
Submitted:
07 January 2025
Posted:
08 January 2025
You are already at the latest version
Abstract
Background/Objectives: Cleft palate is a birth defect associated with environmental and genetic factors. It has been reported that disturbance of microRNAs (miRNAs) and exposure to medicinal agents during pregnancy can cause cleft palate. Although an association between medicine-induced cleft palate and miRNAs has been suggested, it remains to be fully elucidated. This study aimed to clarify the molecular mechanism underlying mycophenolate mofetil (MPM)-induced inhibition of cell proliferation and miRNA expression in human embryonic palatal mesenchymal (HEPM) cells. Methods: Cell viability, apoptosis, and cell cycle-related markers were evaluated 48 h after MPM treatment. In addition, miRNA levels and expression of their downstream genes were measured, and a rescue experiment was performed using miR-4680-3p and/or let-7c-5p inhibitors. Results: MPM dose-dependently reduced HEPM cell viability. Additionally, MPM treatment suppressed cyclin-D1, cyclin E1, cyclin-dependent kinase (CDK)-2, and CDK6 expression in HEPM cells. Furthermore, MPM upregulated miR-4680-3p and let-7c-5p expression and downregulated the downstream genes of each miRNA. Moreover, miR-4680-3p and/or let-7c-5p inhibitors alleviated MPM-induced inhibition of cell proliferation. Conclusions: These results suggest that MPM-induced cleft palate is associated with miR-4680-3p and let-7c-5p expression in HEPM cells.
Keywords:
mycophenolate mofetil
; cleft palate
; microRNA
; cell cycle
1. Introduction
Cleft palate (CP) is one of the most common birth defects worldwide, affecting approximately in 1 in every 500 babies in Asia. It has been reported that 70% of cleft lip cases with or without CP (CL/P) are non-syndromic, while the others are syndromic [1]. In humans, the palate forms in two stages, called primary and secondary palates. Secondary palate formation begins during the 6th week of gestation and is completed by approximately the 10th week. This happens through mesenchymal cell proliferation and epithelial cell fusion, wherein the two pieces of tissue, called palatal shelves, grow downward on either side of the tongue around the 6th–7th week. Then, around the 7th–8th week, the tongue is retracted between the palatal shelves, which then elevate and fuse above the tongue and primary palate. Finally, the medial epithelial seam breaks down through either apoptosis or migration toward the epithelial triangles on both the oral and nasal sides, or by epithelial-mesenchymal transition until 10th week. Any delay or alteration in apoptosis and/or cell cycle arrest during 6–10 weeks of gestation can result in CP [1].
The etiology of CP is associated with both genetic and environmental factors [2,3]. Several types of disruptions in the signaling pathway induced by the deletion of genes such those encoding as Sonic Hedgehog protein, WNT, bone morphogenetic protein, fibroblast growth factor, and transforming growth factor result in CP through the inhibition of cell viability [4,5]. Many genes are known to affect proliferation of palatal shelves or bind the each palatal shelf. Li et al reported that 131 genes involved with CP obtained from systematic review [6]. Environmental factors, such as taking medicine, taking alcohol, and smoking, are suggested to increase the risk for CP through the inhibition of crucial genes or signaling pathways [7]. A recent investigation demonstrated that new environmental factors such as PM2.5 are involved with CP have been reported [8,9]. However, the mechanisms that induce CP are not fully understood.
MicroRNAs (miRNAs) are single strand RNAs that negatively modulate gene expression by combining to 3’-untranslated regions, resulting in the inhibition of the protein translation or degradation of mRNA transcripts [10,11]. miRNAs play a vital effect in the formation of palatal shelves [12,13,14]. Recent studies have shown that specific miRNAs are associated with the mesenchymal palatal cell proliferation by modulating its downstream genes [15,16]. Moreover, our group and other researchers have reported that medicines such as all-trans retinoic acid, dexamethasone, phenytoin, and phenobarbital upregulated specific miRNAs and downregulated its downstream genes in human embryonic palatal mesenchymal (HEPM) cells and other related cell types [16,17,18,19]. However, as the literature focusing on miRNAs and medicine-induced CP is limited, other medicines may modulate miRNAs against medicine-induced CP.
Mycophenolate mofetil (MPM) is a mycophenolic acid prodrug that has been approved as an immunosuppressive agent. MPM has been prescribed as an alternative medicine for many autoimmune diseases such as nephrotic syndrome and lupus nephritis [20]. Although MPM shows potent immunosuppressive efficacy and fewer side effects than previously developed anti-proliferative immunosuppressive agents [21], several studies have reported that maternal exposure to MPM causes embryo teratogenicity, including CL/P and microtia [22,23]. Using the GeneCard database, Lin et al reported that several molecules (MDM2, RPL5, and TP53) may be involved in MPM-induced CL/P [24]. However, the involvement of miRNA regulation in MPM-induced CP remains unknown. In the present study, we investigated the involvement of miRNAs in MPM-induced inhibition of cell proliferation using HEPM cells.
2. Materials and Methods
2.1. Cell Culture
HEPM cells were purchased from American Type Culture Collection (CRL-1486, Manassas, VA, USA) and kept in Minimum Essential Medium Eagle-alpha modification medium (αMEM; Fujifilm-Wako Pure Chemical Corporation, Osaka, Japan) supplemented with 10% fetal bovine serum (Millipore-Sigma, St Louis, MO, USA), penicillin (10 U/mL), and streptomycin (10 μg/mL; Fujifilm-Wako Pure Chemical Corporation). The cells were kept at 37 °C in a humidified atmosphere containing 5% CO2.
2.2. Cell Viability Assay
HEPM cells were seeded in 5000 cells per 96 well plate (n=6) and treated with several concentrations (0–10 μM) of MPM (Tokyo Kasei Co. Ltd., Tokyo, Japan) after 24 h of cell seeding. After 48 h of treatment, the cell viability was measured using Alamar Blue (Bio-Rad Laboratories, Hercules, CA, USA).
2.3. Apoptosis Assay
HEPM cells were seeded in 10000 cells per 8-well chamber slides (Biomedical Sciences Inc., Tokyo, Japan) and treated with 1 μM MPM or vehicle after 24 h seeding. After 48 h of treatment, apoptosis-positive cells were calculated using ApoTracker Green (BioLegend, San Diego, CA, USA) according to our previous litearture [19]. Copper dichloride was used as a positive control [25] and Hoechst 33342 (Nacalai Tesque, Kyoto, Japan) was used as a nuclear counterstaining.
2.4. Western Blotting
HEPM cells were seeded in 200000 cells per 35 mm dish and treated with 1 μM MPM or vehicle after 24 h seeding. After 48 h of treatment, we washed PBS twice. To collect the protein, we added 100 μL ice-cold RIPA buffer (Nacalai Tesque) containing a protease inhibitor (Nacalai Tesque) and waited on ice. After 5 min, we scraped the cells and sonicated (15%, 5 sec, 3 times, Branson, Danbury, CR, USA). We subsequently centrifuged (20,000 × g for 20 min at 4°C) and collected supernatant as protein. The protein concentration was calculated as previously described [25,26,27]. Protein samples (10 μg) were applied to gradient (5-20 %) precast sodium dodecyl sulfate-polyacrylamide gel (ATTO, Tokyo, Japan) and transferred onto polyvinylidene difluoride (PVDF) membranes using Trans Turbo Blot (Bio-Rad Laboratories). The antibodies used are listed in Table 1. Band intensity was measured using Image J software (NIH, Bethesda, MD)
2.5. Bromodeoxyuridine (BrdU) Incorporation Assay
HEPM cells were seeded in 10000 cells per 8-well chamber slides (Biomedical Sciences Inc.) and treated with 1 μM MPM or vehicle (0.1% DMSO). After 48 h of treatment, the cells were incubated with BrdU (100 μg/mL, Sigma-Millipore) as we previously described [19].
2.6. Quantitative RT-PCR
HEPM cells were seeded in 200000 cells per 35 mm dish and treated with 1 μM MPM or vehicle after 24 h seeding. After 48 h of treatment, we washed PBS twice. To collect the total RNA, we extracted RNA from HEPM cells using a QIAshredder and miRNeasy Mini Kit (QIAGEN, Valencia, CA, USA) (n=3–5) [28,29]. miRNA expression was tested according to our previous investigation [30]. The expression level of target miRNAs was standardized to U6 expression levels.
