Submitted:
02 February 2025
Posted:
03 February 2025
You are already at the latest version
Abstract
Platelets, which have been traditionally associated with hemostasis and thrombosis functions, now receive attention for their role in immune responses that may affect vaccine development and effectiveness. Through their interactions with immune cells and modulation of inflammation alongside their role in antigen presentation, platelets become integral components of both innate and adaptive immune systems. New research shows platelets can improve vaccine effectiveness while reducing adverse side effects. During vaccine administration, platelets release cytokines and chemokines, which attract and stimulate immune cells to the injection site. Platelets work together with dendritic cells and T cells to support antigen processing and presentation, which leads to strong immune activation. Platelets' pro-inflammatory mediators strengthen local immune responses to boost protective immunity generation. Significant attention has been given to platelet involvement in vaccine-related thrombotic events, including vaccine-induced immune thrombotic thrombocytopenia (VITT). The rarity and severity of these events demonstrate the need to investigate the complex interplay between vaccine mechanisms and platelet activation. Exploration of the platelet-immune axis can lead to new methods for improving both the effectiveness and safety of vaccines. Researchers are working on creating innovative approaches for treatments that target platelet receptors and thrombosis pathways without interfering with the regular hemostatic functions of platelets. New vaccine development methods and personalized immunization strategies can emerge from targeting platelets with adjuvants and immune modulators.
Keywords:
Platelets
; Innate immunity
; adaptive immunity
; vaccine
; granules
; cytokines
; vaccine-induced immune thrombotic thrombocytopenia
1. Introduction
Platelets are small, non-nucleated, discoid-shaped blood components produced from megakaryocytes in the bone marrow along with erythrocytes (red blood cells), and most leukocytes (white blood cells) [1]. Megakaryocytes are large bone marrow cells, and they mature, and fragment to release over 1,000 platelets per megakaryocyte. The key hormone regulating megakaryocyte development is thrombopoietin (TPO). Platelets have a diameter of 2-3 μm and a 7-10 days lifespan in circulation [2]. Despite lacking a nucleus, they are equipped with a complex molecular machinery that enables their participation in immune responses [3]. Platelets possess a cytoskeleton composed of actin filaments and microtubules, which allows shape changes during activation [4]. Their plasma membrane is enriched with receptors such as glycoproteins (e.g., GPVI, GPIbα) and integrins (e.g., αIIbβ3) that facilitate interactions with extracellular matrix proteins and immune cells [5].
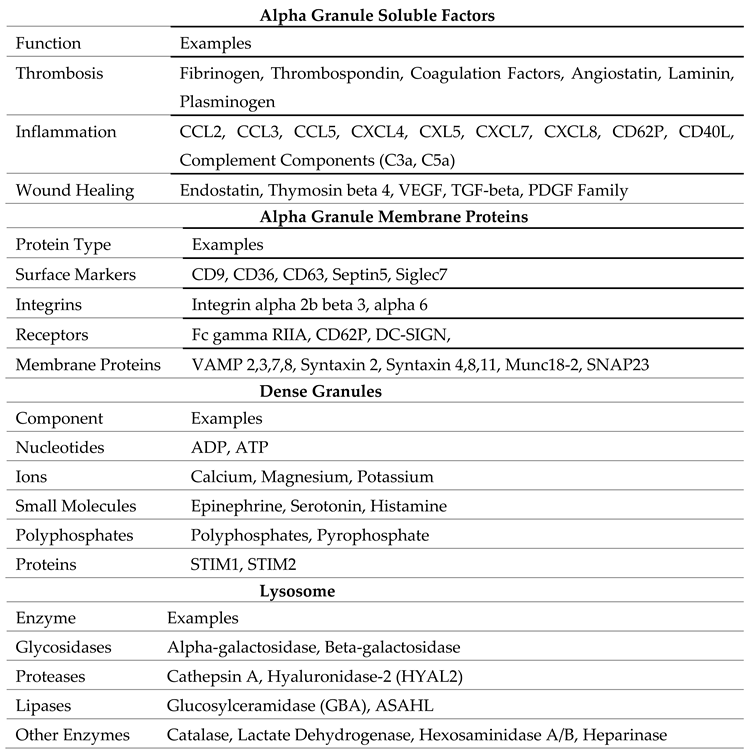
Platelets contain 3 major types of granules which store a wide array of bioactive molecules critical for all the immune functions performed by platelets. These granules are-
1.1. Alpha Granules
Alpha granules are the most abundant type of granules in platelets, numbering approximately 50–80 per platelet. In megakaryocytes (MKs), alpha granules (AGs) are formed through endosomal pathways involving multivesicular bodies (MVBs) [6]. AG biogenesis relies on newly synthesized cargos from the trans-Golgi network (TGN) and endocytosed proteins, with key roles played by VPS33B, VPS16B, and NBEAL2 [7]. They are large, membrane-bound structures that store a variety of proteins crucial for coagulation, inflammation, and tissue repair. Alpha granules contain a wide range of proteins that make up the majority of the platelet secretome [8].(watkins, J 2010) These proteins include adhesion molecules such as P-selectin, von Willebrand factor (vWF), and fibrinogen, critical for platelet aggregation and interaction with endothelial cells and leukocytes [9]. Growth factors such as vascular endothelial growth factor (VEGF), platelet-derived growth factor (PDGF), and transforming growth factor-beta (TGF-β), which promote tissue repair, angiogenesis, and immune modulation are also released from alpha granules upon activation. Cytokines and chemokines like interleukin-1β (IL-1β), CXCL4 (platelet factor 4), and RANTES (CCL5), are also present within the granules [10]. Some of these proteins are synthesized by megakaryocytes and packaged into granules, while others (e.g., fibrinogen and Factor V) are believed to be endocytosed by circulating platelets and subsequently transported to α granules. The release of alpha granule contents is a highly regulated process [11]. When platelets get activated upon agonists and receptors interactions, P-selectin is released and translocated to the surface, facilitating interactions with leukocytes and endothelial cells. This interaction promotes leukocyte recruitment and adhesion, that amplifies the inflammatory response [12]. The alpha granules contents also contribute to the repair of damaged tissues and the resolution of inflammation.
1.2. Dense Granules
Dense granules are smaller in size and less numerous than alpha granules, with an average of 3–8 granules per platelet [6]. DG formation involves cargo sorting by the AP-3 complex, stabilized by protein complexes, BLOC-1, with BLOC-2 and BLOC-3 suspected to function downstream. Rab38/Rab32 facilitate cargo trafficking to mature DGs, aided by BLOC-3, which serves as a guanine nucleotide exchange factor for these Rabs [13]. These granules contain low molecular weight compounds, including adenosine diphosphate (ADP) and adenosine triphosphate (ATP), which amplify platelet activation and recruit additional platelets to the site of injury, calcium ions (Ca2+) which is essential for platelet activation and coagulation cascade signaling, small molecules like serotonin, a vasoactive molecule that modulates vascular tone and permeability. These also contain polyphosphates, that enhance coagulation and support innate immune responses [11]. Dense granules are released rapidly upon platelet activation and likely contribute to the recruitment and activation of several immune cells such as neutrophils and monocytes [14]. Their contents also play a role in promoting vasodilation and increasing vascular permeability, facilitating the extravasation of immune cells to sites of injury, infection or inflammation [15].
