Submitted:
09 January 2025
Posted:
10 January 2025
You are already at the latest version
Abstract
Sex steroids and brain-derived neurotrophic factor (BDNF) participate in neural tissue formation, phenotypic differentiation, and neuroplasticity. All these processes are essential for the health and maintenance of the central nervous system. Aim: To elucidate the interaction mechanisms between BDNF and sex steroids in neuronal function. Method: A series of searches were performed using the Mesh terms for androgen/receptor, estrogens/receptors, and BDNF/receptor, and a collection of the scientific data available on PubMed up to February 2022, on the mechanical interactions between BDNF and sex steroids was included in this literature review. Discussion: Sex steroids influence the formation and/or maintenance of neural circuits via different mechanisms, including the regulation of BDNF expression and signaling. Estrogens exert a time- and region-specific effect on BDNF synthesis. The nuclear estrogen receptor can directly regulate BDNF expression, independently of estrogen presence, in neuronal cells; whereas, progesterone and testosterone upregulate BDNF expression via their specific nuclear receptors. In addition, testosterone has a positive effect on BDNF release by glial cells, which lack androgen receptors.
Keywords:
Estrogen
; Estradiol
; Testosterone
; Progesterone
; Brain-derived neurotrophic factor
; Nervous system
1. Introduction
Sex steroid hormones, i.e., androgens, estrogens and progesterone, are well-known for their role in stimulating the reproductive organs and the development of secondary sexual characteristics in female and male [1]. They share similar structures and can reach the brain across the blood-brain barrier, when released from peripheral steroidogenic organs [2], or be synthesized de novo within the brain [3] whereby they regulate neuronal functions. The synthesis of neuroactive steroids occurs primarily in neurons, but also in microglia and astrocytes, and requires the translocation of cholesterol across the mitochondrial membrane through a molecular complex formed by the translocator protein 18 kDa, the steroidogenic acute regulatory protein, the voltage-dependent anion channel protein and the adenine nucleotide transporter protein. Cholesterol is converted to pregnenolone by enzymatic activity in the mitochondria, and pregnenolone diffuses into the cytosol where it is further metabolized by different enzymes into different neuroactive steroids (see Figure 1). [4,5].
In addition to sexual dimorphism, present in several neural circuits, sex steroids generally play a neuroprotective role and impact differently on the brain’s function [6]. Two main effects of the steroid hormones on the brain are described to produce sexual dimorphism in vertebrates: the first is organizational, associated with changes during sensitive phases of development, such as prenatal and/or perinatal sexual differentiation, and adolescence; and the second referring to activational effects associated with sexually dimorphic phenotypic changes during puberty and throughout adulthood [7]. Furthermore, experimental studies show that learning responses are influenced differently by sex steroids in males and females through various mechanisms underlying neuroplasticity, i.e., learning-induced and hormone-driven brain region- and cell type-specific cellular activity and morphology [8]. Exposure of undifferentiated neural stem cells to different sex steroids results in phenotypic changes with alteration in functional and behavioral responses [9,10].
The brain-derived neurotrophic factor (BDNF) is a conserved neurotrophin that participates in multiple processes related to brain development and functioning, i.e., neuronal cell migration and differentiation, axonal growth and synaptogenesis. The actions of sex steroids overlap with those of BDNF in various neural plasticity processes, however, the mechanisms underlaying such interactions are not fully elucidated [11,12,13,14]. Further, a possible time- and region- specificity for sex steroid effects on BDNF activity adds an extra complexity to this topic [15,16].
The presence of estrogen responsive element (ERE) in BDNF gene sequence started the investigations on the participation of sex hormones in the development and organization of the nervous system. The interaction between estrogen and BDNF was initially evidenced by the co-localization of BDNF and TrkB mRNAs in estrogen-sensitive neurons of the developing cerebral cortex [17]. Next, Singh, Meyer, and Simpkins [18] pointed out that estrogens were more effective in maintaining BDNF levels in the hippocampus than in the cortex of female rats. Which supports the thesis that steroids regulation of BDNF expression is influenced by the neurosteroids availability.
Estrogens activation and translocation of the nuclear estrogen receptors (ERs) to cell nuclei can directly regulate the expression of BDNF via its ERE sequence. This can be called as the ‘genomic mechanism’. Additionally, membrane-associated steroid receptors can activate extra nuclear-initiated kinase signaling and modify intracellular mechanisms amplifying steroids’ effect on gene expression [19,20]. For instance, estrogens activation of G-protein coupled membrane ER activates the mitogen-activated protein kinase (MAPK) and phosphoinositide 3-kinase (PI3K) pathways and, as the kinases downstream MAPK and protein kinase (PKA) pathways increase ER phosphorylation, this potentiate estrogens signal [21].
BDNF signaling via tyrosine kinase (Trk) B receptors within neurons membrane elicits various intracellular signaling pathways including MAPK/ extracellular signal-regulated protein kinase (ERK), phospholipase Cγ (PLCγ) and PI3K pathways that induce the synthesis of proteins that regulate neuronal survival and synaptic maintenance [22]. This implies that BDNF function in synaptic plasticity is partially shared with that of estrogen.
In spite of a great number of studies have addressed the estrogen’s influence on BDNF activity, the regulation of BDNF by androgen and progesterone is also seen to participate in neuroplasticity processes in females and males. Progesterone exerts its effects on BDNF through genomic and non-genomic mechanisms. While the induction of BDNF expression requires the nuclear PR, the release of BDNF can be mediated by activation of a distinct membrane-associated PR. Additionally, some membrane-associated PRs may lead to the activation of ERK1/2 which may exert an inhibitory influence on BDNF expression [23].
In light of the above mentioned, in this literature review we collected experimental evidence available up to February 2022 on interactions between sex steroids (estrogens, testosterone, progesterone) and BDNF from studies in vitro, in vivo and ex vivo, and provided a qualitative synthesis of the mechanisms by which sex steroids participate in neuroplasticity via BDNF modulation.
2. Estrogen and BDNF Interactions
Sex steroids hormones regulate BDNF activity in many neuronal circuits and at different phases of the lifespan. They act on brain tissues at different stages of development, specifically during the perinatal sensitive period, and the adolescence to organize sexual differentiation, and to produce sex-specific behaviors throughout the whole life [1]. The scientific data supporting sex steroids interactions with BDNF in neural tissues are going to be discussed in the following subheadings, and a brief summary of findings is displayed in Table 1, Table 2 and Table 3.
A conserved co-regulation of genes such as ER, BDNF and TrkB through different species indicates that the genes have closely associated functions and are part of a same pathway [24,25]. The patterns of these genes expression in developmental estrogen targets neurons were the first evidences that BDNF expression is related to neurons sensitivity to estrogen [17]. In line, Singh et al. [18] showed that ovariectomy (OVX) results in reduction in BDNF levels in cortical tissue. Associations between reductions in ER and BDNF levels have been reported by several studies.
From another perspective, activation of ERs were seen to increase BDNF levels and synaptogenesis in the cerebellum of neonatal mice [26], the pre-frontal cortex of rats [27], and in zebrafish [28]. Such regulatory role of ERs over BDNF result in positive cognitive outcomes [26,27,28]. Indeed, estrogens directly regulate the expression of BDNF in neural cells [29,30,31,32,33,34,35,36,37,38,39]. Additionally, ER can regulate BDNF expression and function via transactivation of TrkB receptors [40,41,42,43,44,45]. In addition to EREs, activation of TrkB receptors also potentiate estrogen’s effects via ERα phosphorylation downstream TrkB/MAPK/ERK pathway. Meanwhile, the activity of the PI3K/AKT pathway constitutively inhibits basal transcription at EREs and inhibits TrkB-dependent transcriptional activation at EREs [46]. This grants an homeostatic equilibrium to an estrogens regulation on BDNF function. For instance, disruption in estrogens regulation of BDNF expression leads to an elevation in TrkB phosphorylation [47,48,49,50].
The functional interaction between estradiol and BDNF participates in activity-dependent dendritogenesis. Murphy, Cole, and Segal [51] detected a short-term decrease in BDNF concentrations in hippocampal neurons treated with estradiol in which BDNF declined up to 24 hr and recovered within 48 hr. This affected the activity-dependent formation of dendritic spines in GABAergic hippocampal neurons.
Zhu et al. [52] investigated whether estradiol regulates BDNF expression in the hypothalamus during the estrous cycle in rats. BDNF expression transiently increase in the ventromedial nucleus of hypothalamus following the estradiol peak. Balasubramanian et al. [53] added that a chronic exposition of the amygdala to estradiol leads to a decrease in BDNF expression levels and behavioral implications. Alternatively, Cavus and Duman [54] had reported that BDNF levels decrease in the dentate gyrus and the medial prefrontal cortex, when estradiol levels are highest during estrus. And that acute estradiol treatment decreased hippocampal BDNF expression in acutely OVX rats, with no effect in chronically OVX rats. The findings indicate that the effect of cells exposition to estradiol on BDNF expression shall be influenced by time and intensity and involve selective mechanisms according to the neuronal population.
Nuclear ER (α and β) are present in cells from multiple brain regions, e.g., hippocampus, hypothalamus, amygdala, and prelimbic cortex [55,56,57,58,59,60,61,62,63]. Activation of estrogen-related receptor γ also regulate BDNF expression in dopaminergic neurons [64]. The study by Wu et al., [65] reported that neither OVX nor estradiol treatment altered BDNF expression in the hippocampus of BDNF Val66Met heterozygote mice. OVX reduced the expression of BDNF only in the hippocampus of wild-type mice and the estradiol-induced BDNF increase was resumed to the dorsal hippocampus of wild type OVX mice. The control of BDNF function requires the processing of the pro-apoptosis pro-BDNF isoform into the mature BDNF. The Val66Met polymorphism negatively affects BDNF expression and processing into secretory pathways, altering the released pro-BDNF/BDNF [66,67]. It might be so that Val66Met heterozygote mice already presented altered hippocampal levels of BDNF in a way that these levels were not substantially changed by OVX or estradiol treatments.
The enzymatic activity of aromatase plays a relevant role in the regulation of BDNF expression by estrogens [68,69]. Aromatase is a membrane-bound enzyme, located in the endoplasmic reticulum in estrogen-producing cells, which catalyzes the desaturation (aromatization) of the ring A of C19 androgens and converts them to C18 estrogens. Dittrich et al. [70] demonstrated that estradiol-induced BDNF expression in the forebrain song control nucleus of male juvenile zebra finches is decreased by the selective aromatase inhibitor fadrozole.
In line, aging importantly effects estrogens regulation on BDNF. Data from Jezierski and Sohrabji [71,72] study reports that estrogen increases the expression of TrkB receptor in the brain of young, but not in older animals. Data also identified that aging impairs estrogens regulation on pro-apoptotic p75 neurotrophic receptor expression, favouring its increase and influencing on neuronal apoptosis. It is possible that the ERE in BDNF gene becomes less sensitive to estrogen with age, and this leads to a loss in BDNF regulation of TrkB receptors. Moreno-Piovano et al. [73] comparison of mouse with short-term vs. long-term OVX concluded that estradiol effect on BDNF expression decays in time. A possible negative feedback of ER expression to long-term exposure to estrogens might contribute to the loss of estrogens effect on BDNF regulation.
From a physiological perspective, exercise was shown to improve estradiol regulation on BDNF expression [74]; and to restore hippocampal levels of BDNF in OVX animals. These preliminary studies report that rats with a 14-day free-wheel access, or running on a treadmill at low-intensity for 30 min during 20 days; or else, combining 3 days of resistance training and 3 days of running for 8 weeks displayed of improvements in the sex steroids regulation on BDNF expression in their hippocampi [75,76,77,78].
3. Progesterone and BDNF Regulation
The early experiments of Gibbs et al., [79,80] pointed that the levels of BDNF fluctuate across the estrous cycle, and increase in response to acute hormones replacement. Murphy et al. [51,81] identified that estradiol transiently reduces BDNF and GABA synthesis in hippocampal cells and, consequently, reduces inhibitory GABAergic connections and leads to a brief enhancement in neuronal activity, after which, BDNF returns to normal levels and the inhibitory tone is restored. They detected that progesterone was able to blocks the increase in CREB phosphorylation and prevent estradiol-induced increase in cell activity and spine density. In line, Aguirre et al., [82] showed that progesterone reverses estradiol-induced increase in ER and BDNF levels, and eliminates estradiol effect against glutamatergic excitotoxicity in hippocampal cultures. It seems that progesterone inhibition on CREB activity has a negative influence on estradiol effects on BDNF in GABAergic neurons. Additionally, Franklin & Perrot-Sinal, [83] reported that young adult female rats treated with estradiol have higher hippocampal BDNF levels than those treated with estradiol and progesterone combination.

Table 2.
