Submitted:
27 January 2025
Posted:
28 January 2025
You are already at the latest version
Abstract
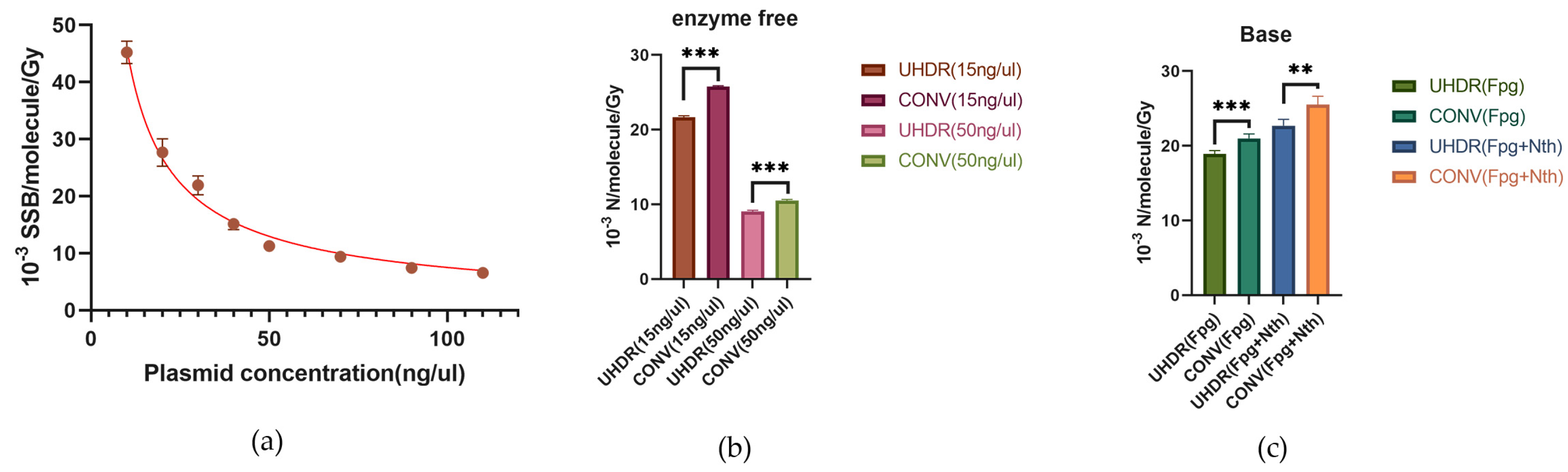
Background: The study of different types of DNA damage after ultra-high dose rate irradiation (UHDR) is of great significance for further understanding the mechanism of FLASH effect. Methods: pBR322 plasmid DNA was irradiated by electron FLASH beam. The content of each subtype of plasmid DNA was measured by gel electrophoresis, and the extent of DNA double strand break (DSBs) and single strand break (SSBs) under UHDR and conventional dose rate irradiation (CONV) was quantitatively compared. Further, by adding the endonuclease Nth and Fpg, the extent of base damage in UHDR and CONV group was quantitatively analyzed. In addition, the effects of different plasmid concentrations on the damage degree were also studied. Results: The induction rates of SSBs (×10−3 SSB/Gy/molecule) of UHDR and CONV were 21.7±0.4 and 25.8±0.3, respectively. When treated with Fpg and Nth enzyme, base damage induction rates (×10−3 SSB/Gy/molecule) of UHDR and CONV were 43.3±2.0 and 58.4±4.5, respectively. Additionally, it was found that plasmid concentration also affected the degree of damage, with lower SSB induction at higher plasmid concentration for both FLASH and CONV. For high-concentration plasmids, the induction rates of SSBs of UHDR was still 14% lower than that of CONV. Conclusions: UHDR has significant effect on reducing SSB and base damage compared to CONV regardless of plasmid concentration.
