Submitted:
29 January 2025
Posted:
30 January 2025
You are already at the latest version
Abstract
Nephrin is an essential constituent of the slit-diaphragm of the kidney filtering unit. Loss of nephrin expression leads to protein leakage into the urine, one of the hallmarks of kidney damage. Autoantibodies against nephrin have been reported in patients with minimal change disease and recurrent focal segmental glomerulosclerosis. Understanding the mechanism of nephrin loss may help improve or lead to the development of a novel treatment strategies.
In this study we demonstrated the important function of miR-204-5p expression on the protection of nephrin from anti-nephrin antibodies present in nephrotoxic serum (NTS). In addition, we identified that the aspartyl protease cathepsin D is one enzyme that may be involved in nephrin enzymatic degradation and that cathepsin D is a direct target of miR-204-5p gene regulation. The regulation of miR-204-5p expression was determined to be regulated by the long non-coding RNA Josd1-ps. In an NTS in vivo animal model, treatment with the pan aspartic proteases inhibitor Pepstatin A ameliorated the renal damage. Finally, we showed that the expression of miR-204-5p had a nephrin protecting function in vitro. Developing a method of delivery of miR-204-5p specifically to podocytes in vivo may provide a novel method of nephroprotection against nephrin autoantibodies.
Keywords:
Podocytes
; nephrin
; miR-204-5p
; nephrotoxic serum
; lncJosd1-ps
; Pepstatin A
; autoantibodies
1. Introduction
Podocytes are key constituents of the glomerular filtration barrier (GFB) and podocyte loss leads to progressive and often irreversible decline in kidney function. The filtration barrier, which prevents leakage of proteins and molecules larger than the size of albumin into the urine, is a complex structure dependent on the correct localization of several structural and signaling proteins [1]. Nephrin, a 150 kDa trans-membrane protein and member of the immunoglobulin superfamily, is one of the most important components of the GFB. Loss of functional nephrin in podocytes leads to significant proteinuria and irreversible renal decline [2]. Nephrin, however, can also be lost or mislocalized in acquired forms of kidney disease, particularly in immune mediated kidney diseases [3,4]. A recent study also found the presence of antinephrin autoantibodies in a significant proportion of patients with minimal change disease [5]. Antinephrin autoantibody levels correlated with disease activity, suggesting that these autoantibodies are pathogenic. In addition, immunization of mice with the ectodomain of murine nephrin resulted in nephrin autoantibody formation and development of significant proteinuria together with down-regulation of proteins necessary to maintain the filtration barrier. The mechanisms underlying how nephrin autoantibodies result in nephrin degradation and damage to the filtration barrier remain unknown.
MicroRNAs (miRNAs) are 18-22 nucleotide noncoding RNAs that that regulate gene expression by binding of a “seed” sequence in the miRNA to the 3’ or on some occasions the 5’ UTR of a target mRNA, thereby inducing degradation or transcriptional repression of the target [6,7]. Of note is that small changes in microRNA expression can have significant effects on their target genes. Previous studies have shown that a microRNA miR-204 which is expressed in the kidney is protective in hypertensive and diabetic kidney disease [8]. In addition, several studies have shown that decreased expression of miR-204 exacerbates kidney injury in various animal models of renal diseases [9].
In this study, we evaluated whether miR-204 is protective in an in vitro model of podocyte stress induced by treating cultured murine podocytes with nephrotoxic serum (NTS) which contains antinephrin antibodies. We show that after NTS treatment, miR-204-5p overexpression protects nephrin from enzymatic degradation by cathepsin D, thereby preserving nephrin expression. We also find that lysosomal-associated membrane protein 1 (LAMP1) and cathepsin D are direct miR-204-5p targets. In addition, we demonstrate that the long non-coding RNA Josd1-ps regulates miR-204-5p function. We also show that induction of nephrotoxic serum nephritis, an in vivo model of immune mediated kidney disease, results in decreased kidney expression of miR-204 and that proteinuria and histologic damage can be significantly ameliorated by treatment with Pepstatin A, an inhibitor of cathepsin D. Taken together, our results suggest that after an immune mediated insult miR-204 protects nephrin from cathepsin D mediated degradation.
2. Materials and Methods
2.1. Reagents
Nephrin antibody (R&D Systems Minneapolis, MN, USA, cat# AF3159, lot# CBIK0422122). Neph1 antibody (NovusBio Centennial, CO, USA, cat# NBP3-03509, lot# 0107180201). Podocin antibody (Invitrogen ThermoFisher, Waltham, MA, USA, cat#P A5-37904, lot# ZD4291857). LAMP1 antibody (abcam, Boston, MA, USA, cat# AB208943, lot# 1015275-34). Actin antibody (SigmaAldrich, St. Louis, MO, USA, cat# A1978, lot# 109M4849V), C5b-9 antibody (bioorbyt, Durham, NC, USA, cat# orb100655, lot# RB70). Alexa-635 phalloidin (Invitrogen ThermoFisher, Waltham, MA, USA, cat# A34054, lot# 2127282). Cathepsin E antibody (NovusBio Centennial, CO, USA, cat# NB400-152SS, lot# B-6).
2.2. Cell Culture
Immortalized mouse podocytes, generated from C57BL/6J mice as described [10], were cultured from frozen stock in DMEM supplemented with 10% FBS and 50 U/mL penicillin and 50 μg/mL streptomycin and grown at 37 °C and 5 % CO2.
2.3. miRNA Transfection and NTS Treatment
MicroRNA-204-5p mimic, inhibitor and negative control sequences were purchased from (ThermoFisher, Waltham, MA, USA, mimic cat# 4464066, lot# ASO2LE3J; inhibitor cat# 4464084, lot# ASO2KMY0; control cat# 4464076, lot# ASO2JI01). The cells were transfected using HiPerFect (Qiagen, Hilden, Germany, cat# 301705, lot# 172028439) and 50 nM of each microRNA sequences mimic, inhibitory, or control were added to the cells dropwise and incubated for further 72 h at 37 °C. The podocytes were then treated with nephrotoxic serum (NTS) (Probetex San Antonio, TX, USA, cat# PTX-001S-Ms, lot# 612-3T) at 1:200 dilution and incubated overnight at 37 °C and 5% CO2. The next day the cells were washed with PBS and the cell lysates were collected using RIPA buffer (50 mM Tris-HCl pH 7.4, 150 mM NaCl, 1 mM EDTA, and 1% Triton X-100) supplemented with protease inhibitor cocktail (Roche, Mannheim, Germany, cat# 04693159001).
2.4. SDS-PAGE and Western Blot
The cell lysates concentration was estimated using Pierce BCA protein assay and 10 ug was resolved in 10% SDS-PAGE gel. The proteins were transferred onto a PVDF membrane and blocked with 5% skim milk in PBST. The primary antibodies were added at 1:1000 dilution prepared in blocking buffer and incubated overnight at 4 °C. The membranes were washed 3X with PBST for 30 min and anti-species secondary antibodies were added at 1:10000 dilution for 1 hour at RT. The membranes were washed again with PBS for 30 min and developed using Immobilon Forte Western Blot Substrate (Millipore, Burlington MA, USA, cat# WBLUF0100, lot# 221543) and exposed to an X-Ray film.
2.5. Immunofluorescence
Mouse podocytes were seeded on 35 mm glass bottom MatTek (Ashland, MA) dishes. The cells were transfected with 50 nM miR 204 mimic or inhibitory sequences or control sequence using HiPerFect (Qiagen, Hilden, Germany, cat# 301705, lot# 172028439) for 72 hours at 37 °C and 5% CO2. The cells were then fixed with 4% paraformaldehyde (Electron Microscopy Sciences, Hatfield, PA) and stained with nephrin and phalloidin-488. The images were obtained using Olympus 1X81 (Tokyo, Japan) confocal microscope available at the Advanced Light Microscopy Core Facility at the Colorado University Anschutz Medical Campus. The images were deconvoluted using Huygens Essential 23.10 software (Scientific Volume Imaging, Hilversum, The Netherland) under the conservative setting.
2.6. Lentivirus-Mediated LncJosd1-ps Overexpression and Antisense Oligos Knockdown
The long non-coding RNA Josd1-ps (GenBank: NG_065344.1) full sequence was cloned into the lentivirus plasmid VVPW (kind gift from Gabriele Luca Gusella, Icahn School of Medicine Mount Sinai Hospital, NY City, NY, USA) and validated by sequencing. The VVPW plasmid along with the packaging plasmid psPAX2 (Addgene, Watertown, MA, USA) and the envelope plasmid pCMV-VSV-G (Addgene) were combined with FUGENE HD (Promega, Madison, WI, USA, cat# E2311, lot# 0000553944) transfection reagent at the 3:2:1 ratio and added to ~70% confluent 293T cells. The cells supernatants were collected after 48 and 72 hours post transfection and validated for LncJosd1-ps overexpression by qPCR using RNA isolated from cells infected with 20% lentivirus supernatants. The mouse podocytes were infected with LncJosd1-ps lentivirus particles in addition to 8 μg/mL polybrene. The cells were used in experiments after 72 hours of infection. LncJosd1-ps knockdown was performed using antisense oligos (ASO). The knockdown was validated using two different ASOs at 1nM delivered using HiPerFect (Qiagen, Hilden, Germany, cat# 301705, lot# 172028439) and the experiments were performed 72 hours after transfection.
