
Submitted:
26 January 2025
Posted:
26 January 2025
You are already at the latest version
Abstract
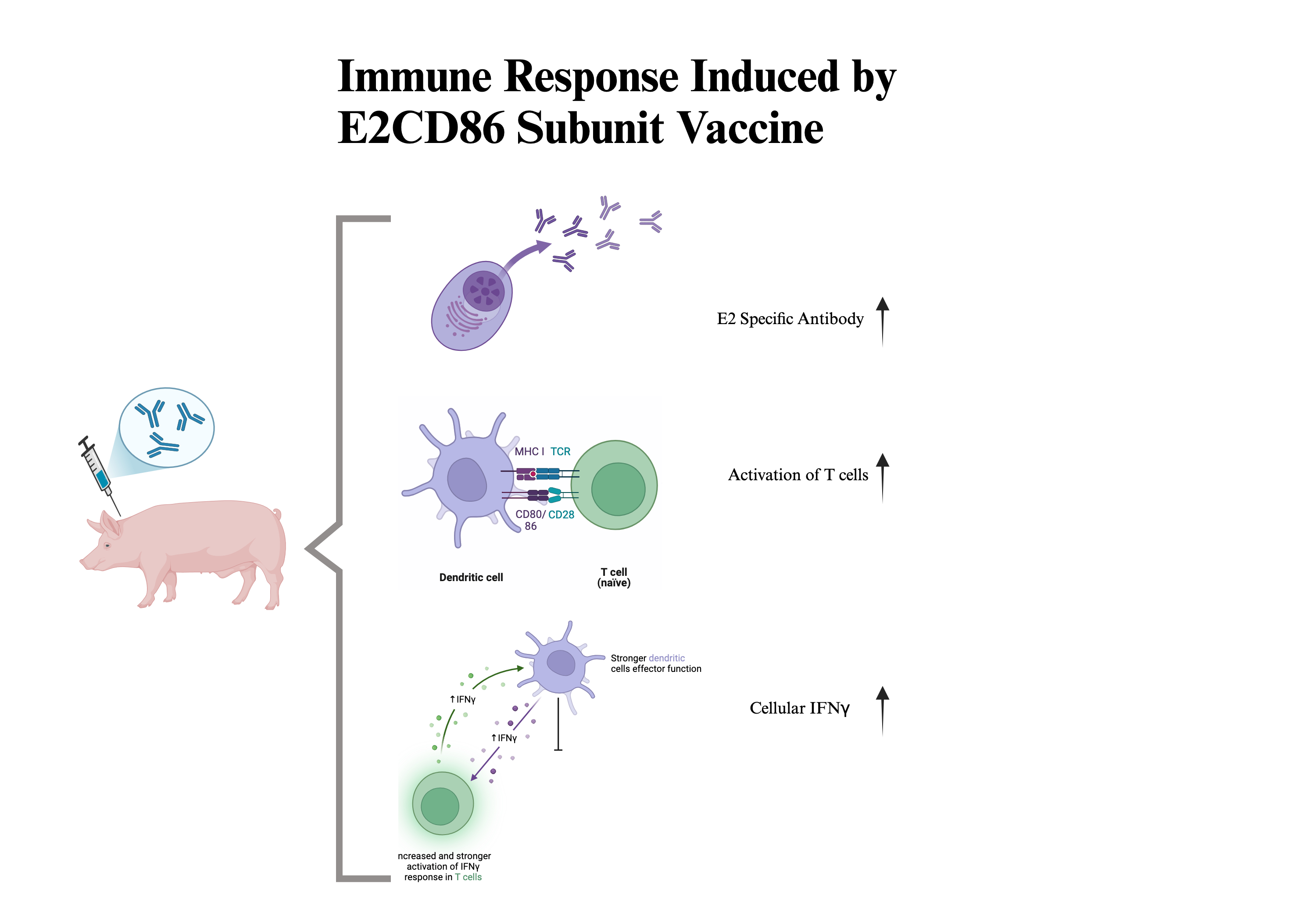
Background: The study of specific T-cell responses to vaccines against CSFV is important for understanding long-term immunity and infection management. The administration of a subunit vaccine against swine pathogens has been demonstrated to be safe and effective in infected pigs. However, subunit vaccines usually need an effective immunomodulatory adjuvant to stimulate active immune responses. Objectives: CD86 plays a crucial role in the immune response by enhancing the ability of APCs to stimulate the maturation and activation of T lymphocytes. Therefore, CD86 may act as a potential molecular adjuvant due to its ability to increase cytokine expression, which is protective against T-cell activation in pathogens. Methods: In this study, the CD86 coding sequence was linked with the CSFV E2 antigen sequence and then expressed in CHO cells for production of the fusion protein as an E2-CD86 vaccine. Commercial pigs aged 5–6 weeks were randomized into four groups: those inoculated with E2, E2-CD86, commercial Bayovac® CSF-E2 vaccines and the saline group. All animals were boosted 2 weeks after primary vaccination. Results: After vaccination, we evaluated the humoral response and compared it with that of T cells with the aim of correlating both types of responses and increasing CSFV-specific E2 antibody detection. The results revealed that the percentages of CD3+CD4+/CD8+, CD4+IL2+, IFNγ+ and CD4+CD8+ (DP) T cells among the PBMC 14 days after primary vaccination were significantly greater in the E2-CD86 group than in the other groups. The ELISpot results revealed a significant increase in the number of E2-specific IFNγ+ cells on D21 after vaccination. Conclusions: These results suggest that CD86 may enhance cellular immunity by increasing the levels of IL-2 and IFNγ-positive cells. Therefore, CD86 can act as a potential immuno-modulatory adjuvant in the development of subunit vaccines because of its protective efficacy.
Keywords:
CD86
; interferon gamma
; CSFV
; cellular immunity
; subunit vaccine
; molecular
1. Introduction
Classical swine fever (CSF) has been deemed a destructive disease for the pig industry through economic and health issues in many countries [1]. The causative agent is CSF virus (CSFV), an enveloped, single-stranded RNA virus belonging to the Pestivirus genus and Flaviviridae family [2]. In prevalent CSFV regions, vaccination with CSFV live attenuated viruses (LAVs) is still a rapid, effective and relatively strong way to protect against this disease [3]. However, modified live vaccines (MLVs) do not allow for the differentiation of infected animals within a vaccinated population, for example, differentiating infected from vaccinated animals (DIVA). Additionally, MLVs require a severe cold chain of delivery, which can be a general problem for low-income countries [4]. Hence, vaccination has been one of the most cost-effective health investments, with verified approaches that make it available to even the most hard-to-reach and vulnerable animals. To overcome the drawbacks of MLVs, subunit recombinant marker vaccines have been developed as promising preventive agents against CSFV [5].
CSFV E2, which is one of the major structural and immunogenic glycoproteins, mediates virus entry into target cells, for example, through adsorption to host cells and interaction with several host proteins. The E2 glycoprotein has been effectively used to induce a protective neutralizing antibody (NAb antibody) response against CSFV [6,7,8]. Several commercial vaccines based on the E2 glycoprotein have been developed [9,10]. However, compared with those of MLVs, the late induction of adaptive immunity and poorer efficacy of subunit CSFV E2 recombinant vaccines provide only partial prevention of vertical transmission [11,12,13]. Several nonstructural proteins of CSFV such as NS2, NS3, and NS4A [14,15], have potential cytotoxic T lymphocyte (CTL) epitopes. However, the NS3 protein to the E2 subunit vaccine does not obviously improve antibody and T-cell responses [16]. The use of cytokines to provoke T-cell responses is another option additionally. Vaccination with the CSFV-E2 antigen coformulated with interferon-α (IFN-α) prevent viremia and clinical symptoms at 7 days postvaccination (dpv) [17]. This information suggests a potential trend for future vaccine improvement. As E2 vaccines can induce antibody responses, enhancing the T-cell immunity of subunit vaccines is necessary.
The initiation of the T cell immune response is a complex process that involves the engagement of T cells with specialized antigen-presenting cells (APCs), including macrophages, dendritic cells and B cells. The APCs usually express high level of class I and II MHC, costimulatory and adhesion molecules. Moreover, the APCs deliver serious secondary signals to T cells through the B7 costimulatory molecules with CD28/CTLA-4 present on T cells. These costimulatory signals are important for the clonal expansion and T cells differentiation. Among different costimulatory molecules, CD80 and CD86 have been showed to provide effective immune signals [18,19]. CD80/CD86 also play an important role in producing cytokines which are critical for T cell activation and efficient antigen presentation. The finding for the combination of anti-CD80 and anti-CD86 antibodies induced anergy that further supports the role of both molecules in primary response [20,21]. The ligation between CD80/CD86 and CD28 provides co-stimulatory signals to activate T cells and promote proliferation of T-cells, production of interleukin 2 (IL-2), and induction of the anti-apoptotic factor Bcl-xL [22,23]. Though CD80/CD86 interact with CTLA-4 or CD28 to regulate T-cell immunity, the main ligand for CTLA4 is CD80, while CD86 is the main ligand for CD28 [24,25,26]. On the other hand, CD86 is expressed constitutively by APCs while CD80 is expressed only after activation of these cells [27,28,29]. CD80 is commonly absent on unstimulated cells, whereas CD86 is constitutively expressed on DCs, macrophages, and monocytes [28]. Thus, CD86 may be critical in the early interactions between APCs and T cells during the induction phase of the immune response. Additionally, CD86 is expressed at low levels on monocytes and is upregulated through interferon-γ (IFN-γ) stimulation. IFN-γ increases the expression of CD86 on peripheral blood monocytes, peritoneal macrophages and B cells [29,30,31]. This may offer the potential benefit of improving the immune response and avoiding multidose reactogenicity or antivector immune responses. On the basis of these studies, the use of CD86 as a vaccine adjuvant to induce the immune response of CD8+ CTLs was investigated [32]. CD86 is composed of an immunopotentiator and a delivery system to induce the immune-stimulating activity of vaccine formulators [33].
To investigate this issue, we constructed an E2 protein combined with the CD86 molecule to develop a new adjuvant/delivery system and improve existing subunit vaccine immunity deficiency.
