Submitted:
28 January 2025
Posted:
29 January 2025
You are already at the latest version
Abstract
SOS1 is the plant membrane Na+/H+ antiporter of Arabidopsis thaliana and is the major transporter extruding Na+ across the plasma membrane of cells in exchange for an intracellular H+. The first 450 amino acid are the membrane transport domain. We examined nine different mutations that may either change specificity or improve salt tolerance. E261K had minor negative effects on the ability to confer LiCl and NaCl tolerance. Mutation A399V had minor effects lowering LiCl tolerance and slightly improving NaCl tolerance as did the double mutant E261KA399V. Four different mutations of amino acid Y346 had varying effects. The Y346R mutation resulted in a major improvement in LiCl tolerance but did not affect NaCl tolerance. The L375I mutant was impaired in NaCl tolerance and the Q362L mutant had minor effects on salt tolerance. Our results demonstrate that amino acid Y346 is critical in ion selectivity and its mutation can dramatically improve LiCl salt tolerance. Some other mutations showed minor improvements in the ability to confer NaCl tolerance (Y346F, A399V, Y346F and Y346A) leaving open the possibility that such mutations might improve salt tolerance in intact plant species.
Keywords:
Arabidopsis thaliana
; ion transport
; membrane protein
; Na+/H+ antiporter
; salt tolerance
; SOS1
1. Introduction
Most plants including crop plants, are highly sensitive to salt stress. Salt stress causes osmotic stress and water deficit, and produces biochemical problems affecting plant growth rate and survival [1,2,3]. Plants usually maintain high intracellular K+ (100-200 mM) and low Na+ (1-10 mM). For crop plants, soil salinity is a key factor limiting plant growth and agricultural productivity [4]. One important mechanism that plants use to deal with Na+ stress is to use the signaling pathway called the Salt Overlay Sensitive (SOS) pathway that detects salt stress and takes protective measures [5]. An important component of the SOS pathway is SOS1, the sodium transporter of the pathway. It is a plasma membrane Na+/H+ exchanger that exchanges an extracellular H+ for an intracellular Na+ ion. It therefore uses the proton gradient generated by the plasma membrane H+-ATPase to remove the intracellular sodium [6]. Plasma membrane SOS1 is chiefly expressed in the root and xylem of plants [6]. SOS1 can also be expressed in vacuoles where it also contributes to salt tolerance [7].
Other types of cation/H+ antiporters exist in plants including intracellular Na+/H+ antiporters and K+(Na+)/H+ antiporters which also assist in coping with salt stress [8,9]. However, SOS1 is of special interest because of its clear role in permitting salt tolerance [3,10] without a requirement for vesicle trafficking. Overexpression of SOS1 improves salt tolerance in plants including in crop plants [11,12,13]. Similarly, enhancing SOS1 activity through regulatory pathways in plants promotes salt tolerance [14]. Conversely, decreased expression of SOS1 causes loss of salt tolerance in different species [15,16,17,18]. AtSOS1 (Arabidopsis thaliana SOS1) can confer salt tolerance across distant species. S. pombe has one principal plasma membrane salt tolerance protein, Sod2, and its deletion results in a salt-sensitive yeast S. pombe strain [19]. The deficient strain shows improved salt tolerance when AtSOS1 is expressed [20] making it an excellent system for examining effects of mutations on the protein.
Given the importance of SOS1 we thought it important to investigate amino acids important in its function. In addition, we have noted that some amino acid residues have been suggested to enhance or be critical in SOS1 activity. For example, Quintero [21] has shown that mutations E261K and A399V, enhanced activity of SOS1 introduced into Saccharomyces cerevisiae. A different study [22] examined how selection pressure acted on the SOS1 gene in the Populus genus to mediate saline resistance. They found several amino acid sites selected for in salt tolerant plant types, including amino acids 345, 361 and 374. In this study, we examined and further characterized the mutation of the corresponding amino acids in a different system, in the salt-sensitive yeast strain Schizosaccharomyces pombe that has its sole plasma membrane salt tolerance protein sod2 deleted [19]. Earlier [20] we demonstrated that we could express Arabidopsis thaliana SOS1 functionally in this system and used a shortened version of AtSOS1. In this study we used this construct so that we can focus on effects mediated by the more conserved membrane transport domain as opposed to those on the regulatory domain. We characterize effects of specific mutations implicated in improved salt tolerance using the shortened protein. We examine effects on expression, alterations in different cation transport and examine some combinations of mutations. Our results demonstrate that some of these mutations improved salt tolerance in the SOS1 protein membrane domain without the regulatory tail. We also found, that Y346 was an amino acid important in specificity of the SOS1 protein, suggesting it may be a critical component of the ion coordination during ion transport.
2. Results
2.1. SOS1 Alignment and Modeling- Salt Tolerant Strains of Plants Have Evolved Adaptations to Cope with the Presence of Extraneous Sodium.
Meng and Wu [22] did a systematic examination of some of these adaptations in SOS1 in the genus Populus examining the Turanga group which has undergone selection pressure for salt tolerance. Three amino acids were often changed to Arg, Leu and Phe respectively. In this species R345, Leu361 and Ile374 corresponded to these amino acids. Figure 1A compares the sequence of several species including that of Populus euphratica and Arabidopsis thaliana SOS1. The three amino acids often changed to Arg, Leu and Phe respectively in AtSOS1 correspond to amino acids Tyr346, Gln362 and Leu375. This led us to mutate these amino acids to Arg, Leu and Phe respectively (below). Additionally, the mutations E261K and A399V, were earlier shown to enhance activity of SOS1 that was introduced into Saccharomyces cerevisiae [21] so this group of amino acids was chosen for analysis in further studies and this region is also shown in Figure 1A. The residues corresponding to E261 and A399 of AtSOS1 are tightly conserved.
SOS1 was examined in earlier studies by cryo-electron microscopy [25] however that analysis was not at a level able to reveal details of the protein structure. We recently used multiple sequence analysis to predict a topology of 13 transmembrane segments [20]. However, more recently [23,24] the detailed structure of SOS1 was determined. We therefore used this structure (Figure 1B, C) to illustrate the locations of the residues which we examined in this study. Analyses of the cryo-EM structure of A. thaliana SOS1 reveal that Tyr346 is located in the extracellular loop connecting transmembrane helix 10 (TM10) and transmembrane helix 11 (TM11) (Figure 1B, C). TM10 is part of the dimerization domain of the protein, while TM11 belongs to the transport domain [23,24]. Multiple sequence alignment (Figure1) shows that residues between these two helices are not conserved. However, the loop maintains a critical balance between negatively and positively charged residues. Our previous work demonstrated that the extracellular loop's charge is essential for the functioning of Na+/H+ antiporters [26]. Further multiple sequence alignment across a wide range of plant SOS1 proteins indicates that, in most plants, a negatively charged amino acid is paired with a positively charged residue in this loop (Figure 1A). Interestingly, in Populus euphratica SOS1, a salt-tolerant species, the corresponding position of Tyr346 in A. thaliana is occupied by arginine, and the same loop lacks any negatively charged residues. Given P. euphratica’s tolerance to salinity, we attempted to replace Tyr346 of A. thaliana SOS1 with arginine to explore its functional implications. Additionally, the cryo-EM structure and sequence of Oryza sativa SOS1 (OsSOS1) available in the PDB database shows that its TM10-TM11 loop contains an arginine residue preceded by an aspartate in the polypeptide sequence (Figure 1).
Figure 2 illustrates close ups of the residues investigated in this work. The Glu261 interaction network is shown in Figure 2A. Glu261 is located at the intracellular side of the dimerization interface. This acidic residue closely interacts with Asp257 through backbone interaction, whereas it interacts with Ser307 through a side chain interaction, and with Lys304 its interaction is through a side chain backbone interaction. Figure 2B shows the proximity of Glu362 to the discontinues helices and proximity to Trp363 and Asn440. Figure 2C illustrates the extracellular location of Tyr346 in the loop joining TM10 with TM11. The hydrophobic network provided by the residue Leu375 together with Leu131, Leu372, and Phe379 is shown in Figure 2D. The amino acid Ala399 and its juxtaposition to Ile326 and Leu109 in the center of the protein provide a hydrophobic groove and are shown in Figure 2E.
2.2. Expression of Wild Type and Mutant SOS1 in S. pombe-
To study the amino acids critical in SOS1 function we expressed the protein in yeast in a knock out strain (sod2::ura4) deficient in endogenous salt tolerant protein as described earlier [27,28,29]. To confirm that both wild type and mutant SOS1 proteins were expressed we used Western blotting against the GFP tag present on the SOS1 protein C-terminal end. The results (Figure 3) demonstrate that all the mutants and the wild type SOS1s proteins were expressed at similar levels. Mutation of SOS1s did not appear to adversely affect expression levels. The size of the shortened SOS1 protein was the same as that reported earlier [20].
2.3. Salt Tolerance of Wild Type and Mutant SOS1 in S. pombe-
We next examined the ability of wild type SOS1 and SOS1 mutants plus sod2 to return salt tolerance in S. pombe that has endogenous sod2 deleted. LiCl and NaCl are both transported by sod2 and a variety of other Na+/H+ exchangers [28]. LiCl was used for assays in liquid media since it is toxic at lower concentrations, avoiding the osmotic challenge of high concentrations of NaCl. Li also has a smaller ionic radius than Na (0.76 A vs 1.02 A) [30]. Figure 4 panels A-X show the growth of various S. pombe strains containing the indicated mutant. Panels A-L illustrate growth in LiCl of varying concentrations and panels M-X in NaCl of varying concentrations. Table 1 summarizes the results of the experiments.
The growth of the knockout strain (Figure 4A) occurred in 0 and 2 mM LiCl. The positive control, the yeast Na+/H+ exchanger sod2 conferred resistance to all concentrations of LiCl tested, up to 8 mM. The SOS1s construct conferred partial resistance to LiCl, allowing growth in up to 4 and 5 mM LiCl (Figure 4C). Growth of the mutants in Figures 4D- L, (mutants E261K, A399V, E261KA399V, Y346A, Y346F, Y346K, L375I and Q362L) in LiCl showed similarities to the wild type SOS1s but varied in interesting and sometimes significant ways. For SOS1 with the E261K mutation (Figure 4D) the general pattern of growth was similar, but growth at the lower LiCl concentrations (2, 4 and 5 mM) was reduced compared to controls at some time points. In contrast, with the E261K mutation there was a slight improvement in growth at some intermediate time points in 6 and 8 mM LiCl, though these changes were quite small. SOS1 with the A399V mutation (Figure 4E) conveyed similar salt tolerance as the wild type protein with minor changes. In 4 and 5 mM LiCl growth was somewhat reduced. The E261KA399V mutant (Figure 4F) displayed similar changes to the A399V mutant with somewhat slightly reduced growth in lower LiCl containing medium. Both the Y346A (Figure 4G) and the Y346K (Figure 4I) mutant proteins conveyed substantially reduced salt tolerance in lower LiCl containing medium though the Y346K mutant was also slightly ineffective in conferring salt tolerance in higher LiCl concentrations. In contrast to these last two mutations, another mutation at this position, Y346F (Figure 4H), had no effect LiCl tolerance. The greatest effect shown of any mutations, was the Y346R mutation (Figure 4J), which showed a remarkable improvement in LiCl tolerance in both high and low LiCl concentrations. This mutation improved LiCl tolerance greatly over a wide range of concentrations and times. The mutations L375I and Q362L (Figure 4K, L) did not greatly affect LiCl tolerance. The Q362L mutation had a positive effect on growth in 6 mM LiCl but both mutations had an effect of slightly decreasing growth in the absence of LiCl during exponential growth. However, the final growth stage reached in the absence of LiCl have a was not different from cells with wild type SOS1.

Table 1.