Data summary of interactions between Progesterone and BDNF.
| Title | Aim/experiment | Main Findings | |
|---|---|---|---|
| 1 | Treatment with estrogen and progesterone affects relative levels of brain-derived neurotrophic factor mRNA and protein in different regions of the adult rat brain DOI: 10.1016/S0006-8993(99)01880-6 |
To examine the acute effects of estrogen and PROG on levels of BDNF expression and protein in different brain regions in adult mice. | Estrogen or estrogen + PROG increase BDNF expression and protein levels in the pyriform cortex of mice. Increases in BDNF expression in the hippocampus accompanied a decrease in BDNF protein. |
| 2 | Progesterone prevents estrogen-induced dendritic spine formation in cultured hippocampal neurons DOI: 10.1159/000054580 |
To examine PROG effects on estrogen- induced formation of dendritic spines in hippocampal cell cultures. | PROG does not affect the estrogen-induced downregulation of BDNF, but it did block the effect of estrogen on CREB phosphorylation. |
| 3 | Progesterone counteracts estrogen-induced increases in neurotrophins in the aged female rat brain DOI: 10.1097/00001756-200412030-00021 |
To test estrogen and estrogen + PROG effects on neurotrophin levels in cognitive brain regions in aged OVX mice. | Estrogen treatment increased BDNF, NGF, and NT3 levels in the mice entorhinal cortex, and PROG abated these effects dropping BDNF levels to aged OVX non-treated mice. |
| 4 | Progesterone up-regulates neuronal brain-derived neurotrophic factor expression in the injured spinal cord DOI:10.1016/j.neuroscience.2004.02.024 |
To demonstrate that BDNF increases with PROG treatment, in ventral horn motoneurons from spinal cord injured mice. | Spinal cord injury reduces BDNF expression levels by 50% in spinal motoneurons. PROG enhances BDNF in the motoneurons of lesioned spinal cord mice. |
| 5 | Progesterone treatment of spinal cord injury: Effects on Receptors, Neurotrophins, and Myelination DOI: 10.1385/JMN:28:1:3 |
To examine the cellular mechanisms involved in PROG neuroprotection in spinal cord injured rats. | PROG increases BDNF expression and protein in motoneurons in injured rats. These increases correlated with an increased TrkB and the phosphorylated CREB in motoneurons. |
| 6 | Progesterone increases Brain-Derived Neurotrophic Factor Expression and Protects Against Glutamate Toxicity in a Mitogen-Activated Protein Kinase- and Phosphoinositide-3 Kinase-Dependent Manner in Cerebral Cortical Explants Paramjit DOI: 10.1002/jnr.21370 |
To examine frontal and cingulate cerebral cortex explants from mice treated with PROG in vitro. | PROG induces a 75% increase in BDNF expression in explants of the cerebral cortex, with a nearly identical effect on BDNF protein levels. |
| 7 | Progesterone modulates brain-derived neurotrophic factor and choline acetyltransferase in degenerating Wobbler motoneurons DOI:10.1016/j.expneurol.2006.08.019 |
To examined steroid and BDNF expression and protein in the spinal cord, and in muscle atrophy in wobbler rodent. | BDNF expression was in in neuros of steroid-naïve Wobbler mice compared to controls. PROG treatment increased BDNF expression in Wobblers compared to untreated, but not to controls. |
| 8 | Progesterone pre-treatment enhances serotonin-stimulated BDNF gene expression in rat C6 glioma cells through production of 5α-reduced neurosteroids DOI:10.1007/s12031-007-9034-6 |
To investigate the rule of neurotransmitters on glial cell metabolism and function in rat glioma cells in vitro. | BDNF expression levels in both non- treated and PROG-pre-treated glioma cells were similarly elevated by serotonin treatment with a concentration-dependent effect of serotonin on BDNF gene expression. |
| 9 | The differences in neuroprotective efficacy of progesterone and medroxyprogesterone acetate correlate with their effects on brain-derived neurotrophic factor expression DOI: 10.1210/en.2008-1247 |
To determine which type of PROG receptor mediates the neuroprotective effect of PROG on BDNF. | PROG induces increase in the BDNF protein levels in cerebral cortical explants in a concentration-dependent manner. PROG regulates BDNF expression through the classical PROG receptor. |
| 10 | Progesterone, BDNF and Neuroprotection in the Injured CNS DOI:10.1080/00207450903116430 |
To investigate the neuroprotective mechanism of PROG and BDNF. | PROG had no effect on BDNF expression in granule neurons. No neuroprotective role for PROG on BDNF was observed. |
| 11 | Progesterone inhibits estrogen-mediated neuroprotection against excitotoxicity by down-regulating estrogen receptor-β. DOI:10.1111/j.1471-4159.2010.07038.x |
To examine PROG and estrogen treatment in cultured hippocampal slices on levels of ERα and ERβ, and BDNF. | Estrogen elevated ERβ expression and protein levels, did not modify ERα expression, but increased ERα protein levels, and increased BDNF expression levels in hippocampal cells. PROG reversed the estrogen-elicited increases in ERβ, ERα protein, and BDNF expression levels. |
| 12 | Progesterone treatment alters neurotrophin/proneurotrophin balance and receptor expression in rats with traumatic brain injury. DOI:10.3233/RNN-2011-0628 |
To characterize the expression of BDNF isoforms following PROG treatment for traumatic brain injury . | PROG reduces levels of pro-BDNF and TrkB post- brain injury. Mature BDNF was decreased at 24 and 72 h. |
| 13 | Progesterone increases the release of brain-derived neurotrophic factor from glia via progesterone receptor membrane component 1 (Pgrmc1)-dependent ERK5 signaling. DOI: 10.1210/en.2011-2177 |
To study PROG-induced BDNF release and the extracellular signal-regulated kinase 5. | PROG and the membrane-impermeable PROG both induced BDNF release from glial cells and primary astrocytes, which lack the classical nuclear/intracellular PROG receptor but express a membrane-associated PROG receptors. |
| 14 | Progesterone effects on neuronal brain-derived neurotrophic factor and glial cells during progression of Wobbler mouse neurodegeneration. DOI:10.1016/j.neuroscience.2011.11.034 |
To compared PROG regulation of BDNF in motoneurons and oligodendrocytes of Wobbler mice. | PROG upregulated the low levels of BDNF expression in the grey matter regions at the symptomatic stage of disease, and increased BDNF expression in the late stage Wobblers. BDNF protein was normal in steroid-naive symptomatic Wobblers. |
| 15 | Progesterone attenuates several hippocampal abnormalities of the wobbler mouse. DOI: 10.1111/jne.12004 |
To examine the hippocampus of Wobbler mice and their changes in response to PROG treatment. | Wobblers mice display a decreased BDNF expression. PROG did not change the normal parameters in control mice and attenuated hippocampal abnormalities in Wobblers. |
| 16 | Progesterone in the treatment of neonatal arterial ischemic stroke and acute seizures: Role of BDNF/TrkB signaling. DOI:10.1016/j.neuropharm.2016.03.052 |
To examine the effects of PROG on BDNF-TrkB signaling and inflammation following neonatal arterial ischemic stroke in mice. | PROG suppresses the expression of BDNF in seizure mice at day 1, but at day 3, BDNF expression is comparable to controls. PROG treatment first inhibited TrkB expression at day 1 then increased TrkB receptor expression at days 3. |
| 17 | Progesterone modulates post-traumatic epileptogenesis through regulation of BDNF-TrkB signaling and cell survival-related pathways in the rat hippocampus. DOI: 10.1016/j.neulet.2019.134384 |
To study the effect of PROG on post-traumatic epileptogenesis survival-related pathways. | PROG treated animals reduced the duration of seizures and enhanced the amount of BDNF in the ipsilateral hippocampus. |
| 18 | Progesterone’s Effects on Cognitive Performance of Male Mice Are Independent of Progestin Receptors but Relate to Increases in GABAA Activity in the Hippocampus and Cortex. DOI:10.3389/fendo.2020.552805 |
To evaluate PROG effect on the hippocampal and cortical levels of BDNF in mice. | PROG increased BDNF levels in the hippocampus, but not in the cortex, of male mice. |
The modulatory effects of progesterone combination to estradiol on BDNF might be influenced by dosage. Bimonte-Nelson et al., [84] showed that aged female rats receiving estradiol at a 1.5 mg/60 day regime exhibited increases in cortical levels of BDNF, and this was null when the animals received also progesterone at a 200mg/60 day regime. Oppositely, Saland, Schoepfer, and Kabbaj [85] reported that rats under cyclic administration of estradiol and progesterone that received ketamine injection exhibited an increase in hippocampal BDNF levels that did not occur to those receiving only estradiol or progesterone.
The time of exposition might be important for the neuroprotective effects of progesterone regulation on BDNF. Coughlan et al., [86] did not find an effect for progesterone on BDNF expression in the brain of mice who received a single progesterone dose of 8 mg/kg at the onset of a stroke; and Cekic et al., [87] reported that the levels of BDNF and pro-BDNF and TrkB were reduced in the brain of rats treated with progesterone as 8mg/kg intraperitoneal injection at 1 hr, and subcutaneous injections at 6 and 24 h and every 24 h post-injury. Alternatively, Yousuf et al., [88] showed that stroke rats who received progesterone (8 or 16 mg/kg) injections at 2 h, 6 h and every 24 h until day 7 post-occlusion restored BDNF levels at 3 and 7 days post-stroke. Similarly, Atif et al., [89] mice with ischemic stroke that received 8 mg/kg progesterone injections at 1 h post-ligation, at 3 h post-ligation and every 24 h for 6 days showed a transient reduction in BDNF expression at day 1 that increased at day 3 through day 7. Finally, Ghadiri et al., [90] recently showed that male rats had an increase in hippocampal BDNF concentrations with a low-dose progesterone treatment; and that a high-dose progesterone treatment resulted in a BDNF levels decline to lower than those found in brain-injury rats. It is possible that a time gap evidenced in BDNF response to progesterone occurs due to demands on membrane-associate-PRs. And that a critical range in progesterone dosage dictates whether BDNF expression will be induced by progesterone receptors activation or inhibited by CREB inhibition.
Jodhka et al., [91] identified that progesterone regulation of BDNF expression in mice brain is mediated by the classical nuclear PR; Su et al., [92] showed that progesterone induces BDNF release in glial cells and astrocytes, which lack the classical nuclear PR, by activation of the membrane component 1 signalling (Pgrmc1). Nuclear steroid receptors stimulate gene expression by facilitating the assembly of basal transcription factors into a preinitiation complex that requires additional, and sometimes common, coactivators. Which means that activation of ERs might impair PR-dependent gene expression via sequester of the common coactivator of CREB (CREB binding protein) [93]. A competition between these nuclear receptors for a limited concentrations of common coactivators is an integrative mechanism by which sex steroids concur in the genomic regulation of BDNF.
Although the presence of ERs is more abundantly present across brain tissues, and the gene for BDNF contains an ERE sequence so that ERs activation more effectively regulate the neuroprotective effects of BDNF. Progesterone activation of non-nuclear receptors influences on BDNF expression and release in diverse neuronal and non-neuronal tissues, and has been identified to positively regulate BDNF expression in spinal cord motor neurons, striatal neuros, hippocampal and cortical neurons [94,95,96,97,98,99,100].
4. Androgens and BDNF Interactions
Several studies elucidate that testosterone is able to increase the expression levels of BDNF in the high vocal center (HVC) of female and male birds, whereby it mediates recruital and survival of newborn neurons Xu et al., [101], Louissaint et al., [102], Fusani et al., [103], Hartog et al., [104], Ottem et al., [105], Li et al., [106], Fanaei et al., [107] and Falk Dittrich et al., [108]. Throughout life, new neurons arise from the ventricular zone of the adult songbird brain and are recruited to the song control nucleus HVC, from which they extend projections to its target. This process of ongoing circuit integration is modulated by seasonal surges in systemic testosterone, and supported by BDNF [109,110]. However, while It is known that singing upregulates HVC’s BDNF expression, and BDNF mediates androgen-induced HVC neuronal recruitment, the expression of BDNF is found diminished in aromatase-inhibited birds [111]. It is important to point out that brain aromatase converts circulating testosterone to estradiol and thus the HVC is exposed to both androgenic and estrogenic stimulation. Considering a sexual dimorphism in BDNF expression in the canary HVC, with higher levels in the adult male than female and evidence of testosterone-induced increase in BDNF levels in adult female HVC, the indirect effect of testosterone on BDNF expression may occur either via its metabolite estradiol or via testosterone-induced increases in singing activity. Consonantly, Allen et al., [112] experiments in adolescent male rats and macaques added that neither gonadectomy nor testosterone replacement altered BDNF or TrkB expression levels in the hippocampal tissue of the animals. Together, these findings suggest that local changes in BDNF expression likely follow estradiol availability and, possibly, gonadectomy or testosterone replacement did not alter the hippocampal production of estradiol.
Table 3.
