILC3s directly affects their protective role in MIRI. Abnormal ILC3s function may lead to exacerbated inflammatory responses or insufficient myocardial repair capacity. Gene detection and transcriptomics analysis: Analyze the gene expression profile of ILC3s by RNA sequencing (RNA-seq), focusing on genes related to glycolysis, inflammatory responses, and antioxidant functions. Changes in gene expression profiles in clinical practice may indicate metabolic reprogramming or functional dysfunction of ILC3s, providing a basis for targeted treatment.
4.4.2. Clinical Adjustment
Treatment plan adjustment strategies: High lactate levels: Indicates excessive glycolysis, which may worsen myocardial ischemic injury. Treatment strategies include using lactate clearers (such as lactate sodium) or drugs regulating the glycolysis pathway (such as 2-deoxy-D-glucose, 2-DG). Insulin resistance: Indicates metabolic disorders, which may affect the energy supply of ILC3s. Treatment strategies include using insulin sensitizers (such as metformin) or drugs regulating the AMPK signaling pathway. Insufficient IL-22 secretion: Indicates impaired anti-inflammatory and repair functions of ILC3s. Treatment strategies include using recombinant IL-22 protein or activating the glycolysis pathway of ILC3s (such as activating the PI3K/Akt signaling pathway). Excessive IL-17 secretion: Indicates excessive inflammatory response. Treatment strategies include using IL-17 inhibitors (such as secukinumab) or regulating the metabolic state of ILC3s to inhibit the secretion of pro-inflammatory cytokines. Low expression of Nrf2/HO-1 pathway: Indicates insufficient antioxidant capacity. Treatment strategies include using Nrf2 activators (such as sulforaphane) or regulating the expression of antioxidant enzymes. Abnormal PI3K/Akt pathway: Indicates impaired cell survival and metabolic regulation. Treatment strategies include using PI3K/Akt activators (such as insulin) or regulating metabolic sensing molecules (such as AMPK).
4.4.3. Treatment Plan Adjustment Strategies
Individualized adjustment of combined therapy: Drug combination: Adjust drug doses and combinations based on metabolic markers and ILC3s functional status. For example, for patients with metabolic disorders, combine the use of antioxidants and anti-inflammatory drugs. Ischemic preconditioning and postconditioning: For patients with normal ILC3s function, prioritize ischemic preconditioning or postconditioning to enhance the tolerance of cardiomyocytes. Stem cell therapy: For patients with impaired ILC3s function, combine stem cell therapy with metabolic regulation drugs to optimize the myocardial microenvironment.
4.4.4. Monitoring and Feedback
Treatment effect monitoring: Regularly detect metabolic markers and ILC3s functional status during treatment to assess treatment effects. For example, detect lactate levels and IL-22 secretion to determine whether glycolysis and anti-inflammatory functions have improved. Adverse reaction monitoring: Closely monitor possible adverse reactions in patients, such as drug side effects, immune responses, or metabolic disorders. Adjust the treatment plan if necessary. Individualized adjustment: Dynamically adjust the treatment plan according to the monitoring results to ensure the precision and effectiveness of the treatment.
5. Conclusion
MIRI poses a significant threat to patient health, and traditional treatment methods have many limitations. ILC3s and their glycolysis process have shown key roles and great potential in the field of MIRI. ILC3s, originating from common lymphoid progenitors and enriched in various tissues, have multiple functions such as anti-inflammation, immune balance regulation, promotion of cardiomyocyte regeneration, enhancement of cell survival, and activation of endogenous protective mechanisms, all of which rely on ILC3s glycolysis. Their glycolysis involves multiple aspects such as glucose uptake, various metabolic pathways, metabolic sensing molecule regulation, complex signal transduction, cytokine secretion regulation, metabolic product signaling, antioxidant enzyme and reactive oxygen species regulation, and apoptosis signaling pathway control, comprehensively affecting the pathological and physiological process of MIRI and thereby influencing the prognosis of MIRI. In terms of MIRI treatment, key enzymes, metabolic sensing molecules, and many signaling pathways in the glycolysis pathway of ILC3s are expected to become new therapeutic targets, opening the door to new treatment strategies; activating this glycolysis can help cardiomyocyte regeneration and repair, enhance survival capacity, and improve clinical prognosis; combining with other means such as drug therapy, ischemic preconditioning, and stem cell therapy can exert synergistic effects; at the same time, based on the individual differences in glycolysis characteristics and ILC3s functional status of patients, it is hoped to achieve precision and individualized treatment. However, current research still faces many challenges, including incomplete understanding of the specific mechanisms of ILC3s glycolysis, insufficient precision and effectiveness of regulatory means, and clinical transformation limited by patient individual differences and ethical considerations. In the future, it is necessary to strengthen the deep integration of basic and clinical research, overcome these barriers, fully explore the application value of ILC3s glycolysis in MIRI treatment, and open up more effective treatment paths for MIRI patients, improving patients' quality of life and prognosis.