Summary of effect of mutations of SOS1s on ability to confer salt tolerance compared to wild type SOS1. Based on liquid phase assay. (A), growth in liquid media. (B), growth on solid media. - reduced growth, less than SOS1 wild type at some time points at these concentrations; -,- greatly reduced growth compared with SOS1 wild type at some time points at these concentrations; + growth slightly (but significantly) greater than wild type SOS1 at some time points at these concentrations; ++, growth more improved over wild type SOS1 by a greater amount or at more time points at the indicated concentrations; +++, growth greatly improved over wild type SOS1; =, growth not significantly different from wild SOS1s at all, or almost all, time points at these concentrations; *, growth was reduced in LiCl free medium, ^ grown was increased in NaCl free medium.
Table 1.
Summary of effect of mutations of SOS1s on ability to confer salt tolerance compared to wild type SOS1. Based on liquid phase assay. (A), growth in liquid media. (B), growth on solid media. - reduced growth, less than SOS1 wild type at some time points at these concentrations; -,- greatly reduced growth compared with SOS1 wild type at some time points at these concentrations; + growth slightly (but significantly) greater than wild type SOS1 at some time points at these concentrations; ++, growth more improved over wild type SOS1 by a greater amount or at more time points at the indicated concentrations; +++, growth greatly improved over wild type SOS1; =, growth not significantly different from wild SOS1s at all, or almost all, time points at these concentrations; *, growth was reduced in LiCl free medium, ^ grown was increased in NaCl free medium.
| ||||||||
| Mutant | Growth in 2-5 mM LiCl | Growth in 6, 8 LiCl mM | Growth in 200- 500 mM NaCl | Growth in 600, 800 mM NaCl ( | ||||
| E261K | - | + | - | = | ||||
| A399V | - | = | + | = | ||||
| E261KA399V | - | = | + | = | ||||
| Y346A | -,- | = | + | = | ||||
| Y346F | = | = | +, ^ | = | ||||
| Y346K | -,- | - | = | + | ||||
| Y346R | +++ | +++ | = | = | ||||
| L375I | =, * | = | -,- | - | ||||
| Q362L | =, * | +, * | = | = | ||||
| ||||||||
| Mutant | Growth in 2, 4 mM LiCl | Growth in 5, 8 LiCl mM | Growth in 200, 400 mM NaCl | Growth in 500, 800 mM NaCl | ||||
| E261K | - | = | = | = | ||||
| A399V | - | - | = | = | ||||
| E261KA399V | - | - | = | = | ||||
| Y346A | = | = | = | = | ||||
| Y346F | = | = | + | + | ||||
| Y346K | = | = | = | = | ||||
| Y346R | ++ | +++ | = | = | ||||
| L375I | = | -- | = | = | ||||
| Q362L | = | = | = | = | ||||
Tests of NaCl tolerance are shown in Figures 4M-X and summarized in Table 1. The growth of the sod2 knockout strain (Figure 4M) occurred in 0 and 200 mM NaCl with some minimal growth in 400 mM NaCl. Yeast containing the wild type sod2 protein grew robustly in 200 to 800 mM NaCl (Figure 4N). Expression of the SOS1s (Figure 4O) protein improved salt tolerance greatly in 400 and 500 mM NaCl but not in 600 mM NaCl or 800 mM. There were varying effects of mutations of SOS1s on NaCl tolerance which were mostly smaller changes. The E261K (Figure 4P, Table 1) mutation inhibited NaCl resistance somewhat in lower NaCl concentrations (200-500 mM) at the later stages of growth. The mutations A399V, E261KA399V, and Y346A all had slight but significant positive effects on salt tolerance in lower NaCl concentrations (200-500 mM). These were in the middle stages of the growth curve. The mutation of Y346 to Phe (Figure 4T) also had some positive effects in lower concentrations of NaCl and there was also a slight but significant improvement in growth relative to the control, in the absence of NaCl. Other mutations of Y346 had either no effect on growth (Y346R, Figure 4V) or a minor but significant positive effect on growth by mutation with the Y346K mutation (Figure 4U) in 500 mM NaCl. The L375I mutant (Figure 4W) showed only negative effects on growth and cells with SOS1s with this mutation showed NaCl tolerance that was more like the knockout in 500 mM NaCl. The Q362L mutation did not change growth in NaCl containing liquid media.
We next (Figure 5, Table 1) examined salt tolerance in the mutants on solid media using techniques we have described earlier [26] in order to verify results seen in liquid media. Figure 5A and B illustrate the growth in different concentrations of LiCl and Figure 5C and D in different concentrations of NaCl. In LiCl and in NaCl, wild type sod2 protein permitted robust growth in all concentrations tested. S. pombe with knock out of the sod2 protein grew well in up to 2 mM LiCl and 200 mM NaCl but not at higher concentrations. The wild type SOS1 protein improved growth in 400 and 500 mM NaCl and 4 and 5 mM LiCl but growth was still reduced compared to lower cation concentrations. These results mirrored those of the growth curves in liquid media with only some minor differences where there were subtle changes in salt tolerance (Table 1). The various mutations of amino acid Y346 were largely similar to that of wild type SOS1, with the notable exception of yeast with SOS1 with the Y346R mutation, which improved growth greatly in up to 8 mM LiCl. In both liquid and solid media, there were also smaller improvements in NaCl tolerance with the Y346F mutation. Mutations of A399V and the double mutant cause impaired growth in LiCl. Other cells containing mutations of the SOS1 protein did not show great changes in cation tolerance on solid phase media, with one minor exceptions. In the case of the L375I mutant, growth was inhibited at higher LiCl concentrations.
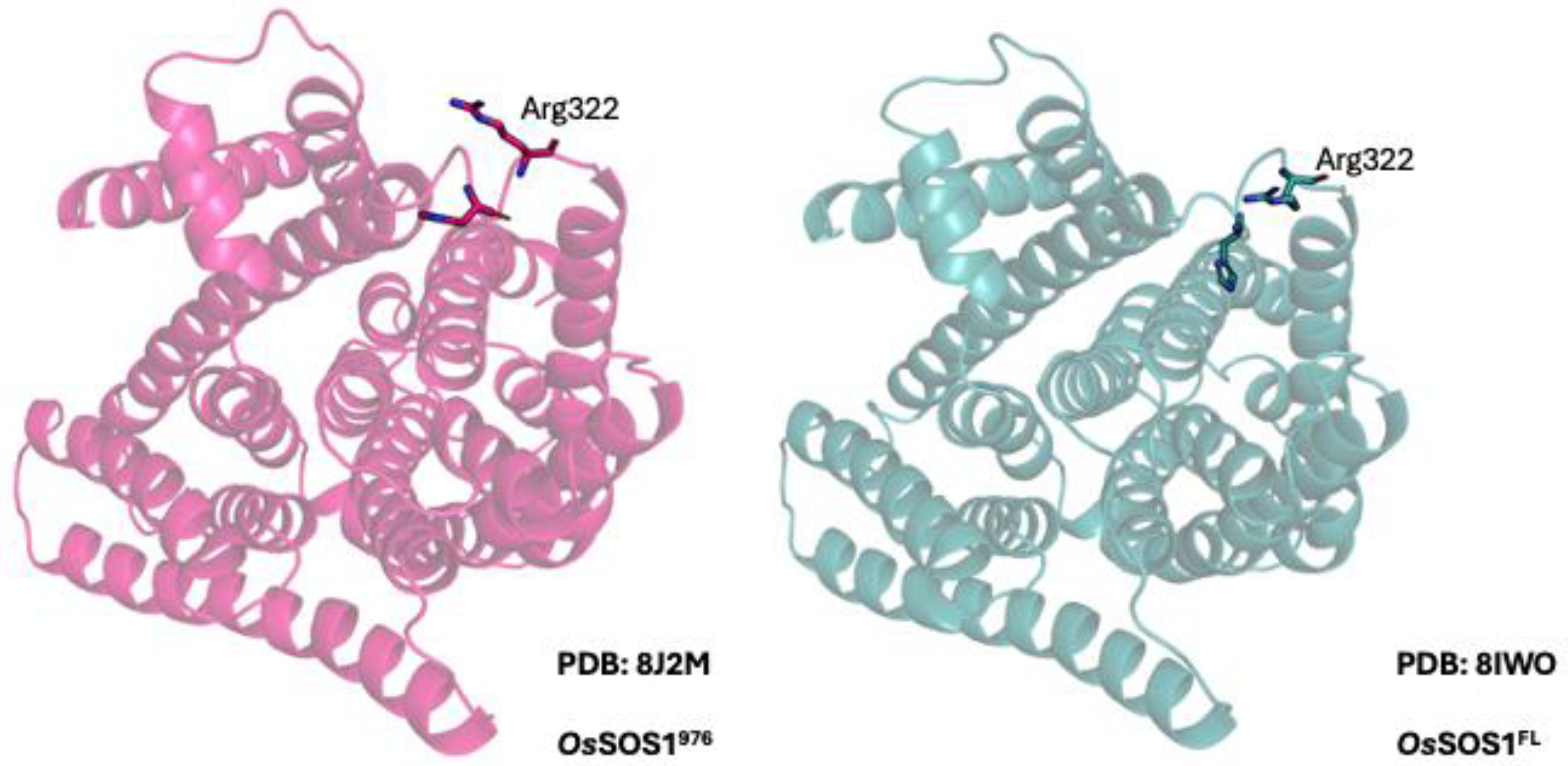
Because we found extremely interesting effects when we mutated amino acid Y346 of Arabidopsis thaliana SOS1, we examined the structure in the region of Arg322 of Oryza sativa SOS1 (the truncated rice Na+/H+ exchanger). This amino acid residue of Oryza sativa SOS1 corresponds to Y346 in Arabidopsis thaliana SOS1 (Figure 1). Figure 6 illustrates two different structures of the TM10-TM11 loop which contains Arg322. Both structures show that that loop is comparatively flexible and contain at least two positively charged residues and one negatively charged residue (Figure 6). The different positions of Arg322 indicate that there is flexibility in Oryza sativa SOS1 TM10-TM11 loop harboring the Arginine residue.
3. Discussion
Soil salinity is a key factor limiting plant growth and agricultural productivity [4]. As noted above, SOS1 is a key transporter involved in salt tolerance in plants and it extrudes an intracellular Na+ ion in exchange for an extracellular H+ [5]. It is obviously desirable to enhance SOS1 activity in this regard to enhance salt tolerance, plant growth SOS1 and agricultural activity. Knowledge of the transport mechanism and critical amino acids involved in transport would undoubtedly be useful in this regard. SOS1 of Arabidopsis was used as a model system in this study and it has a very large internal regulatory tail. Earlier we [14] examined effects of overexpression of SOS1 cytosolic tail fragments which was believed to lead to sequestration of inhibitory proteins and elevation of SOS1 activity, with a resultant increase in salt tolerance. Others, have shown in a variety of plant species, that overexpression of SOS1 or SOS1 like proteins can result in elevated salt tolerance and deletion results in salt sensitivity [6,11,31,32,33]. However, in addition to other regulatory proteins, SOS1 contains a regulatory tail with regions inhibitory to activity and this can confound effects of simple overexpression of the protein [13]. Also, the carboxyl terminal regulatory region is more variable between species and subject to many regulatory factors which are likely to be variable from one plant species to another. Our goal in this study was to characterize ion transport itself in the membrane domain including an examination of amino acids involved in Na+ vs Li+ transport. Additionally, we attempted to identify putative changes in the SOS1 protein that might improve transport and might be widely applicable in different species. We therefore studied the membrane domain which is more conserved and responsible for cation binding and transport and can mediate salt tolerance in yeast [20,34].