2.7. Rescue Experiments Using miRNA Inhibitor
To rescue the effect of MPM, we used miRNA inhibitor was treated. We obtained miRNAs from BIONEER Ltd or Integrated DNA Technologies (IDT), respectively. HEPM cells were seeded in 5000 cells per 96 well plate (n=6). After 6 h of cell seeding, the HEPM cells were treated with hsa-7c-5p inhibitor (3 pmol; BIONEER Ltd., Daejeon, Korea), control miR inhibitor (3 pmol; BIONEER Ltd.), hsa-iR-4680-3p inhibitor (3 pmol: IDT, Coralville, IA, USA), or control miR inhibitor (3 pmol: IDT), using FuGENE SI Transfection Reagent (Promega, Madison, WI, USA), according to the manufacturer's protocol. Cells were treated with 1 μM MPM 24 h after transfection. After 48 h of treatment, cell viability was evaluated using Alamar Blue.
2.8. Statistical Analysis
Comparisons between two or more groups were conducted using Student’s t-test or Tukey’s test, respectively. All statistical analyses were conducted using IBM SPSS Statistics 26.0 for Windows (IBM Corp., Armonk, NY, USA). Values of P<0.05 were regarded statistically significant.
3. Results
3.1. MPM Inhibits Cell Proliferation via G1 Arrest in HEPM Cells
First, we performed a cell viability assay to determine whether an MPM-induced reduction in cell viability in HEPM cells. MPM treatment inhibited the viability of HEPM cells in a dose-dependent manner (Figure 1). We selected 1 μM for the following experiments since the suppression effect plateaued at this dose.
Proliferation of HEPM cells treated with MPM (0.01, 0.03, 0.1, 0.3, 1, 3, and 10 μM) for 48 h. Data are presented as the mean ± standard deviation (SD). *p < 0.05, **p < 0.01 and ***p < 0.001 versus control (n=6).
Several studies have reported that medicine-induced inhibition of mesenchymal palatal cell proliferation induces apoptosis and cell cycle arrest [14,31,32]. Therefore, we tested the effect of apoptosis and cell cycle arrest in MPM-treated HEPM cells. As shown in Figure 2A, treatment with MPM did not increase apoptosis-positive cells monitored by Apotracher, whereas copper dichloride (positive control) treatment activated apoptosis-positive cells. To support the result of Apotracker, we demonstrated that the protein expression of cleaved caspase-3, an indicator of apoptosis, was not increased by MPM treatment (Figure 2B).
Next, we monitored the cell cycle progression using a BrdU incorporation assay and found that the number of BrdU-positive cells was significantly reduced by MPM treatment (Figure 3A). To further investigate the molecular mechanisms underlying MPM-induced cell cycle arrest (G1-arrest), we tested cyclins and cyclin-dependent kinases (CDK) and found that MPM treatment reduced CCND1, CCNE, CDK2, and CDK6 levels, whereas those of CDK4 were comparable (Figure 3B). These results indicate that MPM induces cell cycle arrest (G1-arrest) by suppressing CCND1/CDK6 and CCNE/CDK2 in HEPM cells.
3.2. MPM Modulates let-7c-5p/miR-4680-3p and Its Downstream Genes in HEPM Cells.
Recently, miRNAs have been suggested to be associated with the CL/P etiology [12,13]. Suzuki and Li et al. identified miRNAs associated with human CP-related genes by performing systematic reviews, bioinformatic reviews, cell proliferation assays, and qPCR [6,33]. Moreover, Fu et al identified two miRNAs from a CL/P patient database and in vitro experiments [15]. In the present study, we measured the expression of seven miRNAs (let-7c-5p, miR-133b, miR-140-5p, miR-193a-3p, miR-374a-5p, miR-381-3p, and miR-4680-3p) using qPCR. We found that MPM treatment significantly increased let-7c-5p and miR-4680-3p expression, whereas the expression of the other five miRNAs was unaltered in HEPM cells (Figure 4A). To further investigate the effects of let-7c-5p and miR-4680-3p on downstream genes (BACH1 and PAX3 for let-7c-5p and ERBB2 and JADE1 for miR-4680-3p) [30,34], we conducted western blot analysis and found that MPM treatment suppressed BACH1, PAX3, ERBB2, and JADE levels (Figure 4B). These findings indicate that MPM modulates let-7c-5p and miR-4680-3p upregulation and inhibition of their downstream genes.
3.3. Inhibition of let-7c-5p and/or miR-4680-3p Alleviated MPM-Induced Cell Proliferation Activity in HEPM Cells.
To further investigate the contributions of let-7c-5p and miR-4680-3p, we transfected HEPM cells with let-7c-5p and/or miR-4680-3p inhibitors to examine whether let-7c-5p and/or miR-4680-3p alleviated the inhibition of HEPM cell proliferation following MPM treatment. Transfection with let-7c-5p and miR-4680-3p inhibitors suppressed expression of their respective targets by more than 80% under our experimental conditions (Figure 5A). Finally, we treated let-7c-5p and/or miR-4680-3p inhibitor whether MPM-induced cell proliferation inhibition was attenuated or not. We found that the let-7c-5p or miR-4680-3p inhibition partially alleviated the MPM-induced reduction in cell proliferation (Figure 5B). Moreover, treatment with both let-7c-5p and miR-4680-3p inhibitors fully protected MPM-induced cell proliferation inhibition in HEPM cells (Figure 5B). These results suggest that let-7c-5p and miR-4680-3p were associated with MPM-induced inhibition of HEPM cell proliferation.
4. Discussion
In this study, we examined the role of miRNAs in MPM-induced toxicity in HEPM cells. MPM treatment reduced the cell viability in a dose-dependent manner (Fig. 1) and downregulated the expression of CCND1, CCNE, CDK2, and CDK6 in HEPM cells (Fig. 3B). Several reports have suggested that MPM-induced toxicity derives from its inhibition purine synthesis (inosine-5’-monophosphate dehydrogenase) [35,36]; therefore, it is feasible that MPM induces cell cycle arrest (Fig. 3A, 3B). Additionally, MPM treatment induced let-7c-5p and miR-4680-3p expression and downregulated the expression of their downstream genes (Fig. 4B). Notably, the inhibition of let-7c-5p or miR-4680-3p partially alleviated MPM-induced cell inhibition and inhibition of both let-7c-5p and miR-4680-3p fully protected HEPM cells against MPM-induced cell proliferation inhibition (Fig. 5C).
Cyclins and CDKs play crucial roles in the regulation of cell cycle events [37]. When cells in the G0 phase enter the cell cycle, CDK4/CDK6 forms active complexes with CCND and other proteins, such as phosphorylated retinoblastoma protein (pRb), which activates the transition step from G1 to S phase [38]. Moreover, the CDK2 and CCNE complex phosphorylates and inactivates Rb family members [39]. The subsequent release of transcription factors (e.g., E2F) allows the cells to transition from G1 to S phase. Many cancers are known to increase CCND levels, and CCND activates signaling pathways, such as the MAPK kinase and PI3K/Akt pathways [40]. CCNE overexpression accelerates G1 phase progression in cancer patients [41]. CDK2 inhibition reduces the viability of human colon cancer cells [42]. CDK4/6 inhibitors, such as palbociclib, ribociclib, and abemaciclib have been approved for patients with HER2 negative breast cancer [43]. We demonstrated the number of BrdU-incorporated cells were significantly reduced with MPM treatment (Fig. 3A). Since the BrdU incorporation assay is an indicator of S phase, we investigated the molecular mechanism related to G1 phase proteins and found that MPM reduced CCND1, CDK6, CCNE, and CDK2 levels (Fig. 3B). Therefore, it is reasonable to conclude that MPM-induced cell viability reduction is G1-arrest through the suppression of CCND1/CDK6 and CCNE/CDK2 in HEPM cells.
