Submitted:
16 January 2025
Posted:
16 January 2025
You are already at the latest version
Abstract
Allergens from pollen, mites, and moulds often sensitize patients simultaneously, posing challenges for developing stable and effective combination vaccines. Alternaria alternata, a major source of indoor and outdoor allergens, is strongly linked to asthma development and contains proteolytic enzymes that can degrade other allergens, potentially reducing vaccine efficacy. This study aimed to evaluate the safety, efficacy, and stability of polymerized A. alternata extracts (allergoids) compared to native extracts and their compatibility with pollen extracts (Phleum pratense). Allergoids were prepared using glutaraldehyde and characterized through SDS-PAGE, LC-MS/MS, NMR, and gas chromatography. Their immunogenicity and IgE-binding properties were assessed via Western blot and ELISA competition assays, while enzymatic activity was analysed using ApiZym kits. Mice immunization experiments were conducted to evaluate antibody responses. Polymerized extracts exhibited reduced IgE-binding capacity while maintaining IgG-binding and immunogenicity. Mice immunized with allergoids generated antibodies that efficiently blocked IgE binding in allergic patients. Proteolytic activity was significantly reduced in allergoids, and pollen extracts remained stable when combined with them. These findings demonstrate that A. alternata allergoids are a safer, more stable alternative for immunotherapy, offering enhanced efficacy and reduced injections for polysensitized patients. This study provides critical insights for designing optimized combination vaccines.
Keywords:
alternaria alternata
; fungi allergy
; allergen specific immunotherapy
; allergen extract
; allergoid
1. Introduction
Fungi are the fourth most frequent cause of allergic respiratory diseases and are particularly associated with poor asthma prognosis [1,2]. Allergic immunotherapy (AIT) reduces the risk of developing asthma in children with allergenic rhinitis due to pollen and mites [3,4,5]. However, despite the long-standing use of AIT with mould extracts, evidence supporting its efficacy in treating respiratory allergic diseases remains limited [6,7]. Furthermore, immunotherapy for mould allergies is not completely exempt of risks. The administration of specific native fungal protein extracts carries a high risk of adverse reactions [8,9,10]. Consequently, several allergy and asthma organizations did not recommend AIT for fungi in children due to insufficient evidence regarding the efficacy and safety of native fungal extracts [11,12].
One of the primary goals of AIT is to increase safety by minimising secondary adverse events while preserving efficacy [13,14]. The production of allergoids, or polymerised allergenic extracts with glutaraldehyde, supposes a valid alternative to AIT with unmodified extracts. Due to their reduced allergenicity, while retaining immunogenicity, polymerised extracts (PE) are widely used in the treatment of pollen and mite allergies, offering an efficient and safer alternative to conventional native unmodified extracts.
The chemical cross-linking of allergens during polymerisation produces larger molecules, which reduces accessibility to or recognition of IgE epitopes (decreasing allergenicity) while preserving IgG and linear T-cell epitopes (ensuring immunogenicity). This property of allergoids allows for faster up-dosing during immunotherapy without increasing the risk of adverse reactions [15]. Numerous allergens presenting proteolytic activity in different allergenic sources, mainly fungi, mites and cockroaches, which are particularly rich in proteases have been described [16]. Over 50% of allergens with proteolytic activity accepted by the WHO/IUIS are derived from moulds, with serine proteases being the most prominent group [17]. The allergenicity of these enzymatic proteins is thought to be linked to the development of IgE-mediated immune responses (Th2), possibly as an evolved defence mechanism against parasitic infections that depend on enzymes for tissue invasion and migration [18,19]. Mould allergens, in particular, can exacerbate asthma due to their enzymatic activity. This activity disrupts protein junctions within epithelial cells, facilitating the entry of other allergens across epithelial barriers [20,21,22,23,24,25,26]. Furthermore, the enzymatic properties of these proteins can alter the allergenicity and immunogenicity of other proteins by modifying their structure and carbohydrate content [9].
PEs have demonstrated their safety and efficacy in various studies involving pollens, mites, and animal epithelium allergens, benefiting both adult and paediatric patients [27,28,29,30,31,32,33,34]. The European Survey on Adverse Systemic Reactions in Allergen Immunotherapy (EASSI) reported a lower incidence and risk of systemic reactions during subcutaneous immunotherapy (SCIT) with allergoids compared to native extracts [35]. While allergoids have been developed for several allergen sources, their application to fungal allergens has been limited until recently [36,37].
The aim of this study was to develop and characterise polymerised allergenic extracts from the mould Alternaria alternata, evaluating their safety and efficacy. Additionally, we assessed the stability of grass pollen (Phleum pratense) in a mould-grass allergoid mixture compared to its combination with native unmodified A. alternata extracts. These findings provide valuable insights into the feasibility of integrating mould allergen extracts into AIT formulations alongside other allergen sources, without compromising their immunogenic properties.
2. Materials and Methods
2.1. A. alternata Allergen Extraction and Polymerization
Metabolic and somatic phases from A. alternata cultures (Inmunotek Laboratories, Madrid, Spain) were obtained and extracted in 1/20 w/v 0.01 M PBS under magnetic continuous stirring, for 40 h at 4ºC. The resulting supernatant was filtered through 0.2 µm membranes (Sartorius Stedim Biotech S. A.) and diafiltrated by tangential ultrafiltration using a 5 kDa polyether sulfone Omega membrane (Pall Life Sciences), to remove low molecular weight impurities. The resulting purified extracts were freeze-dried (native extracts, NE) and subsequently reconstituted in PBS and polymerized with glutaraldehyde (25% Sigma-Aldrich). The reactions were performed during 18h at 4 °C in continuous stirring and stopped by the addition of 1.25 M glycine. The resulting allergoids (polymerised extract, PE) were diafiltered through a 100 kDa membrane, to remove non-polymerised molecules. The allergoids were lyophilized and kept at 4ºC until use. The protein content of the native extracts and allergoids was measured by the Bradford protein assay (Bio-Rad Laboratories, Madrid, Spain).
2.2. Analysis of the Protein and Allergenic Profile of the Extracts: SDS-PAGE and Immunoblotting
The native and polymerised extracts, (NE and PE, respectively) were separated by SDS-PAGE under reducing conditions using a Mini-PROTEAN II device (Bio-Rad, Hercules, Calif., USA). Ten micrograms of sample were loaded per lane and separated in a custom polyacrylamide gel (Any kD Mini-PROTEAN TGX Stain-Free, Bio-Rad) at 300 volts. The gels were stained with GelCode Blue Stain Reagent (ThermoFisher Scientific, Madrid, Spain). Precision Plus Protein All Blue Standards (Bio-Rad) was used as molecular weight marker. For immunoblotting, proteins were electrotransferred to nitrocellulose membranes using a Trans-Blot Semi-Dry Electrophoresis Transfer Cell at 20 v for 30 minutes (BioRad). After blocking with 5% bovine serum albumin in PBS-0.25% Tween 20 (PBS-T) the membranes were incubated with a commercial 1:4 diluted serum pool prepared with the sera from patients sensitized to Alternaria alternata (Plasmalab International, WA, USA), for 16 h at 4° C. In IgE inhibition western blot assays, this incubation step with sera from sensitised patients was preceded by the incubation with 1:10,000 diluted sera from the mice immunised for 4 h. Afterwards, the membranes were incubated with 1:2,000 diluted horseradish peroxidase (HRP)-conjugated mouse anti-human IgE Fc (Southern Biotech, Birmingham, Ala., USA) for 1 h at 4° C. After washing with PBS-T, the membranes were visualised using Amersham ECL Prime Western Blotting Detection Reagent (Amersham, GE Healthcare, Little Chalfont, UK), in a GeneGnome device (Syngene, Cambridge, UK). Precision Plus Protein WesternC Standards (Bio-Rad) in the presence of Precision Protein StrepTactinHRP Conjugate (Bio-Rad) was used as molecular weight markers.
2.3. Physicochemical Analysis and Chemical Composition of the Extracts: Analysis of Amino Acid Content, Nuclear Magnetic Resonance and Mass Spectrometry
The amino acid composition of the native and polymerised extracts was analysed by Gas chromatography and compared to evaluate the presence of free amino acids in the NE and their implication in the polymerisation process. NE and PE were first hydrolysed with 6N HCl under vacuum at 110 °C for 24 h. Afterwards, the amino acid content was assessed in a quantitative amino acid analyser (Biochrom 30; Biochrom Ltd., Cambridge, UK). Norleucine was used as internal standard.
