Submitted:
31 January 2025
Posted:
03 February 2025
You are already at the latest version
Abstract
Bovine respiratory syncytial virus (BRSV) is a major pathogen of the bovine respiratory disease complex (BRD) and causes regular severe winter outbreaks of respiratory disease in cattle. It is therefore responsible for important economic losses for the farming industry. In this study, the genetic diversity of circulating BRSV strains in Belgium, which has not been assessed since the end of the 1990’s, was investigated. We analysed 51 BRSV-positive samples collected from 2015 to 2023. Genetic analysis of the BRSV G partial gene sequences revealed the presence of BRSV strains belonging to the subgroups VII and VIII recently identified in Europe. Furthermore, co-circulation of subgroups II and III were recorded in the same period. Four commercially available vaccine strains marketed for Belgium were also included in the analysis and they clustered with subgroups II or III. Our findings that different strains of BRSV are circulating in Belgium highlight the importance of ensuring that available vaccines efficiently protect against strains from circulating subgroups and of assessing potential circulation of attenuated vaccine strains.
Keywords:
vaccine strains
; brsv
; genotype
; veterinary viruses
; bovine viruses
1. Introduction
Bovine respiratory syncytial virus (BRSV or Bovine Orthopneumovirus) is a member of the Orthopneumovirus genus within the Pneumoviridae family. It is a major pathogen of the Bovine Respiratory Disease complex (BRD) in beef and dairy calves. The peak incidence of severe disease concerns mainly calves younger than 6 months of age [1,2] and it causes regular severe winter outbreaks of respiratory disease in cattle [3]. BRSV is closely related to the human respiratory syncytial virus (HRSV), the most important cause of lower respiratory tract disease (LRTD) in children [4]. BRSV and HRSV share common epidemiological, pathological and clinical characteristics [5]. Originally discovered in Europe [6], BRSV is now distributed worldwide [7,8,9]. Its impact on the cattle industry is associated with economic losses as a result of morbidity, mortality, costs of treatment and prevention, loss of production, and reduced carcass value [10].
The genomic heterogeneity of the negative-stranded RNA BRSV genome and its low fidelity in replication are important drivers of viral evolution and escape from vaccine protection [11]. Therefore, the analysis of BRSV strain diversity is essential for the development and update of efficacious vaccines. The gene encoding the attachment glycoprotein G has the highest reported mutation rate among BRSV strains and is commonly used as a target for phylogenetic analyses [5]. Based on the sequence variability of the G ORF within essential immunodominant regions, BRSV has been classified for a long time into six different genetic subgroups [5,12,13]. With better surveillance of BRSV circulation and given its high prevalence around the world, the number of subgroups has steadily increased; over the past two decades, the number of BRSV subgroups has ranged from six to eight subgroups [14,15], and more recently, to ten subgroups [16]. An extensive study of the diversity of circulating BRSV strains in Europe is therefore needed to characterize the up-to-date circulation of distinct genetic subgroups on the continent.
In Belgium, the genetic diversity of circulating BRSV strains has not been assessed since the end of the 1990’s. The Wallonia region of southern Belgium plays an important role in the overall European livestock industry with one of the largest livestock densities, the birth place of the Belgian Blue breed and a central spatial position in Western Europe, sharing borders with France, Germany, Luxembourg and The Netherlands - all of which are major livestock producers on the continent [17]. An outbreak of any livestock disease in Wallonia would therefore have a high potential to spread to these countries.
The aim of this study was to provide an update on the genetic diversity of circulating BRSV strains in the Wallonia region of Belgium.
2. Materials and Methods
2.1. Field Samples
The objective of the study was to collect as many BRSV-positive samples as possible, either from the lungs of deceased animals or from bronchoalveolar lavage. The collection was conducted in collaboration with the necropsy room of the University of Liège and the laboratory of the Regional Association for Animal Identification and Health (ARSIA) between 2015 and 2023. Along with the age of the animals detected as positive for the virus, information on their vaccination status was collected. The proportion of vaccinated animals was compared between the main genetic BRSV subgroups using a Z-test for two proportions.
2.2. BRSV Vaccines
To evaluate the phylogenetic relationship between circulating viral strains and vaccine strains, we included the vaccine strains in our analysis to determine their positioning relative to the circulating strains.
Four modified-live viral (MLV) vaccine strains marketed in Belgium were obtained from vaccine bottles: MLV-A, MLV-B, MLV-C and MLV-D.
2.3. Extraction of BRSV RNA
For each sample, 50 mg of lung tissue or 200 μL of nasal swab preservation fluid were set in a 2 mL Eppendorf tube, along 500 µl of Nucleozol solution (Macherey-Nagel). The tubes with lung tissue samples were homogenized with a 5 mm steel bead (Qiagen) using the Qiagen Tissuelyser II device (3-minute cycle at a frequency of 30Hz). The homogenate of all samples was centrifuged, with the supernatant being removed for subsequent RNA purification using the Nucleozol RNA extraction kit (Macherey-Nagel), following the manufacturer’s instructions.
RNA from 3 samples with low quality extraction (B22.0873, B19.223592 and B21.0944) was isolated by using TANBead® Nucleic Acid Extraction kit (Taiwan Advanced Nanotech Inc.) according to the manufacturer’s instructions.