2. Materials and Methods
2.1. E2CD86 Vaccine Candidates
Briefly, the active ingredient of E2CD86 is a chimeric protein formed by the fusion of the extracellular region of the E2 glycoprotein in the Taiwan CSFV strain (patent number: US-2024209033-A1 and TW-I841147) and the extracellular segment of the swine CD86 molecule. The E2CD86 protein was expressed in a Chinese hamster ovary (CHO) cell expression system. The CSFV-E2 recombinant protein is encoded by a nucleic acid sequence from the E2 gene fragment (nucleotide positions 1021-2294) of CSFV subgroup 2.1a (AY526726.1) [34] and is modified for proper expression. The tertiary protein structures of E2 and CD86 were predicted following a modified approach based on a previously described methodology (Figure S1) [35]. The E2CD86 protein was synthesized by Genomics BioSci & Tech Co., Ltd. and named PTP-E2 and PTP-PCV2d. CHO cells were transfected with PTP-E2/pcDNA3.4 and PTP-PCV2d/pcDNA3.4 plasmid DNA according to the instructions of the commercially available transfection kit (ExpiFectamine™ CHO Transfection Kit, Thermo Fisher Scientific Inc., USA). The E2CD86 protein was formulated in MontanideTM ISA206 (SEPPIC, France) using a 50/50 proportion of aqueous/oil phase. The water-in-oil emulsion was produced with an SD-41 homogenizer (IKA, Germany) under good manufacturing practice (GMP) conditions. The concentration of E2CD86 in the final emulsion was 60 μg/mL.
2.2. Experimental Animals and Immunization Schedules
Healthy 5–6-week-old 25–30 kg swine belonging to a nonvaccinated and specific pathogen-free (SPF) herd were used (Agricultural Technology Research Institute, ATRI, Taiwan) to evaluate the safety and efficacy of the E2CD86 vaccine. According to the Guide for the Care and Use of Laboratory Animals, the swine were under good care and management. Vaccinated animals were carefully evaluated daily for clinical signs, inappetence, prostration and inflammatory reactions at the vaccination site, or other alterations that may be related, or not, to vaccination. The procedure for the animal experiment is presented in Figure 1.
A total of 14 swine were randomly separated into vaccinated E2CD86/ISA206 (n=4), E2/ISA206 (n=4), and placebo groups (10 mM NaH2PO4 pH 7.4, saline, n=2) and compared with the commercial Bayovac® CSF-E2 (Bay ®-E2, n=4) vaccine. The animals were immunized twice by intramuscular injection with 1 ml of formulation (60 µg of purified E2CD86 antigen). The vaccines were administered on day 0 and boosted on day 14. Clinical signs were observed daily, and the rectal temperature was measured after vaccination. Blood samples were collected via sterile tubes with or without anticoagulant from ophthalmic venous sinus punctures on days 0 (preimmune serum), 7, 14, 21 and 28 to evaluate specific E2 titers via ELISA and examine the immune response via flow cytometry and ELISPOT.
All animal experiments in this study were performed in accordance with the relevant guidelines and regulations. The experimental procedures involving pigs were reviewed and approved by the Institutional Animal Care and Use Committees (IACUC) of the National Pingtung University of Science and Technology (NPUST-IACUC NPUST-111–01).
2.3. Peripheral Blood Mononuclear Cell (PBMC) Isolation
Peripheral blood mononuclear cells (PBMC) were isolated from blood samples via density gradient centrifugation as previously described [36]. The use of 96% Ficoll-Paque™ is recommended. For Ficoll‒Paque density gradient centrifugation, the blood samples were treated with anticoagulant (EDTA) and diluted with PBS (10 mM NaH2PO4, pH 7.4), which takes advantage of differences in the cell density of the different blood components. Density gradient centrifugation of diluted whole blood layered over a density gradient medium yields PBMC. For visualization, 10 mL of diluted blood was layered onto 5 mL of Ficoll-Paque™. The samples were subsequently centrifuged at 400xg for 40 min at room temperature with the brake off. The PBMC were harvested after centrifugation and washed. The viability of the PBMC was determined via Trypan blue (Sigma‒Aldrich, St. Louis, MO, USA) staining, and more than 90% viability was detected. PBMC were subjected to an IFN-γ ELISPOT assay, and the cellular immune response was detected via a flow cytometry assay as described below.
2.4. CSFV Neutralizing Antibody Detection
Serum samples were isolated and measured for their ability to neutralize the E2 antibody against CSFV via a commercial ELISA kit, the IDEXX CSFV Ab Test Kit (IDEXX Laboratories, Inc., USA). The assay is a blocking ELISA for the detection of CSFV antibodies in swine serum and follows the steps recommended by the manufacturer. The CSFV ELISA antibody levels are expressed as the percentage of bacteria that were blocked and those with values of < 30 and > 40 were considered negative and positive, respectively. A blocking percentage between 30% to 40% requires further testing. The neutralization antibody titer against CSFV was measured in triplicate wells.
2.5. Flow Cytometry Analysis
Single-cell suspensions from PBMC were analyzed via a FACS II (BD Biosciences) flow cytometer equipped with two lasers (488 and 633 nm). To detect immune protein expression on the surface of the PBMC, we incubated 1X106 PBMC with ABflo® 488 rabbit anti-Pig CD4 mAb (A22774), FITC anti-porcine CD8 (76-2-11, SouthernBiotech), FITC anti-porcine interferon gamma (IFN-r) (P2C11, Mabtech, Sweden), anti-pig IL2 (MT264/267, Mabtech, Sweden) and APC anti-porcine CD3 (PPT3, SouthernBiotech) antibodies overnight at 4 °C. The cells were directly stained with 100 μL of an antibody containing APC-conjugated goat anti-mouse IgG (405308, Biolegend)-conjugated anti-pig IL2. The cells were subsequently washed once with FACS buffer by centrifugation at 400 × g for 5 min at 4 °C. Samples live cells were discarded to ensure convincing gating. Flow cytometry was performed at least two or three times for each individual experiment.
2.6. IFN-γ Enzyme-Linked Immunospot (ELISpot) Assay
IFN-γ ELISpot assays were performed via a porcine IFN-γ ELISpot ALP kit (3130-4APW-10, Mabtech, Sweden) according to the manufacturer’s protocol. The ELISpot assay was performed using PBMC drawn for vaccination after day 28. Briefly, prewetted filter plates were coated overnight at 4°C with the capture antibody P2C11 directed against porcine IFN-γ. The assay plate was rehydrated by washing with 1X PBS and blocked with RPMI media for 1 hour at RT. After being washed and blocked, PBMC were diluted in RPMI media to a concentration of 2.0 × 106 PBMC/mL and plated at 100 µl/well in triplicate for each stimulus (1.0 × 105 PBMC/well). The cells were stimulated with phytohemagglutinin-L (PHA, 5 μg/ml; Sigma‒Aldrich, St. Louis, MO, USA) and E2 protein (10 μg/ml) for 18–24 hours at 37°C in 5% CO2. PHA was used as a positive control stimulant. The spots were visualized after incubation with the substrate BCIP/NBT-plus for 15± 5 min at RT. Spots were counted via an automated spot counter (AID ELISPOT Reader, AID GmbH, Strassberg, Germany) within 24–72 h, and spot counts were analyzed as spot-forming units (SFUs) per well. The raw data of each well for visual quality checking and the corresponding spot-forming unit (SFU) counts per well.
2.7. Statistical Analysis
The t-test or ANOVA test was used for continuous data analyses, Chi-square test for nominal variables among group comparisons. Generalized linear model (GLM) was used to estimate the data analysis of the vaccine pigs for each sample. All data management and statistical analyses were performed using SAS® 9.4 software (version 9.4, SAS Institute Inc., Cary, NC, USA) and GraphPad Prism 8 (GraphPad Software, USA). Data were representing the mean values ± SEM and error bars indicate SEM. The Shapiro-wilk test was used to check the normal distribution of data by SAS® 9.4 software. All statistical tests with a two-sided p value of < 0.05 were considered statistically significant.
3. Results
3.1. The E2CD86 Vaccine Provides Enhanced Protection in Piglets
Body weight gain and rectal temperatures were measured at 24 and 48 hours after vaccination. The mean rectal temperatures in the E2CD86, Bay-E2, and E2-vaccinated and placebo (saline) groups were all less than 40.5 ± 0.1 °C before the 1st vaccination. Compared with those in the saline group, the rectal temperatures in the vaccinated group did not increase significantly, and no fever was observed within 48 h after the 1st and 2nd vaccinations (data not shown). No injection site reactions such as reddish lesions were observed after 12 hours of the 2nd vaccination. In addition, there were no significant differences in mean body weight gain or loss of appetite among the groups after injection.
At 7 days after vaccination, the pigs in all the groups were seronegative for CSFV. E2-specific antibodies started to appear at 14 dpv in the E2 and E2CD86 groups (Figure 2). In contrast, that of the E2CD86 group was greater than that of the E2 and Bay®-E2 groups. The data showed that the E2CD86 vaccine is a single-dose vaccination that can effectively induce E2-specific antibodies for 14 days. The increases in the antibody response among the E2, E2CD86 and Bay®-E2 groups were similar at 21 dpv, whereas all the pigs from the saline group were negative. The serological data indicated that E2-CD86 vaccines induced high titers of CSFV-specific NAbs in pigs.
3.2. The E2CD86 Vaccine Provides Enhanced Protection in Piglets
3.2.1. Immunogenicity of E2CD86 in Piglets with High Activation of T Cells
CD86 and its ligands are well known as major T-cell costimulatory pathways that regulate the T-cell immune response [37]. PBMC were isolated at 14 dpv to examine the cellular immune response. The E2CD86 group had the highest percentage of CD4+ T cells among all vaccine groups (E2CD86: 22.53 ± 1.72%, Bay®-E2: 19.8 ± 3.15% and saline: 18.25 ± 2.33%) (Figure 3A). Interleukin-2 (IL-2) is an important cytokine for T-cell immunity. IL-2-responsive T-cell subsets play various proinflammatory and anti-inflammatory biological roles. The major sources of IL-2 are activated CD4+ T and CD8+ T cells. Our preliminary data (data not shown) revealed that CD4+IL-2+ expression was activated at 3 dpv. Here, we examined the expression of IL-2 in CD4+ T cells after the E2CD86 vaccine. The data revealed that the E2CD86 group presented greater CD4+IL-2+ T-cell expression than did the Bay®-E2 group at 14 dpv (E2CD86: 0.9 ± 0.30%, Bay®-E2: 0.8 ± 0.57% and saline: 0.40 ± 0.42%). CD8+ T cells are very important for the immune system against pathogens, including viruses and bacteria. We also examined the ability of CD8+ T-cell populations to effectively enhance the ability of the E2CD86 vaccine to enhance the cellular immunity of cytotoxic T cells (CTLs). Compared with those of the Bay®-E2 group, those of the E2CD86 group were greater (E2CD86: 24.38 ± 4.00%, Bay®-E2: 23.68 ± 3.40% and saline: 17.50 ± 1.98%) at 14 dpv (Figure 3A). These results suggest that both CD8+ and CD4+ T cells elicited by the E2CD86 vaccine play critical roles in immunity by contributing to the induction of IL-2. On the other hand, CD4+ T cells are essential for the formation of protective memory CD8+ T cells following infection or immunization [38]. The E2CD86 vaccine induced memory T cells to provide protection against subsequent infections caused by the same pathogen or antigen.