2.7. Cytoplasmic and Nuclear RNA Extraction
The RNA from the cytoplasm and the nucleus of mouse podocytes were isolated using the NORGEN BIOTEK CORP. kit (Thorold, ON, Canada, cat# 21000, lot# 605095) as per manufacturer’s instructions using cells growing in monolayer protocol.
2.8. Quantitative PCR and microRNA Analysis
The RNA was extracted using the TRIzol (Life technologies, Carlsbad, CA, USA, cat# 15596026, lot# 349906) method and quantified using NanoDrop 2000 (ThermoScientific). One microgram of total RNA was reversed transcribed into cDNA using LunaScript (NE Biolabs, Ipswich, MA, USA, cat# E3010L, lot# 10175742). SYBR Green master mix (ThermoFisher, Waltham, MA, USA, cat# A25742, lot# 2905743) was used and the PCR reaction was amplified using BioRad SFX96 Thermocycler. The antisense oligos (ASO) were purchased from Integrated DNA Technologies (IDT) (Morrisville, NC, USA). Gene expression was normalized to Valosin containing protein (VCP) and asparagine-linked protein homolog 9 (ALG9). The results are reported as log10 (2-DDCt). The primers were ordered from IDT (Coralville, IA, USA) and the list of primers and anti-sense oligos used in this study are listed in table S1 of supplementary data. The microRNA RT-PCR was performed the miRCURY LNA system (Qiagen Hilden, Germany). miRCURY LNA RT kit (cat# 339340, lot# 77203282). miRCURY LNA SYBR Green PCR kit (cat# 339346, lot# 77203504). miRCURY LNA miRNA PCR Assay primers: miR-204-5p (cat# YP00206072, lot# 20304364-2), miR-16-5p (cat# YP00205702, lot# 40501706-2).
2.9. Protease Inhibitors Assay
Mouse podocytes transfected with miR-204 inhibitory sequence were loaded with the following protease inhibitors: 0.25 mM AEBSF (SigmaAldrich, St. Louis, MO, USA, cat# SBR00015, lot# 0000188562), 1 mM Pepstatin A (SigmaAldrich, St. Louis, MO, USA, cat# SAE0154, lot# P318, lot# 0000144172), 20 mM E64 (SigmaAldrich, St. Louis, MO, USA, cat# SAE0154, lot# 0000105689), and 1X protease inhibitor cocktail (Roche, Mannheim, Germany, cat# 04693159001) for 60 min at 37 °C and 5 % CO2. NTS was added at 1:200 dilution and the cells were incubated overnight. The next day the cells were washed 2X with PBS and cell lysates was collected and resolved using SDS-PAGE and Western blot.
2.10. Cathepsin D ELISA
Cathepsin D was detected using mouse cathepsin D SimpleStep ELISA kit (abcam Waltham, MA, USA, cat# ab239420) as per manufacturer’s instructions, and the plate was read at 450 nm using BioTek Synergy 2 plate reader.
2.11. Nephrin Digestion
Nephrin digestion with cathepsin D was carried out as described previously [11]. Briefly, 10 μg of recombinant mouse nephrin (R & D Systems Minneapolis, MN, USA, cat# 3159-NN-050, lot# NZA0422021) was incubated with 10 μg of recombinant mouse cathepsin D (R & D Systems Minneapolis, MN, USA, cat# 1029-AS-010, lot# FVE0122021) in 100 mM sodium citrate, 200 mM NaCl buffer (pH 3.5) for 3 hours at 37 °C.
2.12. Silver Stain
The digested nephrin was resolved on an SDS-PAGE gel and stained using silver stain (ThermoFisher Waltham, MA, USA, silver reagent cat# 1851130, lot# VB297147, stabilizer base reagent cat# 1851190, lot# VC297149, and reducer aldehyde reagent, cat# 1851170, lot# VB297148) as per manufacturer’s instructions.
2.13. Site Directed Mutagenesis
The mutation of miR-204-5p binding sites in LAMP1, cathepsin D, and Josd1-ps was performed according to New England Biolabs Q5 ® site-directed mutagenesis kit (Ipswich, MA, USA, cat# E0554S, lot# 10164452) according to the manufacturer’s instructions. The primers used to insert the mutated sequence into the lentivirus plasmids were designed using NEBaseChanger™ tool available at New England Biolabs website. The resulting mutant plasmids were validated by sequencing that was performed by Quintara Biosciences (Denver, CO, USA).
2.14. Luciferase Assay
LAMP1, cathepsin D 3’UTR and Josd1-ps sequences were cloned into pIS2 luciferase plasmid (a gift from David Bartel, plasmid 12177; Addgene) and validated by sequencing. Mouse podocytes were transfected with 1 ug of pIS2 plasmid using ViaFect transfection reagent (Promega, Madison, WI, USA, cat# 4981, lot# 0000514926) as well as the cells were transfected with 50 nM miR control sequence or miR-204-5p mimic or inhibitor sequences using the transfection reagent HiPerFect (Qiagen, Hilden, Germany, cat# 301705, lot# 172028439). The luciferase activity was determined 72 h post transfection using the Renilla luciferase assay system (Promega, Madison, WI, USA, cat# E183A, lot# 0000537514) as described in the manufacturer’s product manual. The luciferase substrate’s background fluorescence was subtracted from all the samples readings. The luciferase assay data were normalized to protein concentrations (reporter activity/total protein).
2.15. Identification of miR-204 Long Non-Coding RNA Binding Partners
miR-204 interaction with long non-coding RNA was analyzed using RNA22 and miRCode prediction tools to identify potential targets with at least 6 nucleotides interacting with miR-204-5p seed region. Other prediction tools used to predict miR-204 target genes including TargetScan and miRDB.
2.16. In Vivo Experiments
All animal experiments were approved by the University of Colorado Institutional Animal Care and Use Committee (IACUC) and performed according to the PHS Policy on Humane Care and Use of Laboratory Animals and IACUC.
The nephrotoxic serum nephritis mouse model was performed as previously described [12]. Male C57Bl/6 mice 8-12 weeks old were injected with 100 μL of sheep anti-rat glomerular basement membrane antibodies (NTS) purchased from Probetex Inc. (Probetex San Antonio, TX, USA, cat# PTX-001S-Ms, lot# 612-3T) via tail vein. The mice were euthanized after seven days. Blood and urine were collected prior to euthanasia for blood urea nitrogen (BUN) and proteinuria analysis. For Pepstatin A experiment, following the NTS injection Pepstatin A was administered as described previously [13]. Briefly, male C57Bl/6 mice received 20 mg/kg Pepstatin A by intraperitoneal injection on day 1, 3, and 5 after the NTS injection and the mice were euthanized on day 7. Control mice received 100 μL of ethanol on day 1, 3 and 5 post NTS delivery and the mice were euthanized on day 7.
2.17. MTT Cell Toxicity Assay
Immortalized mouse podocytes were seeded in a 96-well tissue culture plate and grown at 37 °C and 5% CO2 overnight. The next the cells were transfected with miR-204-5p mimic, inhibitory sequences or a negative control sequence and treated with NTS as described in section 2.3. MTT (Thiazolyl Blue Tetrazolium Bromide, Tocris cat# 298-93-1, lot# 8A/261332) was added at 10 μL/well of 5 mg/mL and the plate was incubated for 4 hours. The medium was removed and the formazan crystals were solubilized with 100 μL/well dimethyl sulfoxide (DMSO) and the plate was read using absorbance of 570 nm BioTek Synergy 2 plate reader.
2.18. Statistics
The statistical analysis was performed using GraphPad Prism 10 for macOS, Version 10.4.0 (Boston, MA, USA). The datasets were analyzed using one-way ANOVA multiple comparisons to compare the mean of each column with the mean of every other column. P < 0.05 was considered significant.
3. Results
3.1. Nephrin Is a Target of Nephrotoxic Serum when miR-204-5p Is Inhibited
When conditionally immortalized mouse podocytes were treated with NTS 1:200 dilution for varying amounts of time there were no changes in nephrin expression (Figure S1). However, when the mouse podocyte cell line was transfected with miR-204-5p mimic, inhibitory, or control sequences and then treated with 1:200 dilution of NTS overnight (~16 hours) nephrin expression changed. The cell lysates assayed by SDS-PAGE and Western blot for the podocyte markers nephrin, neph1, and podocin showed that treatment of podocytes with miR-204-5p inhibitory sequence and NTS resulted in a significant decrease in nephrin expression (Figure 1 A), but did not affect the expression levels of neph1 and podocin (Figure 1 B, C, & D). The transfection efficiency of miR-204-5p (mimic & inhibitory) and control sequences were analyzed by real-time PCR (Figure 1 E). In addition, we demonstrate that the nephrotoxic serum contains antibodies against nephrin. Recombinant mouse nephrin was resolved on an SDS-PAGE gel and transferred to a PVDF membranes under native and/or reduced and denatured conditions. The membranes were then probed using the NTS serum followed by anti-sheep IgG. As a result, nephrin was detected on the membrane under native as well as reduced & denatured gels (Figure 1 F). Since the nephrotoxic serum contains anti-nephrin antibody we tested whether the complement system is activated. Using immunofluorescence stain we detected some C5b-9 present in the cytoplasm of control cells whereas the cells that were treated with NTS C5b-9 complex was observed on the cell surface as well as inside the cells (Figure S2 A) suggesting that the complement system is involved in podocyte stress induced by NTS treatment. However, complement system activation did not result in cellular death (Figure S2 B). The effect of NTS on miR204 expression is shown in (G). Representative images show the expression of nephrin in mouse podocytes under normal culturing conditions (NT) and in cells treated with NTS and one of the miRNA sequences: control (miR C), miR204 mimic (Mimic) and miR204 inhibitor (Inhibitor).
