Submitted:
28 January 2025
Posted:
28 January 2025
You are already at the latest version
Abstract
Ovarian cancer is the leading cause of cancer-related death among women, which ranks as the eighth most common gynecological cancer. Although several cytoreductive surgeries and chemotherapies have been attempted to address ovarian cancer, the disease still shows poor prognosis and survival rate due to prevalent metastasis. Peritoneal metastasis is recognized as the primary route of metastatic progression in ovarian cancer, leading to severe symptoms in patients and is generally difficult to detect at an early stage. Current anti-cancer therapy is insufficient to completely treat ovarian cancer, particularly in cases where the disease has progressed to metastasis, due to its high rates of recurrence and resistance. Therefore, it is imperative to develop effective strategies for treating metastatic ovarian cancer. Given that various signaling pathways, including TGF-β, NF-κB, and PI3K/AKT/mTOR pathways, influence cancer progression, their interactions within the tumor microenvironment (TME) are particularly significant. TME plays a critical role in driving metastatic features in ovarian cancer through altered immunologic interactions. Recent therapeutic advances have focused on targeting these distinct features to improve treatment outcomes. Deciphering the complex interaction between signaling pathways and immune populations contributing to metastatic ovarian cancer provides an opportunity to enhance anti-cancer efficacy. Hereby, this review highlights the mechanisms of signaling pathways in metastatic ovarian cancer and immunological interactions to understand improved immunotherapy against ovarian cancer.
Keywords:
ovarian cancer
; peritoneal metastasis
; tropism
; signaling pathway
; tumor microenvironment
; immunotherapy
1. Introduction
Ovarian cancer (OC) is one of the most common gynecologic cancers and has the highest mortality rate due to prevalent malignant metastasis [1]. A hallmark of OC metastasis is its preferential spread to the peritoneal cavity, a process referred as peritoneal metastasis [2]. Unlike hematogenous dissemination common in other cancers, OC cells dissociate form the primary tumor, persist within the ascitic fluid, and adhere to the mesothelial layer of the peritoneum [3]. This process is facilitated by versatile interactions between tumor cells and the peritoneal microenvironment, including mesothelial cells, immune infiltrates, and extracellular matrix (ECM) components [4,5]. These metastatic mechanisms make peritoneal dissemination an important contributor to the poor prognosis of OC [5]. Despite continuous developments in diagnostic techniques, surgical interventions, and the advancement of therapeutic approaches, the five-year survival rate for OC remains unsatisfactory [6]. Currently, standard treatment, which combines maximal cytoreductive surgery and postoperative platinum-based chemotherapy, frequently leads to high recurrence rates and limited long-term benefits [7,8]. Moreover, the cumulative toxicity of chemotherapy often results in severe adverse effects, further deteriorating patient outcomes [9]. Considering these challenges, it is crucial to develop advanced therapeutic strategies targeting metastatic OC [10].
Metastatic OC arises from intricate signaling networks that not only regulate tumor progression but also confer resistance to existing treatments [11,12]. The transforming growth factor-β (TGF-β) pathway exerts contrasting roles in OC pathogenesis, serving as a tumor suppressor in the initial stages of the disease while adopting tumor-promoting characteristics in advanced metastatic stages [13]. Loss of tumor-suppressive TGF-β signaling promotes epithelial-to-mesenchymal transition (EMT), a critical process in peritoneal dissemination and metastasis [14]. In chemoresistant OC cells, TGF-β signaling is often impaired, particularly through the disruption of suppressor of mothers against decapentaplegic 3 (SMAD3) phosphorylation, reducing its tumor-suppressive effects [15]. Beyond its canonical SMAD-mediated pathway, TGF-β activates non-SMAD pathways, such as phosphoinositide 3-kinase (PI3K)/protein kinase B (AKT)/mammalian target of rapamycin (mTOR) and mitogen-activated protein kinase (MAPK)/extracellular signal-regulated kinase (ERK) cascades, which are essential for sustaining tumor cell survival, proliferation, and invasive traits [16,17,18]. Notably, the hyperactivation of the PI3K/AKT/mTOR axis is a representative feature of OC, resulting in chemoresistance by enhancing cellular growth, metabolic reprogramming, and autophagy [19,20]. This multifaceted participation of signaling transductions highlights its therapeutic significance in managing OC progression. The nuclear factor of kappa-light chain of enhancer-activated B cells (NF-κB) pathway also plays a pivotal role in OC metastasis by building the immunosuppressive tumor microenvironment (TME) [21]. Tumor-associated macrophages (TAMs) release tumor necrosis factor-alpha (TNF-α), creating a positive feedback loop that sustains NF-κB activation [22]. It promotes tumor metastasis, immune evasion, and cancer stem cell proliferation [23]. Elevated NF-κB signaling correlates with increased tumor aggressiveness and poor clinical outcomes, indicating the potential of signaling pathway as a therapeutic target. Concertedly, it is sophisticated that the critical roles of TGF-β, NF-κB, and PI3K/AKT/mTOR signaling transductions in inducing metastatic progression and resistance in OC, underlining the desire for improved therapeutic strategies targeting these networks.
Although immunotherapy has rapidly evolved as an anti-cancer treatment, its efficacy in OC has been limited, with clinical trials often reporting suboptimal response rates [24]. Conventional immunotherapy such as immune checkpoint inhibitor fails to adequately overcome the immunosuppressive barriers established by the OC TME [25]. It has prompted interest in combination therapies that integrate immunotherapy with targeted inhibitors of major signaling pathways including TGF-β, NF-κB, and PI3K/AKT/mTOR. Such approaches aim to disrupt the pro-metastatic and immunosuppressive signaling networks while enhancing anti-tumor immune responses [26].
In this review, we discuss the molecular mechanisms associated with OC metastasis, focusing on the vital signaling pathways that produce tumor progression and therapeutic resistance. Furthermore, this paper explores emerging strategies to address these pathways, with an emphasis on combination therapies that is promising for overcoming the present limitations of current treatments.
2. Ovarian Cancer Tropism
OC is characterized by its unique tropism and metastatic behavior, leading to poor survival and high recurrence rates. Understanding tropism in cancer cells provides valuable insights into the mechanisms of metastasis, particularly in explaining why cancer cells preferentially metastasize to specific organs [27]. Metastatic organotropism is regulated by molecular signaling pathways, tumor-intrinsic factors, and interactions with the TME [28]. OC metastasis often spreads from the primary tumor site to adjacent organs through three major routes: transcoelomic, lymphatic, and hematogenous dissemination [2] (Figure 1). Transcoelomic metastasis, facilitated by peritoneal fluid or ascites, is known to be predominant [29]. Due to the proximity of the peritoneal cavity to the ovaries and the absence of a physical barrier between the tumor and the peritoneum, this provides a preferential route for metastasis to the omentum or peritoneum [2]. Lymph nodes connected to tumor vascularization can act as drainage sites for cancer cells, allowing them to shed, circulate, and spread via the lymphatic system [30]. Hematogenous metastasis is the least common route of OC dissemination and is associated with the presence of circulating tumor cells in the bloodstream. However, the lack of effective methods for detecting circulating tumor has hindered a comprehensive understanding of the mechanisms underlying hematogenous spread [31].