3.2.2. Increased Number of CD4+CD8+ Double-Positive cells After the E2CD86 Vaccine
The T-cell population in peripheral blood and tissues expressing both CD4+ and CD8+ T cells simultaneously (CD4+CD8+ double-positive DP T cells) has been described [39]. Circulating DP T cells are composed of subpopulations with a variety of characteristics and functions [40], such as cytotoxic ability and memory immunity. The data presented herein demonstrated that DP T cells were enhanced by the E2CD86 vaccine at 14 dpv (E2CD86: 11.83 ± 0.68%, Bay®-E2: 10.67 ± 1.16% and saline: 10.25 ± 0.78%) and 28 dpv (E2CD86: 7.37 ± 0.58%, Bay®-E2: 6.83 ± 0.58% and saline: 6%) (Figure 3B). However, with the recognition of CD4/CD8 DP T cells as a diverse lymphocyte population, the composition of DP T-cell function needs further study [41].
3.3. Immunogenicity of E2CD86 in Piglets with High Cellular IFNγ Expression
To assess antigen-specific T-cell responses to the E2CD86 subunit vaccine, PBMC were isolated from vaccinated and saline-injected pigs at 10 and 21 dpv. The cells were plated onto ELISpot wells coated with an anti-IFNγ capture antibody and stimulated with E2 recombinant protein encoded by the E2CD86 vaccine. The quantitative data of representative wells are shown in Figure 4A, with a medium, a negative control group (unstimulated) and PHA as a positive control. The number of IFN-γ-positive spot-forming units (SFUs) in each well was calculated for all vaccine groups. In the unimmunized pigs, the response to the E2CD86 vaccine was the highest at 10 dpc (Figure 4A). Compared with 10 SFUs/106 PBMC across the BayE2 vaccine pigs, the IFN-γ ELISpot in response to stimulation with the E2 protein spanning the E2CD86 vaccine averaged 172.5 SFU/106 PBMC across the animals in the vaccine group compared to 10 SFUs/106 PBMC across the BayE2 vaccine pigs (Figure 4A). In addition, the flow cytometry data revealed that the IFN-γ frequency in the E2CD86 group was higher than that in the Bay®-E2 group (E2CD86: 2.3 ± 0.5% and Bay®-E2: 1.53 ± 1.11%) (Figure 4B).
3.2. Figures, Tables and Schemes
Figure 1.
Diagram of the animal experimental design. To compare the protective efficacy of the E2CD86 vaccine via the IM. A total of 60 µg of the E2CD86 vaccine was used to immunize pigs intramuscularly (i.m.) via the prime-boost strategy. A total of 28 days after immunization, the serum and PBMC were harvested and analyzed to assess protection.
Figure 1.
Diagram of the animal experimental design. To compare the protective efficacy of the E2CD86 vaccine via the IM. A total of 60 µg of the E2CD86 vaccine was used to immunize pigs intramuscularly (i.m.) via the prime-boost strategy. A total of 28 days after immunization, the serum and PBMC were harvested and analyzed to assess protection.
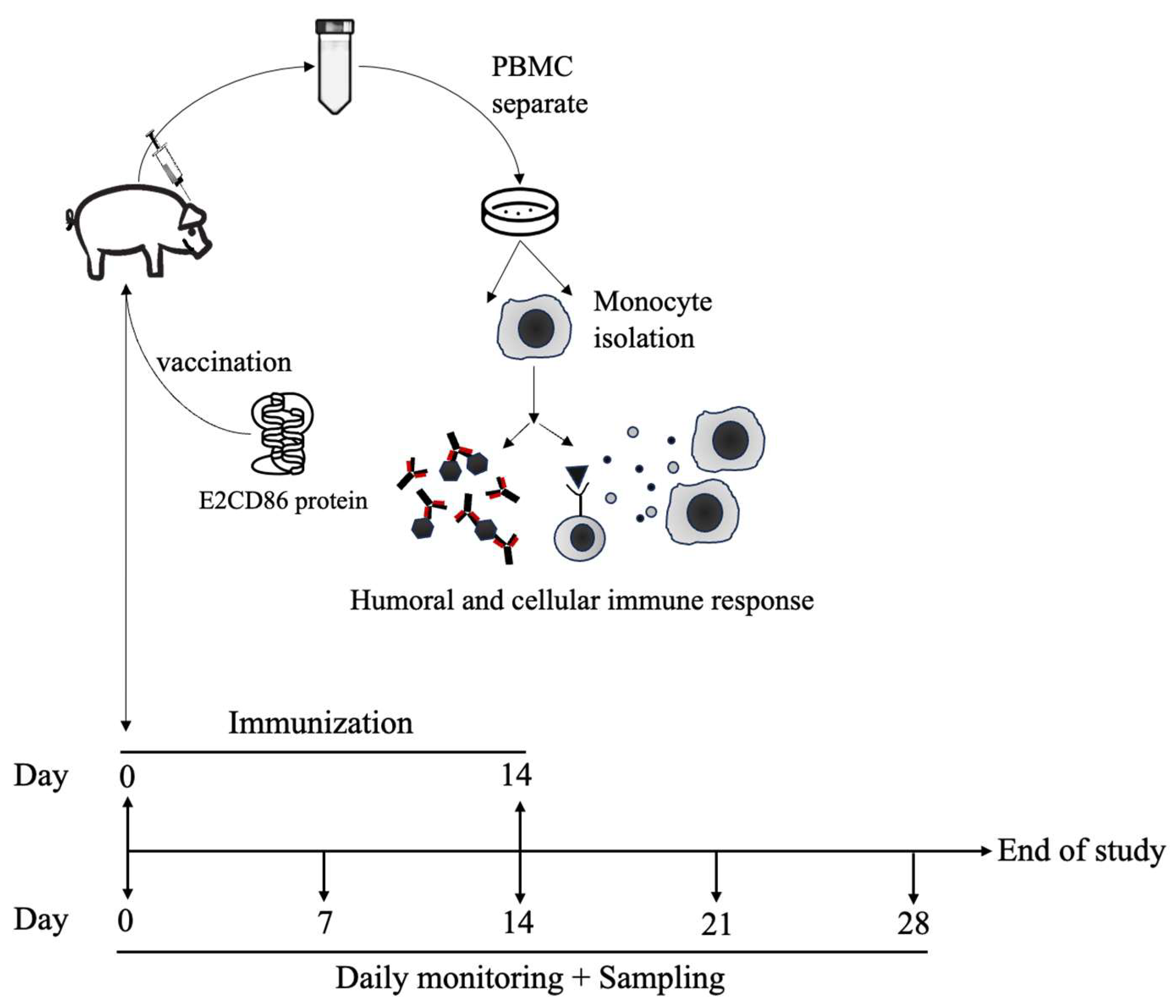
Figure 2.
E2 antibody response of the E2-CD86 subunit vaccine as determined by ELISA. All the serum samples were collected at 0, 7, 14, 21 and 28 days after vaccination. Groups enriched with E2 or E2-CD86 reacted positively (over 40%) on day 14. Compared with those of the E2 and E2-CD86 groups, the Bay®-E2 vaccine group presented lower percentage 14 days later.
Figure 2.
E2 antibody response of the E2-CD86 subunit vaccine as determined by ELISA. All the serum samples were collected at 0, 7, 14, 21 and 28 days after vaccination. Groups enriched with E2 or E2-CD86 reacted positively (over 40%) on day 14. Compared with those of the E2 and E2-CD86 groups, the Bay®-E2 vaccine group presented lower percentage 14 days later.
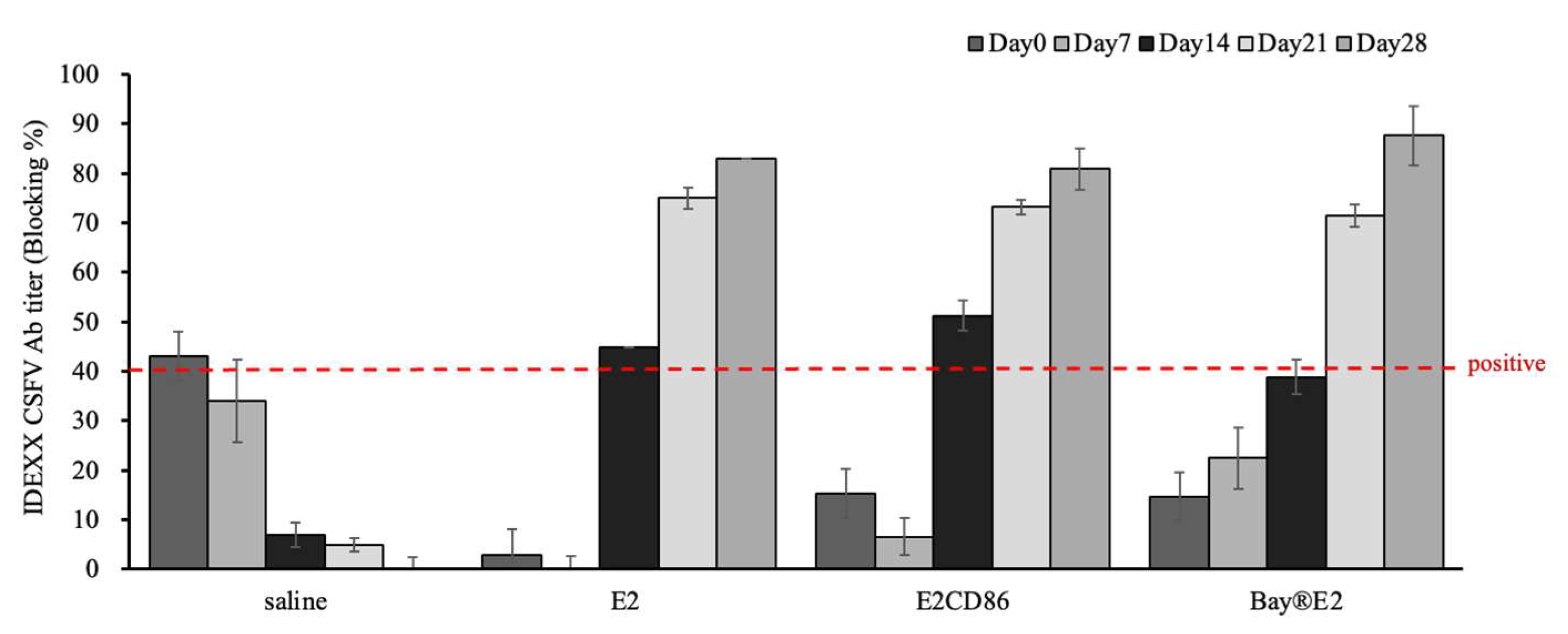
Figure 3.