Abbreviations
| Abbreviation | Full Form |
| AIF | apoptosis-inducing factor |
| AMPK | AMP-activated protein kinase |
| AICAR | 5-Aminoimidazole-4-carboxamide ribonucleotide |
| ATP | adenosine triphosphate |
| Bax | Bcl-2-associated X protein |
| CPB-CABG | Cardiopulmonary Bypass - Coronary Artery Bypass Grafting |
| Cyt c | cytochrome c |
| DEHP | di(2-ethylhexyl) phthalate |
| ELISA | enzyme-linked immunosorbent assay |
| EndoG | endonuclease G |
| ERK | Extracellular Signal-Regulated Kinase |
| FasL | Fas ligand |
| G-3-P | glyceraldehyde-3-phosphate |
| GPR81 | G Protein-Coupled Receptor 81 |
| GLUT1 | glucose transporter 1 |
| HK | hexokinase |
| ILC3s | Group 3 Innate Lymphoid Cells |
| IL-1β | interleukin-1 beta |
| IL-17 | interleukin-17 |
| IL-22 | interleukin-22 |
| IPostC | ischemic postconditioning |
| IPC | Ischemic preconditioning |
| IRE1 | inositol-requiring enzyme 1 |
| JAK | Janus kinase |
| Keap1 | Kelch-like ECH-associated protein 1 |
| MCT | monocarboxylate transporters |
| mTOR | mammalian target of rapamycin |
| MVD | microvascular density |
| MDH2 | malate dehydrogenase 2 |
| MPTP | mitochondrial permeability transition pore |
| NAC | N-acetylcysteine |
| NAD+ | nicotinamide adenine dinucleotide |
| NADPH | nicotinamide adenine dinucleotide phosphate |
| Nrf2 | nuclear factor erythroid 2-related factor 2 |
| NOX | NADPH oxidase |
| OP-CABG | Off-Pump Coronary Artery Bypass Grafting |
| PDK4 | pyruvate dehydrogenase kinase 4 |
| PD-1 | programmed cell death 1 |
| PFK | phosphofructokinase |
| PIP2 | phosphatidylinositol 4,5-bisphosphate |
| PK | pyruvate kinase |
| PTP | permeability transition pores |
| ROS | reactive oxygen species |
| RORγt | retinoic acid receptor-related orphan receptor gamma |
| SOD | superoxide dismutase |
| STAT3 | signal transducer and activator of transcription 3 |
| TNF-α | tumor necrosis factor-alpha |
| VEGF | vascular endothelial growth factor |
| Zbtb46 | zinc finger and BTB domain-containing protein 46 |
| 2-DG | 2-deoxy-D-glucose |
References
- Zhang, S.; Yan, F.; Luan, F.; Chai, Y.; Li, N.; Wang, Y.-W.; Chen, Z.-L.; Xu, D.-Q.; Tang, Y.-P. The pathological mechanisms and potential therapeutic drugs for myocardial ischemia reperfusion injury. Phytomedicine 2024, 129, 155649. [Google Scholar] [CrossRef] [PubMed]
- Xiang, M.; Lu, Y.; Xin, L.; Gao, J.; Shang, C.; Jiang, Z.; Lin, H.; Fang, X.; Qu, Y.; Wang, Y.; et al. Role of Oxidative Stress in Reperfusion following Myocardial Ischemia and Its Treatments. Oxid. Med. Cell. Longev. 2021, 2021, 6614009. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Q.; Yi, X.; Zhu, X.-H.; Wei, X.; Jiang, D.-S. Regulated cell death in myocardial ischemia-reperfusion injury. Trends Endocrinol. Metab. TEM 2024, 35, 219–234. [Google Scholar] [CrossRef]
- Hofmann, C.; Serafin, A.; Schwerdt, O.M.; Fischer, J.; Sicklinger, F.; Younesi, F.S.; Byrne, N.J.; Meyer, I.S.; Malovrh, E.; Sandmann, C.; et al. Transient Inhibition of Translation Improves Cardiac Function After Ischemia/Reperfusion by Attenuating the Inflammatory Response. Circulation 2024, 150, 1248–1267. [Google Scholar] [CrossRef]
- Xu, G.-E.; Yu, P.; Hu, Y.; Wan, W.; Shen, K.; Cui, X.; Wang, J.; Wang, T.; Cui, C.; Chatterjee, E.; et al. Exercise training decreases lactylation and prevents myocardial ischemia-reperfusion injury by inhibiting YTHDF2. Basic Res. Cardiol. 2024, 119, 651–671. [Google Scholar] [CrossRef] [PubMed]
- Gumpper-Fedus, K.; Park, K.H.; Ma, H.; Zhou, X.; Bian, Z.; Krishnamurthy, K.; Sermersheim, M.; Zhou, J.; Tan, T.; Li, L.; et al. MG53 preserves mitochondrial integrity of cardiomyocytes during ischemia reperfusion-induced oxidative stress. Redox Biol. 2022, 54, 102357. [Google Scholar] [CrossRef]
- Tan, M.; Yin, Y.; Chen, W.; Zhang, J.; Jin, Y.; Zhang, Y.; Zhang, L.; Jiang, T.; Jiang, B.; Li, H. Trimetazidine attenuates Ischemia/Reperfusion-Induced myocardial ferroptosis by modulating the Sirt3/Nrf2-GSH system and reducing Oxidative/Nitrative stress. Biochem. Pharmacol. 2024, 229, 116479. [Google Scholar] [CrossRef]
- Dhalla, N.S.; Elmoselhi, A.B.; Hata, T.; Makino, N. Status of myocardial antioxidants in ischemia-reperfusion injury. Cardiovasc. Res. 2000, 47, 446–456. [Google Scholar] [CrossRef]