Nuclear Magnetic Resonance (NMR) was conducted to evaluate the molecular size of the NE and PE. The spectra were obtained for native extracts and allergoid samples in D2O at 298 K in a Bruker Avance 600 MHz spectrometer (Bruker Ltd., Germany). The two-dimension diffusion ordered spectroscopy (2D-DOSY) experiments were developed by recording 64-128 scans for each gradient step, a linear gradient of 16 steps between 2 % and 95 %, a diffusion time (D) between 0.2 and 0.4 s, and length of the diffusion encoding gradient pulses (δ) between 2-4 ms. The spectra were processed with the protocols implemented in TopSpin 3.0 software (Bruker Ltd.).
The protein content in the native and polymerised extracts was determined by liquid chromatography–mass spectrometry (LC-MS/MS). Briefly, samples were first precipitated with methanol/chloroform and submitted to a reduction-alkylation process (50 mM TCEP and 200 mM MMTS) before a digestion step with trypsin (1:20 enzyme/protein). The digested samples were then purified in a SEP-PAK C18 column and peptides were separated, depending on their polarity, by Liquid Chromatography (90 minutes, reverse phase column C-18). The eluted peptides obtained were fragmented in a mass spectrometer (5600 TRIPLE-TOF). NCBI database search was performed with the data obtained, through the MASCOT search engine. The sequences of the proteins identified with at least two peptides and presenting a protein score superior to 20 were compared with the WHO/IUIS protein database.
2.4. Analysis of the Allergenic Activity: IgE/IgG ELISA Competition Assays
Biological IgE and IgG potency of the A. alternata native extracts and their corresponding allergoids (50% inhibition) were determined by ELISA competition assays. Microplates were coated o/n with 1 μg of native extract (NE) or polymerised extract (PE) in 0.05 M carbonate/bicarbonate buffer (pH 9.6). 50 μl of serially diluted competitors and 50 μl of human sera pool from patients sensitized to A. alternata (Plasmalab International, WA, USA) were added to the wells and incubated o/n at 4º C (1:4 diluted for IgE and 1:1,024 for IgG binding detection). After washing, microplates were incubated with secondary antibody (1:2,000 diluted HRP-conjugated mouse anti-human IgE Fc; Southern Biotech, or 1:5,000 diluted HRP-conjugated rabbit anti-human IgG Fc) for 1h. Finally, microplates were washed and incubated with o-phenylenediamine dihydrochloride (OPD, Sigma-Aldrich) for 30 minutes. Reaction was stopped with 1/10 HCl and absorbance was read at 492 nm. All samples were assayed in duplicate. Blank controls were included in the assays and Non-inhibition controls (sera pool without competitor) were included. Percent inhibition were calculated using the following equation: percent inhibition = 100 – [(OD of serum with competitor/OD of serum without competitor) × 100].
2.5. Analysis of the Immunogenicity: Mice Immunization
Eighteen BALB/c mice were divided into 3 groups (6 mice each) and were immunized: 1) Control group: immunized with SPB, 2) NE group: immunized with 20 µg of NE total protein, 3) PE group: immunized with 20 µg of PE total protein. In total, the animals received four subcutaneous injections every two weeks. Seven days after the last immunization, blood was collected, and mice were sacrificed. To determine specific IgG production induced by immunization with NE or PE, total specific IgG levels were analysed in the sera of the three groups of mice. ELISA plates were coated with NE, or PE overnight. Afterwards, sera were added and let to stand overnight. For detection of total IgG, sera were diluted 1:10,000 and incubated o/n. Bound antibodies were detected with 1:2,000 diluted HRP-conjugated goat anti-mouse IgG (Bethyl Laboratories, Texas, USA). OPD was added and the reaction was measured at 492 nm.
To analyse the IgE blocking capacity of the IgG antibodies, Western blot inhibition assays were developed. The assays consisted in the incubation of membranes with 1:10,000 diluted mice sera (immunized with NE or PE), to allow the union of IgG and consecutively, the incubation with the commercial (human) sera pool, from patients sensitized to A. alternata. After washing with SPBT, the membranes were incubated for 2 h with 1:2,000 diluted HRP-conjugated mouse anti-human IgE Fc (SouthernBiotech) and developed as previously described.
2.6. Analysis of the Enzymatic Activity of Allergen Native Extracts and Allergoids
The enzymatic activity of the NE was analysed by qualitative and quantitative methods. To analyse the qualitative enzymatic activity, nineteen enzymatic activities were analysed in the NE, using ApiZym galleries (Biomérieux, Marcy-l’Étoile, France).
The proteolytic activity of the NE and PE was assessed by analysing the hydrolytic capacity of different types of enzymes: trypsin, serine protease, cysteine protease and acid and alkaline phosphatases. Specific substrates were used for each enzyme: Azocoll, S-2288 peptide, L-1460 peptide and NPHP peptide. The absorbances of the coloured products obtained by the hydrolysis of these substrates were measured in a spectrophotometer and the values were extrapolated in a trypsin, papain and acid or alkaline phosphatase standard curve, as corresponding. The PEs were also analysed and compared to the native extract results.
To evaluate the effect of the proteolytic activity of the Alternaria extracts over grass allergen extracts, different mixtures of Alternaria and Phleum pratense (Inmunotek) were prepared and studied after incubation at different intervals of time.
2.7. Stability of Grass Extracts in Mixture with Alternaria Allergoids
Aqueous mixtures of P. pratense native extracts with native or polymerised A. alternata extracts were prepared. Three different types of extracts were prepared: 1) 500 μg/ml of the grass native extract dissolved in deionized water (used as controls); 2) 500 μg/ml of the grass native extract mixed with 25 μg/ml of the Alternaria NE; 3) 500 μg/ml of the grass native extract with 25 μg/ml of the mould PE. The grass extracts were kept at 4ºC, and their stability was evaluated, as described before, by protein quantification (Bradford method), protein profile (SDS-PAGE), Western blot and ELISA competition, after 1 hour, 1 week, 1 month and 3 months of incubation. The immunodetection assays were developed using a commercial pool of sera (Plasmalab International, WA, USA) from patients containing specific IgE antibodies against the grass allergens, as corresponding.
3. Results
3.1. Protein and Allergenic Profile of the Extracts: SDS-PAGE and Immunoblotting
The protein content of the native extract (NE) displayed a broad range of protein sizes, with molecular weights ranging from <10 kDa to >100 kDa, as revealed by SDS-PAGE analysis (Figure 1A, left). At least six distinct protein bands showed IgE reactivity when probed with sera from allergic patients, as demonstrated by Western blotting (Figure 1A, right). Following polymerisation with 25% glutaraldehyde, the polymerised extract (PE) appeared as a diffuse smear in SDS-PAGE, with most proteins exceeding 100 kDa. Although some IgE reactivity was retained, virtually all protein bands below 50 kDa, including the 16 kDa band likely corresponding to Alt a 1, were no longer detectable after polymerisation (Figure 1A).
3.2. Physicochemical Composition Analysis of the Extracts: Amino acid Content, Nuclear Magnetic Resonance and Mass Spectrometry
Upon addition of glutaraldehyde to the NE, the amine groups of proteins, primarily lysine (Lys), arginine (Arg), asparagine/aspartate (Asx), and glutamine/glutamate (Glx), underwent modification. These residues became undetectable in the PE by gas chromatography. Specifically, gas chromatography revealed reductions of 87.5% for Lys, 76.9% for Asx, 75.9% for Glx, and 46.9% for Arg in the PE, indicating successful polymerisation (Figure 1B).
NMR-DOSY spectra of the NE revealed three distinct populations (1NE, 2NE, and 3NE), representing heterogeneous protein particles of varying molecular weights (Figure 1C). In contrast, the PE exhibited a single, homogeneous population (1PE) with a larger hydrodynamic radius than any NE population, indicating the formation of a uniform high molecular weight molecule. Despite structural changes induced by polymerisation, both the NE and PE exhibited similar carbohydrate (δ = 3.5–4.5 ppm) and protein (δ aliphatic = 0–2.5 ppm; δ aromatic = 4.5–5.5 ppm) composition profiles, as determined by NMR-DOSY. These findings suggest that glutaraldehyde polymerisation altered the protein structure without significantly affecting the overall chemical composition of the NE. Mass spectrometry analysis identified over 1,000 proteins in the extracts. Seven allergens were detected in the NE, including Alt a 1, Alt a 3, Alt a 6, Alt a 10, Alt a 13, Alt a 14, and Alt a 15.
