Submitted:
11 January 2025
Posted:
14 January 2025
You are already at the latest version
Abstract
Root-knot nematodes (Meloidogyne spp.) are major pests of vegetable crops, causing significant yield losses in numerous species worldwide. Their widespread distribution and polyphagous nature make them particularly challenging to manage effectively. The use of biological agents and botanical extracts has emerged as a sustainable alternative to chemical nematicides. The present study evaluated the effectiveness of aqueous extracts from Moringa oleifera and a novel strain of plant growth-promoting rhizobacteria, Bacillus australimaris strain LWD73, native to Florida, in reducing Meloidogyne incognita infection and enhancing plant growth. The combined application of M. oleifera extracts and B. australimaris LWD73 significantly reduced the number of galls, eggs per root system, second-stage juveniles (J2) per 60 cc of soil, and the nematode reproduction factor. Moreover, it improved key growth parameters in tomato cultivar HM1824. Fourier Transform Infrared Spectroscopy (FTIR) and Gas Chromatography-Mass Spectrometry (GC-MS) analyses of M. oleifera extracts identified antimicrobial and nematicidal phytochemicals. Similarly, solid-phase microextraction GC-MS (SPME-GC/MS) analysis of B. australimaris LWD73 revealed the presence of volatile organic compounds (VOCs), including 2-nonanone and 2-undecanone, which are known for their potent nematicidal properties. The synergistic application of M. oleifera and B. australimaris LWD73 demonstrated enhanced nematode suppression and improved plant growth, offering a promising, eco-friendly approach to managing root-knot nematodes in vegetable crops.
Keywords:
root-knot nematodes
; biological control
; Meloidogyne incognita
; Moringa oleifera
; rhizobacteria
; bacterial VOCs
; FTIR
; GC-MS
1. Introduction
Root-knot nematodes (Meloidogyne spp.) have a devastating impact on vegetable crops globally, causing significant yield losses and compromising overall plant health. These polyphagous nematodes have an extensive host range, infecting over 3,000 plant species [1,2]. Upon infection, they induce the formation of root galls, which disrupt normal root functions and impair the ability of plant to efficiently absorb water and nutrients from the soil. This leads to stunted growth and reduced crop yields. Moreover, root-knot nematode infections often predispose plants to secondary infections by soil-borne pathogens, exacerbating the damage and further diminishing plant health [3,4].
The severity of the damage caused by root-knot nematodes depends on various factors, including environmental conditions, nematode population density, species virulence, and the susceptibility of the host plant. Managing these nematodes is particularly challenging due to their high reproductive potential and remarkable adaptability [5]. Although chemical nematicides have proven effective in controlling root-knot nematodes, their adverse environmental impacts have driven a shift toward developing alternative, eco-friendly management strategies [6,7].
The use of biological control agents, primarily bacteria and fungi, offers a promising and sustainable approach to suppressing nematode populations [5]. Likewise, plant-based natural products have also been reported to be effective against a wide range of pathogens [8,9,10,11,12]. Botanical management of root-knot nematodes using plant-based formulations has also demonstrated significant potential. Incorporating macerated leaves of Cannabis sativa and Azadirachta indica into the soil significantly reduced Meloidogyne javanica populations and infection severity in peach plants in a dose-dependent manner [13]. Similarly, the combined application of dried neem leaves and the fungus Trichoderma harzianum on tomato plants resulted in a greater reduction in root galls, egg mass production, and egg hatching compared to their individual applications. These combined effects led to increased plant height and fresh shoot weight, with outcomes strongly influenced by both dosage and exposure time [14].
Plant growth-promoting rhizobacteria (PGPR) play a vital role in mitigating plant stress and enhancing plant growth, either directly or indirectly. PGPR offer numerous benefits, including increased mineral availability, production of phytohormones, mitigation of heavy metal stress, and biocontrol of plant pathogens. These attributes contribute to improved nutrient uptake, enhanced plant growth, and higher crop yields [15,16,17].
For sustainable rice and sugarcane production, bacteria belonging to the genera Bacillus, Rhizobium, Comamonas, Cyanobacteria, Nodosilinea, Levinella, and Pseudomonas have been identified as efficient producers of nitrogen and solubilizers of inorganic phosphate, potassium, and other macro- and micronutrients [15].
Moreover, a consortium of Bacillus cereus AR156, Bacillus subtilis SM21, and Serratia sp. XY21 has shown significant efficacy in reducing the severity of root-knot disease while also increasing the yield and fruit quality of cucumber in field trials [18].
Innovative approaches, such as gene editing to confer resistance in crops against nematode infestations, the exploration of bacteria, fungi, and botanicals as potential natural nematicides, and the implementation of cultural practices, are all sustainable strategies for suppressing nematode populations in soil without relying on chemical nematicides [5]. These methods aim to minimize environmental impact while effectively managing plant-parasitic nematodes.
Moringa oleifera has emerged as a promising natural alternative for managing root-knot nematodes. The bioactive compounds in M. oleifera, such as alkaloids, flavonoids, and phenolic compounds, exhibit strong nematicidal properties, disrupting the life cycle of nematodes by impairing egg hatching and larval development. Studies have shown that both aqueous and methanolic extracts of M. oleifera leaves and seeds significantly reduce nematode population densities in the soil and gall formation on roots. Furthermore, its use improves plant vigor and yield by minimizing nematode-induced stress. When combined with other biocontrol agents, such as beneficial rhizobacteria, M. oleifera further enhances the suppression of root-knot nematodes, providing a synergistic and eco-friendly approach to nematode management in sustainable agriculture.
The current study was, therefore, conducted to evaluate the nematicidal potential of M. oleifera, both individually and in combination with various plant-growth-promoting rhizobacteria. The primary objective was to identify an effective and eco-friendly solution for managing plant-parasitic nematodes, thereby contributing to sustainable agricultural practices.
2. Results
2.1. Molecular Characterization of Rhizobacteria
The molecular characterization of rhizobacterial isolates was conducted using the 16S rRNA gene region amplified with universal primers 27F and 1492R. The amplification successfully yielded amplicons of 1500 base pairs, as visualized in gel electrophoresis (Figure 1). This fragment size corresponds to the expected region of the 16S rRNA gene, widely used for bacterial identification and phylogenetic studies.
The sequencing data obtained from the amplified products were compared against the National Center for Biotechnology Information (NCBI) database using the BLAST algorithm to confirm the identity of the isolates at the species level. The rhizobacterial isolates, along with their respective GenBank accession numbers, are detailed in Table 1. The high sequence similarity (>99%) with reference sequences in the database validated the taxonomic assignment of the isolates.
2.2. Effect of Rhizobacteria and Plant Extracts on Plant Growth of Tomato
The effect of different treatments of M. oleifera, both singly and in combination, was evaluated against M. incognita on plant growth parameters (plant height, root length, fresh shoot weight, dry shoot weight, fresh root weight, dry root weight) and M. incognita infection parameters (root gall ratings, eggs per root system, J2/60cc soil, reproduction factor).
Plant height and root length were significantly increased in nearly all treatments compared to the control (Figure 2). The maximum plant height and root length were recorded as 96.94 cm and 30.48 cm, respectively, in T3 (BA-LWD73 plus M. oleifera), while the lowest plant height and root length (63.50 and 22.44 cm, respectively) were observed in T7 (BT-WAG41 plus M. oleifera).