Data summary of interactions between Testosterone and BDNF.
| Title | Experiment | Main Findings | |
|---|---|---|---|
| 1 | Brain-derived neurotrophic factor regulates expression of androgen receptors in perineal motoneurons DOI: 10.1073/pnas.94.4.1521 |
To examine steroid receptor expression in motoneurons of the SNB in mice. | Axonal transport disruption downregulates AR expression in motoneurons and BDNF treatment reverses it. |
| 2 | Estrogen-inducible, sex-specific expression of brain-derived neurotrophic factor mRNA in a forebrain song control nucleus of the juvenile zebra finch DOI: 10.1073/pnas.96.14.8241 |
To examine the expression of AR, BDNF, and TrkB in the HVC, neostriatum, and archistriatum in Zebra finches. | BDNF expression is increased in the HVC of male, but not female, zebra finches. Estrogen and aromatase inhibition induce a premature stimulation and an inhibition on the increase patterns of BDNF expression, respectively, in juvenile males. |
| 3 | BDNF mediates the effects of testosterone on the survival of new neurons in an adult brain DOI:10.1016/S0896-6273(00)80678-9 |
To examine BDNF responses to testosterone treatment in the HVC of male canaria. | Testosterone treatment increases BDNF levels in the HVC of adult canaria. BDNF antibody blocked the testosterone-induced increase in new neurons. |
| 4 | BDNF regulation of androgen receptor expression in axotomized SNB motoneurons of adult male rats DOI:10.1016/S0006-8993(99)02225-8 |
To examine BDNF effects on axotomy- induced loss of AR expression in SNB motoneurons in rats. | Delayed application of BDNF to axotomized SNB motoneurons restored AR expression the intact levels. |
| 5 | Blockade of endogenous neurotrophic factors prevents the androgenic rescue of rat spinal motoneurons DOI: 10.1523/jneurosci.21-12-04366.2001 |
To exploit motoneuron cell death in the SNB of mice and androgens effect. | Blockage of TrkB activity prevented the androgenic sparing of SNB motoneurons. This did not reduce SNB motoneuron number. |
| 6 | Coordinated interaction of neurogenesis and angiogenesis in the adult songbird brain DOI: 10.1016/S0896-6273(02)00722-5 |
To investigated testosterone-related angiogenesis and neuronal recruitment in adult songbird neostriatum. | HVC endothelial cells produce BDNF in a testosterone-dependent manner. |
| 7 | Aromatase inhibition affects testosterone-induced masculinization of song and the neural song system in female canaries DOI: 10.1002/neu.10141 |
To investigated the role of estrogen in controlling the development of the song structure (HVC) in female canaries. | Aromatase inhibition of testosterone-induced song motor development correlate with inhibition of BDNF in HVC of adult female canaries and alters the song pattern. |
| 8 | Brain-Derived Neurotrophic Factor and Androgen Interact in the Maintenance of Dendritic Morphology in a Sexually Dimorphic Rat Spinal Nucleus DOI: 10.1210/en.2003-0853 |
To test BDNF and testosterone effects on dendritic morphology in motoneurons of the SNB in rats. | Testosterone or BDNF failed to support dendritic length or distribution. Treatment with testosterone plus BDNF restores dendritic morphology to the level of controls. |
| 9 | Androgen regulates trkB immunolabeling in spinal motoneurons DOI: 10.1002/jnr.21122 |
To examine gonadal hormones regulation of BDNF systems in rodents spinal motoneurons. | TrkB receptors regulation is androgen sensitive in motoneurons on the SNB. Castration- induced changes in SNB motoneurons are prevented by testosterone replacement. |
| 10 | Androgen-dependent regulation of brain-derived neurotrophic factor and tyrosine kinase B in the sexually dimorphic SNB DOI: 10.1210/en.2007-0308 |
To investigate the androgens regulation of BDNF protein in SNB motoneurons. |
SNB motoneurons and the non-androgen-responsive motoneurons of the adjacent retrodorsolateral nucleus express BDNF and trkB. Testosterone regulates BDNF protein in SNB, but not in the retrodorsolateral nucleus dendrites. |
| 11 | Differential expression and regulation of brain-derived neurotrophic factor mRNA isoforms in androgen-sensitive motoneurons of the rat lumbar spinal cord. DOI:10.1016/j.mce.2010.07.001 |
To examine the specific BDNF transcripts regulated by androgens in the SNB motoneurons of male rats. | BDNF isoforms containing exon VI were decreased in SNB motoneurons in an androgen-dependent manner, but unaffected in retrodorsolateral motoneurons. |
| 12 | Androgen regulates brain-derived neurotrophic factor in spinal motoneurons and their target musculature. DOI: 10.1210/en.2009-1036 |
To examine androgens regulation of BDNF in quadriceps and SNB motoneurons, and their corresponding target musculature in male rats. | Castration reduced BDNF protein in the quadriceps and SNB motoneurons, and their target musculature, and this was prevented with testosterone replacement. |
| 13 | Modulatory Effects of Sex Steroid Hormones on Brain-Derived Neurotrophic Factor-Tyrosine Kinase B Expression during Adolescent Development in C57Bl/6 Mice. DOI:10.1111/j.1365-2826.2012.02277.x |
To examine sex steroid hormones and neurotrophic signalling during adolescent development in a mouse model. | Castration and testosterone or DHT replacement had a receptor-dependent effect on BDNF-TrkB signalling in the forebrain and hippocampal regions of the adolescent animals. Females changes in BDNF-TrkB signalling do not correspond with changes in serum estrogen. |
| 14 | Androgen action at the target musculature regulates brain-derived neurotrophic factor protein in the SNB. DOI: 10.1002/dneu.22083 |
To examine if testosterone regulates BDNF in SNB motoneurons of male rats by acting locally at the bulbocavernosus muscle. | Testosterone directly to the bulbocavernosus muscle maintains BDNF levels in SNB motoneurons intact, after castration. AR blockage decreases BDNF, compared with animals treated with intramuscular testosterone. |
| 15 | Regulatory mechanisms of testosterone-stimulated song in the sensorimotor nucleus HVC of female songbirds. DOI:10.1186/s12868-014-0128-0 |
To examine the effects of testosterone on the anatomy and the song control nucleus HVC of female European robins. | Testosterone induced differentiation, angiogenesis and neuron projection morphogenesis. BDNF functions as a common mediator of the testosterone effects in HVC. |
| 16 | Testosterone enhances functional recovery after stroke through promotion of antioxidant defenses, BDNF levels and neurogenesis in male rats. DOI:10.1016/j.brainres.2014.02.028 |
To evaluate the effects of testosterone on BDNF and neurogenesis in a castrated male rat model of focal cerebral ischemia. | Testosterone increased BDNF levels and neurogenesis after focal cerebral ischemia. |
| 17 | The effect of adolescent testosterone on hippocampal BDNF and TrkB mRNA expression: Relationship with cell proliferation. DOI:10.1186/s12868-015-0142-x |
To examine the molecular mechanism underlying testosterone actions on postnatal neurogenesis and BDNF/TrkB levels in rhesus macaque and rat. | Gonadectomy or steroids replacement did not alter BDNF or TrkB expression levels in young adult male rat or rhesus macaque hippocampus. There was a positive correlation between cell proliferation and TrkB expression, only when steroids are present. |
| 18 | Effects of testosterone on synaptic plasticity mediated by androgen receptors in male SAMP8 mice. DOI:10.1080/15287394.2016.1193113 |
To study the protective role of testosterone on cognitive performance in an Alzheimer disease animal model. | The expression of BDNF and cyclic-AMP response element binding protein (CREB)/CREB levels were elevated in testosterone treated animals. |
| 19 | Hedonic sensitivity to low-dose ketamine is modulated by gonadal hormones in a sex-dependent manner. DOI: 10.1038/SREP21322 |
To investigate the testosterone contribution to the rapid antidepressant-like effects of ketamine. | Testosterone treatment responsiveness was associated with higher hippocampal BDNF levels in female rats. |
| 20 | TrkB is necessary for male copulatory behavior in the Syrian Hamster (Mesocricetus auratus). DOI:10.1016/j.yhbeh.2017.10.016 |
To examine how TrkB and BDNF mediate testosterone effects on the medial preoptic nucleus in Hamsters. | Testosterone treatment increased BDNF expression levels and conversely lowered the expression of TrkB receptors in the medial preoptic area of animals. |
| 21 | Prenatal Androgenization Induces Anxiety-Like Behavior in Female Rats, Associated with Reduction of Inhibitory Interneurons and Increased BDNF in Hippocampus and Cortex. DOI:10.1155/2019/3426092 |
To evaluated the influence of maternal hyperandrogenemia on offspring levels of BDNF in the hippocampus and cerebral cortex. | BDNF expression was increased in the hippocampus and cerebral cortex of Prenatal hyperandrogenization offspring in comparison with the controls. |
| 22 | Deficiency in Androgen Receptor Aggravates the Depressive-Like Behaviors in Chronic Mild Stress Model of Depression. DOI: 10.3390/cells8091021 |
To exploit how AR and stress influence the onset of the major depressive disorder. | Loss of AR affects depressive-like behaviours by modulate BDNF expression. |
| 23 | Effect of adolescent androgen manipulation on psychosis-like behaviour in adulthood in BDNF heterozygous and control mice. DOI:10.1016/j.yhbeh.2019.03.005 |
To examine how adolescent androgens influence psychosis-like behaviour in the adulthood and the BDNF rule, in mice. | Testosterone and DHT treatment reduce the expression of dopamine transporter in the medial prefrontal cortex of mice. These effects are absent in BDNF heterozygous mice. |
| 24 | Dose-dependent effects of testosterone on spatial learning strategies and brain-derived neurotrophic factor in male rats. DOI:10.1016/j.psyneuen.2020.104850 |
To investigate the effect of different doses of testosterone on spatial learning strategies in male rats. | Low testosterone dose increased total BDNF in the striatum, and high dose increased total BDNF in the hippocampus. |
Interaction between BDNF and testosterone seems to be necessary to maintain dendritic morphology in SNB motoneurons. Testosterone can regulate the expression of AR and TrKB in spinal motoneurons [113], and BDNF signaling through TrkB is able to regulate the expression of androgen receptors (AR) [114,115,116,117]. Furthermore, BDNF expression is shown to be upregulated by testosterone signalling in AR expressing motoneurons of the vastus lateralis and the spinal nucleus of the bulbocavernosus and their corresponding muscles [118,119]. Together these findings suggest a mechanistic interaction between BDNF and testosterone whereby testosterone improves BDNF signaling and, consequently its expression, which in turn improves testosterone sensitivity via AR regulation. In the experiment by Yang, Verhovshek, and Sengelaub [120], treatment of castrated male rats with testosterone or BDNF alone was not sufficient to promote dendritic length and distribution in SNB motoneurons; whereas, combined treatment with testosterone and BDNF was able to restore the SNB dendritic morphology. Additionally, Zhang et al., [121] data indicate that different doses of testosterone may differently affect BDNF expression levels within the brain. In their experiment, young adult rats that received a low dose of testosterone (0.125 mg) showed increased BDNF concentrations in the striatum, and those who received a high dose (0.500 mg) had increased BDNF in their hippocampus.
The study by Hill et al., [122] reported a region-specific and time-dependent sex differences in BDNF-TrkB expression and signaling during adolescence. They described that serum testosterone levels increase in males mice from week 3 to 12, peaking at week 8 and then declining, and brain BDNF expression levels were increasing in a positive correlation with serum testosterone, peaking from weeks 7 to 10. A tendency for TrkB expression to decrease when BDNF levels were increasing was noted and castration did not affect BDNF expression in the mice brains. In females, BDNF expression did not change from week 3 to 12, however, TrkB activity was seen to increase from week 4, peaked at week 6, and then declined. OVX resulted in an increase in BDNF expression levels, that was not affected by estrogen replacement, and a decrease in TrkB activity in these adolescent mice. In Brague et al., [123] experiment, testosterone also increased the expression of BDNF while reduced TrkB levels in the hypothalamus of male Syrian hamster. Recently, the study by Rankov Petrovic et al., [124] revealed that the adult female offspring from testosterone-treated pregnant female rats had increased BDNF levels in the hippocampus and cortex.
5. Summary and Conclusion
Sex steroids play an essential role in neuronal circuits formation and maintenance via regulation of BDNF expression in neuronal proliferation, differentiation and homeostasis at important areas of the brain i.e., cortex, hippocampus, hypothalamus and upper areas of the midbrain, and cerebellum [18,26,29,41,51,58]. This occurs from the developmental stages throughout life.
Estrogens regulation of BDNF either via classical nuclear receptors or via activation of membrane-associated ERs by second messenger signalling pathways [125,126] is essential to neuronal function and is suggestibly a main regulator of TrkB expression. Whereas, excessive concentrations of BDNF reportedly downregulate TrkB expression levels [127]. Further, BDNF autocrine and paracrine signal transduction regulates its own expression within neighbour cells [128], and this enhances ERs activity, which in turn, increases BDNF mRNA transcription in a synergistic manner [17,32,36,40,47,48,50,56,129]. That implies that the estrogen/ER system is able to regulate the BDNF/TrkB system; while, the inhibitory effect of TrkB/PI3K/AKT pathway on ERE transcription indicates that BDNF signal is important to limit and control BDNF expression [46,47,48,50]. Furthermore, the time- and region- specific influence found for sex steroids on BDNF function likely depend on the presence and concentrations of estrogens [30,101,102,103,104,105,106,109,113,118,119,120,121,124,130].
A neuroprotective effect for the estrogens regulation of BDNF is consistently seen in individuals with various pathological conditions such as stress, hypertension, ischemic brain injury, Parkinson’s and Alzheimer’s diseases [27,28,31,33,42,59,74,75,76,77,78,83,86,89,107,110].
Although much less research is present on interactions between progesterone and BDNF. A direct regulation of BDNF expression was evidenced in cortical cells expressing the classical nuclear PR [75,91]. In addition, progesterone is able to increase BDNF synthesis and enhance neuroprotective cell responses by different ways [52,79,80,91,92,93,94,95,96,97,98,131]. For example, progesterone enhances anti-inflammatory processes and tissue recovery from neural injury [87,88,89,132,133,134,135].
Figure 2.
Schematic illustration of direct (complete arrow) and indirect (dot arrow) interactions between sex steroids and BDNF systems via activation of nuclear receptors in neuronal cells within the brain. Created with BioRender.com.
Figure 2.