- Jiang, L.; Zhou, X.; Zhao, X.; Wang, Z.; Huang, A.; Huang, Y.; Sun, H.; Guan, F.; Jiang, W. Tetrandrine downregulates TRPV2 expression to ameliorate myocardial ischemia/reperfusion injury in rats via regulation of cardiomyocyte apoptosis, calcium homeostasis and mitochondrial function. Eur. J. Pharmacol. 2024, 964, 176246. [Google Scholar] [CrossRef]
- Xiong, J.; Wang, Q.; Xue, F.-S.; Yuan, Y.-J.; Li, S.; Liu, J.-H.; Liao, X.; Zhang, Y.-M. Comparison of cardioprotective and anti-inflammatory effects of ischemia pre- and postconditioning in rats with myocardial ischemia–reperfusion injury. Inflamm. Res. 2011, 60, 547–554. [Google Scholar] [CrossRef]
- Al-Azab, M.; Idiiatullina, E.; Safi, M.; Hezam, K. Enhancers of mesenchymal stem cell stemness and therapeutic potency. Biomed. Pharmacother. 2023, 162, 114356. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Shi, W.; Sun, H.; Ji, Y.; Chen, Y.; Guo, X.; Sheng, H.; Shu, J.; Zhou, L.; Cai, T.; et al. Activation of DR3 signaling causes loss of ILC3s and exacerbates intestinal inflammation. Nat. Commun. 2019, 10, 3371. [Google Scholar] [CrossRef]
- Wang, W.; Li, N.; Guo, X. The crosstalk between ILC3s and adaptive immunity in diseases. FEBS J. 2024, 291, 3965–3977. [Google Scholar] [CrossRef]
- Jakob, M.O.; Spari, D.; Sànchez Taltavull, D.; Salm, L.; Yilmaz, B.; Doucet Ladevèze, R.; Mooser, C.; Pereyra, D.; Ouyang, Y.; Schmidt, T.; et al. ILC3s restrict the dissemination of intestinal bacteria to safeguard liver regeneration after surgery. Cell Rep. 2023, 42, 112269. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Zhou, L.; Zhou, J.; Chu, C.; Zhang, C.; Sockolow, R.E.; Eberl, G.; Sonnenberg, G.F. Zbtb46 defines and regulates ILC3s that protect the intestine. Nature 2022, 609, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Shen, X.; Liu, K.; Lu, C.; Fan, Y.; Xu, Q.; Meng, X.; Hong, S.; Huang, Z.; Liu, X.; et al. The Transcription Factor ThPOK Regulates ILC3 Lineage Homeostasis and Function During Intestinal Infection. Front. Immunol. 2022, 13. [Google Scholar] [CrossRef]
- The Transcription Factor RORα Preserves ILC3 Lineage Identity and Function during Chronic Intestinal Infection | The Journal of Immunology | American Association of Immunologists. Available online: https://journals.aai.org/jimmunol/article/203/12/3209/110026/The-Transcription-Factor-ROR-Preserves-ILC3 (accessed on 18 January 2025).
- CCR6+ group 3 innate lymphoid cells accumulate in inflamed joints in rheumatoid arthritis and produce Th17 cytokines | Arthritis Research & Therapy | Full Text. Available online: https://arthritis-research.biomedcentral.com/articles/10.1186/s13075-019-1984-x?utm_source=xmol&utm_medium=affiliate&utm_content=meta&utm_campaign=DDCN_1_GL01_metadata (accessed on 18 January 2025).
- Role of Nutrient-sensing Receptor GPRC6A in Regulating Colonic Group 3 Innate Lymphoid Cells and Inflamed Mucosal Healing | Journal of Crohn’s and Colitis | Oxford Academic. Available online: https://academic.oup.com/ecco-jcc/article/16/8/1293/6521543 (accessed on 18 January 2025).
- Vitamin D downregulates the IL-23 receptor pathway in human mucosal group 3 innate lymphoid cells - ScienceDirect. Available online: https://www.sciencedirect.com/science/article/abs/pii/S0091674917306577 (accessed on 18 January 2025).
- Activation of DR3 signaling causes loss of ILC3s and exacerbates intestinal inflammation | Nature Communications. Available online: https://www.nature.com/articles/s41467-019-11304-8?utm_source=xmol&utm_medium=affiliate&utm_content=meta&utm_campaign=DDCN_1_GL01_metadata (accessed on 18 January 2025).
- Musculin is highly enriched in Th17 and IL-22-producing ILC3s and restrains pro-inflammatory cytokines in murine colitis - Yan - 2021 - European Journal of Immunology - Wiley Online Library. Available online: https://onlinelibrary.wiley.com/doi/full/10.1002/eji.202048573 (accessed on 18 January 2025).
- Wang, B.; Lim, J.-H.; Kajikawa, T.; Li, X.; Vallance, B.A.; Moutsopoulos, N.M.; Chavakis, T.; Hajishengallis, G. Macrophage β2-Integrins Regulate IL-22 by ILC3s and Protect from Lethal Citrobacter rodentium-Induced Colitis. Cell Rep. 2019, 26, 1614–1626.e5. [Google Scholar] [CrossRef]
- P5374Genetic deletion of IL-22 increased cardiac rupture after myocardial infarction in mice | European Heart Journal | Oxford Academic. Available online: https://academic.oup.com/eurheartj/article-abstract/40/Supplement_1/ehz746.0337/5597811?redirectedFrom=fulltext&login=false (accessed on 18 January 2025).