We have previously developed an expression and mutation system for S. pombe [20]. In this species elimination of only one salt tolerance protein, sod2, results in salt sensitivity and SOS1 has earlier been shown to restore salt resistance after removal of sod2 [13,20,35]. We have also earlier shown that a shortened version of SOS1, without the inhibitory/regulatory domain is functional in this system [20]. We therefore used this system in the present study, to examine the effects of several amino acids that were suggested to be important in salt tolerance. One group of such amino acids we examined was amino acids Tyr346, Gln362 and Leu375. A phylogenetic analysis had earlier suggested that the corresponding amino acids were important in salt tolerance in Populus euphratica [22]. Figure 1A shows a comparison of amino acids 314-393 of Arabidopsis thaliana SOS1 with the corresponding sequence of Populus euphratica and other related species. Amino acids mutated in this study, Tyr346, Gln362 and Leu375, are indicated and correspond to those suggested to be important in salt tolerance in Populus euphratica. Another pair of amino acids E261 and A399 had earlier [21] also been suggested to be significant in mediating salt tolerance and we examined these and also attempted to examine the effect of inserting a pair of putative salt improving mutations in the same system, in an attempt to further increase salt tolerance (Table 1).
Results with amino acids E261 and A399 were varied. Results with liquid media were more easily quantified. Results with solid media are less sensitive or quantifiable, but they generally agreed with those found in liquid media notably agreeing with the larger more easily viewed changes. In our system, the individual mutation of E261K did not result in an improvement in tolerance to NaCl but did result in a minor improvement in LiCl tolerance in liquid media. There was also a minor reduction in NaCl tolerance. This is in contrast to results found earlier [21] and we can only attribute this to the different system we used. In the case of the A399V mutation, this did result in an improvement in NaCl tolerance. Thus, we can state that there was a minor effect of this mutation which agrees with the earlier report [21] but the effect was not large. It seems as though this mutation is a more robust one which might be favored for further studies in an intact plant model. The combined E261K/A399V mutations showed an effect similar to that of the A399V mutation alone in liquid media, so there was no additional advantage of adding the A399V mutation to the mutant E261K. Why the earlier report showed greater effects than our study is not clear, though this may be due to either the different system used, or due to the absence of the regulatory tail. Whatever the cause, the potency of the mutations effects was not easily transferred to our system. This might mean that they are not easily transferrable to other plant systems. However, further experiments are necessary to determine this.
The Ala399 amino acid is on TM12 and is in the unwound region mid membrane which is a part of the characteristic Na+/H+ exchanger fold of SOS1 (Fig. 2E). The fold has been shown to be critical to Na+/H+ exchanger activity [36]. Thus, it seems reasonable that a mutation in this fold might affect transport. We found mixed results with this amino acid and how this improved transport in an earlier study is not yet clear [21]. In contrast Glu261 is present on the N-terminal of TM11 close to the cytoplasmic side of the protein near amino acids Lys304 and Ser307 (Fig. 2A). There it could play a part in attraction of cations to the transport core. It is difficult to understand how a change from a negatively charged glutamic acid to a positively charged lysine can enhance transport of LiCl. Li has a smaller ionic radius than Na, so perhaps a change to the positive charge excluded this amino acid from playing a role in electrostatic environment mediating cation coordination, allowing the balance of negatively charged residues to release Li more quickly. Alternatively, removing one point of coordination may more suitably coordinate this smaller ion, in comparison to Na+. These theories have yet to be tested.
Mutation of amino acid Q362 to leucine did have a positive effect on growth in higher concentrations of LiCl in liquid media. Q362 is found in relatively close proximity (Fig. 2I, J) to the NhaA fold [37,38] that is critical in cation transport. Thus, it is possible that it affects cation coordination, an effect more evident at higher concentrations. Since this was not found with NaCl, and only at higher LiCl concentrations, it is likely that the effect is a change in cation coordination rather than an enhancement of turnover rate.
Mutation of L375 to isoleucine had a pronounced negative effect on growth in liquid medium containing NaCl at various concentrations. This is in contrast to a previous report [22] that suggested that the corresponding mutation in Poplus enhances salt tolerance. We suggest that the beneficial effect of changing this amino acid to isoleucine may be species specific, and not easily transferrable to the SOS1 protein of Arabidopsis thaliana. This makes it a less desirable target as a method of increasing salt tolerance in many plant species.
The most interesting effect we found was with mutation of amino acid Y346. As noted above, the corresponding amino acid in Poplus euphratica is an arginine residue, which was associated with a fitness advantage in saline environments in a genetic analysis of this species. We therefore mutated this residue to arginine to mimic the residue in Poplus. Our preliminary experiments were interesting therefore we made a number of other mutations of this residue to amino acids, alanine, phenylalanine and lysine. Only mutation of this residue to arginine resulted in a strong positive effect, and that effect was on LiCl resistance. There was a dramatic increase in growth in LiCl containing medium at both high and low concentrations of LiCl (Figure 4 and Figure 5, Table 2). The effect was quite specific. A change to lysine, that has the same charge but is a different size, could not mimic this effect and even had a negative effect on LiCl tolerance. (It is however notable there was one slight improvement in NaCl tolerance at one high concentration of NaCl, though this was a small effect and only at one concentration of NaCl.) Other mutations to alanine or phenylalanine did show some improvements in salt tolerance that were not as dramatic. There were small but significant improvements in NaCl tolerance shown with the Y346 to alanine and the Y346 to phenylalanine mutation at lower NaCl concentrations. While not huge, these improvements at concentrations between 200-500 mM, might be significant if they occurred in plants encountering salt challenge in the environment.
Comparative analysis of two OsSOS1 structures reveals that the TM10-TM11 loop exhibits significant flexibility [23] (Fig. 6). The location of the amino acid Y346 is within this same TM10-TM11 loop of the A. thaliana SOS1 protein (Fig. 1, 2C). Multiple sequence alignment across a wide range of plant SOS1 proteins (Fig. 1) indicated that, in most plants, a negatively charged amino acid is paired with a positively charged residue in this loop (Fig. 1). As noted above, in salt-tolerant Populus euphratica SOS1 the corresponding position of Tyr346 in A. thaliana is occupied by arginine, and the same loop lacks any negatively charged residues. We replaced Tyr346 of A. thaliana SOS1 with arginine to explore its functional implications. Additionally, the cryo-EM structure of Oryza sativa SOS1 (OsSOS1) available in the PDB database shows that its TM10-TM11 loop contains an arginine residue preceded by an aspartate in the polypeptide sequence (Fig. 1). Our results show that AtSOS1 Y346R exhibits increased tolerance to Li+ while maintaining similar Na+ resistance compared to wild-type AtSOS1. Since Tyr346 is not directly involved in metal ion coordination, this improved Li+ transport may result from an indirect effect of the substitution. In A. thaliana, Tyr346 is located near the upstream lysine residue (Lys343). Replacing Tyr346 with arginine increases the positive charge density at this position, potentially causing two effects. First, the positive charges of Lys343 and Arg346 may repel each other, pushing the arginine side chain away from lysine. Flexibility in this region could facilitate this effect. Interestingly, structural comparison shows that the corresponding loop in yeast plasma membrane Na+/H+ antiporter contains large amino acids such as tryptophan, phenylalanine and proline residues making it less flexible [29]. Second, Arg346 may interact with the membrane lipid head groups, possibly restricting the TM10-TM11 loop’s movement and widening the extracellular pore opening. This structural adjustment could enhance Li+ transport specificity. Additionally, two hydrophobic residues are located just upstream to the substituted Tyr346Arg, isoleucine (Ile344) and alanine (Ala345). Previous studies suggest that Li+ may have a stronger affinity for hydrophobic surfaces [39]. Furthermore, Na+ has a larger hydration sphere than Li+, potentially requiring more protein interactions and coordination for effective transport. In contrast, the wider extracellular pore created by the Tyr346Arg substitution may better accommodate ions with smaller hydration radii, favoring Li+ transport. This is consistent with previous findings that ion hydration radii influence ion selectivity in ion channels [40].
While there was no improvement in NaCl tolerance with the Y346R mutation, there was a consistent minor effect with the Y346F mutation and a minor effect on NaCl tolerance with the Y346A mutation. As noted above with the L375I mutation, others found differing results [21]. Again, we suggest that the beneficial effect of changing these amino acids may be species specific, and not transferrable to the SOS1 protein expressed in our system. This makes these mutations less desirable target as a method of increasing salt tolerance in many plant species.
Table 2.
Oligonucleotides used for site-directed mutagenesis of SOS1. Mutated nucleotides are indicated in lowercase. Codons of changed amino acids are indicated in boldface type. Restriction sites introduced are underlined, and wherever indicated (-) a site was removed. Only the forward oligonucleotide of the pairs used for mutagenesis is shown.
Table 2.
Oligonucleotides used for site-directed mutagenesis of SOS1. Mutated nucleotides are indicated in lowercase. Codons of changed amino acids are indicated in boldface type. Restriction sites introduced are underlined, and wherever indicated (-) a site was removed. Only the forward oligonucleotide of the pairs used for mutagenesis is shown.
| Mutation | Oligonucleotide | Restriction Site |
| E261K | CAATGACACTGTtATAaAGATTACTCTTACAATTGC | Psi1 |
| Y346A | AGTGATAAGATTGCCgcaCAAGGGAAcTCATGGCGATTTC | -EcoR1 |
| Y346F | AGTGATAAGATTGCCTtcCAAGGGAAcTCATGG | -EcoR1 |
| Y346K | AGTGATAAGATTGCCaagCAAGGGAAcTCATGGCGATTTC | -EcoR1 |
| Y346R | GATAAGATTGCCcgCCAAGGGAAcTCATGGCGATTTC | -EcoR1 |
| L375I | GGAGTTCTATATCCAaTTcTgTGcagaTTTGGCTATGGTTTG | BsgI |
| Q362L | CTATACGTTTACATCCtcCTcTCGCGTGTTGTTG | BseR1 |
| A399V | GGTTTGAGGGGCgTcGTGGCTCTTGCAC | -Bts1 |
| E261KA399V | caatgacactgttataaAgattactcttacaattgcGGTTTGAGGGGCgTcGTGGCTCTTGCAC | Psi & Bts1 |
4. Materials and Methods
4.1. Materials
Restriction enzymes were purchased from New England Biolabs, Inc. (Mississauga ON, Canada). PWO DNA polymerase was purchased from Roche Applied Science (Roche Molecular Biochemicals, Mannheim, Germany). Other chemicals were of analytical grade and from Fisher Scientific (Ottawa, ON), Sigma or BDH (Toronto, ON).
4.2. Plasmids and Site-Directed Mutagenesis
The SOS1 expression plasmid used in this study (pREP41SOSSGFP) was described earlier [20]. Briefly, SOS1 was cloned into the plasmid pREP41GFP [41,42]. The plasmid contains a Gly-Ala linker preceding GFP which contains the Ser65Thr mutation and has a NdeI site removed with a silent mutation. This “short” plasmid (referred to as SOS1short or SOS1s) construct expresses amino acids 1-481 of SOS1 fused to GFP. It was used for all mutagenesis and expression experiments with SOS1. The plasmid pREP41sod2GFP [26] was used as a positive control and is a similar construct except expressing Sod2, the native Na+/H+ antiporter of S. pombe, which we have characterized earlier [27]. Mutations to the pREP41SOSGFP plasmid containing the shortened SOS1 were by PCR amplification using synthetic oligonucleotides (See Table 1) as described earlier [43]. Mutations were designed to create or remove a restriction site that was used in screening mutants. DNA sequencing confirmed the accuracy of the mutations and fidelity of the resultant plasmid.
To examine expression and activity of wild type and mutant SOS1 protein S. pombe with the sod2 gene disruption (sod2::ura4) was used to host transformations of yeast and where indicated transformation was with the positive control of the same plasmid with the sod2 insert [28]. The knockout strain was maintained on low sodium minimal KMAL medium or on yeast extract adenine (YEA) [28,44]. For growth curves 2 X 106 cells were used from an exponentially growing overnight culture to inoculate 2.5 ml of fresh media liquid. Cultures were grown at 30oC in a rotary shaker. At the times indicated cells were harvested and growth at A600 was determined. Growth curve experiments were determined in triplicate a minimum of three times. Growth on plates was determined in media supplemented with NaCl or LiCl at the concentrations indicated.