2.4. RT-PCR Targeting the BRSV G Gene
Primers corresponding to a 541 bp fragment of the BRSV Glycoprotein (G) open reading frame were used, as previously described [5]. The primers used were as follow (5’ to 3’): VG1, CCACCCTAGCAATGATAACCTTGAC; VG4, GCTAGTTCTGTGGTGGATTGTTGTC. First strand cDNA synthesis was generated using the Luna® Universal Probe One-Step RT-qPCR Kit (New England Biolabs), where 10-15 µg of total extracted RNA were incubated with 0.4 µM of both VG1 and VG4 primers, using reaction buffers and enzymes according to the manufacturer’s instructions. The amplification conditions were 55oC for 20min for RT, denaturation at 95°C for 5min, followed by 50 cycles at 95°C for 30 s, 55°C for 30 s, and 72°C for 60 s and a final elongation at 72°C for 5 min. A second set containing one designed primer (Geneious 10.2.6 software, Biomatters Ltd) corresponding to a 177 bp fragment were applied to 11 samples to extend the 5’end. The primers used were as follow (5’ to 3’): FNew CCAAACACTCCCCYATGTGCCTTG associated with the same VG4. We increased the primer hybridization temperature to 62°C to accommodate the minimum temperature requirement while keeping the rest of the amplification conditions unchanged. The PCR reactions were then run on a 1% agarose gel, stained with a fluorescent reagent (Midori Green Advance; Nippon Genetics), and detected under blue-green LED light. Bands corresponding to the theoretical molecular weight (i.e. 541 bp or 177 bp, respectively) being cut out of the gel for DNA purification (Figure 1).
2.5. DNA Purification and Sequencing of BRSV G Gene
The amplicons were gel purified using the Nucleospin Gel and PCR Clean-Up kit (Macherey-Nagel), following the manufacturer’s instructions. Sanger sequencing was performed by Eurofins Genomics, using VG1/FNew and VG4 primers. The nucleotide sequences used in this study have been submitted to GenBank and assigned the accession numbers PP538078 – PP538081 and PP538083 – PP538133.
2.6. Phylogenetic Analysis
All partial G ORF cDNAs were aligned by the ClustalW multiple alignment tool using the Geneious 10.2.6 software (Biomatters Ltd). The experimental and vaccine sequences were compared with reference sequences of BRSV subgroups I-VIII selected from GenBank. All the experimental and reference sequences were equally trimmed to 418 bp. A reference sequence from ovine respiratory syncytial virus (ORSV) was used as an outgroup for subsequent phylogenetic analyses.
The phylogenetic tree was generated with the MegaX software package [19], using the Maximum Likelihood method with Tamura-Nei with gamma distribution, as determined by a model prediction analysis. The analysis was performed on untranslated cDNA sequences with 1000 bootstrap replications.
2.7. Amino Acid Sequences Analysis
Deduced amino acids from the major immunodominant zone (aa159-aa186) of both experimental and reference sequences were aligned in Geneious 10.2.6 software (Biomatters Ltd). Residues at positions 159-186 correspond to the central conserved region of the extracellular of the BRSV G glycoprotein, which is an important antigenic site [20].
3. Results
3.1. Sample Collection
Lung tissue samples (n=48) or nasal swabs (n=3) from 51 BRSV-infected cattle were obtained from the “laboratory and diagnosis” department of the Regional Association for Animal Health and Identification (n=17) and from the necropsy room of the veterinary Faculty of Liège (n=34) (Table 1). The mean age of our cohort was 5.5 months (standard deviation: 4.2 months). The vaccination status was obtained for 42 animals. Twenty-one (50%) were vaccinated. For the subgroup II, the proportion of vaccinated animals was 10/17 (58,8%). The proportion was lower for the subgroup VIII reaching 10/24 (41,6%). However, the Z-test applied to this two proportions showed no significant difference (p-value = 1.09 > 0.05).
3.2. Sequencing Results
Virus isolates belong to three different phylogenetic subgroups in Wallonia (II, III and VIII). The majority (94,1%) of samples were isolated from lung tissue after necropsy. Age, vaccine status of animals and the accession numbers of the sequences are reported in Table 1.
3.3. Phylogenetic Analysis
Phylogenetic tree was constructed using the Maximum Likelihood methodo allow comparison and clustering of the nucleotide sequences of experimental, vaccine and reference isolates (Figure 2).
Almost all field isolates phylogenetically clustered with reference strains from either subgroup II or subgroup VIII; out of the 51 field isolates, 20 clustered with the subgroup II reference sequences, while 30 isolates clustered with subgroup VIII. One field isolate (B19.207920) clustered with the reference sequences from subgroup III.
Among the four live-attenuated vaccine strains included in the study (Table 1), two vaccines clustered with the subgroup II reference sequences (MLV-A and MLV-B), while the other two (MLV-C and MLV-D) clustered with the subgroup III. The MLV-D vaccine strain and the B19.207920 experimental isolate seem to be phylogenetically very close with only one nucleotide substitution (silent substitution).
3.4. Amino Acid Sequences Analysis
The translated sequences of the cloned BRSV cDNAs were analysed for variability at specific amino acid positions within the major immunodominant zone located in the central conserved region of ectodomain (aa159-aa186) (Figure 3).