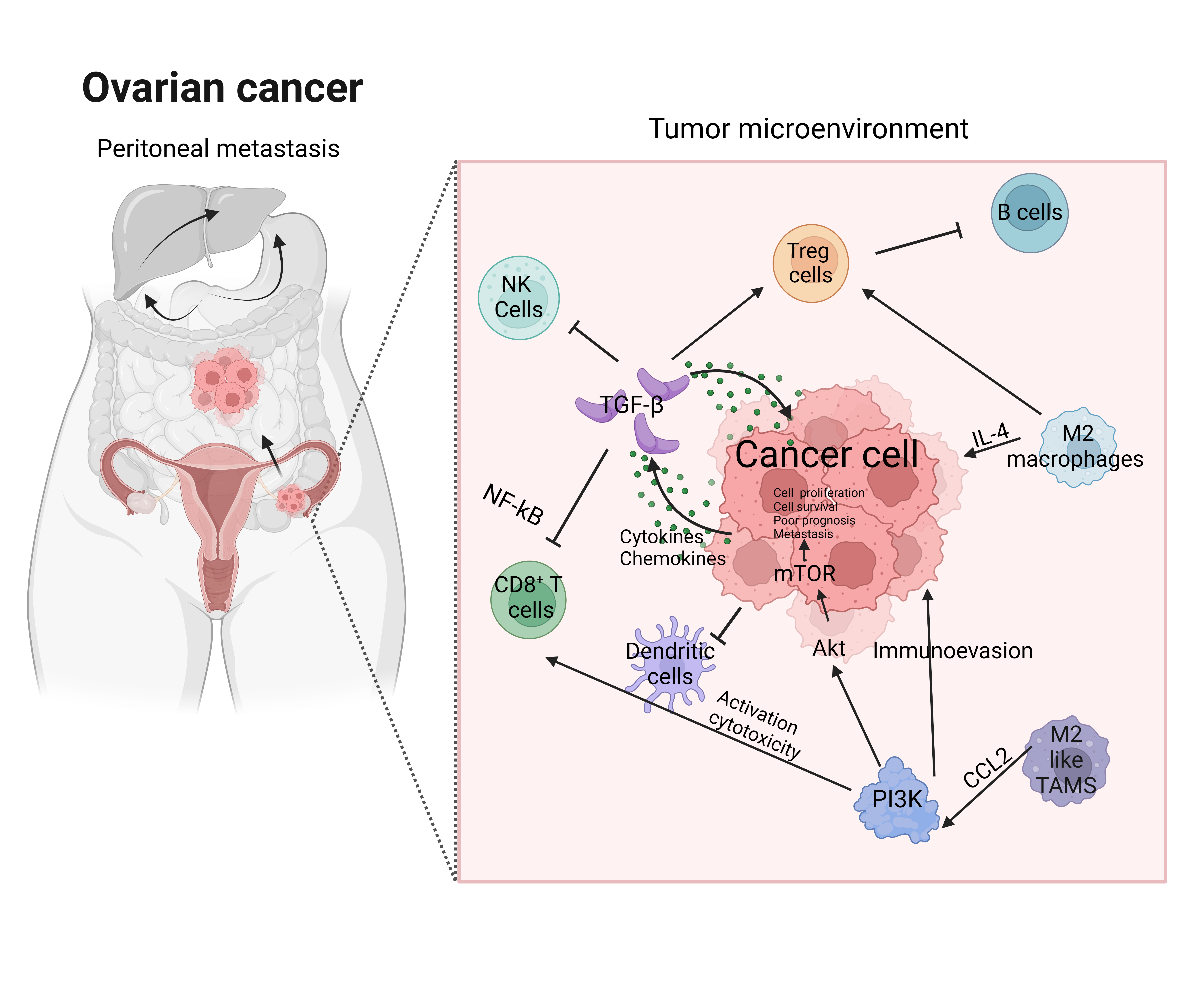
As mentioned above, peritoneal metastasis accounts for the majority of OC cases and involves specific processes, including separation from the primary tumor, spread to the peritoneum, colonization, and progression into metastatic tumors in the peritoneum [32]. The peritoneal membrane, composed of epithelial-like peritoneal mesothelial cells (PMCs) supported by an ECM-rich stroma, can play an essential role in tumor colonization [33]. This process is driven by integrins and ECM remodeling, which provide essential metabolic substrates, along with the immune landscape of the peritoneal cavity that shapes the metastatic niche [2,34]. The peritoneal immune microenvironment is dominated by cancer-associated fibroblasts (CAFs), TAMs, regulatory T cells (Tregs), and myeloid-derived suppressor cells (MDSCs), all of which contribute to immunosuppression [35,36]. CAFs contribute to OC peritoneal metastasis by promoting immune cell recruitment, ECM remodeling, and the secretion of cytokines and chemokines that rebuild the TME. In malignant ascites, CAFs express certain genes related to immune system, such as complement factors, chemokines (e.g., CXCL1/2/10/12), and cytokines (e.g., IL6, IL10), which subsequently enhance tumor formation, invasion, and progression through the activation cadherins and integrins [36]. TAMs in ascites facilitate tumor progression via ECM degradation, angiogenesis, and immune evasion through cytokine secretion such as IL-10 and TGF-β [37]. Tregs suppress cytotoxic T-cell activity of immune effector cells, thereby enabling tumor survival and proliferation [38]. Therefore, these immune populations generally induce a permissive environment that facilitates peritoneal metastasis. Given that OC exhibits versatile immune dysregulation and metastatic organotropism, it is required to elucidate the pivotal role of various signaling pathways regarding OC tropism and metastasis. Additionally, this understanding can offer potential therapeutic targets for addressing peritoneal metastasis.
3. Metastatic Signaling Pathways in Ovarian Cancer
The metastatic progression of OC is regulated by a complex network of signaling pathways related to EMT, immune evasion, and tumor development [39]. EMT, a hallmark of metastasis, enables epithelial cancer cells to acquire mesenchymal traits that promote their mobility and invasiveness [40]. This process is closely correlated with immune regulation and metabolic reprogramming within the TME, further reinforcing the metastatic potential of cancer cells [41]. Given that EMT is essential for cancer cells to migrate and settle in distant sites such as peritoneum, understanding EMT-associated signaling pathways is critical for unraveling the mechanisms of peritoneal dissemination in OC.
3.1. TGF-β Signaling Pathway
The TGF-β family plays a dual role in tumorigenesis, acting as a tumor suppressor in the early stages and as a promoter of metastasis in advanced stages [13]. In OC, TGF-β signaling functions as an early EMT inducer that regulates cancer cell metastasis and development [42]. EMT provides metastatic characteristics through the reversible transition of epithelial cells into mesenchymal cells. This process is characterized by the downregulation of transcriptional epithelial markers, such as E-cadherin, and the upregulation of mesenchymal markers, such as N-cadherin and vimentin [43]. TGF-β can impair the immune responses of NK cells, cytotoxic T lymphocytes (CTLs), and CD8+ T cells, allowing cancer cells to evade immune surveillance [44]. Within the TME, activated TGF-β signaling polarizes macrophages and neutrophils into TAMs, further facilitating immune evasion. Beyond immune suppression, TGF-β induces CAFs, which recruit tumor-promoting factors that, in turn, accelerate cancer progression [17]. TGF-β also contributes to tumor invasion and migration by promoting the detachment of OC cells from the primary tumor, especially in the absence of TGF-β receptors and SMAD proteins [17,44]. Stimulated TGF-β/SMAD signaling enhances peritoneal invasion through the secretion of matrix metalloproteinases (MMPs), which degrade the ECM and enable cancer cells to invade surrounding tissues [45]. Additionally, MMPs activate various growth factors in the ECM, contributing to tumor proliferation and angiogenesis [46]. Importantly, TGF-β-induced EMT through both SMAD and non-SMAD pathways is a canonical process in metastasis, as it produces transcription modulators such as SNAIL, ZEB, and TWIST and triggers other EMT-related signaling pathways [47]. TGF-β/SMAD signaling also enhances the PI3K/AKT pathway, promoting cancer invasion and migration [48]. Furthermore, the MAPK cascade is activated, contributing to tumorigenesis as dephosphorylation of SMAD at the MAPK site reduces its nuclear stability and activity, counteracting the tumor-suppressive effects of TGF-β [48]. The interaction between OC and its EMT induced by TGF-β not only enhances cellular invasiveness but also suppresses immune responses, and thus understanding the pivotal mechanisms underlying this process is crucial for identifying therapeutic targets aimed at the metastatic downstream pathways in OC.
3.2. NF-κB Signaling Pathway
The NF-κB pathways is a central regulator of inflammation, cell survival, and proliferation, playing a critical role in driving cancer metastasis, including OC dissemination and peritoneal metastasis [49]. NF-κB contributes to tumorigenesis in various way. When damaged DNA strands are detected, NF-κB promotes tumorigenesis by activating mutagenic enzymes, such as cytidine deaminase, which induces mutations by converting cytosine to thymine during cell cycle progression [49]. These mutations provide cancer cells with the genetic plasticity required for adaptation and survival [50]. Furthermore, NF-κB promotes EMT by regulating EMT-associated transcription factors, such as SNAIL and TWIST, thereby enhancing the migratory capacity and stemness of cancer cells [51]. In addition, NF-κB drives the secretion of cytokines such as vascular endothelial growth factor (VEGF) and TGF-β. VEGF induces angiogenesis and vascular remodeling, enabling cancer cells to disseminate into the peritoneum. Concurrently, TGF-β upregulates metastatic genes and MMPs, amplifying ECM remodeling and enhancing hypoxic conditions that support tumor progression [49,52]. This dual function establishes a pro-tumorigenic environment that facilitates OC metastasis. Within the TME, NF-κB signaling is activated by pro-inflammatory cytokines such as TNF-α and IL-1β, which are prevalent in the peritoneal microenvironment. This persistent activation triggers NF-κB-driven inflammatory responses and cytokine production, maintaining a TME conducive to immune evasion, angiogenesis, and ECM remodeling, collectively driving ovarian cancer metastasis [53]. Additionally, NF-κB signaling in the TME fosters immune evasion by polarizing macrophages into TAMs and inducing CAFs, both of which facilitate peritoneal dissemination [54,55]. Thus, NF-κB signaling interacts with EMT-associated transcription factors, further serving the metastatic potential of OC.