Immortalized mouse podocytes were treated with miRNA control, miR-204-5p mimic or inhibitory sequences or left untreated prior to the addition of 1:200 nephrotoxic serum NTS. The cell lysates were analyzed by Western blot (A) and probed for the podocyte’s markers nephrin, neph1, and podocin. Actin was used as a loading control. The blots were quantified in B, C & D. The miR-204-5p sequences expressions were analyzed by real-time PCR and the data were normalized to the expression of miR-16-5p (E). The presence of anti-nephrin antibodies in NTS were analyzed by Western blot (F) under native and reduced and denatured conditions. The immunofluorescence images in (G) show mouse podocytes transfected with miR-204-5p mimic, inhibitor or control sequences prior to NTS treatment. The images were obtain using Stedycon Abberior (STED) and Olympus 1X81 confocal microscope and 100× objective lens. The scale bar represents 10 μm. The experiments were repeated 4 times (n = 4) and p values < 0.05 were considered significant.
3.2. Nephrin Is Degraded by an Aspartyl Protease
To determine the nature of the enzyme responsible for nephrin degradation, podocytes were transfected with miR-204-5p mimic and/or inhibitory sequences or control and then pretreated with 1X protease inhibitor cocktail before the addition of the nephrotoxic serum at 1:200 dilution overnight. The next day, cell lysates were collected and assayed by SDS-PAGE and Western blot. As shown in Figure 2 A & B, the decrease in nephrin expression after miR-204 inhibition in the presence of NTS was abrogated suggesting that nephrin is degraded by an unknown protease that can be blocked by a protease inhibitor. In order to identify the enzyme responsible for nephrin degradation, we transfected mouse podocytes with miR-204-5p inhibitor and after 72 hours the cells were treated with various protease inhibitors including the pan-cysteine protease inhibitor E64 (20 μM), the aspartyl protease inhibitor Pepstatin A (1 μM), the serine protease inhibitor AEBSF (250 μM), or DMSO. Thereafter, the cells were treated with nephrotoxic serum overnight after which the cell lysates were collected and assayed by SDS-PAGE and western blot. Treatment with E64, AEBSF or DMSO did not rescue nephrin from degradation. However, the aspartyl protease inhibitor Pepstatin A protected nephrin to the same level of expression observed in the non-treated cells (Figure 2 C & D), indicating that nephrin is degraded by an aspartyl protease.
Immortalized mouse podocytes were transfected with miR-204-5p mimic, inhibitor or control sequences and treated with protease inhibitor cocktail before treatment with NTS. As shown by the Western blot (A) and quantitation (B) protease inhibition prevented NTS-induced degradation of nephrin even when miR-204 was inhibited. Mouse podocytes treated with miR-204-5p inhibitory sequence and various enzymes inhibitors before NTS treatment showed that the aspartic acid protease inhibitor Pepstatin A rescued nephrin form degradation (C) and Western blot quantitation (D). The experiments were repeated 4 times (n = 4). P values < 0.05 were considered significant.
3.3. miR-204-5p Regulates the Expression of LAMP1 and Cathepsin D
We searched in silico for candidate aspartyl proteases that could potentially degrade nephrin and could also be a direct target of miR-204-5p. Using TargetScan we identified cathepsin D as a candidate aspartyl protease. We also identified lysosomal-associated membrane protein 1 (LAMP1) as a miR-204-5p target. We chose cathepsin E as a control since it is also an aspartyl protease but not predicted to be a target of miR-204-5p. Mouse podocytes were transfected with miR-204 mimic, inhibitory and control sequences, cell lysates were extracted and resolved using SDS-PAGE and Western blot. As Figure 3 (A, B) shows, the expression of nephrin was not affected in the presence of miR-204 mimic or inhibitory sequences in the absence of NTS. Interestingly, the overexpression of miR-204-5p significantly decreased the expression of LAMP1 (Figure 3 A & C), indicating that LAMP1 is a target of miR-204-5p. However, the expression of cathepsin E was not affected (Figure 3 A & D). Since cathepsin D expression could not be detected by Western blot we determined the effect of miR-204-5p overexpression or inhibition on cathepsin D by ELISA. Cathepsin D expression was significantly down regulated in podocytes transfected with miR-204-5p mimic sequence, indicating that cathepsin D is a target of miR-204-5p. Inhibition of miR-204-5p slightly increased cathepsin D expression but it was not statistically significant (Figure 3 E). In order to validate the degradation of nephrin by cathepsin D, we incubated recombinant mouse nephrin in the presence or absence of mouse cathepsin D and the reactions were resolved on an SDS-PAGE gel and stained with silver stain. Nephrin samples incubated with cathepsin D showed several bands of nephrin fragments, indicating that cathepsin D can degrade nephrin (Figure 3 F).
3.4. LncJosd1-ps Regulates the Expression of miR-204-5p
MicroRNAS can themselves be regulated by long noncoding RNAs (lncRNAs) and binding of lncRNAs to their target miRNAs can induce degradation and thus decreased expression of the target miRNA. To determine whether miR-204-5p is itself a target of lncRNAs we screened potential candidates using several prediction tools including DIANA, RNA22, and miRCode. Several lncRNAs were identified and screened for expression in mouse kidneys isolated from animals treated with NTS. Amongst all the screened lncRNAs, only lncJosd1-ps was upregulated when miR-204-5p expression was reduced (Figure 6 A & B and Figure S3). To further validate the potential lncJosd1-ps and miR-204-5p interaction, lncJosd1-ps was cloned into the lentivirus plasmid VVPW and virus was produced. In addition, lncJosd1-ps antisense oligos (ASO) were designed to suppress lncJosd1-ps expression. As shown in Figure 4 A and B, LV Josd1-ps induced significant overexpression of lncJosd1-ps RNA in mouse podocytes assayed 72 hours post infection whereas ASO Josd1-ps induced significant down regulation of lncJosd1-ps RNA. As expected, LV Josd1-ps significantly reduced miR-204-5p expression whereas ASO Josd1-ps significantly increased miR-204-5p expression (Figure 4 C and D). We also determined the subcellular location of lncJosd1-ps and found that ~60% of lncJosd1-ps expression is in the nucleus and ~40% is in the cytoplasm (Figure 4 E).
3.5. Validation of miR-204-5p Interaction with LAMP1, Cathepsin D and Josd1-ps
The direct interaction between miR-204-5p and LAMP1, cathepsin D and Josd1-ps was determined by Renilla luciferase assay. The 3’UTRs of LAMP1 and cathepsin D as well as the entire Josd1-ps sequence were cloned into the pIS2 luciferase plasmid. MiR-204-5p binding site(s) was/were mutated using Q5 ® site-directed mutagenesis kit and specific mutant primers. Mouse podocytes were infected with luciferase plasmid carrying intact or mutant miR-204-5p binding site(s), followed by transfection with miR-204-5p mimic, inhibitory, or control sequences. Figure 5 A shows a schematic representation of LAMP1, cathepsin D and Josd1-ps nucleotide sequences that contain miR-204-5p binding sites. The interaction between miR-204-5p and LAMP1 3’UTR sequence is shown in Figure 5 B. The overexpression of miR-204-5p sequence significantly reduced luciferase activity indicating the binding of miR-204-5p to LAMP1 3’UTR and subsequent degradation of LAMP1. However, this effect was abrogated in cells that were infected with LAMP1 mutant plasmid. We identified two potential miR-204-5p binding sites within the cathepsin D 3’UTR region. Luciferase analysis of this interaction is shown in Figure 5 C and demonstrates that miR-204-5p binds preferably to site 2 where mutation of the nucleotide sequence of site 2 prevented miR-204-5p binding and did not affect the luciferase activity. However, the mutation of site 1 did not result in luciferase activity reduction indicating that site 2 is the main miR-204-5p binding site within cathepsin D 3’UTR. The double mutation of sites 1 and 2 completely abrogated miR-204-5p binding and the degradation of cathepsin D. The interaction of miR-204-5p and josd1-ps is shown in Figure 5 D. Again, two potential miR-204-5p binding sites within josd1-ps nucleotide sequence were identified. Mutation of site 1 and site 2 individually resulted in statistically insignificant reductions in luciferase activity as compared to the other treatments indicating miR-204-5p binding to both sites. As expected, the josd1-ps double mutant plasmid prevented miR-204-5p binding and the luciferase activity was restored to normal levels.