The four cysteine residues of the conserved central hydrophobic part of the ectodomain of the G glycoprotein of all Belgian BRSV strains were conserved (i.e. positions 173, 176, 182 and 186) [11]. Some mutations were found within the central conserved region of some experimental and reference samples from different subgroups. Among the samples from subgroup II, glutamate at position 177 was replaced by a lysine residue (E177 → K177) and leucine in position 183 replaced by serine (L183 → S183). The mutations found within subgroup VIII were as follows: P156 → S156, H162 → Q162, N163 → T163 and S165 → L165. Concerning vaccine strains (Table 1), several variations were observed within the immunodominant region. MLV-C strain contains phenylalanine at position 165. As described for field strains, reference strains and MLV-A vaccine strain from subgroup II, the glutamate in position 177 was replaced by a lysine (E177 → K177). On the contrary, the other subgroup II-related MLV-B vaccine strain retained glutamate at this position (Figure 3). It should be noted that the MLV-C vaccine strain and the isolate strain B19.207920 belonging to subgroup III differed by only one amino acid (T102 → G102, not shown).
4. Discussion
The genetic diversity of circulating BRSV strains in Belgium over a period of eight years (2015-2023) was analyzed based on the sequence coding for the extracellular domain of the glycoprotein G. Interestingly, the results of our phylogenetic analyzes confirmed the current classification of BRSV strains into eight subgroups, as observed recently elsewhere in Europe [15]. Sequences of subgroups IX [21] and X [16], which have been described respectively in Brazil and Japan, were not included in our analysis due to their short size (318 bp).
A majority of the strains (58.8%) clustered with subgroup VIII, a subgroup recently described on the European territory, more particularly in Croatia, but with a lower prevalence (37,5%; 9/24) [15]. Circulating BRSV strains in Belgium belong to the new subgroup VIII identified herein for the first time. This genetic evolution of BRSV could be heightened by herd immunity and in particular by the vaccine pressure. It has already been suggested that BRSV strains from countries with intensive vaccination appear to evolve faster [5]. The BRSV vaccine programs are quite intensive in Belgium [5]. However, although the population of our study is limited, the proportion of vaccinated animals in the different genetic subgroups in this study did not support the hypothesis of a subgroup II mainly found in non-vaccinated animals and a subgroup VIII, which evades immunity more frequently, found in vaccinated animals.
Phylogenetic analysis of vaccine strains available on the Belgian market indicates that they belong to either subgroup II or III. Considering that the majority of field samples were classified as subgroup VIII, this is of particular concern, as it is currently not known whether subgroup VIII is antigenically related to subgroup II or III. Therefore, further studies are required to determine whether the immunity generated by using these vaccines can efficiently protect the host from an infection with the subgroup VIII strain of BRSV.
In addition, 39.2% of field strains belonged to subgroup II, which also contains older Belgian strains (≤1983) and two vaccine strains currently marketed. Contemporary strains from the Nordic countries (Norway, Sweden and Denmark) mostly belong to subgroup II [13,22,23]. However, Belgium does not import cattle from those countries. Interestingly, viral strains belonging to subgroups II have been identified in calves vaccinated with vaccine strains belonged to the same subgroup II. These subgroups II viral and vaccine strains are different but very close with 94 to 96% identity. In that case, an escape from vaccine immunity due to a genetic evolution of the virus is not an option. These results suggest that vaccination in the field faces some practical issues which can decrease the efficiency of commercial vaccines.
As previously mentioned, several BRSV subgroups have been found in Europe, including subgroups I, II, IV, V and VI, VII and VIII [5,14,15]. While most of our experimental samples classified into subgroup II or VIII, one sample (i.e. B19.207920) clearly clustered with subgroup III. This is of particular interest, since subgroup III strains were originally observed in the Americas [5] or, more recently, in Turkey [24] and China [25]. There were no imports of cattle from these countries and their neighboring countries since at least 1989 (unpublished data). According to the breeder, the cattle was vaccinated by intramuscular injection with MLV-C vaccine, also belonging to subgroup III. Since only one synonymous substitution was observed between this isolate and the MLV-C vaccine strain, a vaccine origin seems most likely.
Subgroup V strains were described as being dominant in Belgium in the late 1990s [5]. However, in our study, we did not detect any isolates that belonged to this subgroup. This observation is also in favor of the shifting of the dominant subgroup of circulating BRSV strains in Wallonia.
The 4 cysteines present in the central conserved region of glycoprotein G of BRSV lead to the formation of a central loop maintained by two disulfide bridges (Cys173- Cys186 and Cys176-Cys182) at the top of which is a major epitope site. Amino acids located at position 177, 180, 183 and 184 strongly contribute to antibody binding directed against BRSV [26]. Mutations to these positions change the local antigenic protein surface and influence the antibody reactivity of BRSV [15,16,26].
As shown in Figure 3, all field sequences conserved the 4 cysteines. However, field strains from subgroup II as well as the vaccine strain B24 belonging to this subgroup undergone a mutation at position 177 (E177 → K177) and strains from subgroups I, V, VI and VII show a substitution at position 180 (L180 → P180). All of the strains (field and vaccine) of subgroup II had their leucine replaced by a serine in position 183 (L183 → S183). These differences may be involved in the pattern of immunogenicity evolution and should be investigated further for the development of future vaccines against BRSV. Subgroup VIII strains do not show mutations at the aforementioned antibody binding sites and 60,9% of cattle from which these strains are derived are unvaccinated. Future experiments should be performed to determine if marketed subgroup II and III vaccines are protective against strains of subgroup VIII and the potential role played by the different residues in the cross-protection between subgroups. The mutations found within subgroup VIII (H162 → Q162, N163 → T163 and S165 → L165) were also described in the study of Krešić (2018). However, these residues do not seem to have an antigenic role or influence the three-dimensional structure of the protein [20].