Schematic illustration of direct (complete arrow) and indirect (dot arrow) interactions between sex steroids and BDNF systems via activation of nuclear receptors in neuronal cells within the brain. Created with BioRender.com.
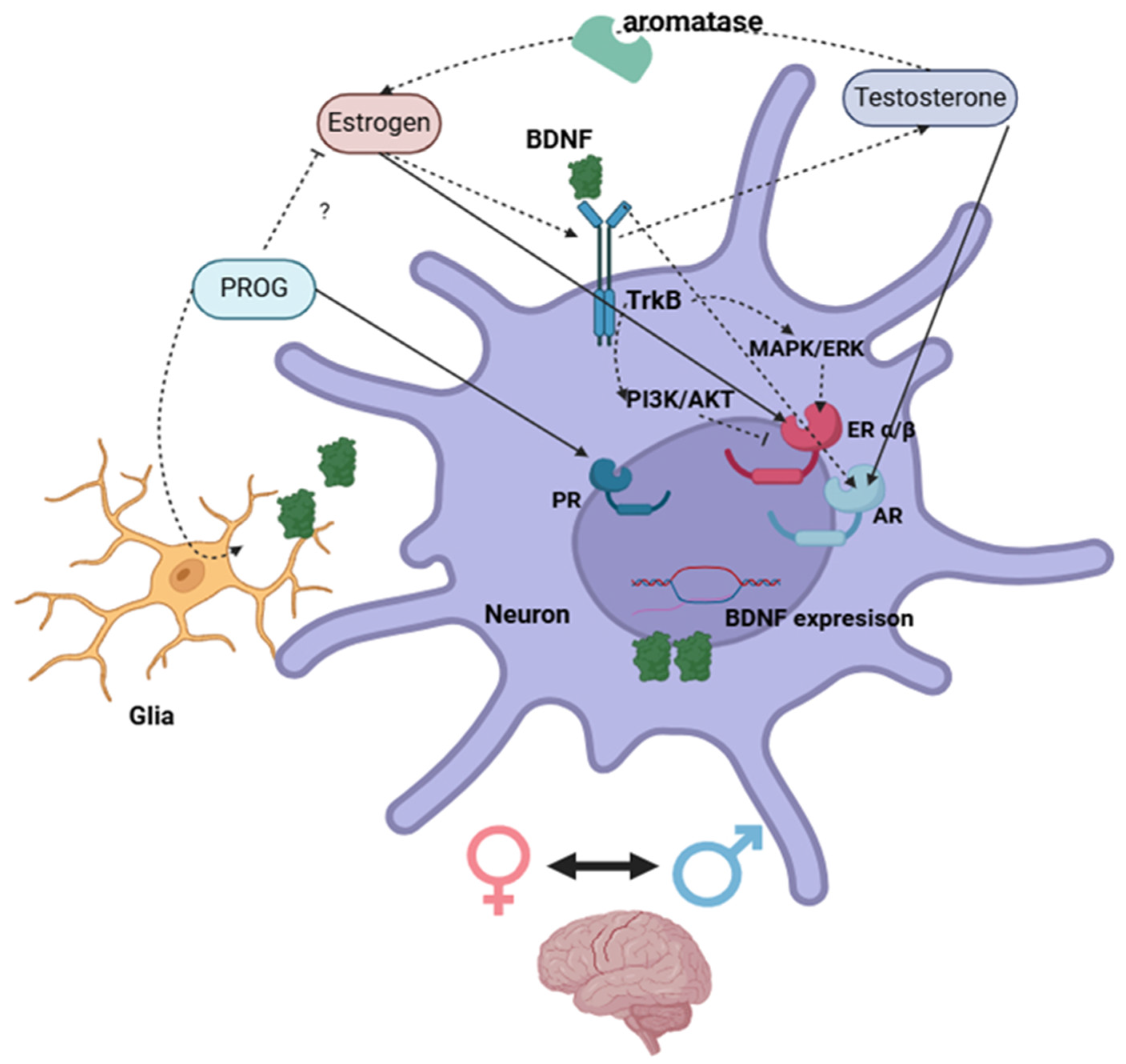
Testosterone does not regulate BDNF gene directly, but rather via activation of ARs which translocate to the nucleus and start gene expression at androgen responsive elements sequences of androgen-responsive genes upstream BDNF [108,114,115,136]. This regulation appears to be influenced by local factors. For instance, the findings of Zhang et al., [121] report that a low-dose of testosterone increase BDNF levels in the striatum while a high-dose testosterone increase BDNF levels in the hippocampus of male rats. Interestingly, the BDNF/TrkB system positively regulates the expression of AR in neurons, potentiating testosterone regulation on BDNF [120].
Regarding progesterone, studies report various effects of interactions between progesterone and estradiol combination on BDNF expression [43,45,81,84,85,94,95,99]. It is possible that their competitive activation of nuclear receptors might deplete co-activators, or their signaling through membrane-associated receptors result in a gain or a loss of effect on BDNF regulation. Further studies on the mechanistic interactions between sex steroids in the regulation of gene expression would be necessary to elucidate of progesterone and estradiol.
Sex steroids regulate the expression of BDNF either directly via nuclear receptors or by altering intracellular pathways that affect genomic regulation of BDNF. This in times can represent concurrent mechanisms.
Author Contributions
Conceptualization, G.G.d.A.; methodology, G.G.d.A. and E.M-C.; investigation, G.G.d.A., M.B.C.d.S. and E.M-C.; writing—original draft preparation, G.G.d.A. and M.B.C.d.S.; resources, E.M-C.; supervision, M.B.C.d.S.; project administration, G.G.d.A.; funding acquisition, E.M-C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Acknowledgments
I thank my professors and colleagues Dr. Maria Bernadete and Eugenia Murawska-Cialowicz for the indispensable support to make this publication possible.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| CREB | cAMP response element-binding protein |
References
- Lenz, K. M., Nugent, B. M. & McCarthy, M. M. Sexual differentiation of the rodent brain: Dogma and beyond. Front. Neurosci. 0, 26 (2012). [CrossRef]
- Giatti, S. et al. Sex differences in steroid levels and steroidogenesis in the nervous system: Physiopathological role. Front. Neuroendocrinol. 56, (2020). [CrossRef]
- Corpechot, C., Robel, P., Axelson, M., Sjövall, J. & Baulieu, E. E. Characterization and measurement of dehydroepiandrosterone sulfate in rat brain. Proc. Natl. Acad. Sci. U. S. A. 78, 4704–4707 (1981). [CrossRef]
- Selvaraj, V., Stocco, D. M. & Tu, L. N. Minireview: Translocator protein (TSPO) and steroidogenesis: A reappraisal. Mol. Endocrinol. 29, 490–501 (2015). [CrossRef]
- Porcu, P. et al. Neurosteroidogenesis Today: Novel Targets for Neuroactive Steroid Synthesis and Action and Their Relevance for Translational Research. J. Neuroendocrinol. 28, 1–19 (2016). [CrossRef]
- Frick, K. M., Kim, J., Tuscher, J. J. & Fortress, A. M. Sex steroid hormones matter for learning and memory: estrogenic regulation of hippocampal function in male and female rodents. Learn. Mem. 22, 472–493 (2015). [CrossRef]
- Anderson, N. K. et al. Activational vs. organizational effects of sex steroids and their role in the evolution of reproductive behavior: Looking to foot-flagging frogs and beyond. Horm. Behav. 146, 105248 (2022). [CrossRef]
- Fleischer, A. W. & Frick, K. M. New perspectives on sex differences in learning and memory. Trends Endocrinol. Metab. 34, 526–538 (2023). [CrossRef]
- Bouron, A., Boisseau, S., De Waard, M. & Peris, L. Differential down-regulation of voltage-gated calcium channel currents by glutamate and BDNF in embryonic cortical neurons. Eur. J. Neurosci. 24, 699–708 (2006). [CrossRef]
- Ibarra, I. L. et al. Comparative chromatin accessibility upon BDNF stimulation delineates neuronal regulatory elements. Mol. Syst. Biol. 18, 1–25 (2022). [CrossRef]
- Reichlin, S. Neuroendocrinology. N. Engl. J. Med. 269, 1246–1250 (1963).
- Hammes, S. R. & Levin, E. R. Impact of estrogens in males and androgens in females. Journal of Clinical Investigation vol. 129 1818–1826 (2019). [CrossRef]
- Segal, S. Hormones, Amino-Acid Transport and Protein Synthesis. Nature 203, 17–19 (1964). [CrossRef]
- Tomkins, G. M. & Maxwell, E. S. Some Aspects of Steroid Hormone Action. Annu. Rev. Biochem. 32, 677–708 (1963). [CrossRef]
- Arnold, A. P. & Gorski, R. A. Gonadal steroid induction of structural sex differences in the central nervous system. Annu. Rev. Neurosci. VOL. 7, 413–442 (1984). [CrossRef]
- Toran-Allerand, C. D. On the Genesis of Sexual Differentiation of the Central Nervous System: Morphogenetic Consequences of Steroidal Exposure and Possible Role of α-Fetoprotein. Prog. Brain Res. 61, 63–98 (1984).
- Miranda, R. C., Sohrabji, F. & Toran-Allerand, D. Interactions of estrogen with the neurotrophins and their receptors during neural development. Horm. Behav. 28, 367–375 (1994). [CrossRef]
- Singh, M., Meyer, E. M. & Simpkins, J. W. The effect of ovariectomy and estradiol replacement on brain-derived neurotrophic factor messenger ribonucleic acid expression in cortical and hippocampal brain regions of female Sprague-Dawley rats. Endocrinology 136, 2320–2324 (1995). [CrossRef]
- Levin, E. R. Plasma membrane estrogen receptors. Trends in Endocrinology and Metabolism vol. 20 477–482 (2009).
- Hammes, S. R. & Levin, E. R. Extranuclear steroid receptors: Nature and actions. Endocrine Reviews vol. 28 726–741 (2007). [CrossRef]
- Kulkoyluoglu, E. & Madak-Erdogan, Z. Nuclear and extranuclear-initiated estrogen receptor signaling crosstalk and endocrine resistance in breast cancer. Steroids 114, 41–47 (2016). [CrossRef]
- Hans, A. et al. Persistent, noncytolytic infection of neurons by Borna disease virus interferes with ERK 1/2 signaling and abrogates BDNF-induced synaptogenesis. FASEB J. 18, 863–865 (2004).
- Singh, M., Su, C. & Ng, S. Non-genomic mechanisms of progesterone action in the brain. Front. Neurosci. 7, 1–7 (2013). [CrossRef]
- van Noort, V., Snel, B. & Huynen, M. A. Predicting gene function by conserved co-expression. Trends Genet. 19, 238–242 (2003). [CrossRef]
- Teichmann, S. A. & Babu, M. M. Conservation of gene co-regulation in prokaryotes and eukaryotes. Trends Biotechnol. 20, 407–410 (2002). [CrossRef]
- Haraguchi, S. et al. Estradiol promotes purkinje dendritic growth, spinogenesis, and synaptogenesis during neonatal life by inducing the expression of BDNF. Cerebellum 11, 416–417 (2012). [CrossRef]
- Niu, Y. et al. Sex-dependent aberrant PFC development in the adolescent offspring rats exposed to variable prenatal stress. Int. J. Dev. Neurosci. 80, 464–476 (2020). [CrossRef]
- Naderi, M., Salahinejad, A., Attaran, A., Niyogi, S. & Chivers, D. P. Rapid effects of estradiol and its receptor agonists on object recognition and object placement in adult male zebrafish. Behav. Brain Res. 384, (2020). [CrossRef]
- Liu, Y. et al. Expression and estrogen regulation of brain-derived neurotrophic factor gene and protein in the forebrain of female prairie voles. J. Comp. Neurol. 433, 499–514 (2001). [CrossRef]
- Krizsan-Agbas, D., Pedchenko, T., Hasan, W. & Smith, P. G. Oestrogen regulates sympathetic neurite outgrowth by modulating brain derived neurotrophic factor synthesis and release by the rodent uterus. Eur. J. Neurosci. 18, 2760–2768 (2003). [CrossRef]
- Zhou, J., Zhang, H., Cohen, R. S. & Pandey, S. C. Effects of estrogen treatment on expression of brain-derived neurotrophic factor and cAMP response element-binding protein expression and phosphorylation in rat amygdaloid and hippocampal structures. Neuroendocrinology 81, 294–310 (2005). [CrossRef]
- Takuma, K. et al. 17Β-Estradiol Attenuates Hippocampal Neuronal Loss and Cognitive Dysfunction Induced By Chronic Restraint Stress in Ovariectomized Rats. Neuroscience 146, 60–68 (2007). [CrossRef]
- Pietranera, L., Lima, A., Roig, P. & De Nicola, A. F. Involvement of brain-derived neurotrophic factor and neurogenesis in oestradiol neuroprotection of the hippocampus of hypertensive rats. J. Neuroendocrinol. 22, 1082–1092 (2010). [CrossRef]
- Pietranera, L. et al. 17α-Oestradiol-Induced Neuroprotection in the Brain of Spontaneously Hypertensive Rats. J. Neuroendocrinol. 26, 310–320 (2014). [CrossRef]
- Fortress, A. M., Kim, J., Poole, R. L., Gould, T. J. & Frick, K. M. 17β-estradiol regulates histone alterations associated with memory consolidation and increases Bdnf promoter acetylation in middle-aged female mice. Learn. Mem. 21, 457–467 (2014). [CrossRef]
- Gresack, J. E. & Frick, K. M. Environmental enrichment reduces the mnemonic and neural benefits of estrogen. Neuroscience 128, 459–471 (2004). [CrossRef]
- Kiss, Á. et al. 17β-Estradiol replacement in young, adult and middle-aged female ovariectomized rats promotes improvement of spatial reference memory and an antidepressant effect and alters monoamines and BDNF levels in memory- and depression-related brain areas. Behav. Brain Res. 227, 100–108 (2012). [CrossRef]
- Kight, K. E. & McCarthy, M. M. Sex differences and estrogen regulation of BDNF gene expression, but not propeptide content, in the developing hippocampus. J. Neurosci. Res. 95, 345–354 (2017).