3.3. PI3K/AKT/mTOR Signaling Pathway
The PI3K/AKT/mTOR pathway is frequently dysregulated in OC, playing a significant role in cell survival, proliferation, resistance to apoptosis, and chemotherapeutic resistance [56]. Malfunction in this pathway supports tumor growth and adaptability, particularly under the metabolic and immune pressures of the OC TME [57]. Key aberrants alteration in OC include PIK3CA amplification as well as PTEN loss through deletion or epigenetic silencing [57]. Studies have demonstrated that components of this pathway, such as phosphorylated AKT and mTOR-associated proteins, are highly expressed in aggressive tumors and are associated with poor prognosis [20]. The PI3K/AKT/mTOR signaling also influences immune interactions and promotes tumor progression within the TME [58]. In high-grade OC, CAFs release hepatocyte growth factor (HGF), activating the PI3K/AKT axis to enhance OC proliferation, including integrin-mediated interactions [59]. It motivates tumor cell adhesion to the mesothelial lining of the peritoneum, a crucial step in initiating peritoneal metastasis [60]. Subsequently, genetic alterations such as mutations or upregulated in PIK3CA, AKT1, and mTOR are significantly observed in OC, underscoring their role in promoting oncogenic signaling [20,61]. Moreover, it facilitates metabolic reprogramming, enabling cancer cells to effectively utilize lipids from omental adipocytes as a primary energy source, a pivotal process for their survival under nutrient-deprived conditions [62,63]. Furthermore, the PI3K/AKT/mTOR pathway regulates angiogenesis by enhancing VEGF expression, which ensures nutrient supply while supporting the formation of blood vessels and oxygen delivery to establish secondary metastatic sites [64]. The activation of AKT directly stabilizes SNAIL, a master regulator of EMT, and promotes the suppression of E-cadherin, thereby reinforcing mesenchymal traits [40]. The role of PI3K/AKT/mTOR in modulating immunoreaction, including the suppression of cytotoxic T cell responses and recruitment of immunosuppressive TAMs, further contributes to OC progression and metastasis [58]. Therefore, the PI3K/AKT/mTOR pathway is involved in EMT-related pathways in cancer cells, controlling cytoskeletal reorganization, enhanced cellular motility, and tumor aggressiveness which are hallmarks of metastatic competence [20,65].
Understanding the metastatic signaling pathways in OC provides a detailed perspective into the molecular mechanisms regarding peritoneal metastasis. Overall, TGF-β, NF-κB, and PI3K/AKT/mTOR pathway closey cooperate within TME as a complex network, influencing EMT, cell proliferation, and immune evasion (Figure 2). Therefore, targeting these pathways has the potential to disrupt the metastatic cascade and improve clinical outcomes for OC patients.
4. Therapeutic Applications for Ovarian Cancer Metastasis
Despite advancements in immunotherapy, the clinical management of metastatic OC remains challenging mainly due to the immunosuppressive TME of OC [66]. Currently, solely use of antibody targeting cytotoxic T lymphocyte-associated protein 4 (CTLA-4) or programmed cell death (PD-1)/programmed cell death ligand (PD-L1) has shown limited efficacy in OC along with poor outcomes [67]. Thus, it is urged to develop advanced treatment strategies, such as combination of traditional immunotherapy with other anti-cancer treatments.
4.1. Recent Advanced Combination Immunotherapy in Ovarian Cancer Metastasis
Recent studies on NK cell therapy using oncolytic viruses demonstrated a promising anti-cancer immune reaction [68]. While TGF-β in ascites exhibits immunosuppressive effect in OC, the proper function of NK cell is compromised by downregulation of NK cell activating receptors [69]. Therefore, recent research has focused not only on the direct killing of cancer cells but also boosting immune cell activity. Oncolytic viruses infect cancer cells and stimulate "danger signals," which help construct an immunogenic TME and regulate inflammatory and immunomodulatory cytokines with transduction molecules [70]. According to studies with experimental cancer models, vesicular stomatitis virus (VSV) and reovirus, both recombinant oncolytic viruses, have successfully induced NK cell activation, resulting in an increased survival rate in peritoneal OC models compared to single-agent therapies [68]. Notably, among the two viruses, infection of VSV enhanced antigen presentation capabilities and activation of IFN-γ generating NK, CD8+, and CD4+ T cells which accelerated anti-cancer immune response [68]. As a result, combination therapy of oncolytic virus and NK cell immunotherapy presents potential for targeting metastatic cancer more effectively than conventional monotherapy.
Several clinical trials are underway to address the challenges of OC metastasis. IMagyn050 (NCT03038100) evaluated the efficacy of atezolizumab, a PD-L1 inhibitor, in combination with chemotherapy and bevacizumab in stage 3/4 OC patients [71,72]. Unfortunately, it has no statistically significant changes of progression-free survival in PD-L1 positive patients [71]. While this clinical trial has not demonstrated remarkable outcomes, it is still ongoing in phase 3, integrating immune checkpoint inhibitors, and no severe side effects have been reported so far. LY2157299 (galunisertib) is a small molecule TGF-β inhibitor which targets type I receptor (TβRI) with decreased toxicity and adverse effect [73]. A phase Ib trial (NCT03206177) is currently investigating the combination therapy of galunisertib with paclitaxel and carboplatin, well-known chemotherapeutic agents, against recurrent or newly diagnosed OC [74].
Bintrafusp alfa (BA) is a cutting-edge dual blockade of TGF-β and PD-L1, demonstrating the anti-tumor efficacy on metastatic OC by promoting tumor-infiltrating CD8+ T cells and provoking the immune response within the TME [75]. As reported in the study using the mouse models, BA treatments markedly hampered the ascites development, generated inflammatory cytokines, and thus prolong long-term survival [76]. This effect is attributed to the increased expression of CD4 and CD8 T effector memory cells and NK cells in the peritoneum [76]. In addition to BA, other dual blockades targeting signaling pathways and immunogenic factors are being explored for metastatic OC. This approach shows promise, given the multifaceted roles of metastatic signaling pathways discussed above.
4.2. Signaling Pathway Targeted Therapy in Ovarian Cancer Metastasis
Given the limitations of current immunotherapy in effectively managing metastatic OC and its recurrence, there is a growing interest in targeting specific pathways that play a pivotal role in tumor progression and the immunosuppressive TME. By integrating signaling pathway inhibitors with present immunotherapeutic approaches, researchers aim to overcome resistance mechanisms and enhance anti-cancer efficacy.
A-83-01 is an inhibitor of the TGF-β which effectively repress transcriptional alterations occurred by TGF-β1 [77]. A-83-01 treatment enable to reverse EMT-gene expression pattern and inhibit MMP2 and phosphorylated SMAD2, along with restraining platelet-induced invasion [78]. The in vitro study revealed that A-83-01 is more effective at reducing cell motility, invasion, and adhesion than cell proliferation, suggesting its potential as a combination therapy with immunotherapies that target cell proliferation. Additionally, A-83-01 has demonstrated anti-tumor efficacy and improved survival rates in vivo [77]. Cordycepin (3’-deoxyadenosine) is a well-established polyadenylation inhibitor with diverse capabilities, including anti-proliferative, anti-inflammatory, and anti-cancer activities as well as immune-activation properties [79]. The study elucidated that cordycepin is related to CCL5-regulated AKT/NF-κB signaling transduction in OC. AKT inactivation by downregulated CCL5 via cordycepin induced apoptosis of OC cells and suppressed expression of nuclear NF-κB [79]. In addition, cordycepin influenced OC cell growth and autophagy in vitro [80]. CMG002, a P13K/mTOR bifunctional inhibitor, can be synergized with paclitaxel or cisplatin and has shown prominent tumor reduction, particularly in chemoresistant OC [81]. It restrained the cancer cell proliferation and provoked apoptosis and G1 cell cycle arrest, suggesting its potential for managing chemoresistant OC [82]. Notably, CMG002 exhibits stronger anti-cancer activity at lower concentrations compared to other PI3K/mTOR inhibitors and re-sensitizes chemoresistant cells to paclitaxel or cisplatin [81,82]. Additionally, the PI3K inhibitor BKM120, when combined with the poly (ADP-ribose) polymerase (PARP) inhibitor Olaparib, exhibited therapeutic effects by blocking cell proliferation, cell growth, migration, and invasion of PIK3CA mutant OC cells. This combination also regulated intraperitoneal dissemination in vivo [83]. Lastly, the mTOR kinase inhibitor CC223 caused the degradation of mTORC1 and mTORC2 complexes, effectively preventing tumor cell proliferation [84]. It also increased reactive oxygen species (ROS) production, thereby inhibiting tumor growth within the intraperitoneal cavity [82,84].
Comprehensively, signaling pathway-targeted therapies, when combined with immunotherapy, present a promising approach to addressing the challenges of metastatic OC (Table 1). By modulating metastatic pathways such as TGF-β, NF-κB, and PI3K/AKT/mTOR signaling, which are involved in tumor progression and metastasis, these combination strategies have the potential to improve clinical outcomes and provide durable responses for OC patients with fewer adverse effects.
5. Conclusions and Perspective
OC remains a significant clinical challenge due to its high metastatic potential, complex signaling pathways, and the immunosuppressive TME. Deciphering the mechanisms related to OC metastasis, particularly though TGF-β, NF-κB, and PI3K/AKT/mTOR signaling transduction, is crucial for developing effective therapeutic strategies. These pathways contribute to EMT, immune evasion, and tumor progression, facilitating the dissemination of OC cells to distant regions, especially the peritoneum. Furthermore, despite versatile immunotherapeutic approaches, the OC TME diminishes the effectiveness of conventional therapies. Within malignant ascites, cancer cells evade immune process by correlation between immune cells and tumor, leading to EMT and chemoresistance [85]. The compromised function of cytotoxic effector T cells weakens both innate and adaptive immunity, reinforcing the immunosuppressive characteristics of OC [86]. As a result, metastasis is exacerbated, and traditional therapies are hindered.