- Bresler, P.; Tejerina, E.; Jacob, J.M.; Legrand, A.; Quellec, V.; Ezine, S.; Peduto, L.; Cherrier, M. T cells regulate lymph node-resident ILC populations in a tissue and subset-specific way. iScience 2021, 24, 102158. [Google Scholar] [CrossRef]
- Frontiers | Biology of Vascular Endothelial Growth Factor C in the Morphogenesis of Lymphatic Vessels. Available online: https://www.frontiersin.org/journals/bioengineering-and-biotechnology/articles/10.3389/fbioe.2018.00007/full (accessed on 18 January 2025).
- Burton, P.B.J.; Owen, V.J.; Hafizi, S.; Barton, P.J.R.; Carr-White, G.; Koh, T.; De Souza, A.; Yacoub, M.H.; Pepper, J.R. Vascular endothelial growth factor release following coronary artery bypass surgery: extracorporeal circulation versus ‘beating heart’ surgery. Eur. Heart J. 2000, 21, 1708–1713. [Google Scholar] [CrossRef]
- Oxidative stress in myocardial ischaemia reperfusion injury: a renewed focus on a long-standing area of heart research: Redox Report: Vol 10, No 4. Available online: https://www.tandfonline.com/doi/abs/10.1179/135100005X57391 (accessed on 18 January 2025).
- Chinese Medical Journal. Available online: https://journals.lww.com/cmj/fulltext/2023/10200/senp2_mediated_serca2a_desumoylation_increases.12.aspx (accessed on 18 January 2025).
- Xuan, X.; Zhou, J.; Tian, Z.; Lin, Y.; Song, J.; Ruan, Z.; Ni, B.; Zhao, H.; Yang, W. ILC3 cells promote the proliferation and invasion of pancreatic cancer cells through IL-22/AKT signaling. Clin. Transl. Oncol. 2020, 22, 563–575. [Google Scholar] [CrossRef]
- Chun, E.; Lavoie, S.; Fonseca-Pereira, D.; Bae, S.; Michaud, M.; Hoveyda, H.R.; Fraser, G.L.; Gallini Comeau, C.A.; Glickman, J.N.; Fuller, M.H.; et al. Metabolite-Sensing Receptor Ffar2 Regulates Colonic Group 3 Innate Lymphoid Cells and Gut Immunity. Immunity 2019, 51, 871–884.e6. [Google Scholar] [CrossRef]
- Hepworth, M.R. Proline fuels ILC3s to maintain gut health. Nat. Metab. 2023, 5, 1848–1849. [Google Scholar] [CrossRef] [PubMed]
- ILC3s integrate glycolysis and mitochondrial production of reactive oxygen species to fulfill activation demands | Journal of Experimental Medicine | Rockefeller University Press. Available online: https://rupress.org/jem/article/216/10/2231/120535/ILC3s-integrate-glycolysis-and-mitochondrial (accessed on 18 January 2025).
- Tan, C.Y.; Theriot, B.S.; Rao, M.V.; Surana, N.K. A commensal-derived exopolysaccharide regulates ILC3-mediated diseases. J. Immunol. 2023, 210, 227.04. [Google Scholar] [CrossRef]
- Montessuit, C.; Lerch, R. Regulation and dysregulation of glucose transport in cardiomyocytes. Biochim. Biophys. Acta BBA - Mol. Cell Res. 2013, 1833, 848–856. [Google Scholar] [CrossRef]
- AMPK: An Epigenetic Landscape Modulator. Available online: https://www.mdpi.com/1422-0067/19/10/3238 (accessed on 27 January 2025).
- Sajiir, H.; Ramm, G.A.; Macdonald, G.A.; McGuckin, M.A.; Prins, J.B.; Hasnain, S.Z. Harnessing IL-22 for metabolic health: promise and pitfalls. Trends Mol. Med. 2024. [Google Scholar] [CrossRef] [PubMed]
- Su, J.-L.; Jiao, Y.-R.; Li, T.; Yao, H.-C. Inhibition of IL-17 might be a novel therapeutic target in the treatment of myocardial ischemia reperfusion injury. Int. J. Cardiol. 2017, 239, 29. [Google Scholar] [CrossRef]
- Che, Y.; Tian, Y.; Chen, R.; Xia, L.; Liu, F.; Su, Z. IL-22 ameliorated cardiomyocyte apoptosis in cardiac ischemia/reperfusion injury by blocking mitochondrial membrane potential decrease, inhibiting ROS and cytochrome C. Biochim. Biophys. Acta Mol. Basis Dis. 2021, 1867, 166171. [Google Scholar] [CrossRef]
- RORγt inhibitors block both IL-17 and IL-22 conferring a potential advantage over anti-IL-17 alone to treat severe asthma | Respiratory Research | Full Text. Available online: https://respiratory-research.biomedcentral.com/articles/10.1186/s12931-021-01743-7?utm_source=xmol&utm_medium=affiliate&utm_content=meta&utm_campaign=DDCN_1_GL01_metadata (accessed on 18 January 2025).
- Dexmedetomidine Ameliorates Myocardial Ischemia-Reperfusion Injury by Inhibiting MDH2 Lactylation via Regulating Metabolic Reprogramming - She - 2024 - Advanced Science - Wiley Online Library. Available online: https://advanced.onlinelibrary.wiley.com/doi/full/10.1002/advs.202409499 (accessed on 18 January 2025).