For fresh shoot weight and dry shoot weight, the maximum fresh shoot weight and dry shoot weight were observed in T2, which is the single treatment of BA-LWD73, with values of 45.43 g and 9.13 g, respectively. The lowest fresh shoot weight was recorded in T7 (BT-WAG41 and M. oleifera combined treatment) at 25.80 g, while the lowest dry shoot weight (5.93 g) was recorded in both T6 and T7 (BT-WAG41 single and combined with M. oleifera).
Fresh root weight and dry root weight were significantly lower in all treatments compared to the control. The minimum fresh root weight was 8.40 g in T1, which is the single treatment of M. oleifera. The minimum dry root weight was 5.83 g in T7, which is the combined treatment of BT-WAG41 and M. oleifera (Figure 2).
2.3. Effect of Rhizobacteria and Plant Extracts on M. incognita Infection
Root gall ratings were significantly lower in most of the treatments compared to the positive control (Figure 3). The minimum root gall rating was 2.67 in T3, which is the combined treatment of BA-LWD73 and M. oleifera, while T8 (M. incognita only, control) showed the highest root gall rating. The single and combined applications of M. oleifera and BA-LWD73 resulted in 11,500, 18,000, and 16,500 eggs per root system, respectively. The lowest numbers of second-stage juveniles were recovered from the single and combined applications of M. oleifera and BA-LWD73 (746.83, 884.33, and 680 J2/60 cm³ of soil, respectively). Similarly, the lowest reproduction factors were observed in the single and combined applications of M. oleifera and BA-LWD73, with values of 3.35, 4.84, and 4.25, respectively (Figure 3).
2.4. FTIR and GC-MS Analysis of Aqueous Extracts of M. oleifera
The FTIR spectra of M. oleifera are shown in Figure 4. The first and second peaks observed in the spectra were at wavelengths 2916.36 cm⁻¹ and 2848.86 cm⁻¹, indicating strong and broad stretching vibrations of hydroxyl groups, predominantly associated with alcohols. The third wavenumber, observed at 1620.20 cm⁻¹, exhibited medium stretching and predominantly indicated the presence of a C=C functional group, typical of alkenes. The fourth wavenumber, at 1415.53 cm⁻¹, showed strong stretching vibrations primarily due to S=O bonds, indicating the presence of sulfates. The fifth wavenumber, ranging from 1315.45 cm⁻¹ to 1022.27 cm⁻¹, exhibited strong stretching vibrations attributable to C-O functional groups, predominantly signifying tertiary alcohols. The final wavenumber observed at 513.06 cm⁻¹ was characteristic of halo compounds.
The GC-MS chromatogram of the aqueous extract of M. oleifera revealed six distinct peaks, identified as six phytocompounds through comparison with the NIST library: 4(1H)-Pyrimidinone (3.02%), 2,6-dimethyl, Acetic acid [(aminocarbonyl)amino]oxo (3.01%), Maltol (9.58%), 4H-Pyran-4-one, 2,3-dihydro-3,5-dihydroxy-6-methyl (10.30%), 5-Hydroxymethylfurfural (71.76%), and 4(1H)-Pyrimidinone, 2-(methylthio) (2.32%) (Table 2). All of these phytocompounds are reported to exhibit antimicrobial and nematicidal properties.
2.5. Detection of VOCs Using SPME-GCMS
3. Discussion
Numerous plant species possess nematicidal constituents that suppress nematode populations and improve plant health. In this study, the aqueous extract of Moringa oleifera, alone and in combination with rhizobacterial strains, effectively reduced Meloidogyne incognita populations and enhanced plant growth. Treatments combining M. oleifera and Bacillus australimaris strain LWD-73 demonstrated superior performance compared to other treatments, significantly reducing nematode infection and promoting plant vigor. All parts of M. oleifera can be utilized as biopesticides due to their ability to suppress pathogens and enhance crop health [20,21].
M. oleifera contains diverse bioactive compounds, including aldehydes, flavonoids, alcohols, phenols, and terpenoids, which may act individually or synergistically to affect nematodes. These compounds disrupt nematode feeding and reproduction, inhibit egg hatching, and exhibit juvenile toxicity. However, the precise mechanisms underlying their nematicidal activity remain unclear [20,21].
Microbes also play a pivotal role in managing polyphagous root-knot nematodes in various crops and soil conditions. They enhance plant health, induce systemic resistance, and combat a wide range of biotic stresses. Rhizobacteria are particularly valuable for their contributions to nutrient uptake, phytohormone production, mitigation of heavy metal stress, and increased crop yields [22,23]. Certain bacterial volatile organic compounds (VOCs), such as dimethyl disulfide, 2-nonanone, 1H-indole, tetradecanol, and 9-hexadecenoic acid, exhibit strong nematicidal activity against RKN [22,24,25].
FTIR and GC-MS analyses of M. oleifera extracts confirmed the presence of flavonoids, alcohols, and phenols, which are known for their antimicrobial and nematicidal properties. The nematicidal activity of M. oleifera was positively correlated with the abundance of these phytochemicals [26,27].
Among the rhizobacteria tested, B. australimaris strain LWD-73 demonstrated the most potent effects against M. incognita. SPME-GC-MS analysis identified key VOCs produced by B. australimaris, including 2-nonanone, 9-hexadecenoic acid, tetradecanol, and 1H-indole, which possess strong antimicrobial, antifungal, and nematicidal properties. For instance, 2-nonanone exhibits significant nematicidal activity by inhibiting egg hatching, disrupting feeding, and reducing nematode populations, thereby limiting root damage over time [28,29]. This compound also shows antifungal activity against pathogens such as Verticillium longisporum and Botrytis cinerea [30,31].
Indole, another prominent VOC, disrupts nematode egg-laying and survival, induces oxidative stress at high concentrations, and triggers methuosis at low concentrations, ultimately causing nematode mortality [32,33]. Moreover, indole acts as a key chemical signal, promoting plant growth and influencing auxin signaling. For example, indole emitted by Escherichia coli has been shown to enhance the root architecture of Arabidopsis thaliana [25,34,35].
9-Hexadecenoic acid (palmitoleic acid) exhibits nematicidal activity by causing toxicity to larvae and eggs, acting as a repellent, and interfering with nematode feeding and reproduction. It may also stimulate plant defenses and influence soil microbial communities, offering a promising approach to integrated nematode management [36].
Tetradecanol, emitted by Paenibacillus polymyxa strain J2-4, has demonstrated strong fumigant activity against M. incognita, further highlighting the potential of VOCs in nematode management strategies [37]. These findings underscore the significance of plant-derived phytochemicals and microbial VOCs as sustainable tools for controlling nematodes and improving crop health.