Analysis of samples from Flemish region would be necessary to carry out a genetic characterization of BRSV strains from the whole Belgian territory
5. Conclusions
This study is the first report of the circulation of BRSV subgroups VIII in Belgium. Altogether, these results enhance the need for a continued genetic surveillance of this virus in other European countries to monitor BRSV evolution and its related risk of BRSV strains emergence that could escape herd immunity and/or vaccine protection. Indeed, mutations within immunodominant region of glycoprotein G indicate continuous BRSV evolution, enabling the virus to escape from the host immune response. The efficacy of available vaccines against the current BRSV strains that circulate in the field should incontestably be investigated in the future.
Funding
This research received no external or public funding and was financed by the University of Liège “fonds spéciaux facultaires de recherche (FSR) FSR 2019 RS-CLIM and FSR 2021 RS-CLIM2”.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Acknowledgments
The authors thank the teams of the “laboratory and diagnosis” department of the Regional Association for Animal Health and Identification (ARSIA) and of the necropsy room of the veterinary Faculty of Liège, for the technical and secretary assistance; and J-F Valarcher for critical discussions.
Conflicts of Interest
The authors declare no competing interests.
References
- Lillie, L. E. The Bovine Respiratory Disease Complex. Can. Vet. J. 1974, 15, 233–242. [Google Scholar]
- Stott, E. J.; Taylor, G. Archives of Virology Respiratory Syncytial Virus Brief Review. Arch. Virol. 1985, 84, 1–52. [Google Scholar] [CrossRef] [PubMed]
- Valarcher, J.; Taylor, G.; Valarcher, J.; Taylor, G. Bovine Respiratory Syncytial Virus Infection To Cite This Version : HAL Id : Hal-00902838 Review Article Bovine Respiratory Syncytial Virus Infection. 2007, 38 (2), 153–180.
- Hall CB, Weinberg GA, Iwane MK, Blumkin AK, Edwards KM, Staat MA, Auinger P, Griffin MR, Poehling KA, Erdman D, Grijalva CG, Zhu Y, S. P. Burden of Respiratory Syncytial Virus Infection in Young Children. N Engl J Med. 2009, 360, 588–598. [Google Scholar] [CrossRef]
- Valarcher, J.-F.; Schelcher, F.; Bourhy, H. Evolution of Bovine Respiratory Syncytial Virus. J. Virol. 2000, 74, 10714–10728. [Google Scholar] [CrossRef] [PubMed]
- Doggett, J. E.; Taylor-Robinson, D.; Gallop, R. G. C. A Study of an Inhibitor in Bovine Serum Active against Respiratory Syncytial Virus. Arch. Gesamte Virusforsch. 1968, 23, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Rossi, C. R.; Kiesel, G. K. Serological Evidence for the Association of Bovine Respiratory Syncytial Virus with Respiratory Tract Disease in Alabama Cattle. Infect. Immun. 1974, 10, 293–298. [Google Scholar] [CrossRef]
- Yaegashi, G.; Seimiya, Y. M.; Seki, Y.; Tsunemitsu, H. Genetic and Antigenic Analyses of Bovine Respiratory Syncytial Virus Detected in Japan. J. Vet. Med. Sci. 2005, 67, 145–150. [Google Scholar] [CrossRef]
- Hay, K. E.; Barnes, T. S.; Morton, J. M.; Gravel, J. L.; Commins, M. A.; Horwood, P. F.; Ambrose, R. C.; Clements, A. C. A.; Mahony, T. J. Associations between Exposure to Viruses and Bovine Respiratory Disease in Australian Feedlot Cattle. Prev. Vet. Med. 2016, 127, 121–133. [Google Scholar] [CrossRef]
- Smith, R. A. Effects of Feedlot Disease on Economics, Production and Carcass Value. Am. Assoc. Bov. Pract. Proc. 2000, 125–128. [Google Scholar] [CrossRef]
- Sarmiento-Silva, R. E.; Nakamura-Lopez, Y.; Vaughan, G. Epidemiology, Molecular Epidemiology and Evolution of Bovine Respiratory Syncytial Virus. Viruses 2012, 4, 3452–3467. [Google Scholar] [CrossRef]
- Valentova, V.; Antonis, A. F. G.; Kovarcik, K. Restriction Enzyme Analysis of RT-PCR Amplicons as a Rapid Method for Detection of Genetic Diversity among Bovine Respiratory Syncytial Virus Isolates. Vet. Microbiol. 2005, 108, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Klem, T. B.; Rimstad, E.; Stokstad, M. Occurrence and Phylogenetic Analysis of Bovine Respiratory Syncytial Virus in Outbreaks of Respiratory Disease in Norway. BMC Vet. Res. 2014, 10, 1–9. [Google Scholar] [CrossRef]
- Bertolotti, L.; Giammarioli, M.; Rosati, S. Genetic Characterization of Bovine Respiratory Syncytial Virus Strains Isolated in Italy: Evidence for the Circulation of New Divergent Clades. J. Vet. Diagnostic Investig. 2018, 30, 300–304. [Google Scholar] [CrossRef] [PubMed]
- Krešić, N.; Bedeković, T.; Brnić, D.; Šimić, I.; Lojkić, I.; Turk, N. Genetic Analysis of Bovine Respiratory Syncytial Virus in Croatia. Comp. Immunol. Microbiol. Infect. Dis. 2018, 58, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, A.; Kawauchi, K.; Andoh, K.; Hatama, S. Sequence and Unique Phylogeny of G Genes of Bovine Respiratory Syncytial Viruses Circulating in Japan. J. Vet. Diagnostic Investig. 2021, 33, 162–166. [Google Scholar] [CrossRef]
- Eurostat. Agri-environmental indicator-livestock patterns.