To examine growth on solid phase media, serial dilutions of cells were inoculated onto agar with KMA medium containing leucine supplemented with either NaCl or LiCl at the indicated concentrations as described earlier [26]. The pREP41sod2GFP plasmid containing sod2 without mutations [45] was used as a control.
Western Blotting Analysis- Western blot analysis of SOS1 compared the levels of protein expression in cell lysates from wild type and mutant SOS1 containing yeast [27,46]. Cell lysates were made from transformed yeast cultures. The yeast cultures were pelleted at 3500 x g, 10 min, then washed with double distilled water. Next, they were then resuspended in lysis buffer containing 50 mM Tris-HCl, 5 mM EDTA, pH 8.0, 1 mM dithiothreitol containing a protease inhibitor cocktail [47]. Cells were lysed by passage through an emulsiflex homogenizer at a pressure of 25000 psi. Non-lysed cells were pelleted at 3500 x g for 5 min, and the supernatant was centrifuged (14000 x g for 10 min). Enriched membranes in the supernatant were centrifuged at 100000 x g for 1 hr, then resuspended in 50 mM Tris-Cl pH 8.0, 150 mM NaCl, 1 mM EGTA, 5 mM EDTA, 1.0% (v/v) NP-40, 0.5% (w/v) deoxycholate and 0.1% (w/v) SDS. Equal amounts of up to 25 μg of samples was resolved on SDS/polyacrylamide gels (10%). Western blotting used as primary antibody an anti-GFP polyclonal antibody (a gift of Dr. Luc Berthiaume, Dept. of Cell Biology, University of Alberta). The secondary antibody was Li-Cor goat anti-rabbit antibody IRDye 680 conjugated, which was detected on a Li-Cor Odyssey Imager.
5. Conclusions
Overall, our results have shown several interesting novel findings. Most notable is the importance of the specific changes of amino acid Y346. There was a striking and dramatic effect on Na+ vs Li+ tolerance when this residue was changed to Arg. Other mutations of amino acids showed minor improvements on NaCl tolerance, such as the mutations Y346F, A399V, Y346F and Y346A. While these changes were not huge, the possible effects of even a slight improvement in agricultural plant growth in arid and saline soils, makes future investigation of these effects in plants of interest.
Author Contributions
“Conceptualization, L.F. and D.D.; methodology, L.F., A.A. and D.D; validation, L.F., A.A. and D.D formal analysis, L.F., A.A. and D.D investigation, L.F., A.A. and D.D.; resources, L.F. and D.D.; data curation, L.F., A.A. and D.D; writing—original draft preparation, L.F.; writing—review and editing, L.F., A.A. and D.D.; visualization, L.F., A.A. and D.D.; supervision, L.F., A.A. and D.D.; project administration, L.F.; funding acquisition, L.F. All authors have read and agreed to the published version of the manuscript.
Funding
This Research was supported by a NSERC grant to LF # 2020 – 03932.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data will be made available on request.
Acknowledgments
We give thanks for the technical assistance of Azan Khan and Mashal Shafi for their assistance with PCR and isolation of plasmids.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| SOS | Salt Overlay Sensitive |
| SOS1s | Salt Overlay Sensitive 1 protein shortened at the C-terminus |
| At | Arabidopsis thaliana |
| TM | Transmembrane |
| GFP | Green fluorescent protein |
References
- Frommer, W. B.; Ludewig, U.; Rentsch, D. Taking transgenic plants with a pinch of salt. Science 1999, 285(5431), 1222–3. [Google Scholar] [CrossRef]
- Apse, M. P.; Blumwald, E. Engineering salt tolerance in plants. Current opinion in biotechnology 2002, 13(2), 146–50. [Google Scholar] [CrossRef]
- Deinlein, U.; Stephan, A. B.; Horie, T.; Luo, W.; Xu, G.; Schroeder, J. I. Plant salt-tolerance mechanisms. Trends in plant science 2014, 19(6), 371–9. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: towards genetic engineering for stress tolerance. Planta 2003, 218(1), 1–14. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.; Pardo, J. M.; Batelli, G.; Van Oosten, M. J.; Bressan, R. A.; Li, X. The Salt Overly Sensitive (SOS) pathway: established and emerging roles. Mol Plant 2013, 6(2), 275–86. [Google Scholar] [CrossRef]
- Shi, H.; Quintero, F. J.; Pardo, J. M.; Zhu, J. K. The putative plasma membrane Na(+)/H(+) antiporter SOS1 controls long- distance Na(+) transport in plants. Plant Cell 2002, 14(2), 465–477. [Google Scholar] [CrossRef]
- Salazar, O. R.; Chen, K.; Melino, V. J.; Reddy, M. P.; Hribova, E.; Cizkova, J.; Berankova, D.; Arciniegas Vega, J. P.; Caceres Leal, L. M.; Aranda, M.; Jaremko, L.; Jaremko, M.; Fedoroff, N. V.; Tester, M.; Schmockel, S. M. SOS1 tonoplast neo-localization and the RGG protein SALTY are important in the extreme salinity tolerance of Salicornia bigelovii. Nat Commun 2024, 15(1), 4279. [Google Scholar] [CrossRef] [PubMed]
- Sze, H.; Chanroj, S. Plant Endomembrane Dynamics: Studies of K(+)/H(+) Antiporters Provide Insights on the Effects of pH and Ion Homeostasis. Plant physiology 2018, 177(3), 875–895. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Rosales, M. P.; Galvez, F. J.; Huertas, R.; Aranda, M. N.; Baghour, M.; Cagnac, O.; Venema, K. Plant NHX cation/proton antiporters. Plant Signal Behav 2009, 4(4), 265–76. [Google Scholar] [CrossRef]
- Hanin, M.; Ebel, C.; Ngom, M.; Laplaze, L.; Masmoudi, K. New Insights on Plant Salt Tolerance Mechanisms and Their Potential Use for Breeding. Front Plant Sci 2016, 7, 1787. [Google Scholar] [CrossRef]
- Yue, Y.; Zhang, M.; Zhang, J.; Duan, L.; Li, Z. SOS1 gene overexpression increased salt tolerance in transgenic tobacco by maintaining a higher K(+)/Na(+) ratio. J Plant Physiol 2012, 169(3), 255–61. [Google Scholar] [CrossRef]
- Shi, H.; Lee, B. H.; Wu, S. J.; Zhu, J. K. Overexpression of a plasma membrane Na+/H+ antiporter gene improves salt tolerance in Arabidopsis thaliana. Nature biotechnology 2003, 21(1), 81–5. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Chen, Z. Z.; Zhou, X. F.; Yin, H. B.; Li, X.; Xin, X. F.; Hong, X. H.; Zhu, J. K.; Gong, Z. , Overexpression of SOS (Salt Overly Sensitive) genes increases salt tolerance in transgenic Arabidopsis. Mol Plant 2009, 2(1), 22–31. [Google Scholar] [CrossRef] [PubMed]
- Duscha, K.; Martins Rodrigues, C.; Muller, M.; Wartenberg, R.; Fliegel, L.; Deitmer, J. W.; Jung, M.; Zimmermann, R.; Neuhaus, H. E., 14-3-3 Proteins and Other Candidates form Protein-Protein Interactions with the Cytosolic C-terminal End of SOS1 Affecting Its Transport Activity. International journal of molecular sciences 2020, 21, (9). [CrossRef]
- El Mahi, H.; Perez-Hormaeche, J.; De Luca, A.; Villalta, I.; Espartero, J.; Gamez-Arjona, F.; Fernandez, J. L.; Bundo, M.; Mendoza, I.; Mieulet, D.; Lalanne, E.; Lee, S. Y.; Yun, D. J.; Guiderdoni, E.; Aguilar, M.; Leidi, E. O.; Pardo, J. M.; Quintero, F. J. A Critical Role of Sodium Flux via the Plasma Membrane Na(+)/H(+) Exchanger SOS1 in the Salt Tolerance of Rice. Plant physiology 2019, 180(2), 1046–1065. [Google Scholar] [CrossRef] [PubMed]
- Oh, D. H.; Leidi, E.; Zhang, Q.; Hwang, S. M.; Li, Y.; Quintero, F. J.; Jiang, X.; D'Urzo, M. P.; Lee, S. Y.; Zhao, Y.; Bahk, J. D.; Bressan, R. A.; Yun, D. J.; Pardo, J. M.; Bohnert, H. J. Loss of halophytism by interference with SOS1 expression. Plant physiology 2009, 151(1), 210–22. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Guan, C.; Wang, P.; Ma, Q.; Bao, A. K.; Zhang, J. L.; Wang, S. M., The Effect of AtHKT1;1 or AtSOS1 Mutation on the Expressions of Na(+) or K(+) Transporter Genes and Ion Homeostasis in Arabidopsis thaliana under Salt Stress. International journal of molecular sciences 2019, 20, (5). [CrossRef]
- Wu, S. J.; Ding, L.; Zhu, J. K. SOS1, a Genetic Locus Essential for Salt Tolerance and Potassium Acquisition. Plant Cell 1996, 8(4), 617–627. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.-P.; McCullough, N.; Martel, R.; Hemmingsen, S.; Young, P. G. Gene amplification at a locus encoding a putative Na+/H+ antiporter confers sodium and lithium tolerance in fission yeast. EMBO J. 1992, 11, 1631–1640. [Google Scholar] [CrossRef]
- Ullah, A.; Dutta, D.; Fliegel, L., Expression and characterization of the SOS1 Arabidopsis salt tolerance protein. Mol Cell Biochem 2016, 415, (1-2), 133-43. [CrossRef]
- Quintero, F. J.; Martinez-Atienza, J.; Villalta, I.; Jiang, X.; Kim, W. Y.; Ali, Z.; Fujii, H.; Mendoza, I.; Yun, D. J.; Zhu, J. K.; Pardo, J. M. Activation of the plasma membrane Na/H antiporter Salt-Overly-Sensitive 1 (SOS1) by phosphorylation of an auto-inhibitory C-terminal domain. Proc Natl Acad Sci U S A 2011, 108(6), 2611–6. [Google Scholar] [CrossRef] [PubMed]
- Meng, K.; Wu, Y. , Footprints of divergent evolution in two Na+/H+ type antiporter gene families (NHX and SOS1) in the genus Populus. Tree Physiol 2018, 38(6), 813–824. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X. Y.; Tang, L. H.; Nie, J. W.; Zhang, C. R.; Han, X.; Li, Q. Y.; Qin, L.; Wang, M. H.; Huang, X.; Yu, F.; Su, M.; Wang, Y.; Xu, R. M.; Guo, Y.; Xie, Q.; Chen, Y. H. Structure and activation mechanism of the rice Salt Overly Sensitive 1 (SOS1) Na(+)/H(+) antiporter. Nat Plants 2023, 9(1), 1924–1936. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Pan, C.; Chen, Q.; Xie, Q.; Gao, Y.; He, L.; Li, Y.; Dong, Y.; Jiang, X.; Zhao, Y. Architecture and autoinhibitory mechanism of the plasma membrane Na(+)/H(+) antiporter SOS1 in Arabidopsis. Nat Commun 2023, 14(1), 4487. [Google Scholar] [CrossRef] [PubMed]
- Nunez-Ramirez, R.; Sanchez-Barrena, M. J.; Villalta, I.; Vega, J. F.; Pardo, J. M.; Quintero, F. J.; Martinez-Salazar, J.; Albert, A. Structural insights on the plant salt-overly-sensitive 1 (SOS1) Na(+)/H(+) antiporter. Journal Mol Biol 2012, 424(5), 283–94. [Google Scholar] [CrossRef] [PubMed]
- Dutta, D.; Ullah, A.; Bibi, S.; Fliegel, L. Functional Analysis of Conserved Transmembrane Charged Residues and a Yeast Specific Extracellular Loop of the Plasma Membrane Na(+)/H(+) Antiporter of Schizosaccharomyces pombe. Sci Rep 2019, 9(1), 6191. [Google Scholar] [CrossRef]
- Dutta, D.; Shin, K.; Rainey, J. K.; Fliegel, L. Transmembrane Segment XI of the Na(+)/H(+) Antiporter of S. pombe is a Critical Part of the Ion Translocation Pore. Sci Rep 2017, 7(1), 12793. [Google Scholar] [CrossRef] [PubMed]
- Dibrov, P.; Young, P. G.; Fliegel, L. Functional analysis of amino acid residues essential for activity in the Na+/H+ exchanger of fission yeast. Biochemistry 1998, 36, 8282–8288. [Google Scholar] [CrossRef] [PubMed]
- Dutta, D.; Fliegel, L. Structure and function of yeast and fungal Na(+) /H(+) antiporters. IUBMB life 2018, 70(1), 23–31. [Google Scholar] [CrossRef]
- Sugiura, Y.; Saito, Y.; Endo, T.; Makita, Y., Effect of the Ionic Radius of Alkali Metal Ions on Octacalcium Phosphate Formation via Different Substitution ModesC. Crystal Growth & Design 2019, 19, (7), 4162-4171.