4. Materials and Methods
4.1. Inoculum Preparation of Nematode
The Meloidogyne incognita culture was initiated from a single egg mass on a susceptible tomato host, cv. HM1824, and identified as M. incognita based on perineal pattern morphology [38]. Small pieces of infected roots were vigorously shaken in a tightly sealed container with 0.5% sodium hypochlorite (NaOCl) solution to release the eggs from the egg masses. The resulting suspension was filtered through a 200-mesh sieve to remove root debris and then through a 500-mesh sieve to collect the eggs, following the described method. The inoculum density was quantified. The eggs were rinsed three times with tap water to remove residual bleach. Freshly hatched second-stage juveniles were obtained by incubating the eggs in extraction trays at 25°C for 48 hours. These juveniles were used for the experiment.
4.2. Preparation of Aqueous Extracts of M. oleifera
Fresh leaves of M. oleifera were carefully washed, blended in a Waring blender with sterile water, and left to stand for 12 hours. The mixture was then filtered sequentially: first through muslin cloth, followed by Whatman filter paper No. 1, and finally through a Millipore filter. Different concentrations (0% to 100%) of the extract were prepared by diluting the standard extract with the requisite amount of distilled water [21].
4.3. Isolation and Purification of Rhizobacteria
Soil samples were collected from the rhizosphere of tomato plants by carefully uprooting the plants and gently shaking the roots to remove excess soil. The rhizobacteria were recovered using the serial dilution method [26]. Dilutions of 10-5, 10-6, and 10-7 were spread onto sterile Petri plates containing solidified autoclaved nutrient agar (NA). The Petri plates were sealed with parafilm and incubated at 26 ± 2°C. After 24 hours, individual bacterial colonies were purified on NA using the streaking method.
4.4. Molecular Characterization of Rhizobacteria Isolates
Genomic DNA was extracted from rhizobacterial isolates using a DNA extraction kit. The extracted DNA was then used for Polymerase Chain Reaction (PCR) analysis. For amplification, the universal primers 27F (5′-AGAGTTTGATCMTGGCTCAG-3′) and 1492R (5′-GGTTACCTTGTTACGACTT-3′), targeting the 16S rRNA molecular marker, were employed [19]. PCR amplification was conducted under the following conditions: initial denaturation at 94°C for 3 min; followed by 35 cycles of 94 °C for 40 s, 60°C for 50 s, and 72°C for 1 min; with a final extension at 72°C for 10 min
The DNA bands were visualized under UV light, and images were captured using a gel documentation system. The amplified DNA was purified using a Qiagen DNA purification kit and quantified with a NanoDrop spectrophotometer. Finally, the amplified products were sent to Eurofins Genomics, USA, for sequencing.
4.5. Sub-Culturing and Preparation of Rhizobacteria Suspensions
The identified rhizobacterial strains were initially cultured on NA plates and subsequently transferred to Luria Bertani broth. The cultures were incubated at 25°C in a rotary shaker for 48 hours to facilitate mass culturing. Cell suspensions of the rhizobacterial strains were prepared by measuring optical density (OD) at 600 nm using a spectrophotometer. The OD was adjusted to 1.0, equivalent to 109 CFU/mL. The cell suspensions were centrifuged at 5000 rpm for 15 minutes at 4°C, after which the supernatant was discarded. The resulting pellet was gently washed with autoclaved distilled water. Following the wash, additional autoclaved distilled water was added to the falcon tube, and the contents were mixed thoroughly using a vortex mixer. The final concentration of each rhizobacterial suspension was adjusted to 109 CFU/mL using a spectrophotometer [39].
4.6. Effect of Rhizobacteria and Plant Extracts on Plant Growth and M. incognita Infection
The soil drench method was employed to assess the effects of rhizobacteria and plant extracts on plant growth parameters and M. incognita infection. To inoculate the plants, 5 mL of suspension containing 5000 freshly hatched second-stage juveniles of M. incognita was applied to 21-day-old tomato plants by creating three small holes around each plant. After 48 hours, 50 mL of rhizobacterial suspension (109 CFU/mL) and 50 mL of an aqueous extract of M. oleifera were applied as a soil drench. Six replicates were conducted for each treatment, with water serving as the control. For the control treatment, 5 mL of M. incognita filtrate was applied to ensure a consistent environment for all plants.
After 45 days, the plants were uprooted to evaluate growth and infection parameters. Plant growth parameters included plant height, tap root weight, dry root weight, shoot length, fresh shoot weight, dry shoot weight, and root length. The M. incognita infection parameters assessed were root gall ratings, eggs per root system, J2 population per 100 cc of soil, and the nematode reproduction factor.
4.7. Detection of Volatile Organic Compounds from Rhizobacteria
B. australimaris LWD73 was cultured overnight at 37°C in LB broth. The bacterial culture was adjusted to an optical density (OD) of 0.132 at 600 nm (equivalent to 0.5 McFarland units) using a spectrophotometer. A 5 mL aliquot of the bacterial culture was transferred to a headspace (HS) vial, and an SPME fiber was inserted into the vial. The HS vial containing the SPME fiber was placed in a water bath at 60 °C for 60 minutes. After incubation, the SPME fiber was immediately exposed to the hot GC injection port for 28 minutes. Each treatment was replicated nine times [40].
4.8. Compound Identification from Plant Extracts
4.9. Statistical Analysis
The data obtained from egg hatch inhibition and juvenile mortality was analyzed by 3-factor factorial in CRD and the data obtained from plant growth parameters and infection parameters was analyzed by 2-factor factorial in CRD. The means were separated by Fischer’s unprotected LSD test, Gen Stat statistical software was used at significance level (P<0.05).
5. Conclusions
The study identified a new bacterial strain, Bacillus australimaris strain LWD-73, with significant nematicidal activity, offering potential for the effective management of Meloidogyne incognita. The integration of this bacterial strain with moringa leaf extract has demonstrated compatibility, enhancing the efficacy of rhizobacteria for nematode control. This combination has yielded improved results in reducing nematode populations. Both moringa and the selected rhizobacteria are rich in phytochemicals with potent nematicidal and antimicrobial properties. These bioactive compounds provide promising solutions for nematode management and can be incorporated into integrated pest management strategies. Small-scale vegetable growers can benefit from using aqueous extracts of moringa leaves as a cost-effective and accessible method for nematode control, contributing to the improved health and productivity of vegetable crops.
Acknowledgments
This research was funded by the higher education commission (HEC) of Pakistan’s “Indigenous Ph.D. Fellowship Program” and the “International Research Support Initiative Program” (IRSIP). Special thanks to Prof. Dr. Larry W. Duncan for funding and facilitating during the IRSIP period at CREC, University of Florida (USA).