- Boxus, M.; Letellier, C.; Kerkhofs, P. Real Time RT-PCR for the Detection and Quantitation of Bovine Respiratory Syncytial Virus. J. Virol. Methods 2005, 125, 125–130. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Langedijk, J. P. M.; Schaaper, W. M. M.; Meloen, R. H.; Van Oirschot, J. T. Proposed Three-Dimensional Model for the Attachment Protein G of Respiratory Syncytial Virus. J. Gen. Virol. 1996, 77, 1249–1257. [Google Scholar] [CrossRef]
- Leme, R. A.; Dall Agnol, A. M.; Balbo, L. C.; Pereira, F. L.; Possatti, F.; Alfieri, A. F.; Alfieri, A. A. Molecular Characterization of Brazilian Wild-Type Strains of Bovine Respiratory Syncytial Virus Reveals Genetic Diversity and a Putative New Subgroup of the Virus. Vet. Q. 2020, 40, 83–96. [Google Scholar] [CrossRef]
- Bidokhti, M. R. M.; Tråvén, M.; Ohlson, A.; Zarnegar, B.; Baule, C.; Belák, S.; Alenius, S.; Liu, L. Phylogenetic Analysis of Bovine Respiratory Syncytial Viruses from Recent Outbreaks in Feedlot and Dairy Cattle Herds. Arch. Virol. 2012, 157, 601–607. [Google Scholar] [CrossRef]
- Giammarioli, M.; Mangili, P.; Nanni, A.; Pierini, I.; Petrini, S.; Pirani, S.; Gobbi, P.; De Mia, G. M. Highly Pathogenic Bovine Respiratory Syncytial Virus Variant in a Dairy Herd in Italy. Vet. Med. Sci. 2020, 6, 740–745. [Google Scholar] [CrossRef] [PubMed]
- Timurkan, M. O.; Aydin, H.; Sait, A. Identification and Molecular Characterisation of Bovine Parainfluenza Virus-3 and Bovine Respiratory Syncytial Virus-First Report from Turkey. J. Vet. Res. 2019, 63, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.; Yao, X.; Yang, Y.; Niu, C.; Zhao, Y.; Zhang, X.; Pan, R.; Jiang, X.; Xiaobo, S.; Qiao, X.; Guan, X.; Xu, Y. Isolation, Identification, and Phylogenetic Analysis of Subgroup III Strain of Bovine Respiratory Syncytial Virus Contributed to Outbreak of Acute Respiratory Disease among Cattle in Northeast China. Virulence 2021, 12, 404–414. [Google Scholar] [CrossRef] [PubMed]
- Langedijk, J. P.; Meloen, R. H.; Taylor, G.; Furze, J. M.; van Oirschot, J. T. Antigenic Structure of the Central Conserved Region of Protein G of Bovine Respiratory Syncytial Virus. J. Virol. 1997, 71, 4055–4061. [Google Scholar] [CrossRef]
Figure 1.
Primer positions and PCR amplicons for BRSV G Gene analysis. All 51 field isolates and four vaccines were amplified using the primer set VG1 and VG4. To enhance the precision of the 5' end sequence of the G gene, 11 samples were further analyzed using a second PCR with an additional forward primer: FNew and VG4.
Figure 1.
Primer positions and PCR amplicons for BRSV G Gene analysis. All 51 field isolates and four vaccines were amplified using the primer set VG1 and VG4. To enhance the precision of the 5' end sequence of the G gene, 11 samples were further analyzed using a second PCR with an additional forward primer: FNew and VG4.