Keywords:
1. Introduction
2. Results
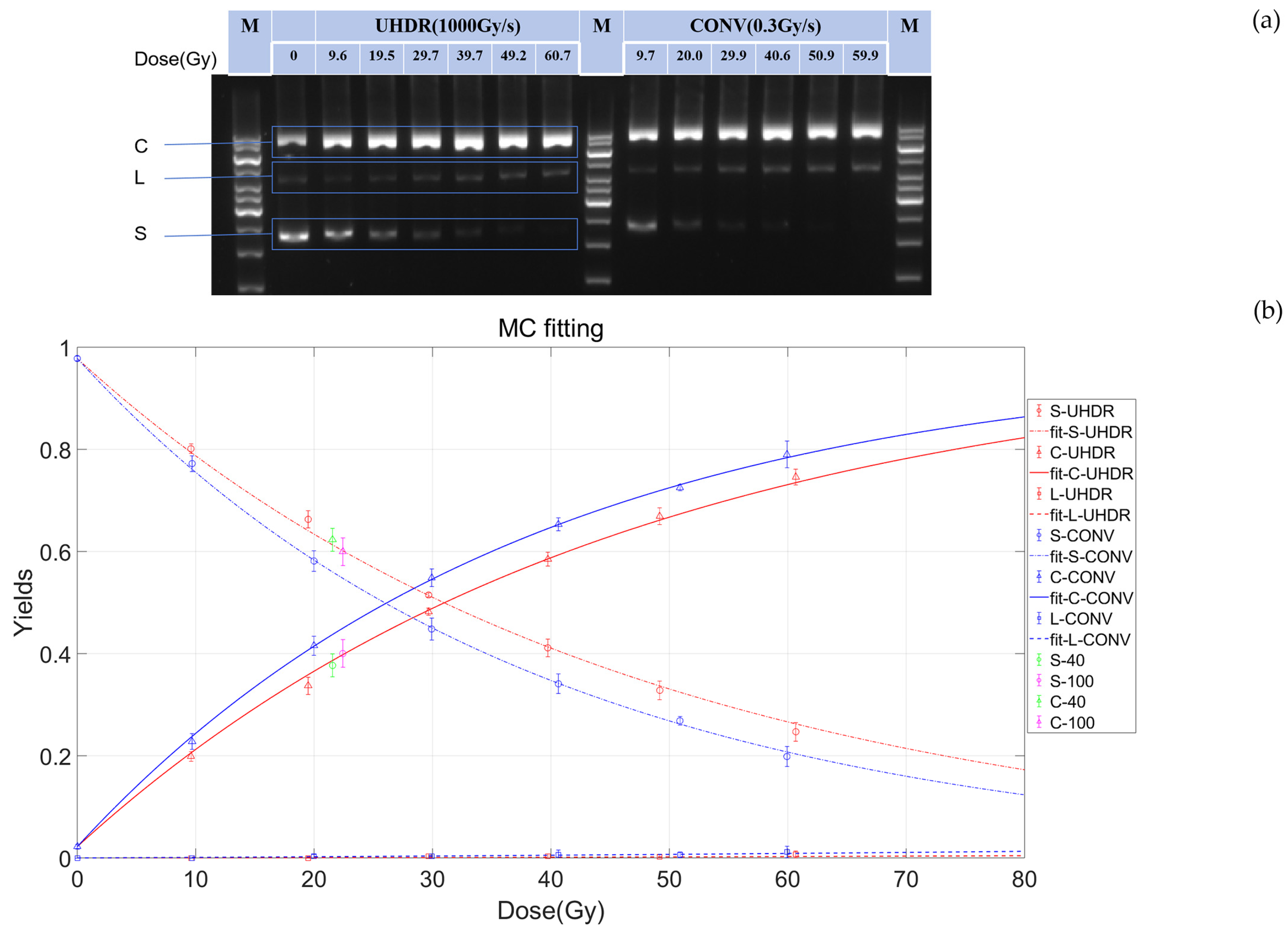
2.1. Comparison of SSBs Between CONV-RT and FLASH-RT
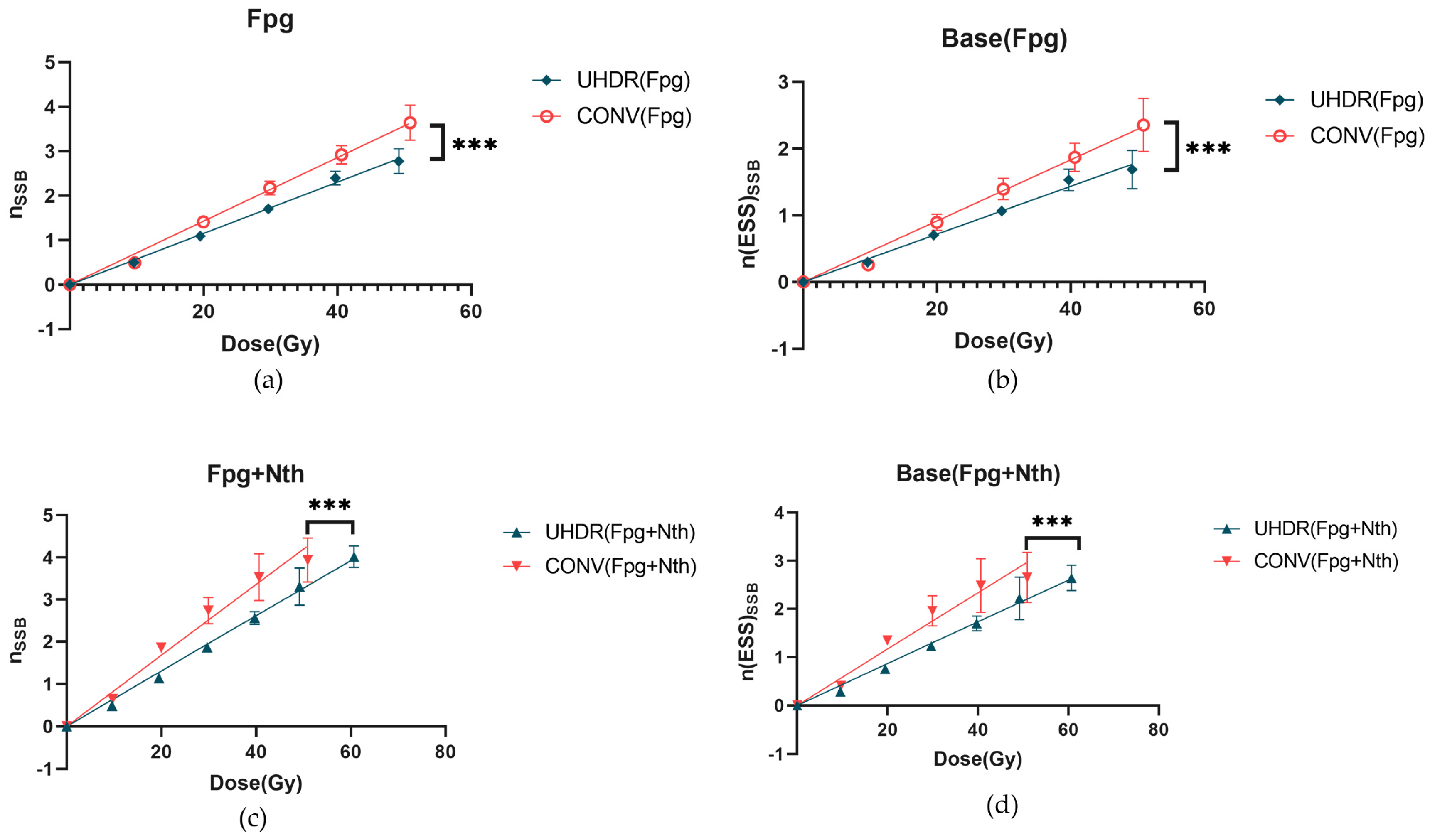
2.2. The Differences in Base Damage Under FLASH and CONV Conditions
2.3. The Effect of Plasmid Concentration on Radiation-Induced SSBs and Base Damage
3. Discussion
4. Materials and Methods
4.1. Sample Preparation
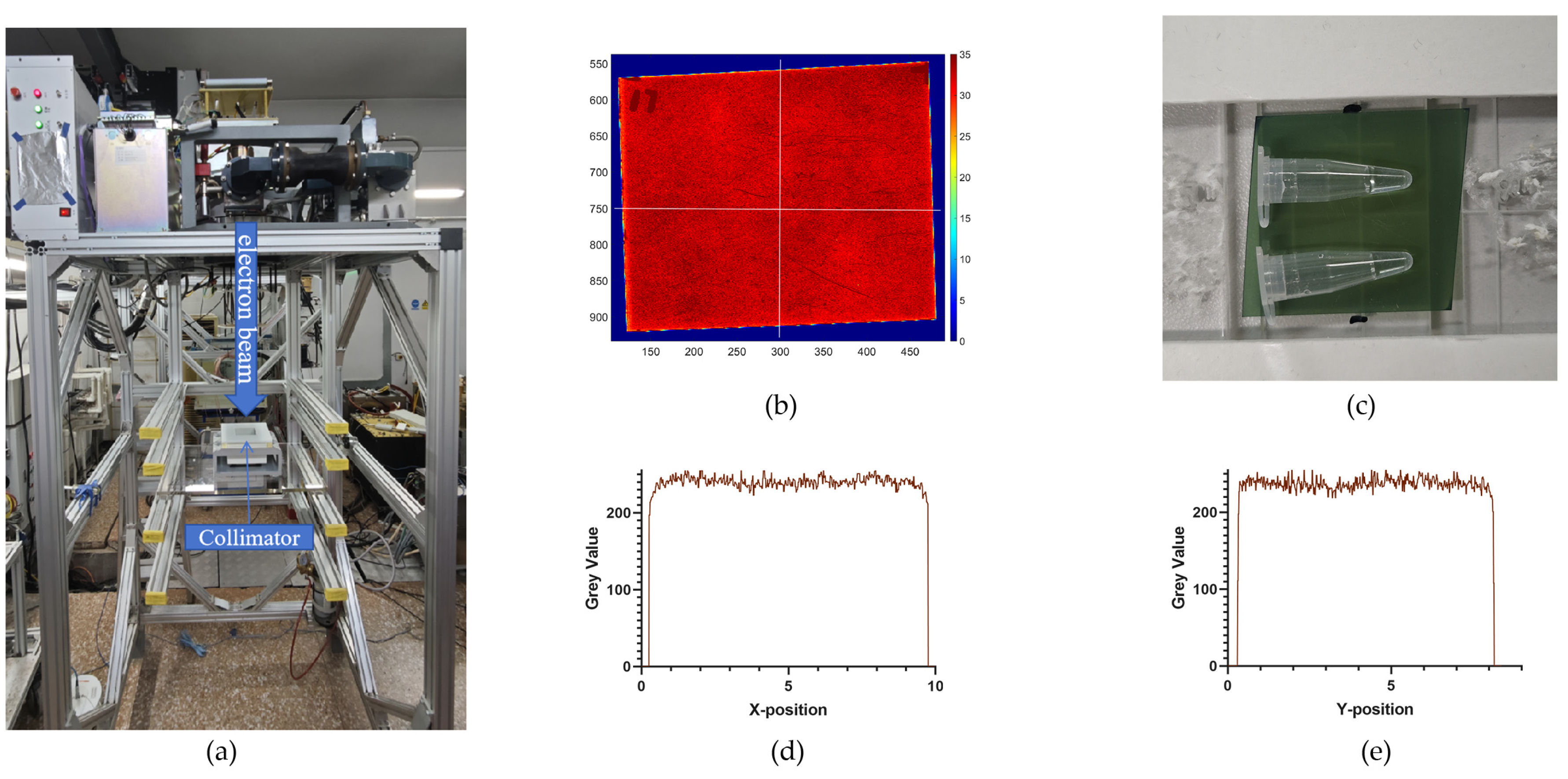
4.2. Irradiation
4.3. Enzyme Treatment
4.4. Agarose Gel Electrophoresis and Quantification of DNA Strand Breaks
4.5. DNA Damage Modeling
4.6. Base Damage Modeling
4.7. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gaide, O.; Herrera, F.; Jeanneret Sozzi, W.; Gonçalves Jorge, P.; Kinj, R.; Bailat, C.; Duclos, F.; Bochud, F.; Germond, J.-F.; Gondré, M.; et al. Comparison of Ultra-High versus Conventional Dose Rate Radiotherapy in a Patient with Cutaneous Lymphoma. Radiother Oncol 2022, 174, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Ohsawa, D.; Hiroyama, Y.; Kobayashi, A.; Kusumoto, T.; Kitamura, H.; Hojo, S.; Kodaira, S.; Konishi, T. DNA Strand Break sInduction of Aqueous Plasmid DNA Exposed to 30 MeV Protons at Ultra-High Dose Rate. J Radiat Res 2022, 63, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Yang, Y.; Zhu, H.; Wang, J.; Xiao, D.; Zhou, Z.; Dai, T.; Zhang, Y.; Feng, G.; Li, J.; et al. First Demonstration of the FLASH Effect with Ultrahigh Dose Rate High-Energy X-Rays. Radiother Oncol 2022, 166, 44–50. [Google Scholar] [CrossRef]