- Gatson, J. W. et al. Estrone is neuroprotective in rats after traumatic brain injury. J. Neurotrauma 29, 2209–2219 (2012). [CrossRef]
- Cambiasso, M. J., Colombo, J. A. & Carrer, H. F. Differential effect of oestradiol and astroglia-conditioned media on the growth of hypothalamic neurons from male and female rat brains. Eur. J. Neurosci. 12, 2291–2298 (2000). [CrossRef]
- Brito, V. I., Carrer, H. F. & Cambiasso, M. J. Inhibition of tyrosine kinase receptor type B synthesis blocks axogenic effect of estradiol on rat hypothalamic neurones in vitro. Eur. J. Neurosci. 20, 331–337 (2004). [CrossRef]
- Wang, P. et al. Additive antidepressant-like effects of fasting with β-estradiol in mice. J. Cell. Mol. Med. 23, 5508–5517 (2019).
- Bourque, M., Morissette, M. & Di Paolo, T. Neuroprotection in Parkinsonian-treated mice via estrogen receptor α activation requires G protein-coupled estrogen receptor 1. Neuropharmacology 95, 343–352 (2015). [CrossRef]
- Tang, Y. P. & Wade, J. 17β-Estradiol Regulates the Sexually Dimorphic Expression of BDNF and TrkB Proteins in the Song System of Juvenile Zebra Finches. PLoS One 7, (2012). [CrossRef]
- Tripanichkul, W., Gerdprasert, O. & Jaroensuppaperch, E. O. Estrogen reduces BDNF level, but maintains dopaminergic cell density in the striatum of MPTP mouse model. Int. J. Neurosci. 120, 489–495 (2010). [CrossRef]
- Wong, J., Woon, H. G. & Weickert, C. S. Full length TrkB potentiates estrogen receptor alpha mediated transcription suggesting convergence of susceptibility pathways in schizophrenia. Mol. Cell. Neurosci. 46, 67–78 (2011). [CrossRef]
- Cikla, U. et al. ERα signaling is required for TrkB-mediated hippocampal neuroprotection in female neonatal mice after hypoxic ischemic encephalopathy. eNeuro 3, 99–114 (2016).
- Spencer-Segal, J. L. et al. Estradiol acts via estrogen receptors alpha and beta on pathways important for synaptic plasticity in the mouse hippocampal formation. Neuroscience 202, 131–146 (2012). [CrossRef]
- Bohm-Levine, N., Goldberg, A. R., Mariani, M., Frankfurt, M. & Thornton, J. Reducing luteinizing hormone levels after ovariectomy improves spatial memory: Possible role of brain-derived neurotrophic factor. Horm. Behav. 118, (2020). [CrossRef]
- Gross, K. S., Alf, R. L., Polzin, T. R. & Frick, K. M. 17β-estradiol activation of dorsal hippocampal TrkB is independent of increased mature BDNF expression and is required for enhanced memory consolidation in female mice. Psychoneuroendocrinology 125, 105110 (2021). [CrossRef]
- Murphy, D. D., Cole, N. B. & Segal, M. Brain-derived neurotrophic factor mediates estradiol-induced dendritic spine formation in hippocampal neurons. Proc. Natl. Acad. Sci. U. S. A. 95, 11412–11417 (1998). [CrossRef]
- Zhu, Z., Liu, X., Senthil Kumar, S. P. D., Zhang, J. & Shi, H. Central expression and anorectic effect of brain-derived neurotrophic factor are regulated by circulating estradiol levels. Horm. Behav. 63, 533–542 (2013). [CrossRef]
- Balasubramanian, P., Subramanian, M., Nunez, J. L., MohanKumar, S. M. J. & MohanKumar, P. S. Chronic estradiol treatment decreases Brain Derived Neurotrophic Factor (BDNF) expression and monoamine levels in the amygdala - Implications for behavioral disorders. Behav. Brain Res. 261, 127–133 (2014). [CrossRef]
- Cavus, I. & Duman, R. S. Influence of estradiol, stress, and 5-HT2A agonist treatment on brain-derived neurotrophic factor expression in female rats. Biol. Psychiatry 54, 59–69 (2003). [CrossRef]
- Jezierski, M. K. & Sohrabji, F. Estrogen Enhances Retrograde Transport of Brain-Derived Neurotrophic Factor in the Rodent Forebrain. Endocrinology 144, 5022–5029 (2003). [CrossRef]
- Aguado-Llera, D. et al. 17β-Estradiol protects depletion of rat temporal cortex somatostatinergic system by β-amyloid. Neurobiol. Aging 28, 1396–1409 (2007). [CrossRef]
- Küppers, E., Krust, A., Chambon, P. & Beyer, C. Functional alterations of the nigrostriatal dopamine system in estrogen receptor-α knockout (ERKO) mice. Psychoneuroendocrinology 33, 832–838 (2008). [CrossRef]
- Solum, D. T. & Handa, R. J. Estrogen Regulates the Development of Brain-Derived Neurotrophic Factor mRNA and Protein in the Rat Hippocampus. J. Neurosci. 22, 2650–2659 (2002). [CrossRef]
- Baumgartner, N. E., Black, K. L., McQuillen, S. M. & Daniel, J. M. Previous estradiol treatment during midlife maintains transcriptional regulation of memory-related proteins by ERα in the hippocampus in a rat model of menopause. Neurobiol. Aging 105, 365–373 (2021). [CrossRef]
- Blurton-Jones, M., Kuan, P. N. & Tuszynski, M. H. Anatomical Evidence for Transsynaptic Influences of Estrogen on Brain-Derived Neurotrophic Factor Expression. J. Comp. Neurol. 468, 347–360 (2004). [CrossRef]
- Meltser, I. et al. Estrogen receptor β protects against acoustic trauma in mice. J. Clin. Invest. 118, 1563–1570 (2008). [CrossRef]
- Chhibber, A., Woody, S. K., Karim Rumi, M. A., Soares, M. J. & Zhao, L. Estrogen receptor β deficiency impairs BDNF–5-HT2A signaling in the hippocampus of female brain: A possible mechanism for menopausal depression. Psychoneuroendocrinology 82, 107–116 (2017). [CrossRef]
- Pietranera, L. et al. Selective Oestrogen Receptor Agonists Rescued Hippocampus Parameters in Male Spontaneously Hypertensive Rats. J. Neuroendocrinol. 28, (2016). [CrossRef]
- Kim, H. I. et al. ERRγ ligand HPB2 upregulates BDNF-TrkB and enhances dopaminergic neuronal phenotype. Pharmacol. Res. 165, 105423 (2021).
- Wu, Y. W. C., Du, X., Van Den Buuse, M. & Hill, R. A. Analyzing the influence of BDNF heterozygosity on spatial memory response to 17β-estradiol. Transl. Psychiatry 5, (2015). [CrossRef]
- de Assis, G. G. & Hoffman, J. R. The BDNF Val66Met Polymorphism is a Relevant, But not Determinant, Risk Factor in the Etiology of Neuropsychiatric Disorders – Current Advances in Human Studies: A Systematic Review. Brain Plast. 8, 133–142 (2022).
- de Assis, G. G. et al. The Val66 and Met66 Alleles-Specific Expression of BDNF in Human Muscle and Their Metabolic Responsivity. Front. Mol. Neurosci. 14, 1–12 (2021).
- Ivanova, T., Kppers, E., Engele, J. & Beyer, C. Estrogen stimulates brain-derived neurotrophic factor expression in embryonic mouse midbrain neurons through a membrane-mediated and calcium-dependent mechanism. J. Neurosci. Res. 66, 221–230 (2001). [CrossRef]
- Zhao, X. et al. Estrogen affects BDNF expression following chronic constriction nerve injury. Neuroreport 14, 1627–1631 (2003). [CrossRef]
- Dittrich, F., Feng, Y., Metzdorf, R. & Gahr, M. Estrogen-inducible, sex-specific expression of brain-derived neurotrophic factor mRNA in a forebrain song control nucleus of the juvenile zebra finch. Proc. Natl. Acad. Sci. U. S. A. 96, 8241–8246 (1999). [CrossRef]
- Jezierski, M. K. & Sohrabji, F. Region- and peptide-specific regulation of the neurotrophins by estrogen. Mol. Brain Res. 85, 77–84 (2000). [CrossRef]
- Jezierski, M. K. & Sohrabji, F. Neurotrophin expression in the reproductively senescent forebrain is refractory to estrogen stimulation. Neurobiol. Aging 22, 311–321 (2001). [CrossRef]
- Moreno-Piovano, G. S., Varayoud, J., Luque, E. H. & Ramos, J. G. Long-term ovariectomy increases BDNF gene methylation status in mouse hippocampus. J. Steroid Biochem. Mol. Biol. 144, 243–252 (2014). [CrossRef]
- Berchtold, N. C., Kesslak, J. P., Pike, C. J., Adlard, P. A. & Cotman, C. W. Estrogen and exercise interact to regulate brain-derived neurotrophic factor mRNA and protein expression in the hippocampus. Eur. J. Neurosci. 14, 1992–2002 (2001). [CrossRef]
- Frye, C. A., Rhodes, M. E. & Dudek, B. Estradiol to aged female or male mice improves learning in inhibitory avoidance and water maze tasks. Brain Res. 1036, 101–108 (2005). [CrossRef]
- Kim, T. W., Kim, C. S., Kim, J. Y., Kim, C. J. & Seo, J. H. Combined exercise ameliorates ovariectomy-induced cognitive impairment by enhancing cell proliferation and suppressing apoptosis. Menopause 23, 18–26 (2016). [CrossRef]
- Rashidy-Pour, A., Bavarsad, K., Miladi-Gorji, H., Seraj, Z. & Vafaei, A. A. Voluntary exercise and estradiol reverse ovariectomy-induced spatial learning and memory deficits and reduction in hippocampal brain-derived neurotrophic factor in rats. Pharmacol. Biochem. Behav. 187, 172819 (2019). [CrossRef]
- Rashidy-Pour, A. et al. Effects of treadmill exercise and sex hormones on learning, memory and hippocampal brain-derived neurotrophic factor levels in transient congenital hypothyroid rats. Behav. Pharmacol. 31, 641–651 (2020). [CrossRef]
- Gibbs, R. B. Treatment with estrogen and progesterone affects relative levels of brain-derived neurotrophic factor mRNA and protein in different regions of the adult rat brain. Brain Res. 844, 20–27 (1999). [CrossRef]
- Gibbs, R. B. Levels of trkA and BDNF mRNA, but not NGF mRNA, fluctuate across the estrous cycle and increase in response to acute hormone replacement. Brain Res. 787, 259–268 (1998). [CrossRef]
- Murphy, D. D. & Segal, M. Progesterone prevents estradiol-induced dendritic spine formation in cultured hippocampal neurons. Neuroendocrinology 72, 133–143 (2000). [CrossRef]
- Aguirre, C., Jayaraman, A., Pike, C. & Baudry, M. Progesterone inhibits estrogen-mediated neuroprotection against excitotoxicity by down-regulating estrogen receptor-β. J. Neurochem. 115, 1277–1287 (2010).
- Franklin, T. B. & Perrot-Sinal, T. S. Sex and ovarian steroids modulate brain-derived neurotrophic factor (BDNF) protein levels in rat hippocampus under stressful and non-stressful conditions. Psychoneuroendocrinology 31, 38–48 (2006). [CrossRef]
- Bimonte-Nelson, H. A., Nelson, M. E. & Granholm, A. C. E. Progesterone counteracts estrogen-induced increases in neurotrophins in the aged female rat brain. Neuroreport 15, 2659–2663 (2004). [CrossRef]
- Saland, S. K., Schoepfer, K. J. & Kabbaj, M. Hedonic sensitivity to low-dose ketamine is modulated by gonadal hormones in a sex-dependent manner. Sci. Rep. 6, 21322 (2016). [CrossRef]
- Coughlan, T., Gibson, C. & Murphy, S. Progesterone, BDNF and neuroprotection in the injured CNS. Int. J. Neurosci. 119, 1718–1740 (2009). [CrossRef]
- Cekic, M., Johnson, S. J., Bhatt, V. H. & Stein, D. G. Progesterone treatment alters neurotrophin/proneurotrophin balance and receptor expression in rats with traumatic brain injury. Restor. Neurol. Neurosci. 30, 115–126 (2012). [CrossRef]
- Yousuf, S., Atif, F., Sayeed, I., Wang, J. & Stein, D. G. Post-stroke infections exacerbate ischemic brain injury in middle-aged rats: Immunomodulation and neuroprotection by progesterone. Neuroscience 239, 92–102 (2013). [CrossRef]
- Atif, F., Yousuf, S. & Stein, D. G. Progesterone in the treatment of neonatal arterial ischemic stroke and acute seizures: Role of BDNF/TrkB signaling. Neuropharmacology 107, 317–328 (2016). [CrossRef]
- Ghadiri, T., Vakilzadeh, G., Hajali, V. & Khodagholi, F. Progesterone modulates post-traumatic epileptogenesis through regulation of BDNF-TrkB signaling and cell survival-related pathways in the rat hippocampus. Neurosci. Lett. 709, (2019).