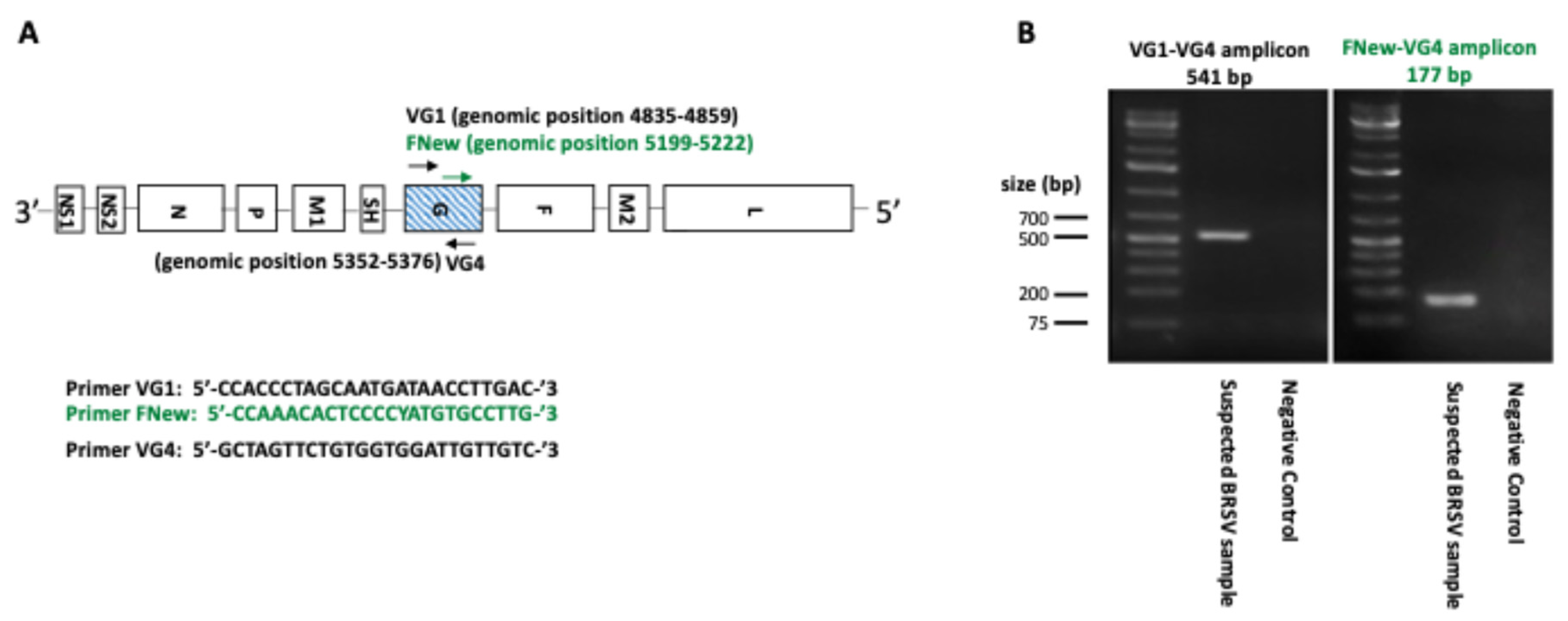
Figure 2.
Maximum Likelihood phylogenetic tree of BRSV G gene partial sequences. All 5 field isolates and 4 vaccine strains were sequenced and trimmed down to a 418 bp segment of the BRSV glycoprotein G ORF and aligned with reference samples from subgroup I-VIII (marked with an asterisk) using ClustalW. Designations at the ends of the branches refer to subgroups. Bootstrap values ≥70 (1000 repetitions) are indicated by the branch roots.
Figure 2.
Maximum Likelihood phylogenetic tree of BRSV G gene partial sequences. All 5 field isolates and 4 vaccine strains were sequenced and trimmed down to a 418 bp segment of the BRSV glycoprotein G ORF and aligned with reference samples from subgroup I-VIII (marked with an asterisk) using ClustalW. Designations at the ends of the branches refer to subgroups. Bootstrap values ≥70 (1000 repetitions) are indicated by the branch roots.
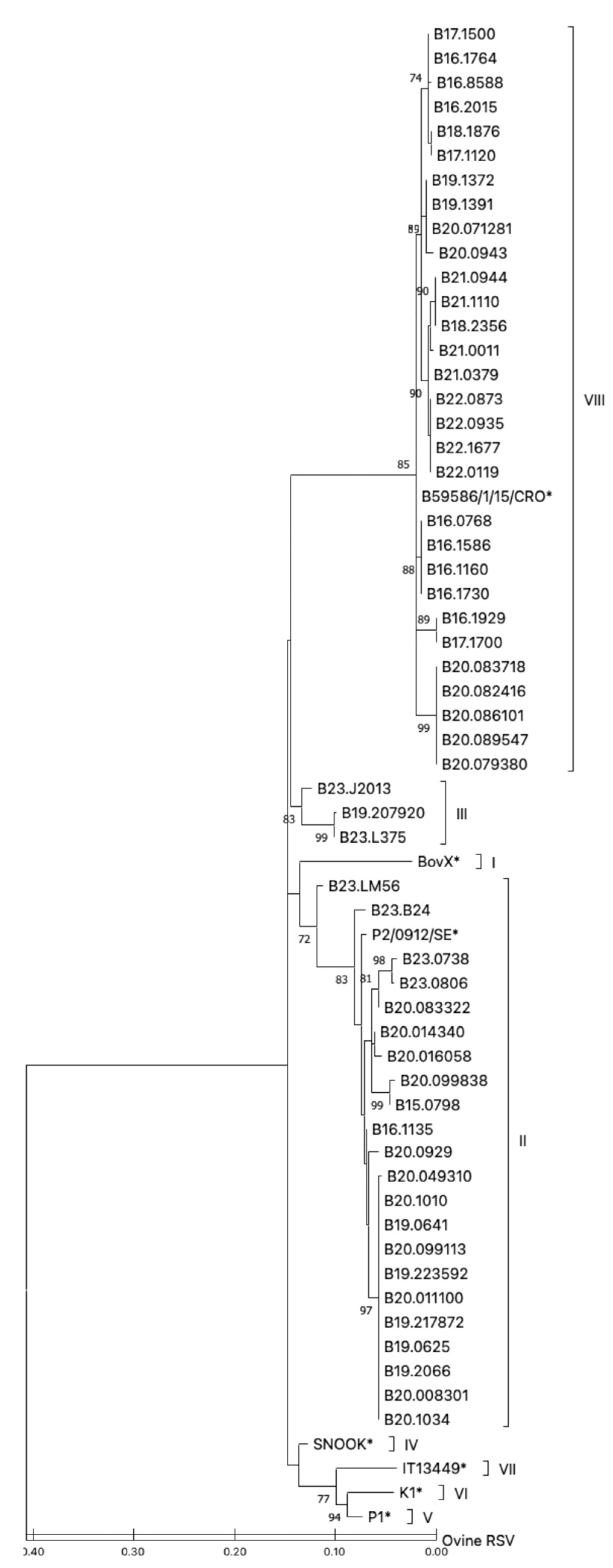
Figure 3.