3.3. Allergenic Activity Analysis: IgE/IgG ELISA Competition Assays
To evaluate the allergenic activity of polymerised and native extracts, we performed ELISA competition assays.
In ELISA competition assays, NE inhibited e 50% of the IgE reactivity (IC50) to A. alternata at 0.046 µg NE/ml, while PE showed a reduced activity, with 0.344 µg PE/ml (Figure 2, left panel). Regarding the IgG reactivity, the NE inhibited 50% at 0.11 µg NE/ml, while PE showed similar results, with 0.13 µg PE/ml to inhibit the 50% of IgG reactivity to A. alternata (Figure 2, right panel). These results suggest that polymerised extracts show an 87% reduction in allergenic activity while its immunogenicity seems to be conserved.
3.4. Immunogenicity and Inhibition of IgE Reactivity by Mouse IgG Antibodies Induced with A. alternata PE
Western blot analysis was performed to assess the immunogenicity of NE and PE extracts of A. alternata, as well as the capacity of antibodies induced by these extracts to inhibit IgE binding from allergic patients. Mouse IgG antibodies induced by immunisation with A. alternata NE and PE were first probed against the respective extracts. In both cases, the antibodies strongly recognised multiple protein bands in the extracts, indicating robust immunogenicity for both NE and PE (Figure 3A).
To evaluate the ability of these IgG antibodies to inhibit IgE binding from human sera, Western blot analysis of A. alternata NE and PE was conducted using human IgE pre-incubated with sera from immunised mice. IgE reactivity to native A. alternata allergens was significantly reduced in the presence of IgG antibodies generated through immunisation with PE (Figure 3B), comparable to the inhibition observed with IgG antibodies generated in mice immunised with NE.
These findings demonstrate that chemical modification of A. alternata extracts through polymerisation does not eliminate critical epitopes necessary for the induction of protective IgG antibodies.
3.5. Enzymatic Activity of Native and Polymerised Extracts
Enzymatic activity analysis using ApiZym kits detected 13 of 19 enzymatic activities in the A. alternata NE (Table 1). Quantitative analysis revealed that serine protease activity was the most abundant, followed by alkaline phosphatase, trypsin, and acid phosphatase activities, while cysteine protease activity was the least abundant. In the PE, all enzymatic activities were significantly reduced compared to the NE. Trypsin and serine protease activities decreased by 99.73% and 90.05%, respectively, while acid and alkaline phosphatase activities dropped by 71.33% and 75.5%. Cysteine protease activity was reduced by 59.15% (Table 2).
3.6. Stability of Grass Extracts in Mixture with Alternaria Allergoids
The reduced proteolytic activity observed in the PE of A. alternata was confirmed in mixtures with grass (Phleum pratense) NE. Protein and allergenic profiles of the P. pratense mixtures were analysed at different incubation times and compared to the grass NE control dissolved in distilled water (Figure 4). In mixtures of the two native extracts, degradation of grass proteins was observed within 1 hour, with the disappearance of bands >37 kDa compared to the control profile. After 3 months, this degradation became more pronounced, as indicated by reduced band intensity. In contrast, when grass NE was mixed with the A. alternata PE, the protein profile remained stable and similar to the grass control throughout the study.
The allergenic profile of the grass control extract and the mixture with A. alternata PE showed a general decrease in band intensity at the 3-month time point.
Biological IgE activity of the mixtures was evaluated by ELISA competition. The potency of the P. pratense control remained stable throughout the study (Figure 5A). However, when mixed with mould NE, grass potency dropped to 67.78% after 1 hour and further declined to 3.36% after 3 months (Figure 5B). In contrast, grass NE mixed with A. alternata PE maintained similar potency to the control throughout the study.
4. Discussion
A pivotal stage in allergen immunotherapy (AIT) involves the selection of extract type, allergen content, and overall allergenic activity. Key factors contributing to treatment failures include inadequate allergen dosage, potential adverse effects, and the prolonged duration of treatment. The use of A. alternata extracts in AIT has been controversial due to challenges in standardisation and the associated high risk of adverse reactions [38,39]. In addition, A. alternata produces a broad and complex array of allergenic proteins [40] that collectively contribute to allergy symptoms [41], potentially limiting the effectiveness of single-allergen immunotherapy in patients sensitised to multiple A. alternata allergens. Native fungal allergenic extracts have a complex composition of proteins, carbohydrates and other components that do not contribute to allergenicity but may produce adverse effects during treatment, so the safety of subcutaneous immunotherapy with fungi native extracts was questioned [41]. To mitigate secondary adverse reactions, strategies such as hypoallergens, peptide-derived vaccines, or purified allergens have been developed. In this study, we demonstrate that glutaraldehyde-treated A. alternata extracts can generate allergoids with reduced IgE-binding capacity while preserving immunogenicity across various allergens, including Alt a 1. These allergoids represent a safer and effective alternative to conventional vaccines for A. alternata allergy treatment.
The mechanisms underlying the advantages of allergoids over native extracts in AIT are not fully elucidated. Current evidence suggests that chemical modification reduces IgE-binding capacity while retaining recognition by IgG and linear T-cell epitopes, facilitating a protective immune response characterised by strong IgG production and/or regulatory IL-10-dependent T-cell responses [42,43,44,45]. However, it is increasingly evident that specific IgG production alone does not guarantee protection. Factors such as epitope specificity and IgG affinity play crucial roles in determining therapeutic efficacy [46,47,48]. Maintaining the native epitope repertoire during allergenic extract modification is critical for generating blocking IgG antibodies that specifically inhibit IgE-allergen interactions.
We demonstrate that mouse IgG antibodies, generated through immunization with A. alternata allergoid, exhibit the capacity to inhibit the reactivity of native allergens present in the extract against human IgE from allergic patients. These results are important because indirectly indicate that the chemical modification did not affect the epitopes or prevent them from inducing a protective immunological response based on the induction of blocking IgG.
The use of a complete source of Alternaria antigens offers an advantage over single-allergen immunotherapy. As aforementioned, Alt a 1 is the most important Alternaria allergen, but it is not the unique sensitizing allergen. There are studies that show the great relevance of some of other minor allergens, such as Alt a 13, Alt a 4 and Alt a 8 [49]. Our immunoblotting results indicate that other Alternaria proteins are recognised by IgE from sensitised patients. Prevalence of sensitisation of other Alternaria allergens and their relevance in symptomatology has not been well studied. As an exception, Armentia et al. found that 46.7% of patients with allergy to fungi were sensitised to Alt a 6 following a molecular analysis [50]. Recently, Rodriguez et al. observed that, among 64 patients, the percentage of recognition for Alt a 3, Alt a 4, and/or Alt a 6, Alt a 7, Alt a 8, Alt a 10, and/or Alt a 15 was 1.6%, 21.9%, 12.5%, 12.5%, and 12.5%, respectively. Notably, 30% of the patients exhibited recognition to several Alternaria allergens beyond Alt a 1 [51]. Furthermore, sensitization to these allergens may play a crucial role in pathology. Illustratively, in a study involving 100 atopic dermatitis patients, sensitisation to Alt a 6 was found to be associated with a subgroup of individuals suffering bronchial asthma [52].
Fungal enzymes play an important role in allergy, both indirectly, by facilitating the passage of other allergens across the epithelial barrier, and directly, by acting on dendritic cells and promoting Th2-type responses. Cross-linking of enzymes with GA leads to a series of structural changes in the enzymes that result in decreased proteolytic capacity and possibly their allergenicity. Enzyme analysis of the extracts showed a significant decrease (p<0.001) in proteolytic activity in the polymerised extract compared to the native extract. The most significant decrease was observed in serine protease activity (including trypsins), with more than 90% loss of the proteolytic capacity. Phosphatase activity also showed a 71-75% reduction, while cysteine protease activity (59.15%), which are less abundant in the extract.
The reduced enzymatic activity of A. alternata allergoids has important implications for polysensitised patients. Fungal enzymes in native extracts are known to degrade allergens from other sources when combined in aqueous mixtures [53,54,55,56]. Our findings demonstrate that P. pratense allergens were degraded within an hour when mixed with native A. alternata extracts, resulting in a 33.2% reduction in biological potency after the first hour of incubation. In contrast, mixtures with polymerised A. alternata extracts maintained the stability and potency of grass allergens, as confirmed by electrophoresis and ELISA competition assays. Kordash et al. obtained similar results for extracts of Lolium perenne in a glycerol mixture with extracts of the fungus Helmintosporium spp. by observing a decrease in potency, as assessed by RAST. However, the biological activity estimated by skin tests was not altered [57].
