Submitted:
02 January 2025
Posted:
07 January 2025
You are already at the latest version
Abstract
This article delves into the intricate mechanisms underlying the sensory integration in the execu-tive control of movement, encompassing ideomotor activity, predictive capabilities, and motor control systems. It examines the interplay between motor and sensory functions, highlighting the role of cortical and subcortical regions of the central nervous system in enhancing environmental interaction. The acquisition of motor skills, procedural memory, and the representation of actions in the brain are discussed emphasizing the significance of mental imagery and training in motor function. Development of this aspect of sensorimotor integration control can help to advances our understanding of the interactions between executive motor control, cortical mechanisms, and consciousness. Bridging theoretical insights with practical applications, it sets the stage for future innovations in clinical rehabilitation, assistive technology, and education. The ongoing explora-tion of these domains promises to uncover new pathways for enhancing human capability and well-being.
Keywords:
Sensorimotor integration
; Motor control
; Imagery
; Rehabilitation
1. Introduction
A fundamental characteristic of human sensory experience relies in the “unified” percept although sensory information come from different sensory modalities. This integration derives from the ability of combining multiple sources of sensory information by different neural circuits. Multisensory responses of superior colliculus neurons, and multisensory integration firstly observed in the superior colliculus of cats [1], has been now recognized a common feature of numerous brain areas of different species, both developmentally and evolutionarily. Multisensory integration of cross-modal signals has been described in prefrontal cortex which neurons are sensitive to both the synchrony and semantic context of vocalizations, but also in premotor, parietal and temporal cortices. Multisensory transformation has raised new way to understand computational principles like Bayesian inference through which has been possible to link multisensory integration (priors) at the level of neurons and behavior (regularities learning). The perception, thus, is determined by priors and sensory likelihoods, in turn, priors reflet accumulated history of sensory likelihoods. Interestingly, the evolution of multisensory integration models and evaluation of fake sensations like rubber hand illusion and full body illusion (virtual reality) are questioning about peripersonal space and bodily self-consciousness opening new way to understand and approach for psychopathologies like schizophrenia and autism spectrum disorders among others [2,3].
In the following a brief description of recent advancement on sensory integration and sensorimotor integration will be presented.
2. Sensory Integration
Traditionally, the sensory system has been described as one in which each of the five senses has its own specific receptors, dedicated pathways, and target regions. This concept, known as the 'Law of Specific Nerve Energies,' posits that each sense operates independently, following its own 'labeled line' [1]. However, subsequent research has revealed that different submodalities and cross-modal information can be spatially integrated and temporally summated, leading to the concept of multisensory integration. This paradigm shift has advanced our understanding of how sensory processing generates more coherent activation patterns across large-scale networks, thus enhancing the efficiency of information processing. Tactile signals, for example, are transmitted from the skin to the spinal cord and brainstem via two pathways: the 'direct' (paucisynaptic or monosynaptic) and 'indirect' (polysynaptic) routes. Recent evidence shows that these pathways converge onto the same brainstem neurons, supporting the idea that tactile sensation is produced by a combination of inputs [4], as observed in other sensory systems. The extent of this convergence onto individual brainstem relay neurons determines the type of information relayed to various brain regions. The discovery that specific sensory and multisensory neurons possess overlapping receptive fields was a significant step in advancing the understanding of subcortical structures, such as the superior colliculus and thalamus, in sensory processing. The superior colliculus, for instance, plays a crucial role in visual processing for gaze orientation. The superficial layers of the superior colliculus represent visual space, while neurons in the deep layers respond to visual, auditory, and somatosensory stimuli. These distinct sensory inputs are organized into somatotopic maps that are spatially aligned, ensuring that different sensory signals arising from the same event, spatially and temporally concordant, are mapped to the same point on the motor map. This alignment facilitates sensorimotor transformations essential for the superior colliculus role in detection, localization, and orientation behavior [1]. All sensory information, except for olfaction, passes through thalamic relay centers before reaching the sensory cortex. Thalamus mediates interactions between cortical and subcortical systems. Reciprocal connections exist between cortex and thalamus that enables the thalamus to function as an integrative hub. First-order thalamic nuclei receive their primary inputs from subcortical sources. The higher-order nuclei are primarily driven by cortical inputs. The thalamic reticular nucleus (TRN) serves as the principal source of intrathalamic inhibitory input. All sensory nuclei within the thalamus exhibit modality-specific topographical organization. External inputs to the thalamus modulate thalamic firing patterns via the TRN, inducing distinct oscillatory rhythms that regulate brainwave activity and states of arousal. Sensory information relayed from the thalamus to the cortex maintains the modality-specific topographic organization, ensuring accurate sensory mapping. In contrast, higher-order thalamic neurons, along with first-order neurons, project extensively to multiple cortical areas. This projection pattern forms an indirect trans-thalamic communication pathway between cortical regions, facilitating cross-modal sensory integration. Consequently, individual sensory cortical areas can receive input from a diverse array of thalamic nuclei. Cortico-thalamic inputs originate from various cortical regions and project to subcortical structures, such as the superior colliculus and basal ganglia. These reciprocal connections between the cortex and thalamus underpin the spatial and temporal modulation of incoming sensory information. Spatial modulation is governed by the feedback and feedforward interactions between cortico-thalamic and thalamo-cortical pathways. The topographic organization and spatial specificity of cortico-thalamic fibers enable mechanisms like surround suppression, where the center-surround retinotopic excitatory receptive fields are flanked by larger regions that suppress responses to visual stimuli. Cortical feedback can amplify and refine these responses, a process observed in both the auditory and somatosensory systems. Such modulation also occurs in the reverse direction. Through dynamic, bidirectional interactions between feedforward thalamocortical inputs and feedback corticothalamic inputs, sensory tuning is achieved. Temporal modulation is mediated by the TRN. When cortico-thalamic neurons fire at low frequencies, their activity is transiently enhanced before being suppressed by TRN-mediated thalamocortical inhibition. In contrast, high-frequency firing of cortical neurons leads to short-term facilitation, while TRN-mediated thalamocortical synapses undergo short-term depression. This mechanism allows TRN-induced depression to be overcome, resulting in sustained enhancement of thalamic activity, constrained within specific temporal windows. The temporal patterning of thalamic inputs significantly influences the cortical discrimination and detection of sensory stimuli [5]. The spatial and temporal principles of multisensory integration suggest that the closer two unisensory stimuli are (in space or time) the more likely they are to be integrated into a unified percept [3]. Beyond processing, stimuli coming from various cortical networks must be integrated each other to form coherent perception. Integration of information across sensory modalities occurs through the transient synchronization of neural oscillations. Oscillatory neural activity reflects the synchronized fluctuations in the summed postsynaptic activity of large neuronal populations. These oscillations can be broken down into distinct frequency components, each defined by specific amplitudes and phases. Various types of oscillatory responses, each linked to different forms of neural synchronization, can be identified. Research has shown that bottom-up processing primarily involves local networks operating in the gamma frequency band (>30 Hz), while top-down control, involving long-range integrative processing, engages lower frequency bands (<30 Hz) [6]. Multisensory processing depends on the coordinated activity of different cortical regions. Key mechanisms include local neural oscillations and functional connectivity between distant cortical areas. Emerging evidence indicates that neural oscillations in distinct frequency bands correspond to specific mechanisms in multisensory processing. It is proposed that oscillations in different frequency bands play roles in feedforward-feedback processing, attention modulation, and predictive coding during multisensory integration. These mechanisms may also operate concurrently [6]. Through the process of multisensory integration, not only are the external environment and the observer’s body unified, but also the individual who perceives the environment and the agent that interacts with the world are unified. In other words, multisensory integration plays a crucial role in shaping human–environment interactions from both the 'environment' and 'individual' perspectives. By expanding the discussion, the process of multisensory integration is rooted in the structure of bodily self-consciousness [3], concours at learning education and express cultural heritage when considered a significant aspect of the society [7]. The combination of signals from different sensory modalities can enhance perception and facilitate behavioral responses. In other words, the synthesis of complementary information coming from multiple sensory modalities improve perceptual performance, leading to more accurate and faster responses. Crossmodal interactions are governed by neural oscillations in different frequency bands that can occur at both early and late stages of processing and involve bottom-up and top-down mechanisms. Recent studies have examinated crossmodal facilitation of response speed, as manifested in the speeding of visual responses by concurrent task-irrelevant auditory information. These data reported that the crossmodal facilitation of response speed was associated with reduced early beta power in multisensory association and secondary visual areas. This beta power modulation has been supposed to reflect an early stage of behaviorally relevant crossmodal feedback influence from higher multisensory areas and secondary visual areas improving visual processing through attentional gating [8].