E2CD86 vaccine-induced T cells display a sustained cellular immune response. PBMC from pigs vaccinated with subunit vaccines after 14 days and a further 28 days. The number of T cells was assessed via flow cytometry. Representative dot plots showing quadrant gates defining the expression profiles of (a) CD4, CD3 and IL-2 cytokines in CD4+ T cells. (b) CD4+CD8+ DP T-cell frequency was induced by the E2CD86 vaccine at 14 and 28 dpv.
Figure 3.
E2CD86 vaccine-induced T cells display a sustained cellular immune response. PBMC from pigs vaccinated with subunit vaccines after 14 days and a further 28 days. The number of T cells was assessed via flow cytometry. Representative dot plots showing quadrant gates defining the expression profiles of (a) CD4, CD3 and IL-2 cytokines in CD4+ T cells. (b) CD4+CD8+ DP T-cell frequency was induced by the E2CD86 vaccine at 14 and 28 dpv.
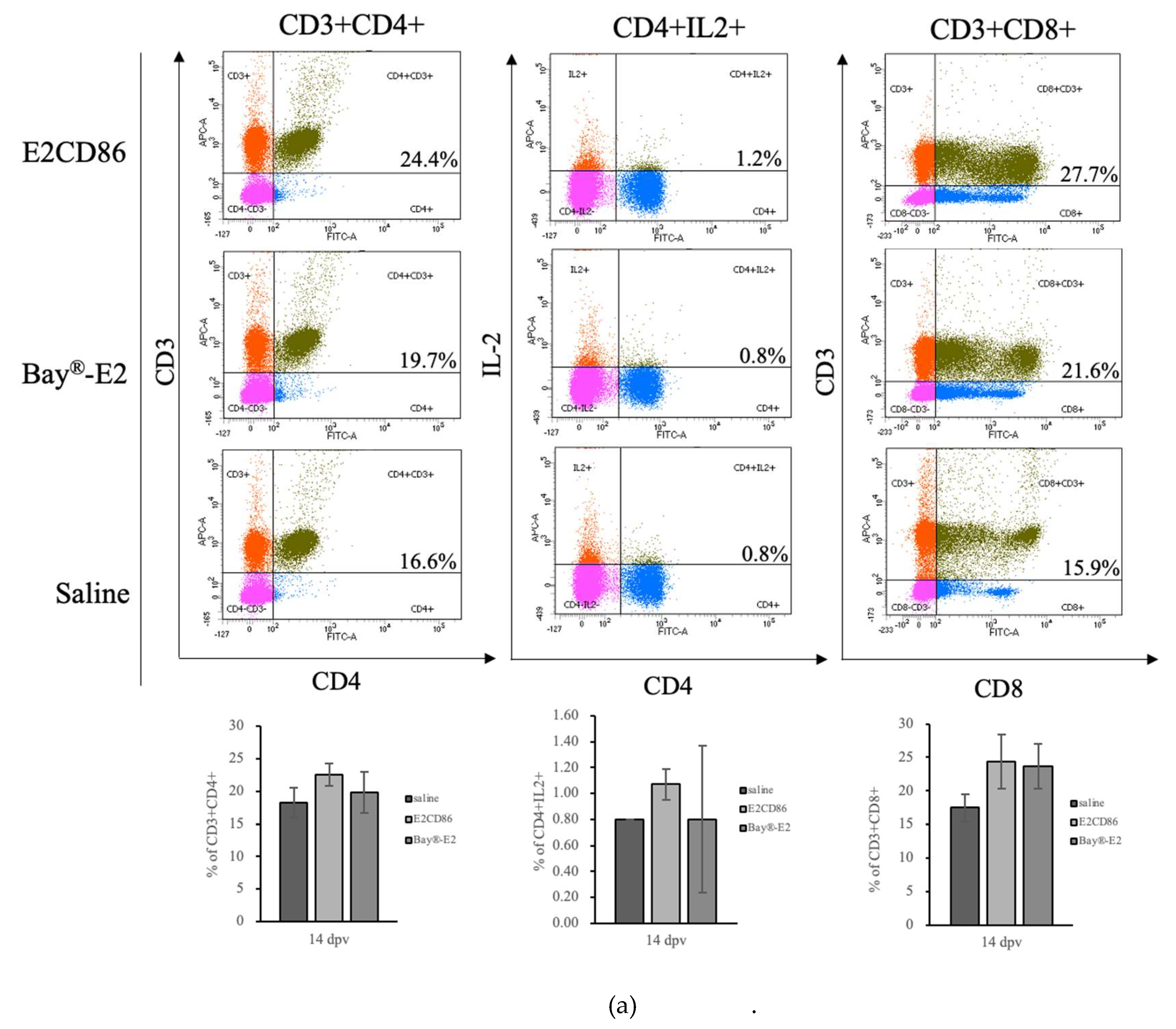
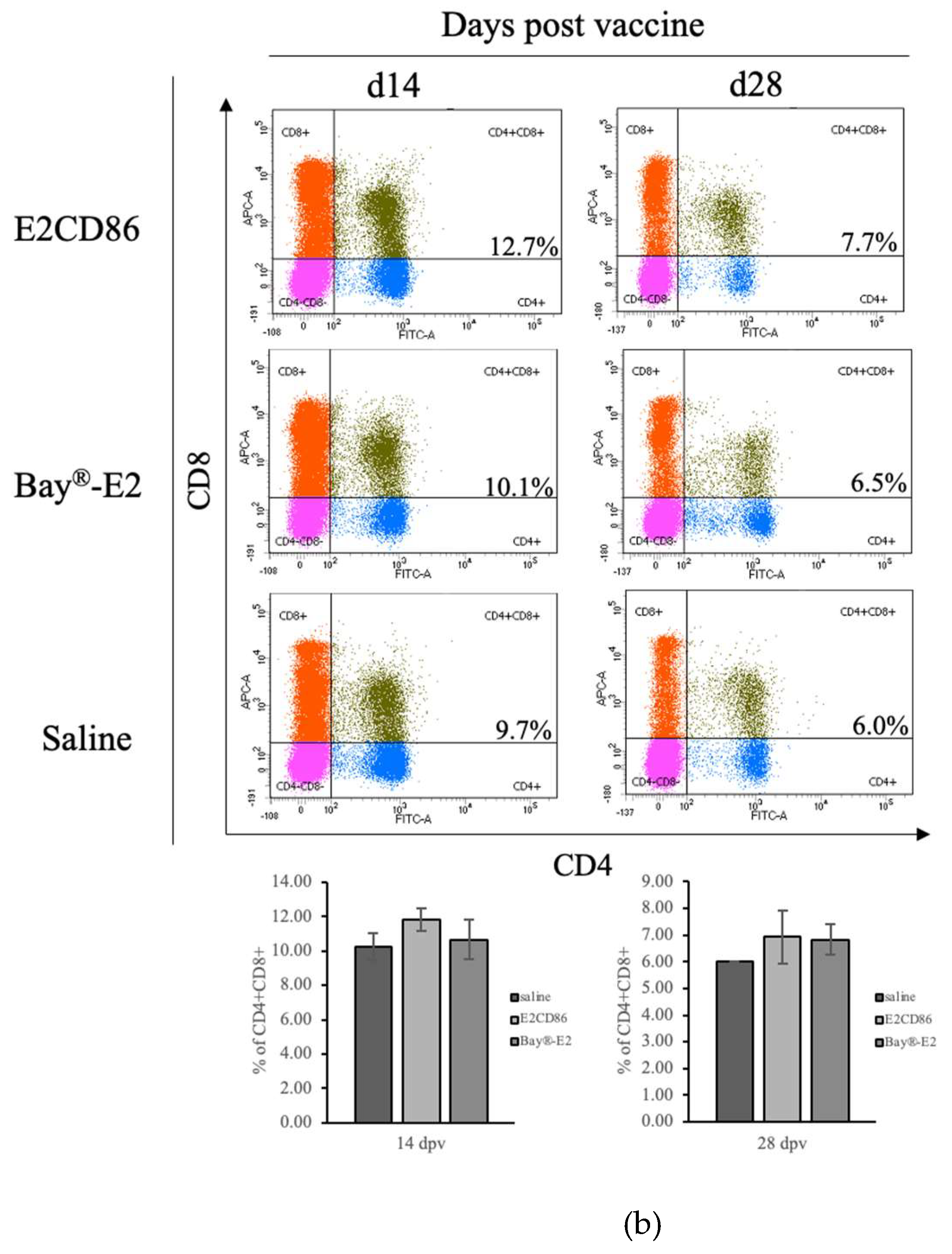
Figure 4.
Functional and phenotypic characterization of CSFV-specific IFN-γ expression.