5. Conclusions
Polymerising A. alternaria native extracts results in hypoallergenic allergoids that induce robust IgG responses that can potentially regulate the allergic response generated after natural exposure to A. alternaria. These polymerised allergens constitute an alternative to replace native allergenic extracts for AIT and can be considered to be co-administered in mixture with grass allergy treatments.
Author Contributions
Conceptualization, E.A.-F. and E.F-C.; methodology, E.A.-F.; writing—original draft preparation, E.A.-F.; writing—review and editing, E.F.-C. and S.I. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The animal study was approved by Dirección General de Agricultura, Ganadería y Alimentación de la Comunidad de Madrid, under the corresponding PROEX protocol. The study was conducted in accordance with the local legislation and institutional requirements.
Informed Consent Statement
Not applicable.
Data Availability Statement
This study did not generate publicly archive dataset.
Conflicts of Interest
E.A.-F. and S.I. are employees in Inmunotek S. L.
References
- López Couso, V.P.; Tortajada-Girbés, M.; Rodriguez Gil, D.; Martínez Quesada, J.; Palacios Pelaez, R. Fungi Sensitization in Spain: Importance of the Alternaria alternata Species and Its Major Allergen Alt a 1 in the Allergenicity. J Fungi (Basel) 2021, 7. [Google Scholar] [CrossRef] [PubMed]
- Masaki, K.; Fukunaga, K.; Matsusaka, M.; Kabata, H.; Tanosaki, T.; Mochimaru, T.; Kamatani, T.; Ohtsuka, K.; Baba, R.; Ueda, S. Characteristics of severe asthma with fungal sensitization. Annals of Allergy, Asthma & Immunology 2017, 119, 253–257. [Google Scholar] [CrossRef]
- Möller, C.; Dreborg, S.; Ferdousi, H.A.; Halken, S.; Høst, A.; Jacobsen, L.; Koivikko, A.; Koller, D.Y.; Niggemann, B.; Norberg, L.A.; et al. Pollen immunotherapy reduces the development of asthma in children with seasonal rhinoconjunctivitis (the PAT-study). J Allergy Clin Immunol 2002, 109, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Klain, A.; Senatore, A.A.; Licari, A.; Galletta, F.; Bettini, I.; Tomei, L.; Manti, S.; Mori, F.; Miraglia del Giudice, M.; Indolfi, C. The Prevention of House Dust Mite Allergies in Pediatric Asthma. Children 2024, 11, 469. [Google Scholar] [CrossRef]
- Tosca, M.A.; Licari, A.; Olcese, R.; Marseglia, G.; Sacco, O.; Ciprandi, G. Immunotherapy and Asthma in Children. Frontiers in Pediatrics 2018, 6. [Google Scholar] [CrossRef]
- Di Bona, D.; Frisenda, F.; Albanesi, M. Efficacy and safety of allergen immunotherapy in patients with allergy to molds: A systematic review. 2018, 48, 1391–1401. [CrossRef]
- Brehler, R.; Rabe, U. Allergen-specific immunotherapy for mold allergies. Allergo Journal International 2024, 33, 101–105. [Google Scholar] [CrossRef]
- Ostergaard, P.A.; Kaad, P.H.; Kristensen, T. A prospective study on the safety of immunotherapy in children with severe asthma. Allergy 1986, 41, 588–593. [Google Scholar] [CrossRef]
- Twaroch, T.E.; Curin, M.; Valenta, R.; Swoboda, I. Mold allergens in respiratory allergy: From structure to therapy. Allergy, asthma and immunology research 2015, 7, 205–220. [Google Scholar] [CrossRef]
- Kaad, P.H.; Ostergaard, P.A. The hazard of mould hyposensitization in children with asthma. Clinical allergy 1982, 12, 317–320. [Google Scholar] [CrossRef]
- Jutel, M.; Agache, I.; Bonini, S.; Burks, A.W.; Calderon, M.; Canonica, W.; Cox, L.; Demoly, P.; Frew, A.J.; O'Hehir, R.; et al. International Consensus on Allergen Immunotherapy II: Mechanisms, standardization, and pharmacoeconomics. J Allergy Clin Immunol 2016, 137, 358–368. [Google Scholar] [CrossRef]
- Alvaro-Lozano, M.; Akdis, C.A.; Akdis, M.; Alviani, C.; Angier, E.; Arasi, S.; Arzt-Gradwohl, L.; Barber, D.; Bazire, R.; Cavkaytar, O.; et al. EAACI Allergen Immunotherapy User's Guide. Pediatr. Allergy Immunol. Off. Publ. Eur. Soc. Pediatr. Allergy Immunol. 2020, 31 (Suppl. S25), 1–101. [Google Scholar] [CrossRef]
- Akdis, M.; Akdis, C.A. Mechanisms of allergen-specific immunotherapy: Multiple suppressor factors at work in immune tolerance to allergens. J Allergy Clin Immunol 2014, 133, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, L.; Valovirta, E. How strong is the evidence that immunotherapy in children prevents the progression of allergy and asthma? Current opinion in allergy and clinical immunology 2007, 7, 556–560. [Google Scholar] [CrossRef] [PubMed]
- Ricketti, P.A.; Alandijani, S.; Lin, C.H.; Casale, T.B. Investigational new drugs for allergic rhinitis. Expert opinion on investigational drugs 2017, 26, 279–292. [Google Scholar] [CrossRef]
- Ouyang, X.; Reihill, J.A.; Douglas, L.E.J.; Martin, S.L. Airborne indoor allergen serine proteases and their contribution to sensitisation and activation of innate immunity in allergic airway disease. Eur Respir Rev 2024, 33. [Google Scholar] [CrossRef]
- Kespohl, S.; Raulf, M. Mould allergens: Where do we stand with molecular allergy diagnostics?: Part 13 of the series Molecular Allergology. Allergo journal international 2014, 23, 120–125. [Google Scholar] [CrossRef]
- Kobayashi, T.; Iijima, K.; Radhakrishnan, S.; Mehta, V.; Vassallo, R.; Lawrence, C.B.; Cyong, J.C.; Pease, L.R.; Oguchi, K.; Kita, H. Asthma-related environmental fungus, Alternaria, activates dendritic cells and produces potent Th2 adjuvant activity. Journal of immunology 2009, 182, 2502–2510. [Google Scholar] [CrossRef]
- Cardona Villa, R.; Yépes Núñez, J.J.; Salgado Vélez, H.; Montoya Guarín, C.J. Aspectos básicos de las reacciones de hipersensibilidad y la alergia. In Alergia. Abordaje clínico, diagnóstico y tratamiento, Panamericana, Ed. 2010; p. 736.