1.3. Lysosomes
Lysosomes in platelets contain hydrolytic enzymes, including proteases, glycosidases, and lipases [9]. These enzymes are primarily involved in degrading extracellular matrix components and removing damaged or infected tissues [9]. While lysosomes are less studied than alpha and dense granules, their role in immune responses and tissue remodeling is increasingly recognized.
These granules release regulation is crucial for platelets function. Platelet secretion may need to be differential, especially when comparing the release of dense granule and α-granule cargo, but it remains unclear whether this is required for specific platelet functions at injury sites [16]. Further analysis is needed to understand whether platelet secretion is controlled by differences in release kinetics, differential solubility of cargo proteins, or other mechanisms, as platelets may have more complexities to reveal in their biology [17].
Italiano et al. proposed that platelets selectively store and release granular cargo based on specific physiological needs, such as promoting angiogenesis by releasing VEGF or inhibiting it by releasing angiostatin from distinct granule subsets [18]. This idea stems from two key observations: (1) pro- and anti-angiogenic factors were localized in separate compartments, and (2) Ma et al. demonstrated that different agonists (e.g., PAR1 and PAR4 peptides) triggered the selective release of specific cargo proteins. This hypothesis has gained significant attention, suggesting that platelets function as “smart” delivery systems capable of context-dependent cargo release [19,20]. While the hypothesis of differential granule exocytosis positions platelets as highly adaptable effector cells, questions remain about the extent of α granule diversity and whether the secretory machinery in platelets is sophisticated enough to enable this level of selective cargo release. Further research is required to fully understand these mechanisms and their physiological relevance.
2. Platelets Activation
Platelets, traditionally known as small cellular fragments circulating in blood and assisting in hemostasis and thrombosis, are being acknowledged as dynamic immune cells. These small, anucleate components of the blood are not merely bystanders in the immune system but active participants that influence innate and adaptive immunity [21]. Platelets endorsement as immune sentinels and mediators of immune surveillance and vascular remodeling has shifted our understanding of platelets, leading to profound implications for immunology. Platelets also have anti-inflammatory potential by interacting with regulatory T cells (Tregs), macrophages, and leukocytes and releasing pro-resolving mediators (ResolvinE1, Lipoxin, Maresin1) (Figure 1). Thus, the dual nature of platelets as both pro-inflammatory and anti-inflammatory entities underscores their complexity and adaptability in various physiological and pathological contexts [22].
Platelet activation triggers numerous essential processes for maintaining hemostasis, regulating immune responses, and influencing various physiological and pathological functions (Figure 1). These can be grouped into the following key areas:
Hemostasis and Thrombosis: Activated platelets adhere to vascular injury sites, change shape, release granule contents, and aggregate to form a hemostatic plug, preventing excessive bleeding and maintaining vascular integrity. Dysregulated activation can lead to thrombosis, causing vessel occlusion and events like heart attacks or strokes [23].
Inflammatory Responses: Platelets interact with leukocytes and endothelial cells, recruiting inflammatory cells to injury or infection sites. They release cytokines, chemokines, and growth factors that drive immune cell function, inflammation, tissue repair, and regeneration [24].
Angiogenesis: Platelets release factors like VEGF and PDGF that stimulate endothelial cell growth, migration, and new blood vessel formation, crucial for tissue repair and regeneration [25].
Wound Healing: Platelet-derived growth factors (e.g., TGF-β, FGF) promote cell proliferation, formation of extracellular matrix, and tissue remodeling, which are essential for healing and wound closure [26].
Immune Modulation: Platelets modulate immune responses by interacting with immune cells, influencing cytokine production, and presenting antigens to T cells. This contributes to immune activation and regulation [27].
Vascular Function: Platelets release vasoactive substances like serotonin and thromboxane A2, inducing vasoconstriction and aggregation. They also interact with endothelial cells to regulate vascular tone, permeability, and inflammation [28,29].
Overall, platelet activation effects are diverse and multifaceted, constituting their primary roles in hemostasis and inflammation (Figure 1). They also assist in angiogenesis and immune modulation. Understanding and addressing these functions is essential for unraveling the complex roles of platelets in health and different infectious and inflammatory situations, such as cardiovascular disease and autoimmune disorders, and developing targeted therapeutic interventions for platelet-related disorders.
3. Synergy of Platelets and Macrophages in Immune Regulation
Platelets and macrophages synergize to perform immunomodulatory functions; regulate innate immune response and inflammation (Figure 1). This interaction enhances the inflammatory response and facilitates pathogen clearance. Activated platelets release a variety of cytokines, chemokines, and growth factors, such as platelet factor 4 (PF4) and transforming growth factor-beta (TGF-β), which recruit and activate macrophages at sites of injury or infection [28,29,30,31,32]. Macrophages, in turn, respond by releasing pro-inflammatory cytokines like tumor necrosis factor-alpha (TNF-α) and interleukin-6 (IL-6), further amplifying the inflammatory cascade [21,33,34,35,36,37].
Moreover, platelets significantly influence macrophage polarization during inflammation, infection, and injury, guiding them toward either the pro-inflammatory M1 phenotype or the anti-inflammatory M2 phenotype based on specific environmental cues [38,39,40,41,42]. This dynamic interaction contributes to various physiological and pathological processes, including tissue repair, chronic inflammation, and thrombo-inflammatory disorders. For instance, platelet-derived epidermal growth factor receptor (EGFR) has been shown to induce M1 macrophage polarization by increasing the expression of inducible nitric oxide synthase (iNOS) and CD64, thereby enhancing pro-inflammatory responses and bacterial clearance [43,44]. Conversely, platelet-rich plasma (PRP), which is abundant in anti-inflammatory mediators and growth factors, has been observed to suppress M1 polarization and promote M2 polarization, facilitating tissue repair and reducing inflammation [45]. This dual capacity highlights the pivotal role of platelets in immune responses modulation across various physiological and pathological contexts. The complex interplay between platelets and macrophages, highlights the context-dependent effects of platelets on macrophage polarization and the subsequent implications for immune responses and tissue homeostasis.
However, dysregulated platelet-macrophage interactions can exacerbate conditions like atherosclerosis and autoimmune diseases, emphasizing the need for targeted therapies to modulate this synergy. Understanding the mechanisms underlying platelet-macrophage communication provides insights into novel treatment strategies for inflammation-driven diseases [21,28,32,35].
4. Platelets as Mediator of Innate Immunity
Platelets play an essential role in innate immunity by acting as sentinels that recognize and respond to pathogens (Figure 1). Neutrophils are key innate immune responders, and platelet-derived factors are crucial in their recruitment. P-selectin and the serotonin metabolite, 5-Hydroxy indoleacetic acid (5-HIAA) promote neutrophil migration via PSGL1 and GPR35, respectively. Platelet TLR4, p110β, and CLEC-2 signaling further regulate neutrophil infiltration in inflammation, tissue repair, and anti-tumor responses, underscoring the importance of platelets in neutrophil mobilization [19,20,46,47,48,49].