Several studies have reported that miRNA-gene networks involved with craniofacial development [44,45]. Li et al. showed that overexpression and/or knock out mice of mmu-miR-17-92 cluster leads to CL and CP by modulating the BMP signaling pathway [46]. Polymorphisms in miR-140 and miR-4260 are associated with non-syndromic CL/P in humans [47,48]. Li and Suzuki et al. showed that several miRNAs (hsa-miR-133b, hsa-miR-140-5p, hsa-miR-374a-5p, hsa-miR-381a-3p, and hsa-miR-4680-3p) involved with development of human palate by combinating multi experiments such as systematic reviews, bioinformatics analyses, and cell viability assays [6,33]. Fu et al. reported that hsa-let-7c-5p and hsa-miR-193a-3p involved with CP using the patients with CP and in vitro experiment using HEPM cells [15]. We measured the seven (hsa-let-7c-5p, hsa-miR-133b, hsa-miR-140-5p, hsa-miR-193a-3p, hsa-miR-374a-5p, hsa-miR-381a-3p, and hsa-miR-4680-3p) miRNAs and demonstrated that MPM significantly induced let-7c-5p and miR-4680-3p expression level in HEPM cells (Fig. 4A). In addition, let-7c-5p or miR-4680-3p specific inhibitors partially alleviated and combined treatment with let-7c-5p and miR-4680-3p inhibitors fully protected against MPM-induced suppression of HEPM cell proliferation (Fig. 5B), indicating that let-7c-5p and miR-4680-3p play an important role in MPM-induced toxicity. Human let-7c-5p is located on chromosome-21 and let-7c-5p are involved in cell proliferation [49]. Overexpression of let-7c-5p suppresses human breast cancer and osteoblasts by downregulating CCDN1 [50,51]. Moreover, we previously reported that let-7c-5p and its inhibitor partially attenuated the phenobarbital-induced cell viability reduction by modulating BACH1 and PAX3 in HEPM cells [30]. Human miR-4680-3p is located on chromosome-10 and miR-4680-3p are expressed in gastric cancer [52]. miR-4680-3p and miR-4680-3p inhibitors partially attenuated the all-trans retinoic acid- and phenytoin-induced inhibition of cell proliferation through the regulation of ERBB2 and JADE1 [19,34]. Since these miRNAs are associated with several genes, they may play an important role in palate development by modulating downstream genes.
miRNAs negatively regulate downstream genes [53]. We previously identified that let-7c-5p inhibitor induces two CP-associated genes (BACH1 and PAX3) in humans [30]. As expected, both genes were downregulated following MPM treatment (Fig. 4B). BACH1 is ubiquitously expressed in mammals and is involved in multiple events, such as cell cycle and proliferation, through the modulation of the Wntfig/β-catenin signaling pathway [54,55]. PAX3 is essential for neural crest development. Knockout of PAX3 results in CP via downregulation of the BMP signaling pathway [56], and a PAX3 variant is associated with non-syndromic CL/P in humans [57]. As for miR-4680-3p, ERBB2 and JADE1 have been reported as downstream genes in HEPM cells [34]. We found that the expression of both genes was attenuated by MPM treatment in HEPM cells (Figure 4B). ERBB2 (HER2) is a member of the ERBB-receptor tyrosine kinase family, which includes epidermal growth factor receptor (EGFR). ERBB2 is a target in ERBB2 positive breast cancer [58]. ERBB2 is associated with cell proliferation, migration, and differentiation through the modulation of signaling cascades such as the MAPK/ERK and PI3K/AKT/mTOR pathways [59]. ERBB2 downregulation by all-trans retinoic acid or siRNA reduced cell viability via the ERK1/2 signaling pathway in HEPM cells [34]. JADE1 (PHF17) is a transcription factor and its inhibition reduces cell viability in cultured epithelial cell lines and primary fibroblasts [60]. JADE1 is known to regulate the Wnt/β-catenin signaling pathway [61]. JADE1 inhibition by siRNA reduces HEPM cell viability [34]. As these genes are associated with several signaling pathways related to proliferation, these miRNA-mRNA networks may play a crucial role in palate development by modulating these signaling pathways.
5. Conclusions
In conclusion, we demonstrated that MPM inhibits cell proliferation by inducing the expression of let-7c-5p and miR-4680-3p and downregulating their downstream genes. This is the first report to show the involvement of miRNAs in MPM-induced inhibition of palate cells. Since miRNAs are potential therapeutic agents [62], our present investigation may contribute to clinical protocols against CP development. Although further investigation is needed to understand how let-7c-5p and miR-4680-3p regulate the G1 phase, our findings may aid in understanding the etiology of CP.
Author Contributions
Conceptualization, H.Y. and K.O.; methodology, H.Y. and K.O.; software, H.K. and H.Y.; validation, H.H., H.Y. and Y.T.; formal analysis, Y.T. and H.Y.; investigation, H.H., A.O., and H.Y.; resources, A.O.. K.O. and H.Y.; data curation, Y.T, H.H. and H.Y.; writing—original draft preparation, Y.T. and H.H.; writing—review and editing, H.K., A.O., K.O. and H.Y.; visualization, H.K. and H.Y.; supervision, K.O. and H.Y.; project administration, H.Y.; funding acquisition, A.O. and H.Y. All authors have read and agreed to the published version of the manuscript.”.
Funding
This research was funded by Gifu University of Medical Science research grant A.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All relevant data are within the manuscript.
Acknowledgments
The authors thank Dr. Nobuaki Matsui and Dr. Naoki Inagaki (Gifu University of Medical Science, Japan) and Dr. Hyogo Horiguchi (Kitasato University, Japan) for his kind suggestions.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Babai, A.; Irving, M., Orofacial Clefts: Genetics of Cleft Lip and Palate. Genes (Basel) 2023, 14, (8). [CrossRef]
- Gonseth, S.; Shaw, G. M.; Roy, R.; Segal, M. R.; Asrani, K.; Rine, J.; Wiemels, J.; Marini, N. J., Epigenomic profiling of newborns with isolated orofacial clefts reveals widespread DNA methylation changes and implicates metastable epiallele regions in disease risk. Epigenetics 2019, 14, (2), 198-213. [CrossRef]
- Martinelli, M.; Palmieri, A.; Carinci, F.; Scapoli, L., Non-syndromic Cleft Palate: An Overview on Human Genetic and Environmental Risk Factors. Front Cell Dev Biol 2020, 8, 592271. [CrossRef]
- Yoshioka, H.; Tsukiboshi, Y.; Horita, H.; Kurita, H.; Ogata, A.; Ogata, K.; Horiguchi, H., Sasa veitchii extract alleviates phenobarbital-induced cell proliferation inhibition by upregulating transforming growth factor-beta 1. Tradit Kampo Med 2024, 11 (3), 192-199. [CrossRef]
- Meng, L.; Bian, Z.; Torensma, R.; Von den Hoff, J. W., Biological mechanisms in palatogenesis and cleft palate. J Dent Res 2009, 88, (1), 22-33. [CrossRef]
- Li, A.; Jia, P.; Mallik, S.; Fei, R.; Yoshioka, H.; Suzuki, A.; Iwata, J.; Zhao, Z., Critical microRNAs and regulatory motifs in cleft palate identified by a conserved miRNA-TF-gene network approach in humans and mice. Brief Bioinform 2020, 21, (4), 1465-1478.
- Vieira, A. R., Genetic and environmental factors in human cleft lip and palate. Front Oral Biol 2012, 16, 19-31.
- Ulschmid, C. M.; Sun, M. R.; Jabbarpour, C. R.; Steward, A. C.; Rivera-Gonzalez, K. S.; Cao, J.; Martin, A. A.; Barnes, M.; Wicklund, L.; Madrid, A.; Papale, L. A.; Joseph, D. B.; Vezina, C. M.; Alisch, R. S.; Lipinski, R. J., Disruption of DNA methylation-mediated cranial neural crest proliferation and differentiation causes orofacial clefts in mice. Proc Natl Acad Sci U S A 2024, 121, (3), e2317668121. [CrossRef]
- Zhou, Y.; Gilboa, S. M.; Herdt, M. L.; Lupo, P. J.; Flanders, W. D.; Liu, Y.; Shin, M.; Canfield, M. A.; Kirby, R. S., Maternal exposure to ozone and PM(2.5) and the prevalence of orofacial clefts in four U.S. states. Environ Res 2017, 153, 35-40.