- Borger, P.; Koeter, G.H.; Timmerman, J.A.; Vellenga, E.; Tomee, J.F.; Kauffman, H.F. Proteases from Aspergillus fumigatus induce interleukin (IL)-6 and IL-8 production in airway epithelial cell lines by transcriptional mechanisms. The Journal of infectious diseases 1999, 180, 1267–1274. [Google Scholar] [CrossRef]
- Kauffman, H.F.; Tomee, J.F.C.; van de Riet, M.A.; Timmerman, A.J.B.; Borger, P. Protease-dependent activation of epithelial cells by fungal allergens leads to morphologic changes and cytokine production. Journal of Allergy and Clinical Immunology 2000, 105, 1185–1193. [Google Scholar] [CrossRef]
- Tai, H.Y.; Tam, M.F.; Chou, H.; Peng, H.J.; Su, S.N.; Perng, D.W.; Shen, H.D. Pen ch 13 allergen induces secretion of mediators and degradation of occludin protein of human lung epithelial cells. Allergy 2006, 61, 382–388. [Google Scholar] [CrossRef]
- Lamhamedi-Cherradi, S.-E.; Martin, R.E.; Ito, T.; Kheradmand, F.; Corry, D.B.; Liu, Y.-J.; Moyle, M. Fungal proteases induce Th2 polarization through limited dendritic cell maturation and reduced production of IL-12. The Journal of Immunology 2008, 180, 6000–6009. [Google Scholar] [CrossRef] [PubMed]
- Boitano, S.; Flynn, A.N.; Sherwood, C.L.; Schulz, S.M.; Hoffman, J.; Gruzinova, I.; Daines, M.O. Alternaria alternata serine proteases induce lung inflammation and airway epithelial cell activation via PAR2. American journal of physiology. Lung cellular and molecular physiology 2011, 300, L605–L614. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.C.; Chuang, J.G.; Su, Y.Y.; Chiang, B.L.; Lin, Y.S.; Chow, L.P. The protease allergen Pen c 13 induces allergic airway inflammation and changes in epithelial barrier integrity and function in a murine model. The Journal of biological chemistry 2011, 286, 26667–26679. [Google Scholar] [CrossRef] [PubMed]
- Snelgrove, R.J.; Gregory, L.G.; Peiro, T.; Akthar, S.; Campbell, G.A.; Walker, S.A.; Lloyd, C.M. Alternaria-derived serine protease activity drives IL-33-mediated asthma exacerbations. The Journal of allergy and clinical immunology 2014, 134, 583–592. [Google Scholar] [CrossRef]
- Casanovas, M.; Fernandez-Caldas, E.; Alamar, R.; Basomba, A. Comparative study of tolerance between unmodified and high doses of chemically modified allergen vaccines of Dermatophagoides pteronyssinus. Int Arch Allergy Immunol 2005, 137, 211–218. [Google Scholar] [CrossRef]
- Chaker, A.M.; Al-Kadah, B.; Luther, U.; Neumann, U.; Wagenmann, M. An accelerated dose escalation with a grass pollen allergoid is safe and well-tolerated: A randomized open label phase II trial. Clinical and translational allergy 2015, 6, 4. [Google Scholar] [CrossRef]
- Himly, M.; Carnes, J.; Fernandez-Caldas, E.; Briza, P.; Ferreira, F. Characterization of allergoids. Arbeiten aus dem Paul-Ehrlich-Institut (Bundesinstitut fur Impfstoffe und biomedizinische Arzneimittel) Langen/Hessen 2009, 96, 61–69. [Google Scholar]
- Manzano, A.I.; Javier Canada, F.; Cases, B.; Sirvent, S.; Soria, I.; Palomares, O.; Fernandez-Caldas, E.; Casanovas, M.; Jimenez-Barbero, J.; Subiza, J.L. Structural studies of novel glycoconjugates from polymerized allergens (allergoids) and mannans as allergy vaccines. Glycoconj J 2016, 33, 93–101. [Google Scholar] [CrossRef]
- Morales, M.; Gallego, M.; Iraola, V.; Taules, M.; de Oliveira, E.; Moya, R.; Carnes, J. In vitro evidence of efficacy and safety of a polymerized cat dander extract for allergen immunotherapy. BMC immunology 2017, 18, 10. [Google Scholar] [CrossRef]
- Passalacqua, G.; Pasquali, M.; Ariano, R.; Lombardi, C.; Giardini, A.; Baiardini, I.; Majani, G.; Falagiani, P.; Bruno, M.; Canonica, G.W. Randomized double-blind controlled study with sublingual carbamylated allergoid immunotherapy in mild rhinitis due to mites. Allergy 2006, 61, 849–854. [Google Scholar] [CrossRef]
- Rodriguez Mosquera, M.; Sola Martinez, F.J.; Montoro de Francisco, A.; Chivato Perez, T.; Sanchez Moreno, V.; Rodriguez-Marco, A.; Hernandez-Pena, J. Satisfaction and perceived effectiveness in patients on subcutaneous immunotherapy with a high-dose hypoallergenic pollen extract. European annals of allergy and clinical immunology 2016, 48, 188–193. [Google Scholar] [PubMed]
- Roger, A.; Depreux, N.; Jurgens, Y.; Heath, M.D.; Garcia, G.; Skinner, M.A. A novel and well tolerated mite allergoid subcutaneous immunotherapy: Evidence of clinical and immunologic efficacy. Immunity, inflammation and disease 2014, 2, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez Del Río, P.; Vidal, C.; Just, J.; Tabar, A.I.; Sanchez-Machin, I.; Eberle, P.; Borja, J.; Bubel, P.; Pfaar, O.; Demoly, P.; et al. The European Survey on Adverse Systemic Reactions in Allergen Immunotherapy (EASSI): A paediatric assessment. Pediatr. Allergy Immunol. Off. Publ. Eur. Soc. Pediatr. Allergy Immunol. 2017, 28, 60–70. [Google Scholar] [CrossRef]
- Abel-Fernández, E. Producción y caracterización de extractos alergénicos polimerizados de Alternaria alternata. Evaluación de su alergenicidad e inmunogenicidad. Universidad Complutense de Madrid, Madrid, España, 2017.
- Morales, M.; Gallego, M.T.; Iraola, V.; Moya, R.; Santana, S.; Carnes, J. Preclinical safety and immunological efficacy of Alternaria alternata polymerized extracts. Immunity, inflammation and disease 2018, 6, 234–244. [Google Scholar] [CrossRef]
- Di Bona, D.; Albanesi, M.; Macchia, L. Is immunotherapy with fungal vaccines effective? Current opinion in allergy and clinical immunology 2019, 19, 646–653. [Google Scholar] [CrossRef]
- Bozek, A.; Pyrkosz, K. Immunotherapy of mold allergy: A review. Human vaccines & immunotherapeutics 2017, 13, 2397–2401. [Google Scholar] [CrossRef]
- Gabriel, M.F.; Postigo, I.; Tomaz, C.T.; Martinez, J. Alternaria alternata allergens: Markers of exposure, phylogeny and risk of fungi-induced respiratory allergy. Environment international 2016, 89-90, 71–80. [Google Scholar] [CrossRef]
- Abel-Fernández, E.; Martínez, M.J.; Galán, T.; Pineda, F. Going over Fungal Allergy: Alternaria alternata and Its Allergens. J Fungi (Basel) 2023, 9. [Google Scholar] [CrossRef]
- Cirkovic, T.D.; Bukilica, M.N.; Gavrovic, M.D.; Vujcic, Z.M.; Petrovic, S.; Jankov, R.M. Physicochemical and immunologic characterization of low-molecular-weight allergoids of Dactylis glomerata pollen proteins. Allergy 1999, 54, 128–134. [Google Scholar] [CrossRef]
- Kahler, H.; Stuwe, H.; Cromwell, O.; Fiebig, H. Reactivity of T cells with grass pollen allergen extract and allergoid. Int Arch Allergy Immunol 1999, 120, 146–157. [Google Scholar] [CrossRef]
- López-Matas, M.A.; Gallego, M.; Iraola, V.; Robinson, D.; Carnés, J. Depigmented allergoids reveal new epitopes with capacity to induce IgG blocking antibodies. BioMed research international 2013, 2013, 284615. [Google Scholar] [CrossRef] [PubMed]
- Starchenka, S.; Bell, A.J.; Mwange, J.; Skinner, M.A.; Heath, M.D. Molecular fingerprinting of complex grass allergoids: Size assessments reveal new insights in epitope repertoires and functional capacities. World Allergy Organ J 2017, 10, 17. [Google Scholar] [CrossRef] [PubMed]
- Shamji, M.H.; Francis, J.N.; Würtzen, P.A.; Lund, K.; Durham, S.R.; Till, S.J. Cell-free detection of allergen-IgE cross-linking with immobilized phase CD23: Inhibition by blocking antibody responses after immunotherapy. J Allergy Clin Immunol 2013, 132, 1003–1005.e4. [Google Scholar] [CrossRef]
- Reithofer, M.; Böll, S.L.; Kitzmüller, C.; Horak, F.; Sotoudeh, M.; Bohle, B.; Jahn-Schmid, B. Alum-adjuvanted allergoids induce functional IgE-blocking antibodies. 2018, 48, 741–744. [CrossRef]
- Rauber, M.M.; Wu, H.K.; Adams, B.; Pickert, J.; Bohle, B. Birch pollen allergen-specific immunotherapy with glutaraldehyde-modified allergoid induces IL-10 secretion and protective antibody responses. 2019, 74, 1575–1579. [CrossRef]
- Abel-Fernández, E.; Fernández-Caldas, E. Allergy to fungi: Advances in the understanding of fungal allergens. Mol Immunol 2023, 163, 216–223. [Google Scholar] [CrossRef]
- Armentia, A.; Martín-Armentia, S.; Moral, A.; Montejo, D.; Martin-Armentia, B.; Sastre, R.; Fernández, S.; Corell, A.; Fernandez, D. Molecular study of hypersensitivity to spores in adults and children from Castile & Leon. Allergol Immunopathol (Madr) 2019, 47, 350–356. [Google Scholar] [CrossRef]
- Rodríguez, D.; Tabar, A.I.; Castillo, M.; Martínez-Gomariz, M.; Dobski, I.C.; Palacios, R. Changes in the Sensitization Pattern to Alternaria alternata Allergens in Patients Treated with Alt a 1 Immunotherapy. J Fungi (Basel) 2021, 7. [Google Scholar] [CrossRef]
- Celakovska, J.; Vankova, R.; Bukac, J.; Cermakova, E.; Andrys, C.; Krejsek, J. Atopic Dermatitis and Sensitisation to Molecular Components of Alternaria, Cladosporium, Penicillium, Aspergillus, and Malassezia—Results of Allergy Explorer ALEX 2. Journal of Fungi 2021, 7, 183. [Google Scholar] [CrossRef]
- Nelson, H.S.; Ikle, D.; Buchmeier, A. Studies of allergen extract stability: The effects of dilution and mixing. The Journal of allergy and clinical immunology 1996, 98, 382–388. [Google Scholar] [CrossRef]
- Grier, T.J.; LeFevre, D.M.; Duncan, E.A.; Esch, R.E. Stability of standardized grass, dust mite, cat, and short ragweed allergens after mixing with mold or cockroach extracts. Ann. Allergy Asthma Immunol. Off. Publ. Am. Coll. Allergy Asthma Immunol. 2007, 99, 151–160. [Google Scholar] [CrossRef]
- Esch, R.E. Allergen immunotherapy: What can and cannot be mixed? The Journal of allergy and clinical immunology 2008, 122, 659–660. [Google Scholar] [CrossRef]
- Coop, C.A. Immunotherapy for mold allergy. Clinical reviews in allergy and immunology 2014, 47, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Kordash, T.R.; Amend, M.J.; Williamson, S.L.; Jones, J.K.; Plunkett, G.A. Effect of mixing allergenic extracts containing Helminthosporium, D. farinae, and cockroach with perennial ryegrass. Annals of allergy 1993, 71, 240–246. [Google Scholar]
Figure 1.