References
- Khairy, D. , Refaei, A, and Mostafa, F.: ‘Management of Meloidogyne incognita infecting eggplant using moringa extracts, vermicompost, and two commercial bio-products’. Egyptian J. Agronema., 2021, 20, 1–16. [Google Scholar]
- Saeed, M. , Mukhtar, T. Root-knot nematodes (Meloidogyne spp.) infecting peach (Prunus persica L.) in the Pothwar region of Pakistan. Journal of Agricultural Science and Technology 2024, 26, 897–908. [Google Scholar]
- Yaseen, I. and Mukhtar, TImpact of sequential and concurrent inoculations of Meloidogyne incognita and Fusarium oxysporum f. sp. vasinfectum on the growth performance of diverse okra cultivars. Plant Protection 2024, 8, 303–313. [Google Scholar]
- Yaseen, I. , Mukhtar, T, Kim, H.-T. and Arshad, B. Interactive effects of Meloidogyne incognita and Fusarium oxysporum f.sp. vasinfectum on okra cultivars. Bragantia 2024, 83, e20230266. [Google Scholar] [CrossRef]
- Afzal, A. and Mukhtar, TRevolutionizing nematode management to achieve global food security goals-An overview. Heliyon 2024, 10, e25325. [Google Scholar] [CrossRef]
- Ayub, H.M. , Akram, M, Mehak, A., Ajmal, M., Ilyas, I., Seerat, W., Tatar, M., Ali, A., Sarwar, R., Abbasi, M. and Rahman, A. A preliminary study on the interaction between Meloidogyne incognita and some strains of Pseudomonas son growth performance of tomato under greenhouse conditions. Plant Protection, 2024, 8, 341–350. [Google Scholar]
- Shahid, M. , Gowen, SR., Burhan, M., Niaz, M.Z., Anwar-ul-Haq, M. and Mehmood, K. Differential responses of Meloidogyne sto Pasteuria isolates over crop cycles. Plant Protection 2024, 8, 257–267. [Google Scholar]
- Aslam, N. , Atiq, M, Rajput, N.A., Akram, A., Arif, A.M., Kachelo, G.A., Nawaz, A., Jahangir, M.M., Jabbar, A. and Ijaz, A. Explicating Botanical Bactericides as an Intervention Tool towards Citrus Canker. Plant Protection, 2024, 8, 25–32. [Google Scholar]
- Atiq, M. , Malik, L, Rajput, N.A., Shabbir, M., Ashiq, M., Matloob, M.J., Waseem, A., Jabbar, A., Ullah, A., Zawar, R., Majeed, A. Evaluating the efficacy of plant-mediated copper-silver nanoparticles for controlling Cercospora leaf spot in mung beans. Plant Protection 2024, 8, 661–670. [Google Scholar]
- Hassan, A. , Abbas, H, Abbasi, W.M., u Rahman, M.S., Akram, W., Rafiq, F. and Murad, M.T. Evaluation of rhizospheric Pseudomonas sfor the management of Fusarium wilt of tomato in Cholistan, Pakistan. Plant Protection 2024, 8, 433–445. [Google Scholar]
- Manzoor, F. , Atiq, M, Aleem, M., Naveed, K., Khan, N.A., Kachelo, G.A., Ali, M.U., Ali, F., Haris, M. and Rajput, N.A. Appraisal of antifungal potential of chemicals and plant extracts against brown leaf spot of soybean caused by Septoria glycine. Plant Protection 2024, 8, 447–455. [Google Scholar]
- Taha, Z.R. , Altaai, AF., Mohammad, T.H., Khajeek, T.R. Efficacy of silver nanoparticles from Fusarium solani and mycorrhizal inoculation for biological control of Fusarium wilt in tomato. Plant Protection 2024, 8, 635–648. [Google Scholar]
- Saeed, M. , Mukhtar, T., and Khan, M.A. Assessment of nematicidal potential of Cannabis sativa and Azadirachta indica in the management of root-knot nematode (Meloidogyne javanica) on peach’, Pak. J. Agric. Sci. 2021, 58. [Google Scholar]
- Azeem, W.; Mukhtar, T.; Hamid, T. Evaluation of Trichoderma harzianum and Azadirachta indica in the management of Meloidogyne incognita in Tomato’. Pak. J. Zoo. 2021, 53, 119–125. [Google Scholar] [CrossRef]
- Khandagale, P. , Kansara, S, Padsala, J., and Patel, P.: ‘Plant Growth Promoting Rhizobacteria for Sustainable Production of Sugarcane and Rice’, Int. J. Plant Sci., 2024, 36, 298–305. [Google Scholar]
- Zhang, L.-N. , Jiang, C-H., Si, F., Song, N., Yang, W., Zhu, Y., Luo, Y., and Guo, J.-H.: ‘Long-term field application of a plant growth-promoting rhizobacterial consortium suppressed root-knot disease by shaping the rhizosphere microbiota’, Plant Dis., 2024, 108, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Abbas, M. , Saadedin, S, Suleiman, A. Cloning and overexpression of Zea mays cystatin 2 (ccii) gene in Bacillus subtilis to reduce root-knot nematode infection in cucumber plants in Iraq. Plant Protection, 2024, 8, 621–633. [Google Scholar] [CrossRef]
- Zhang, L.-N. , Jiang, C-H., Si, F., Song, N., Yang, W., Zhu, Y., Luo, Y., and Guo, J.-H.: ‘Long-term field application of a plant growth-promoting rhizobacterial consortium suppressed root-knot disease by shaping the rhizosphere microbiota’, Plant Dis., 2024, 108, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S. , Ramanjini Gowda, P, Saikia, B., Debbarma, J., Velmurugan, N., and Chikkaputtaiah, C.: ‘Screening of tomato genotypes against bacterial wilt (Ralstonia solanacearum) and validation of resistance linked DNA markers’, Australasian Plant Pathology, 2018, 47, 365–374. [Google Scholar] [CrossRef]
- Yaseen, A. , and Hájos, MT.: ‘Study on moringa tree (Moringa oleifera Lam.) leaf extract in organic vegetable production: A review’, Research on Crops, 2020, 21, 402–414. [Google Scholar]
- Mukhtar, T. , Kayani, MZ., and Hussain, M.A.: ‘Nematicidal activities of Cannabis sativa L. and Zanthoxylum alatum Roxb. against Meloidogyne incognita’, Ind. Crops Prod., 2013, 42, 447–453. [Google Scholar] [CrossRef]