3.6. miR-204-5p Is Down Regulated in Mice Kidneys Treated with Nephrotoxic Serum
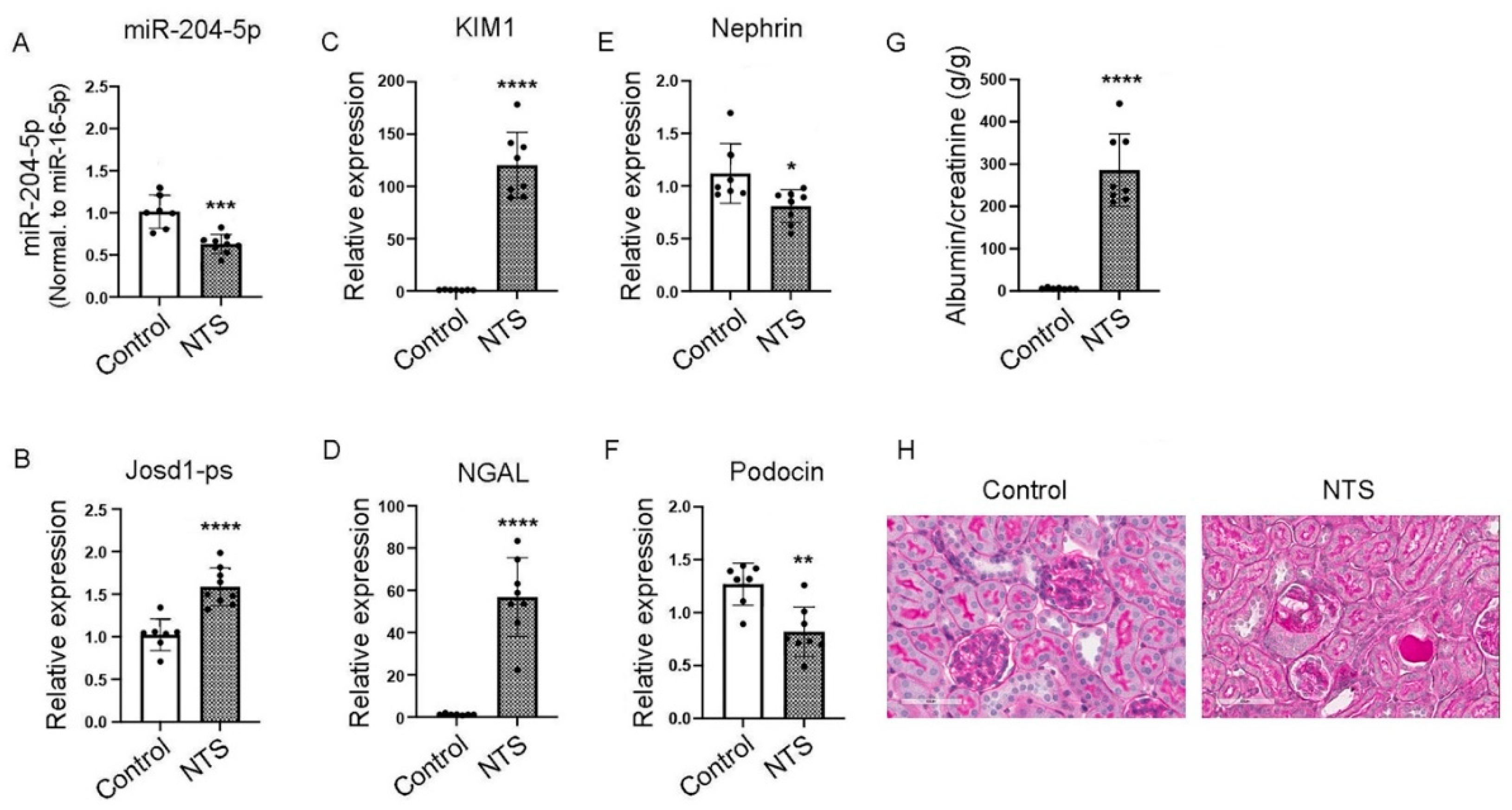
One week after treatment with nephrotoxic serum, real-time PCR showed a significant down regulation of miR-204-5p expression in mice treated with NTS as compared to control (Figure 6 A), whereas the expression level of josd1-ps was significantly increased (Figure 6 B). The kidney injury markers KIM1 and NGAL were up regulated significantly in NTS versus control mice (Figure 6 C & D). The podocyte markers nephrin and podocin were down regulated in the mice that received the NTS treatment compared to control (Figure 6 E & F). Albuminuria was also significantly increased in NTS treated mice compared to control (Figure 6 G). Histologic analysis also demonstrates increased glomerulosclerosis in NTS treated mice compared to control (Figure 6 H).
Figure 6.
The effects of NTS on kidney function in vivo. Male C57Bl/6 mice were injected with 100 μL NTS via tail vein and euthanized after 1 week. Kidneys were excised and total RNA was extracted and analyzed by real-time PCR for the expression levels of miR-204-5p (A), lncJosd1 (B), KIM1 (C), NGAL (D), nephrin (E) and podocin (F). Albuminuria was significantly increased in mice treated with NTS compared to control (G). Figure (H) shows glomerular damage induced by NTS (PAS stain, scale bar 50 µm). Seven to eight animals were included per group. P values < 0.05 were considered significant.
Figure 6.
The effects of NTS on kidney function in vivo. Male C57Bl/6 mice were injected with 100 μL NTS via tail vein and euthanized after 1 week. Kidneys were excised and total RNA was extracted and analyzed by real-time PCR for the expression levels of miR-204-5p (A), lncJosd1 (B), KIM1 (C), NGAL (D), nephrin (E) and podocin (F). Albuminuria was significantly increased in mice treated with NTS compared to control (G). Figure (H) shows glomerular damage induced by NTS (PAS stain, scale bar 50 µm). Seven to eight animals were included per group. P values < 0.05 were considered significant.
3.7. Pepstatin A Decreases Kidney Damage in NTS-Treated Mice by Preserving miR-204-5p Expression
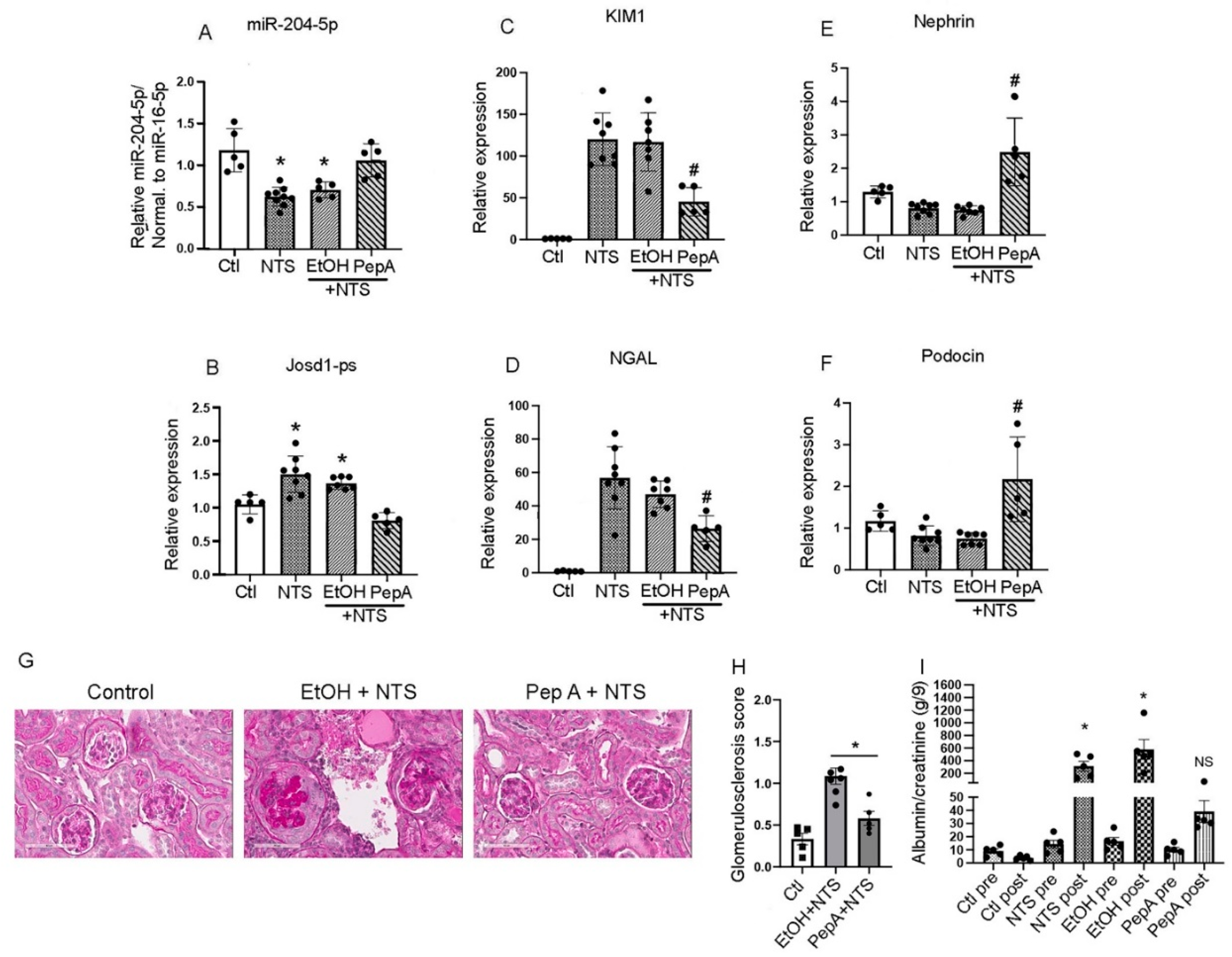
The effect of Pepstatin A on miR-204-5p expression and kidney function in vivo was determined. Mice received either Pepstatin A (20 mg/kg every other day for 3 doses) or vehicle alone (ethanol) post NTS injection. Analysis of mouse kidneys one week after NTS injection showed that Pepstatin A treatment protected miR-204-5p expression comparable to control mice whereas NTS, and ethanol + NTS groups had significantly reduced levels of miR-204-5p expression (Figure 7 A). In contrast, josd1-ps expression was increased in the NTS and ethanol + NTS groups whereas Pepstatin A treatment decreased josd1-ps expression (Figure 7 B). In addition, Pepstatin A treatment significantly reduced the expression of the kidney injury markers KIM1 and NGAL as compared to the mice groups that received NTS or ethanol + NTS (Figure 7 C & D). Nephrin and podocin were significantly upregulated in the mice treated with Pepstatin A prior to the administration of NTS as compared to the mice that received only NTS (Figure 7 E & F). Mice pretreated with Pepstatin A had significantly lower glomerulosclerosis scores as compared to the EtOH + NTS group (Figure 7 G and H) as well as a significant reduction in albuminuria compared to mice in the NTS, EtOH or NTS + EtOH groups (Figure 7 I). Taken together, these results indicate that Pepstatin A significantly reduces kidney damage induced by NTS by protecting miR-204-5p from degradation and reducing expression of josd1-ps.