Platelets express all major classes of pattern recognition receptors (PRRs), including Toll-like receptors (TLRs), C-type lectin receptors (CLRs) such as DC-SIGN, and NOD-like receptors (NLRs). Platelets can be activated by bacterial pathogens through a TLR4-dependent mechanism, which triggers neutrophil activation and NETosis without inducing classical platelet aggregation [50]. However, TLR4 signaling in nucleated immune cells primarily relies on Myd88. Studies by using murine models of Klebsiella pneumoniae infection showed that platelet-specific Myd88 deficiency did not alter outcomes. This indicates that the role of platelet TLR4 signaling may vary depending on the context or operate via a Myd88-independent pathway [51]. NOD2, a NOD-like receptor expressed by platelets, recognizes muramyl dipeptide (MDP), a bacterial peptidoglycan motif. It mediates effects via Rip2, contributing to platelet responses during sepsis and promoting thrombus formation in arterial thrombosis models [52]. Platelet TLR4 signaling activates platelets, promoting the release of IL-1β-rich microparticles (MPs) and facilitating interactions with other cells. These interactions, particularly with adherent neutrophils, induce the formation of neutrophil extracellular traps (NETs), which can capture bacteria [50,53].
Platelets are a major source of IL-1β, which is not stored in granules but produced upon stimulation. Surprisingly, platelets express pre-mRNA for IL-1β, which is spliced and translated into pro-IL-1β, then processed by caspase-1 to release functional IL-1β over several hours. In a mouse model of severe malaria, platelet-derived IL-1β plays a critical role in initiating the acute phase response, with platelets localized to hepatic sinusoids post-Plasmodium infection, suggesting a contact-dependent mechanism [54,55].
Platelets also express the FcγRIIIa receptor, which has low affinity for monomeric IgG but can bind to immune complexes and IgG-coated pathogens and to autoantibodies that target a subset of platelet membrane proteins [56,57]. This applies to interactions with Influenza (H1N1), Streptococci, Bacillus anthracis. FcγRIIA plays a key role in the pathology of heparin-induced thrombocytopenia (HIT) by recognizing anti-PF4/heparin immune complexes and, in cooperation with GPIIbIIIa receptor, activating platelets through a Src/Syk-dependent pathway, leading to thrombosis [58,59,60,61]. FcγRIIa has been shown to interact with αIIbβ3, the primary integrin on platelets, this interaction helps transmit “outside-in” signals across the membrane when the activated integrin binds to ligands like fibrinogen or von Willebrand factor (vWR). These signals are essential for full platelet activation, including degranulation and clot retraction [56]. Another receptor expressed by platelets- C-type lectin receptor 2 (CLEC-2), which recognizes podoplanin, a transmembrane receptor glycoprotein, plays a critical role in platelet-mediated blood-lymphatic separation during development. It also modulates inflammation in experimental peritoneal sepsis, most likely through interactions with macrophage-expressed podoplanin, promoting an anti-inflammatory macrophage phenotype. Contrarywise, during systemic Salmonella typhimurium infection, hepatic podoplanin expression increases in the subacute phase, activating CLEC-2 and driving infection-associated thrombus formation. These findings highlight the dual roles of CLEC-2 in mediating both inflammatory and anti-inflammatory effects depending on the timing and nature of the stimulus [62,63].
Platelets, like any nucleated cells, utilize innate immune pathways to support host defense mechanisms. The stimulator of interferon genes (STING) regulates platelet activity through STXBP2, facilitating granule release via SNARE complex formation. Platelet-specific STING deficiency reduces NET formation and intravascular thrombosis in experimental sepsis, with blocking STING-STXBP2 interactions yielding similar protective effects [64,65]. Additionally, platelets express the NLRP3 inflammasome, which activates Caspase-1 and processes pro-IL-1β into IL-1β, playing a role in platelet-driven inflammatory responses. Platelets can also mediate type I interferon (IFN) production via the cGAS-STING pathway, activated by cytosolic DNA, amplifying antiviral and pro-inflammatory immune responses [43,66].
Neutrophils defend against pathogens through NETosis, expelling nuclear and mitochondrial contents to form antimicrobial extracellular traps (NETs). Although platelets lack a nucleus, they release mitochondria upon activation, either as free organelles or in microparticles. These mitochondria can trigger inflammation through proinflammatory lipid mediators and mitochondrial DNA (mtDNA), which also stimulates thrombin generation and neutrophil activation. In systemic lupus erythematosus (SLE), platelets contribute to extracellular mtDNA accumulation and antibody recognition via FcγRIIa, potentially driving immunopathology. Platelet microparticles (PMPs) further modulate inflammation and access sites like synovial and lymph fluid, but their role in host defense remains unclear due to the absence of PMP-deficient models [67,68,69,70,71,72].
5. Platelets as Adaptive Immune Regulation
Host defense involves a multi-layered process, where innate immune cells take on all intruders providing immediate protection, followed by T and B lymphocytes coming into play ensuring specific and long-lasting responses or adaptive immunity [73]. While adaptive immunity takes days to develop, it is essential for long-term specific protection. Platelets play a significant role in adaptive immunity (Figure 1), influencing antigen trafficking and presentation by dendritic cells (DCs) and modulating T and B cell signaling, maturation, and polarization [74]. Platelets affect adaptive immune cells by activating monocytes, macrophages, T and B cells, DCs, and NK cells [75].
Lymph nodes are crucial for antigen presentation and lymphocyte trafficking [76], with CLEC-2 expressed by platelets playing a key role in their development and maintenance. Platelet-specific CLEC-2 deletion leads to blood-filled lymph nodes, fibrosis, and impaired antibody production [77]. Beyond supporting lymph node function for effective adaptive immunity, platelets also directly influence antigen-presenting dendritic cells (DCs) [75]. They promote DC differentiation via the P-selectin-PSGL-1 axis and enhance type I interferon production through CD40L [78]. CD40L-driven DC activation and interferon production contribute to autoimmunity in lupus [79]., where innate immune activation and coagulation crosstalk are amplified by endothelial protein C receptor (EPCR) and tissue factor signaling.
CLEC-2, expressed by platelets, plays a key role in the lymph nodes’ development and maintenance, the deletion of which leads to blood-filled lymph nodes, fibrosis, and impaired antibody production [62,80]. Platelets also promote dendritic cell differentiation via the P-selectin-PSGL-1 axis and enhance type I interferon production through CD40L [78]. Platelets exert diverse effects on lymphocyte subpopulations. They can suppress cytokine secretion and immunosuppressive responses of T helper cells while enhancing cytotoxic T cell proliferation and activity. Activated platelets can promote B cell isotype switching and antibody production but reduce the cytolytic activity of natural killer (NK) cells [81].