Protein and physicochemical characterisation of A. alternata native and polymerised extracts. A) Protein (SDS-PAGE) and allergenic (Western blot) profiles of native and polymerised extracts. Bands of varying molecular weights, corresponding to allergenic and non-allergenic proteins, were detected in the native extract (NE). In the polymerised extract (PE), proteins appeared as a diffuse smear, predominantly >100 kDa. MW: Molecular weight marker; NE: Native extract; PE: Polymerised extract. B) Amino acid molar composition of native and polymerised extracts (%). The polymerised extract (PE) showed reduced detection of Lys, Arg, Asx, and Glx compared to the native extract (NE). The Gly content increased in the PE due to its use in stopping the polymerisation reaction. NE: Native extract; PE: Polymerised extract. C) NMR analysis of Alternaria alternata native and polymerised extracts. Superimposed ¹H NMR spectra and DOSY experiments are shown. The diffusion coefficient (D, m²/s) is displayed on the y-axis, with more negative values indicating larger hydrodynamic radii. The chemical shift (δ, ppm) is shown on the x-axis, highlighting carbohydrate regions (3.5–4.5 ppm) and protein regions, including aliphatic (0–3 ppm) and aromatic (4.5–5.5 ppm). NE: Native extract (blue); PE: Polymerised extract (black).
Figure 1.
Protein and physicochemical characterisation of A. alternata native and polymerised extracts. A) Protein (SDS-PAGE) and allergenic (Western blot) profiles of native and polymerised extracts. Bands of varying molecular weights, corresponding to allergenic and non-allergenic proteins, were detected in the native extract (NE). In the polymerised extract (PE), proteins appeared as a diffuse smear, predominantly >100 kDa. MW: Molecular weight marker; NE: Native extract; PE: Polymerised extract. B) Amino acid molar composition of native and polymerised extracts (%). The polymerised extract (PE) showed reduced detection of Lys, Arg, Asx, and Glx compared to the native extract (NE). The Gly content increased in the PE due to its use in stopping the polymerisation reaction. NE: Native extract; PE: Polymerised extract. C) NMR analysis of Alternaria alternata native and polymerised extracts. Superimposed ¹H NMR spectra and DOSY experiments are shown. The diffusion coefficient (D, m²/s) is displayed on the y-axis, with more negative values indicating larger hydrodynamic radii. The chemical shift (δ, ppm) is shown on the x-axis, highlighting carbohydrate regions (3.5–4.5 ppm) and protein regions, including aliphatic (0–3 ppm) and aromatic (4.5–5.5 ppm). NE: Native extract (blue); PE: Polymerised extract (black).
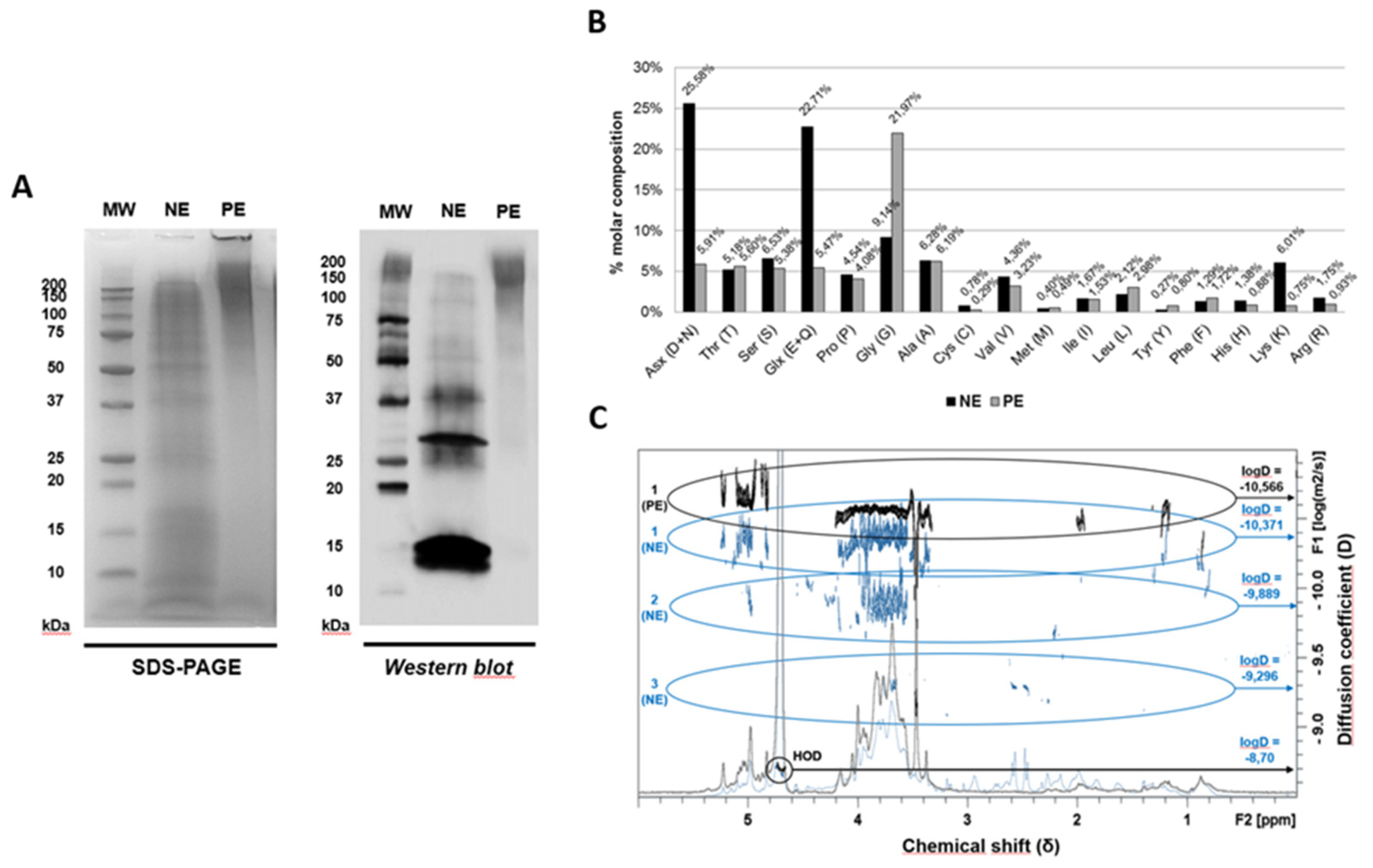
Figure 2.