- Vozenin, M.-C.; De Fornel, P.; Petersson, K.; Favaudon, V.; Jaccard, M.; Germond, J.-F.; Petit, B.; Burki, M.; Ferrand, G.; Patin, D.; et al. The Advantage of FLASH Radiotherapy Confirmed in Mini-Pig and Cat-Cancer Patients. Clin Cancer Res 2019, 25, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Favaudon, V.; Caplier, L.; Monceau, V.; Pouzoulet, F.; Sayarath, M.; Fouillade, C.; Poupon, M.-F.; Brito, I.; Hupé, P.; Bourhis, J.; et al. Ultrahigh Dose-Rate FLASH Irradiation Increases the Differential Response between Normal and Tumor Tissue in Mice. Sci Transl Med 2014, 6, 245ra93. [Google Scholar] [CrossRef] [PubMed]
- Saade, G.; Bogaerts, E.; Chiavassa, S.; Blain, G.; Delpon, G.; Evin, M.; Ghannam, Y.; Haddad, F.; Haustermans, K.; Koumeir, C.; et al. Ultrahigh-Dose-Rate Proton Irradiation Elicits Reduced Toxicity in Zebrafish Embryos. Adv Radiat Oncol 2023, 8, 101124. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Yang, Y.; Zhang, W.; Wang, J.; Xiao, D.; Ren, H.; Wang, T.; Gao, F.; Liu, Z.; Zhou, K.; et al. FLASH X-Ray Spares Intestinal Crypts from Pyroptosis Initiated by cGAS-STING Activation upon Radioimmunotherapy. Proc Natl Acad Sci U S A 2022, 119, e2208506119. [Google Scholar] [CrossRef]
- Fouillade, C.; Curras-Alonso, S.; Giuranno, L.; Quelennec, E.; Heinrich, S.; Bonnet-Boissinot, S.; Beddok, A.; Leboucher, S.; Karakurt, H.U.; Bohec, M.; et al. FLASH Irradiation Spares Lung Progenitor Cells and Limits the Incidence of Radio-Induced Senescence. Clin Cancer Res 2020, 26, 1497–1506. [Google Scholar] [CrossRef]