3. Sensorimotor Integration
The integration of sensory and motor functions underlies the brain's ability to plan and execute actions based on environmental cues. This process involves the continuous updating of motor plans based on sensory feedback, ensuring that movements are accurate and goal-directed. The ability to adapt motor actions in response to changing sensory information is essential for effective interaction with the environment. The execution of goal-oriented behaviors requires a spatially coherent alignment between sensory and motor maps. It has recently described a direct connection between collicular motor units and kinetic visual features that enable goal-directed behavior organizer to rapidly intercept moving and static targets [9]. Multisensory processing is critical for coordinating complex motor tasks. The brain integrates information from multiple sensory sources, such as vision, hearing, and touch, to create a comprehensive understanding of the environment [10]. This multisensory integration allows for more accurate and efficient motor control, enabling individuals to perform tasks that require the coordination of different sensory modalities. Sensory inputs play a vital role in guiding motor actions, with parieto-frontal circuits mediating visuomotor transformations. The brain integrates visual, auditory, and somatosensory information to guide movements, ensuring that sensory information is effectively translated into motor commands. Specific cortical areas are involved in visuomotor integration. The ventral intraparietal area (VIP) and the anterior intraparietal area (AIP) are reciprocally connected with F4 and F5 premotor area of the monkey’s frontal cortex. The VIP-F4 circuit is responsible for movements of reaching, approaching, and withdrawing with the arm, trunk, and face/mouth. Similarly, the AIP-F5 circuit guides visually directed grasping movements, integrating visual information about objects with motor commands to the hand of monkeys [11,12,13]. The most part of parietal cortex plays a crucial role in integrating sensory inputs with motor plans. It receives and processes information from various sensory modalities, creating a cohesive representation of the external environment. This information is then transmitted to motor areas, where it is used to guide actions. Due to interactions within the corticothalamic systems, the brain exhibits rhythmic neural activity [14]. Various oscillation patterns are associated with different brain functions, ranging from sensory and motor to cognitive tasks. Although there is not complete agreement [15], the phase of oscillations influences brain function, and the synchronization of oscillations within specific frequency bands—known as the communication-through-coherence model—can alter perception, attention, and working memory [16]. For instance, alpha oscillations in the parietal area are linked to controlling visual perception across different spaces. In contrast, beta and theta oscillations in the parietal region are involved in perceiving local versus global image features, with theta and beta oscillations respectively associated with the encoding of this information. These observations support the idea that brain oscillations facilitate a complex pattern of information processing and communication between different brain areas [16]. Beta oscillations with frequencies ranging from 13 to 30 Hz are thought to be involved in somatosensory processing and motor control and have been predominantly observed in sensorimotor cortices and basal ganglia. The origin of beta synchrony has been extensively discussed and associated with oscillations of the whole basal ganglia-cortical loop. Changes in beta oscillation power have been functionally linked to phases of movement. Typically, beta oscillations are prominent during stable postures and diminish during active states such as movement planning and execution. Beta oscillation power is spontaneously and bilaterally reduced in the premovement period and increased close to the premovement period. Similarly, the motor cortex shows a decrease in beta power during ipsilateral execution movements [17]. More specifically, supports for emerging of synchronous activity in the beta-band associated with pallido-striatal connections during static conditions, and in the gamma-band associated with pallido-subthalamic feedback loops during movement have been provided in a silica-based model [18]. A higher level of beta synchrony has been found linked to dopamine levels in the cortico-basal ganglia loop. Parkinson’s disease, a hypokinetic movement disorder resulting from the loss of nigral dopaminergic input to the striatum, is associated with disruption of beta-band oscillatory firing of the globus pallidum, subthalamic nucleus and pars reticulata of the substantia nigra [19]. These dynamics into the basal ganglia can contribute to the desynchronization of the beta frequency band in the motor cortex through the thalamo-cortical pathway [17]. A notable reduction in beta oscillation amplitude occurs in sensorimotor areas also during motor imagery or when observing actions. In conditions where motor function is functionally decreased as in aging, or pathologically impaired such as in Parkinson's disease, there is a corresponding disruption in beta oscillations, indicating an altered motor release. These oscillations have different roles, including reducing inhibition to initiate motor plans, maintaining current motor states against internal and external noises disruptions, and facilitating motor learning [20]. The hierarchical organization of motor control systems ensures that simple movements engage minimal cortical areas (partial autonomy of lower-level motor control [21], while complex movements activate broader networks, including the supplementary motor area, premotor cortex, parietal cortex, basal ganglia, and cerebellum. This hierarchical control allows for the smooth execution of movements and the ability to adapt to new and changing environments.
In the following a brief description of principal subcortical and cortical areas involved in motor control and sensorimotor integration are presented.
4. Basal Ganglia and Sensorimotor Integration
Most voluntary actions necessitate choosing a single action from a broad array of potential motor programs. This intricate process of action selection involves merging various signals, such as sensory and contextual data from the surroundings, previous experiences, and self-generated goals. All these information often conflict, requiring a system to choose the most adaptive and suitable action for varying circumstances. Such a system would need to preserve and individually represent both goals and signals, allowing for their independent modification and manipulation, and determine and execute one action from among the alternatives through downstream motor structures. These functions are performed by basal ganglia. Receiving input from virtually all cortical areas, the striatum is located at the interface for mediating cortico-subcortical interactions. The striatum converts a variety of signals from the cortex—including sensory, cognitive, and motor signals—into action selection cues that either inhibit or activate relevant motor programs within specific motor modules. Receiving dopaminergic inputs from the pars compacta of the substantia nigra, the striatum processes those environmental stimuli marked as significant ones during motor learning. The striatal internal structure, including the “direct” and “indirect” pathways identified by Alexander and Crutcher [22], along with the “striosomes” and “matrix” compartmentalization [23], illustrates the intricate framework of basal ganglia processing. This complexity is further detailed by the “funnelling” of the extensive cortical inputs into sensorimotor, limbic, and cognitive parallel loops [24]. These loops have been thought to originate from and return to the same input sites, showcasing the extensive potential for processing within the basal ganglia. On the other hand, it has been recently described that dorsolateral striatum receives excitatory inputs from both sensory and motor cortical regions. Both motor and sensory information are modulated by direct and indirect cortico-basal ganglia-thalamo-cortical pathway and altered by dopamine-depletion [25]. The direct pathway acts by disinhibiting the thalamus, ultimately facilitating the motor behavior being executed. The indirect pathway acts by promoting the tonic suppression of thalamic activity, thereby inducing the cessation of motor activity. According to this model, a selective loss of indirect projections would result in an increase in spontaneous involuntary movements, as seen in hemiballismus. Conversely, an increase in neuronal activity involved in the indirect pathway, along with the loss of direct connections, would lead to a reduction in thalamo-cortical facilitation, resulting in the bradykinetic and akinetic condition characteristic of parkinsonism. A third “hyperdirect” pathway connect the subthalamic neurons with the cortical inputs, bypassing the striatum, and excite pallidal output neurons so increasing their inhibitory output (Figure 1). The functional property of the hyperdirect pathway to inhibit a cortical motor program would enable the direct pathway of the basal ganglia to adjust the motor program according to the situation [26,27], and the subthalamic nucleus to dynamically control the effective decision limit over time [28]. The dopaminergic neurons of primate’s substantia nigra exhibit increased firing activity in response to “significant” sensory stimuli. This neuronal activity is linked to the nature of the reward that the animal expects to receive after executing a given command and is not directly related to the movement itself. The D1-D2 dopaminergic receptor subtypes are specifically inserted on striatal projecting neurons of the direct and indirect pathway and are respectively exited or inhibited when a significant stimulus evokes a shot of dopamine from the pars compacta of the substantia nigra [29]. These responses gradually diminish as the animal learns and automates the task. Thus, the dopaminergic system of the basal ganglia, by transmitting signals indicative of the incentive value of a given environmental stimulus, influences the acquisition of the conditioned response at the striatal level and, ultimately, modulates the behavior at the onset of learning and over time with the maintenance of the “success” that behavior has produced [30]. Striatal interneurons are actively involved in these processes. They gradually acquire a conditioned excitatory response as conditioning progresses, which is retained in long-term memory and used when the conditioned behavior is fully acquired and automated. Cholinergic interneurons in the striatum, also named tonically active neurons, detect coincidences between sensory inputs and behavioral context, adjusting the sensitivity of other striatal neurons to cortical inputs (Figure 1) [31,32,33]. Therefore, the sensory responses processed into the striatum can be modulated by motor activity thus accounting for the ability of the basal ganglia to control movement in relation to the contextual variability [25]. The sensorimotor cortical regions and the dorsolateral region of the human putamen are suggested to be the long-term storage structures where well-learned movement sequences are preserved. During motor learning, a concordant shifting of activations from anterior to posterior regions of basal ganglia that mirror changes in the motor areas of the frontal cortex has been described. Initially, new learning activates the dorsolateral prefrontal cortex and striatum; selected movements engage the premotor cortex and mid-putamen; and automatic movements activate the sensorimotor cortex and posterior putamen. Focusing on well-practiced actions, activation returns to the dorsolateral prefrontal cortex and striatum. Notably, the cerebellum remains inactive during new decisions, focused actions, or when selecting movements [34].
The consistent projection relaying basal ganglia with prefrontal areas implied in cognitive processing and mirror system suggest that basal ganglia contribute to higher levels of cognitive functions, such as planning, syllogistic reasoning, and mathematical problem solving. To this regard, using a gradient functional connectivity approach [35], have recently described a double orientation of human caudate functional relations with cortical networks confirming previous non-human data. A medial organization of these functional relations have been associated with internal orienting behavior, whereas the lateral extent of caudate is more related to external oriented behavior [36]. Noteworthy, recent evidence includes a clustered representations of five separate higher-order association networks within the caudate, positioned side-by-side, and linked to higher-order networks underlining language and social functions. These clusters are spatially separate and asymmetrically distributed paralleling the cortical lateralization [37].