6. Platelets in Vaccine-Induced Immune Thrombotic Thrombocytopenia
Platelets play a critical role as effector cells in adverse immune responses to vaccines, with a notable example being vaccine-induced immune thrombotic thrombocytopenia (VITT) [82]. For most individuals, administering an adenovirus-based COVID-19 vaccine effectively triggers the production of antibodies against the SARS-CoV-2 spike protein, thereby providing strong protection against COVID-19 [83]. However, in rare cases, the vaccine can induce VITT. Platelet-activating antibodies targeting platelet factor 4 (PF4), a platelet-associated chemokine, triggered this condition. These antibodies form immune complexes with PF4, which are then recognized by platelet Fc gamma IIa receptors, leading to pathological platelet activation, initiating a cascade of coagulating events, leading to blood clots or thrombosis and thrombocytopenia or reduction in platelet count [84]. While VITT represents a severe pathological reaction, platelets may also exert intended immune response upon SARS-CoV-2 vaccination [85].; probably programmed cell death pathways activation to control viral replication. This raises an intriguing question about how platelets respond to the vaccine and potentially contribute to effective immunization, that underscores the importance of understanding immune responses to vaccination.
In the review by Hirsch et al., they effectively explore the interaction between platelets and neutrophils, with a particular focus on COVID-19 and VITT [86]. They highlight the key roles of hyperactivated platelets and neutrophil extracellular traps (NETs) in the coagulopathy seen in COVID-19 and discuss the development of a new thrombophilia paradigm triggered by auto-antibody formation following adenoviral vector vaccinations [86]. In another study by Ostrowski et al., a direct comparison between the Oxford/AstraZeneca [ChAdOx1] (AZ) and mRNA vaccines was done that revealed enhanced inflammation, platelet activation, and thrombin generation following AZ vaccination, further supporting potential triggers and mechanisms underlying complications like VITT [87].
Rapid treatment is essential for VITT patients, as delays can be life-threatening. Since FcγRIIa mediates VITT, immune-suppressive therapies are effective. American Society of Hematology, recommends Intravenous immunoglobulin (IVIG) as the first-line treatment at 1 g/kg daily for two days, often supplemented with corticosteroids like dexamethasone. Studies have shown that IVIG significantly reduces mortality in VITT patients, validating its use as a key treatment strategy [88,89,90]. Targeting fibrinolysis may offer a promising treatment for VITT. Enhancing fibrinolysis has shown effectiveness in a mouse model of HIT [91].
7. Therapeutic Implications of Platelets
Beyond their usual physiological and pathological functions, platelets offer significant therapeutic potential in many clinical applications. One of the primary therapeutic uses of platelets is in transfusion medicine [92]. Platelet transfusions are essential for patients with thrombocytopenia or platelet dysfunction to prevent or control bleeding. In oncology, platelet transfusions are critical for managing bleeding risks in patients undergoing chemotherapy or bone marrow transplantation [93]. Platelets also hold promise in regenerative medicine and wound healing. Platelet-rich plasma (PRP), derived from concentrated platelets, is widely used to promote tissue repair and regeneration [94]. Growth factors such as platelet-derived growth factor (PDGF) and vascular endothelial growth factor (VEGF) in PRP stimulate cell proliferation, angiogenesis, and extracellular matrix formation [94], making it valuable for treating sports injuries, chronic wounds, and osteoarthritis.
Antiplatelet drugs are the primary treatment for cardiovascular disease and atherothrombosis prevention [94]. Current therapies have limitations, with few antiplatelet drugs available and limited options for managing platelet-related bleeding. Treatments are mostly restricted to increasing platelet count (e.g., thrombopoietin) or platelet transfusions for conditions like thrombocytopenia [95]. Aspirin (acetylsalicylic acid) is the most commonly prescribed drug for cardiovascular disease, working by irreversibly inhibiting cyclooxygenase-1 and blocking thromboxane A2 formation [96]. Despite an associated bleeding risk, low-dose aspirin is recommended for primary prevention in individuals at high cardiovascular risk. For secondary prevention in acute coronary syndrome or post-coronary stent implantation, dual antiplatelet therapy (DAPT), combining aspirin with P2Y12 receptor blockers, is the standard [97]. Irreversible P2Y12 inhibitors include clopidogrel and prasugrel, while ticagrelor and cangrelor provide faster, reversible effects. For symptomatic peripheral artery disease, single antiplatelet therapy with aspirin or clopidogrel is preferred due to a better benefit-risk ratio compared to DAPT, although the risk of recurrent thrombosis remains significant in these patients [98,99,100,101].
The relevance of platelet immunology extends beyond basic science to the realm of drug development and therapeutics. Platelets represent both a target and a tool in the quest to modulate immune responses. Antiplatelet therapies, traditionally used to prevent thrombosis, are being repurposed to mitigate inflammation in autoimmune and cardiovascular diseases. Drugs such as aspirin and P2Y12 inhibitors, which block TXA2 formation and inhibits ADP agonist receptors respectively [102]. (Table 2) causing inhibition of platelet activation, have shown promise in reducing immune-mediated vascular damage and improving outcomes in inflammatory diseases. In addition, platelet-derived biomarkers such as proteins and RNA present inside the platelets are emerging as valuable tools for cancer diagnosis, and monitoring and confronting immune-related inflammatory conditions [103]. Platelet microparticles, cytokines, and extracellular vesicles provide insights into disease activity and therapeutic efficacy, paving the way for personalized medicine.
Recent advancement in platelet biology highlights their potential to secrete several pro- and anti-inflammatory mediators that modulate immune functions and which could be further utilized for designing and developing therapies for autoimmune diseases and inflammatory conditions. Additionally, emerging evidence suggests targeting platelet activation pathways for treating thrombotic disorders, such as heart attacks and strokes [104,105,106,107]. Overall, platelets represent a versatile tool in therapeutic interventions, with ongoing research expanding their applications in both acute and chronic medical conditions.

Table 2.
Current and potential anti-platelet therapies.
| Molecule | Target | Inhibitory Mechanism |
|---|---|---|
| Aspirin and NSAIDs | Cyclooxygenase1 | Blocks TXA2 formation |
| Clopidogrel | P2Y12 | Irreversibly inhibits ADP receptors |
| Ticagrelor | P2Y12 | Reversibly inhibits ADP receptors |
| Cangrelor | P2Y12 | Reversibly inhibits ADP receptors |
| Prasugrel | P2Y12 | Irreversibly inhibits ADP receptors |
| Tirofiban | GPIIbIIIa | Blocks integrin |
| Eptifibatide | GPIIbIIIa | Blocks integrin |
| Abciximab | GPIIbIIIa | Blocks integrin |
| Vorapaxar | PAR1 | Blocked thrombin receptors |
| Iloprost | PGI2 analogue | Increases platelet cAMP levels, thus acting as an intravenous reversible antiplatelet agent. |
| Cilostazol | PDE3A | Inhibits adenosine cellular uptake |
| Dipyridamole | PDE3/5 | Scavenge peroxy radicals and increase interstitial adenosine levels |
| Revacept | CLEC2/GPVI | Competes with platelet GPVI for binding to collagen |
| Quercetin | PDI | PI3K/Akt inactivation, cAMP elevation |
8. Conclusions
In a nutshell, platelets are not merely cellular fragments or hemostatic agents but versatile players in the immune system with profound implications for health and many diseases. The ability of these small anucleated blood cells to draw a connection between inflammation and immune responses makes them key targets for therapeutics aimed at thromboinflammation, autoimmunity, and infection. As our understanding of platelet immunobiology continues to evolve, so too will the opportunities to harness their potential for improving patient outcomes in a wide range of immune-mediated conditions.