- Shi, H.; Ishitani, M.; Kim, C.; Zhu, J. K. The Arabidopsis thaliana salt tolerance gene SOS1 encodes a putative Na+/H+ antiporter. PNAS U S A 2000, 97(12), 6896–6901. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wang, D.; Dong, W.; Song, Z.; Dou, K. Over-expression of Na(+)/H (+) exchanger 1 and its clinicopathologic significance in hepatocellular carcinoma. Med Oncol 2009. [Google Scholar] [CrossRef] [PubMed]
- Shang, G. A. O.; Li, Y. U. A. N.; Hong, Z. H. A. I.; LIU, C. L.; HE, S. Z.; LIU, Q. C., Overexpression of SOS genes enhanced salt tolerance in sweetpotato. Journal of Integrative Agriculture, 11(3), 378-386. Journal of Integrative Agriculture 2012, 11, (3), 378-386.
- Dutta, D.; Esmaili, M.; Overduin, M.; Fliegel, L. Expression and detergent free purification and reconstitution of the plant plasma membrane Na(+)/H(+) antiporter SOS1 overexpressed in Pichia pastoris. Biochim Biophys Acta Biomembr 2020, 1862(3), 183111. [Google Scholar] [CrossRef]
- Hahnenberger, K. M.; Jia, Z.; Young, P. G. Functional expression of the Schizosaccharomyces pombe Na+/H+ antiporter gene, sod2, in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 1996, 93, 5031–5036. [Google Scholar] [CrossRef]
- Padan, E. The enlightening encounter between structure and function in the NhaA Na+-H+ antiporter. Trends Biochem Sci 2008, 33(9), 435–43. [Google Scholar] [CrossRef] [PubMed]
- Rimon, A.; Amartely, H.; Padan, E. The crossing of two unwound transmembrane regions that is the hallmark of the NhaA structural fold is critical for antiporter activity. Sci Rep 2024, 14(1), 5915. [Google Scholar] [CrossRef] [PubMed]
- Padan, E.; Kozachkov, L.; Herz, K.; Rimon, A. NhaA crystal structure: functional-structural insights. J Expl Biol 2009, 212 Pt 11, 1593–603. [Google Scholar] [CrossRef]
- Beauchamp, D. L.; Khajehpour, M., The effect of lithium ions on the hydrophobic effect: does lithium affect hydrophobicity differently than other ions? Biophys Chem 2012, 163-164, 35-43.
- Li, H.; Francisco, J. S.; Zeng, X. C. Unraveling the mechanism of selective ion transport in hydrophobic subnanometer channels. Proc Natl Acad Sci U S A 2015, 112(35), 10851–6. [Google Scholar] [CrossRef]
- Fliegel, L. Functional analysis of conserved polar residues important for salt tolerance of the Na+/H+ exchanger of Schizosaccharomyces pombe. Mol. Cell. Biochem. 2004, 43, 16477–16486. [Google Scholar]
- Fliegel, L. The Na+/H+ exchanger isoform 1. Int. J. Biochem. Cell Biol. 2005, 37(1), 33–37. [Google Scholar] [CrossRef]
- Ullah, A.; El-Magd, R. A.; Fliegel, L. Functional role and analysis of cysteine residues of the salt tolerance protein Sod2. Mol Cell Biochem 2014, 386(1-2), 85–98. [Google Scholar] [CrossRef]
- Ndayizeye, M.; Touret, N.; Fliegel, L. Proline 146 is critical to the structure, function and targeting of sod2, the Na+/H+ exchanger of Schizosaccharomyces pombe. Biochim Biophys Acta 2009, 1788(5), 983–92. [Google Scholar] [CrossRef]
- Fliegel, L.; Wiebe, C.; Chua, G.; Young, P. G. Functional expression and cellular localization of the Na+/H+ exchanger Sod2 of the fission yeast Schizosaccharomyces pombe. Canadian journal of physiology and pharmacology 2005, 83(7), 565–72. [Google Scholar] [CrossRef]
- Slepkov, E. R.; Chow, S.; Lemieux, M. J.; Fliegel, L. Proline residues in transmembrane segment IV are critical for activity, expression and targeting of the Na+/H+ exchanger isoform 1. Biochem J 2004, 379 Pt 1, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Silva, N. L.; Wang, H.; Harris, C. V.; Singh, D.; Fliegel, L. Characterization of the Na+/H+ exchanger in human choriocarcinoma (BeWo) cells. Pflugers Archiv Eur J Physiol 1997, 433, 792–802. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D. M. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol Biol Evol 2013, 30(4), 772–80. [Google Scholar] [CrossRef]
- Robert, X.; Gouet, P., Deciphering key features in protein structures with the new ENDscript server. Nucleic acids research 2014, 42, (Web Server issue), W320-4. [CrossRef]
- DeLano, W. L. Pymol: An open-source molecular graphics tool. CCP4 Newsl. Protein Crystallogr 2002, 82(1), 82–92. [Google Scholar]
Figure 1.
Analysis of Arabidopsis thaliana (At) SOS1 protein. (A) Multiple sequence alignment of plant SOS1 demonstrating the segments harboring A. thaliana SOS1 amino acids investigated in this manuscript. The same amino acids are shown using an asterisk. The red background white colored amino acids are conserved amino acids. The red colored amino acids are amino acids of similar types. TM# denotes the transmembrane segments. The coils on the top of the alignment represent the helices of the A. thaliana SOS1 structure. Amino acid numbers are indicated at left and are aligned with Arabidopsis thaliana SOS1 sequence beginning at amino acid 253, 335 and 372. At, Arabidopsis thaliana (NP_178307.2); Th, Thellungiella halophila (ABN04857.1); Sl, Solanum lycopersicum (NP_001234698.2); Gh, Gossypium hirsutum (AMY98958.1); Gb, Gossypium barbadense (A0A5J5SE13); Pt, Populus trichocarpa (UniRef100_U5G5K6); Pe, Populus euphratica (DQ517530); Nt, Nitraria tangutorum (AGW30210.1); Pd, Phoenix dactylifera (XP_008798100.1); Ta, Triticum aestivum (Q4L224); Os, Oryza sativa (Q5ICN3). The red background resides are conserved residues. The red colour residues are similar type amino acids. The residues subjected to investigation are highlighted using asterisk. The transmembrane segments correspond to a particular amino acid stretch are mentioned on the top. The square box indicates the second segment of the alignment. B and C, Cryogenic electron microscopy structure of the AtSOS1s (amino acid 32 – 481) dimer (modified from the PDB 7Y3E) [23,24]. The N-terminal residues (1 – 31) are not shown for clarity. Two different color sets are used to refer to each monomer of the dimer chain A (magenta and cyan) and chain B (orange and blue). The dimerization domains (cyan and blue) and transport domains (magenta and orange) are highlighted. (B) view of AtSOS1 parallel to the membrane plane. (C) view of AtSOS1 perpendicular to the membrane from the extracellular side.
Figure 1.
Analysis of Arabidopsis thaliana (At) SOS1 protein. (A) Multiple sequence alignment of plant SOS1 demonstrating the segments harboring A. thaliana SOS1 amino acids investigated in this manuscript. The same amino acids are shown using an asterisk. The red background white colored amino acids are conserved amino acids. The red colored amino acids are amino acids of similar types. TM# denotes the transmembrane segments. The coils on the top of the alignment represent the helices of the A. thaliana SOS1 structure. Amino acid numbers are indicated at left and are aligned with Arabidopsis thaliana SOS1 sequence beginning at amino acid 253, 335 and 372. At, Arabidopsis thaliana (NP_178307.2); Th, Thellungiella halophila (ABN04857.1); Sl, Solanum lycopersicum (NP_001234698.2); Gh, Gossypium hirsutum (AMY98958.1); Gb, Gossypium barbadense (A0A5J5SE13); Pt, Populus trichocarpa (UniRef100_U5G5K6); Pe, Populus euphratica (DQ517530); Nt, Nitraria tangutorum (AGW30210.1); Pd, Phoenix dactylifera (XP_008798100.1); Ta, Triticum aestivum (Q4L224); Os, Oryza sativa (Q5ICN3). The red background resides are conserved residues. The red colour residues are similar type amino acids. The residues subjected to investigation are highlighted using asterisk. The transmembrane segments correspond to a particular amino acid stretch are mentioned on the top. The square box indicates the second segment of the alignment. B and C, Cryogenic electron microscopy structure of the AtSOS1s (amino acid 32 – 481) dimer (modified from the PDB 7Y3E) [23,24]. The N-terminal residues (1 – 31) are not shown for clarity. Two different color sets are used to refer to each monomer of the dimer chain A (magenta and cyan) and chain B (orange and blue). The dimerization domains (cyan and blue) and transport domains (magenta and orange) are highlighted. (B) view of AtSOS1 parallel to the membrane plane. (C) view of AtSOS1 perpendicular to the membrane from the extracellular side.