IgE and IgG binding capacities of native and polymerised A. alternata extracts. Graphical representation of IgE and IgG ELISA competition results for native and polymerised extracts. The 50% inhibition values for each extract are summarised in the accompanying tables (µg/ml).
Figure 2.
IgE and IgG binding capacities of native and polymerised A. alternata extracts. Graphical representation of IgE and IgG ELISA competition results for native and polymerised extracts. The 50% inhibition values for each extract are summarised in the accompanying tables (µg/ml).
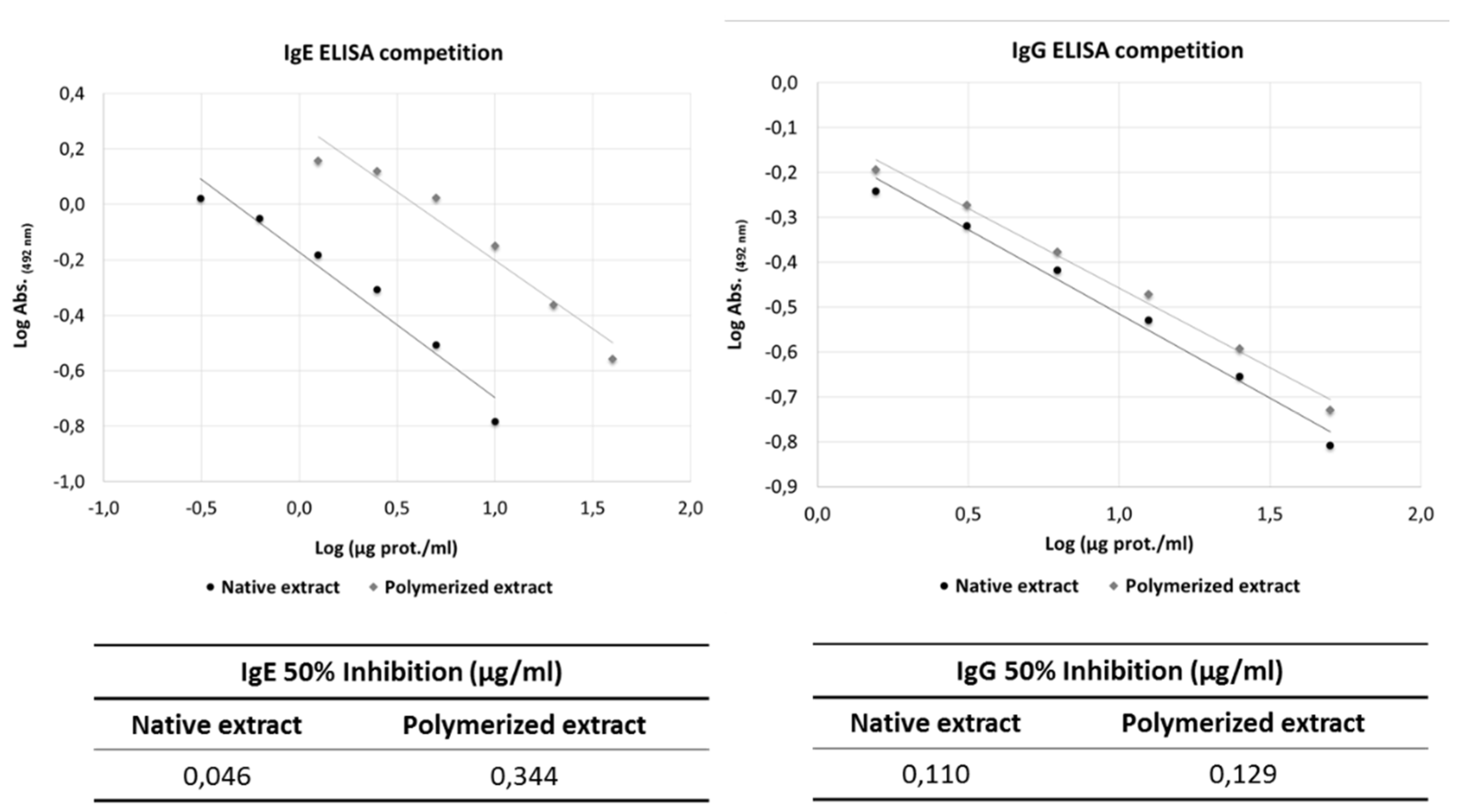
Figure 3.
Inhibition of human IgE binding by mouse IgG induced with A. alternata allergoids A) Western blot analysis of IgG from mice immunised with native (NE) or polymerised (PE) extracts, probed with NE and PE. B) Western blot analysis of human IgE binding to NE and PE, inhibited with sera from mice immunised with NE or PE. Immunisation with both native and polymerised extracts induced antibodies capable of blocking the binding of IgE from allergic patient.
Figure 3.
Inhibition of human IgE binding by mouse IgG induced with A. alternata allergoids A) Western blot analysis of IgG from mice immunised with native (NE) or polymerised (PE) extracts, probed with NE and PE. B) Western blot analysis of human IgE binding to NE and PE, inhibited with sera from mice immunised with NE or PE. Immunisation with both native and polymerised extracts induced antibodies capable of blocking the binding of IgE from allergic patient.
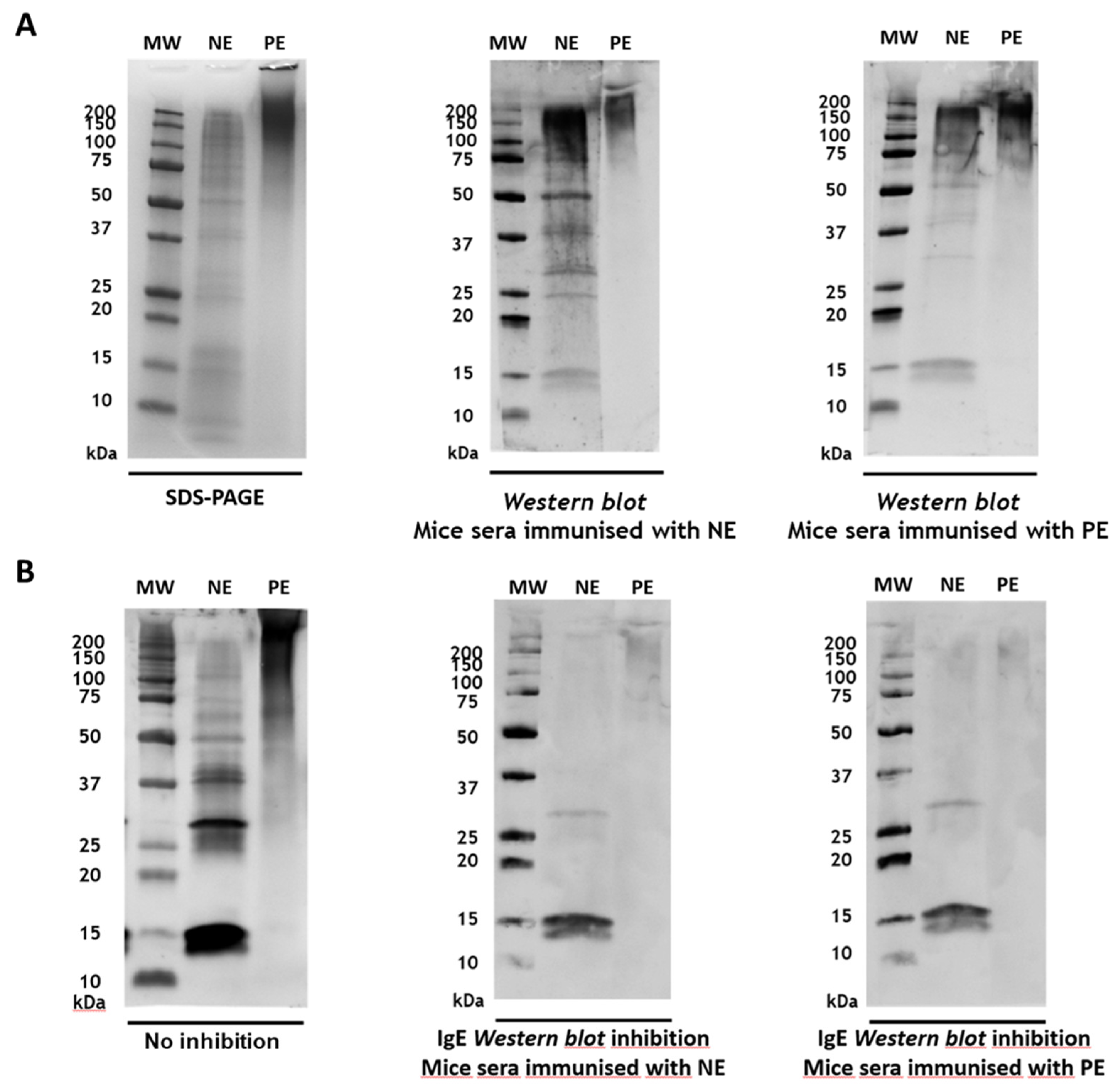
Figure 4.