- Fu, Q.-B.; Zhang, Y.; Wang, Y.-C.; Huang, T.-C.; Zhu, H.-Y.; Deng, X.-W. Systematic Analysis and Modeling of the FLASH Sparing Effect as a Function of Dose and Dose Rate. NUCL SCI TECH 2024, 35, 171. [Google Scholar] [CrossRef]
- Wanstall, H.C.; Korysko, P.; Farabolini, W.; Corsini, R.; Bateman, J.J.; Rieker, V.; Hemming, A.; Henthorn, N.T.; Merchant, M.J.; Santina, E.; et al. VHEE FLASH Sparing Effect Measured at CLEAR, CERN with DNA Damage of pBR322 Plasmid as a Biological Endpoint. Sci Rep 2024, 14, 14803. [Google Scholar] [CrossRef]
- Luo, H.; Yuan, Q.-G.; Phyllis, Z.; et al. Ultra-high dose rate irradiation induced DNA strand break in plasmid DNA[J]. Chin J Radiol Med Prot,2023,43(03):161-167. [CrossRef]
- Konishi, T.; Kusumoto, T.; Hiroyama, Y.; Kobayashi, A.; Mamiya, T.; Kodaira, S. Induction of DNA Strand Breaks and Oxidative Base Damages in Plasmid DNA by Ultra-High Dose Rate Proton Irradiation. Int J Radiat Biol 2023, 99, 1405–1412. [Google Scholar] [CrossRef] [PubMed]
- Perstin, A.; Poirier, Y.; Sawant, A.; Tambasco, M. Quantifying the DNA-Damaging Effects of FLASH Irradiation With Plasmid DNA. Int J Radiat Oncol Biol Phys 2022, 113, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Vozenin, M.-C.; Bourhis, J.; Durante, M. Towards Clinical Translation of FLASH Radiotherapy. Nat Rev Clin Oncol 2022, 19, 791–803. [Google Scholar] [CrossRef]