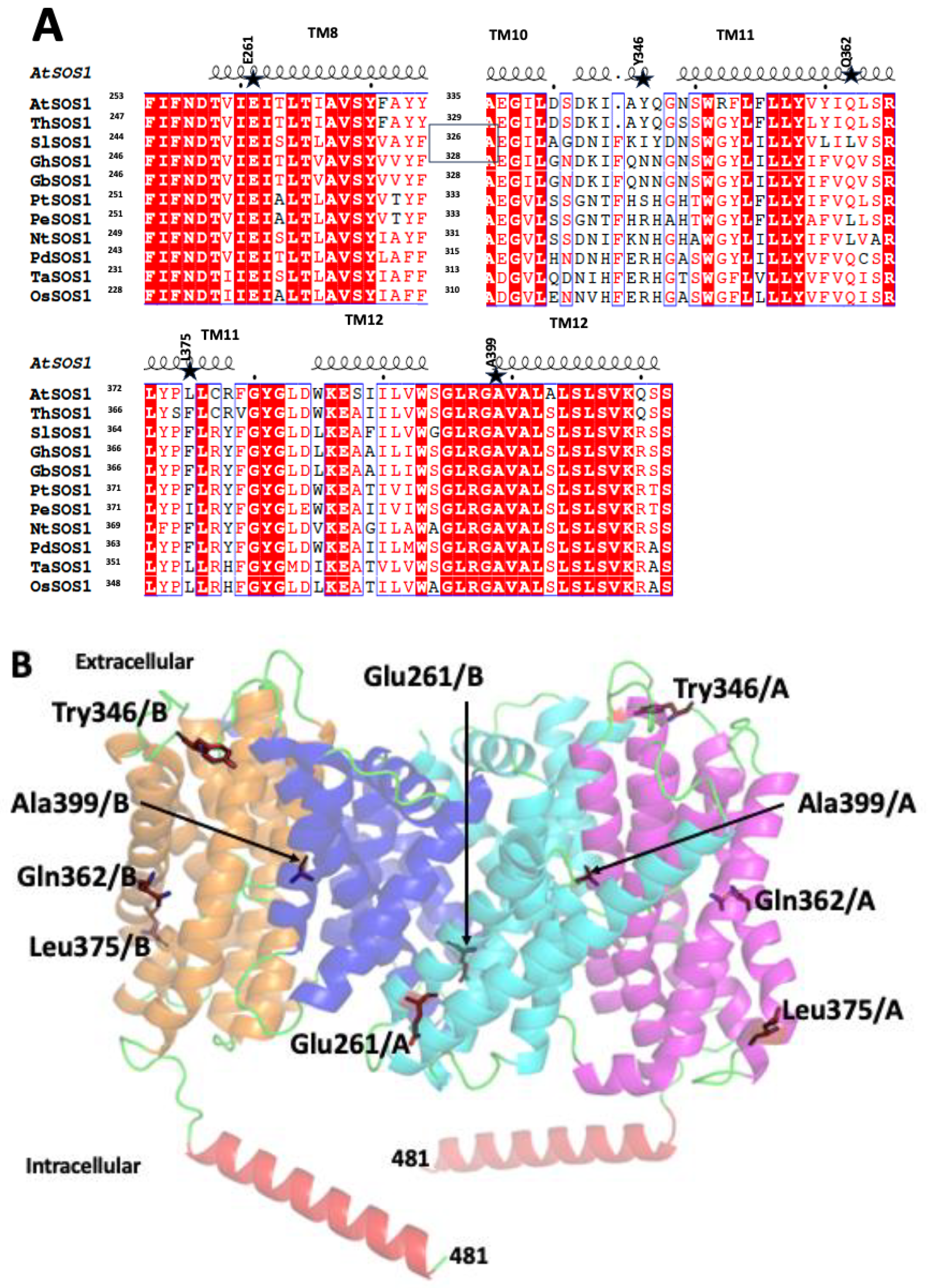
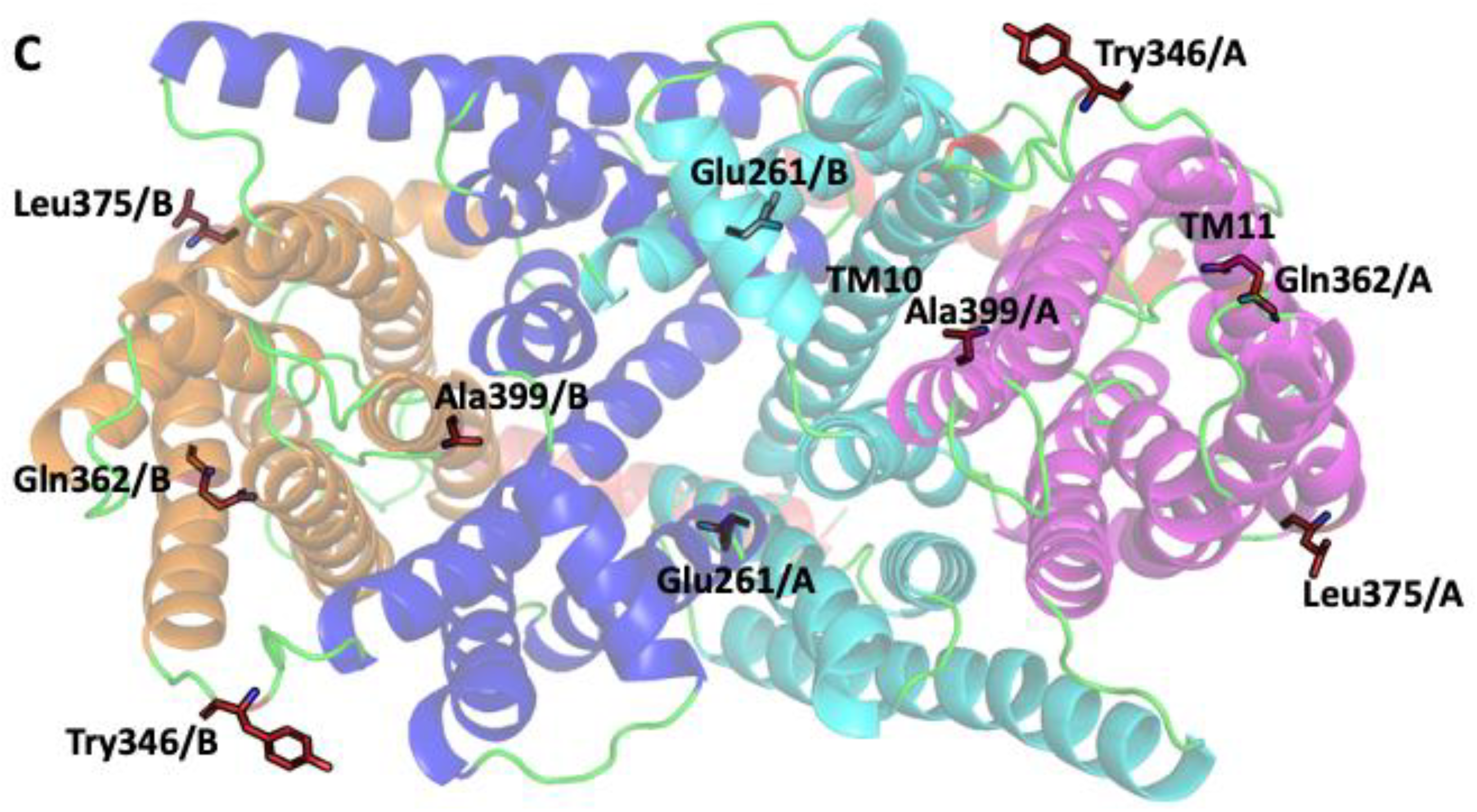
Figure 2.
Close up view of the residues subjected to investigation. A. thaliana SOS1 (PDB 7Y3E) is used for the representation. The mutated residues are shown in firebrick color. Residues from the loop are shown in green where the magenta and cyan colors represent transport and dimerization domains respectively. (A) Glu261 and its interacting distance with the Lys304 and Ser307. (B) Proximity of Gln362 close to Trp363 and Asn440. (C) Tyr346 is shown residing on the extracellular loop joining the TM10 and TM11. TM10 and TM11 are the parts of the dimerization domain and the transport domain respectively. In polypeptide chain, Tyr346 is preceded by the residues Lys343, Ile344, and Ala345. (D) Leu375 side chain is surrounded by the hydrophobic side residues such as Leu131, Leu372, and Phe379. (E) Ala399 is located at the center of the protein and is located at the discontinued helices of the transport domain.
Figure 2.
Close up view of the residues subjected to investigation. A. thaliana SOS1 (PDB 7Y3E) is used for the representation. The mutated residues are shown in firebrick color. Residues from the loop are shown in green where the magenta and cyan colors represent transport and dimerization domains respectively. (A) Glu261 and its interacting distance with the Lys304 and Ser307. (B) Proximity of Gln362 close to Trp363 and Asn440. (C) Tyr346 is shown residing on the extracellular loop joining the TM10 and TM11. TM10 and TM11 are the parts of the dimerization domain and the transport domain respectively. In polypeptide chain, Tyr346 is preceded by the residues Lys343, Ile344, and Ala345. (D) Leu375 side chain is surrounded by the hydrophobic side residues such as Leu131, Leu372, and Phe379. (E) Ala399 is located at the center of the protein and is located at the discontinued helices of the transport domain.
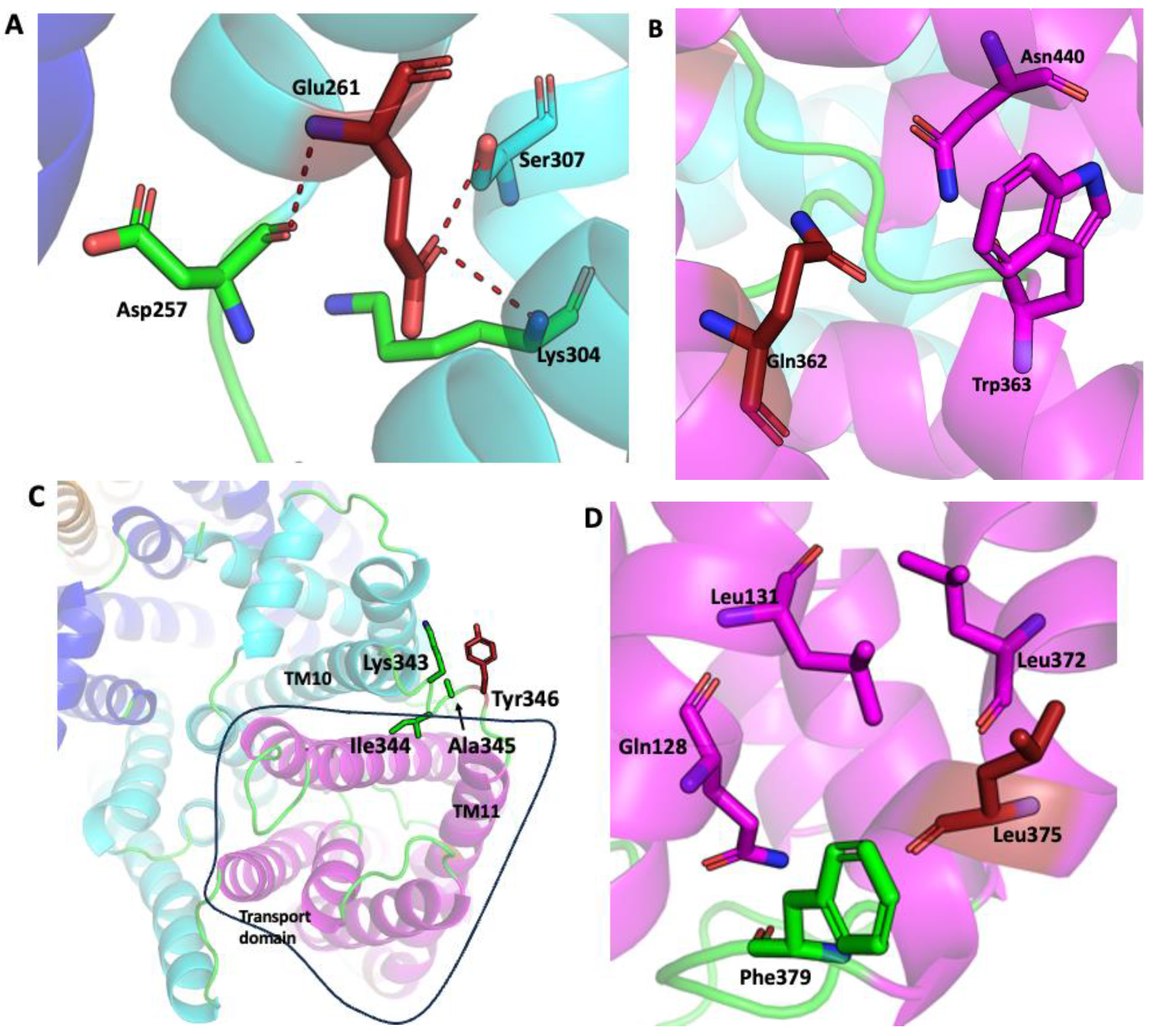
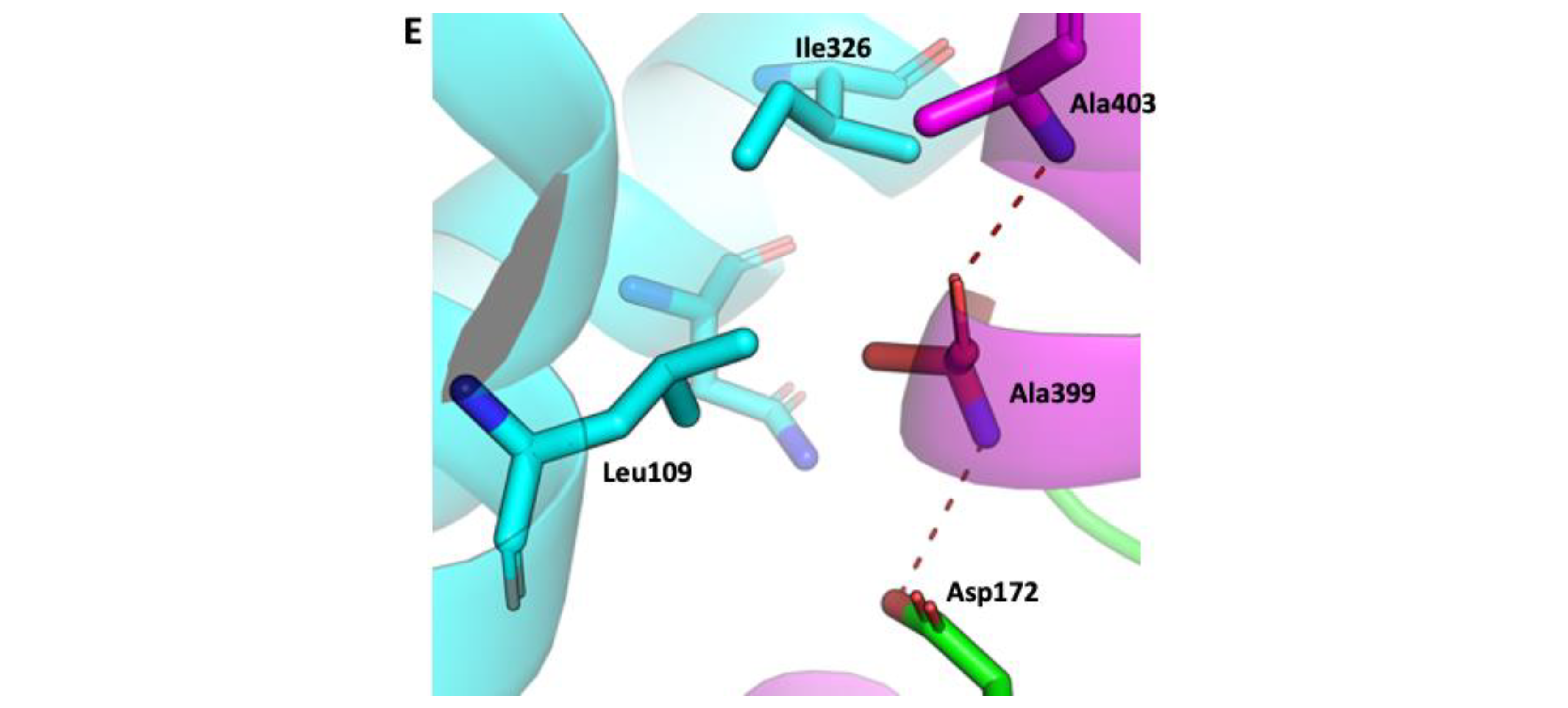
Figure 3.