- Bartel, D. P., MicroRNAs: target recognition and regulatory functions. Cell 2009, 136, (2), 215-33. [CrossRef]
- Wang, W.; Chen, X.; Li, C.; Zhao, R.; Zhang, J.; Qin, H.; Wang, M.; Su, Y.; Tang, M.; Han, L.; Sun, N., The single nucleotide polymorphism rs1814521 in long non-coding RNA ADGRG3 associates with the susceptibility to silicosis: a multi-stage study. Environ Health Prev Med 2022, 27, 5. [CrossRef]
- Schoen, C.; Aschrafi, A.; Thonissen, M.; Poelmans, G.; Von den Hoff, J. W.; Carels, C. E. L., MicroRNAs in Palatogenesis and Cleft Palate. Front Physiol 2017, 8, 165. [CrossRef]
- Wang, S.; Sun, C.; Meng, Y.; Zhang, B.; Wang, X.; Su, Y.; Shi, L.; Zhao, E., A pilot study: Screening target miRNAs in tissue of nonsyndromic cleft lip with or without cleft palate. Exp Ther Med 2017, 13, (5), 2570-2576. [CrossRef]
- Yoshioka, H.; Suzuki, A.; Iwaya, C.; Iwata, J., Suppression of microRNA 124-3p and microRNA 340-5p ameliorates retinoic acid-induced cleft palate in mice. Development 2022, 149, (9), dex200476. [CrossRef]
- Fu, C.; Lou, S.; Zhu, G.; Fan, L.; Yu, X.; Zhu, W.; Ma, L.; Wang, L.; Pan, Y., Identification of New miRNA-mRNA Networks in the Development of Non-syndromic Cleft Lip With or Without Cleft Palate. Front Cell Dev Biol 2021, 9, 631057.
- Yoshioka, H.; Jun, G.; Suzuki, A.; Iwata, J., Dexamethasone Suppresses Palatal Cell Proliferation through miR-130a-3p. Int J Mol Sci 2021, 22, (22) 12453. [CrossRef]
- Zhang, W.; Shen, Z.; Xing, Y.; Zhao, H.; Liang, Y.; Chen, J.; Zhong, X.; Shi, L.; Wan, X.; Zhou, J.; Tang, S., MiR-106a-5p modulates apoptosis and metabonomics changes by TGF-beta/Smad signaling pathway in cleft palate. Exp Cell Res 2020, 386, (2), 111734.
- Yoshioka, H.; Ramakrishnan, S. S.; Suzuki, A.; Iwata, J., Phenytoin Inhibits Cell Proliferation through microRNA-196a-5p in Mouse Lip Mesenchymal Cells. Int J Mol Sci 2021, 22, (4) 1746. [CrossRef]
- Tsukiboshi, Y.; Horita, H.; Mikami, Y.; Noguchi, A.; Yokota, S.; Ogata, K.; Yoshioka, H., Involvement of microRNA-4680-3p against phenytoin-induced cell proliferation inhibition in human palate cells. J Toxicol Sci 2024, 49, (1), 1-8. [CrossRef]
- Appel, G. B.; Radhakrishnan, J.; Ginzler, E. M., Use of mycophenolate mofetil in autoimmune and renal diseases. Transplantation 2005, 80, (2 Suppl), S265-71. [CrossRef]
- Perez-Aytes, A.; Marin-Reina, P.; Boso, V.; Ledo, A.; Carey, J. C.; Vento, M., Mycophenolate mofetil embryopathy: A newly recognized teratogenic syndrome. Eur J Med Genet 2017, 60, (1), 16-21. [CrossRef]
- Coscia, L. A.; Armenti, D. P.; King, R. W.; Sifontis, N. M.; Constantinescu, S.; Moritz, M. J., Update on the Teratogenicity of Maternal Mycophenolate Mofetil. J Pediatr Genet 2015, 4, (2), 42-55. [CrossRef]
- Tjeertes, I. F.; Bastiaans, D. E.; van Ganzewinkel, C. J.; Zegers, S. H., Neonatal anemia and hydrops fetalis after maternal mycophenolate mofetil use. J Perinatol 2007, 27, (1), 62-4. [CrossRef]
- Lin, Y.; Song, T.; Ronde, E. M.; Ma, G.; Cui, H.; Xu, M., The important role of MDM2, RPL5, and TP53 in mycophenolic acid-induced cleft lip and palate. Medicine (Baltimore) 2021, 100, (21), e26101. [CrossRef]
- Tominaga, S.; Yoshioka, H.; Yokota, S.; Tsukiboshi, Y.; Suzui, M.; Nagai, M.; Hara, H.; Maeda, T.; Miura, N., Copper-induced diurnal hepatic toxicity is associated with Cry2 and Per1 in mice. Environ Health Prev Med 2023, 28, 78. [CrossRef]
- Yoshioka, H.; Tominaga, S.; Suzui, M.; Shinohara, Y.; Maeda, T.; Miura, N., Involvement of Npas2 and Per2 modifications in zinc-induced acute diurnal toxicity in mice. J Toxicol Sci 2022, 47, (12), 547-553. [CrossRef]
- Yoshioka, H.; Tominaga, S.; Amano, F.; Wu, S.; Torimoto, S.; Moriishi, T.; Tsukiboshi, Y.; Yokota, S.; Miura, N.; Inagaki, N.; Matsushita, Y.; Maeda, T., Juzentaihoto alleviates cisplatin-induced renal injury in mice. Tradit Kampo Med 2024, 11, (2), 147-155. [CrossRef]
- Yoshioka, H.; Wu, S.; Moriishi, T.; Tsukiboshi, Y.; Yokota, S.; Miura, N.; Yoshikawa, M.; Inagaki, N.; Matsushita, Y.; Nakao, M., Sasa veitchii extract alleviates nonalcoholic steatohepatitis in methionine–choline deficient diet-induced mice by regulating peroxisome proliferator-activated receptor alpha. Tradit Kampo Mede 2023, 10, (3), 259-268. [CrossRef]
- Yoshioka, H.; Yokota, S.; Tominaga, S.; Tsukiboshi, Y.; Suzui, M.; Shinohara, Y.; Yoshikawa, M.; Sasaki, H.; Sasaki, N.; Maeda, T.; Miura, N., Involvement of Bmal1 and Clock in Bromobenzene Metabolite-Induced Diurnal Renal Toxicity. Biol Pharm Bull 2023, 46, (6), 824-829. [CrossRef]
- Tsukiboshi, Y.; Noguchi, A.; Horita, H.; Mikami, Y.; Yokota, S.; Ogata, K.; Yoshioka, H., Let-7c-5p associate with inhibition of phenobarbital-induced cell proliferation in human palate cells. Biochem Biophys Res Commun 2024, 696, 149516. [CrossRef]
- Dhulipala, V. C.; Welshons, W. V.; Reddy, C. S., Cell cycle proteins in normal and chemically induced abnormal secondary palate development: a review. Hum Exp Toxicol 2006, 25, (11), 675-82. [CrossRef]
- Smane, L.; Pilmane, M.; Akota, I., Apoptosis and MMP-2, TIMP-2 expression in cleft lip and palate. Stomatologija 2013, 15, (4), 129-34.