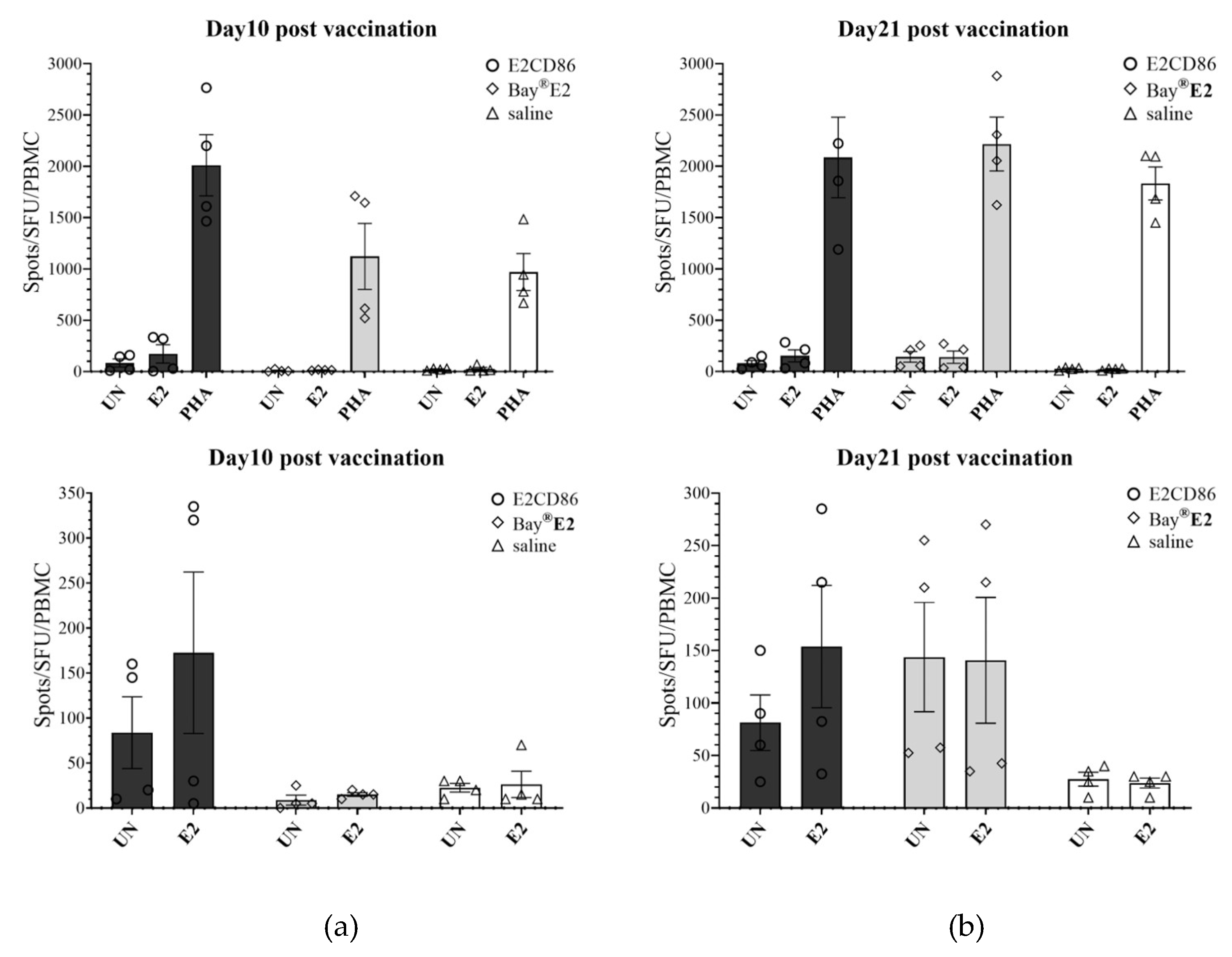
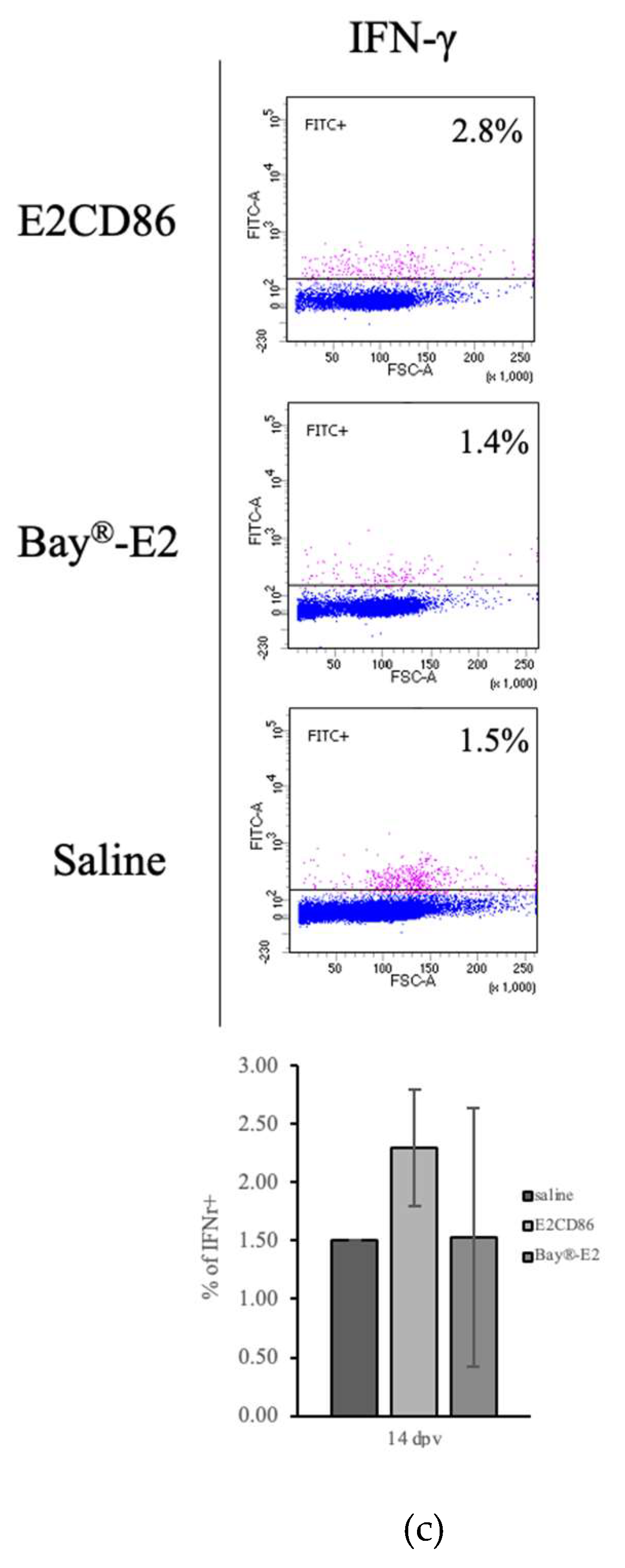
E2-specific IFN-γ expression was detected via an ELISpot assay. IFN-γ expression in unstimulated (UN) or stimulated E2 protein (10 µg/ml) and PHA (5 µg/ml) for 16 hours in PBMC after the E2CD86 subunit vaccine was injected into pigs on days 10 (a) and 21 (b). PBMC was assessed by flow cytometry. The IFN-γ frequency was the highest at 14 dpv among the subunit vaccine groups. Representative dot plots showing quadrant gates defining the expression of IFN-γ.
4. Discussion
CSF has a serious negative economic impact on the swine industry and is highly questionable from an ethical point of view worldwide [42]. A previous study showed that a commercial E2 subunit vaccine provides protection against different strains of CSFV [43]. The effectiveness and long-term immunity of CSFV vaccines are still issues that need to be addressed. In the present study, we demonstrated the immunogenicity of a novel CSFV subunit vaccine with a molecular adjuvant in vaccinated pigs. The results presented herein support further development into more effectively subunit vaccines.
A previous study revealed that a commercial E2 subunit vaccine provides protection to pigs against lethal CSFV challenge with different strains [44]. To obtain excellent immunogenicity for subunit vaccines, a crucial step is to choose an appropriate adjuvant that can activate the immune regulation pathway, enhance the immune response of subunit vaccines, and improve vaccine efficacy [45]. Several studies have shown that the fusion or coexpression of cytokines with protective antigens is synergistic and that the induction of immunity is greater than that of antigens alone [46]. Effective immunity has always been an issue with subunit vaccines. However, the E2CD154 subunit vaccine was evaluated for inducible immunogenicity and safety [47]. There is rarely advanced discussion on the application of other molecular adjuvants. Our group has developed a CSFV subunit vaccine candidate based on the chimeric recombinant protein E2-CD86 mixed with Montanide™ ISA206. CD86 is preferentially expressed on dendritic cells, Langerhans cells, macrophages, germinal center B cells and memory B cells [48]. The role of the costimulatory molecule CD86 in the differentiation and activation of T cells has also been investigated in studies of virus infections, such as HBV [49], influenza virus [50] and HIV [51].
This study confirms that the subunit vaccine model, E2CD86, has the potential to induce a strong cellular immune response after the first dose of vaccination on day 14. For humoral immunity, we measured E2 antibodies in vaccine pigs. Compared with the E2-only and Bay®-E2 groups, the first dose of the E2CD86 vaccine induced E2-specific antibodies at 14 dpv. The response of the animals after vaccination indicates that 75% of the animals were able to respond effectively to the vaccination or that these vaccinated animals can be protected from future exposure to contact infection. In our study, the CD86 antigen acted as a molecular adjuvant to induce both humoral and cellular immune responses against the E2 viral glycoprotein (Figure 2, Figure 3 and Figure 4).
CD4+ T cells help cells contribute to the induction of tissue-resident memory CD8+ T cells during virus infection by helping CD8+ T cells enter a microenvironment where they can be exposed to signals for their continued maintenance within the tissue. Following vaccination, CD4+ T-cell activation is necessary and important for the efficient induction of memory CD8+ T cells to mediate protective immunity. We used flow cytometric tools to monitor the responses of specific T-cell subsets. PBMC were isolated to examine T-cell expression after vaccination. E2CD86 was associated with the highest percentage of CD4+ T cells among all the groups after vaccination. Additional studies are needed to determine whether nonclassical MHC molecules are involved in the presentation of such epitopes.
The secretion of IL-2 is important and a key marker of the early activation process of CD4+ T cells. IL-2 signaling mediates several biological processes that support immune system function. Compared with the commercial vaccine, the E2CD86 vaccine resulted in a greater frequency of CD4+IL-2+ T cells. IL-2 signaling is required to enhance the ability of antigen-stimulated CD8+ T cells to differentiate and become cytotoxic as "better effector cells", with greater antitumor and antiviral properties in mice [52,53]. CD8 + cytotoxic T lymphocytes can identify and respond to highly conserved antigens via major histocompatibility complex (MHC) class I presentation pathways and are involved in mediating protection against viruses in preclinical models such as influenza [54,55] and CSFV studies [56]. When pigs were vaccinated with E2CD86, the number of CD8+ T cells was obviously greater than that in pigs vaccinated with the commercial E2 vaccine. These data suggested that the E2CD86 vaccine induced not only humoral but also cellular immunity. However, the use of IL-2 as an early marker needs to be confirmed in further experiments.