- Limoli, C.L.; Vozenin, M.-C. Reinventing Radiobiology in the Light of FLASH Radiotherapy. Annu Rev Cancer Biol 2023, 7, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Perstin, A.; Poirier, Y.; Sawant, A.; Tambasco, M. Quantifying the DNA-Damaging Effects of FLASH Irradiation With Plasmid DNA. Int J Radiat Oncol Biol Phys 2022, 113, 437–447. [Google Scholar] [CrossRef]
- Souli, M.P.; Nikitaki, Z.; Puchalska, M.; Brabcová, K.P.; Spyratou, E.; Kote, P.; Efstathopoulos, E.P.; Hada, M.; Georgakilas, A.G.; Sihver, L. Clustered DNA Damage Patterns after Proton Therapy Beam Irradiation Using Plasmid DNA. International Journal of Molecular Sciences 2022, 23, 15606. [Google Scholar] [CrossRef] [PubMed]
- Hatahet, Z.; Kow, Y.W.; Purmal, A.A.; Cunningham, R.P.; Wallace, S.S. New Substrates for Old Enzymes. 5-Hydroxy-2’-Deoxycytidine and 5-Hydroxy-2’-Deoxyuridine Are Substrates for Escherichia Coli Endonuclease III and Formamidopyrimidine DNA N-Glycosylase, While 5-Hydroxy-2’-Deoxyuridine Is a Substrate for Uracil DNA N-Glycosylase. J Biol Chem 1994, 269, 18814–18820. [Google Scholar] [PubMed]
- Dizdaroglu, M.; Laval, J.; Boiteux, S. Substrate Specificity of the Escherichia Coli Endonuclease III: Excision of Thymine- and Cytosine-Derived Lesions in DNA Produced by Radiation-Generated Free Radicals. Biochemistry 1993, 32, 12105–12111. [Google Scholar] [CrossRef] [PubMed]