- Suzuki, A.; Li, A.; Gajera, M.; Abdallah, N.; Zhang, M.; Zhao, Z.; Iwata, J., MicroRNA-374a, -4680, and -133b suppress cell proliferation through the regulation of genes associated with human cleft palate in cultured human palate cells. BMC Med Genomics 2019, 12, (1), 93. [CrossRef]
- Yoshioka, H.; Ramakrishnan, S. S.; Shim, J.; Suzuki, A.; Iwata, J., Excessive All-Trans Retinoic Acid Inhibits Cell Proliferation Through Upregulated MicroRNA-4680-3p in Cultured Human Palate Cells. Front Cell Dev Biol 2021, 9, 618876. [CrossRef]
- Schmidt, F.; Eckardt, K.; Shakibaei, M.; Glander, P.; Stahlmann, R., Effects of mycophenolic acid alone and in combination with its metabolite mycophenolic acid glucuronide on rat embryos in vitro. Arch Toxicol 2013, 87, (2), 361-70. [CrossRef]
- Jiang, L. L.; Liu, M. H.; Li, J. Y.; He, Z. H.; Li, H.; Shen, N.; Wei, P.; He, M. F., Mycophenolic Acid-Induced Developmental Defects in Zebrafish Embryos. Int J Toxicol 2016, 35, (6), 712-718. [CrossRef]
- Sherr, C. J.; Roberts, J. M., Living with or without cyclins and cyclin-dependent kinases. Genes Dev 2004, 18, (22), 2699-711. [CrossRef]
- Fassl, A.; Geng, Y.; Sicinski, P., CDK4 and CDK6 kinases: From basic science to cancer therapy. Science 2022, 375, (6577), eabc1495. [CrossRef]
- Ettl, T.; Schulz, D.; Bauer, R. J., The Renaissance of Cyclin Dependent Kinase Inhibitors. Cancers (Basel) 2022, 14, (2). [CrossRef]
- Lukasik, P.; Zaluski, M.; Gutowska, I., Cyclin-Dependent Kinases (CDK) and Their Role in Diseases Development-Review. Int J Mol Sci 2021, 22, (6), 2935. [CrossRef]
- Ohtsubo, M.; Theodoras, A. M.; Schumacher, J.; Roberts, J. M.; Pagano, M., Human cyclin E, a nuclear protein essential for the G1-to-S phase transition. Mol Cell Biol 1995, 15, (5), 2612-24. [CrossRef]
- Lee, Y. M.; Sicinski, P., Targeting cyclins and cyclin-dependent kinases in cancer: lessons from mice, hopes for therapeutic applications in human. Cell Cycle 2006, 5, (18), 2110-4.
- Huang, J.; Zheng, L.; Sun, Z.; Li, J., CDK4/6 inhibitor resistance mechanisms and treatment strategies (Review). Int J Mol Med 2022, 50, (4), 128.
- Garland, M. A.; Sun, B.; Zhang, S.; Reynolds, K.; Ji, Y.; Zhou, C. J., Role of epigenetics and miRNAs in orofacial clefts. Birth Defects Res 2020, 112, (19), 1635-1659. [CrossRef]
- Iwata, J., Gene-Environment Interplay and MicroRNAs in Cleft Lip and Cleft Palate. Oral Sci Int 2021, 18, (1), 3-13. [CrossRef]
- Wang, J.; Bai, Y.; Li, H.; Greene, S. B.; Klysik, E.; Yu, W.; Schwartz, R. J.; Williams, T. J.; Martin, J. F., MicroRNA-17-92, a direct Ap-2alpha transcriptional target, modulates T-box factor activity in orofacial clefting. PLoS Genet 2013, 9, (9), e1003785.
- Li, L.; Meng, T.; Jia, Z.; Zhu, G.; Shi, B., Single nucleotide polymorphism associated with nonsyndromic cleft palate influences the processing of miR-140. Am J Med Genet A 2010, 152A, (4), 856-62. [CrossRef]
- Zhu, G.; Zhang, C.; Wang, Y.; Wang, Y.; Li, D.; Yu, X.; Zhu, W.; Fu, C.; Lou, S.; Fan, L.; Ma, L.; Wang, L.; Pan, Y., Variants in miRNA regulome and their association with the risk of nonsyndromic orofacial clefts. Epigenomics 2020, 12, (13), 1109-1121. [CrossRef]
- Johnson, C. D.; Esquela-Kerscher, A.; Stefani, G.; Byrom, M.; Kelnar, K.; Ovcharenko, D.; Wilson, M.; Wang, X.; Shelton, J.; Shingara, J.; Chin, L.; Brown, D.; Slack, F. J., The let-7 microRNA represses cell proliferation pathways in human cells. Cancer Res 2007, 67, (16), 7713-22.
- Fu, X.; Mao, X.; Wang, Y.; Ding, X.; Li, Y., Let-7c-5p inhibits cell proliferation and induces cell apoptosis by targeting ERCC6 in breast cancer. Oncol Rep 2017, 38, (3), 1851-1856. [CrossRef]
- Luo, K.; Qin, Y.; Ouyang, T.; Wang, X.; Zhang, A.; Luo, P.; Pan, X., Let-7c-5p Regulates CyclinD1 in Fluoride-Mediated Osteoblast Proliferation and Activation. Toxicol Sci 2021, 182, (2), 275-287. [CrossRef]
- Xing, Y.; Chen, H.; Guo, Z.; Zhou, X., Circular RNA circ0007360 Attenuates Gastric Cancer Progression by Altering the miR-762/IRF7 Axis. Front Cell Dev Biol 2022, 10, 789073.
- Valinezhad Orang, A.; Safaralizadeh, R.; Kazemzadeh-Bavili, M., Mechanisms of miRNA-Mediated Gene Regulation from Common Downregulation to mRNA-Specific Upregulation. Int J Genomics 2014, 2014, 970607.
- Zhang, X.; Guo, J.; Wei, X.; Niu, C.; Jia, M.; Li, Q.; Meng, D., Bach1: Function, Regulation, and Involvement in Disease. Oxid Med Cell Longev 2018, 2018, 1347969.
- Liu, C.; Yu, J.; Liu, B.; Liu, M.; Song, G.; Zhu, L.; Peng, B., BACH1 regulates the proliferation and odontoblastic differentiation of human dental pulp stem cells. BMC Oral Health 2022, 22, (1), 536. [CrossRef]
- Wu, M.; Li, J.; Engleka, K. A.; Zhou, B.; Lu, M. M.; Plotkin, J. B.; Epstein, J. A., Persistent expression of Pax3 in the neural crest causes cleft palate and defective osteogenesis in mice. J Clin Invest 2008, 118, (6), 2076-87. [CrossRef]
- Liang, W.; Huang, W.; Sun, B.; Zhong, W.; Zhang, Y.; Zhang, J.; Zhou, Z.; Lin, J.; Chen, F., A Novel PAX3 Variant in a Chinese Pedigree with Nonsyndromic Cleft Lip With or Without Palate. Genet Test Mol Biomarkers 2021, 25, (12), 749-756. [CrossRef]
- Tan, M.; Yu, D., Molecular mechanisms of erbB2-mediated breast cancer chemoresistance. Adv Exp Med Biol 2007, 608, 119-29.
- Kirouac, D. C.; Du, J.; Lahdenranta, J.; Onsum, M. D.; Nielsen, U. B.; Schoeberl, B.; McDonagh, C. F., HER2+ Cancer Cell Dependence on PI3K vs. MAPK Signaling Axes Is Determined by Expression of EGFR, ERBB3 and CDKN1B. PLoS Comput Biol 2016, 12, (4), e1004827.
- Havasi, A.; Haegele, J. A.; Gall, J. M.; Blackmon, S.; Ichimura, T.; Bonegio, R. G.; Panchenko, M. V., Histone acetyl transferase (HAT) HBO1 and JADE1 in epithelial cell regeneration. Am J Pathol 2013, 182, (1), 152-62. [CrossRef]
- Borgal, L.; Habbig, S.; Hatzold, J.; Liebau, M. C.; Dafinger, C.; Sacarea, I.; Hammerschmidt, M.; Benzing, T.; Schermer, B., The ciliary protein nephrocystin-4 translocates the canonical Wnt regulator Jade-1 to the nucleus to negatively regulate beta-catenin signaling. J Biol Chem 2012, 287, (30), 25370-80.
- Lu, T. X.; Rothenberg, M. E., MicroRNA. J Allergy Clin Immunol 2018, 141, (4), 1202-1207.
Figure 1.
MPM inhibited cell proliferation in HEPM cells.