IFN-γ, a member of the interferon family, is a host-specific cytokine that is produced by mammalian cells infected with viruses and triggers the immune system to activate T cells and macrophages [57], thereby playing an antiviral role. The coexpression of IFN-γ and antigens often synergistically affects the antigen immune response [58,59]. This study was based on a CHO cell system in which the E2 protein of CSFV was coexpressed with CD86. To confirm whether T-cell mediated immunity contributes to vaccine-induced protective immunity in pigs, we also analyzed IFN-γ expression in PBMC samples from vaccinated E2CD86 and commercial E2. To increase the value of E2CD86 as an important subunit vaccine model for measuring immunity, we used the IFN-γ ELISpot to quantity the T-cell response to antigenic determinants. We isolate and process blood sampling to peripheral T cells, which allows clear enumeration of IFN-γ-positive spot-forming units following E2 protein (antigenic peptide) stimulation. The further advantage of blood sampling requires repeated immunizations, which is valuable in evaluating vaccine immunogenicity and duration of T-cell immunity across treatment regimens. We demonstrated that IFN-γ was highly facilitated in vaccinated E2CD86-producing cells via both flow cytometry and ELISpot assays. Otherwise, IFN-γ acts on T cells to promote CD8+ T-cell differentiation into cytotoxic T lymphocytes, which secrete cytokines to clear virus infected cells [60]. This finding is also consistent with our observation of increased CD8+ T cells. This study revealed increased IFN-γ expression after the E2CD86 vaccine.
Moreover, circulating CD4+CD8+ double-positive (DP) T cells are related to multiple diseases. Many studies have shown that the DP T-cell population is associated with several disorders, including cancer [61], autoimmune disorders [62], graft rejection [63], and viral infections [64,65]. Above all, studies have reported that DP T cells are increased in humans with viral infections, such as HIV and COVID-19, suggesting a potential role of DP T-cell populations related to the clearance of viruses [66,67,68]. These DP-T cells are rich in blood or target organs in most cases, suggesting that DP-T cells are involved in the pathological process. Consequently, peripheral DP T cells may be either differentiated resting memory or effector T cells that derived from CD4+ or CD8+ T cells in response to chronic inflammatory diseases [69]. Otherwise, DP T-cell populations may exhibit pleiotropic properties, such as regulatory [70], follicular helper (fh) [71], cytotoxic, innate or adaptive functions [72], depending on their expression of CD4 or CD8 coreceptors. The data presented herein demonstrate that circulating DP T-cell subpopulations are highly expressed after the E2CD86 vaccine until day 28. A study indicated that approximately 54% of peripheral DP T cells exhibit an effector memory T-cell phenotype, whereas approximately 8% present a central memory phenotype [73].
Moreover, DP T cells exhibit more differentiated phenotypes, for instance, CD4dim+CD8bright+ T cells exhibit the phenotype of CCR7-CD45RA- EM, whereas CD4bright+CD8dim+ T cells are mainly CCR7+CD45RA- memory T cells [65]. Circulating DP T cells are effector cells with antiviral activity. CD57 was found expression on DP T cells as well as considers a terminal differentiation marker on single positive T cells. It was suggested that DP T cells could present some characters of terminally differentiated T cells [74]. In addition, DP T cells in HIV-infected patients produce high levels of proinflammatory cytokines such as TNF and IFN-γ [75]. Also, in response to HIV and CMV antigens, it was indicated CD4bright+CD8dim+ cells with specific MHC class I-restricted cytotoxic activity to against tumor cells and showed increased cytokine expression [76]. Graham SP et. al. indicated a population of DP T follicular helper cells that regulate the humoral immunity in chronic inflammatory lesions [77], revealing novel and unknown functions of DP T cells.
Other studies have shown that CD3+ CD4− CD8hi T-cell populations are the first and major foundation of CSFV-specific IFN-γ [78,79], whereas innate immune cells such as NK and gamma-delta (gd)-T cells may not be involved in the development of rapid protection [80].
In this study, we demonstrated that the E2CD86 subunit vaccine facilitated CSFV-specific IFN-γ expression. The percentages of CD4+, CD8+, IL-2+ and DP T cells were increased with the E2CD86 vaccine. The increased IL-2 and IFN-γ cytokine profiles of CD4+ T cells related to different viral infections and antigen loads during viral infections have recently been studied. IL-2 and IFN-γ expressions in PBMC at different levels, highlighting the important role of these cells and their regulated responsiveness to key cytokines in the cross-talk between adaptive and innate immunity. The levels of many proinflammatory cytokines, such as IL-1β, IL-6, IL-12, TNF-α, and other interferons, and the cell-mediated immune response in E2CD86-vaccinated pigs need to be explored. However, these data revealed that the new CD86 adjuvant was effective for pig immunization. Therefore, to achieve better immunogenicity of subunit vaccine, the indispensable step is to select an appropriate adjuvant to enhance cellular immune response, activate the immune regulation pathway and increase the efficacy [81]. The experimental results revealed that E2CD86 is a promising candidate CSFV subunit vaccine.
5. Conclusions
E2CD86 is the first reported subunit CSFV vaccine to induce robust cellular immunity, which is efficacious for protection.
Funding
This work was funded by Higher Education SPROUT Project by the Ministry of Education (MOE) in Taiwan.
Acknowledgments
We would like to thank all colleagues and students who contributed to this study. We are grateful to Li-Wu Cheng, who assisted with bioinformatics. We thank the editors and reviewers for constructive comments.
Conflicts of Interest
“The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results”.
References
- Ganges, L.; Crooke, H.R.; Bohórquez, J.A.; Postel, A.; Sakoda, Y.; Becher, P.; Ruggli, N. Classical swine fever virus: The past, present and future. Virus Res. 2020, 289, 198151. [Google Scholar] [CrossRef] [PubMed]
- Postel, A.; Smith, D.B.; Becher, P. Proposed Update to the Taxonomy of Pestiviruses: Eight Additional Species within the Genus Pestivirus, Family Flaviviridae. Viruses. 2021, 13, 1542. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Blome, S.; Staubach, C.; Henke, J.; Carlson, J.; Beer, M. Classical Swine Fever-An Updated Review. Viruses. 2017, 9, 86. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Blome, S.; Meindl-Böhmer, A.; Nowak, G.; Moennig, V. Disseminated intravascular coagulation does not play a major role in the pathogenesis of classical swine fever. Vet Microbiol. 2013, 162, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Beer, M.; Reimann, I.; Hoffmann, B.; Depner, K. Novel marker vaccines against classical swine fever. Vaccine. 2007, 25, 5665–5670. [Google Scholar] [CrossRef] [PubMed]
- Bouma, A.; de Smit, A.J.; de Kluijver, E.P.; Terpstra, C.; Moormann, R.J. Efficacy and stability of a subunit vaccine based on glycoprotein E2 of classical swine fever virus. Vet Microbiol. 1999, 66, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Jasna, P.R.; Radoslav, D.; Vladimir, P.; Tamaš; P; Igor, S.; Radomir, R.; Miroslav, V. Classical Swine Fever: Active Immunisation of Piglets with Subunit (E2) Vaccine in the Presence of Different Levels of Colostral Immunity (China Strain). Acta Veterinaria, 2014, Sciendo, vol. 64 no. 4, pp. 493–509. [CrossRef]
- Madera, R.; Gong, W.; Wang, L.; Burakova, Y.; Lleellish, K.; Galliher-Beckley, A.; Nietfeld, J.; Henningson, J.; Jia, K.; Li, P.; Bai, J.; Schlup, J.; McVey, S.; Tu, C.; Shi, J. Pigs immunized with a novel E2 subunit vaccine are protected from subgenotype heterologous classical swine fever virus challenge. BMC Vet Res. 2016, 12, 197. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Moormann, R.J.; Bouma, A.; Kramps, J.A.; Terpstra, C.; De Smit, H.J. Development of a classical swine fever subunit marker vaccine and companion diagnostic test. Vet Microbiol. 2000, 73, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Sordo-Puga, Y.; Suárez-Pedroso, M.; Naranjo-Valdéz, P.; Pérez-Pérez, D.; Santana-Rodríguez, E.; Sardinas-Gonzalez, T.; Mendez-Orta, M.K.; Duarte-Cano, C.A.; Estrada-Garcia, M.P.; Rodríguez-Moltó, M.P. Porvac® Subunit Vaccine E2-CD154 Induces Remarkable Rapid Protection against Classical Swine Fever Virus. Vaccines (Basel). 2021, 9, 167. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Madera, R.; Gong, W.; Wang, L.; Burakova, Y.; Lleellish, K.; Galliher-Beckley, A.; Nietfeld, J.; Henningson, J.; Jia, K.; Li, P.; Bai, J.; Schlup, J.; McVey, S.; Tu, C.; Shi, J. Pigs immunized with a novel E2 subunit vaccine are protected from subgenotype heterologous classical swine fever virus challenge. BMC Vet Res. 2016, 12, 197. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- van Aarle, P. Suitability of an E2 subunit vaccine of classical swine fever in combination with the E(rns)-marker-test for eradication through vaccination. Dev Biol (Basel) 2003, 114, 193–200. [Google Scholar]