- Bhat, A.A. , Shakeel, A, Waqar, S., Handoo, Z.A., and Khan, A.A.: ‘Microbes vs. nematodes: Insights into biocontrol through antagonistic organisms to control root-knot nematodes’, Plants, 2023, 12, 451. [Google Scholar] [CrossRef] [PubMed]
- Groover, W. , Held, D, Lawrence, K., and Carson, K.: ‘Plant growth-promoting rhizobacteria: a novel management strategy for Meloidogyne incognita on turfgrass’. Pest Manag. Sci., 2020, 76, 3127–3138. [Google Scholar] [CrossRef] [PubMed]
- Agisha, V.N. , Kumar, A, Eapen, S.J., Sheoran, N., and Suseelabhai, R.: ‘Broad-spectrum antimicrobial activity of volatile organic compounds from endophytic Pseudomonas putida BP25 against diverse plant pathogens’. Biocontrol Sci. Technol., 2019, 29, 1069–1089. [Google Scholar] [CrossRef]
- Bhattacharyya, D. , Garladinne, M, and Lee, Y.H.: ‘Volatile indole produced by rhizobacterium Proteus vulgaris JBLS202 stimulates growth of Arabidopsis thaliana through auxin, cytokinin, and brassinosteroid pathways’, J. Plant Growth Regul., 2015, 34, 158–168. [Google Scholar] [CrossRef]
- Alam, E.A. , and Nuby, AS.E.: ‘Phytochemical and nematicidal screening on some extracts of different plant parts of egyptian Moringa oleifera L.’, Pak. J. Phytopath., 2022, 34, 293–306. [Google Scholar] [CrossRef]
- Anekwe, I.I. , Ifenyinwa, CC., Amadi, E.S., Ursulla, N.N., and Chukwuebuka, I.F.: ‘Characterization of Aqueous Extract of Moringa oleifera Leaves using GC-MS Analysis’. J. Adv. Microbiol., 2023, 23, 8–18. [Google Scholar] [CrossRef]
- Cheng, W. , Yang, J, Nie, Q., Huang, D., Yu, C., Zheng, L., Cai, M., Thomashow, L.S., Weller, D.M., and Yu, Z.: ‘Volatile organic compounds from Paenibacillus polymyxa KM2501-1 control Meloidogyne incognita by multiple strategies’. Sci. Rep., 2017, 7, 16213. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y. , Xu, C, Ma, L., Zhang, K., Duan, C., and Mo, M.: ‘Characterisation of volatiles produced from Bacillus megaterium YFM3. 25 and their nematicidal activity against Meloidogyne incognita’. Eur. J. Plant Pathol., 2010, 126, 417–422. [Google Scholar] [CrossRef]
- Rybakova, D. , Rack-Wetzlinger, U, Cernava, T., Schaefer, A., Schmuck, M., and Berg, G.: ‘Aerial warfare: a volatile dialogue between the plant pathogen Verticillium longisporum and its antagonist Paenibacillus polymyxa’. Front. Plant Sci., 2017, 8, 278579. [Google Scholar] [CrossRef]
- Huang, R. , Li, G, Zhang, J., Yang, L., Che, H., Jiang, D., and Huang, H.: ‘Control of postharvest Botrytis fruit rot of strawberry by volatile organic compounds of Candida intermedia’, Phytopathology, 2011, 101, 859–869. [Google Scholar] [CrossRef] [PubMed]
- Bommarius, B. , Anyanful, A, Izrayelit, Y., Bhatt, S., Cartwright, E., Wang, W., Swimm, A.I., Benian, G.M., Schroeder, F.C., and Kalman, D.: ‘A family of indoles regulate virulence and Shiga toxin production in pathogenic E. coli’, PLoS One, 2013, 8, e54456. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H. , Kim, YG., Kim, M., Kim, E., Choi, H., Kim, Y., and Lee, J.: ‘Indole-associated predator–prey interactions between the nematode Caenorhabditis elegans and bacteria’, Environ. Microbiol., 2017, 19, 1776–1790. [Google Scholar] [CrossRef] [PubMed]
- Elzey, B. , Pollard, D , and Fakayode, S.O.: ‘Determination of adulterated neem and flaxseed oil compositions by FTIR spectroscopy and multivariate regression analysis’. Food Control, 2016, 68, 303–309. [Google Scholar] [CrossRef]
- Ahmed, D.; Shahab, S.; Ahmad, M.; Pathak, N. Mahua Oilseed Cake: Chemical Compounds and Nematicidal Potential’: ‘Oilseed Cake for Nematode Management’ (CRC Press, 2024), 81-88.
- Wollum, A. ‘Cultural methods for soil microorganisms’, Methods of soil analysis: part 2 chemical and microbiological properties. 1982, 9, 781–802. [Google Scholar]
- Nikoo, F.S. , Sahebani, N., Aminian, H., Mokhtarnejad, L., and Ghaderi, R.: ‘Induction of systemic resistance and defense-related enzymes in tomato plants using Pseudomonas fluorescens CHAO and salicylic acid against root-knot nematode Meloidogyne javanica’, J. Plant Prot. Res. 2014. [Google Scholar]
- Taylor, D.P.; Netscher, C. ‘An improved technique for preparing perineal patterns of Meloidogyne spp’, Nematologica 1974, 20, 268–269. [Google Scholar] [CrossRef]
- Nikoo, F.S.; Sahebani, N., Aminian; Ghaderi, R. Induction of systemic resistance and defense-related enzymes in tomato plants using Pseudomonas fluorescens CHAO and salicylic acid against root-knot nematode Meloidogyne javanica. J. Plant Prot. Res 2014. [Google Scholar]
- Tait, E.; Perry, J.D., Stanforth. Identification of volatile organic compounds produced by bacteria using HS-SPME-GC–MS. J. Chromatogr. Sci. 2014, 52, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Ramanjini Gowda, P.; Saikia, B., Debbarma; Chikkaputtaiah, C. Screening of tomato genotypes against bacterial wilt (Ralstonia solanacearum) and validation of resistance linked DNA markers. Australasian Plant Pathology 2018, 47, 365–374. [Google Scholar] [CrossRef]
Figure 1.
Molecular validation of rhizobacterial isolates using 16S rRNA Primers. Lane 1-8 represents the rhizobacterial isolates, 9 as control and L represents 1kb Ladder.
Figure 1.
Molecular validation of rhizobacterial isolates using 16S rRNA Primers. Lane 1-8 represents the rhizobacterial isolates, 9 as control and L represents 1kb Ladder.
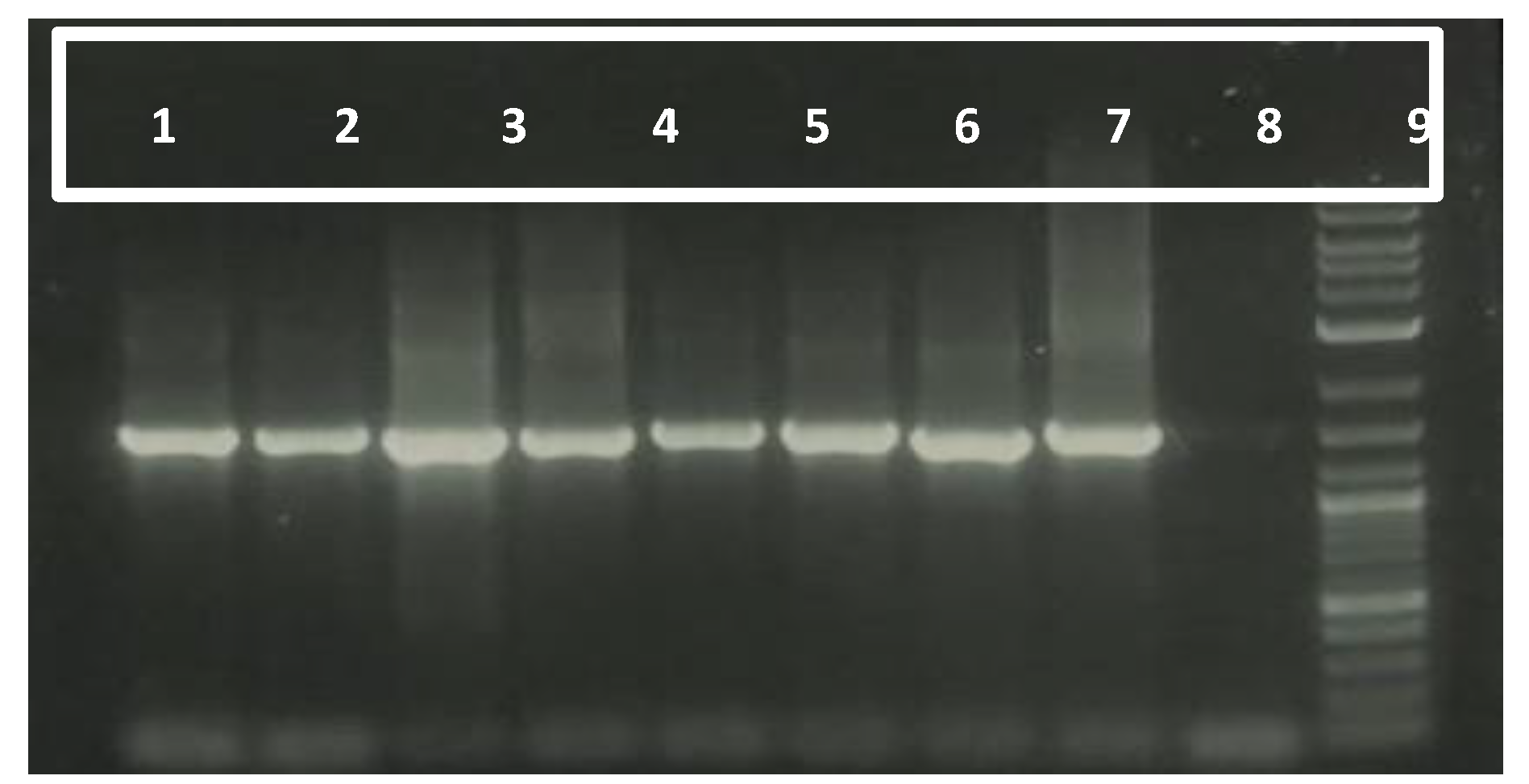
Figure 2.