Recent advances in combination therapies targeting both signaling pathways and the immune system, such as oncolytic virus therapy, NK cell therapy, and immune checkpoint inhibitors, hold promise for overcoming these challenges Additionally, as the aforementioned signaling pathways serve as key modulators in metastasis, combination therapies are anticipated to exhibit synergistic effects against metastatic OC. These approaches are expected to improve clinical outcomes by disrupting the metastatic cascade, reducing recurrence, and providing more durable responses with fewer side effects. Further research is required to optimize these therapies to maximize their long-term effects on patient outcomes. Also, signaling pathways are not only involved in specific tumorigenic processes but are also intricately interconnected, exerting complex effects. Thus, identifying molecules to inhibit these pathways requires delicate consideration, as targeting one pathway may inadvertently affect others, potentially causing undesirable consequences. In summary, addressing the complex networks of metastasis, immune evasion, and tumor progression offers a promising strategy to improve the prognosis and survival of OC patients. Therefore, this review highlights the mechanisms of OC metastasis and the major pathways involved, demonstrating how targeting these pathways presents a promising approach to combating OC metastasis.
Author Contributions
SC mainly designed and wrote the manuscript; SC and MJ wrote the manuscript; HY supervised, organized, and wrote the manuscript; all authors revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Catholic University of Korea, Research Fund, 2022 (# M2022B00080009) and the Ministry of Food and Drug Safety in Korea (22213MFDS421).
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| TME | Tumor microenvironment |
| ECM | Extracellular matrix |
| TGF-β | Transforming growth factor-β |
| EMT | Epithelial-to-mesenchymal transition |
| SMAD | Suppressor of mothers against decapentaplegic |
| PI3K | Phosphoinositide 3-kinase |
| mTOR | Mammalian target of rapamycin |
| MAPK | Mitogen-activated protein kinase |
| ERK | Extracellular signal-regulated kinase |
| NF-κB | Nuclear factor of kappa-light chain of enhancer-activated B cells |
| TAMs | Tumor-associated macrophages |
| TNF-α | Tumor necrosis factor-alpha |
| PMCs | Peritoneal mesothelial cells |
| CAFs | Cancer-associated fibroblasts |
| Tregs | Regulatory T cells |
| MDSCs | Myeloid-derived suppressor cells |
| NK | Natural killer |
| CTLs | Cytotoxic T lymphocytes |
| MMPs | Matrix metalloproteinases |
| VEGF | Vascular endothelial growth factor |
| HGF | Hepatocyte growth factor |
| CTLA-4 | Cytotoxic T lymphocyte-associated protein 4 |
| PD-1 | Programmed cell death |
| PD-L1 | Programmed cell death ligand |
| VSV | Vesicular stomatitis virus |
| TβRI | TGF-β receptor type I |
| BA | Bintrafusp alfa |
| PARP | Poly (ADP-ribose) polymerase |
| ROS | Reactive oxygen species |
| CTC | Circulating tumor cell |
| MSCs | Mesenchymal stem cells |
| RTK | Receptor tyrosine kinase |
| PIP2 | Phosphatidylinositol 4,5-bisphosphate |
| PIP3 | Phosphatidylinositol (3,4,5)-triphosphate |
| PTEN | Phosphatase and tensin homolog |
| INPP4B | Inositol polyphosphate 4-phosphatase B |
| PDK1 | Phosphoinositide-dependent protein kinase 1 |
References
- Momenimovahed, Z.; Tiznobaik, A.; Taheri, S.; Salehiniya, H. Ovarian cancer in the world: epidemiology and risk factors. Int J Womens Health 2019, 11, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Mei, S.; Chen, X.; Wang, K.; Chen, Y. Tumor microenvironment in ovarian cancer peritoneal metastasis. Cancer cell international 2023, 23((1)), 11. [Google Scholar] [CrossRef] [PubMed]
- Uno, K.; Iyoshi, S.; Yoshihara, M.; Kitami, K.; Mogi, K.; Fujimoto, H.; Sugiyama, M.; Koya, Y.; Yamakita, Y.; Nawa, A. Metastatic voyage of ovarian cancer cells in ascites with the assistance of various cellular components. International Journal of Molecular Sciences 2022, 23((8)), 4383. [Google Scholar] [CrossRef] [PubMed]
- Yonemura, A.; Semba, T.; Zhang, J.; Fan, Y.; Yasuda-Yoshihara, N.; Wang, H.; Uchihara, T.; Yasuda, T.; Nishimura, A.; Fu, L., Mesothelial cells with mesenchymal features enhance peritoneal dissemination by forming a protumorigenic microenvironment. Cell Reports 2024, 43, (1).
- Laurent-Issartel, C.; Landras, A.; Agniel, R.; Giffard, F.; Blanc-Fournier, C.; Cruz, E. D. S.; Habes, C.; Leroy-Dudal, J.; Carreiras, F.; Kellouche, S. Ascites microenvironment conditions the peritoneal pre-metastatic niche to promote the implantation of ovarian tumor spheroids: Involvement of fibrinogen/fibrin and αV and α5β1 integrins. Experimental Cell Research 2024, 441((1)), 114155. [Google Scholar] [CrossRef]
- Gutic, B.; Bozanovic, T.; Mandic, A.; Dugalic, S.; Todorovic, J.; Dugalic, M. G.; Sengul, D.; Detanac, D. A.; Sengul, I.; Detanac, D. Preliminary outcomes of five-year survival for ovarian malignancies in profiled Serbian Oncology Centre. Clinics 2023, 78, 100204. [Google Scholar] [CrossRef]
- Garzon, S.; Laganà, A. S.; Casarin, J.; Raffaelli, R.; Cromi, A.; Franchi, M.; Barra, F.; Alkatout, I.; Ferrero, S.; Ghezzi, F. Secondary and tertiary ovarian cancer recurrence: what is the best management? Gland Surg 2020, 9((4)), 1118–1129. [Google Scholar] [CrossRef]
- Song, M.; Cui, M.; Liu, K. Therapeutic strategies to overcome cisplatin resistance in ovarian cancer. Eur J Med Chem 2022, 232, 114205. [Google Scholar] [CrossRef]
- Elies, A.; Rivière, S.; Pouget, N.; Becette, V.; Dubot, C.; Donnadieu, A.; Rouzier, R.; Bonneau, C. The role of neoadjuvant chemotherapy in ovarian cancer. Expert Rev Anticancer Ther 2018, 18((6)), 555–566. [Google Scholar] [CrossRef]
- Chen, C.; Ge, X.; Zhao, Y.; Wang, D.; Ling, L.; Zheng, S.; Ding, K.; Wang, J.; Sun, L. Molecular Alterations in Metastatic Ovarian Cancer From Gastrointestinal Cancer. Front Oncol 2020, 10, 605349. [Google Scholar] [CrossRef]
- Cui, M.; Liu, Y.; Cheng, L.; Li, T.; Deng, Y.; Liu, D. Research progress on anti-ovarian cancer mechanism of miRNA regulating tumor microenvironment. Frontiers in Immunology 2022, 13, 1050917. [Google Scholar] [CrossRef]
- Dai, W.; Zhou, J.; Chen, T. Unraveling the extracellular vesicle network: insights into ovarian cancer metastasis and chemoresistance. Molecular Cancer 2024, 23((1)), 201. [Google Scholar] [CrossRef] [PubMed]
- Baba, A. B.; Rah, B.; Bhat, G. R.; Mushtaq, I.; Parveen, S.; Hassan, R.; Hameed Zargar, M.; Afroze, D. Transforming growth factor-beta (TGF-β) signaling in cancer-A betrayal within. Frontiers in pharmacology 2022, 13, 791272. [Google Scholar] [CrossRef] [PubMed]
- Jinesh, G. G.; Brohl, A. S. Classical epithelial-mesenchymal transition (EMT) and alternative cell death process-driven blebbishield metastatic-witch (BMW) pathways to cancer metastasis. Signal transduction and targeted therapy 2022, 7((1)), 296. [Google Scholar] [CrossRef] [PubMed]