- Jun, J.H.; Shim, J.-K.; Oh, J.E.; Shin, E.-J.; Shin, E.; Kwak, Y.-L. Protective Effect of Ethyl Pyruvate against Myocardial Ischemia Reperfusion Injury through Regulations of ROS-Related NLRP3 Inflammasome Activation. Oxid. Med. Cell. Longev. 2019, 2019, 4264580. [Google Scholar] [CrossRef]
- Das, A.; Martinez-Ruiz, G.U.; Bouladoux, N.; Stacy, A.; Moraly, J.; Vega-Sendino, M.; Zhao, Y.; Lavaert, M.; Ding, Y.; Morales-Sanchez, A.; et al. Transcription factor Tox2 is required for metabolic adaptation and tissue residency of ILC3 in the gut. Immunity 2024, 57, 1019–1036.e9. [Google Scholar] [CrossRef]
- Glucose metabolism and acute myocardial infarctionThe opinions expressed in this article are not necessarily those of the Editors of the European Heart Journal or of the European Society of Cardiology. | European Heart Journal | Oxford Academic. Available online: https://academic.oup.com/eurheartj/article-abstract/27/11/1264/456905?redirectedFrom=fulltext&login=false (accessed on 19 January 2025).
- Liu, X.; Zeng, J.; Yang, Y.; Qi, C.; Xiong, T.; Wu, G.; Zeng, C.; Wang, D. DRD4 Mitigates Myocardial Ischemia/Reperfusion Injury in Association With PI3K/AKT Mediated Glucose Metabolism. Front. Pharmacol. 2021, 11. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, S.; Jia, T.; Feng, Y.; Xu, X.; Zhang, D. T Cell Protein Tyrosine Phosphatase in Glucose Metabolism. Front. Cell Dev. Biol. 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- JCI Insight - Glucose metabolism controls disease-specific signatures of macrophage effector functions. Available online: https://insight.jci.org/articles/view/123047 (accessed on 19 January 2025).
- A New Paradigm in Catalase Research - ScienceDirect. Available online: https://www.sciencedirect.com/science/article/abs/pii/S096289242030252X (accessed on 19 January 2025).
- Targeted delivery and ROS-responsive release of Resolvin D1 by platelet chimeric liposome ameliorates myocardial ischemia–reperfusion injury | Journal of Nanobiotechnology | Full Text. Available online: https://jnanobiotechnology.biomedcentral.com/articles/10.1186/s12951-022-01652-x?utm_source=xmol&utm_medium=affiliate&utm_content=meta&utm_campaign=DDCN_1_GL01_metadata (accessed on 19 January 2025).
- Li, X.; Jin, Y. Inhibition of miR-182-5p attenuates ROS and protects against myocardial ischemia-reperfusion injury by targeting STK17A. Cell Cycle Georget. Tex 2022, 21, 1639–1650. [Google Scholar] [CrossRef]
- BCL2-regulated apoptotic process in myocardial ischemia-reperfusion injury (Review). Available online: https://www.spandidos-publications.com/10.3892/ijmm.2020.4781 (accessed on 19 January 2025).
- Liu, H.; Li, S.; Jiang, W.; Li, Y. MiR-484 Protects Rat Myocardial Cells from Ischemia-Reperfusion Injury by Inhibiting Caspase-3 and Caspase-9 during Apoptosis. Korean Circ. J. 2020, 50, 250–263. [Google Scholar] [CrossRef]
- Location matters: hexokinase 1 in glucose metabolism and inflammation - ScienceDirect. Available online: https://www.sciencedirect.com/science/article/abs/pii/S1043276022001400 (accessed on 20 January 2025).
- Fulghum, K.L.; Audam, T.N.; Lorkiewicz, P.K.; Zheng, Y.; Merchant, M.; Cummins, T.D.; Dean, W.L.; Cassel, T.A.; Fan, T.W.M.; Hill, B.G. In vivo deep network tracing reveals phosphofructokinase-mediated coordination of biosynthetic pathway activity in the myocardium. J. Mol. Cell. Cardiol. 2022, 162, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Jeon, J.-H.; Min, B.-K.; Ha, C.-M.; Thoudam, T.; Park, B.-Y.; Lee, I.-K. Role of the Pyruvate Dehydrogenase Complex in Metabolic Remodeling: Differential Pyruvate Dehydrogenase Complex Functions in Metabolism. Diabetes Metab. J. 2018, 42, 270–281. [Google Scholar] [CrossRef]
- Cai, X.; Li, H.; Wang, M.; Chu, E.; Wei, N.; Lin, J.; Hu, Y.; Dai, J.; Chen, A.; Zheng, H.; et al. mTOR participates in the formation, maintenance, and function of memory CD8+T cells regulated by glycometabolism. Biochem. Pharmacol. 2022, 204, 115197. [Google Scholar] [CrossRef]
- Ni, J.; Liu, Z.; Jiang, M.; Li, L.; Deng, J.; Wang, X.; Su, J.; Zhu, Y.; He, F.; Mao, J.; et al. Ginsenoside Rg3 ameliorates myocardial glucose metabolism and insulin resistance via activating the AMPK signaling pathway. J. Ginseng Res. 2022, 46, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Coupling of GABA Metabolism to Mitochondrial Glucose Phosphorylation | Neurochemical Research. Available online: https://link.springer.com/article/10.1007/s11064-021-03463-2?utm_source=xmol&utm_medium=affiliate&utm_content=meta&utm_campaign=DDCN_1_GL01_metadata (accessed on 20 January 2025).