4. Discussion
Podocyte injury is detrimental to proper kidney function and at the core of various glomerulopathies [2]. Identifying the different mechanisms that lead to podocyte injury aids in the development of novel therapeutic measures to treat podocytopathies.
Several podocyte antigens such as PLA2R1, THSD7A, NELL1, HTRA1, and nephrin are the primary targets of deleterious autoantibodies in glomerulonephritis patients [14,15,16,17,18]. Of all those antigens nephrin is an integral component of the slit diaphragm and is directly involved in preventing large proteins from leaking into the urine [19]. Various in vivo studies demonstrated the importance of miR-204 expression for the preservation of the kidney function. The reported mechanisms involve the decreased expression of certain long-noncoding RNAs that block miR-204 activity such as MALAT1 [20] and Kcnq1ot1 [21] or by the activation/deactivation of signaling pathways including HMX1 [22], Smad5 [23], NLRP3 inflammasome [21], and Fas/FasL [9]. However, a direct link between miR-204 function and podocytes has not been reported. In this study, we describe the mechanisms underlying miR-204-5p protection of nephrin after NTS-induced stress.
The nephrotoxic serum model is widely used as an in vivo model of acute and chronic glomerulonephritis [24]. However, the specific effect of NTS on podocytes in vitro is not clear. Given the reported function of miR-204 in protecting kidney function, we sought to explore the effect of miR-204 modulation on podocytes in the presence of NTS. Initial experiments with an NTS time course in a mouse podocyte cell line did not show a change in nephrin expression. However, when podocytes were transfected with miR-204-5p inhibitory sequence and treated with NTS nephrin expression was significantly decreased whereas levels of neph1 and podocin remained unchanged. This result suggests that miR-204-5p protects nephrin from the toxic effects of NTS. Interestingly, when podocytes were grown on a glass cover slip for immunofluorescence staining the effect of miR-204-5p inhibitory sequence and NTS was so drastic that the majority of cells did not survive and the remaining cells were almost devoid of nephrin. In contrast, the presence of miR-204-5p was protective at even a low level of expression in the cells that received microRNA control sequence and the miR-204-5p mimic sequence showed a strong protective effect on nephrin expression and the overall viability of the cells. Since the NTS contains anti-nephrin antibody, it suggested that the complement system is activated. Immunofluorescence staining of control and NTS-treated cells showed intracellular location of C5b-9 in control cells however the NTS-treated cells displayed surface as well as cytoplasmic C5b-9 locations. Notwithstanding the complement activation there was no detectable cell lysis even though C5b-9 deposition on the cell surface was observed. It is not surprising that podocytes can resist complement-mediated cell killing. Previous reports also showed that in vitro cultured podocytes can withstand complement activation in a mechanism that involves several complement regulatory molecules such as factor H, CD46, CD55, and CD59 that the podocytes possess [25,26].
The loss of nephrin in podocytes treated with NTS and miR-204-5p inhibitory sequence can be attributed to an enzymatic activity as we observed a rescue of nephrin expression when the cells were pretreated with a protease inhibitor cocktail prior to treatment with NTS and miR-204-5p inhibitory sequence. In the search for a potential enzyme group that is responsible for nephrin degradation we pretreated the podocytes with various enzyme inhibitors including: the cysteine proteases inhibitor E64, the aspartyl enzymes inhibitor Pepstatin A, and the serine proteases inhibitor AEBSF. Nephrin was only rescued by Pepstatin A. The results suggested the involvement of an aspartic enzyme(s), a group of proteases that include pepsins, renins and cathepsins [27]. Amongst this group of proteases, the cathepsins seemed more likely candidates to be involved in nephrin degradation. Screening the 3’UTR of the two cathepsin aspartic proteases cathepsin D and E, we identified cathepsin D as a potential target of miR-204-5p that was validated using a luciferase assay. The interaction between cathepsin D and nephrin was demonstrated by the cleavage of recombinant mouse nephrin by mouse cathepsin D. This result suggested that in the absence of miR-204-5p expression cathepsin D, under cellular stress such as that incurred by NTS, can act upon nephrin to reduce its expression. In line with our observation, in diabetic nephropathy nephrin is internalized inside endocytic vesicles mediated by dynein light chain 1 (DYNLL1) and eventual nephrin is degraded in the lysosome. In the absence of DYNLL1 expression endocytosed nephrin was recycled back to the plasma membrane [28]. Another mechanism of nephrin degradation may also involve the SEL1L-HRD1 protein complex of endoplasmic reticulum-associated degradation (ERAD) which is highly expressed in podocytes where nephrin is an endogenous substrate [29]. However, this degradation pathway was not investigated in this study.
The expression level of mir-204-5p was down regulated in mice treated with NTS suggesting that a long noncoding RNA may regulate miR-204-5p activity. Several lncRNA reportedly regulate miR-204-5p function including MALAT1, KCNQ1OT, and SPANXA1-OT1 [20,21,23] along with other potential in silico identified LncRNA partners of miR-204-5p were assayed for expression in mice treated with NTS. Amongst all the assayed lncRNAs only LncJosd1-ps was up regulated suggesting a potential interaction with miR-204-5p that merited further validation. No reports were found that described a function for lncJosd1-ps. We cloned lncJosd1 into a lentivirus vector and designed antisense oligos in order to manipulate its expression and determine whether it has any effect on miR-204-5p expression. The results clearly showed a link between lncJosd1-ps expression and that of miR-204-5p suggesting a potential binding partnership. Indeed, lncJosd1-ps overexpression led to miR-204-5p suppression and lncJosd1-ps inhibition increased miR-204-5p expression. This binding partnership was further validated and confirmed using a luciferase assay. The cellular location of lncJosd1-ps is important for its “sponging” of microRNA function. Although around 60 % of lncJosd1-ps transcripts are present in the nucleus, the remaining 40 % are available in the cytoplasm for a potential sponging activity.
There is a general agreement in the scientific literature that the expression of miR-204 is protective in various kidney injury models [8,9,20,21,22,23,30]. However, the nephroprotective function of miR-204 has never previously been linked to nephrin. Here we show a correlation between miR-204 and nephrin expression and ultimately kidney function. In the NTS animal model miR-204-5p expression was significantly downregulated which coincided with nephrin’s reduced expression. As a result, the renal injury was severe as measured by the elevated expressions of the kidney injury markers KIM1 and NGAL as well as the albumin/creatinine ratio with noticeable glomerular injury. In contrast, Pepstatin A treatment in vivo protected miR-204-5p and nephrin expression and overall kidney function. Inhibiting aspartyl proteases had a beneficial effect on kidney function after NTS treatment but other potential effects of inhibiting aspartyl proteases remain to be determined. It has proved very challenging to deliver microRNAs effectively to podocytes. However, once miRNAs can be delivered to podocytes with high efficiency, miR-204 remains an excellent candidate for potential renal therapy for glomerular diseases in which nephrin is degraded. Our findings are of special relevance given recent findings that anti-nephrin autoantibodies are present in a significant number of patients with minimal change disease [4]. However, the mechanisms underlying how these anti-nephrin autoantibodies lead to loss of nephrin from the slit diaphragm remain to be determined.
5. Conclusion
The data presented in this study showed the importance of the miR-204-5p expression in protecting nephrin from enzymatic degradation by the aspartic enzyme cathepsin D under stress conditions caused by the nephrotoxic serum. We demonstrated that cathepsin D and LAMP1 are direct targets of miR-204-5p which is in turn is regulated by the long noncoding RNA Josd1-ps. The inhibition of aspartyl proteases by Pepstatin A in vivo protected the kidney from the deleterious effect of NTS and preserved its function. Our results suggest that miR-204-5p is nephroprotective and devising a strategy to deliver this micro-RNA specifically to podocytes could have a therapeutic potential in the future.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
GH experiments design, data collection and analysis, and manuscript writing. JB experiments design and manuscript editing.
Funding
This study was supported by the National Institute of Health (NIDDK R01 DK104264).