- Sicard, A. A.; Dao, T.; Suarez, N. G.; Annabi, B. Diet-Derived Gallated Catechins Prevent TGF-β-Mediated Epithelial-Mesenchymal Transition, Cell Migration and Vasculogenic Mimicry in Chemosensitive ES-2 Ovarian Cancer Cells. Nutr Cancer 2021, 73((1)), 169–180. [Google Scholar] [CrossRef]
- Zhou, J.; Jiang, W.; Huang, W.; Ye, M.; Zhu, X. Prognostic Values of Transforming Growth Factor-Beta Subtypes in Ovarian Cancer. BioMed Research International 2020, 2020((1)), 2170606. [Google Scholar] [CrossRef]
- Deng, Z.; Fan, T.; Xiao, C.; Tian, H.; Zheng, Y.; Li, C.; He, J. TGF-β signaling in health, disease, and therapeutics. Signal transduction and targeted therapy 2024, 9((1)), 61. [Google Scholar]
- Ali, S.; Rehman, M. U.; Yatoo, A. M.; Arafah, A.; Khan, A.; Rashid, S.; Majid, S.; Ali, A.; Ali, M. N. TGF-β signaling pathway: Therapeutic targeting and potential for anti-cancer immunity. European Journal of Pharmacology 2023, 947, 175678. [Google Scholar] [CrossRef]
- Panwar, V.; Singh, A.; Bhatt, M.; Tonk, R. K.; Azizov, S.; Raza, A. S.; Sengupta, S.; Kumar, D.; Garg, M. Multifaceted role of mTOR (mammalian target of rapamycin) signaling pathway in human health and disease. Signal transduction and targeted therapy 2023, 8((1)), 375. [Google Scholar] [CrossRef]
- Glaviano, A.; Foo, A. S.; Lam, H. Y.; Yap, K. C.; Jacot, W.; Jones, R. H.; Eng, H.; Nair, M. G.; Makvandi, P.; Geoerger, B. PI3K/AKT/mTOR signaling transduction pathway and targeted therapies in cancer. Molecular cancer 2023, 22((1)), 138. [Google Scholar] [CrossRef]
- Baghban, R.; Roshangar, L.; Jahanban-Esfahlan, R.; Seidi, K.; Ebrahimi-Kalan, A.; Jaymand, M.; Kolahian, S.; Javaheri, T.; Zare, P. Tumor microenvironment complexity and therapeutic implications at a glance. Cell Communication and Signaling 2020, 18, 1–19. [Google Scholar] [CrossRef]
- Harrington, B. S.; Annunziata, C. M. NF-κB signaling in ovarian cancer. Cancers 2019, 11((8)), 1182. [Google Scholar] [CrossRef]
- Kaltschmidt, B.; Witte, K. E.; Greiner, J. F.; Weissinger, F.; Kaltschmidt, C. Targeting NF-κB signaling in cancer stem cells: a narrative review. Biomedicines 2022, 10((2)), 261. [Google Scholar] [CrossRef] [PubMed]
- Yin, N.; Li, X.; Zhang, X.; Xue, S.; Cao, Y.; Niedermann, G.; Lu, Y.; Xue, J. Development of pharmacological immunoregulatory anti-cancer therapeutics: current mechanistic studies and clinical opportunities. Signal Transduction and Targeted Therapy 2024, 9((1)), 126. [Google Scholar] [CrossRef]
- Siminiak, N.; Czepczyński, R.; Zaborowski, M. P.; Iżycki, D. Immunotherapy in ovarian cancer. Archivum Immunologiae et Therapiae Experimentalis 2022, 70((1)), 19. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Wang, X.; Yao, W.; Shi, D.; Shao, X.; Lu, Z.; Chai, Y.; Song, J.; Tang, W.; Wang, X. Mechanism insights and therapeutic intervention of tumor metastasis: latest developments and perspectives. Signal transduction and targeted therapy 2024, 9((1)), 192. [Google Scholar] [CrossRef]
- Dunbar, K. J.; Efe, G.; Cunningham, K.; Esquea, E.; Navaridas, R.; Rustgi, A. K. Regulation of metastatic organotropism. Trends in Cancer 2024. [Google Scholar] [CrossRef]
- Gao, Y.; Bado, I.; Wang, H.; Zhang, W.; Rosen, J. M.; Zhang, X. H.-F. Metastasis organotropism: redefining the congenial soil. Developmental cell 2019, 49((3)), 375–391. [Google Scholar] [CrossRef]
- Ford, C. E.; Werner, B.; Hacker, N. F.; Warton, K. The untapped potential of ascites in ovarian cancer research and treatment. British journal of cancer 2020, 123((1)), 9–16. [Google Scholar] [CrossRef]
- Pal, S.; Bhowmick, S.; Sharma, A.; Sierra-Fonseca, J. A.; Mondal, S.; Afolabi, F.; Roy, D., Lymphatic vasculature in ovarian cancer. Biochimica et Biophysica Acta (BBA)-Reviews on Cancer 2023, 188950.
- Szczerba, A.; Śliwa, A.; Pieta, P. P.; Jankowska, A. The role of circulating Tumor cells in Ovarian Cancer Dissemination. Cancers 2022, 14((24)), 6030. [Google Scholar] [CrossRef]
- Yousefi, M.; Dehghani, S.; Nosrati, R.; Ghanei, M.; Salmaninejad, A.; Rajaie, S.; Hasanzadeh, M.; Pasdar, A. Current insights into the metastasis of epithelial ovarian cancer-hopes and hurdles. Cellular Oncology 2020, 43, 515–538. [Google Scholar] [CrossRef]
- Pascual-Antón, L.; Cardeñes, B.; Sainz de la Cuesta, R.; González-Cortijo, L.; López-Cabrera, M.; Cabañas, C.; Sandoval, P. Mesothelial-to-mesenchymal transition and exosomes in peritoneal metastasis of ovarian cancer. International journal of molecular sciences 2021, 22((21)), 11496. [Google Scholar] [CrossRef] [PubMed]
- Ramos, C.; Gerakopoulos, V.; Oehler, R. Metastasis-associated fibroblasts in peritoneal surface malignancies. British Journal of Cancer 2024, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Shi, H.; Zhang, B.; Ou, X.; Ma, Q.; Chen, Y.; Shu, P.; Li, D.; Wang, Y. Myeloid-derived suppressor cells as immunosuppressive regulators and therapeutic targets in cancer. Signal transduction and targeted therapy 2021, 6((1)), 362. [Google Scholar] [CrossRef] [PubMed]
- Rakina, M.; Kazakova, A.; Villert, A.; Kolomiets, L.; Larionova, I. Spheroid formation and peritoneal metastasis in ovarian cancer: the role of stromal and immune components. International journal of molecular sciences 2022, 23((11)), 6215. [Google Scholar] [CrossRef]
- Xu, T.; Yu, S.; Zhang, J.; Wu, S., Dysregulated tumor-associated macrophages in carcinogenesis, progression and targeted therapy of gynecological and breast cancers. Journal of hematology & oncology 2021, 14, 1-20.
- Li, C.; Jiang, P.; Wei, S.; Xu, X.; Wang, J. Regulatory T cells in tumor microenvironment: new mechanisms, potential therapeutic strategies and future prospects. Molecular cancer 2020, 19, 1–23. [Google Scholar] [CrossRef]
- Wang, X.; Xue, X.; Pang, M.; Yu, L.; Qian, J.; Li, X.; Tian, M.; Lyu, A.; Lu, C.; Liu, Y. Epithelial–mesenchymal plasticity in cancer: signaling pathways and therapeutic targets. MedComm 2024, 5((8)), e659. [Google Scholar] [CrossRef]
- Huang, Y.; Hong, W.; Wei, X., The molecular mechanisms and therapeutic strategies of EMT in tumor progression and metastasis. Journal of hematology & oncology 2022, 15, (1), 129.
- Chou, M.-Y.; Yang, M.-H. Interplay of immunometabolism and epithelial–mesenchymal transition in the tumor microenvironment. International Journal of Molecular Sciences 2021, 22((18)), 9878. [Google Scholar] [CrossRef]
- Wang, X.; Eichhorn, P. J. A.; Thiery, J. P. In TGF-β, EMT, and resistance to anti-cancer treatment, Seminars in Cancer Biology, 2023; Elsevier: 2023; pp 1-11.
- Sun, L.; Xing, J.; Zhou, X.; Song, X.; Gao, S., Wnt/β-catenin signalling, epithelial-mesenchymal transition and crosslink signalling in colorectal cancer cells. Biomedicine & Pharmacotherapy 2024, 175, 116685.
- Yang, Y.; Ye, W.-L.; Zhang, R.-N.; He, X.-S.; Wang, J.-R.; Liu, Y.-X.; Wang, Y.; Yang, X.-M.; Zhang, Y.-J.; Gan, W.-J., The role of TGF-β signaling pathways in cancer and its potential as a therapeutic target. Evidence-Based Complementary and Alternative Medicine 2021, 2021, (1), 6675208.