- Jodhka, P. K., Kaur, P., Underwood, W., Lydon, J. P. & Singh, M. The differences in neuroprotective efficacy of progesterone and medroxyprogesterone acetate correlate with their effects on brain-derived neurotrophic factor expression. Endocrinology 150, 3162–3168 (2009). [CrossRef]
- Su, C., Cunningham, R. L., Rybalchenko, N. & Singh, M. Progesterone increases the release of brain-derived neurotrophic factor from glia via progesterone receptor membrane component 1 (Pgrmc1)-dependent ERK5 signaling. Endocrinology 153, 4389–4400 (2012). [CrossRef]
- Smith, C. L., Oñate, S. A., Tsai, M. J. & O’Malley, B. W. CREB binding protein acts synergistically with steroid receptor coactivator-1 to enhance steroid receptor-dependent transcription. Proc. Natl. Acad. Sci. U. S. A. 93, 8884–8888 (1996). [CrossRef]
- González, S. L. et al. Progesterone up-regulates neuronal brain-derived neurotrophic factor expression in the injured spinal cord. Neuroscience 125, 605–614 (2004). [CrossRef]
- Gonzalez Deniselle, M. C. et al. Progesterone modulates brain-derived neurotrophic factor and choline acetyltransferase in degenerating Wobbler motoneurons. Exp. Neurol. 203, 406–414 (2007). [CrossRef]
- Meyer, M. et al. Progesterone effects on neuronal brain-derived neurotrophic factor and glial cells during progression of Wobbler mouse neurodegeneration. Neuroscience 201, 267–279 (2012). [CrossRef]
- Meyer, M. et al. Progesterone attenuates several hippocampal abnormalities of the wobbler mouse. J. Neuroendocrinol. 25, 235–243 (2013). [CrossRef]
- Kaur, P. et al. Progesterone increases brain-derived neurotrophic factor expression and protects against glutamate toxicity in a mitogen-activated protein kinase- and phosphoinositide-3 kinase-dependent manner in cerebral cortical explants. J. Neurosci. Res. 85, 2441–2449 (2007).
- Litim, N., Morissette, M. & Di Paolo, T. Effects of progesterone administered after MPTP on dopaminergic neurons of male mice. Neuropharmacology 117, 209–218 (2017). [CrossRef]
- Frye, C. A., Lembo, V. F. & Walf, A. A. Progesterone’s Effects on Cognitive Performance of Male Mice Are Independent of Progestin Receptors but Relate to Increases in GABAA Activity in the Hippocampus and Cortex. Front. Endocrinol. (Lausanne). 11, 552805 (2021). [CrossRef]
- Xu, J., Gingras, K. M., Bengston, L., Di Marco, A. & Forger, N. G. Blockade of endogenous neurotrophic factors prevents the androgenic rescue of rat spinal motoneurons. J. Neurosci. 21, 4366–4372 (2001). [CrossRef]
- Louissaint, A., Rao, S., Leventhal, C. & Goldman, S. A. Coordinated interaction of neurogenesis and angiogenesis in the adult songbird brain. Neuron 34, 945–960 (2002). [CrossRef]
- Fusani, L., Metzdorf, R., Hutchison, J. B. & Gahr, M. Aromatase inhibition affects testosterone-induced masculinization of song and the neural song system in female canaries. J. Neurobiol. 54, 370–379 (2003). [CrossRef]
- Hartog, T. E. et al. Brain-derived neurotrophic factor signaling in the HVC is required for testosterone-induced song of female canaries. J. Neurosci. 29, 15511–15519 (2009). [CrossRef]
- Ottem, E. N., Poort, J. E., Wang, H., Jordan, C. L. & Breedlove, S. M. Differential expression and regulation of brain-derived neurotrophic factor (BDNF) mRNA isoforms in androgen-sensitive motoneurons of the rat lumbar spinal cord. Mol. Cell. Endocrinol. 328, 40–46 (2010). [CrossRef]
- Li, M., Masugi-Tokita, M., Takanami, K., Yamada, S. & Kawata, M. Testosterone has sublayer-specific effects on dendritic spine maturation mediated by BDNF and PSD-95 in pyramidal neurons in the hippocampus CA1 area. Brain Res. 1484, 76–84 (2012). [CrossRef]
- Fanaei, H. et al. Testosterone enhances functional recovery after stroke through promotion of antioxidant defenses, BDNF levels and neurogenesis in male rats. Brain Res. 1558, 74–83 (2014). [CrossRef]
- Dittrich, F. et al. Regulatory mechanisms of testosterone-stimulated song in the sensorimotor nucleus HVC of female songbirds. BMC Neurosci. 15, 1–16 (2014). [CrossRef]
- Rasika, S., Alvarez-Buylla, A. & Nottebohm, F. BDNF mediates the effects of testosterone on the survival of new neurons in an adult brain. Neuron 22, 53–62 (1999). [CrossRef]
- Jia, J. xin et al. Effects of testosterone on synaptic plasticity mediated by androgen receptors in male SAMP8 mice. J. Toxicol. Environ. Heal. - Part A Curr. Issues 79, 849–855 (2016). [CrossRef]
- Chen, Z., Ye, R. & Goldman, S. A. Testosterone modulation of angiogenesis and neurogenesis in the adult songbird brain. Neuroscience 239, 139–148 (2013). [CrossRef]
- Allen, K. M., Purves-Tyson, T. D., Fung, S. J. & Shannon Weickert, C. The effect of adolescent testosterone on hippocampal BDNF and TrkB mRNA expression: Relationship with cell proliferation. BMC Neurosci. 16, (2015). [CrossRef]
- Osborne, M. C., Verhovshek, T. & Sengelaub, D. R. Androgen regulates trkB immunolabeling in spinal motoneurons. J. Neurosci. Res. 85, 303–309 (2007). [CrossRef]
- Al-Shamma, H. A. & Arnold, A. P. Brain-derived neurotrophic factor regulates expression of androgen receptors in perineal motoneurons. Proc. Natl. Acad. Sci. U. S. A. 94, 1521–1526 (1997). [CrossRef]
- Yang, L. Y. & Arnold, A. P. BDNF regulation of androgen receptor expression in axotomized SNB motoneurons of adult male rats. Brain Res. 852, 127–139 (2000). [CrossRef]
- Halievski, K. et al. Androgen-dependent loss of muscle BDNF mRNA in two mouse models of SBMA. Exp. Neurol. 269, 224–232 (2015). [CrossRef]
- Du, X., McCarthny, C. R., Notaras, M., van den Buuse, M. & Hill, R. A. Effect of adolescent androgen manipulation on psychosis-like behaviour in adulthood in BDNF heterozygous and control mice. Horm. Behav. 112, 32–41 (2019). [CrossRef]
- Verhovshek, T. & Sengelaub, D. R. Androgen action at the target musculature regulates brain-derived neurotrophic factor protein in the spinal nucleus of the bulbocavernosus. Dev. Neurobiol. 73, 587–598 (2013). [CrossRef]
- Verhovshek, T., Cai, Y., Osborne, M. C. & Sengelaub, D. R. Androgen regulates brain-derived neurotrophic factor in spinal motoneurons and their target musculature. Endocrinology 151, 253–261 (2010). [CrossRef]
- Yang, L. Y., Verhovshek, T. & Sengelaub, D. R. Brain-Derived Neurotrophic Factor and Androgen Interact in the Maintenance of Dendritic Morphology in a Sexually Dimorphic Rat Spinal Nucleus. Endocrinology 145, 161–168 (2004). [CrossRef]
- Zhang, K. J., Ramdev, R. A., Tuta, N. J. & Spritzer, M. D. Dose-dependent effects of testosterone on spatial learning strategies and brain-derived neurotrophic factor in male rats. Psychoneuroendocrinology 121, (2020). [CrossRef]
- Hill, R. A., Wu, Y. W. C., Kwek, P. & Van den Buuse, M. Modulatory Effects of Sex Steroid Hormones on Brain-Derived Neurotrophic Factor-Tyrosine Kinase B Expression during Adolescent Development in C57Bl/6 Mice. J. Neuroendocrinol. 24, 774–788 (2012). [CrossRef]
- Brague, J. C., Zinn, C. R., Granot, D. Y., Feathers, C. T. & Swann, J. M. TrkB is necessary for male copulatory behavior in the Syrian Hamster (Mesocricetus auratus). Horm. Behav. 97, 162–169 (2018). [CrossRef]
- Rankov Petrovic, B. et al. Prenatal Androgenization Induces Anxiety-Like Behavior in Female Rats, Associated with Reduction of Inhibitory Interneurons and Increased BDNF in Hippocampus and Cortex. Biomed Res. Int. 2019, (2019). [CrossRef]
- Lannigan, D. A. Estrogen receptor phosphorylation. Steroids 68, 1–9 (2003). [CrossRef]
- Anbalagan, M. & Rowan, B. G. Estrogen receptor alpha phosphorylation and its functional impact in human breast cancer. Mol. Cell. Endocrinol. 418, 264–272 (2015). [CrossRef]
- Frank, L. BDNF down-regulates neurotrophin responsiveness, TrkB protein and TrkB mRNA levels in cultured rat hippocampal neurons. Eur. J. Neurosci. 8, 1220–1230 (1996). [CrossRef]
- Anerillas, C. et al. A BDNF-TrkB autocrine loop enhances senescent cell viability. Nat. Commun. 13, 6228 (2022).
- Sohrabji, F., Miranda, R. C. G. & Toran-Allerand, C. D. Identification of a putative estrogen response element in the gene encoding brain-derived neurotrophic factor. Proc. Natl. Acad. Sci. U. S. A. 92, 11110–11114 (1995). [CrossRef]
- Mirabella, N. et al. Effects of castration on the expression of brain-derived neurotrophic factor (BDNF) in the vas deferens and male accessory genital glands of the rat. Cell Tissue Res. 323, 513–522 (2006). [CrossRef]
- Morita, K. & Her, S. Progesterone pretreatment enhances serotonin-stimulated BDNF gene expression in rat C6 glioma cells through production of 5α-reduced neurosteroids. J. Mol. Neurosci. 34, 193–200 (2008). [CrossRef]
- Sanchez-Bezanilla, S. et al. Growth Hormone Increases BDNF and mTOR Expression in Specific Brain Regions after Photothrombotic Stroke in Mice. Neural Plast. 2022, 1–13 (2022). [CrossRef]
- Eyileten, C. et al. The Relation of the Brain-Derived Neurotrophic Factor with MicroRNAs in Neurodegenerative Diseases and Ischemic Stroke. Mol. Neurobiol. 58, 329–347 (2021). [CrossRef]
- Koh, S. H. & Park, H. H. Neurogenesis in Stroke Recovery. Transl. Stroke Res. 8, 3–13 (2017). [CrossRef]
- Chen, A., Xingo, L., Tong, Y. & Mao, M. The neuroprotective roles of BDNF in hypoxic ischemic brain injury. Biomed. Reports 1, 167–176 (2013). [CrossRef]
- Cole, R., Pascal, L. E. & Wang, Z. The classical and updated models of androgen receptor nucleocytoplasmic trafficking. Am J Clin Exp Urol 9, 287–291 (2021).
- Morgan, R. L., Whaley, P., Thayer, K. A. & Schünemann, H. J. Identifying the PECO: A framework for formulating good questions to explore the association of environmental and other exposures with health outcomes. Environ. Int. 121, 1027–1031 (2018). [CrossRef]
Figure 1.
Overview of the sex steroids de novo synthesis within mitochondria. NOTES: Figure does not display all intermediate steroids, pathways, or enzymes. DHEA = dehydroepiandrosterone.
Figure 1.
Overview of the sex steroids de novo synthesis within mitochondria. NOTES: Figure does not display all intermediate steroids, pathways, or enzymes. DHEA = dehydroepiandrosterone.