Institutional Review Board
The study was conducted according to the guidelines of the Institutional Animal Care at the University of Colorado, and approved by the Institutional review Board of the University of Colorado (protocol code 00085, approved 08/09/2024).
Informed Consent
Not Applicable.
Data availability
All data would be made available to all interested scientists upon request.
Acknowledgements
Not Applicable
Conflict of Interest
GH and JB have nothing to declare.
References
- Daehn, I.S.; Duffield, J.S. The glomerular filtration barrier: a structural target for novel kidney therapies. Nature Reviews Drug Discovery 2021, 20, 770–788. [Google Scholar] [CrossRef]
- Hengel, F.E.; et al. Autoantibodies Targeting Nephrin in Podocytopathies. N Engl J Med 2024, 391, 422–433. [Google Scholar] [CrossRef] [PubMed]
- Doublier, S.; et al. Nephrin redistribution on podocytes is a potential mechanism for proteinuria in patients with primary acquired nephrotic syndrome. Am J Pathol 2001, 158, 1723–1731. [Google Scholar] [CrossRef] [PubMed]
- Meyer-Schwesinger, C.; et al. Nephrotic syndrome and subepithelial deposits in a mouse model of immune-mediated anti-podocyte glomerulonephritis. J Immunol 2011, 187, 3218–3229. [Google Scholar] [CrossRef] [PubMed]
- Hengel, F.E.; et al. Autoantibodies Targeting Nephrin in Podocytopathies. N Engl J Med 2024, 391, 422–433. [Google Scholar] [CrossRef]
- Liu, J.; et al. miR-204: Molecular Regulation and Role in Cardiovascular and Renal Diseases. Hypertension 2021, 78, 270–281. [Google Scholar] [CrossRef]
- Xue, M.; Zhuo, Y.; Shan, B. MicroRNAs, Long Noncoding RNAs, and Their Functions in Human Disease. Methods Mol Biol 2017, 1617, 1–25. [Google Scholar] [CrossRef]
- Cheng, Y.; et al. Endogenous miR-204 Protects the Kidney against Chronic Injury in Hypertension and Diabetes. J Am Soc Nephrol 2020, 31, 1539–1554. [Google Scholar] [CrossRef]
- Zhu, Y.; et al. Overexpression of microRNA-204-5p alleviates renal ischemia-reperfusion injury in mice through blockage of Fas/FasL pathway. Exp Cell Res 2019, 381, 208–214. [Google Scholar] [CrossRef]
- Dylewski, J.; et al. Differential trafficking of albumin and IgG facilitated by the neonatal Fc receptor in podocytes in vitro and in vivo. PLoS One 2019, 14, e0209732. [Google Scholar] [CrossRef]
- Sun, H.; et al. Proteolytic characteristics of cathepsin D related to the recognition and cleavage of its target proteins. PLoS One 2013, 8, e65733. [Google Scholar] [CrossRef]
- Dylewski, J.F.; et al. Podocyte-specific knockout of the neonatal Fc receptor (FcRn) results in differential protection depending on the model of glomerulonephritis. PLoS One 2020, 15, e0230401. [Google Scholar] [CrossRef] [PubMed]
- Fox, C.; et al. Inhibition of lysosomal protease cathepsin D reduces renal fibrosis in murine chronic kidney disease. Sci Rep 2016, 6, 20101. [Google Scholar] [CrossRef] [PubMed]
- Beck, L.H., Jr.; et al. M-type phospholipase A2 receptor as target antigen in idiopathic membranous nephropathy. N Engl J Med 2009, 361, 11–21. [Google Scholar] [CrossRef]
- Tomas, N.M.; et al. Thrombospondin type-1 domain-containing 7A in idiopathic membranous nephropathy. N Engl J Med 2014, 371, 2277–2287. [Google Scholar] [CrossRef]
- Sethi, S.; et al. Neural epidermal growth factor-like 1 protein (NELL-1) associated membranous nephropathy. Kidney Int 2020, 97, 163–174. [Google Scholar] [CrossRef]
- Al-Rabadi, L.F.; et al. Serine Protease HTRA1 as a Novel Target Antigen in Primary Membranous Nephropathy. J Am Soc Nephrol 2021, 32, 1666–1681. [Google Scholar] [CrossRef] [PubMed]
- Watts, A.; et al. Discovery of autoantibodies targeting nephrin in minimal change disease supports a novel autoimmune etiology. J Am Soc Nephrol 2021. [Google Scholar] [CrossRef]
- Martin, C.E.; Jones, N. Nephrin Signaling in the Podocyte: An Updated View of Signal Regulation at the Slit Diaphragm and Beyond. Front Endocrinol (Lausanne) 2018, 9, 302. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.Y.; et al. Knockdown of lncRNA MALAT1 ameliorates acute kidney injury by mediating the miR-204/APOL1 pathway. J Clin Lab Anal 2021, 35, e23881. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; et al. Silencing Long Non-coding RNA Kcnq1ot1 Limits Acute Kidney Injury by Promoting miR-204-5p and Blocking the Activation of NLRP3 Inflammasome. Front Physiol 2021, 12, 721524. [Google Scholar] [CrossRef]
- Li, X.Y.; et al. MiR-204/miR-211 downregulation contributes to candidemia-induced kidney injuries via derepression of Hmx1 expression. Life Sci 2014, 102, 139–144. [Google Scholar] [CrossRef]
- Hu, H.; et al. LncRNA SPANXA2-OT1 Participates in the Occurrence and Development of EMT in Calcium Oxalate Crystal-Induced Kidney Injury by Adsorbing miR-204 and Up-Regulating Smad5. Front Med (Lausanne) 2021, 8, 719980. [Google Scholar] [CrossRef]
- Ougaard, M.K.E.; et al. Murine Nephrotoxic Nephritis as a Model of Chronic Kidney Disease. Int J Nephrol 2018, 2018, 8424502. [Google Scholar] [CrossRef] [PubMed]
- Haddad, G.; et al. Altered glycosylation of IgG4 promotes lectin complement pathway activation in anti-PLA2R1-associated membranous nephropathy. J Clin Invest 2021, 131. [Google Scholar] [CrossRef] [PubMed]
- Bruno, V.; et al. New insights into the immune functions of podocytes: the role of complement. Mol Cell Pediatr 2023, 10, 3. [Google Scholar] [CrossRef] [PubMed]
- Szecsi, P.B. The aspartic proteases. Scand J Clin Lab Invest Suppl 1992, 210, 5–22. [Google Scholar] [CrossRef]
- Sun, H.; et al. Dynein-Mediated Trafficking: A New Mechanism of Diabetic Podocytopathy. Kidney360 2023, 4, 162–176. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; et al. Endoplasmic reticulum-associated degradation is required for nephrin maturation and kidney glomerular filtration function. J Clin Invest 2021, 131. [Google Scholar] [CrossRef]
- Pawluczyk, I.; et al. A Pilot Study to Predict Risk of IgA Nephropathy Progression Based on miR-204 Expression. Kidney Int Rep 2021, 6, 2179–2188. [Google Scholar] [CrossRef]
Figure 1.
NTS targets nephrin when miR-204-5p expression is reduced.

Figure 2.
Nephrin degradation is mediated by an aspartyl enzyme.
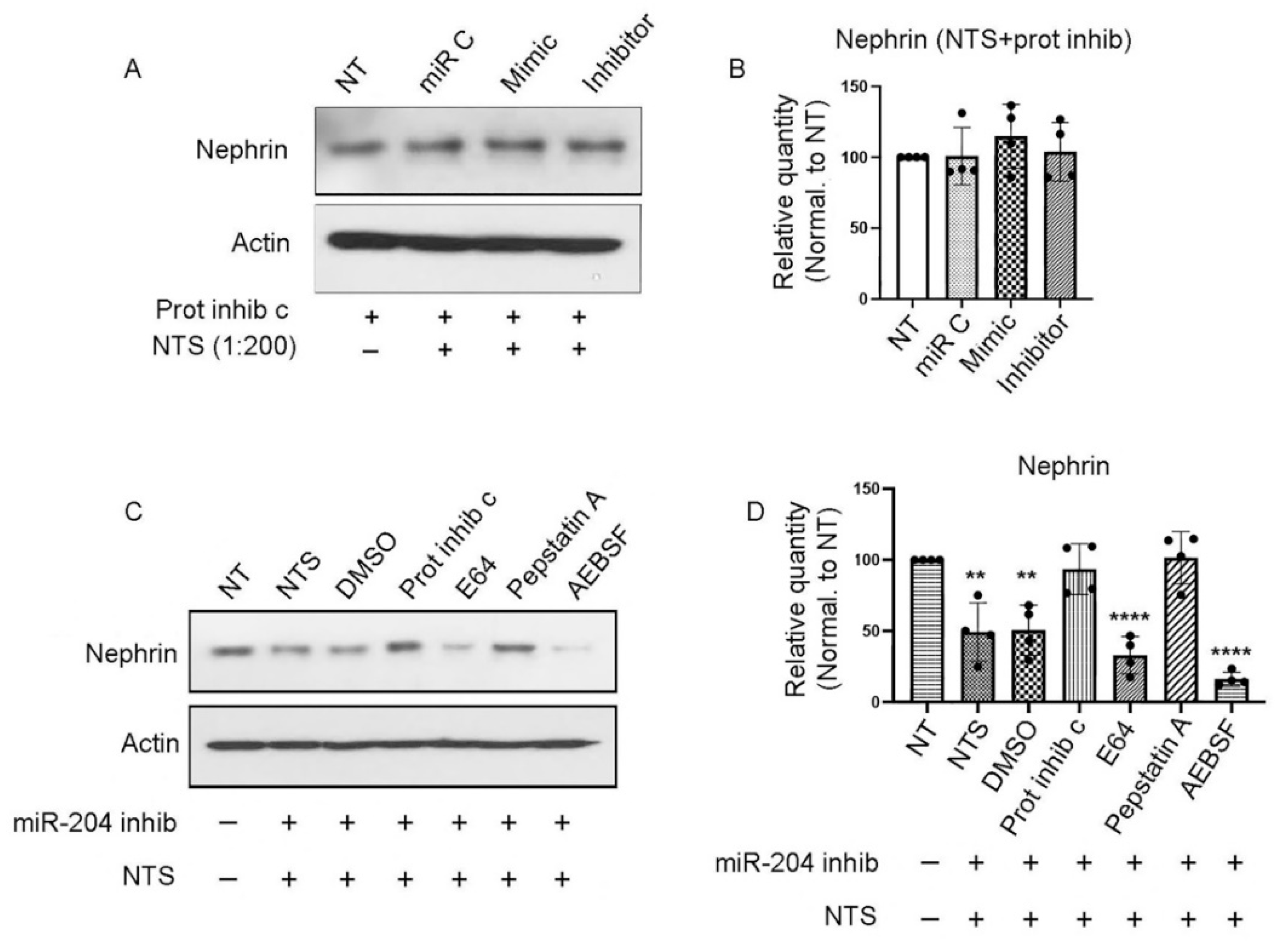
Figure 3.