- Suárez, M.; Sordo, Y.; Prieto, Y.; Rodríguez, M.P.; Méndez, L.; Rodríguez, E.M.; Rodríguez-Mallon, A.; Lorenzo, E.; Santana, E.; González, N.; Naranjo, P.; Frías, M.T.; Carpio, Y.; Estrada, M.P. A single dose of the novel chimeric subunit vaccine E2-CD154 confers early full protection against classical swine fever virus. Vaccine. 2017, 35, 4437–4443. [Google Scholar] [CrossRef] [PubMed]
- Armengol, E.; Wiesmüller, K.H.; Wienhold, D.; Büttner, M.; Pfaff, E.; Jung, G.; Saalmüller, A. Identification of T-cell epitopes in the structural and non-structural proteins of classical swine fever virus. J Gen Virol. 2002, 83(Pt 3):551-560. [CrossRef] [PubMed]
- Ceppi, M.; de Bruin, M.G.M.; Seuberlich, T.; Balmelli, C.; Pascolo, S.; Ruggli, N.; Wienhold, D.; Tratschin, J.D.; McCullough, K.C.; Summerfield, A. Identification of classical swine fever virus protein E2 as a target for cytotoxic T cells by using mRNA-transfected antigen-presenting cells. J Gen Virol. 2005, 86(Pt 9):2525-2534. [CrossRef] [PubMed]
- Rau, H.; Revets, H.; Balmelli, C.; McCullough, K.C.; Summerfield, A. Immunological properties of recombinant classical swine fever virus NS3 protein in vitro and in vivo. Vet Res. 2006, 37, 155–168. [Google Scholar] [CrossRef] [PubMed]
- Toledo, J.R.; Barrera, M.; Farnós, O.; Gómez, S.; Rodríguez, M.P.; Aguero, F.; Ormazabal, V.; Parra, N.C.; Suárez, L.; Sánchez, O. Human αIFN co-formulated with milk derived E2-CSFV protein induce early full protection in vaccinated pigs. Vaccine. 2010, 28, 7907–7914. [Google Scholar] [CrossRef] [PubMed]
- Lanier, L.L.; O'Fallon, S.; Somoza, C.; Phillips, J.H.; Linsley, P.S.; Okumura, K.; Ito, D.; Azuma MCD80, (.B.7.).; CD86 (B70) provide similar costimulatory signals for T cell proliferation cytokine production generation of, C.T.L. J Immunol. 1995, 154, 97–105. [PubMed]
- Linsley, P.S.; Clark, E.A.; Ledbetter, J.A. T-cell antigen CD28 mediates adhesion with B cells by interacting with activation antigen B7/BB-1. Proc Natl Acad Sci U S A. 1990, 87, 5031–5035. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Chen, C.; Nabavi, N. In vitro induction of T cell anergy by blocking B7 and early T cell costimulatory molecule ETC-1/B7-2. Immunity 1994, 1, 147–154. [Google Scholar] [CrossRef]
- Lenschow DJ, Ho SC, Sattar H, Rhee L, Gray G, Nabavi N; et al. Differential effects of anti-B7-1 and anti-B7-2 monoclonal antibody treatment on the development of diabetes in the nonobese diabetic mouse. J Exp Med 1995, 181, 1145–1155. [CrossRef]
- Bour-Jordan, H.; Blueston, J.A. CD28 function: A balance of costimulatory and regulatory signals. J Clin Immunol. 2002, 22, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Beyersdorf, N.; Kerkau, T.; Hünig, T. CD28 co-stimulation in T-cell homeostasis: A recent perspective. Immunotargets Ther. 2015, 4, 111–122. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Carosella, E.D.; Ploussard, G.; LeMaoult, J.; Desgrandchamps, F. A Systematic Review of Immunotherapy in Urologic Cancer: Evolving Roles for Targeting of CTLA-4, PD-1/PD-L1, and HLA-G. Eur Urol. 2015, 68, 267–279. [Google Scholar] [CrossRef] [PubMed]
- Gu, P.; Gao, J.F.; D'Souza, C.A.; Kowalczyk, A.; Chou, K.Y.; Zhang, L. Trogocytosis of CD80 and CD86 by induced regulatory T cells. Cell Mol Immunol. 2012, 9, 136–146. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Miller, J.; Baker, C.; Cook, K.; Graf, B.; Sanchez-Lockhart, M.; Sharp, K.; Wang, X.; Yang, B.; Yoshida, T. Two pathways of costimulation through CD28. Immunol Res. 2009, 45, 159–172. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Freeman, G.J.; Freedman, A.S.; Segil, J.M.; Lee, G.; Whitman, J.F.; Nadler, L.M. B7, a new member of the Ig superfamily with unique expression on activated and neoplastic B cells. J Immunol. 1989, 143, 2714–2722. [Google Scholar] [CrossRef] [PubMed]
- Azuma, M.; Ito, D.; Yagita, H.; Okumura, K.; Phillips, J.H.; Lanier, L.L.; Somoza, C. B70 antigen is a second ligand for CTLA-4 and CD28. Nature. 1993, 366, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Freedman, A.S.; Freeman, G.J.; Rhynhart, K.; Nadler, L.M. Selective induction of B7/BB-1 on interferon-gamma stimulated monocytes: A potential mechanism for amplification of T cell activation through the CD28 pathway. Cell Immunol 1991, 137, 429–437. [Google Scholar] [CrossRef]
- Hathcock, K.S.; Laszlo, G.; Pucillo, C.; Linsley, P.; Hodes, R.J. Comparative analysis of B7-1 and B7-2 costimulatory ligands: Expression and function. J Exp Med 1994, 180, 631–640. [Google Scholar] [CrossRef]
- Stack, R.M.; Lenschow, D.J.; Gray, G.S.; Bluestone, J.A.; Fitch, F.W. IL-4 treatment of small splenic B cells induces costimulatory molecules B7-1 and B7-2. J Immunol 1994, 152, 5723–5733. [Google Scholar] [CrossRef]
- Kim, J.J.; Bagarazzi, M.L.; Trivedi, N.; Hu, Y.; Kazahaya, K.; Wilson, D.M.; Ciccarelli, R.; Chattergoon, M.A.; Dang, K.; Mahalingam, S.; Chalian, A.A.; Agadjanyan, M.G.; Boyer, J.D.; Wang, B.; Weiner, D.B. Engineering of in vivo immune responses to DNA immunization via codelivery of costimulatory molecule genes. Nat Biotechnol. 1997, 15, 641–646. [Google Scholar] [CrossRef] [PubMed]
- Yüksel, S.; Pekcan, M.; Puralı, N.; Esendağlı, G.; Tavukçuoğlu, E.; Rivero-Arredondo, V.; Ontiveros-Padilla, L.; López-Macías, C.; Şenel, S. Development and in vitro evaluation of a new adjuvant system containing Salmonella Typhi porins and chitosan. Int J Pharm. 2020, 578, 119129. [Google Scholar] [CrossRef] [PubMed]
- Pan, C.H.; Jong, M.H.; Huang, T.S.; Liu, H.F.; Lin, S.Y.; Lai, S.S. Phylogenetic analysis of classical swine fever virus in Taiwan. Arch Virol. 2005, 150, 1101–1119. [Google Scholar] [CrossRef] [PubMed]
- Wu, Cheng & Huang, Qiong-Yi & Wang, Pei-Chi & Tsai, Ming-An & Chen, shih-chu. Transcriptome and pathophysiological analysis during Bacillus cereus group infection in Pelodiscus sinensis uncovered the importance of iron and toll like receptor pathway. Aquaculture. 2024, 594, 741424. [CrossRef]
- Yanjuan Jia, Hui Xu, Yonghong Li, Chaojun Wei, Rui Guo, Fang Wang, Yu Wu, Jing Liu, Jing Jia, Junwen Yan, Xiaoming Qi, Yuanting Li, Xiaoling Gao. A Modified Ficoll-Paque Gradient Method for Isolating Mononuclear Cells from the Peripheral and Umbilical Cord Blood of Humans for Biobanks and Clinical Laboratories. Biopreserv Biobank. 2018, 16, 82–91. [CrossRef] [PubMed]
- Tai, Y.; Wang, Q.; Korner, H.; Zhang, L.; Wei, W. Molecular Mechanisms of T Cells Activation by Dendritic Cells in Autoimmune Diseases. Front Pharmacol. 2018, 9, 642. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Laidlaw, B.J.; Craft, J.E.; Kaech, S.M. The multifaceted role of CD4(+) T cells in CD8(+) T cell memory. Nat Rev Immunol. 2016, 16, 102–111. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Overgaard, N.H.; Jung, J.W.; Steptoe, R.J.; Wells, J.W. CD4+/CD8+ double-positive T cells: More than just a developmental stage? J Leukoc Biol. 2015, 97, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.M.; Park, H.J.; Choi, E.A.; Jung, K.C.; Lee, J.I. Cellular heterogeneity of circulating CD4+CD8+ double-positive T cells characterized by single-cell RNA sequencing. Sci Rep. 2021, 11, 23607. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Van Kaer, L.; Rabacal, W.A.; Scott Algood, H.M.; Parekh, V.V.; Olivares-Villagómez, D. In vitro induction of regulatory CD4+CD8α+ T cells by TGF-β, IL-7 and IFN-γ. PLoS ONE. 2013, 8, e67821. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Saatkamp, H.W.; Berentsen, P.B.; Horst, H.S. Economic aspects of the control of classical swine fever outbreaks in the European Union. Vet Microbiol. 2000, 73, 221–237. [Google Scholar] [CrossRef] [PubMed]
- Dahle, J.; Liess, B. Assessment of safety and protective value of a cell culture modified strain "C" vaccine of hog cholera/classical swine fever virus. Berl Munch Tierarztl Wochenschr. 1995, 108, 20–25. [Google Scholar] [PubMed]
- Gong, W.; Li, J.; Wang, Z.; Sun, J.; Mi, S.; Xu, J.; Cao, J.; Hou, Y.; Wang, D.; Huo, X.; Sun, Y.; Wang, P.; Yuan, K.; Gao, Y.; Zhou, X.; He, S.; Tu, C. Commercial E2 subunit vaccine provides full protection to pigs against lethal challenge with 4 strains of classical swine fever virus genotype 2. Vet Microbiol. 2019, 237, 108403. [Google Scholar] [CrossRef] [PubMed]
- Perrie, Y.; Mohammed, A.R.; Kirby, D.J.; McNeil, S.E.; Bramwell, V.W. Vaccine adjuvant systems: Enhancing the efficacy of sub-unit protein antigens. Int J Pharm. 2008, 364, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Genmei, L.; Manlin, L.; Ruiai, C.; Hongliang, H.; Dangshuai, P. Construction and immunogenicity of recombinant adenovirus expressing ORF2 of PCV2 and porcine IFN gamma. Vaccine. 2011, 29, 8677–8682. [Google Scholar] [CrossRef] [PubMed]
- Suárez-Pedroso, M.; Sordo-Puga, Y.; Sosa-Teste, I.; Rodriguez-Molto, M.P.; Naranjo-Valdés, P.; Sardina-González, T.; Santana-Rodríguez, E.; Montero-Espinosa, C.; Frías-Laporeaux, M.T.; Fuentes-Rodríguez, Y.; Pérez-Pérez, D.; Oliva-Cárdenas, A.; Pereda, C.L.; González-Fernández, N.; Bover-Fuentes, E.; Vargas-Hernández, M.; Duarte, C.A.; Estrada-García, M.P. Novel chimeric E2CD154 subunit vaccine is safe and confers long lasting protection against classical swine fever virus. Vet Immunol Immunopathol. 2021, 234, 110222. [Google Scholar] [CrossRef] [PubMed]