Effect of M. oleifera and rhizobacteria on Plant Growth Parameters of Tomato: (a) Plant Height; (b) Fresh Shoot Weight; (c) Dry Shoot Weight; (d) Fresh Root Weight; (e) Dry Root Weight and (f) Root Length.
Figure 2.
Effect of M. oleifera and rhizobacteria on Plant Growth Parameters of Tomato: (a) Plant Height; (b) Fresh Shoot Weight; (c) Dry Shoot Weight; (d) Fresh Root Weight; (e) Dry Root Weight and (f) Root Length.
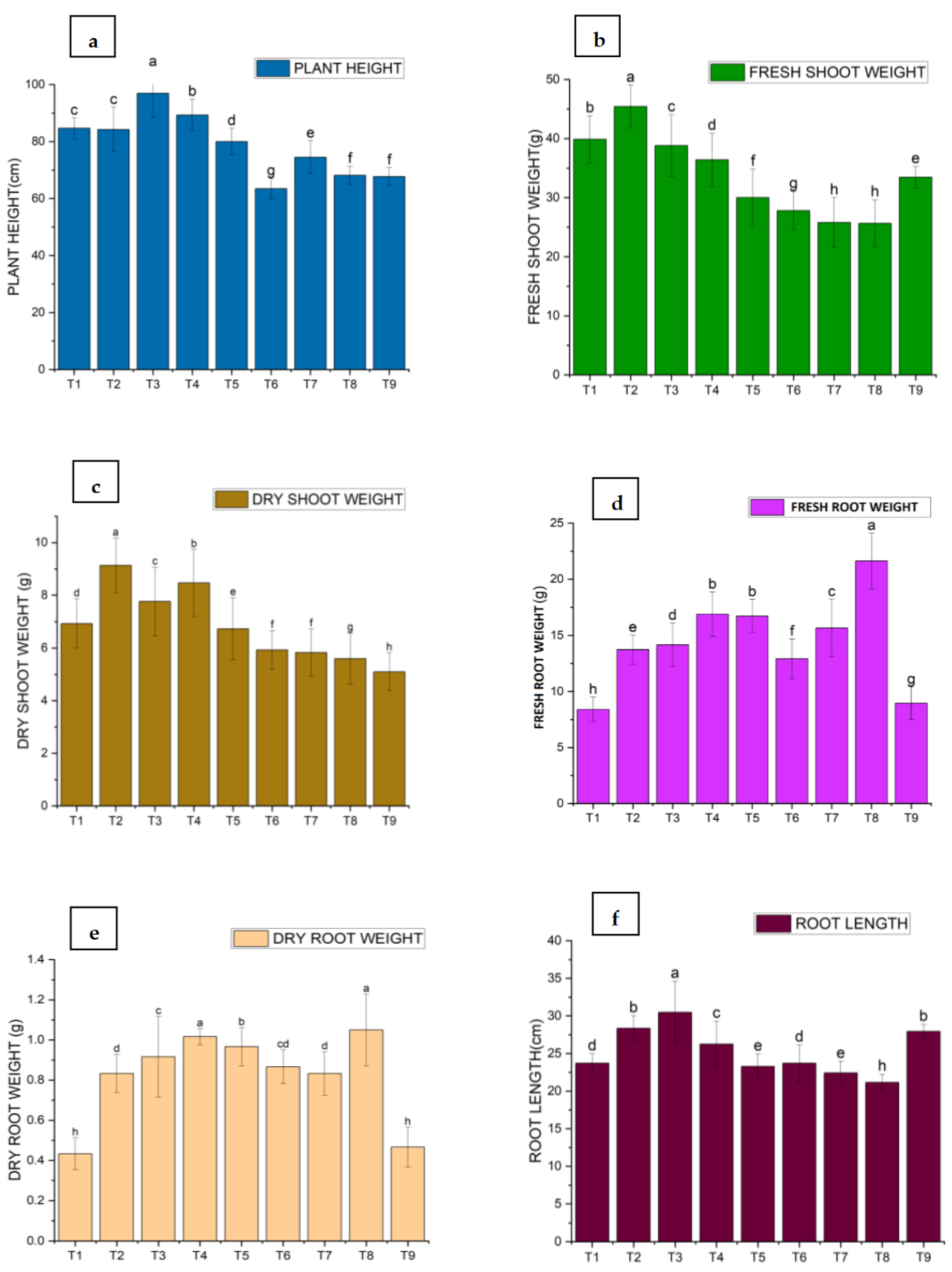
Figure 3.
Effect of M. oleifera and rhizobacteria on Infection parameters of M. incognita: (a) Root Gall Ratings; (b) Eggs per Root System; (c) J2/60cc Soil and (d) Reproduction Factor. There was no nematode infection in T9.
Figure 3.
Effect of M. oleifera and rhizobacteria on Infection parameters of M. incognita: (a) Root Gall Ratings; (b) Eggs per Root System; (c) J2/60cc Soil and (d) Reproduction Factor. There was no nematode infection in T9.