- Cai, J.; Shi, G.; Zhang, Y.; Zheng, Y.; Yang, J.; Liu, Q.; Gong, Y.; Yu, D.; Zhang, Z. Taxifolin ameliorates DEHP-induced cardiomyocyte hypertrophy via attenuating mitochondrial dysfunction and glycometabolism disorder in chicken. Environ. Pollut. 2019, 255, 113155. [Google Scholar] [CrossRef]
- Wende, A.R.; Schell, J.C.; Ha, C.-M.; Pepin, M.E.; Khalimonchuk, O.; Schwertz, H.; Pereira, R.O.; Brahma, M.K.; Tuinei, J.; Contreras-Ferrat, A.; et al. Maintaining Myocardial Glucose Utilization in Diabetic Cardiomyopathy Accelerates Mitochondrial Dysfunction. Diabetes 2020, 69, 2094–2111. [Google Scholar] [CrossRef]
- Cogliati, S.; Cabrera-Alarcón, J.L.; Enriquez, J.A. Regulation and functional role of the electron transport chain supercomplexes. Biochem. Soc. Trans. 2021, 49, 2655–2668. [Google Scholar] [CrossRef]
- Oherle, K.; Acker, E.; Bonfield, M.; Wang, T.; Gray, J.; Lang, I.; Bridges, J.; Lewkowich, I.; Xu, Y.; Ahlfeld, S.; et al. Insulin-like Growth Factor 1 Supports a Pulmonary Niche that Promotes Type 3 Innate Lymphoid Cell Development in Newborn Lungs. Immunity 2020, 52, 275–294.e9. [Google Scholar] [CrossRef]
- Lennicke, C.; Cochemé, H.M. Redox regulation of the insulin signalling pathway胰岛素信号通路的氧化还原调节. Redox Biol. 2021, 42, 101964. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y.S.; Jun, H.-S. Effects of Glucagon-Like Peptide-1 on Oxidative Stress and Nrf2 Signaling. Int. J. Mol. Sci. 2018, 19, 26. [Google Scholar] [CrossRef]
- Desideri, E.; Vegliante, R.; Cardaci, S.; Nepravishta, R.; Paci, M.; Ciriolo, M.R. MAPK14/p38α-dependent modulation of glucose metabolism affects ROS levels and autophagy during starvation. Autophagy 2014, 10, 1652–1665. [Google Scholar] [CrossRef]
- Pai, P.; Lai, C.J.; Lin, C.-Y.; Liou, Y.-F.; Huang, C.-Y.; Lee, S.-D. Effect of superoxide anion scavenger on rat hearts with chronic intermittent hypoxia. J. Appl. Physiol. 2016, 120, 982–990. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.; Park, M.; Kang, C.; Dilmen, S.; Kang, T.H.; Kang, D.G.; Ke, Q.; Lee, S.U.; Lee, D.; Kang, P.M. Hydrogen Peroxide-Responsive Nanoparticle Reduces Myocardial Ischemia/Reperfusion Injury. J. Am. Heart Assoc. 2016, 5, e003697. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.G.; Fearnhead, H.O. Apocytochrome c blocks caspase-9 activation and Bax-induced apoptosis. J. Biol. Chem. 2002, 277, 50834–50841. [Google Scholar] [CrossRef]
- Xue, L.; Borné, Y.; Mattisson, I.Y.; Wigren, M.; Melander, O.; Ohro-Melander, M.; Bengtsson, E.; Fredrikson, G.N.; Nilsson, J.; Engström, G. FADD, Caspase-3, and Caspase-8 and Incidence of Coronary Events. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 983–989. [Google Scholar] [CrossRef]
- Zhuang, Q.; Li, M.; Hu, D.; Li, J. Recent advances in potential targets for myocardial ischemia reperfusion injury: Role of macrophages. Mol. Immunol. 2024, 169, 1–9. [Google Scholar] [CrossRef]
- role of neutrophils in myocardial ischemia–reperfusion injury | Cardiovascular Research | Oxford Academic. Available online: https://academic.oup.com/cardiovascres/article-abstract/43/4/860/340838?redirectedFrom=fulltext&login=false (accessed on 20 January 2025).
- Tian, H.; Zhao, X.; Zhang, Y.; Xia, Z. Abnormalities of glucose and lipid metabolism in myocardial ischemia-reperfusion injury. Biomed. Pharmacother. 2023, 163, 114827. [Google Scholar] [CrossRef]
- Mohr, M.E.; Li, S.; Trouten, A.M.; Stairley, R.A.; Roddy, P.L.; Liu, C.; Zhang, M.; Sucov, H.M.; Tao, G. Cardiomyocyte-fibroblast interaction regulates ferroptosis and fibrosis after myocardial injury. iScience 2024, 27. [Google Scholar] [CrossRef] [PubMed]
- Pharmacological inhibition of GLUT1 as a new immunotherapeutic approach after myocardial infarction - ScienceDirect. Available online: https://www.sciencedirect.com/science/article/abs/pii/S0006295221002033 (accessed on 21 January 2025).