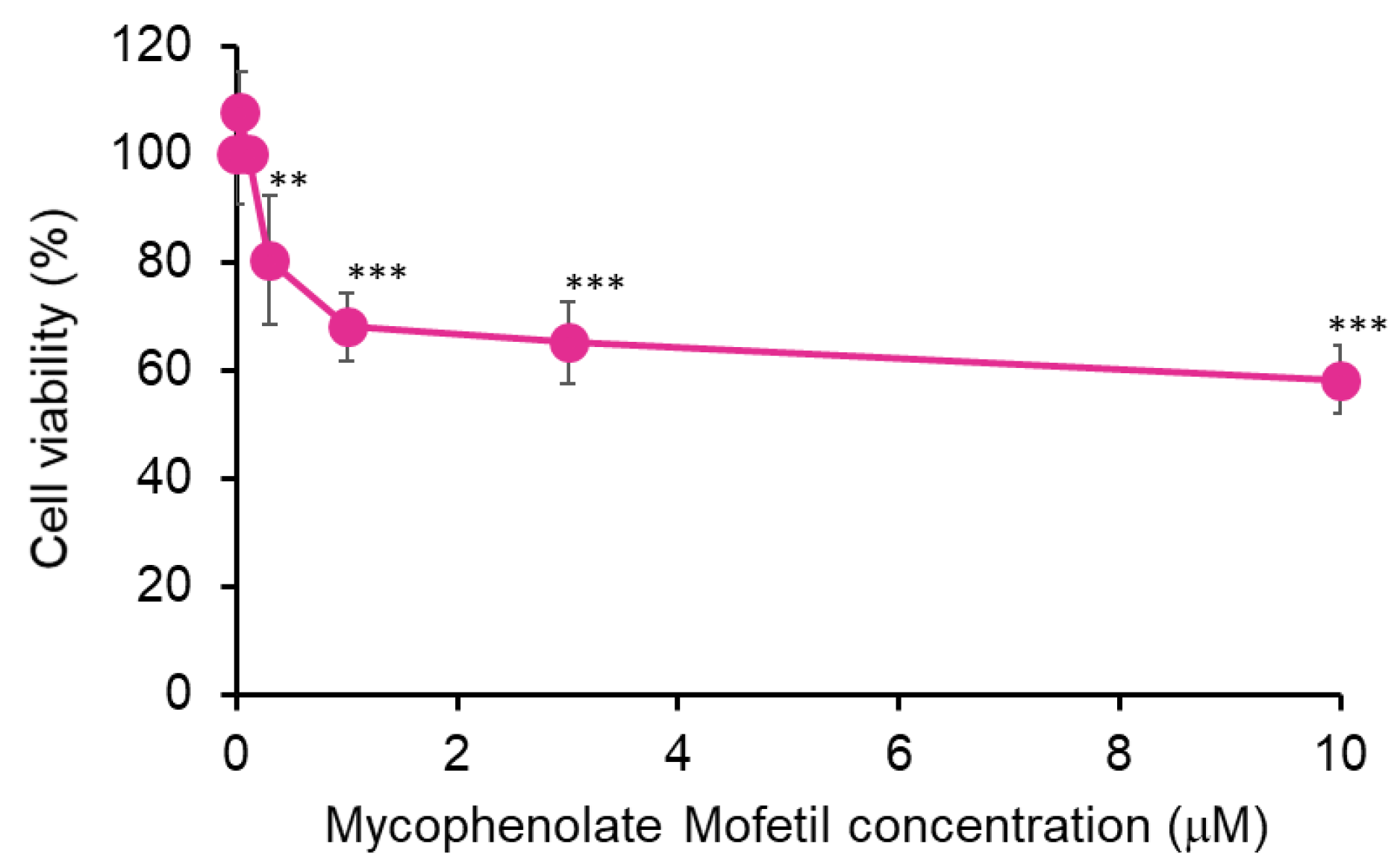
Figure 2.
MPM-induced cell proliferation is not associated with apoptosis in HEPM cells. (A)Apotracker staining of HEPM cells after treatment with 1 μM MPM for 48 h. The nuclei were counterstained with Hoechst 33342. Copper dichloride (500 μM 24 h) was used as a positive control. Scale bar, 50 μm. (B) Western blotting of HEPM cells treated with 1 μM MPM for 48 h. β-ACTIN was served as an internal control. Copper dichloride (500 μM M 24 h) was used as the positive control (P.C.).
Figure 2.
MPM-induced cell proliferation is not associated with apoptosis in HEPM cells. (A)Apotracker staining of HEPM cells after treatment with 1 μM MPM for 48 h. The nuclei were counterstained with Hoechst 33342. Copper dichloride (500 μM 24 h) was used as a positive control. Scale bar, 50 μm. (B) Western blotting of HEPM cells treated with 1 μM MPM for 48 h. β-ACTIN was served as an internal control. Copper dichloride (500 μM M 24 h) was used as the positive control (P.C.).
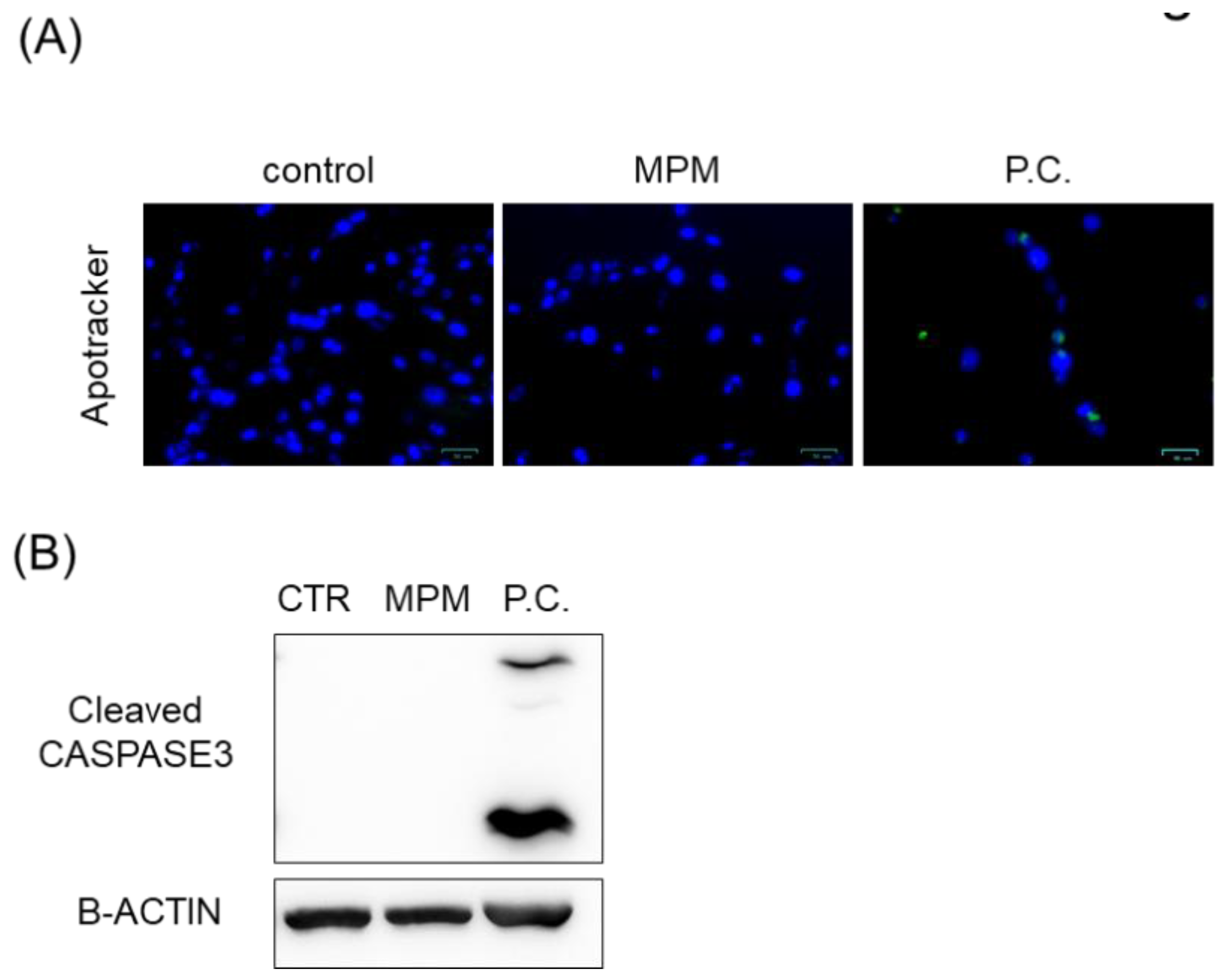
Figure 3.
MPM-induced cell proliferation is associated with G1 arrest in HEPM cells. (A) BrdU staining (green) of HEPM cells after treatment with 1 μM MPM for 48 h. The nuclei were counterstained with Hoechst 33342 (blue). Scale bar, 50 μm. Graph shows the quantification of BrdU positive cells. Data are presented as the mean ± standard deviation (SD). **p < 0.01 (n=6). (B) Western blotting of HEPM cells treated with 1 μM MPM for 48 h. β-ACTIN was served as an internal control. *p < 0.05, **p < 0.01, ***p < 0.001 (n=3).