Despite significant progress in antiplatelet therapy, atherothrombotic events remain a major cause of death worldwide. Current treatments provide incomplete protection and pose bleeding risks. Efforts are now directed at creating therapies that target platelet receptors and thrombosis pathways while preserving normal hemostatic functions. Continued research is crucial to overcome these challenges and enhance cardiovascular disease management.
Author Contributions
S.R. conceived the review article topic, conducted the literature search, drafted the initial manuscript, and revised the final version. S.M.Y. reviewed the literature, provided substantial feedback on the article structure and content, and helped with final edits. All authors reviewed and approved the final review article.
Funding
This article received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| VITT | Vaccine-induced immune thrombotic thrombocytopenia |
| TPO | Thrombopoietin |
| MK | Megakaryocytes |
| MVB | Multivesicular bodies |
| AG | Alpha Granules |
| TGN | Trans-Golgi network |
| VPS33B | Vacuolar Protein Sorting 33B |
| VPS16B | Vacuolar Protein Sorting 16B |
| NBEAL2 | Neurobeachin-like 2 |
| vWF | von Willebrand factor |
| PDGF | Platelet-derived growth factor |
| VEGF | vascular endothelial growth factor |
| TGF-β | Transforming growth factor-beta |
| DG | Dense Granules |
| BLOC | Biogenesis of lysosome-related organelles complex |
| ADP | Adenosine diphosphate |
| ATP | Adenosine triphosphate |
| VAMP | Vesicle-associated membrane protein |
| SNAP23 | Synaptosome-associated protein 23 |
| STIM | Stromal Interaction Molecule |
| PAR | Protease-Activated Receptor |
| PF4 | Platelet factor 4 |
| EGFR | Epidermal growth factor receptor |
| PRP | Platelet-rich plasma |
| 5-HIAA | 5-Hydroxy indoleacetic acid |
| PRR | Pattern recognition receptors |
| TLRs | Toll-like receptors |
| CLRs | C-type lectin receptors |
| NLRs | NOD-like receptors |
| NET | Neutrophil extracellular traps |
| STING | Stimulator of interferon genes |
| SLE | Systemic lupus erythematosus |
| DCs | Dendritic cells |
| EPCR | Endothelial protein C receptor |
| IVIG | Intravenous immunoglobulin |
| HIT | Heparin-Induced Thrombotic Thrombocytopenia |
| DAPT | Dual antiplatelet therapy |
References
- Scherlinger, M.; et al. The role of platelets in immune-mediated inflammatory diseases. Nat Rev Immunol. 2023, 23, 495–510. [Google Scholar] [CrossRef]
- Maouia, A.; et al. The Immune Nature of Platelets Revisited. Transfus Med Rev 2020, 34, 209–220. [Google Scholar] [CrossRef]
- Franco, A.T.; Corken, A.; Ware, J. Platelets at the interface of thrombosis, inflammation, and cancer. Blood 2015, 126, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Machlus, K.R.; Italiano, J.E. Jr. The incredible journey: From megakaryocyte development to platelet formation. J Cell Biol 2013, 201, 785–796. [Google Scholar] [PubMed]
- Bergmeier, W.; Stefanini, L. Platelet ITAM signaling. Curr Opin Hematol 2013, 20, 445–450. [Google Scholar] [CrossRef]
- Chen, Y.; Yuan, Y.; Li, W. Sorting machineries: how platelet-dense granules differ from alpha-granules. Biosci Rep 2018, 38. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.H.; et al. alpha-granule biogenesis: from disease to discovery. Platelets 2017, 28, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Flaumenhaft, R. alpha-granules: a story in the making. Blood 2012, 120, 4908–4909. [Google Scholar] [CrossRef] [PubMed]
- Sharda, A.; Flaumenhaft, R. The life cycle of platelet granules. F1000Res 2018, 7, 236. [Google Scholar] [CrossRef]
- Smith, C.W. Release of alpha-granule contents during platelet activation. Platelets 2022, 33, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Golebiewska, E.M.; Poole, A.W. Platelet secretion: From haemostasis to wound healing and beyond. Blood Rev, 2015, 29, 153–162. [Google Scholar] [CrossRef]
- Etulain, J.; et al. P-selectin promotes neutrophil extracellular trap formation in mice. Blood 2015, 126, 242–246. [Google Scholar] [CrossRef] [PubMed]
- Ambrosio, A.L.; Boyle, J.A.; Di Pietro, S.M. Mechanism of platelet dense granule biogenesis: study of cargo transport and function of Rab32 and Rab38 in a model system. Blood 2012, 120, 4072–4081. [Google Scholar] [CrossRef] [PubMed]
- King, S.M.; et al. Platelet dense-granule secretion plays a critical role in thrombosis and subsequent vascular remodeling in atherosclerotic mice. Circulation 2009, 120, 785–791. [Google Scholar] [CrossRef] [PubMed]
- Thon, J.N.; Italiano, J.E. Platelets: production, morphology and ultrastructure. Handb Exp Pharmacol 2012, 3–22. [Google Scholar]
- Heijnen, H.; van der Sluijs, P. Platelet secretory behaviour: as diverse as the granules... or not? J Thromb Haemost 2015, 13, 2141–2151. [Google Scholar] [CrossRef]
- Rendu, F.; Brohard-Bohn, B. The platelet release reaction: granules’ constituents, secretion and functions. Platelets 2001, 12, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Italiano, J.E., Jr.; Battinelli, E.M. Selective sorting of alpha-granule proteins. J Thromb Haemost 2009, 7 Suppl 1, 173–176. [Google Scholar] [CrossRef]
- Silvestre-Roig, C.; et al. Neutrophils as regulators of cardiovascular inflammation. Nat Rev Cardiol 2020, 17, 327–340. [Google Scholar] [CrossRef]
- Smyth, S.S.; et al. Beta(3)-integrin-deficient mice but not P-selectin-deficient mice develop intimal hyperplasia after vascular injury: correlation with leukocyte recruitment to adherent platelets 1 hour after injury. Circulation 2001, 103, 2501–2507. [Google Scholar] [CrossRef] [PubMed]
- Morrell, C.N.; et al. Emerging roles for platelets as immune and inflammatory cells. Blood 2014, 123, 2759–2767. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, N.; et al. Platelets at the Crossroads of Pro-Inflammatory and Resolution Pathways during Inflammation. Cells 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; et al. Platelets in hemostasis and thrombosis: Novel mechanisms of fibrinogen-independent platelet aggregation and fibronectin-mediated protein wave of hemostasis. J Biomed Res 2015, 29, 437–444. [Google Scholar]