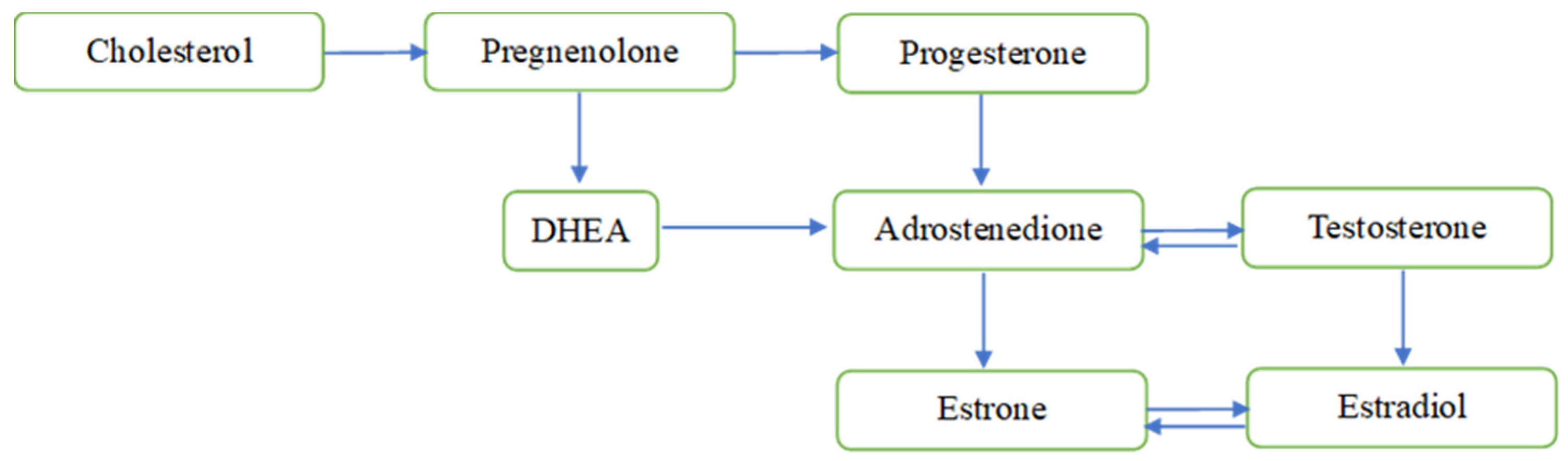
Table 1.
Data summary of interactions between Estrogens and BDNF.
| Title | Aim/experiment | Main Findings | |
|---|---|---|---|
| 1 | Interactions of estrogen with the neurotrophins and their receptors during neural development. DOI:10.1006/hbeh.1994.1033 |
To examine interactions between estrogen and neurotrophins in cortical neurons. | Cortical neurons co-express BDNF, p75 and TrkB, and basal forebrain neurons only express neurotrophin receptors. |
| 2 | The effect of ovariectomy and estrogen replacement on brain-derived neurotrophic factor messenger ribonucleic acid expression in cortical and hippocampal brain regions of female Sprague-Dawley rats. DOI:10.1210/endo.136.5.7720680 |
To investigate whether estradiol affects cholinergic function by modulating the levels of neurotrophic factors. | There was a reduction in BDNF levels in the rats frontal, parietal and temporal cortices 28 after OVX. |
| 3 | Levels of trkA and BDNF mRNA, but not NGF mRNA, fluctuate across the estrous cycle and increase in response to acute hormone replacement DOI:10.1016/S00068993(97)01511-4 |
To examine the levels of BDNF in mice hippocampus, and physiological changes in circulating sex steroids. | BDNF expression levels in CA1 and CA3/4 fluctuate across the estrous cycle. BDNF expression increased in the dentate granule cell layer, region CA1, and region CA3/4 in OVX animals 53 h after estrogen and 5 h after PROG treatment. |
| 4 | Brain-derived neurotrophic factor mediates estrogen-induced dendritic spine formation in hippocampal neurons DOI:10.1073/pnas.95.19.11412 |
To examine estrogen and BDNF regulation of glutamic acid decarboxylase expression in hippocampal cultures. | Estrogen decreases BDNF expression in hippocampal neurons within 24 hr, which suppress inhibition and increases the excitatory tone, leading to an increase in dendritic spine density in pyramidal neurons. |
| 5 | Region- and peptide-specific regulation of the neurotrophins by estrogen DOI: 10.1016/S0169-328X(00)00244-8 |
To examine the effect of estrogen on BDNF expression in the olfactory bulb and the cingulate cortex in OVX mice. | Estrogen regulation of BDNF is region-specific. It increases BDNF expression in mice olfactory bulb and diagonal band of Broca, but decreases in the cingulate cortex. |
| 6 | Neurotrophin expression in the reproductively senescent forebrain is refractory to estrogen stimulation DOI: 10.1016/S0197-4580(00)00230-X |
To compare estrogen regulation of neurotrophin ligands and receptors in young adult and senescent mice diagonal band of Broca. | Estrogen increases BDNF and TrKB expression in the olfactory bulb and horizontal limb; decreases p75NRT expression in young, and increases it in senescent mice. Senescent mice have higher ERα expression, but very low expression of the steroid receptor coactivator - SRC-1 in the olfactory bulb. |
| 7 | Expression and estrogen regulation of brain-derived neurotrophic factor gene and protein in the forebrain of female prairie voles DOI: 10.1002/cne.1156 |
To map the BDNF immunoreactive staining and mRNA labelling throughout the forebrain in female prairie voles. | Estrogen-treated mice have higher levels of BDNF in the DG and CA3 regions of the hippocampus, as well as in the basolateral nucleus of the amygdala, than controls. |
| 8 | Estrogen and exercise interact to regulate brain-derived neurotrophic factor mRNA and protein expression in the hippocampus DOI: 10.1046/j.0953-816x.2001.01825.x |
To investigate estrogen and exercise interaction in BDNF regulation. | Exercise increases hippocampal BDNF expression and protein levels in female mice, and this is reduced in the absence of estrogen, in a time-dependent manner. |
| 9 | Estrogen stimulates brain-derived neurotrophic factor expression in embryonic mouse midbrain neurons through a membrane-mediated and calcium-dependent mechanism DOI: 10.1002/jnr.1214 |
To investigate if estrogen influences on dopaminergic cells differentiation through a BDNF-dependent mechanism in mice midbrain. | Estrogen upregulates BDNF expression in mice midbrain; and has a stimulatory effect on dopaminergic neurons differentiation by coordinating BDNF expression. |
| 10 | Estrogen Regulates the Development of Brain-Derived Neurotrophic Factor mRNA and Protein in the Rat Hippocampus DOI: 10.1523/jneurosci.22-07-02650.2002 |
To examine gonadectomy and estrogen replacement effects on BDNF system in mice developing hippocampus. | ERα and BDNF co-localize in cells within developing hippocampus. BDNF expression levels reduce within 7-day in postnatal gonadectomized male rats, and estrogen treatment restores BDNF levels to intact animals. No changes were found in TrkB levels. |
| 11 | Estrogen enhances retrograde transport of Brain-Derived Neurotrophic Factor in the Rodent Forebrain DOI: 10.1210/en.2003-0724 |
To examine estrogen effect on the retrograde transport of BDNF in the diagonal band of Broca and its forebrain target in mice. | Estrogen-treated animals had greater numbers of neurons with retrogradely labelled BDNF than controls. |
| 12 | Influence of estradiol, stress, and 5-HT2A agonist treatment on brain-derived neurotrophic factor expression in female rats DOI: 10.1016/S0006-3223(03)00236-1 |
To examine the estrous cycle, and BDNF expression in the hippocampus and cortex of mice. | BDNF expression levels in the DG and the medial prefrontal cortex decrease when estradiol levels are highest. Acute estradiol treatment decreased hippocampal BDNF expression in acutely OVX, but had no effect in chronically OVX animals. |
| 13 | Oestrogen regulates sympathetic neurite outgrowth by modulating brain derived neurotrophic factor synthesis and release by the rodent uterus DOI: 10.1111/j.1460-9568.2003.03029.x |
To examine the role of neurotrophins and estrogen in uterine sympathetic nerve remodelling. | Estrogen increase BDNF expression and protein in the myometrium and endometrium of OVX mice. |
| 14 | Estrogen affects BDNF expression following chronic constriction nerve injury DOI: 10.1097/00001756-200308260-00017 |
To investigate the effect of estrogen on BDNF expression in neuropathic pain in a chronic constriction injury model of mice. | Mice with higher estrogen were more sensitive to thermal stimuli and higher levels of BDNF expression and protein levels. |
| 15 | Anatomical evidence for transsynaptic influences of estrogen on brain-derived neurotrophic factor expression DOI: 10.1002/cne.10989 |
To examined the localization of estrogen receptors and BDNF in the adult mice brain. | ERα and BDNF colocalize in the hypothalamus, amygdala, prelimbic cortex and ventral hippocampus. ERβ and BDNF do not colocalize in any brain regions. |
| 16 | Environmental enrichment reduces the mnemonic and neural benefits of estrogen DOI:10.1016/j.neuroscience.2004.06.011 |
To observed environmental factors that influence on mnemonic and neural response to estrogen in mice. | Estrogen decreased hippocampal BDNF in mice in standard, but not enriched condition. |
| 17 | Inhibition of tyrosine kinase receptor type B synthesis blocks axogenic effect of estrogen on rat hypothalamic neurones in vitro DOI:10.1111/j.1460-9568.2004.03485.x |
To examine the estrogen- induced axogenic response and upregulation of TrkB in neuronal and glial cultures of male mace. | Increase in TrkB is necessary for estrogen to exert its axogenic effect in male-derived neurones. |
| 18 | Effects of estrogen treatment on expression of brain-derived neurotrophic factor and cAMP response element-binding protein expression and phosphorylation in rat amygdaloid and hippocampal structures DOI:10.1159/000088448 |
To examine the estrogen effect of on CREB expression and phosphorylation, and BDNF expression, in the amygdala and hippocampus of mice. | Estrogen increased BDNF expression levels in mice amygdala, CA1, and CA3 regions of the hippocampus; and increased pCREB in the medial and basomedial, but not central or basolateral amygdala. |
| 19 | Estradiol to aged female or male mice improves learning in inhibitory avoidance and water maze tasks DOI:10.1016/j.brainres.2004.12.014 |
To evaluate the mnemonic effects of post-training estradiol in aged male mice. | BDNF levels decreased in the hippocampus of trained mice 1 h following estradiol exposure. |
| 20 | Sex and ovarian steroids modulate brain-derived neurotrophic factor (BDNF) protein levels in rat hippocampus under stressful and non-stressful conditions DOI:10.1016/j.psyneuen.2005.05.008 |
To investigate stress and sex hormones, and BDNF protein levels in CA1, CA3, and DG subregions of mice hippocampus. | Females have higher levels of BDNF in CA3, and lower levels in DG, relative to males. Stress decreases BDNF in CA3 in all animals. Stress increases BDNF levels in DG of PROG-treated OVX mice while decreases in controls. |
| 21 | 17β-estrogen Attenuates Hippocampal Neuronal Loss and Cognitive Dysfunction Induced By Chronic Restraint Stress in Ovariectomized Rats DOI:10.1016/j.neuroscience.2007.01.017 |
To evaluate the effect of estrogen on cognitive function in rodent under stress environment. | OVX or chronic stress decrease the levels of hippocampal BDNF expression in the CA3 region. Estrogen attenuates the stress-induced decrease in hippocampal BDNF expression levels in OVX rats. |
| 22 | Mode of action and functional significance of estrogen-inducing dendritic growth, spinogenesis, and synaptogenesis in the developing Purkinje cell DOI:10.1523/JNEUROSCI.0710-07.2007 |
To analyze the expression of BDNF and estrogen in Purkinje cells of neonatal and cytochrome P450 aromatase knock-out rodents. | Estrogen induces the expression of BDNF in mouse cerebella and promotes dendritic growth of Purkinje cells during development. |
| 23 | 17β-estrogen protects depletion of rat temporal cortex somatostatinergic system by β-amyloid DOI:10.1016/j.neurobiolaging.2006.06.009 |
To investigated estrogen treatment in amyloid-beta related changes in neuronal cells in the hippocampi of rodents. | Estrogen increased BDNF expression in hippocampal cells both in the absence or presence of estrogen. |
| 24 | Estrogen receptor β protects against acoustic trauma in mice DOI: 10.1172/JCI32796 |
To examine the role of ERs in response to auditory trauma. | BDNF expression was more pronounced in wild mice compared to ER deficient mice. |
| 25 | β-estrogen induces synaptogenesis in the hippocampus by enhancing brain-derived neurotrophic factor release from dentate gyrus granule cells DOI:10.1016/j.brainres.2007.02.093 |
To examined the effect of estrogen on synaptogenesis in hippocampal neuronal cell cultures. | The effects of estrogen in the hippocampal and subregional hippocampal neurons were independent of nuclear ERs and dependent on BDNF. Estrogen enhanced BDNF release from DG granule cells via nuclear ER-independent and PKA-dependent mechanisms. |
| 26 | Brain-derived neurotrophic factor signaling in the HVC is required for testosterone-induced song of female canaries DOI:10.1523/JNEUROSCI.2564-09.2009 |
To examine BDNF and T4-dependent development of the song system in female canaries. | T4-induced song is blocked by a concurrent inhibition of the vascular endothelial growth factor receptor tyrosine kinase, and this is reversed by BDNF. |
| 27 | Involvement of Brain-Derived Neurotrophic Factor and Neurogenesis in Oestrogen Neuroprotection of the Hippocampus of Hypertensive Rats. DOI: 10.1111/j.1365-2826.2010.02058.x |
To evaluate estrogen treatment in hypertensive rats and BDNF expression. | Hypertensive rats exhibit decreased expression and protein levels of BDNF in the DG, without changes in CA1 or CA3 pyramidal cell layers. Estrogen increases BDNF expression in the DG, and BDNF protein in whole hippocampus. |
| 28 | Estrogen reduces BDNF level, but maintains dopaminergic cell density in the striatum of MPTP mouse model. DOI:10.3109/00207451003721892 |
To examined the effects of estrogen treatment on BDNF expression and density of DA neurons in the striatum of MPTP mouse. | Estrogen impaired dopaminergic denervation and decreased the striatal BDNF upregulation triggered by MPTP. |
| 29 | Full length TrkB potentiates estrogen receptor alpha mediated transcription suggesting convergence of susceptibility pathways in schizophrenia. DOI:10.1016/j.mcn.2010.08.007 |
To examine ERα interaction with TrkB in neuronal and non-neuronal cell-lines. | TrkB activation increases transcription at EREs, independent of exogenous estrogen; and further potentiates the effect of estrogen-ERα-mediated transcription. |
| 30 | 17β-Estrogen replacement in young, adult and middle-aged female ovariectomized rats promotes improvement of spatial reference memory and an antidepressant effect and alters monoamines and BDNF levels in memory- and depression-related brain areas. DOI:10.1016/j.bbr.2011.10.047 |