5. Cerebellum and Sensorimotor Integration
The other important subcortical hub for sensorimotor integration is represented by cerebellum that might function expanding the range of sensorimotor associations in accordance with the dynamics shaping the behavioral condition, while adjusting the input-output coupling rules to fit the broader context [38]. Classically, the cerebellum has been primarily involved in motor learning including error detection and correction essential for refining motor skills over time. When a new motor act is performed, the cerebellum adjusts the timing and intensity of muscle activations to improve performance prevalently based on response feedback. With repetition and practice, these adjustments become more precise, resulting in smoother and more coordinated movements. Traditionally, two models have faced about cerebellar motor functions. The theory of “cerebellar control mechanism” [39], and the “cerebellar motor learning” mechanism [40,41]. Briefly, the motor learning model would function by modulating the responsiveness of Purkinje cells, thereby serving as a teaching or error-correction signal [42]. According to Llinás' model [43], the cerebellum is believed to function as a control mechanism, utilizing the rhythmic activity of the inferior olive to synchronize Purkinje cell populations, thereby fine-tuning coordination [41]. The most accepted role of cerebellum has classically regarded the “error encoding” function played by the Purkinje cells system under the climbing fibers driving information about sensory information incongruency as “complex spiking”. This way the cerebellum function as a comparator of inferior olive signals with respect to the cortical upcoming commands enabling corrections of motor plans whenever incongruences between them are detected [44]. This mechanism, imply that cerebellum, developing new input-output associations, is responsible for motor learning. Motor learning thus depends on cerebellar ability to map outgoing motor commands to the predicted sensory feedback resulting from those movements. The ability to learn through sensorimotor adaptation may be linked to the impairment in updating motor commands as result in patients affected by cerebellar disorders [45]. On the other hand, findings have shown that climbing fibers conveys a variety of non-error signals too. It has been observed that the “simple spike” activity of Purkinje cells predicts performance errors and conveys feedback signals related to movement kinematics. This perspective has strengthened the notion of a forward internal model represented by simple spike, which is updated on changes in the behavioral state through input from climbing fibers [46]. Beyond from the on-line motor control, cerebellar processing has been reported for persistent representation of information in motor areas of the frontal cortex. During motor planning, preparatory activity has been described in both cerebellum and motor frontal cortex. After cerebellar processing efferent projections reach the frontal cortex again through the thalamus, closing a cortico-cerebellar loop able to work on time scale of seconds [47]. Conceivably, Boven and Cerminara [48] have found that the ability of the cerebellum to support millisecond timescales might be intrinsic to cerebellar circuitry, and the ability to support supra-second timescales might result from cerebellar interactions with other brain regions, such as the prefrontal cortex. This way, the cerebellum would make predictions (Figure 2) and provide feedback over multiple time horizons spamming from milliseconds when engaged in motor control, to seconds or more when involved in decision-making, or long-term motor planning [49].
Hence, the neural mechanisms underlying sensory integration and motor control involve the coordinated activity of various cortical and subcortical regions. All these areas are highly interconnected, with extensive communication pathways that facilitate the integration of sensory and motor functions from planning to execution. The parietal cortex, the premotor cortex, and cerebellum work together to process sensory inputs and generate appropriate motor responses. The prefrontal cortex, the supplementary motor area and basal ganglia work together to process inputs and respond to demanding environment. The functional dissociation between these two internally vs externally guided loops have been broadly studied in the control of movements [50].
6. Cortical Areas and Sensorimotor Integration
Motor planning and control rely heavily on the brain's ability to predict the outcomes of movements and adjust subsequent actions accordingly. This predictive framework involves the representation of the intended movement and its consequences, thus forming the basis for abstract behaviors and logical movement sequences [51]. Motor schemas, which are sequences of muscle activations necessary to perform an action, are learned through experience and stored in procedural memory. The premotor cortex and supplementary motor area play critical roles in representing and executing movements (Figure 3) [52].
During the initial stages of motor learning, the premotor cortex, with its extensive connections to the parietal cortex, integrates external information to develop new motor programs. As the expertise becomes more refined, motor control shifts to the supplementary motor area, which is involved in executing learned and internally driven movements. The hierarchical motor control system engages various cortical and subcortical regions, ensuring smooth and adaptive motor performance. The primary motor cortex is primarily responsible for the detailed execution of movements, while higher-order areas like the premotor cortex and supplementary motor area plan and coordinate complex actions. The cerebellum and basal ganglia, as mentioned, are crucial for refining motor skills and ensuring precise execution. The cerebellum is involved in timing and coordinating movements, detecting and correcting errors, and adapting motor commands based on sensory feedback. The basal ganglia facilitate the selection and initiation of appropriate motor programs, playing a critical role in procedural memory and motor learning. The medial prefrontal cortex and the striatum are involved in matching results of behavior with expectation based on a predictive ability to represent the consequences of an action [53]. To this regard, it has been recently demonstrated that during learning of a rewarded task the default mode network (DMN) and dorsal attention network (DAN) underwent shifting of their connectivity. At the beginning of the learning, an increased functional coupling between several sensorimotor areas of the DAN (Premotor cortex) with salience/ventral attention network (anterior insula/inferior frontal gyrus and anterior cingulate cortex) has been recorded. On the contrary, during late learning, those DAN sensorimotor areas switch their connectivity to DMN areas (medial PF cortex) detecting a transition in the functional coupling of sensorimotor to transmodal cortex when adapting behavior. This shifting might reflect a transition from more exploration to exploitation activity when decision making can be made on established sensorimotor contingencies [54].
Predictive representations in the brain enable the anticipation of movement outcomes, which is essential for adjusting actions in real-time. The cerebellar role in error detection and correction involves monitoring the accuracy of movements and making necessary adjustments to improve performance [55]. This process is vital for learning new motor skills and refining existing ones, particularly based on the ability of the cerebellum to respond to reward-related error signals [56]. The ability to adapt motor plans based on sensory feedback is a hallmark of effective motor control. The integration of sensory inputs with motor commands allows the brain to continuously refine and optimize movements, ensuring that actions are goal-directed and efficient. This adaptability is crucial for responding to changing environmental conditions and achieving complex motor tasks. The parallel and synchronized interaction between cortical and subcortical structures allows mental representation of action and its consequences, the comprehension of other’s motor acts and temporal abstraction. A huge of learning practice, clinical therapeutics and technological applications have been started from these perspectives. Relevant in this last area is the concept that synthetic systems, like AI based devices, need to “learn” from motor behavior too. The relationship between imagination and reality is evident in the immediate neural adaptations that accompany physical exercise but also motivational and affective outcomes [57].
Because of what previously described, appear sharp to define a separation between motor and sensory function. Such a division has been vanished also in those cerebral areas ever engaged in motor control such as the primary motor cortex. During the execution of a varying location manual interception task, the persistent activity of most monkey’s neurons in the primary motor cortex resulted mixed and related both to reach-direction and target motion. The neural population activity exhibited a dynamical orbital geometry distributed along the reach-direction which slope was dependent by target-motion conditions. As result, primary motor neurons showed sensory modulation of motor output probably related to predictive sensorimotor control [58]. The seminal study of Rizzolatti and coworkers [59,60] had provided new insight about the complex organization of cortical and subcortical areas, traditionally implied in “pyramidal” and “extrapyramidal” loops, and their functional roles in sensorimotor integration. The posterior (F1-F5) parieto-dependent motor area relates to primary motor cortex and send direct projections to the spinal cord. The rich sensory information originating from parietal lobe enable the parieto-dependent motor area to process sensory-motor transformation. Contrarily, the anterior (F6-F7) prefronto-dependent areas do not send fibers to the primary motor cortex and show diffused connections with other motor areas receiving higher order cognitive information, relating this circuitry with motor planning and motivation. Particularly, the sensorimotor transformation by F5 neurons unable this area to categorize for specific actions by which is possible to plan movement starting by coding the action at different degrees of abstraction (prototypes of actions) that facilitates the association between sensory properties of the objects (affordances) and the appropriate motor plan to interact with these objects. These connections and functional evidence had permitted to postulate the principle by which there is direct coupling between action observation and action execution. The F5 neurons exhibiting such a property have been collected as integral part of the mirror system whose functional roles are implicated in action recognition and imitation [59]. As in monkey, several fMRI, TMS, EEG and electrophysiological studies have reported that also in human a fronto-parietal mirror system is operating. The cortical areas including inferior parietal lobule, ventral premotor cortex and the inferior frontal gyrus are involved in goal and intention of observed motor act. Based on activation of inferior frontal gyrus, an intransitive movement mirror system has been described as the basis of the imitation capacity [61]. The left sided human speech motor centers have been described as able to elicit motor representation of the heard phonemes in the corresponding motor representation of the same sound involved in the comprehension of word meaning [61,62,63]. Still, the representation of certain types of emotion has been found to activate the insula and cingulate sites able to evoke emotion and visceromotor expression of the same emotions [61,62,64]. These findings have raised the hypothesis that other’s emotions are recognized through the activation of that neural centers mediating the feeling of that emotions in the observer [65]. This way, specific body motor areas, hand and mouth principally, which representations are somatotopically replicated in different cortical areas (frontal, posterior parietal regions) are mirrored. The specific activity of some neurons placed in these cortical areas relates to general goals of motor actions. The consequent categorizations of different motor actions [63] enable the nervous system to code different motor acts, to generate a repertoire of actions and to activate their motor representation whenever observing the movements of others. These mechanisms are at the basis of the ability to understand what the others are doing mirroring the agent’s motion into the observer’s internal representation of that motion, that is to understand what goal the agent is reaching through that action, or to understand the intention of others [60,61,66]. Similarly, the aptitude to imitate others motor actions pertain to these mechanisms. Imitation learning, that consists of the ability of copying a behavior that is not previously present in the motor repertoire of the observer, is operated by dissecting the component of agent’s motion into string of component motor acts. The observer can “recognize” and code the sequence of each component in its own repertoire so that the new imitated actions become similar to the agent’s motion (ideomotor theory of action). The integration between internal imagery ability and external cues information enable to target different “goals” attributing them specific functional meanings [67].