- Yuan, Z.; Li, Y.; Zhang, S.; Wang, X.; Dou, H.; Yu, X.; Zhang, Z.; Yang, S.; Xiao, M. Extracellular matrix remodeling in tumor progression and immune escape: from mechanisms to treatments. Molecular cancer 2023, 22((1)), 48. [Google Scholar] [CrossRef]
- Peltanova, B.; Raudenska, M.; Masarik, M. Effect of tumor microenvironment on pathogenesis of the head and neck squamous cell carcinoma: a systematic review. Molecular cancer 2019, 18((1)), 63. [Google Scholar] [CrossRef]
- Hao, Y.; Baker, D.; Ten Dijke, P. TGF-β-mediated epithelial-mesenchymal transition and cancer metastasis. International journal of molecular sciences 2019, 20((11)), 2767. [Google Scholar] [CrossRef]
- Xue, V. W.; Chung, J. Y.-F.; Córdoba, C. A. G.; Cheung, A. H.-K.; Kang, W.; Lam, E. W.-F.; Leung, K.-T.; To, K.-F.; Lan, H.-Y.; Tang, P. M.-K. Transforming growth factor-β: a multifunctional regulator of cancer immunity. Cancers 2020, 12((11)), 3099. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Ma, C.; Zhang, Z.; Zhang, H.; Hu, H. NF-κB signaling in inflammation and cancer. MedComm 2021, 2((4)), 618–653. [Google Scholar] [CrossRef] [PubMed]
- Bhat, G. R.; Sethi, I.; Sadida, H. Q.; Rah, B.; Mir, R.; Algehainy, N.; Albalawi, I. A.; Masoodi, T.; Subbaraj, G. K.; Jamal, F. Cancer cell plasticity: From cellular, molecular, and genetic mechanisms to tumor heterogeneity and drug resistance. Cancer and Metastasis Reviews 2024, 43((1)), 197–228. [Google Scholar] [CrossRef]
- Oh, A.; Pardo, M.; Rodriguez, A.; Yu, C.; Nguyen, L.; Liang, O.; Chorzalska, A.; Dubielecka, P. M. NF-κB signaling in neoplastic transition from epithelial to mesenchymal phenotype. Cell Communication and Signaling 2023, 21((1)), 291. [Google Scholar] [CrossRef]
- Quintero-Fabián, S.; Arreola, R.; Becerril-Villanueva, E.; Torres-Romero, J. C.; Arana-Argáez, V.; Lara-Riegos, J.; Ramírez-Camacho, M. A.; Alvarez-Sánchez, M. E. Role of matrix metalloproteinases in angiogenesis and cancer. Frontiers in oncology 2019, 9, 1370. [Google Scholar] [CrossRef]
- Chen, S.; Liu, Y.; Zhong, Z.; Wei, C.; Liu, Y.; Zhu, X. Peritoneal immune microenvironment of endometriosis: Role and therapeutic perspectives. Frontiers in Immunology 2023, 14, 1134663. [Google Scholar] [CrossRef]
- Mao, X.; Xu, J.; Wang, W.; Liang, C.; Hua, J.; Liu, J.; Zhang, B.; Meng, Q.; Yu, X.; Shi, S. Crosstalk between cancer-associated fibroblasts and immune cells in the tumor microenvironment: new findings and future perspectives. Molecular cancer 2021, 20, 1–30. [Google Scholar] [CrossRef]
- Han, J.; Dong, L.; Wu, M.; Ma, F. Dynamic polarization of tumor-associated macrophages and their interaction with intratumoral T cells in an inflamed tumor microenvironment: from mechanistic insights to therapeutic opportunities. Frontiers in Immunology 2023, 14, 1160340. [Google Scholar] [CrossRef]
- Rinne, N.; Christie, E. L.; Ardasheva, A.; Kwok, C. H.; Demchenko, N.; Low, C.; Tralau-Stewart, C.; Fotopoulou, C.; Cunnea, P. Targeting the PI3K/AKT/mTOR pathway in epithelial ovarian cancer, therapeutic treatment options for platinum-resistant ovarian cancer. Cancer Drug Resistance 2021, 4((3)), 573. [Google Scholar] [CrossRef]
- Ediriweera, M. K.; Tennekoon, K. H.; Samarakoon, S. R. In Role of the PI3K/AKT/mTOR signaling pathway in ovarian cancer: Biological and therapeutic significance, Seminars in cancer biology, 2019; Elsevier: 2019; pp 147-160.
- Mafi, S.; Mansoori, B.; Taeb, S.; Sadeghi, H.; Abbasi, R.; Cho, W. C.; Rostamzadeh, D. mTOR-mediated regulation of immune responses in cancer and tumor microenvironment. Frontiers in immunology 2022, 12, 774103. [Google Scholar] [CrossRef]
- Ritch, S. J.; Telleria, C. M. The transcoelomic ecosystem and epithelial ovarian cancer dissemination. Frontiers in Endocrinology 2022, 13, 886533. [Google Scholar] [CrossRef] [PubMed]
- van Baal, J. O.; van Noorden, C. J.; Nieuwland, R.; Van de Vijver, K. K.; Sturk, A.; van Driel, W. J.; Kenter, G. G.; Lok, C. A., Development of peritoneal carcinomatosis in epithelial ovarian cancer: a review. Journal of Histochemistry & Cytochemistry 2018, 66, (2), 67-83.
- Maioru, O.-V.; Radoi, V.-E.; Coman, M.-C.; Hotinceanu, I.-A.; Dan, A.; Eftenoiu, A.-E.; Burtavel, L.-M.; Bohiltea, L.-C.; Severin, E.-M. Developments in genetics: better management of ovarian cancer patients. International Journal of Molecular Sciences 2023, 24((21)), 15987. [Google Scholar] [CrossRef] [PubMed]
- Schiliro, C.; Firestein, B. L. Mechanisms of metabolic reprogramming in cancer cells supporting enhanced growth and proliferation. Cells 2021, 10((5)), 1056. [Google Scholar] [CrossRef]
- Fu, Y.; Zou, T.; Shen, X.; Nelson, P. J.; Li, J.; Wu, C.; Yang, J.; Zheng, Y.; Bruns, C.; Zhao, Y. Lipid metabolism in cancer progression and therapeutic strategies. MedComm 2021, 2((1)), 27–59. [Google Scholar] [CrossRef]
- Tufail, M.; Jiang, C.-H.; Li, N. Altered metabolism in cancer: Insights into energy pathways and therapeutic targets. Molecular Cancer 2024, 23((1)), 203. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhanghuang, C.; Mi, T.; Jin, L.; Liu, J.; Li, M.; Wu, X.; Wang, J.; Li, M.; Wang, Z., The PI3K-AKT-mTOR signaling pathway mediates the cytoskeletal remodeling and epithelial-mesenchymal transition in bladder outlet obstruction. Heliyon 2023, 9, (11).
- Yang, C.; Xia, B.-R.; Zhang, Z.-C.; Zhang, Y.-J.; Lou, G.; Jin, W.-L. Immunotherapy for ovarian cancer: adjuvant, combination, and neoadjuvant. Frontiers in immunology 2020, 11, 577869. [Google Scholar] [CrossRef]
- Ghisoni, E.; Imbimbo, M.; Zimmermann, S.; Valabrega, G. Ovarian cancer immunotherapy: turning up the heat. International journal of molecular sciences 2019, 20((12)), 2927. [Google Scholar] [CrossRef]
- Gebremeskel, S.; Nelson, A.; Walker, B.; Oliphant, T.; Lobert, L.; Mahoney, D.; Johnston, B., Natural killer T cell immunotherapy combined with oncolytic vesicular stomatitis virus or reovirus treatments differentially increases survival in mouse models of ovarian and breast cancer metastasis. Journal for immunotherapy of cancer 2021, 9, (3).
- Hoogstad-van Evert, J. S.; Bekkers, R.; Ottevanger, N.; Jansen, J. H.; Massuger, L.; Dolstra, H. Harnessing natural killer cells for the treatment of ovarian cancer. Gynecologic oncology 2020, 157((3)), 810–816. [Google Scholar] [CrossRef]
- Apolonio, J. S.; de Souza Gonçalves, V. L.; Santos, M. L. C.; Luz, M. S.; Souza, J. V. S.; Pinheiro, S. L. R.; de Souza, W. R.; Loureiro, M. S.; de Melo, F. F. Oncolytic virus therapy in cancer: A current review. World journal of virology 2021, 10((5)), 229. [Google Scholar] [CrossRef]
- Pignata, S.; Bookman, M.; Sehouli, J.; Miller, A.; Penson, R. T.; Taskiran, C.; Anderson, C.; Hietanen, S.; Myers, T.; Madry, R. Overall survival and patient-reported outcome results from the placebo-controlled randomized phase III IMagyn050/GOG 3015/ENGOT-OV39 trial of atezolizumab for newly diagnosed stage III/IV ovarian cancer. Gynecologic oncology 2023, 177, 20–31. [Google Scholar] [CrossRef]
- Moore, K.; Bookman, M.; Sehouli, J.; Miller, A.; Anderson, C.; Scambia, G.; Myers, T.; Taskiran, C.; Robison, K.; Maenpaa, J. LBA31 Primary results from IMagyn050/GOG 3015/ENGOT-OV39, a double-blind placebo (pbo)-controlled randomised phase III trial of bevacizumab (bev)-containing therapy+/-atezolizumab (atezo) for newly diagnosed stage III/IV ovarian cancer (OC). Annals of Oncology 2020, 31, S1161–S1162. [Google Scholar] [CrossRef]
- Huang, C.-Y.; Chung, C.-L.; Hu, T.-H.; Chen, J.-J.; Liu, P.-F.; Chen, C.-L., Recent progress in TGF-β inhibitors for cancer therapy. Biomedicine & Pharmacotherapy 2021, 134, 111046.
- Makker, V.; Green, A. K.; Wenham, R. M.; Mutch, D.; Davidson, B.; Miller, D. S. New therapies for advanced, recurrent, and metastatic endometrial cancers. Gynecologic oncology research and practice 2017, 4, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kment, J. L.; Newsted, D.; Young, S.; Vermeulen, M.; Craig, A. W., 544 Coordinated blockade of TGF-β and PD-L1 by bintrafusp alfa promotes survival in preclinical ovarian cancer models by promoting T effector memory responses. In BMJ Specialist Journals: 2023.