- Chen, Y.; Bin, J.; Li, M.; Chen, Y.; Chen, G.; Zheng, H. ENHANCED GLYCOLYSIS PROMOTES POSTNATAL CARDIOMYOCYTE PROLIFERATION AND IMPROVES CARDIAC FUNCTION. J. Am. Coll. Cardiol. 2020, 75, 1042. [Google Scholar] [CrossRef]
- Zhao, R.; Yang, J.; Zhai, Y.; Zhang, H.; Zhou, Y.; Hong, L.; Yuan, D.; Xia, R.; Liu, Y.; Pan, J.; et al. Nucleophosmin 1 promotes mucosal immunity by supporting mitochondrial oxidative phosphorylation and ILC3 activity. Nat. Immunol. 2024, 25, 1565–1579. [Google Scholar] [CrossRef] [PubMed]
- Pedriali, G.; Ramaccini, D.; Bouhamida, E.; Wieckowski, M.R.; Giorgi, C.; Tremoli, E.; Pinton, P. Perspectives on mitochondrial relevance in cardiac ischemia/reperfusion injury. Front. Cell Dev. Biol. 2022, 10. [Google Scholar] [CrossRef]
- Relaxin activates AMPK-AKT signaling and increases glucose uptake by cultured cardiomyocytes | Endocrine. Available online: https://link.springer.com/article/10.1007/s12020-018-1534-3?utm_source=xmol&utm_medium=affiliate&utm_content=meta&utm_campaign=DDCN_1_GL01_metadata (accessed on 27 January 2025).
- mTOR signaling mediates ILC3-driven immunopathology | Mucosal Immunology. Available online: https://www.nature.com/articles/s41385-021-00432-4?utm_source=xmol&utm_medium=affiliate&utm_content=meta&utm_campaign=DDCN_1_GL01_metadata (accessed on 21 January 2025).
- Wang, X.; Cai, J.; Lin, B.; Ma, M.; Tao, Y.; Zhou, Y.; Bai, L.; Jiang, W.; Zhou, R. GPR34-mediated sensing of lysophosphatidylserine released by apoptotic neutrophils activates type 3 innate lymphoid cells to mediate tissue repair. Immunity 2021, 54, 1123–1136.e8. [Google Scholar] [CrossRef]
- Liang, Z.; Tang, Z.; Zhu, C.; Li, F.; Chen, S.; Han, X.; Zheng, R.; Hu, X.; Lin, R.; Pei, Q.; et al. Intestinal CXCR6+ ILC3s migrate to the kidney and exacerbate renal fibrosis via IL-23 receptor signaling enhanced by PD-1 expression. Immunity 2024, 57, 1306–1323.e8. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.-M.; Hu, Y.-Y.; Yang, T.; Wu, N.; Wang, X.-N. Reactive Oxygen Species and Oxidative Stress in Vascular-Related Diseases. Oxid. Med. Cell. Longev. 2022, 2022, 7906091. [Google Scholar] [CrossRef]
- Zhang, J.; Huang, F.; Chen, L.; Li, G.; Lei, W.; Zhao, J.; Liao, Y.; Li, Y.; Li, C.; Chen, M. Sodium Lactate Accelerates M2 Macrophage Polarization and Improves Cardiac Function after Myocardial Infarction in Mice. Cardiovasc. Ther. 2021, 2021, 5530541. [Google Scholar] [CrossRef]
- Saksida, T.; Paunović, V.; Koprivica, I.; Mićanović, D.; Jevtić, B.; Jonić, N.; Stojanović, I.; Pejnović, N. Development of Type 1 Diabetes in Mice Is Associated with a Decrease in IL-2-Producing ILC3 and FoxP3+ Treg in the Small Intestine. Molecules 2023, 28, 3366. [Google Scholar] [CrossRef]
- Tian, R.; Guan, X.; Qian, H.; Wang, L.; Shen, Z.; Fang, L.; Liu, Z. Restoration of NRF2 attenuates myocardial ischemia reperfusion injury through mediating microRNA-29a-3p/CCNT2 axis. BioFactors Oxf. Engl. 2021, 47, 414–426. [Google Scholar] [CrossRef]
- Terse, P.; Joshi, P.; Bordelon, N.; Brys, A.; Patton, K.; Arndt, T.; Sutula, T. 2-Deoxy–D-Glucose (2-DG) Induced Cardiac Toxicity in Rat: NT- proBNP and BNP as Potential Early Cardiac Safety Biomarkers. Int. J. Toxicol. 2016, 35, 284–293. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Li, C.; Zhu, Y.; Song, Y.; Lu, S.; Sun, H.; Hao, H.; Xu, X. TEPP-46-Based AIE Fluorescent Probe for Detection and Bioimaging of PKM2 in Living Cells. Anal. Chem. 2021, 93, 12682–12689. [Google Scholar] [CrossRef] [PubMed]
- Choksey, A.; Carter, R.D.; Thackray, B.D.; Ball, V.; Kennedy, B.W.C.; Ha, L.H.T.; Sharma, E.; Broxholme, J.; Castro-Guarda, M.; Murphy, M.P.; et al. AICAR confers prophylactic cardioprotection in doxorubicin-induced heart failure in rats. J. Mol. Cell. Cardiol. 2024, 191, 12–22. [Google Scholar] [CrossRef] [PubMed]
- The mystery of metformin.,Journal of Biological Chemistry - X-MOL. Available online: https://www.x-mol.com/paper/5659876?adv (accessed on 23 January 2025).