Summarizing, the modality by which motor sequences are linked into new motor arrangements have been attributed to a distributed network composed by presupplementary motor area, supplementary motor area, dorsal premotor cortex, primary motor cortex, primary somatosensory cortex, superior parietal lobule, thalamus, basal ganglia, and cerebellum. Motor schemas, learned through trial and error, become automated and are stored in procedural memory, enabling efficient and coordinated movements. In the skilled sequential movements (those acquired after intensive and repetitive practice) the presupplementary motor area is involved in cognitive aspect of the early learning phase of movement sequences, the supplementary motor area is involved in the performance of memorized movement sequences, meanwhile, dorsal premotor area, ventral premotor area and primary moor area are densely interconnected with each other to a network for the control of hand movements. This organization seems to be in the possibility to give the mirror system the cognitive link for internally guided movements and the temporal organization of the motor sequencing [68]. Expertise resulting from prolonged practice is associated with cognitive adaptation and the development of mental representation that capture the specific relationship between action and contextual demands, which are crucial for the organization of action control [69]. The resulting predictive capabilities about the outcomes of those actions play a crucial role in motor control involving subsequent adjustments accounting for time estimation capability [70]. This prediction-based framework allows for abstract behaviors and logical movement sequencing, bridging the gap between sensory inputs and motor outputs.
7. Mental Imagery and Sensorimotor Integration
Neuroimaging technologies (PET, fMRI) but TMS too, have demonstrated that mental imagery relies on much of the same neural machinery as perception with the same modality. It can also engage mechanisms involved in memory, emotion and motor control. The early human visual cortex (areas 17, 18) for example, activates during imagery indicating that visual imagery engages the same structures involved in perception within their respective modalities [71]. Similarly, imagery of emotional events activates the same brain region involved in specific perceptual experiences including amygdala, insula and autonomic nervous system. Imagery is associated with all perceptual modalities. Odor imagery is associated with activity in the olfactory cortex, auditory imagery with bilateral activity in the secondary auditory cortices, tactile imagery with activity in the pri mary and secondary somatosensory areas and motor imagery with activity in the premotor cortex and somatosensory cortex [72]. On the other hand, motor imagery depends on distinct neural mechanisms although occurs alongside visual imagery. Mirror neurons could be involved in motor imagery, supporting the idea that people often manipulate images by imagining how objects would appear if moved in a particular way. Mental imagery can engage the motor system, which helps explain why ‘mental practice’ can enhance real performance. Imagining movements may not only stimulate relevant brain regions but also create connections among processes implemented in different areas, ultimately facilitating complex performances. Most of the neural processes that underlie modality perception are also used in imagery. Imagery, in many respects, can act as a substitute for perceptual stimulus “re-presenting” it. Imagery not only engages the motor system, but can also affect the body, similarly to actual perceptual experiences [71]. Ideomotor activity is fundamental to cognitive processes, including problem-solving, working memory, action simulation and internal models. Through these mechanisms, anticipatory images are generated. Musicians, for example, often mentally rehearse their finger movements to optimize performance, utilizing internal visual, auditory, and kinesthetic channels facilitating interpersonal coordination too [73]. The ability to generate mental images of actions, depending on internal and external factors, make possible to have accurate predictions about situations and other’s actions and mental states (social prediction) [74]. These mechanisms help the brain to create "internal notebooks" or working memories [75] and are the basis of cause-effect relationships that support the development of thought. The integration between internal imagery ability and external cues information enable to target different “goals” attributing them specific functional meanings [67]. The relationship between imagination and reality is manifest in the immediate neural adaptations that accompany physical exercise but also motivational and affective outcomes [57]. On the other hand, motor imagery varies in its association with motor planning, preparation, and potentially "suppressed" execution. The neural mechanisms involved depend on which stage of motor control is engaged. Additionally, it can differ based on how much sensory input—visual, auditory, kinesthetic, or vestibular—is involved, and whether the imagery is consciously generated or more implicit. Different types of motor imagery activate distinct brain regions. Visual imagery engages visual-related areas and the superior parietal lobe, with first-person perspectives activating sensorimotor areas more than third-person perspectives. Kinesthetic imagery, on the other hand, activates motor-related regions and the inferior parietal lobe. The degree of motor cortex activation is influenced by whether the task involves motor preparation or planning, with preparation leading to greater M1 activity. The premotor and supplementary motor areas are the primary regions activated by motor imagery, as they play crucial roles in the planning and preparation stages of motor control. The involvement of virtual sensory modalities in motor imagery can significantly alter the activity within the posterior parietal cortex. The ventral prefrontal cortex and anterior cingulate cortex have been found to participate in movement suppression during the preparation phase. The prefrontal cortex, anterior cingulate cortex, and premotor cortex are consistently reported as the most frequently activated areas during motor imagery [76]. When individuals visualize running, for instance, their bodies exhibit physiological responses similar to actual running, such as increased heart rate [77]. The supplementary motor area is activated during the mental rehearsal of processing of the spatial-temporal elements of movement sequences, highlighting the close relationship between mental representation and physical execution [78]. This activation occurs even in individuals with amputated limbs, where imagining movements with "phantom" limbs stimulates the motor cortex [79]. The physical practice has been reported to facilitate the corticospinal excitability during mental imagery under the same conditions [80]. Mental imagery is proposed to initiate from the frontal cortex, which then projects back to modality specific regions such as the visual cortex. People suffering with aphantasia, that are unable to voluntary generate mental images, result having deficits affecting all sensory modalities or modality specific. On the contrary, subjects with extremely and atypically vivid mental imagery, hyperphantasia, show a visual specific modality overscore of self reported imagery vividness. The strong functional connectivity in resting state fMRI between the visual occipital network and the prefrontal cortex of aphantasia suffering subjects seems to suggest that imagery impairments can depend on a reduced connectivity between executive control regions and the visual cortex [72]. Then, despite being a cognitive process, motor imagery shares control mechanisms and neural substrates with actual movement and sensorimotor integration, offering insights into how movements are controlled by the brain. Defined as the mental rehearsal of movements without physical execution, motor imagery is used in sports, rehabilitation and robotics.