Stability of P. pratense protein and allergenic profiles in mixtures with native or polymerised A. alternata extracts SDS-PAGE (top) and Western blot IgE (botton) of the P. pratense mixtures and control, after different intervals of incubation. 1: Grass native extract; 2: P. pratense and A. alternata native extracts mixture; 3: P. pratense native extract and A. alternata polymerised extract mixture. MW: Molecular Weight Marker.
Figure 4.
Stability of P. pratense protein and allergenic profiles in mixtures with native or polymerised A. alternata extracts SDS-PAGE (top) and Western blot IgE (botton) of the P. pratense mixtures and control, after different intervals of incubation. 1: Grass native extract; 2: P. pratense and A. alternata native extracts mixture; 3: P. pratense native extract and A. alternata polymerised extract mixture. MW: Molecular Weight Marker.
Figure 5.
Preservation of P. pratense biological potency in mixtures with polymerised A. alternata extracts A) IgE biological activity (50% inhibition) of P. pratense native extracts (controls) over different incubation intervals. These values were set as the 100% potency reference for each interval to evaluate the mixtures’ potency. B) Potency percentage of P. pratense native extract mixed with A. alternata native or polymerised extracts relative to the grass native extract (control) at different incubation intervals.
Figure 5.
Preservation of P. pratense biological potency in mixtures with polymerised A. alternata extracts A) IgE biological activity (50% inhibition) of P. pratense native extracts (controls) over different incubation intervals. These values were set as the 100% potency reference for each interval to evaluate the mixtures’ potency. B) Potency percentage of P. pratense native extract mixed with A. alternata native or polymerised extracts relative to the grass native extract (control) at different incubation intervals.
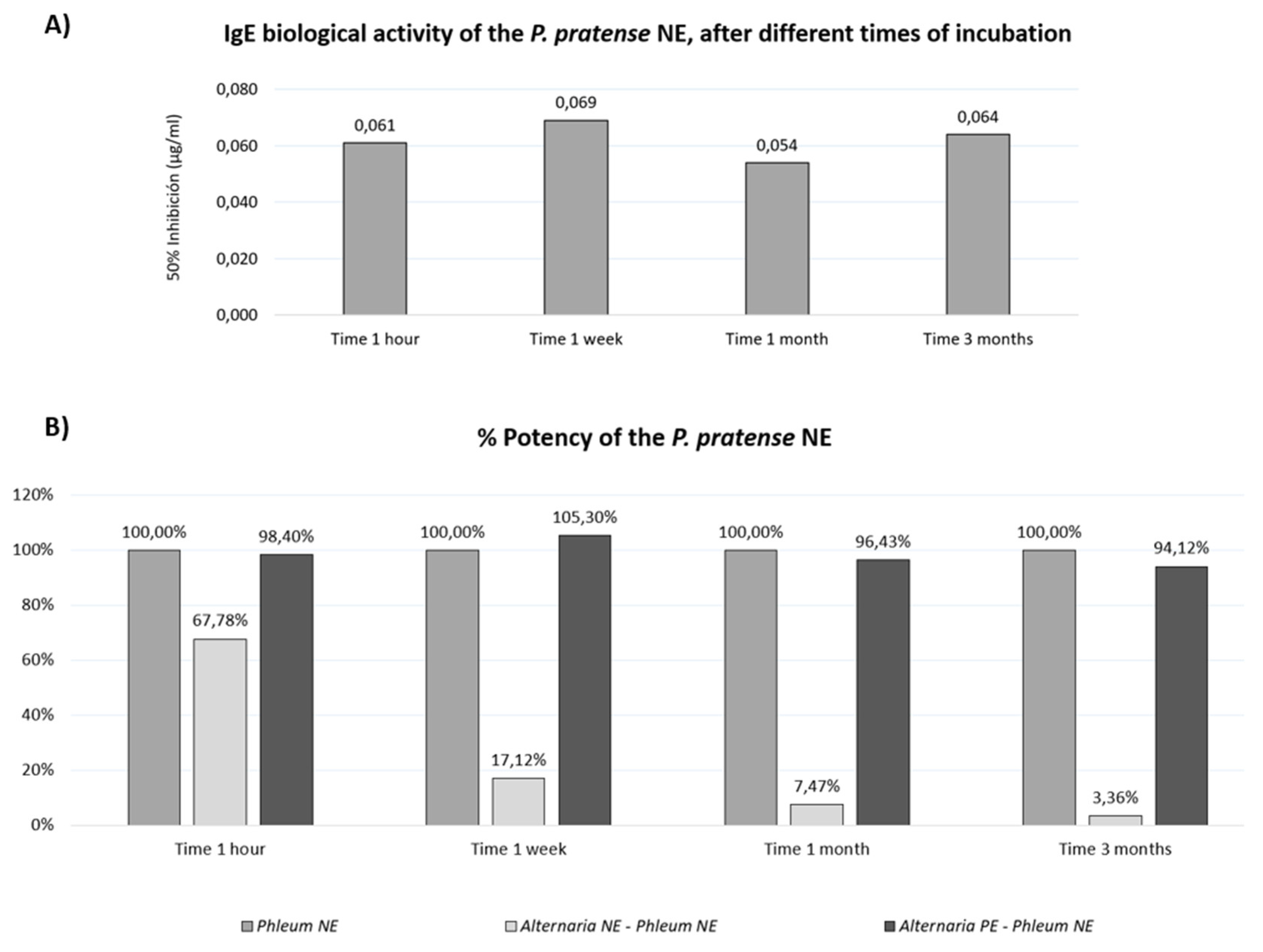

Table 1.
Qualitative determination of the enzymatic activity in A. alternata native extracts (+: detected, -: not detected).
Table 1.
Qualitative determination of the enzymatic activity in A. alternata native extracts (+: detected, -: not detected).
| ENZYMATIC ACTIVITY | NATIVE EXTRACT |
| Alkaline phosphatase | + |
| Esterase (C4) | + |
| Esterase lipase (C8) | + |
| Lipase (C14) | - |
| Leucine arylamidase | + |
| Valine arylamidase | - |
| Cystine arylamidase | - |
| Trypsin | + |
| α-chymotrypsin | - |
| Acid phosphatase | + |
| Naphthol AS-BI-phosphohydrolase | + |
| α-galactosidase | + |
| β -galactosidase | + |
| β-glucuronidase | - |
| α-glucosidase | + |
| β-glucosidase | + |
| N-acetyl-β-glucosaminidase | + |
| β-mannosidase | + |
| α-fucosidase | - |
Table 2.
Results of quantification of the specific proteolytic activities assessed in the A. alternata native and polymerised extracts (expressed in enzyme equivalents/protein mg). The percentage of diminution is referred to the diminution of the proteolytic activity in the polymerised extract regarding to the native extract.
Table 2.
Results of quantification of the specific proteolytic activities assessed in the A. alternata native and polymerised extracts (expressed in enzyme equivalents/protein mg). The percentage of diminution is referred to the diminution of the proteolytic activity in the polymerised extract regarding to the native extract.
| PROTEOLYTIC ACTIVITY | NATIVE EXTRACT | POLYMERISED EXTRACT | % DIMINUTION |
| Trypsin | 119.049 | 0.321 | 99.73 |
| Serine protease | 6172.52 | 614.03 | 90.05 |
| Cysteine protease | 3.55 | 1.45 | 59.15 |
| Acid phosphatase | 98.62 | 28.27 | 71.33 |
| Alkaline phosphatase | 543.86 | 134.80 | 75.21 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