- Labarbe, R.; Hotoiu, L.; Barbier, J.; Favaudon, V. A Physicochemical Model of Reaction Kinetics Supports Peroxyl Radical Recombination as the Main Determinant of the FLASH Effect. Radiother Oncol 2020, 153, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Froidevaux, P.; Grilj, V.; Bailat, C.; Geyer, W.R.; Bochud, F.; Vozenin, M.-C. FLASH Irradiation Does Not Induce Lipid Peroxidation in Lipids Micelles and Liposomes. Radiation Physics and Chemistry 2023, 205, 110733. [Google Scholar] [CrossRef]
- Kong, F.-Q.; Wang, X.; Ni, M.-N.; Sui, L.; Yang, M.-J. Effect of Concentration of DNA and Dose Rate in DNA Damage Induced by γ Ray[J]. Nuclear Physics Review, 2007, 24(2): 103-107. [CrossRef]
- Hanai, R.; Yazu, M.; Hieda, K. On the Experimental Distinction between Ssbs and Dsbs in Circular DNA. Int J Radiat Biol 1998, 73, 475–479. [Google Scholar] [CrossRef] [PubMed]
- McMahon, S.J.; Currell, F.J. A Robust Curve-Fitting Procedure for the Analysis of Plasmid DNA Strand Break Data from Gel Electrophoresis. Radiat Res 2011, 175, 797–805. [Google Scholar] [CrossRef]
- Povirk, L.F.; Wübter, W.; Köhnlein, W.; Hutchinson, F. DNA Double-Strand Breaks and Alkali-Labile Bonds Produced by Bleomycin. Nucleic Acids Res 1977, 4, 3573–3580. [Google Scholar] [CrossRef] [PubMed]

| Radiation quality | Energy (MeV) | Plasmid concentration (ng/ul) | Dose (Gy) | Mean dose rate (Gy/s) | Enzyme |
(×10−3 SSB/Gy/molecule) |
(×10−4 DSB/Gy/molecule) |