- Chen, Y.; et al. Role of platelet biomarkers in inflammatory response. Biomark Res 2020, 8, 28. [Google Scholar] [CrossRef]
- Kisucka, J.; et al. Platelets and platelet adhesion support angiogenesis while preventing excessive hemorrhage. Proc Natl Acad Sci USA 2006, 103, 855–860. [Google Scholar] [CrossRef] [PubMed]
- Sopova, K.; Tatsidou, P.; Stellos, K. Platelets and platelet interaction with progenitor cells in vascular homeostasis and inflammation. Curr Vasc Pharmacol 2012, 10, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; et al. Platelet, a key regulator of innate and adaptive immunity. Front Med (Lausanne) 2023, 10, 1074878. [Google Scholar] [CrossRef] [PubMed]
- Ho-Tin-Noe, B. The multifaceted roles of platelets in inflammation and innate immunity. Platelets 2018, 29, 531–532. [Google Scholar] [CrossRef]
- Ho-Tin-Noe, B.; Boulaftali, Y.; Camerer, E. Platelets and vascular integrity: how platelets prevent bleeding in inflammation. Blood 2018, 131, 277–288. [Google Scholar] [CrossRef]
- Bordon, Y. Innate immunity: Platelets on the prowl. Nat Rev Immunol 2018, 18, 3. [Google Scholar] [CrossRef]
- Nording, H.; Langer, H.F. Complement links platelets to innate immunity. Semin Immunol 2018, 37, 43–52. [Google Scholar] [CrossRef]
- Semple, J.W.; Freedman, J. Platelets and innate immunity. Cell Mol Life Sci 2010, 67, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Kapur, R.; Semple, J.W. Platelets as immune-sensing cells. Blood Adv 2016, 1, 10–14. [Google Scholar] [CrossRef]
- Lam, F.W.; Vijayan, K.V.; Rumbaut, R.E. Platelets and Their Interactions with Other Immune Cells. Compr Physiol 2015, 5, 1265–1280. [Google Scholar] [PubMed]
- Rolfes, V.; et al. Platelets Fuel the Inflammasome Activation of Innate Immune Cells. Cell Rep 2020, 31, 107615. [Google Scholar] [CrossRef] [PubMed]
- Speth, C.; et al. Platelets as immune cells in infectious diseases. Future Microbiol, 2013, 8, 1431–1451. [Google Scholar] [CrossRef] [PubMed]
- von Hundelshausen, P.; Weber, C. Platelets as immune cells: bridging inflammation and cardiovascular disease. Circ Res, 2007, 100, 27–40. [Google Scholar] [CrossRef]
- Bo, Y.; et al. The role of platelets in central hubs of inflammation: A literature review. Medicine (Baltimore), 2024, 103, e38115. [Google Scholar] [CrossRef]
- Cleary, S.J.; Conrad, C. Investigating and imaging platelets in inflammation. Int J Biochem Cell Biol, 2023, 157, 106373. [Google Scholar] [CrossRef] [PubMed]
- Contursi, A.; et al. Platelets as crucial players in the dynamic interplay of inflammation, immunity, and cancer: unveiling new strategies for cancer prevention. Front Pharmacol, 2024, 15, 1520488. [Google Scholar] [CrossRef] [PubMed]
- Rayes, J.; et al. The dual role of platelet-innate immune cell interactions in thrombo-inflammation. Res Pract Thromb Haemost, 2020, 4, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Vulliamy, P.; Armstrong, P.C. Platelets in Hemostasis, Thrombosis, and Inflammation After Major Trauma. Arterioscler Thromb Vasc Biol, 2024, 44, 545–557. [Google Scholar] [CrossRef]
- Carestia, A.; et al. Platelets Promote Macrophage Polarization toward Pro-inflammatory Phenotype and Increase Survival of Septic Mice. Cell Rep, 2019, 28, 896–908. [Google Scholar] [CrossRef] [PubMed]
- Yip, J.Y.; et al. Optimizing intrauterine insemination and spontaneous conception in women with unilateral hydrosalpinx or tubal pathology: A systematic review and narrative synthesis. Eur J Obstet Gynecol Reprod Biol, 2023, 286, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, R.; et al. Effect of Platelet-Rich Plasma on M1/M2 Macrophage Polarization. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- De Giovanni, M.; et al. GPR35 promotes neutrophil recruitment in response to serotonin metabolite 5-HIAA. Cell, 2022, 185, 1103–1104. [Google Scholar] [CrossRef] [PubMed]
- Revenstorff, J.; et al. Role of S100A8/A9 in Platelet-Neutrophil Complex Formation during Acute Inflammation. Cells 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Schrottmaier, W.C.; et al. Platelet p110beta mediates platelet-leukocyte interaction and curtails bacterial dissemination in pneumococcal pneumonia. Cell Rep, 2022, 41, 111614. [Google Scholar] [CrossRef]
- Shang, Y.; et al. Activated platelet membrane nanovesicles recruit neutrophils to exert the antitumor efficiency. Front Chem, 2022, 10, 955995. [Google Scholar] [CrossRef] [PubMed]
- Clark, S.R.; et al. Platelet TLR4 activates neutrophil extracellular traps to ensnare bacteria in septic blood. Nat Med, 2007, 13, 463–469. [Google Scholar] [CrossRef] [PubMed]
- de Stoppelaar, S.F.; et al. The role of platelet MyD88 in host response during gram-negative sepsis. J Thromb Haemost, 2015, 13, 1709–1720. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; et al. Nucleotide-binding oligomerization domain 2 receptor is expressed in platelets and enhances platelet activation and thrombosis. Circulation, 2015, 131, 1160–1170. [Google Scholar] [CrossRef]
- Brown, G.T.; McIntyre, T.M. Lipopolysaccharide signaling without a nucleus: kinase cascades stimulate platelet shedding of proinflammatory IL-1beta-rich microparticles. J Immunol, 2011, 186, 5489–5496. [Google Scholar] [CrossRef] [PubMed]
- Aggrey, A.A.; et al. Platelet induction of the acute-phase response is protective in murine experimental cerebral malaria. J Immunol, 2013, 190, 4685–4691. [Google Scholar] [CrossRef] [PubMed]
- Denis, M.M.; et al. Escaping the nuclear confines: signal-dependent pre-mRNA splicing in anucleate platelets. Cell, 2005, 122, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Boylan, B.; et al. Identification of FcgammaRIIa as the ITAM-bearing receptor mediating alphaIIbbeta3 outside-in integrin signaling in human platelets. Blood, 2008, 112, 2780–2786. [Google Scholar] [CrossRef]
- Zhi, H.; et al. Cooperative integrin/ITAM signaling in platelets enhances thrombus formation in vitro and in vivo. Blood, 2013, 121, 1858–1867. [Google Scholar] [CrossRef]
- Arman, M.; et al. Amplification of bacteria-induced platelet activation is triggered by FcgammaRIIA, integrin alphaIIbbeta3, and platelet factor 4. Blood, 2014, 123, 3166–3174. [Google Scholar] [CrossRef]