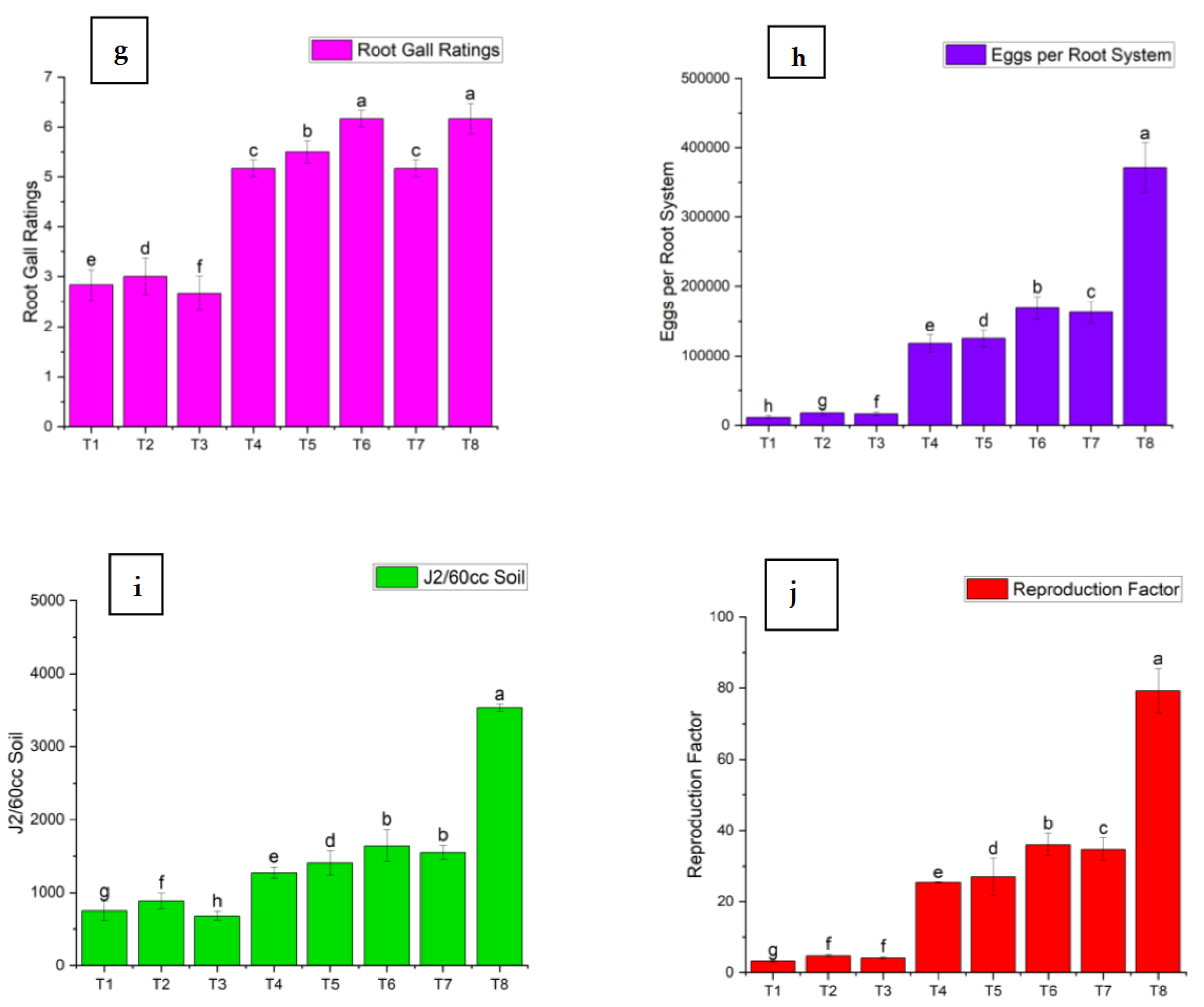
Figure 4.
FTIR Spectra of M. oleifera leaves.
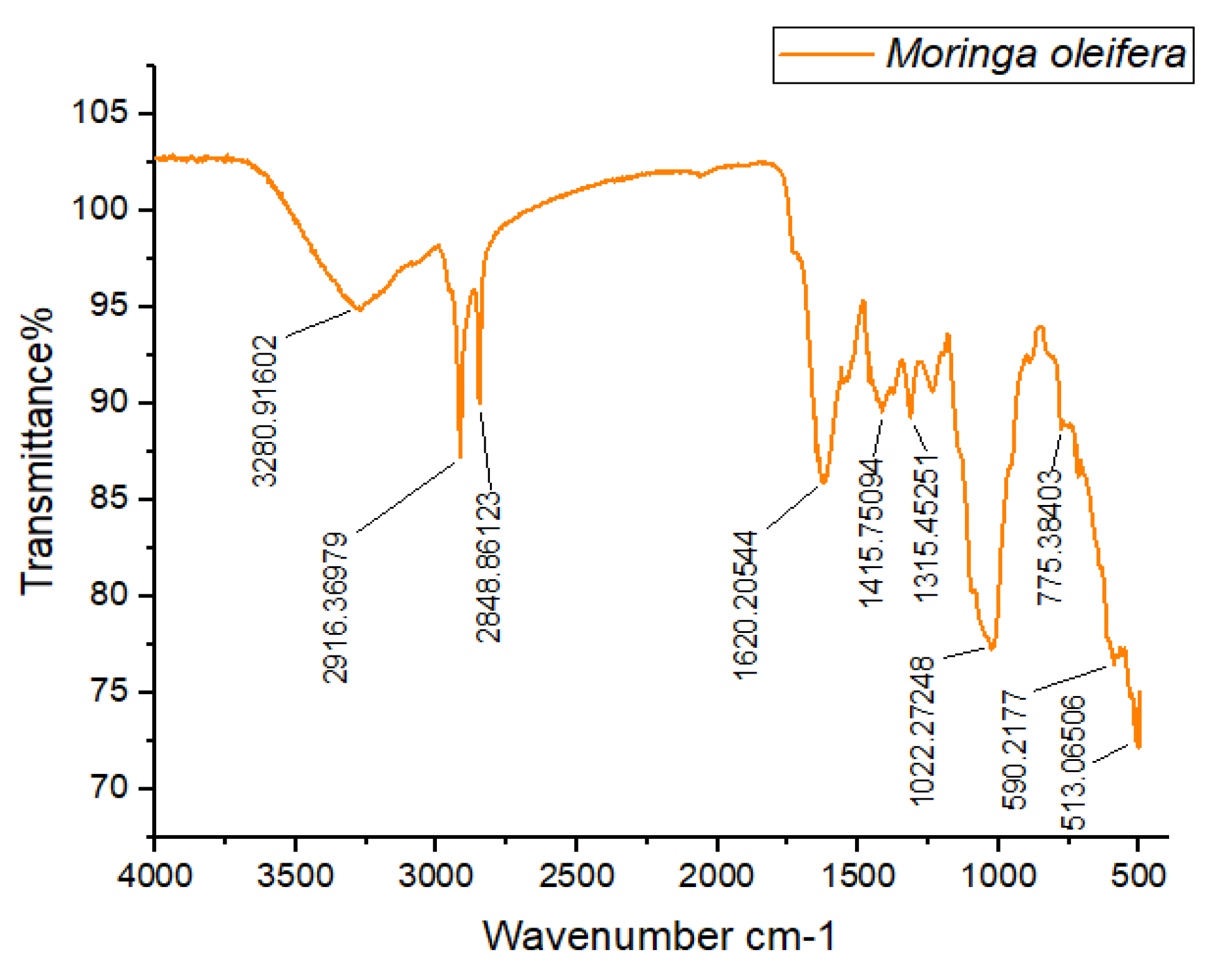
Figure 5.