- VEGF-A in Cardiomyocytes and Heart Diseases. Available online: https://www.mdpi.com/1422-0067/21/15/5294 (accessed on 23 January 2025).
- The Clinical Efficacy of N-Acetylcysteine in the Treatment of ST Segment Elevation Myocardial Infarction. Available online: https://www.jstage.jst.go.jp/article/ihj/62/1/62_20-519/_article/-char/en (accessed on 23 January 2025).
- Talabieke, S.; Yang, J.; Wang, M. Abstract 424: Arachidonic Acid Synergizes With Aspirin In Cardioprotection Following Acute Myocardial Infarction. Arterioscler. Thromb. Vasc. Biol. 2023, 43, A424–A424. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, X. Ischaemic preconditioning-induced serum exosomes protect against myocardial ischaemia/reperfusion injury in rats by activating the PI3K/AKT signalling pathway. Cell Biochem. Funct. 2021, 39, 287–295. [Google Scholar] [CrossRef]
- Catalpol Promotes the Survival and VEGF Secretion of Bone Marrow-Derived Stem Cells and Their Role in Myocardial Repair After Myocardial Infarction in Rats | Cardiovascular Toxicology. Available online: https://link.springer.com/article/10.1007/s12012-018-9460-4?utm_source=xmol&utm_medium=affiliate&utm_content=meta&utm_campaign=DDCN_1_GL01_metadata (accessed on 24 January 2025).
Figure a.
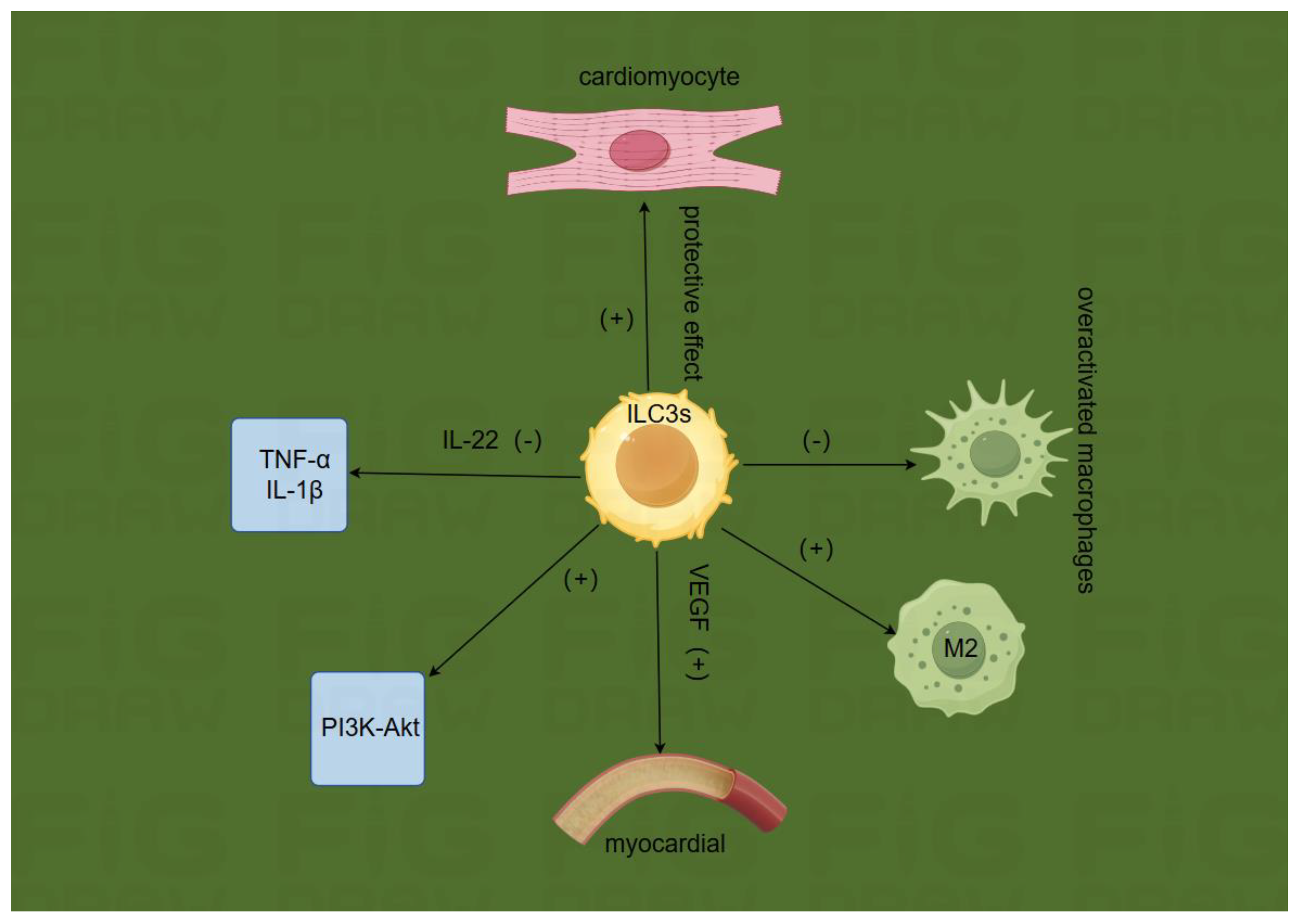
Figure c.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