8. Implications
Motor imagery has gained attention as a tool for neurorehabilitation and brain-machine interfaces (BMIs). Motor imagery, as the ability to identify the congruence between action observation and execution, forms the basis of Action Observation Treatment, AOT, a therapeutic tool in neurodegenerative disease, in brain lesioned patients but also for maintenance of motor skills in elderly people, or amelioration of sport performance [81]. Ideomotor training is a technique that involves mentally rehearsing a movement without physically performing it with the aim of improving motor skills by engaging the brain’s motor planning and control system. Ideomotor training specifically emphasizes mentally rehearsing a movement focusing on brain’s ability to translate imaged movement into physical execution. It is often used in rehabilitation or skill acquisition. As mentioned, motor imagery involves mentally simulating or visualizing a movement without any actual physical movement with the goal to engage motor systems by imagining the movement in detail. It is often used for improving performance in sports, rehabilitation and brain training. The key difference between motor imagery and ideomotor training lies in their focus and application. While both involve mental rehearsal, ideomotor training is more explicitly about the mental-to-physical link and tends to be more directed toward preparing the body for movement through mental practice. Effective ideomotor training requires relaxation and focus, enabling individuals to “experience” themselves performing an action through internal visual, auditory, and kinesthetic sensations. This mental imagery stimulates the relevant muscles, boosting the impact of physical training. However, mental rehearsal alone cannot substitute for physical practice, as the kinesthetic aspect of movement are refined through actual execution. Ideomotor training can enhance muscle strength by up to 20%, underscoring the neural contribution to physical improvement [77]. The training technique of mental rehearsal, or visualization, is a vital aspect of athletic mental training. By vividly imagining performing a sport gesture, athletes can improve their skills without physical movement, provided that this mental practice is complemented by actual physical practice [77]. As a technique that involves visualizing movements without physical execution, ideomotor training, or mental practice, enhances motor performance by activating motor areas and increasing muscle tone, supporting physical practice. Athletes and performers often use mental rehearsal to improve their skills, engaging all sensory channels to vividly imagine the action. Effective ideomotor training involves a combination of relaxation and concentration, allowing individuals to “perceive” themselves performing the action through internal visual, auditory, and kinesthetic channels. This mental imagery pre-activates the relevant muscles, enhancing the effectiveness of physical training. However, mental practice cannot replace the need for physical practice, as the kinesthetic aspects of movement are developed through actual execution. Mental rehearsal is particularly beneficial for complex gestures, where detailed visualization can enhance motor learning and execution. By imagining the movements vividly, individuals can improve muscle memory and coordination, complementing their physical training routines. The combination of mental and physical practice leads to better overall performance and skill acquisition [82]. The physiological mechanisms underlying ideomotor training involve the activation of motor cortical areas similar to those engaged during physical practice. Functional imaging studies have shown that mental rehearsal activates the primary motor cortex, premotor cortex, and supplementary motor area. This neural activation is thought to prime the motor system, making it more responsive to subsequent physical practice [83]. Ideomotor training can significantly improve motor learning and performance. Individuals who engage in mental rehearsal in addition to physical practice exhibit greater improvements in skill acquisition compared to those who rely solely on physical practice. This suggests that mental imagery can enhance the consolidation of motor skills in the brain. Combining mental and physical training is particularly effective for learning complex motor tasks. Mental rehearsal allows individuals to practice movements in their minds, refining their motor plans and improving coordination. When combined with physical practice, this approach leads to more efficient and effective motor learning, resulting in better performance and skill mastery. Also, it has been showed that the simulation of virtual reality may induce a modulation in the motor responses of physical reality [84]. Stroke patients treated for simultaneous observation of repetitive movements displayed on a monitor peripheral nerve stimulation and concurrent application of their own imagination of the sensations arising from physically performing the same action (AOT) have showed measurable improvement in dexterity [85]. Interestingly, studying the temporal accuracy of motor imagery of people with PD, Lambert [86] had indicated that these patients imagine movements differently with respect to their more affected versus less affected side. The side-specific impairments result in an increased dependence on visual and cognitive processes to successfully execute motor imagery involving the more affected side, as a consequence of the lateralized deficits in the accurate processing of kinesthetic information [86]. Mental imagery is used by humans to intuitively solve most daily planning tasks. A novel algorithm based on “imaged” images has been proposed to enable robot to use existing scene to generate action plans. This method, called simulated mental imagery for planning (SiMIP) is different from deep learning and permit robot to operate through parsing process unlike to predict the entire scenes thus enhancing the success rate of the plans whenever a feedback loop is incorporated [87]. Wearable robots, on the other hand, need to be embodied with wearers to facilitate motor and sensory reconstruction and enhancement. Control strategies targeting this embodiment are focused in integrating multisensory information (biomechatronic chips) to generate coherent control parameters configurations and improve human-robot interaction [88].
9. Conclusions
Investigating the functions of different cerebral regions and how they were intermingled, the history of our comprehension of sensory and motor system has been changed to reach an integrated and merged ability of different functions raising from different areas of all cerebral tissue. Nowadays there are growing evidence that the combined functions of all these regions are in the possibility to harmonize sensorimotor integration until to emerging of new motor expression even to transfer this one to external avatar living in the virtual reality. Before we have completely described the sensorimotor basis of the “embodiment” of motor control, it is passing out, overtaken by imagery. Generating internal representations of actions is central to cortical motor function, the external contingencies and motivational factors determine then whether these action representations are transformed into actual (or virtual) actions through decisional processes that lead to action initiation. Related research reveals that motor imagery and action observation activate similar neural circuits as performing the actions themselves. This insight is pivotal for developing therapeutic strategies, especially for rehabilitation in patients with motor control disorders or brain injuries. By leveraging motor imagery, therapists can design interventions that stimulate relevant brain regions, potentially accelerating recovery and enhancing motor learning. Understanding these complex mechanisms can inform strategies for improving motor function and rehabilitation in individuals with motor impairments. The integration of sensory and motor functions is critical for effective interaction with the environment, enabling individuals to adapt their actions based on feedback. The role of predictive capabilities and procedural memory in motor learning highlights the importance of repetition and practice in acquiring and refining motor skills.
Rehabilitation programs can incorporate motor imagery and action observation to stimulate the brain's natural learning processes without physical execution. This approach is particularly beneficial for stroke rehabilitation, where patients often struggle with regaining motor functions. Furthermore, these findings can inform the design of brain-computer interfaces (BCIs) that translate neural signals into actionable commands for prosthetic limbs, enhancing the quality of life for individuals with severe motor impairments.
Conflicts of Interest
The author declares no conflict of interest.
References
- Stein, B.E.; Stanford, T.R.; Rowland, B.A. Multisensory Integration and the Society for Neuroscience: Then and Now. J. Neurosci. 2020, 40. [Google Scholar] [CrossRef] [PubMed]
- Angelaki, D.E.; Gu, Y.; DeAngelis, G.C. Multisensory Integration: Psychophysics, Neurophysiology, and Computation. Curr. Opin. Neurobiol. 2009, 19. [Google Scholar] [CrossRef] [PubMed]
- Noel, J.P.; Blanke, O.; Serino, A. From Multisensory Integration in Peripersonal Space to Bodily Self-Consciousness: From Statistical Regularities to Statistical Inference. Ann. N. Y. Acad. Sci. 2018, 1426. [Google Scholar] [CrossRef] [PubMed]
- Turecek, J; Ginty, D. How Two Intermingled Sensory Pathways Combine to Encode Touch. Nature 2022. [Google Scholar] [CrossRef]
- Cruz, K.G.; Leow, Y.N.; Le, N.M.; Adam, E.; Huda, R.; Sur, M. Cortical-Subcortical Interactions in Goal-Directed Behavior. Physiol. Rev. 2023, 103. [Google Scholar] [CrossRef]
- Keil, J.; Senkowski, D. Neural Oscillations Orchestrate Multisensory Processing. Neuroscientist 2018, 24. [Google Scholar] [CrossRef]
- Parker, M.; Spennemann, D.H.R.; Bond, J. Sensory and Multisensory Perception—Perspectives toward Defining Multisensory Experience and Heritage. J. Sens. Stud. 2024, 39. [Google Scholar] [CrossRef]
- Michail, G.; Senkowski, D.; Holtkamp, M.; Wächter, B.; Keil, J. Early Beta Oscillations in Multisensory Association Areas Underlie Crossmodal Performance Enhancement. Neuroimage 2022, 257. [Google Scholar] [CrossRef]