To evaluated the effects of chronic treatment with estrogen on cognition and depressive-like behaviours in young, adult and middle-aged female rats. | Both young mice and estrogen treated OVX mice have higher BDNF levels. And the young estrogen treated OVX group presented higher BDNF levels compared to adult and middle-aged estrogen treated animals. |
| 31 | 17β-estrogen Regulates the Sexually Dimorphic Expression of BDNF and TrkB Proteins in the Song System of Juvenile Zebra Finches. DOI:10.1371/journal.pone.0043687 |
To examined BDNF isoforms and TrkB expression in the developing song system of juvenile males and females zebra finch treated with estrogen. | Estrogen modulates BDNF expression and TrkB in the song system of juveniles of both sexes. |
| 32 | Estradiol acts via estrogen receptors alpha and beta on pathways important for synaptic plasticity in the mouse hippocampal formation. DOI:10.1016/j.neuroscience.2011.11.035 |
To study estradiol systems and pathways related to plasticity and learning. | Estradiol increased phosphorylated Akt phosphorylated TrkB receptor in the hippocampus. These effects were abolished in ERα and ERβ knockout mice. |
| 33 | Estrone is neuroprotective in rats after traumatic brain injury. DOI:10.1089/neu.2011.2274 |
To studied the role of estrone in traumatic brain injury in rats. | Cortical levels of phospho-ERK1/2 are increased by estrone and this was associated with an increase in phospho- CREB levels and BDNF expression. |
| 34 | Estradiol promotes purkinje dendritic growth, spinogenesis, and synaptogenesis during neonatal life by inducing the expression of BDNF. DIO:10.1007/s12311-011-0342-6 |
To studied estradiol and cerebellar neuronal circuit formation, dendritic growth, spinogenesis, and synaptogenesis in the Purkinje cell of wild vs aromatase KO mice. | Estradiol increased neuroplasticity in all Purkinje cells. ER antagonist decreases BDNF levels in all mice. BDNF administration to ER antagonist- treated mice increased Purkinje dendritic growth. |
| 35 | Central expression and anorectic effect of brain-derived neurotrophic factor are regulated by circulating estrogen levels. DOI:10.1016/j.yhbeh.2013.01.009 |
To study if estradiol modulates the anorectic effect of BDNF in OVX rats. | BDNF expression is elevated in the hypothalamus during oestrus, following the estradiol peak, and after estradiol treatment. |
| 36 | Post-stroke infections exacerbate ischemic brain injury in middle-aged rats: Immunomodulation and neuroprotection by PROG. DOI:10.1016/j.neuroscience.2012.10.017 |
To evaluate the effect of systemic inflammation on stroke outcomes and PROG neuroprotection in middle-aged rats. | Serum BDNF levels decrease in systemic inflammation conditions. PROG decreases cytokine levels and systemic inflammation, and restores BDNF levels withing 3 and 7 days post-stroke. |
| 37 | Long-term OVX increases BDNF gene methylation status in mouse hippocampus. DOI: 10.1016/j.jsbmb.2014.08.001 |
To determine the post-OVX timeframe elapsed before E treatment is critical for the estrogen induction of neurotrophins BDNF in the rodent hippocampus. | Fast estrogen treated animals showed increased BDNF and expression and a higher activity of BDNF II, IV, and V promoters. Late treated animals did not show estrogen induction of neurotrophins and the methylation levels of the regulatory sequences of BDNF gene were higher than in the fast treated animals. |
| 38 | Oestrogen-Induced Neuroprotection in the Brain of Spontaneously Hypertensive Rats. DOI:10.1111/jne.12151 |
To investigate estrogen and pathological changes in the hippocampus and hypothalamus of hypertensive rats. | Estrogen treatment enhanced the number of cells and increased BDNF expression in the CA1 region and DG. |
| 39 | 17β-Estrogen regulates histone alterations associated with memory consolidation and increases Bdnf promoter acetylation in middle-aged female mice. DOI:10.1101/lm.034033.113 |
To examine the effects of estrogen infusion in the mice hippocampal on object recognition and spatial memory. | Estrogen specifically increased acetylation at BDNF promoters pII and pIV in the dorsal hippocampus of young and middle-aged mice, despite age-related decreases. |
| 40 | Chronic estrogen treatment decreases Brain Derived Neurotrophic Factor (BDNF) expression and monoamine levels in the amygdala - Implications for behavioral disorders. DOI:10.1016/j.bbr.2013.12.018 |
To verify whether chronic low estrogen dose cause anxiety-like disorder by altering BDNF and monoamine levels in rats hippocampus and amygdala. | Chronic estrogen treatment decreased BDNF expression and protein levels in the central amygdala that was accompanied by a reduction in dopamine levels. No changes were observed in the hippocampus. |
| 41 | Analyzing the influence of BDNF heterozygosity on spatial memory response to 17β-estrogen. DOI:10.1038/tp.2014.143 |
To test if disruption to the estrogen–parvalbumin pathway alters learning and memory, and BDNF levels in mice. | Estrogen replacement prevented reduction in BDNF and parvalbumin protein levels in the dorsal hippocampus and CA1. BDNF heterozygote mice showed either no response or an opposite response to estrogen treatment. |
| 42 | Aging-induced changes in sex-steroidogenic enzymes and sex-steroid receptors in the cortex, hypothalamus and cerebellum. DOI:10.1007/s12576-015-0363-x |
To examine age-induced changes in sex-steroidogenic enzymes and sex-steroid receptors in 3-, 12-, and 24-month-old male rats cerebral cortex, hypothalamus and cerebellum. | BDNF expression decreased from 3 to 24 m in the cerebral cortex, but increased in the hypothalamus and did not change in the cerebellum. The expression levels of AR, ERα and ERβ were higher in the Hypothalamus than in the cerebral cortex and Cerebellum. |
| 43 | ERα Signaling Is Required for TrkB-Mediated Hippocampal Neuroprotection in Female Neonatal Mice after Hypoxic Ischemic Encephalopathy. DOI:10.1523/ENEURO.0025-15.2015 |
To investigate how hypoxia induces ERα expression in female neonatal hippocampus. | TrkB phosphorylation post-trauma is greater in females than in males, after selective TrkB agonist therapy, and depends on the presence of ERα. TrkB agonist therapy decreases c-caspase-3, only in the presence of ERα. |
| 44 | Combined exercise ameliorates OVX-induced cognitive impairment by enhancing cell proliferation and suppressing apoptosis. DOI:10.1097/GME.0000000000000486 |
To evaluate the effects of exercise on memory deficits, cell proliferation and apoptosis in the hippocampus of OVX rats. | The expression of BDNF and TrkB decreased in the DG, together with memory decrease in OVX rats. These expression levels increased in the exercise group. |
| 45 | Selective Oestrogen Receptor Agonists Rescued Hippocampus Parameters in Male Spontaneously Hypertensive Rats. DOI:10.1111/jne.12415 |
To examine which type of ER is involved in low BDNF expression in the hippocampus of hypertensive rats. | ERα agonist slightly increased BDNF expression but had no effect on the number of doublecortin progenitors. Treatment with ERβ agonist increased BDNF expression and doublecortin progenitors. |
| 46 | Regulation of endometrial cell proliferation by estrogen-induced BDNF signaling pathway. DOI:10.1080/09513590.2017.1295439 |
To investigate the role of the BDNF Val66Met polymorphism in regulating proliferation in endometrial cells treated with estrogen. | BDNF signalling pathway activates with estrogen stimulation. BDNF production is induced by estrogen and the BDNF Val66Met is a loss-of-function polymorphism in the regulation of endometrial cells proliferation. |
| 47 | Sex differences in the effect of chronic mild stress on mouse prefrontal cortical BDNF levels: A role of major ovarian hormones. DOI:10.1016/j.neuroscience.2017.05.020 |
To examine the effect of sex hormones on depression-like phenotypes in mice exposed to a 21-day Chronic stress. | There was a decrease in the BDNF protein levels in intact females, but not in OVX or male mice. Estrogen treatment, and not PROG, increased BDNF expression in prefrontal cortex of the OVX stressed mice. |
| 48 | Sex differences and estrogen regulation of BDNF gene expression, but not propeptide content, in the developing hippocampus. DOI: 10.1002/jnr.23920 |
To examine the downstream effects of estrogen on hippocampal cell proliferation and BDNF expression in male and female neonatal rats. | Estrogen had opposite effects on BDNF expression in different areas of the neonatal hippocampus. In the CA1, BDNF increased, but decreased in DG. Blocking estrogen signaling decreased BDNF expression in the DG in males, but not females. No difference were observed in pro-BDNF protein. |
| 49 | Estrogen receptor β deficiency impairs BDNF–5-HT2A signaling in the hippocampus of female brain: A possible mechanism for menopausal depression. DOI:10.1016/j.psyneuen.2017.05.016 |
To examine the ER subtypes in the regulation of BDNF and serotonin signaling in mice. | BDNF was downregulated in ERβ−/− mice in a brain-region specific manner. There was a reduction in BDNF protein levels in the hippocampus of ERβ−/− mice, and no changes in the cortex and hypothalamus. ERβ agonism enhanced BDNF/TrkB signaling pathways. |
| 50 | Additive antidepressant-like effects of fasting with β-Estrogen in mice. DOI:10.1111/jcmm.14434 |
To evaluated the antidepressant-like effects of acute fasting and estrogen treatment. | BDNF-TrkB signalling pathway was increased in the prefrontal cortex and hippocampus. Serum ghrelin and estrogen were increased by fasting plus estrogen. |
| 51 | Voluntary exercise and estrogen reverse OVX-induced spatial learning and memory deficits and reduction in hippocampal brain-derived neurotrophic factor in rats. DOI:10.1016/j.pbb.2019.172819 |
To investigate voluntary exercise and estrogen replacement in learning and memory deficits and hippocampal BDNF levels in OVX mice. | Either exercise and estrogen alone or their combination recovery the negative effects of OVX on learning and memory performance. Combined treatment does not potentiate the effect of either treatment alone. |
| 52 | Sex-dependent aberrant prefrontal cortex development in the adolescent offspring rats exposed to variable prenatal stress. DOI: 10.1002/jdn.10034 |
To examine neurochemical and behavioural changes in the offspring rats (adolescences) from rats treated with prenatal stress during the third week of gestation. | Prenatal stress increase BDNF levels in prefrontal cortex adolescent females. ER concentration increased with age in all animals. |
| 53 | Rapid effects of estrogen and its receptor agonists on object recognition and object placement in adult male zebrafish. DOI:10.1016/j.bbr.2020.112514 |
To evaluate the effects of post-training estrogen in the consolidation of object recognition and object placement memory in adult male zebrafish. | Post-training estrogen treatment and selective ER agonist induced a upregulation in the BDNF expression levels. BDNF expression increase with high estrogen dose. |
| 54 | Reducing luteinizing hormone levels after OVX improves spatial memory: Possible role of brain-derived neurotrophic factor. DOI:10.1016/j.yhbeh.2019.104590 |
Exploited whether lowering luteinizing hormone increases BDNF expression levels. | Gonadotropin releasing hormone receptor antagonist lowers luteinizing hormone levels and estrogen enhances spatial memory in OVX females; all of which is ineffective in the absence of TrKB. Both hormones increase BDNF expression in the hippocampus. |
| 55 | ERRγ ligand HPB2 upregulates BDNF-TrkB and enhances dopaminergic neuronal phenotype. DOI:10.1016/j.phrs.2021.105423 |
To examine whether ERRγ ligand regulates BDNF signalling and subsequent DAergic neuronal phenotype. | ERRγ agonist increase BDNF expression and protein levels, and TrkB expression in human neuroblastoma, differentiated Lund mesencephalic cells, and primary ventral mesencephalic neurons. Activation of ERK and phosphorylation of CREB induced BDNF upregulation in human neuroblastoma cells. |
| 56 | Previous oestradiol treatment during midlife maintains transcriptional regulation of memory-related proteins by ERα in the hippocampus in a rat model of menopause. DOI:10.1016/j.neurobiolaging.2021.05.022 |
To examine whether previous estradiol treatment increases the levels of nuclear ERα, resulting in transcriptional regulation of proteins. | Continuous or previous estradiol treatments increase gene expression of BDNF. |
| 57 | 17β-Estrogen activation of dorsal hippocampal TrkB is independent of increased mature BDNF expression and is required for enhanced memory consolidation in female mice. DOI:10.1016/j.psyneuen.2020.105110 |
To examine the effects of hippocampal TrkB signalling on estrogen-induced enhancement of memory consolidation in the object placement and recognition tasks. | Dorsal hippocampal estrogen infusion increased levels of phospho-TrkB and BDNF. The Estrogen-induced increase in hippocampal BDNF expression was not required for hippocampal TrkB activation and was not inhibited by TrkB antagonism. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