- Lenschow, D.J.; Su, G.H.; Zuckerman, L.A.; Nabavi, N.; Jellis, C.L.; Gray, G.S.; Miller, J.; Bluestone, J.A. Expression and functional significance of an additional ligand for CTLA-4. Proc Natl Acad Sci U S A. 1993, 90, 11054–11058. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Said, E.A.; Al-Reesi, I.; Al-Riyami, M.; Al-Naamani, K.; Al-Sinawi, S.; Al-Balushi, M.S.; Koh, C.Y.; Al-Busaidi, J.Z.; Idris, M.A.; Al-Jabri, A.A. Increased CD86 but Not CD80 and PD-L1 Expression on Liver CD68+ Cells during Chronic HBV Infection. PLoS ONE. 2016, 11, e0158265. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Sun, R.; Yang, Y.; Gu, Z.; Tang, X.; Zhang, C.; Kou, W.; Wei, P. Silencing of CD86 in dendritic cells by small interfering RNA regulates cytokine production in T cells from patients with allergic rhinitis in vitro. Mol Med Rep. 2019, 20, 3893–3900. [Google Scholar] [CrossRef] [PubMed]
- Köchli, C.; Wendland, T.; Frutig, K.; Grunow, R.; Merlin, S.; Pichler, W.J. CD80 and CD86 costimulatory molecules on circulating T cells of HIV infected individuals. Immunol Lett. 1999, 65, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Niederlova, V.; Tsyklauri, O.; Kovar, M.; Stepanek, O. IL-2-driven CD8+ T cell phenotypes: Implications for immunotherapy. Trends Immunol. 2023, 44, 890–901. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Spolski, R.; Li, P.; Leonard, W.J. Biology and regulation of IL-2: From molecular mechanisms to human therapy. Nat Rev Immunol. 2018, 18, 648–659. [Google Scholar] [CrossRef] [PubMed]
- Epstein, S.L.; Lo, C.Y.; Misplon, J.A.; Bennink, J.R. Mechanism of protective immunity against influenza virus infection in mice without antibodies. J Immunol. 1998, 160, 322–327. [Google Scholar] [CrossRef] [PubMed]
- Yap, K.L.; Ada, G.L.; McKenzie, I.F. Transfer of specific cytotoxic T lymphocytes protects mice inoculated with influenza virus. Nature. 1978, 273, 238–239. [Google Scholar] [CrossRef] [PubMed]
- Franzoni, G.; Kurkure, N.V.; Essler, S.E.; Pedrera, M.; Everett, H.E.; Bodman-Smith, K.B.; Crooke, H.R.; Graham, S.P. Proteome-wide screening reveals immunodominance in the CD8 T cell response against classical swine fever virus with antigen-specificity dependent on MHC class I haplotype expression. PLoS ONE. 2013, 8, e84246. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Moraes, M.P.; de Los Santos, T.; Koster, M.; Turecek, T.; Wang, H.; Andreyev, V.G.; Grubman, M.J. Enhanced antiviral activity against foot-and-mouth disease virus by a combination of type I and II porcine interferons. J Virol. 2007, 81, 7124–7135. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wang, Y.P.; Liu, D.; Guo, L.J.; Tang, Q.H.; Wei, Y.W.; Wu, H.L.; Liu, J.B.; Li, S.B.; Huang, L.P.; Liu, C.M. Enhanced protective immune response to PCV2 subunit vaccine by co-administration of recombinant porcine IFN-γ in mice. Vaccine. 2013, 31, 833–838. [Google Scholar] [CrossRef] [PubMed]
- Eichinger, K.M.; Resetar, E.; Orend, J.; Anderson, K.; Empey, K.M. Age predicts cytokine kinetics and innate immune cell activation following intranasal delivery of IFNγ and GM-CSF in a mouse model of RSV infection. Cytokine. 2017, 97, 25–37. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Whitmire, J.K.; Tan, J.T.; Whitton, J.L. Interferon-gamma acts directly on CD8+ T cells to increase their abundance during virus infection. J Exp Med. 2005, 201, 1053–1059. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Desfrançois, J.; Moreau-Aubry, A.; Vignard, V.; Godet, Y.; Khammari, A.; Dréno, B.; Jotereau, F.; Gervois, N. Double positive CD4CD8 alphabeta T cells: A new tumor-reactive population in human melanomas. PLoS ONE. 2010, 5, e8437. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Parel, Y.; Chizzolini, C. CD4+ CD8+ double positive (DP) T cells in health and disease. Autoimmun Rev. 2004, 3, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Park, H.J.; Park, H.J.; Jung, K.C.; Lee, J.I. CD4hiCD8low Double-Positive T Cells Are Associated with Graft Rejection in a Nonhuman Primate Model of Islet Transplantation. J Immunol Res. 2018, 2018, 3861079. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Nascimbeni, M.; Pol, S.; Saunier, B. Distinct CD4+ CD8+ double-positive T cells in the blood and liver of patients during chronic hepatitis B and C. PLoS ONE. 2011, 6, e20145. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Nascimbeni, M.; Shin, E.C.; Chiriboga, L.; Kleiner, D.E.; Rehermann, B. Peripheral CD4(+)CD8(+) T cells are differentiated effector memory cells with antiviral functions. Blood. 2004, 104, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Ribrag, V.; Salmon, D.; Picard, F.; Guesnu, M.; Sicard, D.; Dreyfus, F. Increase in double-positive CD4+CD8+ peripheral T-cell subsets in an HIV-infected patient. AIDS. 1993, 7, 1530. [Google Scholar] [CrossRef] [PubMed]
- Zahran, A.M.; Zahran, Z.A.M.; Mady, Y.H.; Mahran, E.E.M.O.; Rashad, A.; Makboul, A.; Nasif, K.A.; Abdelmaksoud, A.A.; El-Badawy, O. Differential alterations in peripheral lymphocyte subsets in COVID-19 patients: Upregulation of double-positive and double-negative T cells. Multidiscip Respir Med. 2021, 16, 758. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zou, S.; Tan, Y.; Xiang, Y.; Liu, Y.; Zhu, Q.; Wu, S.; Guo, W.; Luo, M.; Shen, L.; Liang, K. The Role of CD4+CD8+ T Cells in HIV Infection With Tuberculosis. Front Public Health. 2022, 10, 895179. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Akari, H.; Terao, K.; Murayama, Y.; Nam, K.H.; Yoshikawa, Y. Peripheral blood CD4+CD8+ lymphocytes in cynomolgus monkeys are of resting memory T lineage. Int Immunol. 1997, 9, 591–597. [Google Scholar] [CrossRef] [PubMed]
- Van Kaer, L.; Rabacal, W.A.; Scott Algood, H.M.; Parekh, V.V.; Olivares-Villagómez, D. In vitro induction of regulatory CD4+CD8α+ T cells by TGF-β, IL-7 and IFN-γ. PLoS ONE. 2013, 8, e67821. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Quandt, D.; Rothe, K.; Scholz, R.; Baerwald, C.W.; Wagner, U. Peripheral CD4CD8 double positive T cells with a distinct helper cytokine profile are increased in rheumatoid arthritis. PLoS ONE. 2014, 9, e93293. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kerstein, A.; Müller, A.; Pitann, S.; Riemekasten, G.; Lamprecht, P. Circulating CD4+CD8+ double-positive T-cells display features of innate and adaptive immune function in granulomatosis with polyangiitis. Clin Exp Rheumatol. 2018, 36 (Suppl. S111), 93–98. [Google Scholar] [PubMed]
- Waschbisch, A.; Sammet, L.; Schröder, S.; Lee, D.H.; Barrantes-Freer, A.; Stadelmann, C.; Linker, R.A. Analysis of CD4+ CD8+ double-positive T cells in blood, cerebrospinal fluid and multiple sclerosis lesions. Clin Exp Immunol. 2014, 177, 404–411. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Chauhan, N.K.; Vajpayee, M.; Mojumdar, K.; Singh, R.; Singh, A. Study of CD4+CD8+ double positive T-lymphocyte phenotype and function in Indian patients infected with HIV-1. J Med Virol. 2012, 84, 845–856. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Hu, S.; Fu, X.; Li, L. CD4+ Cytotoxic T Lymphocytes in Cancer Immunity and Immunotherapy. Adv Biol (Weinh). 2023, 7, e2200169. [Google Scholar] [CrossRef] [PubMed]
- Murayama, K.; Ikegami, I.; Kamekura, R.; Sakamoto, H.; Yanagi, M.; Kamiya, S.; Sato, T.; Sato, A.; Shigehara, K.; Yamamoto, M.; Takahashi, H.; Takano, K.I.; Ichimiya, S. CD4+CD8+ T follicular helper cells regulate humoral immunity in chronic inflammatory lesions. Front Immunol. 2022, 13, 941385. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Graham, S.P.; Haines, F.J.; Johns, H.L.; Sosan, O.; La Rocca, S.A.; Lamp, B.; Rümenapf, T.; Everett, H.E.; Crooke, H.R. Characterisation of vaccine-induced, broadly cross-reactive IFN-γ secreting T cell responses that correlate with rapid protection against classical swine fever virus. Vaccine. 2012, 30, 2742–2748. [Google Scholar] [CrossRef] [PubMed]
- Franzoni, G.; Kurkure, N.V.; Edgar, D.S.; Everett, H.E.; Gerner, W.; Bodman-Smith, K.B.; Crooke, H.R.; Graham, S.P. Assessment of the phenotype and functionality of porcine CD8 T cell responses following vaccination with live attenuated classical swine fever virus (CSFV) and virulent CSFV challenge. Clin Vaccine Immunol. 2013, 20, 1604–1616. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Franzoni, G.; Edwards, J.C.; Kurkure, N.V.; Edgar, D.S.; Sanchez-Cordon, P.J.; Haines, F.J.; Salguero, F.J.; Everett, H.E.; Bodman-Smith, K.B.; Crooke, H.R.; Graham, S.P. Partial Activation of natural killer and γδ T cells by classical swine fever viruses is associated with type I interferon elicited from plasmacytoid dendritic cells. Clin Vaccine Immunol. 2014, 21, 1410–1420. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Perrie, Y.; Mohammed, A.R.; Kirby, D.J.; McNeil, S.E.; Bramwell, V.W. Vaccine adjuvant systems: Enhancing the efficacy of sub-unit protein antigens. Int J Pharm. 2008, 364, 272–280. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