LAMP1 and cathepsin D are targets of miR-204-5p. The overexpression of miR-204-5p mimic sequence reduced the expression of LAMP1 (A & C) but had no effect on nephrin (A & B) or cathepsin E (A & D) as determined by Western blot analysis. Using cathepsin ELISA the overexpression of miR-204-5p decreased cathepsin D expression (E). The effect of cathepsin D enzymatic activity on nephrin is shown in (F) using SDS-PAGE gel and silver stain. Cathepsin D treatment results in nephrin fragments (arrows). The experiments were repeated 4 times (n = 4). P values < 0.05 were considered significant.
Figure 3.
LAMP1 and cathepsin D are targets of miR-204-5p. The overexpression of miR-204-5p mimic sequence reduced the expression of LAMP1 (A & C) but had no effect on nephrin (A & B) or cathepsin E (A & D) as determined by Western blot analysis. Using cathepsin ELISA the overexpression of miR-204-5p decreased cathepsin D expression (E). The effect of cathepsin D enzymatic activity on nephrin is shown in (F) using SDS-PAGE gel and silver stain. Cathepsin D treatment results in nephrin fragments (arrows). The experiments were repeated 4 times (n = 4). P values < 0.05 were considered significant.
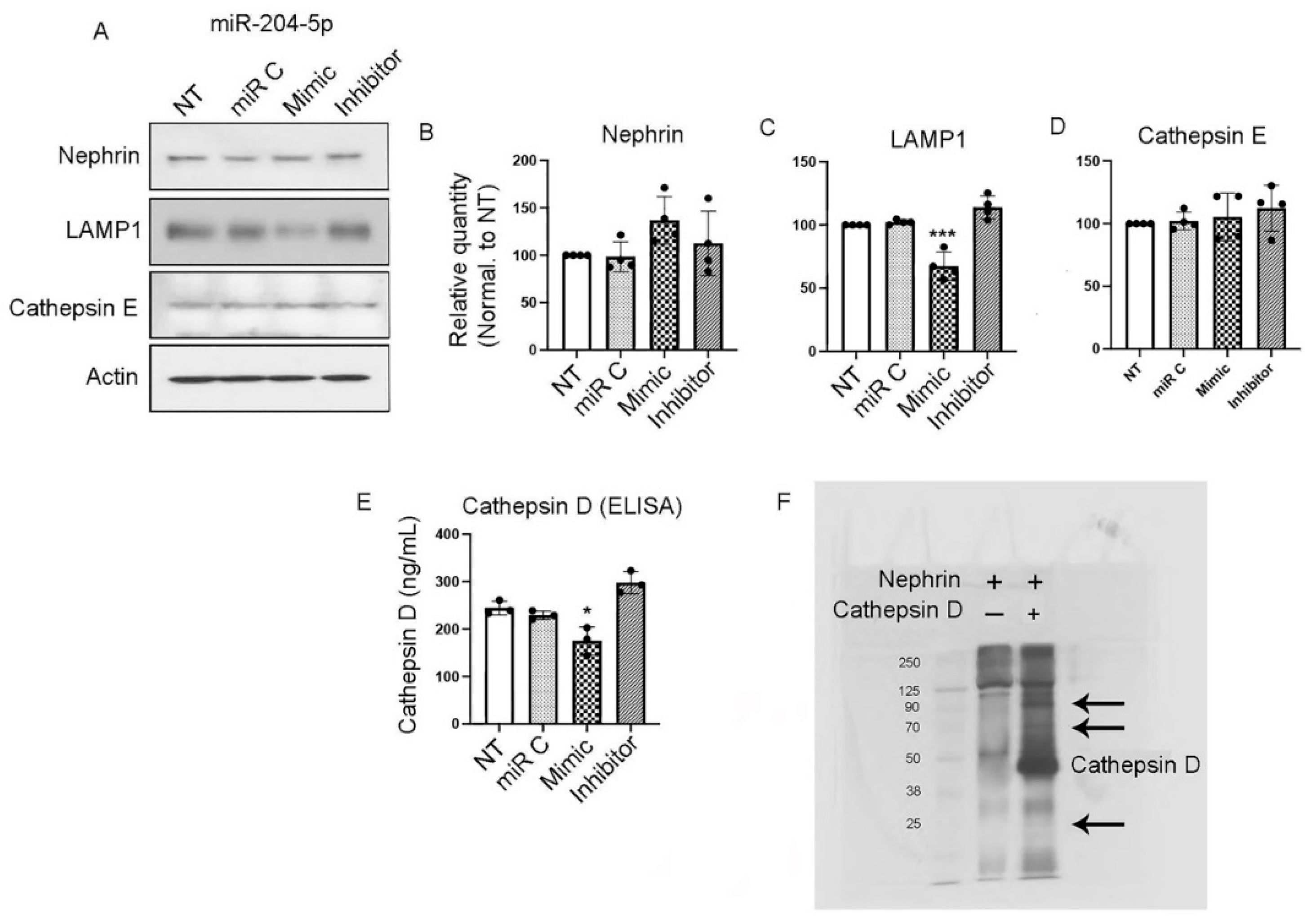
Figure 4.
LncJosd1-ps regulates the expression of miR-204-5p. LncJosd1-ps sequence was cloned into a lentiviral vector (A) to overexpress lncJosd1-ps and an antisense oligo (ASO) was designed (B) to reduce lncJosd1-ps expression. (C) shows that the overexpression of lncJosd1-ps reduced miR-204-5p expression, whereas lncJosd1-ps inhibition increased miR-204-5p expression (D). The subcellular location of lncJosd1-ps was determined and it shows nuclear as well as cytoplasmic expressions (E). The experiments were repeated 4 times (n = 4). P values < 0.05 were considered significant.
Figure 4.
LncJosd1-ps regulates the expression of miR-204-5p. LncJosd1-ps sequence was cloned into a lentiviral vector (A) to overexpress lncJosd1-ps and an antisense oligo (ASO) was designed (B) to reduce lncJosd1-ps expression. (C) shows that the overexpression of lncJosd1-ps reduced miR-204-5p expression, whereas lncJosd1-ps inhibition increased miR-204-5p expression (D). The subcellular location of lncJosd1-ps was determined and it shows nuclear as well as cytoplasmic expressions (E). The experiments were repeated 4 times (n = 4). P values < 0.05 were considered significant.