- González-Rueda, A.; Jensen, K.; Noormandipour, M.; de Malmazet, D.; Wilson, J.; Ciabatti, E.; Kim, J.; Williams, E.; Poort, J.; Hennequin, G.; et al. Kinetic Features Dictate Sensorimotor Alignment in the Superior Colliculus. Nature 2024, 631, 378–385. [Google Scholar] [CrossRef]
- Collins, A.G.E.; McDougle, S.D. Context Is Key for Learning Motor Skills. Nature 2021, 600, 387–388. [Google Scholar] [CrossRef]
- Iacoboni, M.; Dapretto, M. The Mirror Neuron System and the Consequences of Its Dysfunction. Nat. Rev. Neurosci. 2006, 7. [Google Scholar] [CrossRef] [PubMed]
- Rizzolatti, G.; Luppino, G. The Cortical Motor System. Neuron 2001, 31. [Google Scholar] [CrossRef] [PubMed]
- Rizzolatti, G.; Cattaneo, L.; Fabbri-Destro, M.; Rozzi, S. Cortical Mechanisms Underlying the Organization of Goal-Directed Actions and Mirror Neuron-Based Action Understanding. Physiol. Rev. 2014, 94, 655–706. [Google Scholar] [CrossRef] [PubMed]
- Steriade, M. Grouping of Brain Rhythms in Corticothalamic Systems. Neuroscience 2006, 137, 1087–1106. [Google Scholar] [CrossRef]
- Jones, S.R. When Brain Rhythms Aren’t ‘Rhythmic’: Implication for Their Mechanisms and Meaning. Curr. Opin. Neurobiol. 2016, 40. [Google Scholar] [CrossRef]
- Thut, G.; Miniussi, C.; Gross, J. The Functional Importance of Rhythmic Activity in the Brain. Curr. Biol. 2012, 22. [Google Scholar] [CrossRef]
- Villalobos, N.; Almazán-Alvarado, S.; Magdaleno-Madrigal, V.M. Elevation of GABA Levels in the Globus Pallidus Disinhibits the Thalamic Reticular Nucleus and Desynchronized Cortical Beta Oscillations. J. Physiol. Sci. 2022, 72. [Google Scholar] [CrossRef]
- Blenkinsop, A.; Anderson, S.; Gurney, K. Frequency and Function in the Basal Ganglia: The Origins of Beta and Gamma Band Activity. J. Physiol. 2017, 595. [Google Scholar] [CrossRef]
- Alavi, M.; Dostrovsky, J.O.; Hodaie, M.; Lozano, A.M.; Hutchison, W.D. Spatial Extent of Beta Oscillatory Activity in and between the Subthalamic Nucleus and Substantia Nigra Pars Reticulata of Parkinson’s Disease Patients. Exp. Neurol. 2013, 245, 60–71. [Google Scholar] [CrossRef]
- Barone, J.; Rossiter, H.E. Understanding the Role of Sensorimotor Beta Oscillations. Front. Syst. Neurosci. 2021, 15. [Google Scholar] [CrossRef]
- Merel, J.; Botvinick, M.; Wayne, G. Hierarchical Motor Control in Mammals and Machines. Nat. Commun. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Alexander, G.E.; Crutcher, M.D. Functional Architecture of Basal Ganglia Circuits: Neural Substrates of Parallel Processing. Trends Neurosci. 1990, 13. [Google Scholar] [CrossRef] [PubMed]
- Graybiel, A.M. A Stereometric Pattern of Distribution of Acetylthiocholinesterase in the Deep Layers of the Superior Colliculus. Nature 1978, 272. [Google Scholar] [CrossRef] [PubMed]
- Alexander, G.E.; DeLong, M.R.; Strick, P.L. Parallel Organization of Functionally Segregated Circuits Linking Basal Ganglia and Cortex. Annu. Rev. Neurosci. 1986, VOL. 9, 357–381. [Google Scholar] [CrossRef]
- de la Torre-Martinez, R.; Ketzef, M.; Silberberg, G. Ongoing Movement Controls Sensory Integration in the Dorsolateral Striatum. Nat. Commun. 2023, 14. [Google Scholar] [CrossRef]
- Nambu, A.; Tokuno, H.; Takada, M. Functional Significance of the Cortico-Subthalamo-Pallidal ‘hyperdirect’ Pathway. Neurosci. Res. 2002, 43, 111–117. [Google Scholar] [CrossRef]
- Chen, W.; de Hemptinne, C.; Miller, A.M.; Leibbrand, M.; Little, S.J.; Lim, D.A.; Larson, P.S.; Starr, P.A. Prefrontal-Subthalamic Hyperdirect Pathway Modulates Movement Inhibition in Humans. Neuron 2020, 106, 579-588.e3. [CrossRef]
- Ding, L. Contributions of the Basal Ganglia to Visual Perceptual Decisions. Annu. Rev. Vis. Sci. 2023, 9, 385–407. [Google Scholar] [CrossRef]
- Condé, H. Organization and Physiology of the Substantia Nigra. Exp. Brain Res. 1992, 88. [Google Scholar] [CrossRef]
- Schultz, W. Dopamine Reward Prediction-Error Signalling: A Two-Component Response. Nat. Rev. Neurosci. 2016, 17. [Google Scholar] [CrossRef]
- Matityahu, L.; Gilin, N.; Sarpong, G.A.; Atamna, Y.; Tiroshi, L.; Tritsch, N.X.; Wickens, J.R.; Goldberg, J.A. Acetylcholine Waves and Dopamine Release in the Striatum. Nat. Commun. 2023, 14. [Google Scholar] [CrossRef]
- Threlfell, S.; Lalic, T.; Platt, N.J.; Jennings, K.A.; Deisseroth, K.; Cragg, S.J. Striatal Dopamine Release Is Triggered by Synchronized Activity in Cholinergic Interneurons. Neuron 2012, 75. [Google Scholar] [CrossRef] [PubMed]
- Apicella, P. The Role of the Intrinsic Cholinergic System of the Striatum: What Have We Learned from TAN Recordings in Behaving Animals? Neuroscience 2017, 360, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Moretti, R.; Caruso, P.; Crisman, E.; Gazzin, S. Basal Ganglia: Their Role in Complex Cognitive Procedures in Experimental Models and in Clinical Practice. Neurol. India 2017, 65. [Google Scholar]
- O’Rawe, J.F.; Leung, H.C. Topographic Organization of the Human Caudate Functional Connectivity and Age-Related Changes with Resting-State FMRI. Front. Syst. Neurosci. 2022, 16. [Google Scholar] [CrossRef]
- Menon, V.; D’Esposito, M. The Role of PFC Networks in Cognitive Control and Executive Function. Neuropsychopharmacology 2022, 47. [Google Scholar] [CrossRef]
- Kosakowski, H.L.; Saadon-Grosman, N.; Du, J.; Eldaief, M.C.; Buckner, R.L. Human Striatal Association Megaclusters. J. Neurophysiol. 2024, 131, 1083–1100. [Google Scholar] [CrossRef]
- Manzoni, D. The Cerebellum and Sensorimotor Coupling: Looking at the Problem from the Perspective of Vestibular Reflexes. Cerebellum 2007, 6, 24–37. [Google Scholar] [CrossRef]
- Kazantsev, V.B.; Nekorkin, V.I.; Makarenko, V.I.; Llinás, R. Olivo-Cerebellar Cluster-Based Universal Control System. Proc. Natl. Acad. Sci. U. S. A. 2003, 100. [Google Scholar] [CrossRef]
- Freeman, J.H. Cerebellar Learning Mechanisms. Brain Res. 2015, 1621, 260–269. [Google Scholar] [CrossRef]
- Cheron, G.; Márquez-Ruiz, J.; Dan, B. Oscillations, Timing, Plasticity, and Learning in the Cerebellum. Cerebellum 2016, 15. [Google Scholar] [CrossRef]
- Morton, S.M.; Bastian, A.J. Cerebellar Control of Balance and Locomotion. Neuroscientist 2004, 10. [Google Scholar] [CrossRef] [PubMed]
- Llinás, R.R. The Olivo-Cerebellar System: A Key to Understanding the Functional Significance of Intrinsic Oscillatory Brain Properties. Front. Neural Circuits 2014, 7. [Google Scholar] [CrossRef] [PubMed]
- Pisotta, I.; Molinari, M. Cerebellar Contribution to Feedforward Control of Locomotion. Front. Hum. Neurosci. 2014, 8. [Google Scholar] [CrossRef]
- Leech, K.A.; Roemmich, R.T.; Gordon, J.; Reisman, D.S.; Cherry-Allen, K.M. Updates in Motor Learning: Implications for Physical Therapist Practice and Education. Phys. Ther. 2022, 102. [Google Scholar] [CrossRef] [PubMed]
- Streng, M.L.; Popa, L.S.; Ebner, T.J. Cerebellar Representations of Errors and Internal Models. Cerebellum 2022, 21. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Davis, C.; Thomas, A.M.; Economo, M.N.; Abrego, A.M.; Svoboda, K.; De Zeeuw, C.I.; Li, N. A Cortico-Cerebellar Loop for Motor Planning. Nature 2018, 563. [Google Scholar] [CrossRef]
- Boven, E.; Cerminara, N.L. Cerebellar Contributions across Behavioural Timescales: A Review from the Perspective of Cerebro-Cerebellar Interactions. Front. Syst. Neurosci. 2023, 17. [Google Scholar] [CrossRef]
- Boven, E.; Pemberton, J.; Chadderton, P.; Apps, R.; Costa, R.P. Cerebro-Cerebellar Networks Facilitate Learning through Feedback Decoupling. Nat. Commun. 2023, 14. [Google Scholar] [CrossRef]
- Debaere, F.; Wenderoth, N.; Sunaert, S.; Van Hecke, P.; Swinnen, S.P. Internal vs External Generation of Movements: Differential Neural Pathways Involved in Bimanual Coordination Performed in the Presence or Absence of Augmented Visual Feedback. Neuroimage 2003, 19. [Google Scholar] [CrossRef]
- Henschke, J.U.; Pakan, J.M.P. Engaging Distributed Cortical and Cerebellar Networks through Motor Execution, Observation, and Imagery. Front. Syst. Neurosci. 2023, 17. [Google Scholar] [CrossRef]
- Virameteekul, S.; Bhidayasiri, R. We Move or Are We Moved? Unpicking the Origins of Voluntary Movements to Better Understand Semivoluntary Movements. Front. Neurol. 2022, 13. [Google Scholar]
- Klein-Flügge, M.C.; Bongioanni, A.; Rushworth, M.F.S. Medial and Orbital Frontal Cortex in Decision-Making and Flexible Behavior. Neuron 2022, 110. [Google Scholar] [CrossRef] [PubMed]
- Nick, Q.; Gale, D.J.; Areshenkoff, C.; De Brouwer, A.; Nashed, J.; Wammes, J.; Zhu, T.; Flanagan, R.; Smallwood, J.; Gallivan, J. Reconfigurations of Cortical Manifold Structure during Reward-Based Motor Learning. Elife 2023, 12. [Google Scholar] [CrossRef]
- Raymond, J.L. Research on the Cerebellum Yields Rewards. Nature 2020, 579. [Google Scholar] [CrossRef]
- Sendhilnathan, N.; Semework, M.; Goldberg, M.E.; Ipata, A.E. Neural Correlates of Reinforcement Learning in Mid-Lateral Cerebellum. Neuron 2020, 106. [Google Scholar] [CrossRef]
- Simonsmeier, B.A.; Andronie, M.; Buecker, S.; Frank, C. The Effects of Imagery Interventions in Sports: A Meta-Analysis. Int. Rev. Sport Exerc. Psychol. 2021, 14. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, Y.; Wang, T.; Cui, H. Neural Geometry from Mixed Sensorimotor Selectivity for Predictive Sensorimotor Control 2024. [CrossRef]
- Rizzolatti, G.; Luppino, G. The Cortical Motor System. Neuron 2001, 31, 889–901. [Google Scholar] [CrossRef]
- Rizzolatti, G.; Fogassi, L.; Gallese, V. Neurophysiological Mechanisms Underlying the Understanding and Imitation of Action. Nat. Rev. Neurosci. 2001, 2. [Google Scholar] [CrossRef]
- Fabbri-Destro, M.; Rizzolatti, G. Mirror Neurons and Mirror Systems in Monkeys and Humans. Physiology 2008, 23. [Google Scholar] [CrossRef]
- Iacoboni, M. Neurobiology of Imitation. Curr. Opin. Neurobiol. 2009, 19. [Google Scholar] [CrossRef]
- Rizzolatti, G.; Fabbri-Destro, M.; Cattaneo, L. Mirror Neurons and Their Clinical Relevance. Nat. Clin. Pract. Neurol. 2009, 5. [Google Scholar] [CrossRef] [PubMed]
- Rizzolatti, G.; Cattaneo, L.; Fabbri-Destro, M.; Rozzi, S. Cortical Mechanisms Underlying the Organization of Goal-Directed Actions and Mirror Neuron-Based Action Understanding. Physiol. Rev. 2014, 94, 655–706. [Google Scholar] [CrossRef] [PubMed]
- Rizzolatti, G.; Sinigaglia, C. The Functional Role of the Parieto-Frontal Mirror Circuit: Interpretations and Misinterpretations. Nat. Rev. Neurosci. 2010, 11. [Google Scholar] [CrossRef] [PubMed]
- Rizzolatti, G.; Ferrari, P.F.; Rozzi, S.; Fogassi, L. The Inferior Parietal Lobule: Where Action Becomes Perception. In Percept, Decision, Action: Bridging the Gaps; 2008; pp. 129–145 ISBN 9780470034989.