HS-SPME GC-MS chromatogram profiles of VOCs emitted from B. australimaris LWD-73.
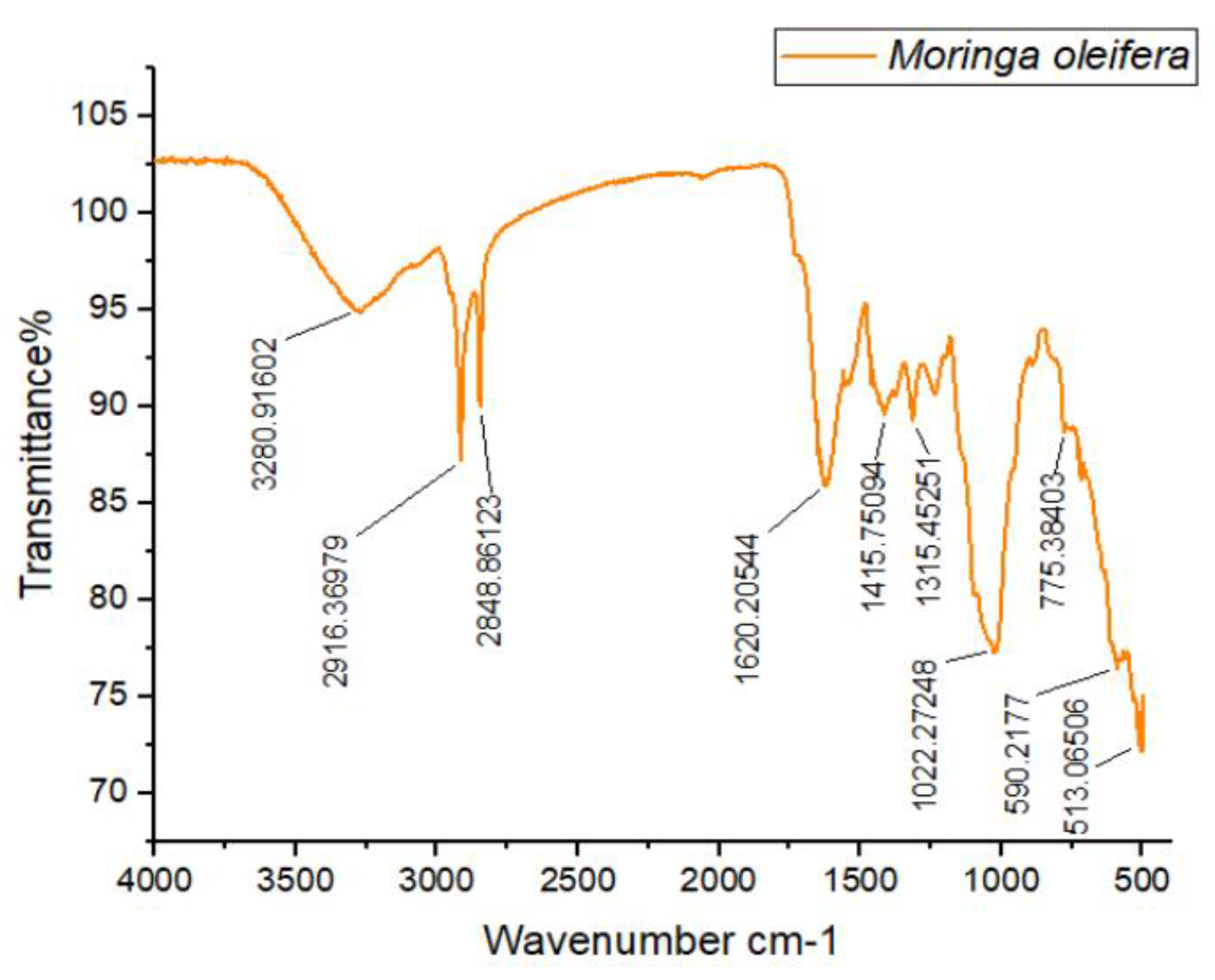

Table 1.
Rhizobacteria Identity with Accession Numbers.
| Sr. No. | Isolate | Rhizobacteria Identified as | Accesion No. |
|---|---|---|---|
| 1 | W2 | Bacillus australimaris strain LWD73 | OQ366704 |
| 2 | W13 | B. cereus strain HR001 | OQ372951 |
| 3 | W44 | B. thuringiensis strain WAG41 | OQ370579 |
Table 2.
Compounds identified from aqueous extract of M. oleifera L. through GCMS analysis.
| Peak No. | Compound Name | Molecular Formula | Molecular Weight (g/mol) | Retention Time (Min) | Peak Area (%) |
|---|---|---|---|---|---|
| 1 | 4(1H)-Pyrimidinone, 2,6-dimethyl | C24H30N2O | 362.5 | 5.310 | 3.02 |
| 2 | Acetic acid, [(aminocarbonyl)amino]oxo | C3H4N2O4 | 132.08 | 5.561 | 3.01 |
| 3 | Maltol | C6H6O3 | 126.11 | 5.705 | 9.58 |
| 4 | 4H-Pyran-4-one, 2,3-dihydro-3,5-dihydroxy-6-methyl | C6H8O4 | 144.12 | 6.155 | 10.30 |
| 5 | 5-Hydroxymethylfurfural | C6H6O3 | 126.11 | 7.636 | 71.76 |
| 6 | 4(1H)-Pyrimidinone, 2-(methylthio)- | C5H6N2OS | 142.18 | 11.364 | 2.32 |
Table 3.
Volatile Organic Compounds of B. australimaris strain LWD-73 detected by solid phase micro extraction gas chromatography.
Table 3.
Volatile Organic Compounds of B. australimaris strain LWD-73 detected by solid phase micro extraction gas chromatography.
| No. | Compound | Molecular Formula | Retention Time (min) | Molecular Weight (g/mol) |
Peak Area | Percent |
|---|---|---|---|---|---|---|
| 1 | 2-Nonanone | C9H18O | 8.35 | 142.24 | 215618130 | 6.54 |
| 2 | 1H-Indole | C8H7N | 11.50 | 117.15 | 2882910162 | 87.46 |
| 3 | Tetradecanol | C14H30O | 13.73 | 214.38 | 32564023 | 0.99 |
| 4 | 9-Hexadecenoic acid | C₁₆H₃₂O₂ | 19.00 | 256.42 | 165322864 | 5.02 |
Table 4.
Treatments for the Effect of Rhizobacteria and Moringa Leaf Extracts on Plant Growth Attributes and M. incognita infection.
Table 4.
Treatments for the Effect of Rhizobacteria and Moringa Leaf Extracts on Plant Growth Attributes and M. incognita infection.
| T1 | M. oleifera + M. incognita |
| T2 | Bacillus aurtralimaris LWD73 + M. incognita |
| T3 | B. aurtralimaris LWD73 + M. oleifera + M. incognita |
| T4 | Bacillus cereus HR001+ M. incognita |
| T5 | B. cereus HR001 + M. oleifera + M. incognita |
| T6 | Bacillus thuringiensis WAG41 + M. incognita |
| T7 | B. thuringiensis WAG41 + M. oleifera + M. incognita |
| T8 | M. incognita only |
| T9 | Healthy Control |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