Figure 3.
MPM-induced cell proliferation is associated with G1 arrest in HEPM cells. (A) BrdU staining (green) of HEPM cells after treatment with 1 μM MPM for 48 h. The nuclei were counterstained with Hoechst 33342 (blue). Scale bar, 50 μm. Graph shows the quantification of BrdU positive cells. Data are presented as the mean ± standard deviation (SD). **p < 0.01 (n=6). (B) Western blotting of HEPM cells treated with 1 μM MPM for 48 h. β-ACTIN was served as an internal control. *p < 0.05, **p < 0.01, ***p < 0.001 (n=3).
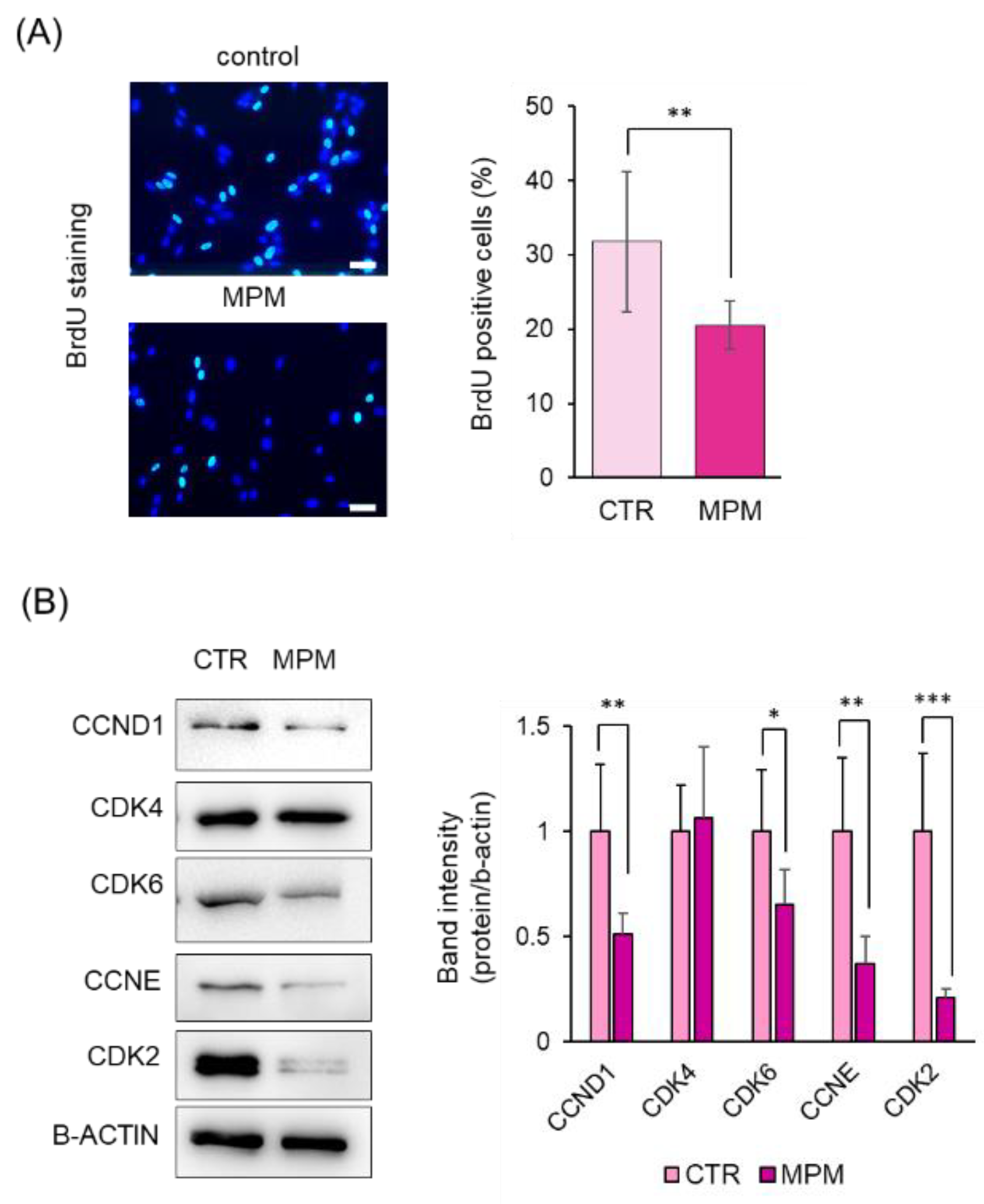
Figure 4.
MPM upregulated let-7c-5p and miR-4680-3p in HEPM cells. (A)Quantitative RT-PCR of let-7c-5p, miR-133b, miR-140-5p, miR-193a-3p, miR-374a-5p, miR-381-3p, and miR-4680-3p after treatment of HEPM cells with 1 μM MPM for 48 h. Data are presented as the mean ± standard deviation (SD). **p < 0.01 and ***p < 0.001 (n=3). (B) Western blotting of HEPM cells treated with 1 μM MPM for 48 h. β-ACTIN was served as an internal control. *p < 0.05, **p < 0.01, ***p < 0.001 (n=3).
Figure 4.
MPM upregulated let-7c-5p and miR-4680-3p in HEPM cells. (A)Quantitative RT-PCR of let-7c-5p, miR-133b, miR-140-5p, miR-193a-3p, miR-374a-5p, miR-381-3p, and miR-4680-3p after treatment of HEPM cells with 1 μM MPM for 48 h. Data are presented as the mean ± standard deviation (SD). **p < 0.01 and ***p < 0.001 (n=3). (B) Western blotting of HEPM cells treated with 1 μM MPM for 48 h. β-ACTIN was served as an internal control. *p < 0.05, **p < 0.01, ***p < 0.001 (n=3).
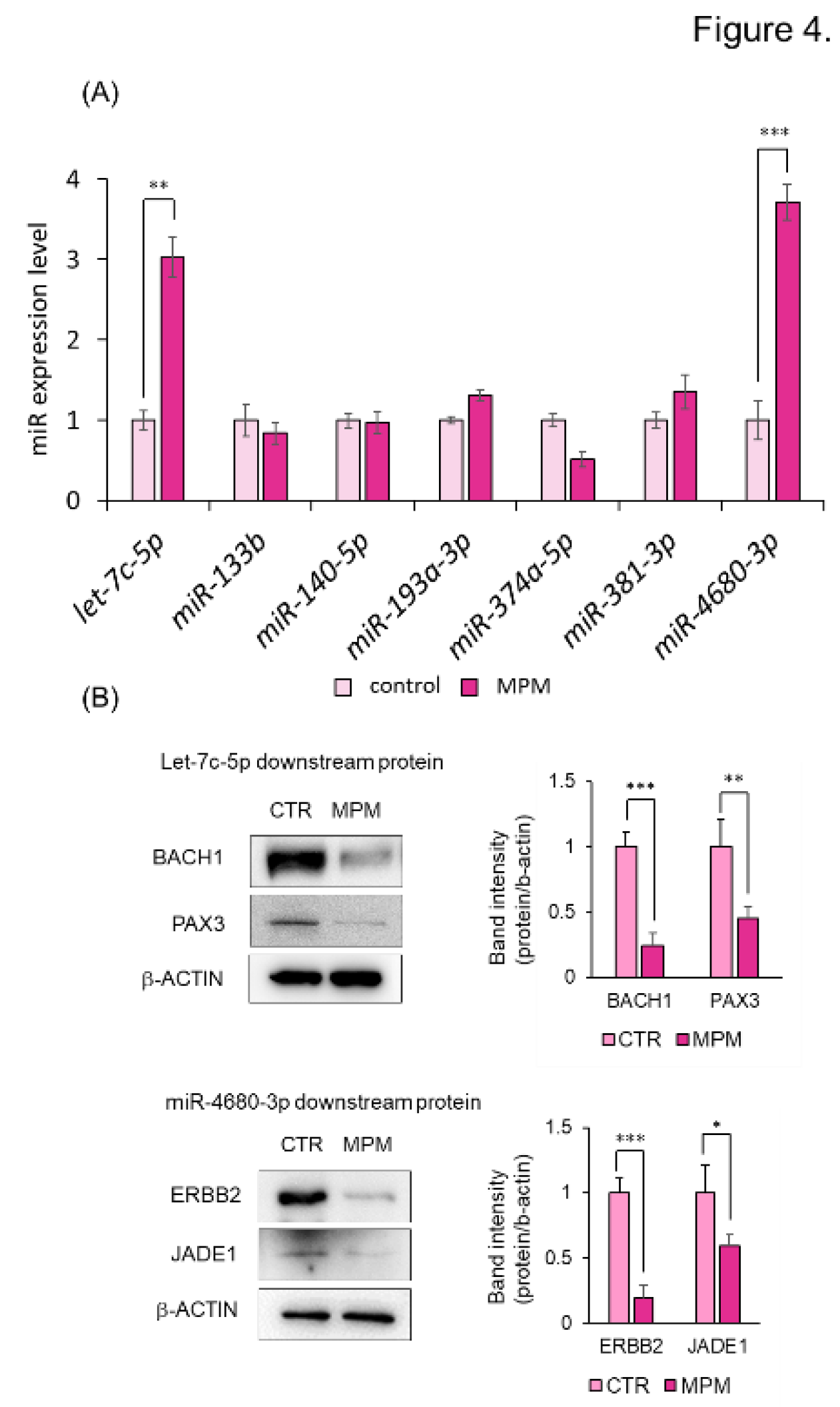
Figure 5.