- Fogassi, L. The Mirror Neuron System: How Cognitive Functions Emerge from Motor Organization. J. Econ. Behav. Organ. 2011, 77. [Google Scholar] [CrossRef]
- Ohbayashi, M. The Roles of the Cortical Motor Areas in Sequential Movements. Front. Behav. Neurosci. 2021, 15. [Google Scholar] [CrossRef]
- Land, W.M.; Volchenkov, D.; Bläsing, B.E.; Schack, T. From Action Representation to Action Execution: Exploring the Links between Cognitive and Biomechanical Levels of Motor Control. Front. Comput. Neurosci. 2013. [Google Scholar] [CrossRef]
- Spapé, M.M.; Serrien, D.J.; Ravaja, N. 3-2-1, Action! A Combined Motor Control-Temporal Reproduction Task Shows Intentions, Motions, and Consequences Alter Time Perception. Heliyon 2023, 9. [Google Scholar] [CrossRef]
- Kosslyn, S.M.; Ganis, G.; Thompson, W.L. Neural Foundations of Imagery. Nat. Rev. Neurosci. 2001, 2. [Google Scholar] [CrossRef]
- Muraki, E.J.; Speed, L.J.; Pexman, P.M. Insights into Embodied Cognition and Mental Imagery from Aphantasia. Nat. Rev. Psychol. 2023, 2. [Google Scholar] [CrossRef]
- Keller, P.E. Mental Imagery in Music Performance: Underlying Mechanisms and Potential Benefits. Ann. N. Y. Acad. Sci. 2012, 1252. [Google Scholar] [CrossRef]
- Thornton, M.A.; Tamir, D.I. Neural Representations of Situations and Mental States Are Composed of Sums of Representations of the Actions They Afford. Nat. Commun. 2024, 15. [Google Scholar] [CrossRef] [PubMed]
- Jeannerod, M. Mental Imagery in the Motor Context. Neuropsychologia 1995, 33. [Google Scholar] [CrossRef] [PubMed]
- Hanakawa, T. Organizing Motor Imageries. Neurosci. Res. 2016, 104. [Google Scholar] [CrossRef] [PubMed]
- Mulder, T. Motor Imagery and Action Observation: Cognitive Tools for Rehabilitation. In Proceedings of the Journal of Neural Transmission; 2007; Vol. 114. [Google Scholar]
- Cona, G.; Semenza, C. Supplementary Motor Area as Key Structure for Domain-General Sequence Processing: A Unified Account. Neurosci. Biobehav. Rev. 2017, 72. [Google Scholar] [CrossRef]
- Raffin, E.; Mattout, J.; Reilly, K.T.; Giraux, P. Disentangling Motor Execution from Motor Imagery with the Phantom Limb. Brain 2012, 135. [Google Scholar] [CrossRef]
- Kitamura, M.; Kamibayashi, K. Changes in Corticospinal Excitability during Motor Imagery by Physical Practice of a Force Production Task: Effect of the Rate of Force Development during Practice. Neuropsychologia 2024, 201. [Google Scholar] [CrossRef]
- Rizzolatti, G.; Fabbri-Destro, M.; Nuara, A.; Gatti, R.; Avanzini, P. The Role of Mirror Mechanism in the Recovery, Maintenance, and Acquisition of Motor Abilities. Neurosci. Biobehav. Rev. 2021, 127. [Google Scholar] [CrossRef]
- Frank, C.; Kraeutner, S.N.; Rieger, M.; Boe, S.G. Learning Motor Actions via Imagery—Perceptual or Motor Learning? Psychol. Res. 2024, 88. [Google Scholar] [CrossRef]
- Frank, C.G.A.V.S. Imagery and Motor Learning: A Special Issue on the Neurocognitive Mechanisms of Imagery and Imagery Practice of Motor Actions. Psychol. Res. 2024, 88, 1785–1789. [Google Scholar] [CrossRef]
- Ida, H.; Fukuhara, K.; Ogata, T. Virtual Reality Modulates the Control of Upper Limb Motion in One-Handed Ball Catching. Front. Sport. Act. Living 2022, 4. [Google Scholar] [CrossRef]
- Seitz, S.; Schuster-Amft, C.; Wandel, J.; Bonati, L.H.; Parmar, K.; Gerth, H.U.; Behrendt, F. Effect of Concurrent Action Observation, Peripheral Nerve Stimulation and Motor Imagery on Dexterity in Patients after Stroke: A Pilot Study. Sci. Rep. 2024, 14. [Google Scholar] [CrossRef] [PubMed]
- Lambert, K.J.M.; Singhal, A.; Leung, A.W.S. The Lateralized Effects of Parkinson’s Disease on Motor Imagery: Evidence from Mental Chronometry. Brain Cogn. 2024, 178. [Google Scholar] [CrossRef]
- Li, S.; Kulvicius, T.; Tamosiunaite, M.; Wörgötter, F. Simulated Mental Imagery for Robotic Task Planning. Front. Neurorobot. 2023, 17. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.; Zhang, Y.; Rajabi, N.; Taleb, F.; Yang, Q.; Kragic, D.; Li, Z. Shaping High-Performance Wearable Robots for Human Motor and Sensory Reconstruction and Enhancement. Nat. Commun. 2024, 15. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The basal ganglia pathway. (a) A simplified diagram of the cortico-basal ganglia-thalamus loop. (b) Multiple types of neuromodulatory inputs to the striatum. Abbreviations: 5HT, serotonin; ACh, acetylcholine; GPe, external segment of the globus pallidus; GPi, internal segment of the globus pallidus; LC, locus coeruleus; NE, norepinephrine; PPN-LDT, pedunculopontine nucleus-laterodorsal tegmental complex; SNc, substantia nigra pars compacta; SNr, substantia nigra pars reticulata; TAN, tonically active neurons; VTA, ventral tegmental area [modified from reference 28].
Figure 1.
The basal ganglia pathway. (a) A simplified diagram of the cortico-basal ganglia-thalamus loop. (b) Multiple types of neuromodulatory inputs to the striatum. Abbreviations: 5HT, serotonin; ACh, acetylcholine; GPe, external segment of the globus pallidus; GPi, internal segment of the globus pallidus; LC, locus coeruleus; NE, norepinephrine; PPN-LDT, pedunculopontine nucleus-laterodorsal tegmental complex; SNc, substantia nigra pars compacta; SNr, substantia nigra pars reticulata; TAN, tonically active neurons; VTA, ventral tegmental area [modified from reference 28].

Figure 2.
Activation in cortical and cerebellar regions during movement execution, observation, and imagery [from reference 51].
Figure 2.
Activation in cortical and cerebellar regions during movement execution, observation, and imagery [from reference 51].