- Boilard, E.; et al. Influenza virus H1N1 activates platelets through FcgammaRIIA signaling and thrombin generation. Blood, 2014, 123, 2854–2863. [Google Scholar] [CrossRef] [PubMed]
- Karas, S.P.; Rosse, W.F.; Kurlander, R.J. Characterization of the IgG-Fc receptor on human platelets. Blood, 1982, 60, 1277–1282. [Google Scholar] [CrossRef]
- Sun, D.; et al. Bacillus anthracis peptidoglycan activates human platelets through FcgammaRII and complement. Blood, 2013, 122, 571–579. [Google Scholar] [CrossRef]
- Benezech, C.; et al. CLEC-2 is required for development and maintenance of lymph nodes. Blood, 2014, 123, 3200–3207. [Google Scholar] [CrossRef] [PubMed]
- Hitchcock, J.R.; et al. Inflammation drives thrombosis after Salmonella infection via CLEC-2 on platelets. J Clin Invest, 2015, 125, 4429–4446. [Google Scholar] [CrossRef]
- Al Hawas, R.; et al. Munc18b/STXBP2 is required for platelet secretion. Blood, 2012, 120, 2493–2500. [Google Scholar] [CrossRef]
- Yang, M.; et al. STING activation in platelets aggravates septic thrombosis by enhancing platelet activation and granule secretion. Immunity, 2023, 56, 1013–1026. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.; et al. The 17beta-estradiol induced upregulation of the adhesion G-protein coupled receptor (ADGRG7) is modulated by ESRalpha and SP1 complex. Biol Open 2019, 8. [Google Scholar]
- Bhagirath, V.C.; Dwivedi, D.J.; Liaw, P.C. Comparison of the Proinflammatory and Procoagulant Properties of Nuclear, Mitochondrial, and Bacterial DNA. Shock, 2015, 44, 265–271. [Google Scholar] [CrossRef]
- Boudreau, L.H.; et al. Platelets release mitochondria serving as substrate for bactericidal group IIA-secreted phospholipase A2 to promote inflammation. Blood, 2014, 124, 2173–2183. [Google Scholar] [CrossRef]
- Melki, I.; et al. Platelets release mitochondrial antigens in systemic lupus erythematosus. Sci Transl Med 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Puhm, F.; Boilard, E.; Machlus, K.R. Platelet Extracellular Vesicles: Beyond the Blood. Arterioscler Thromb Vasc Biol, 2021, 41, 87–96. [Google Scholar] [CrossRef]
- Tavukcuoglu, Z.; et al. Platelet-derived extracellular vesicles induced through different activation pathways drive melanoma progression by functional and transcriptional changes. Cell Commun Signal, 2024, 22, 601. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Itagaki, K.; Hauser, C.J. Mitochondrial DNA is released by shock and activates neutrophils via p38 map kinase. Shock, 2010, 34, 55–59. [Google Scholar] [CrossRef]
- Nish, S.; Medzhitov, R. Host defense pathways: role of redundancy and compensation in infectious disease phenotypes. Immunity, 2011, 34, 629–636. [Google Scholar] [CrossRef]
- Nicolai, L.; Pekayvaz, K.; Massberg, S. Platelets: Orchestrators of immunity in host defense and beyond. Immunity, 2024, 57, 957–972. [Google Scholar] [CrossRef]
- Ali, R.A.; Wuescher, L.M.; Worth, R.G. Platelets: essential components of the immune system. Curr Trends Immunol, 2015, 16, 65–78. [Google Scholar]
- Hirosue, S.; Dubrot, J. Modes of Antigen Presentation by Lymph Node Stromal Cells and Their Immunological Implications. Front Immunol, 2015, 6, 446. [Google Scholar] [CrossRef] [PubMed]
- Tsukiji, N.; Suzuki-Inoue, K. Impact of Hemostasis on the Lymphatic System in Development and Disease. Arterioscler Thromb Vasc Biol, 2023, 43, 1747–1754. [Google Scholar] [CrossRef] [PubMed]
- Han, P.; et al. Platelet P-selectin initiates cross-presentation and dendritic cell differentiation in blood monocytes. Sci Adv 2020, 6, eaaz1580. [Google Scholar] [CrossRef] [PubMed]
- Ramanujam, M.; et al. Phoenix from the flames: Rediscovering the role of the CD40-CD40L pathway in systemic lupus erythematosus and lupus nephritis. Autoimmun Rev, 2020, 19, 102668. [Google Scholar] [CrossRef] [PubMed]
- Hess, P.R.; et al. Platelets mediate lymphovenous hemostasis to maintain blood-lymphatic separation throughout life. J Clin Invest, 2014, 124, 273–284. [Google Scholar] [CrossRef]
- Cognasse, F.; et al. Human platelets can activate peripheral blood B cells and increase production of immunoglobulins. Exp Hematol, 2007, 35, 1376–1387. [Google Scholar] [CrossRef] [PubMed]
- Greinacher, A.; et al. Pathogenesis of vaccine-induced immune thrombotic thrombocytopenia (VITT). Semin Hematol, 2022, 59, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Mendonca, S.A.; et al. Adenoviral vector vaccine platforms in the SARS-CoV-2 pandemic. NPJ Vaccines, 2021, 6, 97. [Google Scholar] [CrossRef]
- Roytenberg, R.; Garcia-Sastre, A.; Li, W. Vaccine-induced immune thrombotic thrombocytopenia: what do we know hitherto? Front Med (Lausanne), 2023, 10, 1155727. [Google Scholar] [CrossRef]
- Thiele, T.; Schwarz, S.L.; Handtke, S. Platelet size as a mirror for the immune response after SARS-CoV-2 vaccination. J Thromb Haemost, 2022, 20, 818–820. [Google Scholar] [CrossRef]
- Hirsch, J.; et al. Platelet-neutrophil interaction in COVID-19 and vaccine-induced thrombotic thrombocytopenia. Front Immunol 2023, 14, 1186000. [Google Scholar] [CrossRef]
- Ostrowski, S.R.; et al. Inflammation and Platelet Activation After COVID-19 Vaccines - Possible Mechanisms Behind Vaccine-Induced Immune Thrombocytopenia and Thrombosis. Front Immunol 2021, 12, 779453. [Google Scholar] [CrossRef]
- Ferro, J.M.; et al. European stroke organization interim expert opinion on cerebral venous thrombosis occurring after SARS-CoV-2 vaccination. Eur Stroke J 2021, 6, CXVI–CXXI. [Google Scholar] [CrossRef] [PubMed]
- Gresele, P.; et al. Management of cerebral and splanchnic vein thrombosis associated with thrombocytopenia in subjects previously vaccinated with Vaxzevria (AstraZeneca): a position statement from the Italian Society for the Study of Haemostasis and Thrombosis (SISET). Blood Transfus 2021, 19, 281–283. [Google Scholar] [PubMed]
- Jacobson, B.F.; et al. Recommendations for the diagnosis and management of vaccine-induced immune thrombotic thrombocytopenia. S Afr Med J 2021, 111, 535–537. [Google Scholar]
- Tutwiler, V.; et al. Platelet transactivation by monocytes promotes thrombosis in heparin-induced thrombocytopenia. Blood.