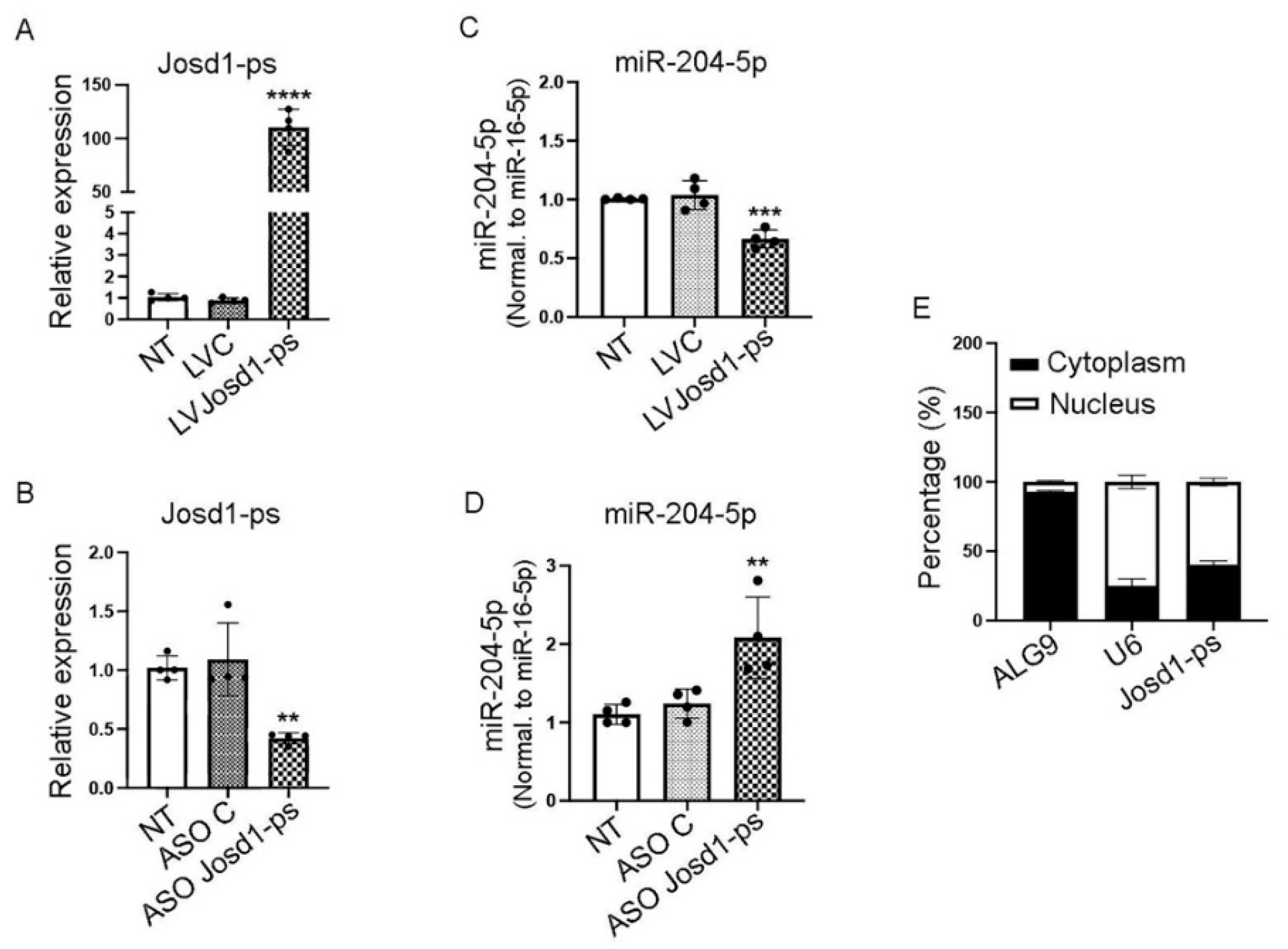
Figure 5.
Validation of miR-204-5p targets by luciferase assay. The miR-204-5p binding sites within LAMP1 and cathepsin D 3’UTR and within lncJosd1-ps nucleotides sequence are shown in (A). miR-204-5p interaction with LAMP1 3’UTR is validated by a luciferase assay using intact LAMP1 or mutated binding sites (B), cathepsin D (C), and lncJosd1-ps (D). The experiments were repeated 3 times (n=3). P values < 0.05 were considered significant.
Figure 5.
Validation of miR-204-5p targets by luciferase assay. The miR-204-5p binding sites within LAMP1 and cathepsin D 3’UTR and within lncJosd1-ps nucleotides sequence are shown in (A). miR-204-5p interaction with LAMP1 3’UTR is validated by a luciferase assay using intact LAMP1 or mutated binding sites (B), cathepsin D (C), and lncJosd1-ps (D). The experiments were repeated 3 times (n=3). P values < 0.05 were considered significant.
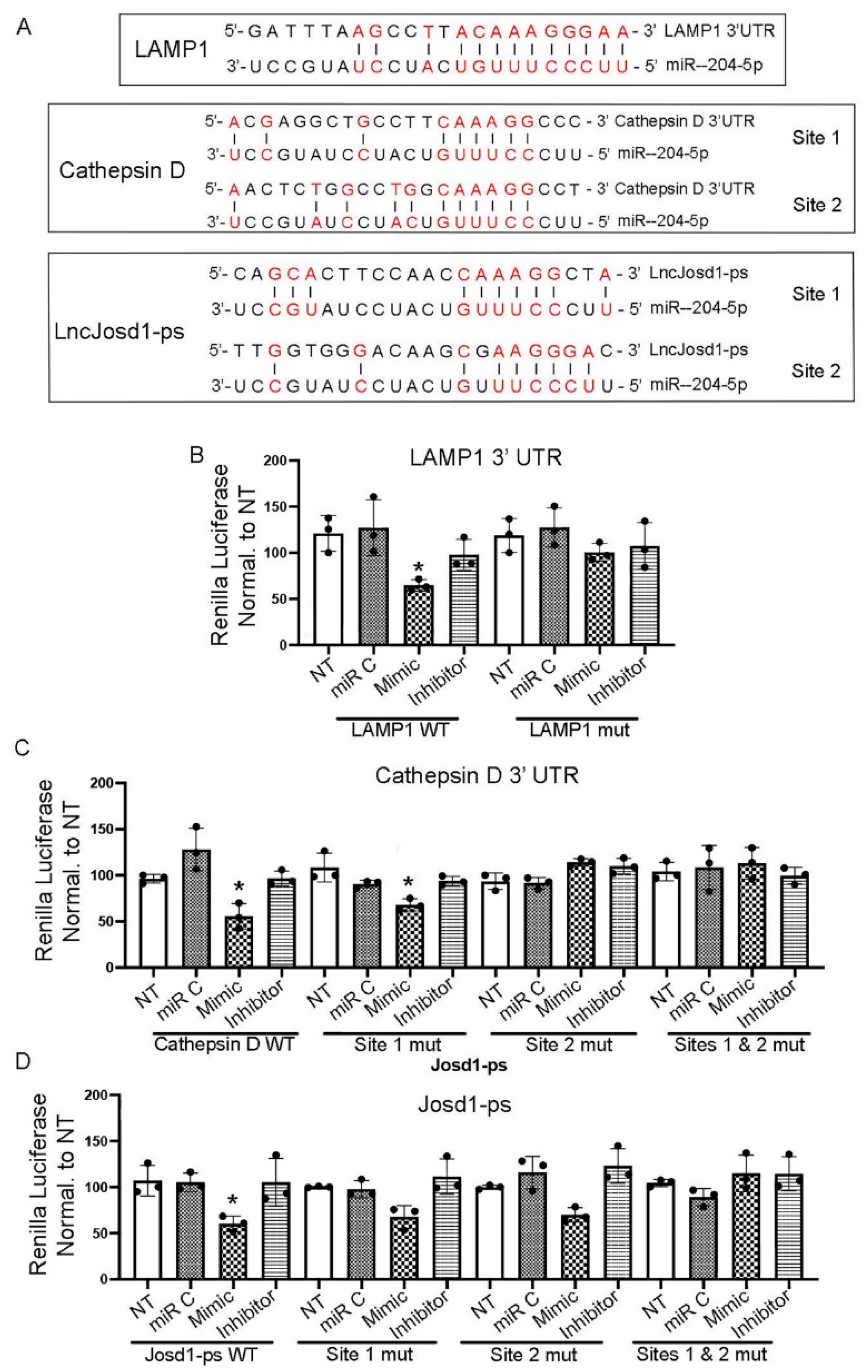
Figure 7.
Pepstatin A reduced kidney injury in the NTS model of immune mediated kidney disease. Mouse kidney total RNA was analyzed for miR-204-5p and lncJosd1-ps expressions in animals treated with NTS followed by IP injection of Pepstatin A (20 mg/Kg) or carrier (100 µl ethanol) on days 1, 3, and 5 (A & B). The kidney injury markers KIM1 (C) and NGAL (D) expression levels were determined at the mRNA level along with the podocyte markers nephrin (E) and podocin (F). Kidney morphology was assessed using PAS stain (G). Glomerulosclerosis was determined using a 0-4 scoring system where 0 = 0 %, 1 = 25 %, 2 = 50 %, 3 = 75 % and 4 = 100 % glomerulosclerosis. All available cortical glomeruli in a PAS-stained tissue section were analyzed (H). Albuminuria was determined using the albumin to creatinine ratio (I). At least 5-8 animals were used per group. P values < 0.05 were considered significant, scale bars 50 µm.
Figure 7.
Pepstatin A reduced kidney injury in the NTS model of immune mediated kidney disease. Mouse kidney total RNA was analyzed for miR-204-5p and lncJosd1-ps expressions in animals treated with NTS followed by IP injection of Pepstatin A (20 mg/Kg) or carrier (100 µl ethanol) on days 1, 3, and 5 (A & B). The kidney injury markers KIM1 (C) and NGAL (D) expression levels were determined at the mRNA level along with the podocyte markers nephrin (E) and podocin (F). Kidney morphology was assessed using PAS stain (G). Glomerulosclerosis was determined using a 0-4 scoring system where 0 = 0 %, 1 = 25 %, 2 = 50 %, 3 = 75 % and 4 = 100 % glomerulosclerosis. All available cortical glomeruli in a PAS-stained tissue section were analyzed (H). Albuminuria was determined using the albumin to creatinine ratio (I). At least 5-8 animals were used per group. P values < 0.05 were considered significant, scale bars 50 µm.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