Expression and of various length SOS1 proteins. (A, B) Western blot analysis of expression of SOS1s proteins. Equal amounts of cell extracts from S. pombe strains expressing various SOS1s-GFP constructs were blotted with anti GFP antibody as described in the materials and methods. SOS1s is the wild type (shortened) protein and Sod2Ura4 is the knockout strain. Other lanes are SOS1s protein with the indicated mutation.
Figure 3.
Expression and of various length SOS1 proteins. (A, B) Western blot analysis of expression of SOS1s proteins. Equal amounts of cell extracts from S. pombe strains expressing various SOS1s-GFP constructs were blotted with anti GFP antibody as described in the materials and methods. SOS1s is the wild type (shortened) protein and Sod2Ura4 is the knockout strain. Other lanes are SOS1s protein with the indicated mutation.
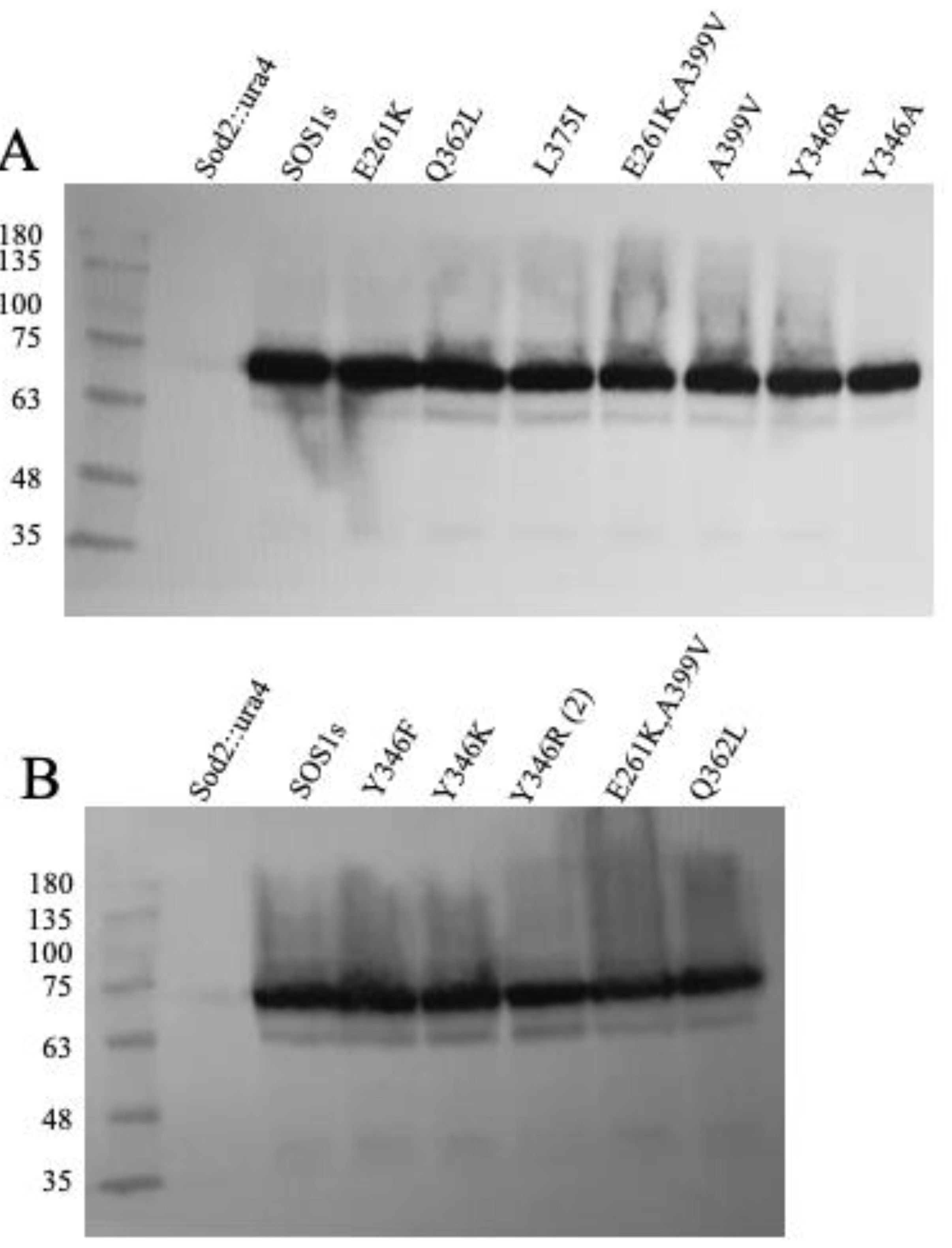
Figure 4.
Growth in liquid medium of sod2 knockout S. pombe and the same strain containing either wild type sod2 or SOS1s protein or SOS1s mutant proteins. LiCl (A-L) or NaCl (M-X) tolerance of strains was assessed by growing 2 X 106 of inoculated cells into 2.5 ml of medium at 30 oC for up to 120 hours. Growth was assessed by measuring the absorbance of the cell suspensions at 600 nm at the times indicated. Results are the mean +/- SE of at least three determinations. S. pombe were grown in the presence of 0, 2, 4, 5, 6 or 8 mM LiCl or 0, 0.2, 0.4, 0.5, 0.6 or 0.8 M NaCl as indicated. (A, M), growth rates of control, Sod2Ura4 cells. Sod2Ura4 refers to S. pombe with the sod2 knockout described earlier [28]. (B, N), S. pombe containing wild type sod2 and C, O, S. pombe containing wild type SOS1s. (B), as in A except in various LiCl containing medium as indicated. Other growth curves (D-L and P-X) are for yeast containing SOS1s with the indicated mutations. *, ^ significantly different from SOS1s mutant at P < 0.05 or 0.01 respectively.
Figure 4.
Growth in liquid medium of sod2 knockout S. pombe and the same strain containing either wild type sod2 or SOS1s protein or SOS1s mutant proteins. LiCl (A-L) or NaCl (M-X) tolerance of strains was assessed by growing 2 X 106 of inoculated cells into 2.5 ml of medium at 30 oC for up to 120 hours. Growth was assessed by measuring the absorbance of the cell suspensions at 600 nm at the times indicated. Results are the mean +/- SE of at least three determinations. S. pombe were grown in the presence of 0, 2, 4, 5, 6 or 8 mM LiCl or 0, 0.2, 0.4, 0.5, 0.6 or 0.8 M NaCl as indicated. (A, M), growth rates of control, Sod2Ura4 cells. Sod2Ura4 refers to S. pombe with the sod2 knockout described earlier [28]. (B, N), S. pombe containing wild type sod2 and C, O, S. pombe containing wild type SOS1s. (B), as in A except in various LiCl containing medium as indicated. Other growth curves (D-L and P-X) are for yeast containing SOS1s with the indicated mutations. *, ^ significantly different from SOS1s mutant at P < 0.05 or 0.01 respectively.