- Stolla, M.; Kapur, R.; Semple, J.W. New Emerging Developments of Platelets in Transfusion Medicine. Transfus Med Rev 2020, 34, 207–208. [Google Scholar] [CrossRef]
- Fletcher, C.H.; DomBourian, M.G.; Millward, P.A. Platelet transfusion for patients with cancer. Cancer Control 2015, 22, 47–51. [Google Scholar] [CrossRef]
- Du, X.; et al. Clinical application of platelet rich plasma to promote healing of open hand injury with skin defect. Regen Ther 2024, 26, 308–314. [Google Scholar] [CrossRef]
- van der Meijden, P.E.J.; Heemskerk, J.W.M. Platelet biology and functions: new concepts and clinical perspectives. Nat Rev Cardiol 2019, 16, 166–179. [Google Scholar] [CrossRef]
- Jones, W.S.; et al. Comparative Effectiveness of Aspirin Dosing in Cardiovascular Disease. N Engl J Med 2021, 384, 1981–1990. [Google Scholar] [CrossRef]
- Alagna, G.; et al. Dual Antiplatelet Therapy with Parenteral P2Y(12) Inhibitors: Rationale, Evidence, and Future Directions. J Cardiovasc Dev Dis 2023, 10. [Google Scholar]
- Halvorsen, S.; et al. Aspirin therapy in primary cardiovascular disease prevention: a position paper of the European Society of Cardiology working group on thrombosis. J Am Coll Cardiol 2014, 64, 319–327. [Google Scholar] [CrossRef]
- McFadyen, J.D.; Schaff, M.; Peter, K. Current and future antiplatelet therapies: emphasis on preserving haemostasis. Nat Rev Cardiol 2018, 15, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Olie, R.H.; van der Meijden, P.E.J.; Cate, H.T. The coagulation system in atherothrombosis: Implications for new therapeutic strategies. Res Pract Thromb Haemost 2018, 2, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Patrono, C.; et al. Antiplatelet Agents for the Treatment and Prevention of Coronary Atherothrombosis. J Am Coll Cardiol 2017, 70, 1760–1776. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, P.C.; et al. In the presence of strong P2Y12 receptor blockade, aspirin provides little additional inhibition of platelet aggregation. J Thromb Haemost 2011, 9, 552–561. [Google Scholar] [CrossRef]
- Best, M.G.; Vancura, A.; Wurdinger, T. Platelet RNA as a circulating biomarker trove for cancer diagnostics. J Thromb Haemost 2017, 15, 1295–1306. [Google Scholar] [CrossRef]
- Xiang, Y.Z.; et al. Platelet activation, and antiplatelet targets and agents: current and novel strategies. Drugs 2008, 68, 1647–1664. [Google Scholar] [CrossRef]
- Duley, L.; et al. Antiplatelet agents for preventing pre-eclampsia and its complications. Cochrane Database Syst Rev 2019, 2019. [Google Scholar] [CrossRef]
- Safouris, A.; Magoufis, G.; Tsivgoulis, G. Emerging agents for the treatment and prevention of stroke: progress in clinical trials. Expert Opin Investig Drugs 2021, 30, 1025–1035. [Google Scholar] [CrossRef] [PubMed]
- Gelbenegger, G.; Jilma, B. Clinical pharmacology of antiplatelet drugs. Expert Rev Clin Pharmacol 2022, 15, 1177–1197. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Multifaceted Roles of Platelets in Health and Diseases.
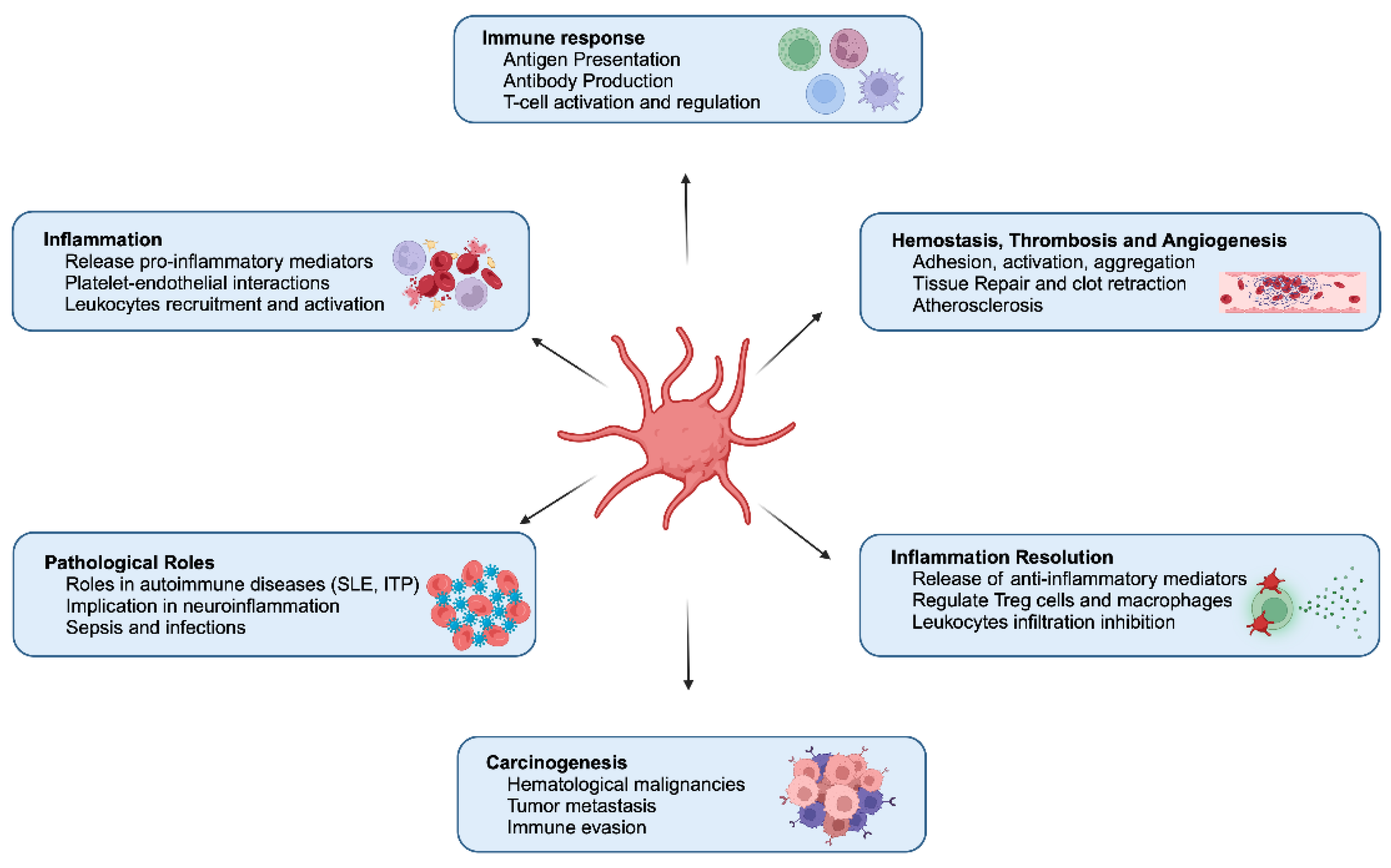
Table 1.
Platelet granules contents.
 |
|---|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