Alignment of deduced amino acids 151 to 198 of the G gene protein of BRSV isolates. The central conserved region of the extracellular domain of BRSV glycoprotein G (positions 159 to 186), considered to be a major immunodominant area has been boxed. The strains were grouped into subgroups (I to VIII) shown on the left side.
Figure 3.
Alignment of deduced amino acids 151 to 198 of the G gene protein of BRSV isolates. The central conserved region of the extracellular domain of BRSV glycoprotein G (positions 159 to 186), considered to be a major immunodominant area has been boxed. The strains were grouped into subgroups (I to VIII) shown on the left side.
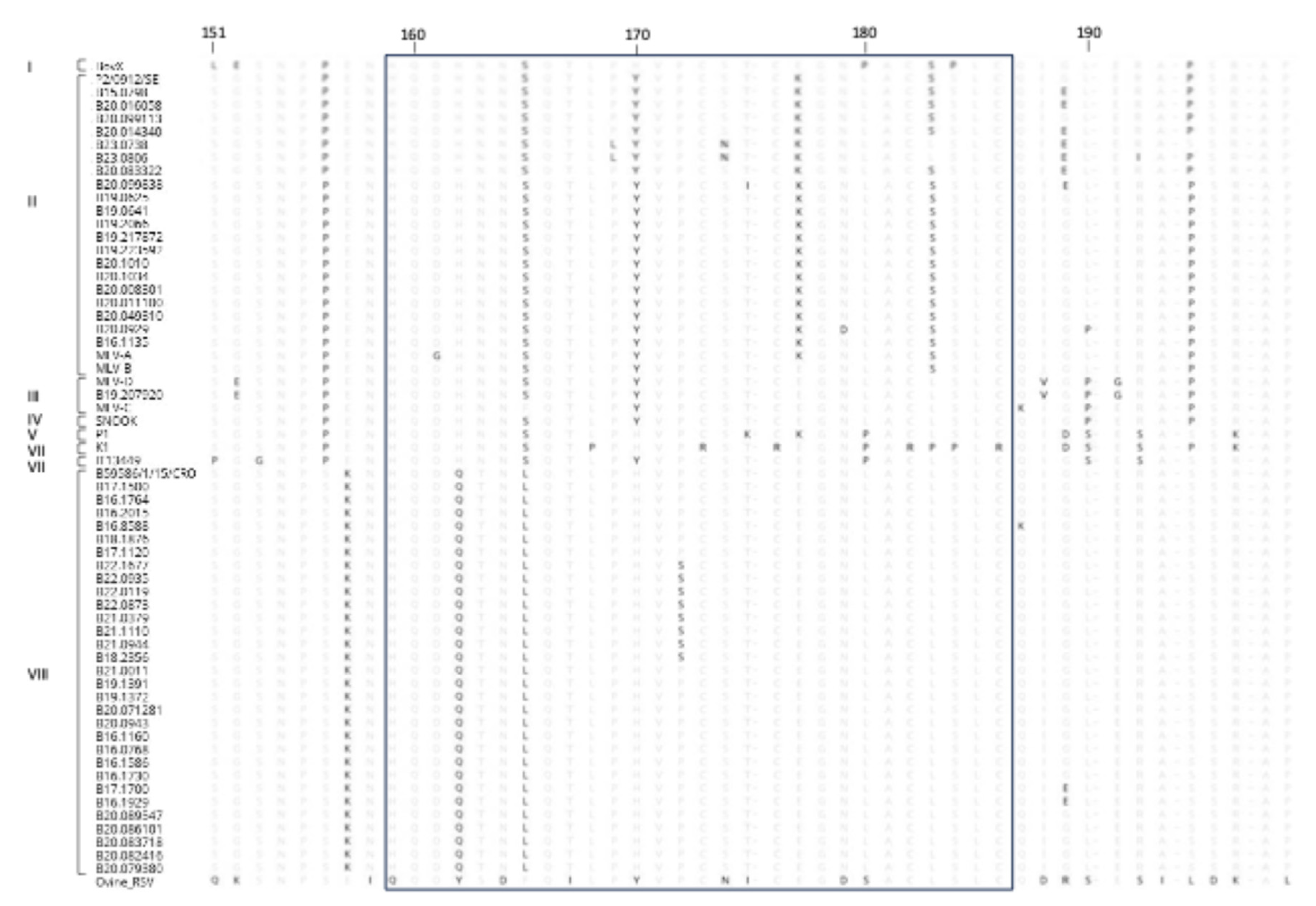

Table 1.
Samples analyzed for the study of bovine respiratory syncytial virus in Wallonia (Belgium).
Table 1.