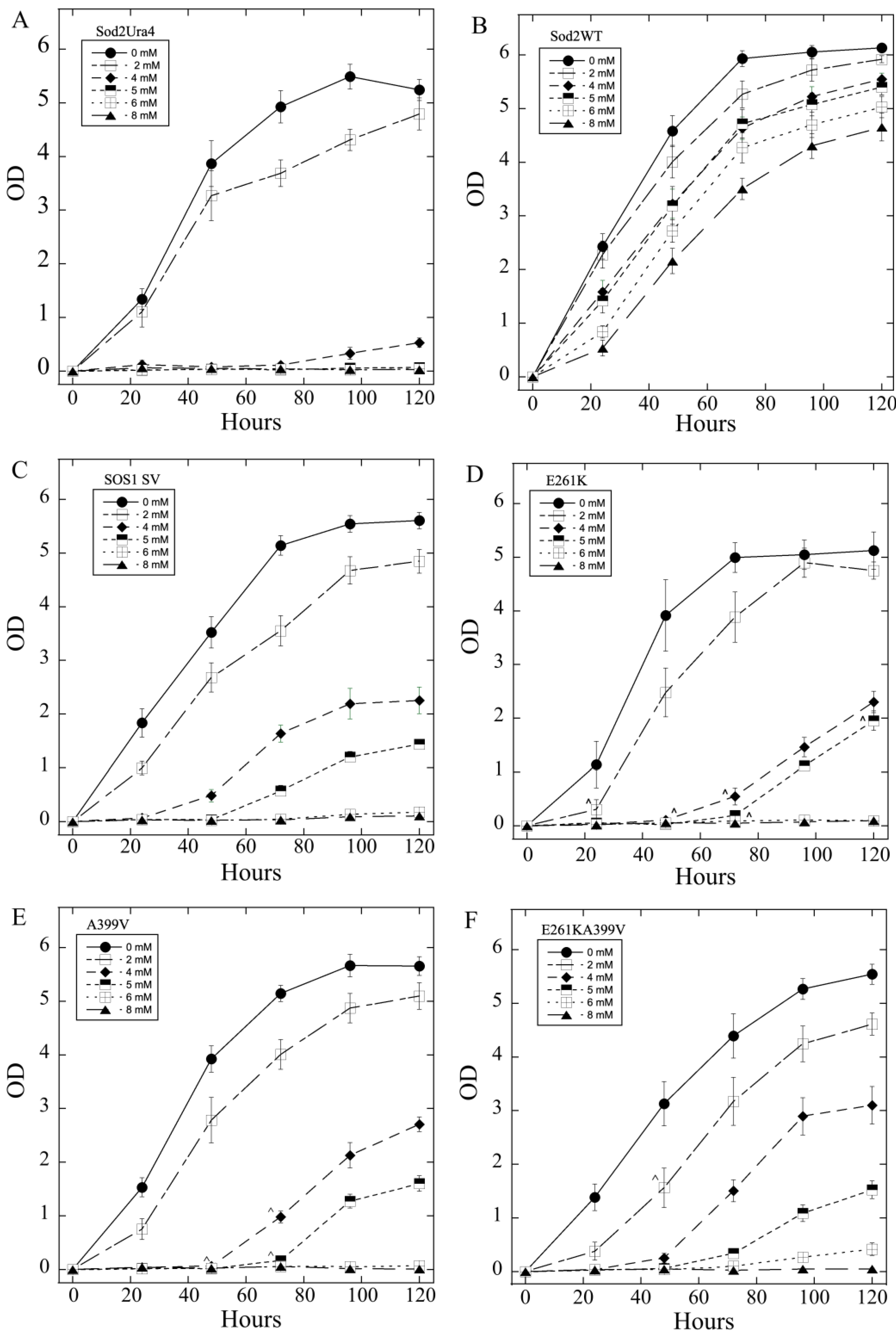
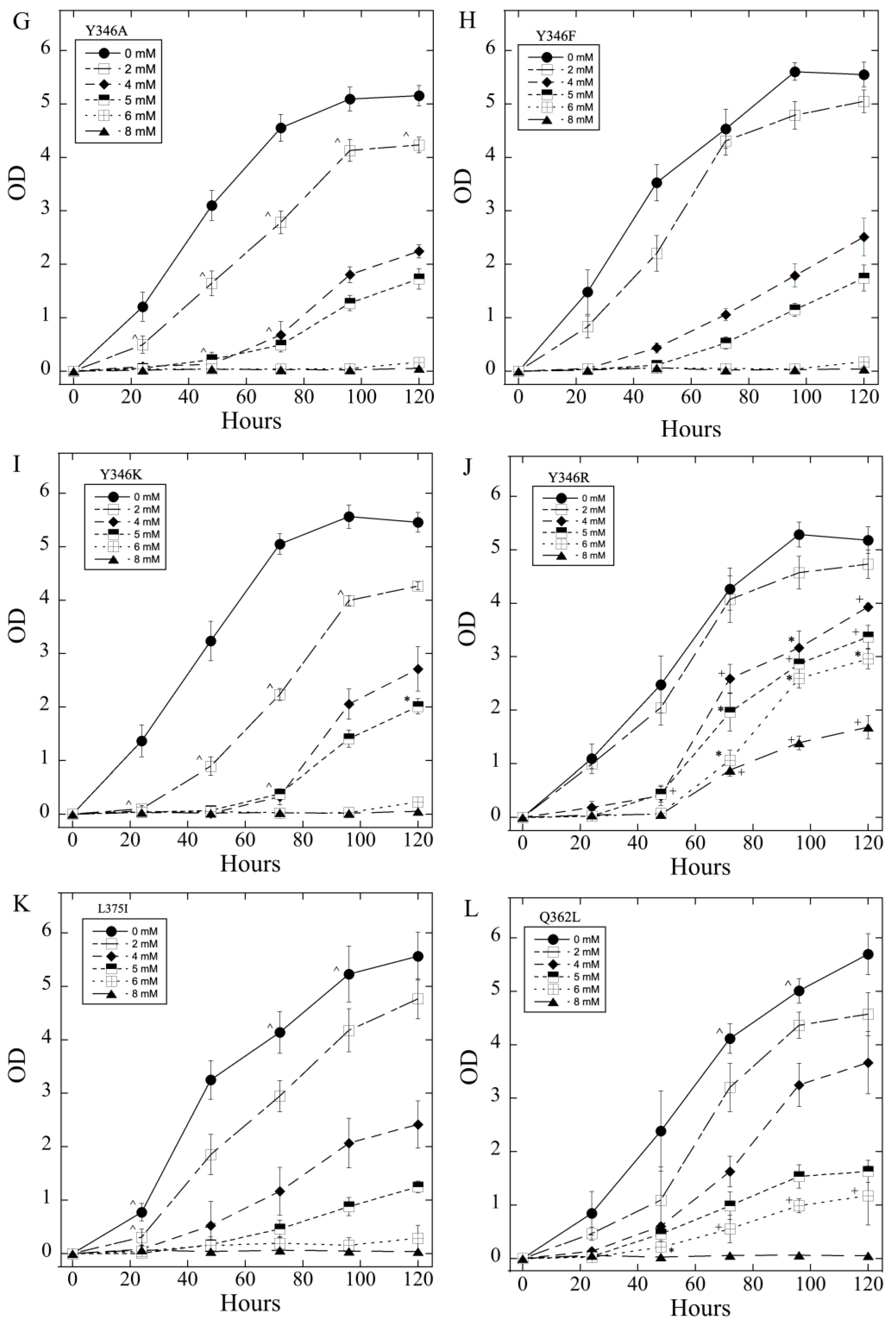
Figure 5.
Growth of wild type (WT) and mutant SOS1 containing S. pombe on solid media in the presence or absence of varying amounts of LiCl (A, B), or NaCl (C, D). Samples of the various strains were from stationary phase cultures that were serially diluted 1:10 repeatedly. They were spotted onto minimal media plates with the indicated concentrations of NaCl or LiCl. Plates were incubated for 72 hours at 30o C as described earlier [26]. S. pombe containing wild type Sod2 was used as a positive control. Results are typical of 3 repeats. KO, S. pombe containing the wild type sod2 knocked out as a negative control. (C, top left, shows growth on KMA plate without Leu. All other growth was done on KMA media plus Leu.).
Figure 5.
Growth of wild type (WT) and mutant SOS1 containing S. pombe on solid media in the presence or absence of varying amounts of LiCl (A, B), or NaCl (C, D). Samples of the various strains were from stationary phase cultures that were serially diluted 1:10 repeatedly. They were spotted onto minimal media plates with the indicated concentrations of NaCl or LiCl. Plates were incubated for 72 hours at 30o C as described earlier [26]. S. pombe containing wild type Sod2 was used as a positive control. Results are typical of 3 repeats. KO, S. pombe containing the wild type sod2 knocked out as a negative control. (C, top left, shows growth on KMA plate without Leu. All other growth was done on KMA media plus Leu.).
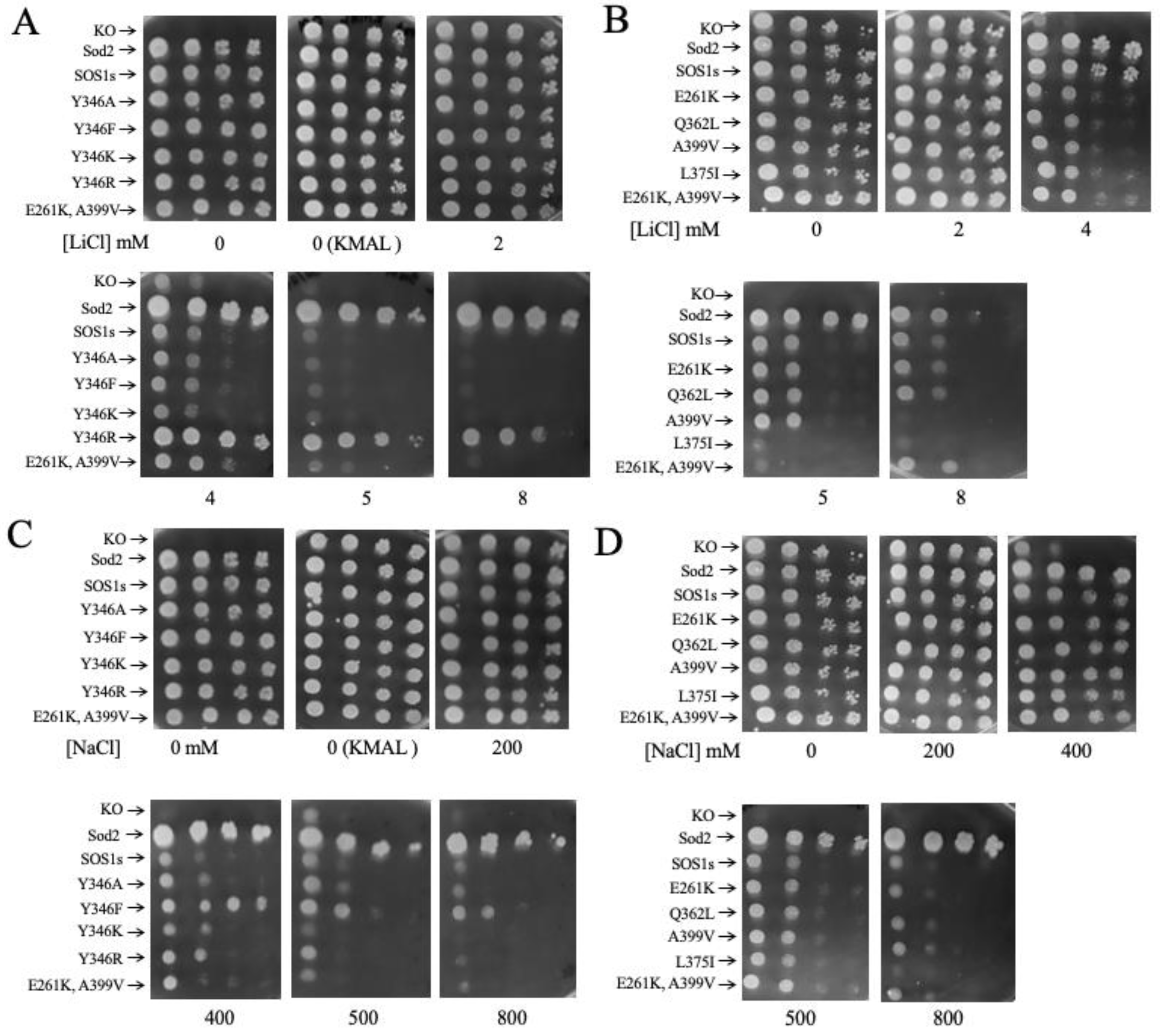
Figure 6.
Comparison of the structures of Oryza sativa SOS1 (residues up to 976) (OsSOS1) PDB:8J2M and full length Oryza sativa SOS1 PDB:8IWO [23]. The structure reveals the flexibility of the TM10 – TM11 loop harboring the residue Arg322, which is present at the corresponding positions of Arabidopsis thaliana SOS1 Try346.
Figure 6.
Comparison of the structures of Oryza sativa SOS1 (residues up to 976) (OsSOS1) PDB:8J2M and full length Oryza sativa SOS1 PDB:8IWO [23]. The structure reveals the flexibility of the TM10 – TM11 loop harboring the residue Arg322, which is present at the corresponding positions of Arabidopsis thaliana SOS1 Try346.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