- Kment, J.; Newsted, D.; Young, S.; Vermeulen, M. C.; Laight, B. J.; Greer, P. A.; Lan, Y.; Craig, A. W. Blockade of TGF-β and PD-L1 by bintrafusp alfa promotes survival in preclinical ovarian cancer models by promoting T effector and NK cell responses. British Journal of Cancer 2024, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Yamamura, S.; Matsumura, N.; Mandai, M.; Huang, Z.; Oura, T.; Baba, T.; Hamanishi, J.; Yamaguchi, K.; Kang, H. S.; Okamoto, T. The activated transforming growth factor-beta signaling pathway in peritoneal metastases is a potential therapeutic target in ovarian cancer. International Journal of Cancer 2012, 130((1)), 20–28. [Google Scholar] [CrossRef]
- Guo, Y.; Cui, W.; Pei, Y.; Xu, D. Platelets promote invasion and induce epithelial to mesenchymal transition in ovarian cancer cells by TGF-β signaling pathway. Gynecologic oncology 2019, 153((3)), 639–650. [Google Scholar] [CrossRef]
- Cui, Z. Y.; Park, S. J.; Jo, E.; Hwang, I.-H.; Lee, K.-B.; Kim, S.-W.; Kim, D. J.; Joo, J. C.; Hong, S. H.; Lee, M.-G. Cordycepin induces apoptosis of human ovarian cancer cells by inhibiting CCL5-mediated Akt/NF-κB signaling pathway. Cell death discovery 2018, 4((1)), 62. [Google Scholar] [CrossRef]
- Jang, H.-J.; Yang, K. E.; Hwang, I.-H.; Huh, Y. H.; Kim, D. J.; Yoo, H.-S.; Park, S. J.; Jang, I.-S. Cordycepin inhibits human ovarian cancer by inducing autophagy and apoptosis through Dickkopf-related protein 1/β-catenin signaling. American journal of translational research 2019, 11((11)), 6890. [Google Scholar]
- Choi, H. J.; Heo, J. H.; Park, J. Y.; Jeong, J. Y.; Cho, H. J.; Park, K. S.; Kim, S. H.; Moon, Y. W.; Kim, J. S.; An, H. J. A novel PI3K/mTOR dual inhibitor, CMG002, overcomes the chemoresistance in ovarian cancer. Gynecologic Oncology 2019, 153((1)), 135–148. [Google Scholar] [CrossRef]
- Ghoneum, A.; Said, N. PI3K-AKT-mTOR and NFκB pathways in ovarian cancer: implications for targeted therapeutics. Cancers 2019, 11((7)), 949. [Google Scholar] [CrossRef]
- Wang, D.; Wang, M.; Jiang, N.; Zhang, Y.; Bian, X.; Wang, X.; Roberts, T. M.; Zhao, J. J.; Liu, P.; Cheng, H. Effective use of PI3K inhibitor BKM120 and PARP inhibitor Olaparib to treat PIK3CA mutant ovarian cancer. Oncotarget 2016, 7((11)), 13153. [Google Scholar] [CrossRef]
- Jin, Z.; Niu, H.; Wang, X.; Zhang, L.; Wang, Q.; Yang, A. Preclinical study of CC223 as a potential anti-ovarian cancer agent. Oncotarget 2017, 8((35)), 58469. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Acharya, S.; Karthikeyan, M.; Biswas, P.; Kumari, S. Limitations and potential of immunotherapy in ovarian cancer. Frontiers in Immunology 2024, 14, 1292166. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R. L.; Cummings, M.; Thangavelu, A.; Theophilou, G.; de Jong, D.; Orsi, N. M. Barriers to immunotherapy in ovarian cancer: metabolic, genomic, and immune perturbations in the tumour microenvironment. Cancers 2021, ((24)), 6231. [Google Scholar] [CrossRef] [PubMed]
- Moore, K. N.; Pignata, S., Trials in progress: IMagyn050/GOG 3015/ENGOT-OV39. A Phase III, multicenter, randomized study of atezolizumab versus placebo administered in combination with paclitaxel, carboplatin, and bevacizumab to patients with newly-diagnosed stage III or stage IV ovarian, fallopian tube, or primary peritoneal cancer. International journal of gynecologic cancer 2019, 29, (2).
Figure 1.
Mechanisms of ovarian cancer tropism. OC metastasizes through multiple routes, including transcoelomic, hematogenous, and lymphatic dissemination. Transcoelomic metastasis involves the direct spread of cancer cells to the peritoneal cavity, where they attach to the mesothelial lining. Hematogenous metastasis occurs when CTCs enter the bloodstream, enabling distant organ dissemination. Lymphatic metastasis implies cancer cells spreading via lymphatic vessels. The TME comprises CAFs, MSCs, Tregs, immune cells, and mesothelial cells, which overall contribute to tumor progression and immune evasion. They play important roles in promoting metastasis through interactions within TME. OC, ovarian cancer; CTC, circulating tumor cell; CAFs, cancer-associated fibroblasts; MSCs, mesenchymal stem cells; Treg, regulatory T cell; TME, tumor microenvironment.
Figure 1.
Mechanisms of ovarian cancer tropism. OC metastasizes through multiple routes, including transcoelomic, hematogenous, and lymphatic dissemination. Transcoelomic metastasis involves the direct spread of cancer cells to the peritoneal cavity, where they attach to the mesothelial lining. Hematogenous metastasis occurs when CTCs enter the bloodstream, enabling distant organ dissemination. Lymphatic metastasis implies cancer cells spreading via lymphatic vessels. The TME comprises CAFs, MSCs, Tregs, immune cells, and mesothelial cells, which overall contribute to tumor progression and immune evasion. They play important roles in promoting metastasis through interactions within TME. OC, ovarian cancer; CTC, circulating tumor cell; CAFs, cancer-associated fibroblasts; MSCs, mesenchymal stem cells; Treg, regulatory T cell; TME, tumor microenvironment.
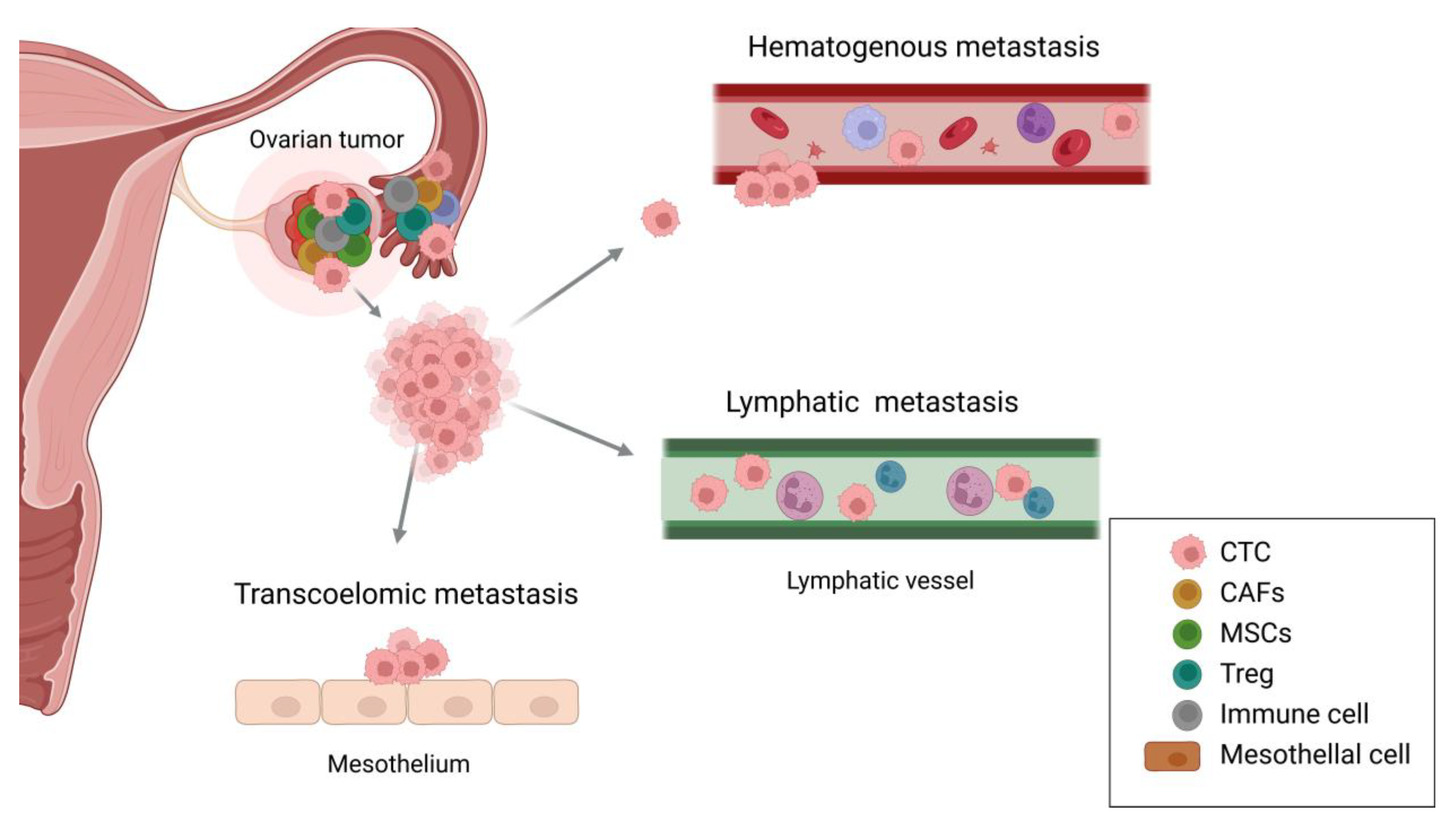
Figure 2.