Samples analyzed for the study of bovine respiratory syncytial virus in Wallonia (Belgium).
| Subgroup | Isolate name | Accession number | Year/month | Tissue | Age (months) | Vaccination |
|---|---|---|---|---|---|---|
| II | B15.0798 | PP538078 | 2015/11 | Lung | 6 | No |
| II | B16.1135 | PP538080 | 2016/01 | Lung | 2.5 | Yesb |
| II | B19.2066 | PP538098 | 2019/03 | Lung | 18 | No |
| II | B19.0641 | PP538095 | 2019/12 | Lung | 6 | Yesb |
| II | B19.0625 | PP538094 | 2019/12 | Lung | 5 | NK |
| II | B19.217872 | PP538100 | 2019/11 | Lung | 16 | Yesb |
| II | B19.223592 | PP538101 | 2019/12 | Lung | 8 | No |
| II | B20.0929 | PP538102 | 2020/02 | Lung | 4 | NK |
| II | B20.1010 | PP538104 | 2020/02 | Lung | 7 | Yesb |
| II | B20.1034 | PP538105 | 2020/02 | Lung | 6 | No |
| II | B20.008301 | PP538106 | 2020/01 | Lung | 1 | Yesa |
| II | B20.016058 | PP538109 | 2020/01 | Lung | 4 | Yesb |
| II | B20.011100 | PP538107 | 2020/01 | Lung | 4 | No |
| II | B20.014340 | PP538108 | 2020/01 | Lung | 1.5 | Yesb |
| II | B20.049310 | PP538110 | 2020/03 | Lung | 4 | No |
| II | B20.083322 | PP538114 | 2020/04 | Lung | 1.5 | Yesb |
| II | B20.099113 | PP538118 | 2020/05 | Lung | 1.5 | Yesb |
| II | B20.099838 | PP538119 | 2020/05 | Lung | 3 | No |
| II | B23.0738 | PP538128 | 2023/02 | Lung | 8 | NK |
| II | B23.0806 | PP538129 | 2023/02 | Lung | 19 | Yesb |
| II | MLV-A | PP538130 | 2021/04 | Vaccine | ||
| II | MLV-B | PP538132 | 2021/04 | Vaccine | ||
| III | B19.207920 | PP538099 | 2019/11 | Lung | 7 | Yesb |
| III | MLV-C | PP538131 | 2021/04 | Vaccine | ||
| III | MLV-D | PP538133 | 2021/04 | Vaccine | ||
| VIII | B16.1160 | PP538081 | 2016/01 | Lung | 5 | NK |
| VIII | B16.1586 | PP538083 | 2016/03 | Lung | 3 | No |
| VIII | B16.1730 | PP538084 | 2016/04 | Lung | 2 | NK |
| VIII | B16.1764 | PP538085 | 2016/04 | Lung | 9 | No |
| VIII | B16.1929 | PP538086 | 2016/05 | Lung | 2 | No |
| VIII | B16.8588 | PP538088 | 2016/05 | Lung | 3 | Yesb |
| VIII | B16.2015 | PP538087 | 2016/05 | Lung | 3 | No |
| VIII | B16.0768 | PP538079 | 2016/11 | Lung | 4 | NK |
| VIII | B17.1500 | PP538090 | 2017/02 | Lung | 9.5 | Yesb |
| VIII | B17.1700 | PP538091 | 2017/03 | Lung | 3 | No |
| VIII | B17.1120 | PP538089 | 2017/12 | Lung | 5.5 | No |
| VIII | B18.1876 | PP538092 | 2018/04 | Lung | 4 | Yesb |
| VIII | B18.2356 | PP538093 | 2018/06 | Lung | 9 | No |
| VIII | B19.1372 | PP538096 | 2019/01 | Lung | 14 | No |
| VIII | B19.1391 | PP538097 | 2019/01 | Lung | 6 | NK |
| VIII | B20.0943 | PP538103 | 2020/02 | Lung | 1 | No |
| VIII | B20.071281 | PP538111 | 2020/04 | Lung | ||
| VIII | B20.079380 | PP538112 | 2020/04 | Nasal swab | 2 | Yesa |
| VIII | B20.082416 | PP538113 | 2020/04 | Nasal swab | 4.5 | No |
| VIII | B20.083718 | PP538115 | 2020/04 | Lung | 0.5 | NK |
| VIII | B20.086101 | PP538116 | 2020/04 | Nasal swab | ||
| VIII | B20.089547 | PP538117 | 2020/05 | Lung | 5 | Yesa |
| VIII | B21.0944 | PP538122 | 2021/02 | Lung | 4 | Yesa |
| VIII | B21.1110 | PP538123 | 2021/03 | Lung | 4.5 | Yesa |
| VIII | B21.0011 | PP538120 | 2021/09 | Lung | 8 | Yesa |
| VIII | B21.0379 | PP538121 | 2021/11 | Lung | 8 | Yesb |
| VIII | B22.0873 | PP538125 | 2022/02 | Lung | 2 | No |
| VIII | B22.0935 | PP538126 | 2022/02 | Lung | 1.5 | No |
| VIII | B22.1677 | PP538127 | 2022/09 | Lung | 12 | Yesb |
| VIII | B22.0119 | PP538124 | 2022/09 | Lung | 8 | No |
Identification country/year of isolation/number, B = Belgium, aCalf vaccinated with MLV-A or MLV-B (subgroup II), Calf vaccinated with MLV-C or MLV-D (subgroup III), NKNot known.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