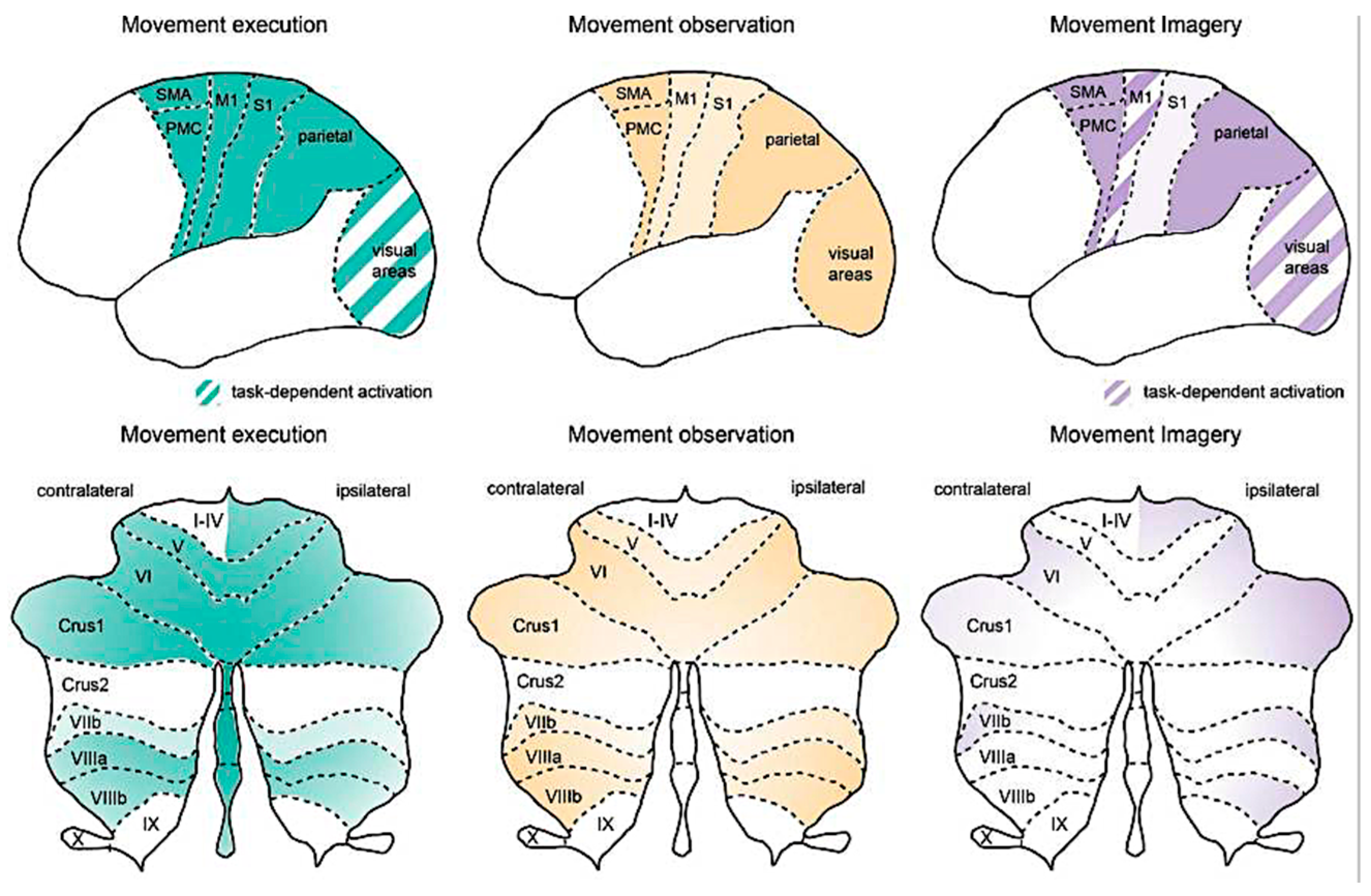
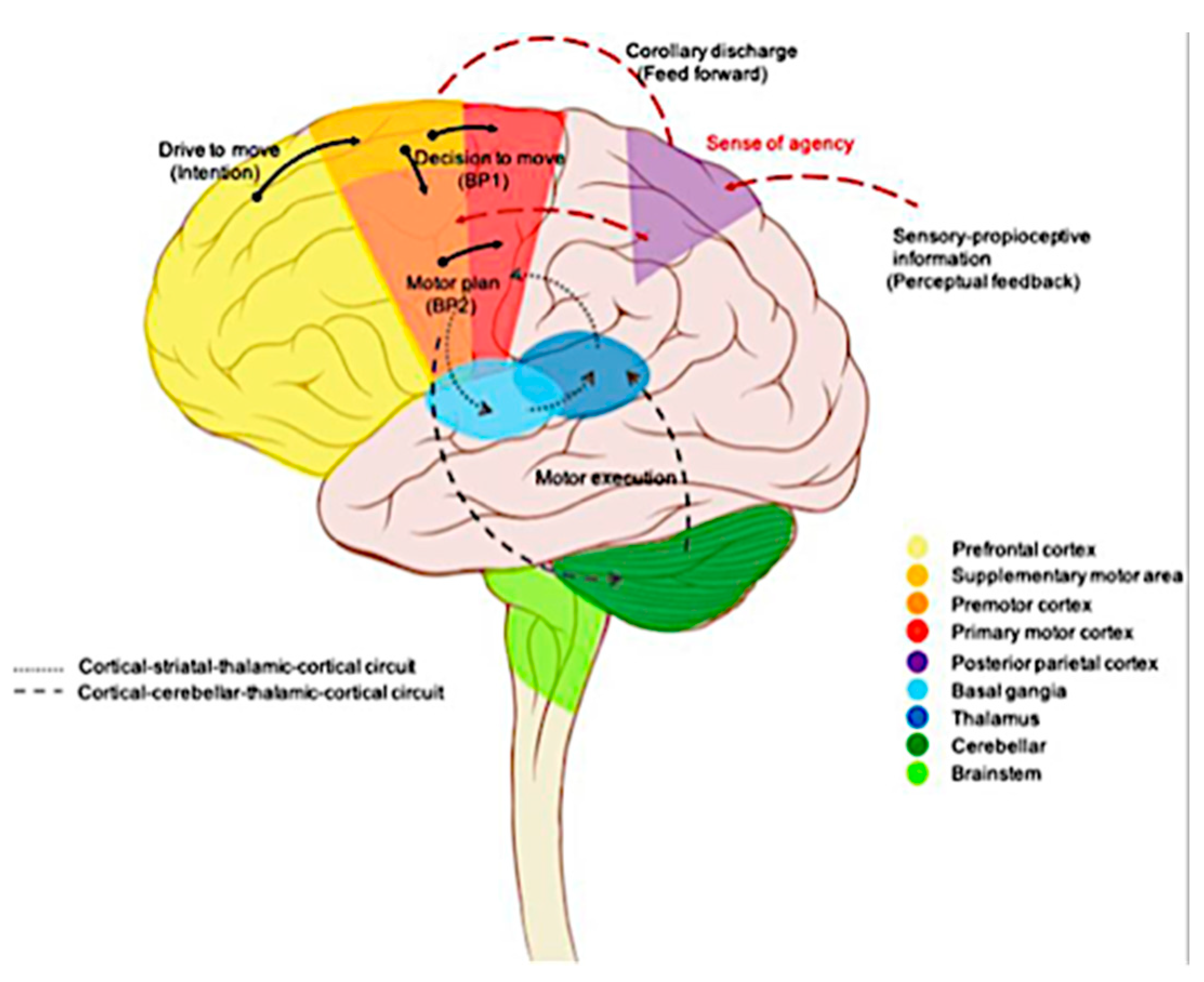
Figure 3.
Schematic representation of voluntary movement circuitry. The intention to move is generated in the prefrontal cortex and limbic area. The complex movement sequences of movements are subsequently programmed in the presupplementary and supplementary motor areas. The premotor cortex is primarily involved in movements selection based on external information from the parietal cortex. The presupplementary and supplementary motor areas, along with the premotor cortex, produce the readiness potentials (BP1) that is transmitted through the motor cortex to the basal ganglia and cerebellum for motor control modulation. The processed information is then sent back to the motor cortex via the thalamus. Simultaneously, a corollary discharge (feedforward model) is generated and directed to the parietal cortex to compare with proprioceptive feedback, thereby creating a sense of agency. Ultimately, the neural signal exits the primary motor cortex (BP2) to the spinal cord and contralateral muscles, initiating the actual movement [from reference 52].
Figure 3.
Schematic representation of voluntary movement circuitry. The intention to move is generated in the prefrontal cortex and limbic area. The complex movement sequences of movements are subsequently programmed in the presupplementary and supplementary motor areas. The premotor cortex is primarily involved in movements selection based on external information from the parietal cortex. The presupplementary and supplementary motor areas, along with the premotor cortex, produce the readiness potentials (BP1) that is transmitted through the motor cortex to the basal ganglia and cerebellum for motor control modulation. The processed information is then sent back to the motor cortex via the thalamus. Simultaneously, a corollary discharge (feedforward model) is generated and directed to the parietal cortex to compare with proprioceptive feedback, thereby creating a sense of agency. Ultimately, the neural signal exits the primary motor cortex (BP2) to the spinal cord and contralateral muscles, initiating the actual movement [from reference 52].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