Inhibition of let-7c-5p and miR-4680-3p alleviated MPM-induced cell proliferation inhibition in HEPM cells. (A)Quantitative RT-PCR analysis of let-7c-5p or miR-4680-3p expression after transfected HEPM cells with let-7c-5p inhibitor or miR-4680-3p inhibitor for 24 h. Data are presented as the mean ± standard deviation (SD). ***p < 0.001 (n=3). (B) Proliferation of HEPM cells treated with 1 μM MPM and/or let-7c-5p inhibitor and/or miR-4680-3p inhibitor for 48 h. Data are presented as the mean ± standard deviation (SD). *p < 0.05, **p < 0.01 and ***p < 0.001 (n=6).
Figure 5.
Inhibition of let-7c-5p and miR-4680-3p alleviated MPM-induced cell proliferation inhibition in HEPM cells. (A)Quantitative RT-PCR analysis of let-7c-5p or miR-4680-3p expression after transfected HEPM cells with let-7c-5p inhibitor or miR-4680-3p inhibitor for 24 h. Data are presented as the mean ± standard deviation (SD). ***p < 0.001 (n=3). (B) Proliferation of HEPM cells treated with 1 μM MPM and/or let-7c-5p inhibitor and/or miR-4680-3p inhibitor for 48 h. Data are presented as the mean ± standard deviation (SD). *p < 0.05, **p < 0.01 and ***p < 0.001 (n=6).
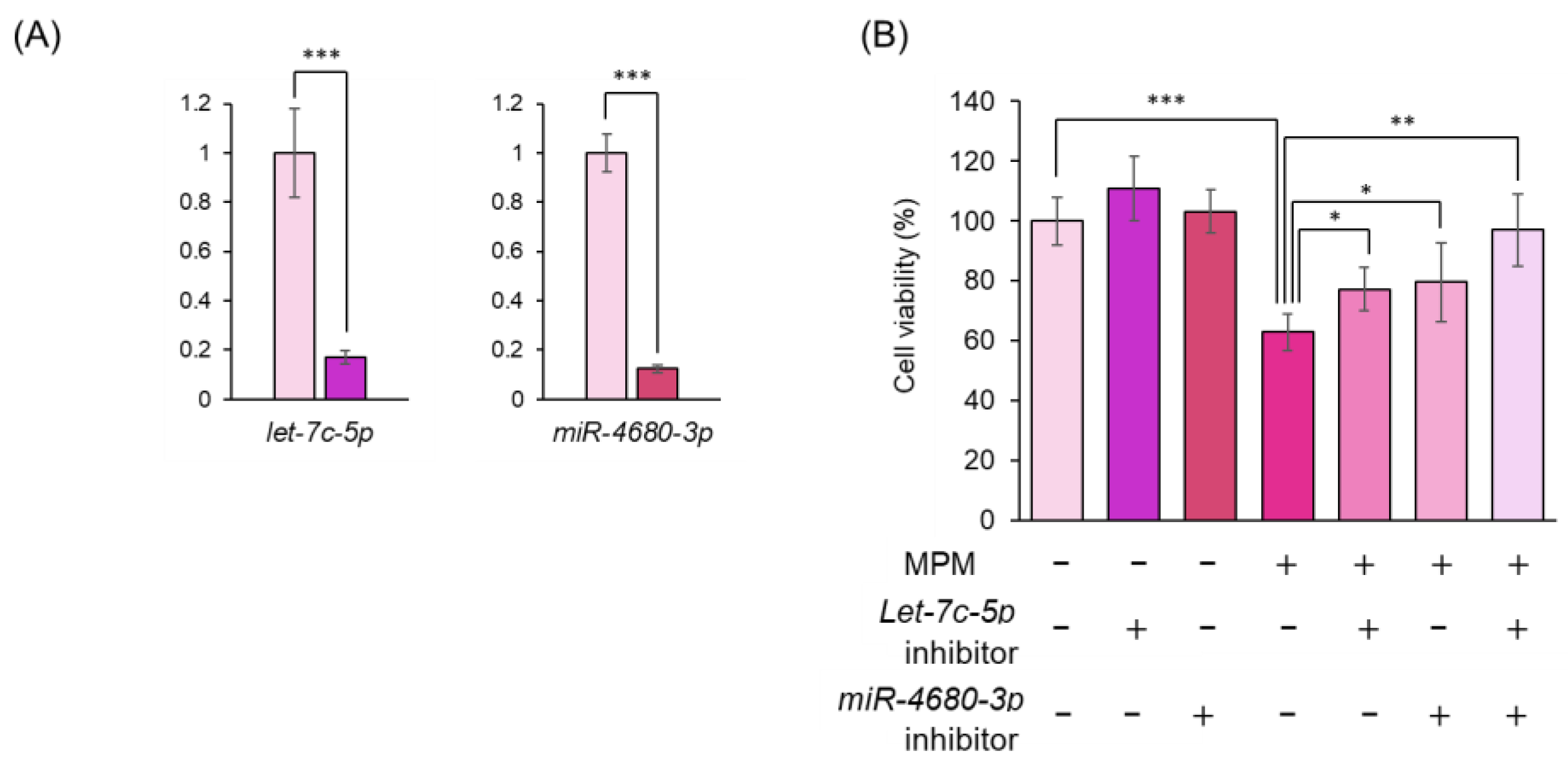

Table 1.
Antibody lists for western blotting.
| Antibody name |
Vendor | Catalog number |
Concentration |
|---|---|---|---|
| -ACTIN | Medical & Biological Laboratories | M177-3 | 1:3000 |
| BAX | Santa Cruz Biotechnology | sc-20067 | 1:1000 |
| Cleaved CASPASE-3 | Cell Signaling Technology | 9661 | 1:3000 |
| CCND1 | Santa Cruz Biotechnology | sc-8396 | 1:500 |
| CCNE | Santa Cruz Biotechnology | sc-377100 | 1:1000 |
| CDK2 | Santa Cruz Biotechnology | sc-6248 | 1:1000 |
| CDK4 | Santa Cruz Biotechnology | sc-56277 | 1:1000 |
| CDK6 | Santa Cruz Biotechnology | sc-53638 | 1:500 |
| BACH1 | Santa Cruz Biotechnology | sc-271211 | 1:500 |
| PAX3 | Santa Cruz Biotechnology | sc-376204 | 1:500 |
| ERBB2 | Santa Cruz Biotechnology | sc-393712 | 1:1000 |
| JADE1 | Proteintech Japan | 28472-1-AP | 1:2000 |
| Rabbit IgG HRP | Cell Signaling Technology | 7074 | 1:10000 |
| Mouse IgG HRP | Cell Signaling Technology | 7976 | 1:10000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