||||
| Mean | 95%CI | P-value | Mean | 95%CI | P-value | ||||||
| Electron* | 6 | 15 | 0-60 | UHDR | \ | 22.1 | ±0.4 | <0.0001 | 0.9 | ±0.4 | <0.0001 |
| CONV | 25.9 | ±0.5 | 1.5 | ±0.6 | |||||||
| UHDR | Fpg | 57.7 | ±1.7 | <0.0001 | 6.9 | ±1.3 | <0.0001 | ||||
| CONV | 71.4 | ±2.5 | 10.2 | ±1.3 | |||||||
| UHDR | Fpg+ Nth |
65.4 | ±2.0 | <0.0001 | 2.5 | ±1.5 | <0.0001 | ||||
| CONV | 84.0 | ±4.6 | 12.8 | ±1.8 | |||||||
| 50 | 0-40 | UHDR | \ | 9.0 | ±0.3 | 0.0004 | |||||
| CONV | 10.5 | ±0.3 | |||||||||
| UHDR | Fpg | 28.0 | ±1.2 | <0.0001 | |||||||
| CONV | 31.5 | ±1.4 | |||||||||
| UHDR | Fpg+ Nth |
31.7 | ±2.0 | 0.0003 | |||||||
| CONV | 36.0 | ±2.4 | |||||||||
| Electron [10] |
201 | 100 | 0-150 | 2E9 | \ | 9.8 | ±0.4 | \ | 2.6 | ±0.1 | \ |
| 0.08 | 13.3 | ±0.6 | 2.2 | ±0.3 | |||||||
| Electron [11] |
9 | 0-30 | 125 | 10.9 | ±0.4 | 1.1 | ±0.4 | ||||
| 0.05 | 11.8 | ±0.4 | 8.8 | ±0.4 | |||||||
| Proton [12] |
59.5 | 50 | 0-65 | 48.6 | 12.5 | ±1,0 | 2.5 | ±0.2 | |||
| 0.057 | 16.9 | ±1.0 | 2.7 | ±0.4 | |||||||
| 48.6 | Fpg | 20.1 | ±1.2 | 4.3 | ±0.4 | ||||||
| 0.057 | 26.2 | ±1.4 | 5.0 | ±0.4 | |||||||
| Proton [2] |
27.5 | 50 | 0-100 | 40 | \ | 8.8 | ±0.3 | 1.1 | ±0.3 | ||
| 0.05 | 10.8 | ±1.3 | 1.2 | ±0.4 | |||||||
| Electron [13] |
16 | 24 | 0-30 | 93.2 | 59.2 | ±4.9 | 5.4 | ±4.3 | |||
| 46.6 | 53.3 | ±3.1 | 5.4 | ±3.3 | |||||||
| 0.167 | 145.0 | ±27.4 | 8.1 | ±5.7 | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