Key signaling pathways involved in ovarian cancer metastasis. It illustrates the molecular interactions among major signaling pathways driving OC metastasis, including TGF-β, NF-κB, and PI3K/AKT/mTOR pathways. Upon activation, TGF-β receptors phosphorylate SMAD proteins, which then associate with SMAD4 and translocate to the nucleus to regulate gene expression, enhancing EMT, angiogenesis, and metastasis. SMAD7 acts as a negative regulator, inhibiting this signal. RTKs and TGF-β activate PI3K, leading to the generation of PIP3 and the recruitment of AKT. Stimulated AKT phosphorylates downstream targets, including mTOR complexes, which modulate cell growth, proliferation, and survival via effectors like 4EBP-1 and S6K-1. PTEN negatively regulates this pathway by dephosphorylating PIP3. Crosstalk between the PI3K/AKT and NF-κB pathways amplifies metastatic processes. AKT activation facilitates NF-κB signaling by promoting the phosphorylation and degradation of IκB, releasing the p50/p65 complex for nuclear translocation. This cascade controls the expression of genes involved in inflammation, immune evasion, EMT, and stemness. OC, ovarian cancer; TGF-β, transforming growth factor-β; NF-κB, nuclear factor of kappa-light chain of enhancer-activated B cells; PI3K, phosphoinositide 3-kinase; AKT, protein kinase B; mTOR, mammalian target of rapamycin; RTK, receptor tyrosine kinase; PIP2, phosphatidylinositol 4,5-bisphosphate; PIP3, phosphatidylinositol (3,4,5)-triphosphate; PTEN, phosphatase and tensin homolog; INPP4B, inositol polyphosphate 4-phosphatase B; PDK1, phosphoinositide-dependent protein kinase 1; SMAD, suppressor of mothers against decapentaplegic; MEK, mitogen-activated protein kinase; ERK, extracellular signal-regulated kinase.
Figure 2.
Key signaling pathways involved in ovarian cancer metastasis. It illustrates the molecular interactions among major signaling pathways driving OC metastasis, including TGF-β, NF-κB, and PI3K/AKT/mTOR pathways. Upon activation, TGF-β receptors phosphorylate SMAD proteins, which then associate with SMAD4 and translocate to the nucleus to regulate gene expression, enhancing EMT, angiogenesis, and metastasis. SMAD7 acts as a negative regulator, inhibiting this signal. RTKs and TGF-β activate PI3K, leading to the generation of PIP3 and the recruitment of AKT. Stimulated AKT phosphorylates downstream targets, including mTOR complexes, which modulate cell growth, proliferation, and survival via effectors like 4EBP-1 and S6K-1. PTEN negatively regulates this pathway by dephosphorylating PIP3. Crosstalk between the PI3K/AKT and NF-κB pathways amplifies metastatic processes. AKT activation facilitates NF-κB signaling by promoting the phosphorylation and degradation of IκB, releasing the p50/p65 complex for nuclear translocation. This cascade controls the expression of genes involved in inflammation, immune evasion, EMT, and stemness. OC, ovarian cancer; TGF-β, transforming growth factor-β; NF-κB, nuclear factor of kappa-light chain of enhancer-activated B cells; PI3K, phosphoinositide 3-kinase; AKT, protein kinase B; mTOR, mammalian target of rapamycin; RTK, receptor tyrosine kinase; PIP2, phosphatidylinositol 4,5-bisphosphate; PIP3, phosphatidylinositol (3,4,5)-triphosphate; PTEN, phosphatase and tensin homolog; INPP4B, inositol polyphosphate 4-phosphatase B; PDK1, phosphoinositide-dependent protein kinase 1; SMAD, suppressor of mothers against decapentaplegic; MEK, mitogen-activated protein kinase; ERK, extracellular signal-regulated kinase.
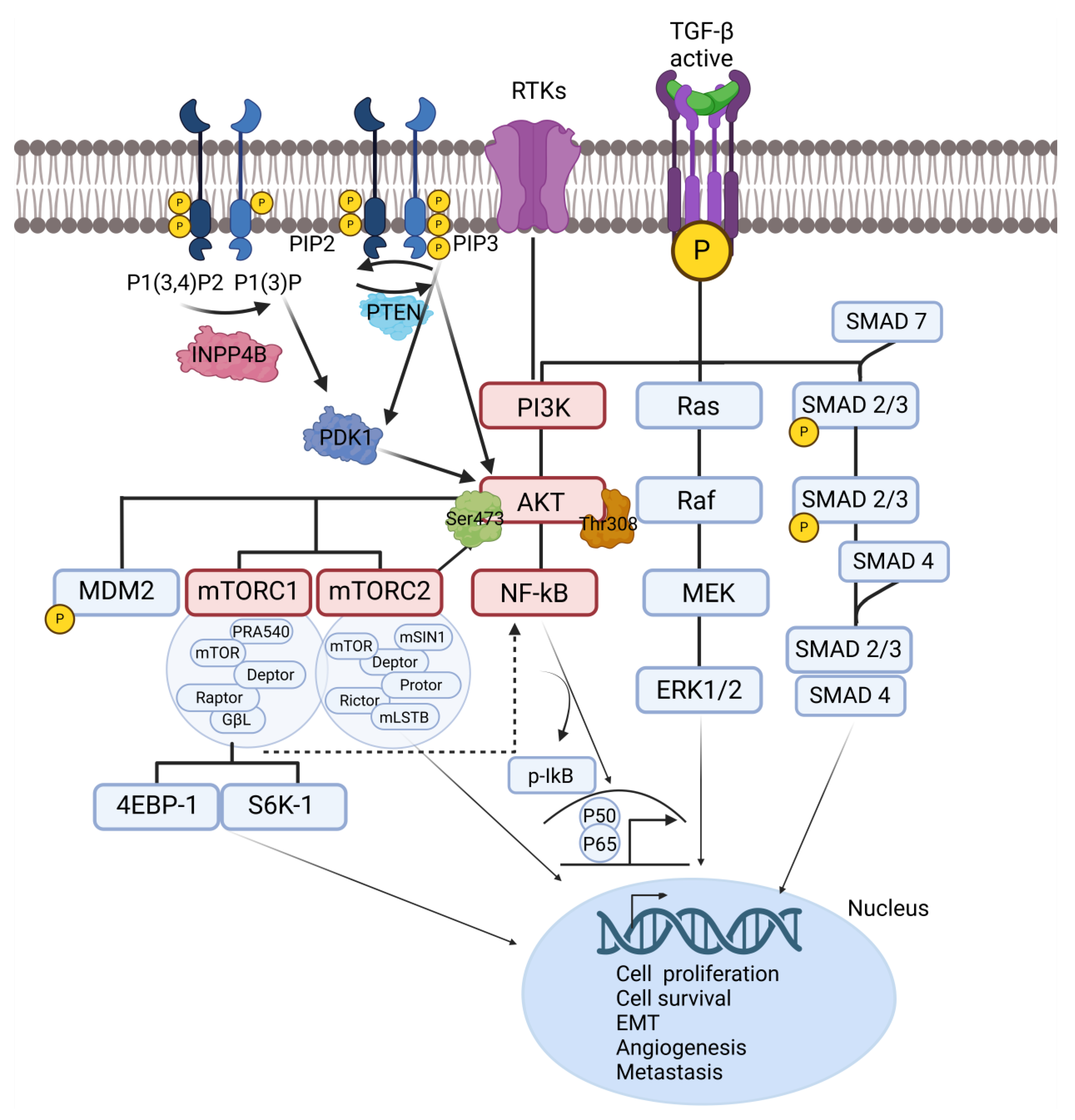

Table 1.
Therapeutic approaches in Ovarian cancer metastasis.
| Type of Therapy | Related Signaling pathway | Drug | Function | Reference |
|---|---|---|---|---|
| Combination immunotherapy |
- | NK cell therapy with oncolytic virus |
Infect cancer cells to enhance immune response and anti-tumor effect | [68] |
| - | IMAgyn050 | Target PD-L1 (atezolizumab) with bevacizumab | [87] | |
| TGF-β | LY2157299 | Small molecule TGF-β inhibitor with paclitaxel and carboplatin | [73] | |
| TGF-β | Bintrafusp alfa | Block both TGF-β and PD-L1 to promote immune reaction including CD4, CD8, and NK T cells | [75] | |
| Signaling pathway targeted therapy |
TGF-β | A-83-01 | Suppress EMT, MMP2, and pSMAD2 to regulate cell invasion and adhesion | [77,78] |
| AKT/NF-κB | Cordycepin | Modulate cell proliferation, inflammation, anti-cancer effect as a polyadenylation inhibitor | [79] | |
| PI3K/mTOR | CMG002 | Induce apoptosis and G1 cell cycle arrest to address chemoresistance | [82] | |
| PI3K | BKM120 | Repress cell growth, migration, and invasion with PARP inhibitor Olaparib | [83] | |
| mTOR | CC223 | Degrade mTORC complexes to inhibit cell proliferation and upregulate ROS generation | [82,84] |
IMagyn050, NCT03038100; LY2157299, galunisertib, NCT03206177; TGF-β, transforming growth factor-β; NF-κB, nuclear factor of kappa-light chain of enhancer-activated B cells; PI3K, phosphoinositide 3-kinase; AKT, protein kinase B; mTOR, mammalian target of rapamycin; PD-L1, programmed cell death ligand; EMT, epithelial-to-mesenchymal transition; NK, natural killer; MMP, matrix metalloproteinases; pSMAD, phosphorylated suppressor of mothers against decapentaplegic; PARP, poly (ADP-ribose) polymerase; ROS, reactive oxygen species.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



